
Submitted:
11 October 2023
Posted:
13 October 2023
You are already at the latest version
Abstract
With rising rates of human obesity, the study aimed to determine the relationship between maternal diet-induced obesity and offspring morphometrics and behaviour in mice. Pregnant and lactating female mice fed a diet high in fat and sugar (HFHS) commonly consumed by human populations showed decreased food, calorie, and protein intake but increased adiposity at the expense of lean mass. The pre-weaning body weight of HFHS offspring was reduced for the first postnatal week but not thereafter with HFHS female offspring having higher body weights by weaning, due to continuing higher fractional growth rates. Post-weaning, there were minor differences in offspring food and protein intake. Maternal diet, however, affected fractional growth rate and total body fat content of male but not female HFHS offspring. Maternal diet did not affect offspring locomotor activity or social behaviour in either sex. Both male and female HFHS offspring displayed reduced anxiety-related behaviours with sex differences in particular aspects of the elevated plus maze task. In the novel object recognition task, performance was impaired in male but not female HFHS offspring. Collectively, the findings demonstrate that maternal obesity alters the growth, adiposity, and behaviour of male and female offspring, with sex-specific differences.
Keywords:
adiposity
; high fat and sugar diet
; dietary intake
; obesity
; behavioural neuroscience
1. Introduction
Human epidemiological and experimental animal studies have shown that adult phenotype can be programmed by sub-optimal intrauterine conditions resulting in intrauterine growth restriction, such as under-nutrition, stress, and/or hypoxia [1,2]. More recently, evidence has grown that adverse intrauterine environments can also affect neurodevelopment with consequences for adult behaviour [3]. With increasing consumption of obesogenic diets globally, there has also been greater emphasis on over-nutrition and maternal obesity in developmental programming as more women enter pregnancy with a high body mass index (BMI). In the UK, approximately 20-28% of women in antenatal care are classified as obese (BMI ≥30kg/m2), while >30% are overweight [4], which has significant implications for a successful pregnancy outcome and long-term child health [5]. Compared to mothers with a healthy BMI (18.5-24.9 kg/m2), pregnant women with a high BMI are more prone to gestational diabetes mellitus and stillbirth, while their infants are at greater risk of abnormal birthweight and adult cardio-metabolic dysfunction [5,6]. More recent epidemiological studies indicate that, by adolescence, these infants are also more likely to develop neurodevelopmental conditions such as intellectual disability, anxiety, autism spectrum disorder, attention deficit hyperactivity disorder and schizophrenia [7,8]. However, whether these behavioural disorders are caused by maternal obesity during pregnancy and lactation and/or by a later obesogenic environment remains unclear.
Critical periods for programming offspring behaviour can be studied more systematically in experimental animals like rodents in which dietary intake, growth and adiposity can be measured precisely throughout the lifespan. Maternal diet-induced obesity in rodents has been shown to influence locomotor activity, anxiety and social-related behaviours and cognition in the offspring. However, these studies show conflicting outcomes [9,10,11,12]. They have also concentrated on high fat diets rather than diets high in both fat and sugar more commonly consumed by obese human populations [13]. Indeed, the high fat diets used experimentally often have a lower sugar content than standard, control diets [14]. Detailed descriptions of maternal and offspring caloric and protein intake, and of somatic growth and adiposity are not often reported in studies investigating the impact of a maternal obesogenic diet on behavioural outcomes of their adult offspring. In addition, many of these behavioural studies concentrate on the male offspring because of the potential variability in female data caused by oestrus cycling [15].
With the greater emphasis now on investigating both sexes simultaneously [16], this murine study tested the hypothesis that maternal obesity induced by a diet high in fat and sugar has differential effects on the adult behaviour of their male and female offspring fed a post-weaning control diet. Offspring behaviour was assessed using the open field, elevated plus maze, social preference and novel object recognition tasks with reference to the biometry, adiposity and dietary intakes of the offspring and their mothers.
2. Materials and Methods
2.1. Experimental Design
All animal experimentation was carried out under the UK Home Office Animals (Scientific Procedures) Act 1986, following ethical approval by the University of Cambridge (project licence number PC6CEFE59). A total of 233 C57Bl/6J mice were studied. Of these, 106 were females purchased commercially for breeding (Charles River, Margate, UK) while the remainder were their offspring. All mice were group-housed (n=3-7 mice per cage) for most of the experimental period under a 12:12 h dark/light photocycle with ad libitum access to food and water. The majority of dams and offspring (n = 178) were fed a standard rodent diet (RM3, Special Dietary Services [SDS], Witham, UK; 11%kcal fat, 62%kcal carbohydrate of which 7%kcal is simple sugar, 27%kcal protein and a water content of 10%). The remaining females used for breeding (n=55) were fed a customised highfat, high sugar (HFHS) diet to induce obesity. This diet was made by combining high-fat diet pellets (D12451 diet, Research Diets Inc., Denmark) with condensed milk (Carnation, Nestle, Gatwick, UK) and water to form patties that were baked at 55°C for 46–48 h, as described previously [17]. The final nutritional composition of the HFHS diet was 38% kcal fat, 45% kcal carbohydrate of which 33% kcal was simple sugars, 17% kcal protein, with a water content of 12%: a diet similar in composition to that consumed by obese women reported previously [18]. Both mouse diets were replenished every 48 h during the period of the study to ensure palatability.
Eight week old female mice were fed the standard or HFHS diet for 6 weeks before mating and throughout pregnancy and lactation. Body weight was measured weekly before, during and after pregnancy on both diets. For a subset of the cages, intakes of food, kilocalories and protein were calculated by weighing food consumed in each cage over 24-48 h periods and totalling the food intake per week. Values are expressed as grams or kilocalories per day per mouse. After 6 weeks on the diet, a subset of animals in each dietary group were killed by cervical dislocation to measure body fat and lean mass by Dual Energy X-ray Absorptiometry (DEXA) scanning (Lunar PIXImus densitometer; GE Healthcare) and the weights of individual fat deposits before pregnancy (n = 7 per group). The remaining females were mated with males fed the control diet. At day 18.5 days of pregnancy when the dams had been on their respective diets for 9-10 weeks (17-18 weeks of age), a further subset from each dietary group was killed by cervical dislocation to measure maternal fat deposit weights and DEXA body composition (term 20.5 days, n=9 per group). The remaining control and HFHS-fed pregnant dams were then single-housed until their pups were weaned at postpartum (PP) day 21. At PP2, litters were reduced to 6 pups per dam. All postpartum dams were killed by cervical dislocation after weaning at approximately 20 weeks of age (after 12-13 weeks on their respective diets), with fat deposit weights and DEXA body composition measured in a subset of each dietary group.
At weaning, the offspring were group housed by sex and maternal diet, and fed the control diet. Pups were sexed and weighed at postnatal (PN) day 2 and then weighed again at PN7, PN14 and PN21 upon weaning. Thereafter, they were weighed weekly until behavioural assessments began at 13 weeks (PN91). Fractional growth rates (FGR) were calculated over specific time periods by dividing the weight increment over the period by the weight at the start of each period. In a subset of the offspring cages, intake of food, kilocalories and protein per mouse was measured weekly as grams or kilocalories per day. A total of 127 offspring were studied (n=64 from control dams, n=32 females, n=32 males; n=63 from HFHS dams, n=32 females, n=31 males). At the end of the behavioural assessments, all offspring were killed by cervical dislocation at 14 weeks (PN98) with DEXA body composition and/or fat deposit weights measured in a subset of both females and males in each dietary group.
2.2. Behavioural testing
At intervals of 3-4 days, offspring carried out two different behavioural tasks. They were tested either in the elevated plus maze (EPM) followed by the novel object recognition (NOR) tasks, or with the combination of the open field (OF) and social preference (SP) tasks. Only one male and one female from a litter were tested in a particular combination of behavioural tasks. Female mice were tested in proestrus to account for the behavioural effects of oestrus cycling [15,19]. Each testing session was recorded with a ceiling-mounted webcam and the videos analysed manually blinded to the cohort where possible.
The OF and EPM tasks were used to assess novelty, locomotion, exploration, and anxiety- and risk-taking behaviours [20]. In the OF arena, the number of grid lines crossed was used as an index of locomotion, while the time spent in the centre and periphery, and the number of rears, were used as a measure of exploration and anxiety-related behaviour [19]. For the EPM, the number of entries and time spent in the open and closed arms, the number of explorations to the end of the open arm, and the total number of rears were used as an index of exploration, anxiety- and risk-taking behaviours [19].
The SP task is designed to assess a mouse’s preference for interacting with a social stimulus versus a non-social stimulus [21]. Test mice were first habituated to the testing arena before being exposed to the unfamiliar sex- and strain-matched intruder mouse, as described previously [19]. Interaction time with the intruder (social stimulus -TS) and non-social stimulus (TNS) was quantified, and the social preference index (SPI) was calculated:
SPI=(TS-TNS)/(TS+TNS)
The NOR task was used to evaluate cognition, particularly recognition memory [22]. During the acquisition phase, two identical objects were placed at diagonal corners of the test box. The retention phase of the test was set up identically to the acquisition phase, except that one of the familiar objects was replaced with a novel object. A discrimination index (DI) was calculated for each animal to indicate which object was favoured as DI=[time spent investigating novel object – time spent investigating familiar object]/total time investigating objects. A preference for the novel or familiar object is indicated by a positive or negative DI respectively.
2.3. Statistical analysis
All statistical analyses were performed using GraphPad Prism (version 9.5.1 for Windows, GraphPad Software, San Diego, CA, USA). A Shapiro–Wilk normality test was used to examine if data were normally distributed. Data are presented as mean ± standard error of the mean (SEM). A t-test or Mann–Whitney non-parametric test was used, as appropriate, to compare biometric and behavioural measures between two groups (control versus HFHS). A mixed effects analysis or two-way ANOVA followed by a Sidak’s post hoc test was used to compare the effects of diet and age on food intake and body mass. Statistical significance was accepted as p≤0.05.
3. Results
3.1. Dietary intake and biometry of the dams
The HFHS diet affected the nutrient intake of dams pre-pregnancy and throughout pregnancy and lactation (Figure 1). Across the 6 weeks pre-pregnancy, the HFHS diet significantly reduced intake of food and protein, with a similar tendency for kilocalorie intake compared to controls (Figure 1A-C). Time per se did not affect any of the dietary intakes pre-pregnancy (Figure 1A–C). During pregnancy, there were significant effects of both diet and pregnancy day on the intakes of food, calories and protein (Figure 1D–F) with no interaction between diet and pregnancy day. Similarly, during lactation, both diet and day had significant effects on the intake of food, calories and protein (Figure 1G–I) with no interaction between factors. Overall, these dietary intakes were lower on the HFHS diet, particularly later in pregnancy and throughout lactation (Figure 1D–I).
The diet affected body weight, total body fat mass and individual fat deposit masses in all three reproductive states (Figure 2; Table 1). Before pregnancy, there was a significant effect of both diet and time on diet on total body mass with a significant interaction between the two factors such that body weight increased more with age on the HFHS diet (Figure 2A). Overall, these factors and their interaction remained significant during pregnancy and lactation (Figure 2B,C). Body weights of HFHF dams remained significantly elevated until day 14.5 of pregnancy but were lower than controls at PP2 and PP7 (Figure 2B,C), although body weight did not differ with diet by the end of pregnancy or lactation (Table 1). Total fat mass and weights of the individual fat deposits were significantly higher on the HFHS than the control diet at the expense of lean mass irrespective of reproductive state (Figure 2G–L, Table 1). In addition, the adrenal glands were heavier in HFHS than control-fed pre-pregnant mice, but not during pregnancy or lactation, while the liver of pregnant but not pre-pregnant or lactating dams was heavier on the HFHS diet (Table 1).
3.2. Dietary intake and biometry of the offspring
Overall, during pre-weaning development, there was a significant effect of age but not diet on body weight (Figure 3A,B). There was a significant interaction between age and diet with significantly lower body weights in HFHS than control pups of both sexes at PN2 and PN7 and higher body weights in females but not males at PN21 (Figure 3A,B). The FGR of the pre-weaning pups decreased significantly with age and was higher overall in HFHS than control pups of both sexes, particularly after PN7, remaining higher for longer in the females (Figure 3C,D).
After weaning, intakes of food, kilocalories and protein increased with age from PN21 to PN91 in both sexes but were only affected by maternal diet in the males with lower intakes in the HFHS than control offspring; there were no interactions between age and diet on the dietary intakes in either sex (Figure 4A–F).
There was a significant effect of increasing offspring age on post-weaning body weight and a significant interaction with maternal diet overall in both sexes (Figure 5A,B), although body weight did not differ with maternal diet in either sex at PN98 (Table 1). Post-weaning FGR decreased with age in both sexes and was also lower in male but not female HFHS offspring during the first week after weaning (Figure 5C,D). At PN98, total fat mass was significantly higher in male but not female HFHS offspring, with no differences in lean mass weight with maternal diet in either sex (Figure 5E–H). Weights of the gonadal, retroperitoneal and perirenal fat deposits were all significantly greater in HFHS than control male offspring whereas, in HFHS females, only perirenal fat weighed more than control values (Table 1) Adrenal weight was less in HFHS than control offspring in males but not females (Table 1). At PN98, maternal diet had no effect on the weights of the brain, liver or heart of the offspring in either sex (Table 1).
3.3. Behaviour and cognition function of the adult offspring
In the OF test, there were no significant differences in offspring locomotion (number of lines crossed), anxiety-related behaviour (entries into the centre or as duration in the centre), or exploratory rearing with maternal diet in either sex (Supplementary Figure S1). In the EPM task, female but not male offspring of HFHS dams spent significantly more time in the open arm (Figure 6A,B), while male but not female offspring of HFHS dams made significantly more full entries into the open arm (Figure 6C,D) than the offspring of control dams. In both sexes, HFHS offspring made more explorations to the end of the open arm than their control counterparts (Figure 6G,H).
Maternal diet has no significant effect on social preference of either sex (Supplementary Figure S2). In the NOR task, there was no significant difference in the time spent sniffing two identical objects during the acquisition phase in either male or female offspring of HFHS dams (Figure 7A–D). In the retention phase of the NOR task, DI differed significantly with maternal diet in the male but not female offspring (Figure 7E–H). Control males and females had a positive DI due to spending more time sniffing the novel object while male HFHS offspring had a DI close to zero, indicating equal time spent sniffing the familiar and novel object (Figure 7E,F). Maternal diet had no effect on the retention phase of the NOR task in the female offspring (Figure 7F,H). This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
4. Discussion
This study demonstrates that feeding female mice a HFHS diet reduces food intake and increases adiposity both before and throughout pregnancy and lactation. This maternal HFHS diet also results in sex-specific effects on the growth trajectory, adiposity, anxiety-related behaviour and cognitive function of their adult offspring, which are summarised in Table 2.
4.1. Dam food intake and biometry
Females fed the HFHS diet consumed significantly less food which resulted in reduced calorie and protein intakes throughout pregnancy and lactation. Despite the decreased dietary intake, females fed the HFHS diet significantly increased their adiposity in all three reproductive states, which was associated with an increased body mass until near the end of pregnancy. Body mass of HFHS dams fell during the early stages of lactation but increased to control values by the end of lactation when body fat content was still 100% greater in HFHS than control dams. Previous studies using similar HFHS diets have also shown increased body weight and adiposity during pregnancy and lactation with some of the greatest increases in fat mass seen in dams that had ad libitum access to the highly calorific sweetened condensed milk [23,24]. In the current study, increased fat accumulation was accompanied by a loss of lean mass irrespective of reproductive state, which may reflect, in part, the reduced dietary and protein intake. With less lean mass and more fat, the HFHS mice may have had a reduced basal metabolic rate and expenditure of energy for processes such as locomotion and thermoregulation, which would contribute to a lower calorie requirement [25,26]. In addition, the enhanced fat deposition in HFHS dams may have increased concentrations of leptin, the anorexigenic peptide, consistent with their reduced food intake. Indeed, increased leptin concentrations have been observed in previous studies of pregnant HFHS dams depending on the stage of pregnancy [17,27].
4.2. Offspring food intake and biometry
In human populations, obesity during pregnancy can affect intrauterine growth, resulting in small for gestational age, growth restricted or large for gestational age infants [28]. In the current study, the body mass of both female and male HFHS pups was reduced during the first week of life, which may reflect the reduced maternal intake kilocalorie and protein intake during pregnancy and/or early lactation as well as the transient fall in maternal weight postpartum with the onset of lactation. Previous studies using maternal low protein and the current HFHS diet have shown fetal growth restriction at the end of gestation in association with impaired placental growth and morphology [17,29]. During the suckling period, HFHS pups exhibited catch-up growth irrespective of sex, a phenomenon commonly seen in neonates growth restricted in utero [30]. The accelerated growth during suckling was maintained for longer in HFHS females and stopped at weaning in both sexes when the offspring were transferred onto the control diet. Compared to the controls, the males but not female HFHS offspring reduced their fractional growth rate in the first week post-weaning and consumed less food, kilocalories and protein throughout the 14 week postnatal study period without a significant decrease in their body mass. Despite this, adult male HFHS offspring had a higher fat mass than controls, while the fat mass of adult HFHS females was unaffected by maternal diet. These differences in offspring growth trajectory and adult body composition suggest that male HFHS offspring, in particular, are probably programmed to be more metabolically efficient.
A number of previous human epidemiological and animal experimental studies have shown increased offspring fat deposition in response to maternal obesity during pregnancy but this is most commonly associated with offspring hyperphagia and an increased body mass from weaning [23,31]. In many of the experimental studies in rodents, maternal obesity was induced by feeding high fat (HF) rather than HFHS diets, with the offspring often studied at older ages than in the current study. Young adult offspring of obese mothers have been shown to increase their fat deposition in the absence of increased body weight in rats and sheep [32,33], in line with the current findings. The outcome of maternal obesity on offspring morphometry, therefore, appears to depend on a range of factors including species, offspring sex and age, and the specific maternal and post-weaning diet. The current findings suggest that offspring appetite, metabolism and/or growth regulation can be programmed in a sex-linked manner by early life exposure to maternal diet-induced obesity.
4.3. Offspring behaviour and cognitive function
The current study clearly shows that aspects of offspring behaviour and cognitive function are altered in both sexes by a maternal HFHS diet and concomitant obesity. Multiple measures obtained from the EPM task indicated reduced anxiety-like behaviour in both male and female HFHS offspring, as evidenced by increased entries into the anxiogenic open arm environment and a greater number of visits to the distal end of the open arm. Previously, increased [34,35], decreased [36,37], or no change [38] in anxiety-related behaviours have been reported for offspring of obese dams, which may relate to offspring age [36,37]. The greater frequency with which the HFHS offspring travelled to the distal ends of the open arm of the EPM in the current study could also be interpreted as increased risk-taking behaviour. In a human study, an association between overweight/obesity and engaging in risky behaviours has been reported in adolescents [39], and warrants further investigation in the HFHS mouse offspring.
Male but not female HFHS offspring displayed a deficit in object recognition memory compared to control offspring. This finding aligns with the available literature on compromised recognition memory in adult offspring of mothers with obesity [40,41,42,43]. In contrast, there was no effect of the maternal HFHS diet on offspring social behaviour. Recent studies support the current finding [36,44], while others have reported maternal HF diet-induced disruptions to social processes in the offspring [45]. The observed effects of maternal diet-induced obesity on offspring social behaviour may be heavily dependent on the age and sex of the offspring tested [44]. An interplay of social and anxiety-related behaviour may explain why HFHS offspring were comfortable interacting with the intruder mouse, with lower anxiety-related behaviour in HFHS offspring facilitating approach to, and interaction with, an unknown intruder mouse. Further behavioural analysis is needed to understand the contribution of anxiety in social situations for HFHS offspring.
In the current study, a maternal HFHS diet from pre-pregnancy through to lactation did not affect offspring locomotor activity in the open field arena. This finding agrees with several previous studies of maternal HF models [42,46]. On the other hand, a maternal HF diet has been shown to lead to hyperactivity in male offspring, along with increased speed of movement within the open field arena [36,41]. However, data from juvenile and adult offspring in experimental studies of maternal obesity suggest that enhanced locomotor activity is only temporary and diminishes with age [10], in agreement with the current findings in the adult HFHS offspring.
Determining the primary mechanism of developmental programming of adult behaviour by the maternal HFHS diet is difficult, due to the complexity of disentangling the maternal obesogenic milieu from the intrauterine and subsequent postnatal environments. Indeed, the current HFHS diet is known to cause sex-specific differences in the feto-placental phenotype with alterations in the fetal nutrient supply and growth [17], which may contribute to the disparities observed postnatally between the male and female offspring reported here. In male HFHS offspring, there were changes in post-weaning dietary intake, growth profile and adult adiposity, while in female HFHS offspring, only the neonatal growth trajectory was altered relative to controls. The changes in anxiety-related behaviours in both sexes of HFHS offspring may, therefore, indicate a dependence on the combined effects of maternal obesity and the postnatal growth profile, independent of offspring adiposity, while the memory deficit of the male offspring may depend more heavily on their increased adiposity. Multiple interrelated mechanisms have been proposed to underlie offspring neurodevelopmental programming by diet-induced maternal obesity including oxidative stress and inflammation, altered hypothalamic-pituitary-adrenal axis activity, dysregulation of dopaminergic and serotonergic signalling and reward circuitry, perturbations in growth factor-mediated synaptic plasticity, and impaired insulin, glucose, and leptin signalling in the brain (see [7]. For instance, the reduction in adrenal mass in male HFHS offspring may affect corticosterone production and/or hypothalamic-pituitary-adrenal axis activity, resulting in blunted stress responsiveness and lowered anxiety. Increased leptin availability due to the increased adiposity of HFHS male offspring may also influence their phenotype [47]. Recent evidence supports the role of leptin in higher cognitive functions, particularly hippocampal-dependent learning and memory [48].
5. Conclusions
Collectively, the current data show that a HFHS diet fed before and throughout mouse pregnancy and lactation alters maternal dietary intake and body composition with sex-specific consequences for offspring somatic growth, adiposity, anxiety-related behaviour and spatial memory. Maternal diet-induced obesity, therefore, has widespread programming effects on offspring development with important implications for the long-term impacts of current obesogenic diets on the phenotypical diversity of human populations.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org, Figure S1: Effect of a maternal obesogenic diet on adult offspring behaviour in the open field; Preprints.org, Figure S2: Effect of a maternal obesogenic diet on adult offspring social interaction behaviour.
Author Contributions
EJC and ALF designed the study; EJC and EJM carried out the experiments; EJM and SH evaluated the behavioural tasks; EJM, ALF, EJC and SJ analysed the data, EJM and SJ created the figures; EJC, ALF and SJ wrote the text, all authors reviewed and revised the paper.
Funding
This research was funded by an MRC STP studentship to Emily J. Mort (Grant number; MR/N013433/1).
Institutional Review Board Statement
The animal study protocol was approved by the University of Cambridge Ethics Committee (project licence number PC6CEFE59).
Data Availability Statement
The data presented in this study are openly available in [repository name e.g., FigShare] at [doi], reference number [reference number].
Acknowledgments
We would like to thank the staff of the University Biomedical Services for their technical assistance and care of the animals.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barker, D.J. The developmental origins of chronic adult disease. Acta paediatrica 2004, 93, 26–33. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in utero and early-life conditions on adult health and disease. N Engl J Med 2008, 359, 61–73. [Google Scholar] [CrossRef]
- Miguel, P.M.; Pereira, L.O.; Silveira, P.P.; Meaney, M.J. Early environmental influences on the development of children's brain structure and function. Dev Med Child Neurol 2019, 61, 1127–1133. [Google Scholar] [CrossRef]
- Denison, F.C.; Aedla, N.R.; Keag, O.; Hor, K.; Reynolds, R.M.; Milne, A.; Diamond, A.; Royal College of, O. Gynaecologists. Care of Women with Obesity in Pregnancy: Green-top Guideline No. 72. BJOG 2019, 126, e62–e106. [Google Scholar] [CrossRef]
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: mechanisms of short term and long term adverse consequences for mother and child. BMJ 2017, 356, j1. [Google Scholar] [CrossRef]
- Fowden, A.; Camm, E.; Sferruzzi-Perri, A. Effects of Maternal Obesity On Placental Phenotype. Curr Vasc Pharmacol 2020, 10.2174/1570161118666200513115316. [CrossRef]
- Edlow, A.G. Maternal obesity and neurodevelopmental and psychiatric disorders in offspring. Prenat Diagn 2017, 37, 95–110. [Google Scholar] [CrossRef]
- Marchi, J.; Berg, M.; Dencker, A.; Olander, E.K.; Begley, C. Risks associated with obesity in pregnancy, for the mother and baby: a systematic review of reviews. Obes Rev 2015, 16, 621–638. [Google Scholar] [CrossRef]
- Lopez-Taboada, I.; Gonzalez-Pardo, H.; Conejo, N.M. Western Diet: Implications for Brain Function and Behavior. Front Psychol 2020, 11, 564413. [Google Scholar] [CrossRef]
- Menting, M.D.; van de Beek, C.; Mintjens, S.; Wever, K.E.; Korosi, A.; Ozanne, S.E.; Limpens, J.; Roseboom, T.J.; Hooijmans, C.; Painter, R.C. The link between maternal obesity and offspring neurobehavior: A systematic review of animal experiments. Neurosci Biobehav Rev 2019, 98, 107–121. [Google Scholar] [CrossRef]
- Clark, T.D.; Crean, A.J.; Senior, A.M. Obesogenic diets induce anxiety in rodents: A systematic review and meta-analysis. Obes Rev 2022, 23, e13399. [Google Scholar] [CrossRef]
- Rivera, H.M.; Christiansen, K.J.; Sullivan, E.L. The role of maternal obesity in the risk of neuropsychiatric disorders. Frontiers in neuroscience 2015, 9, 194. [Google Scholar] [CrossRef]
- Rakhra, V.; Galappaththy, S.L.; Bulchandani, S.; Cabandugama, P.K. Obesity and the Western Diet: How We Got Here. Mo Med 2020, 117, 536–538. [Google Scholar] [PubMed]
- Speakman, J.R. Use of high-fat diets to study rodent obesity as a model of human obesity. Int J Obes (Lond) 2019, 43, 1491–1492. [Google Scholar] [CrossRef] [PubMed]
- Marcondes, F.K.; Miguel, K.J.; Melo, L.L.; Spadari-Bratfisch, R.C. Estrous cycle influences the response of female rats in the elevated plus-maze test. Physiol Behav 2001, 74, 435–440. [Google Scholar] [CrossRef]
- UK Research and Innovation, https://www.ukri.org/councils/mrc/guidance-for-applicants/policies-and-guidance-for-researchers/sex-in-experimental-design/#:~:text=Use%20of%20both%20sexes%20is,use%20of%20immortalised%20cell%20lines. Accessed 10 October 2023.
- Napso, T.; Lean, S.C.; Lu, M.; Mort, E.J.; Desforges, M.; Moghimi, A.; Bartels, B.; El-Bacha, T.; Fowden, A.L.; Camm, E.J.; et al. Diet-induced maternal obesity impacts feto-placental growth and induces sex-specific alterations in placental morphology, mitochondrial bioenergetics, dynamics, lipid metabolism and oxidative stress in mice. Acta Physiol (Oxf) 2022, 234, e13795. [Google Scholar] [CrossRef]
- Ogilvie, A.; Shapses, S.; Schlussel, Y. Diet Quality in Overweight/Obese Women: Higher Protein Intake During Weight Loss (P08-014-19). Current developments in nutrition 2019, 3, nzz044.P008-014-019. [Google Scholar] [CrossRef]
- Mort, E.J.; Fordington, S.; Heritage, S.; Fowden, A.L.; Jones, S.; Camm, E.J. Age and an obesogenic diet affect mouse behaviour in a sex-dependent manner. Eur J Neurosci 2023, 58, 2451–2468. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nature Protocols 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Moy, S.S.; Nadler, J.J.; Perez, A.; Barbaro, R.P.; Johns, J.M.; Magnuson, T.R.; Piven, J.; Crawley, J.N. Sociability and preference for social novelty in five inbred strains: an approach to assess autistic-like behavior in mice. Genes Brain Behav 2004, 3, 287–302. [Google Scholar] [CrossRef]
- Antunes, M.; Biala, G. The novel object recognition memory: neurobiology, test procedure, and its modifications. Cogn Process 2012, 13, 93–110. [Google Scholar] [CrossRef]
- Samuelsson, A.M.; Matthews, P.A.; Argenton, M.; Christie, M.R.; McConnell, J.M.; Jansen, E.H.; Piersma, A.H.; Ozanne, S.E.; Twinn, D.F.; Remacle, C.; et al. Diet-induced obesity in female mice leads to offspring hyperphagia, adiposity, hypertension, and insulin resistance: a novel murine model of developmental programming. Hypertension 2008, 51, 383–392. [Google Scholar] [CrossRef]
- Fernandez-Twinn, D.S.; Alfaradhi, M.Z.; Martin-Gronert, M.S.; Duque-Guimaraes, D.E.; Piekarz, A.; Ferland-McCollough, D.; Bushell, M.; Ozanne, S.E. Downregulation of IRS-1 in adipose tissue of offspring of obese mice is programmed cell-autonomously through post-transcriptional mechanisms. Mol Metab 2014, 3, 325–333. [Google Scholar] [CrossRef]
- John, L.M.; Petersen, N.; Gerstenberg, M.K.; Torz, L.; Pedersen, K.; Christoffersen, B.O.; Kuhre, R.E. Housing-temperature reveals energy intake counter-balances energy expenditure in normal-weight, but not diet-induced obese, male mice. Commun Biol 2022, 5, 946. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, H.R. Homeostatic and non-homeostatic pathways involved in the control of food intake and energy balance. Obesity (Silver Spring) 2006, 14 Suppl 5, 197S–200S. [Google Scholar] [CrossRef] [PubMed]
- Musial, B.; Vaughan, O.R.; Fernandez-Twinn, D.S.; Voshol, P.; Ozanne, S.E.; Fowden, A.L.; Sferruzzi-Perri, A.N. A Western-style obesogenic diet alters maternal metabolic physiology with consequences for fetal nutrient acquisition in mice. J Physiol 2017, 595, 4875–4892. [Google Scholar] [CrossRef]
- Lewandowska, M. Maternal Obesity and Risk of Low Birth Weight, Fetal Growth Restriction, and Macrosomia: Multiple Analyses. Nutrients 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Coan, P.M.; Vaughan, O.R.; McCarthy, J.; Mactier, C.; Burton, G.J.; Constancia, M.; Fowden, A.L. Dietary composition programmes placental phenotype in mice. J Physiol 2011, 589, 3659–3670. [Google Scholar] [CrossRef] [PubMed]
- Drake, A.J.; Reynolds, R.M. Impact of maternal obesity on offspring obesity and cardiometabolic disease risk. Reproduction 2010, 140, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Kirk, S.L.; Samuelsson, A.M.; Argenton, M.; Dhonye, H.; Kalamatianos, T.; Poston, L.; Taylor, P.D.; Coen, C.W. Maternal obesity induced by diet in rats permanently influences central processes regulating food intake in offspring. PLoS One 2009, 4, e5870. [Google Scholar] [CrossRef]
- Buckley, A.J.; Keseru, B.; Briody, J.; Thompson, M.; Ozanne, S.E.; Thompson, C.H. Altered body composition and metabolism in the male offspring of high fat-fed rats. Metabolism 2005, 54, 500–507. [Google Scholar] [CrossRef]
- Long, N.M.; George, L.A.; Uthlaut, A.B.; Smith, D.T.; Nijland, M.J.; Nathanielsz, P.W.; Ford, S.P. Maternal obesity and increased nutrient intake before and during gestation in the ewe results in altered growth, adiposity, and glucose tolerance in adult offspring. J Anim Sci 2010, 88, 3546–3553. [Google Scholar] [CrossRef]
- Sasaki, A.; de Vega, W.C.; St-Cyr, S.; Pan, P.; McGowan, P.O. Perinatal high fat diet alters glucocorticoid signaling and anxiety behavior in adulthood. Neuroscience 2013, 240, 1–12. [Google Scholar] [CrossRef]
- Abuaish, S.; Spinieli, R.L.; McGowan, P.O. Perinatal high fat diet induces early activation of endocrine stress responsivity and anxiety-like behavior in neonates. Psychoneuroendocrinology 2018, 98, 11–21. [Google Scholar] [CrossRef]
- Balsevich, G.; Baumann, V.; Uribe, A.; Chen, A.; Schmidt, M.V. Prenatal Exposure to Maternal Obesity Alters Anxiety and Stress Coping Behaviors in Aged Mice. Neuroendocrinology 2016, 103, 354–368. [Google Scholar] [CrossRef]
- Sasaki, A.; de Vega, W.; Sivanathan, S.; St-Cyr, S.; McGowan, P.O. Maternal high-fat diet alters anxiety behavior and glucocorticoid signaling in adolescent offspring. Neuroscience 2014, 272, 92–101. [Google Scholar] [CrossRef]
- Janthakhin, Y.; Rincel, M.; Costa, A.M.; Darnaudery, M.; Ferreira, G. Maternal high-fat diet leads to hippocampal and amygdala dendritic remodeling in adult male offspring. Psychoneuroendocrinology 2017, 83, 49–57. [Google Scholar] [CrossRef]
- Farhat, T.; Iannotti, R.J.; Simons-Morton, B.G. Overweight, obesity, youth, and health-risk behaviors. Am J Prev Med 2010, 38, 258–267. [Google Scholar] [CrossRef]
- Zanini, P.; Arbo, B.D.; Niches, G.; Czarnabay, D.; Benetti, F.; Ribeiro, M.F.; Cecconello, A.L. Diet-induced obesity alters memory consolidation in female rats. Physiol Behav 2017, 180, 91–97. [Google Scholar] [CrossRef]
- Page, K.C.; Jones, E.K.; Anday, E.K. Maternal and postweaning high-fat diets disturb hippocampal gene expression, learning, and memory function. Am J Physiol Regul Integr Comp Physiol 2014, 306, R527–537. [Google Scholar] [CrossRef]
- Cordner, Z.A.; Khambadkone, S.G.; Boersma, G.J.; Song, L.; Summers, T.N.; Moran, T.H.; Tamashiro, K.L.K. Maternal high-fat diet results in cognitive impairment and hippocampal gene expression changes in rat offspring. Exp Neurol 2019, 318, 92–100. [Google Scholar] [CrossRef]
- Mucellini, A.B.; Laureano, D.P.; Silveira, P.P.; Sanvitto, G.L. Maternal and post-natal obesity alters long-term memory and hippocampal molecular signaling of male rat. Brain Res 2019, 1708, 138–145. [Google Scholar] [CrossRef]
- Zieba, J.; Uddin, G.M.; Youngson, N.A.; Karl, T.; Morris, M.J. Long-term behavioural effects of maternal obesity in C57BL/6J mice. Physiol Behav 2019, 199, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, X.; Xia, B.; Jin, X.; Zou, Q.; Zeng, Z.; Zhao, W.; Yan, S.; Li, L.; Yuan, S.; et al. High-fiber diet mitigates maternal obesity-induced cognitive and social dysfunction in the offspring via gut-brain axis. Cell Metab 2021, 33, 923–938.e926. [Google Scholar] [CrossRef] [PubMed]
- Raygada, M.; Cho, E.; Hilakivi-Clarke, L. High maternal intake of polyunsaturated fatty acids during pregnancy in mice alters offsprings' aggressive behavior, immobility in the swim test, locomotor activity and brain protein kinase C activity. J Nutr 1998, 128, 2505–2511. [Google Scholar] [CrossRef] [PubMed]
- Ingvorsen, C.; Lelliott, C.J.; Brix, S.; Hellgren, L.I. Effects of maternal high-fat/high sucrose diet on hepatic lipid metabolism in rat offspring. Clin Exp Pharmacol Physiol 2020, 48, 86–95. [Google Scholar] [CrossRef]
- Irving, A.; Harvey, J. Regulation of hippocampal synaptic function by the metabolic hormone leptin: Implications for health and disease. Prog Lipid Res 2021, 82, 101098. [Google Scholar] [CrossRef]
Figure 1.
An obesogenic diet alters nutrient intake in pre-pregnant, pregnant and lactating female mice. (A) Food intake (gram of food per day) in female mice fed either a control diet (from n=5 cages, open columns) or a high fat, high sugar (HFHS, from n=5 cages, filled columns) diet for 6 weeks before pregnancy; (B) Kilocalorie intake per day in the same groups of female mice; (C) Protein intake (gram of protein per day) in the same group of female mice; (D-F) Food intake, kcal intake and protein intake in female mice fed either a control diet (from n=7 cages) or a HFHS (from n=6 cages) diet over each week while pregnant. Note that the x-axis scale is non-linear; (G-I) Food intake, kcal intake and protein intake in female mice fed either a control diet (from n=7 cages) or a HFHS (from n=5 cages) diet on specific post-partum (PP) days during lactation. Note that the x-axis scale is non-linear. Two-way ANOVA result is given above each graph (interaction was not significantly different in all cases); post hoc test of diet effect indicated above bars, *p<0.05, **p<0.01, ***p<0.001.
Figure 1.
An obesogenic diet alters nutrient intake in pre-pregnant, pregnant and lactating female mice. (A) Food intake (gram of food per day) in female mice fed either a control diet (from n=5 cages, open columns) or a high fat, high sugar (HFHS, from n=5 cages, filled columns) diet for 6 weeks before pregnancy; (B) Kilocalorie intake per day in the same groups of female mice; (C) Protein intake (gram of protein per day) in the same group of female mice; (D-F) Food intake, kcal intake and protein intake in female mice fed either a control diet (from n=7 cages) or a HFHS (from n=6 cages) diet over each week while pregnant. Note that the x-axis scale is non-linear; (G-I) Food intake, kcal intake and protein intake in female mice fed either a control diet (from n=7 cages) or a HFHS (from n=5 cages) diet on specific post-partum (PP) days during lactation. Note that the x-axis scale is non-linear. Two-way ANOVA result is given above each graph (interaction was not significantly different in all cases); post hoc test of diet effect indicated above bars, *p<0.05, **p<0.01, ***p<0.001.
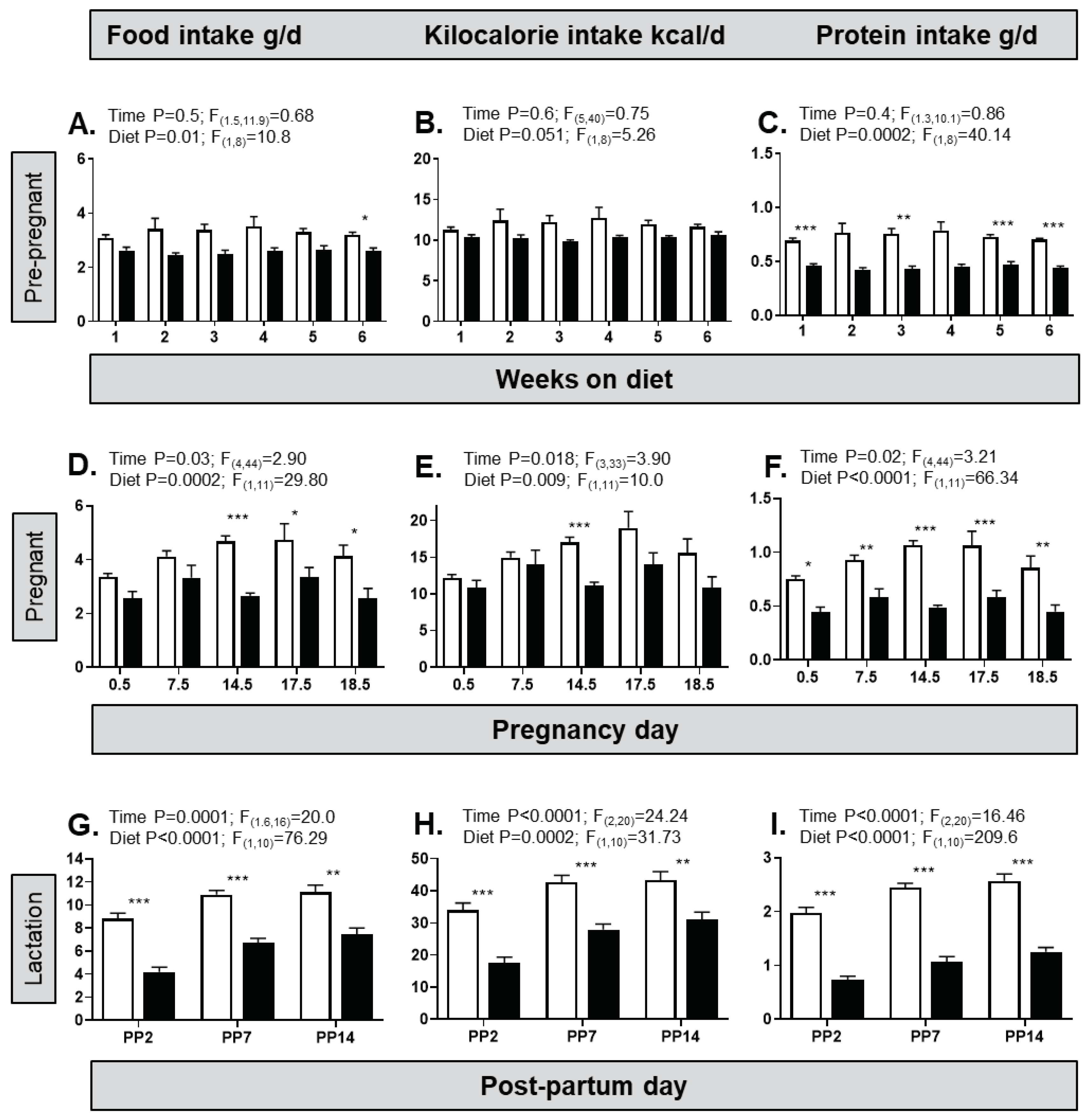
Figure 2.
An obesogenic diet affects growth and adiposity in female mice. (A-C) Total body mass (g) of female mice on either a control (open symbols) or high fat, high sugar (HFHS, closed symbols) diet, either pre-pregnant (A; Control n=45, HFHS n=55), pregnant (B; Control n=35, HFHS n=39) or lactating (C; Control n=27, HFHS n=17). Two-way ANOVA (A) or mixed effects analysis (B, C) results given above graphs; post hoc test of diet effect, *p<0.05, **p<0.01, ***p<0.001; (D) Example DEXA scans of female mice aged 14 weeks on the control diet (left) or HFHS diet (right); (E) Example DEXA scans of female mice aged ~17 weeks on the control diet (left) or HFHS diet (right); (F) Example DEXA scans of female mice aged ~20 weeks on the control diet (left) or HFHS diet (right); (G-I) Total fat mass (g) in pre-pregnant (G; n=7; **p=0.0037, t-test), pregnant (H; n=9; **p=0.0012, t-test) and lactating (I; n=7-9; ***p=0.0002, Mann Whitney) female mice on a control or HFHS diet; (J-L). Total lean mass (g) in pre-pregnant (J; n=7; *p=0.035, t-test), pregnant (K; n=9; ***p<0.0001, t-test) and lactating (L; n=7-9; **p=0.0023, t-test) female mice on a control or HFHS diet.
Figure 2.
An obesogenic diet affects growth and adiposity in female mice. (A-C) Total body mass (g) of female mice on either a control (open symbols) or high fat, high sugar (HFHS, closed symbols) diet, either pre-pregnant (A; Control n=45, HFHS n=55), pregnant (B; Control n=35, HFHS n=39) or lactating (C; Control n=27, HFHS n=17). Two-way ANOVA (A) or mixed effects analysis (B, C) results given above graphs; post hoc test of diet effect, *p<0.05, **p<0.01, ***p<0.001; (D) Example DEXA scans of female mice aged 14 weeks on the control diet (left) or HFHS diet (right); (E) Example DEXA scans of female mice aged ~17 weeks on the control diet (left) or HFHS diet (right); (F) Example DEXA scans of female mice aged ~20 weeks on the control diet (left) or HFHS diet (right); (G-I) Total fat mass (g) in pre-pregnant (G; n=7; **p=0.0037, t-test), pregnant (H; n=9; **p=0.0012, t-test) and lactating (I; n=7-9; ***p=0.0002, Mann Whitney) female mice on a control or HFHS diet; (J-L). Total lean mass (g) in pre-pregnant (J; n=7; *p=0.035, t-test), pregnant (K; n=9; ***p<0.0001, t-test) and lactating (L; n=7-9; **p=0.0023, t-test) female mice on a control or HFHS diet.
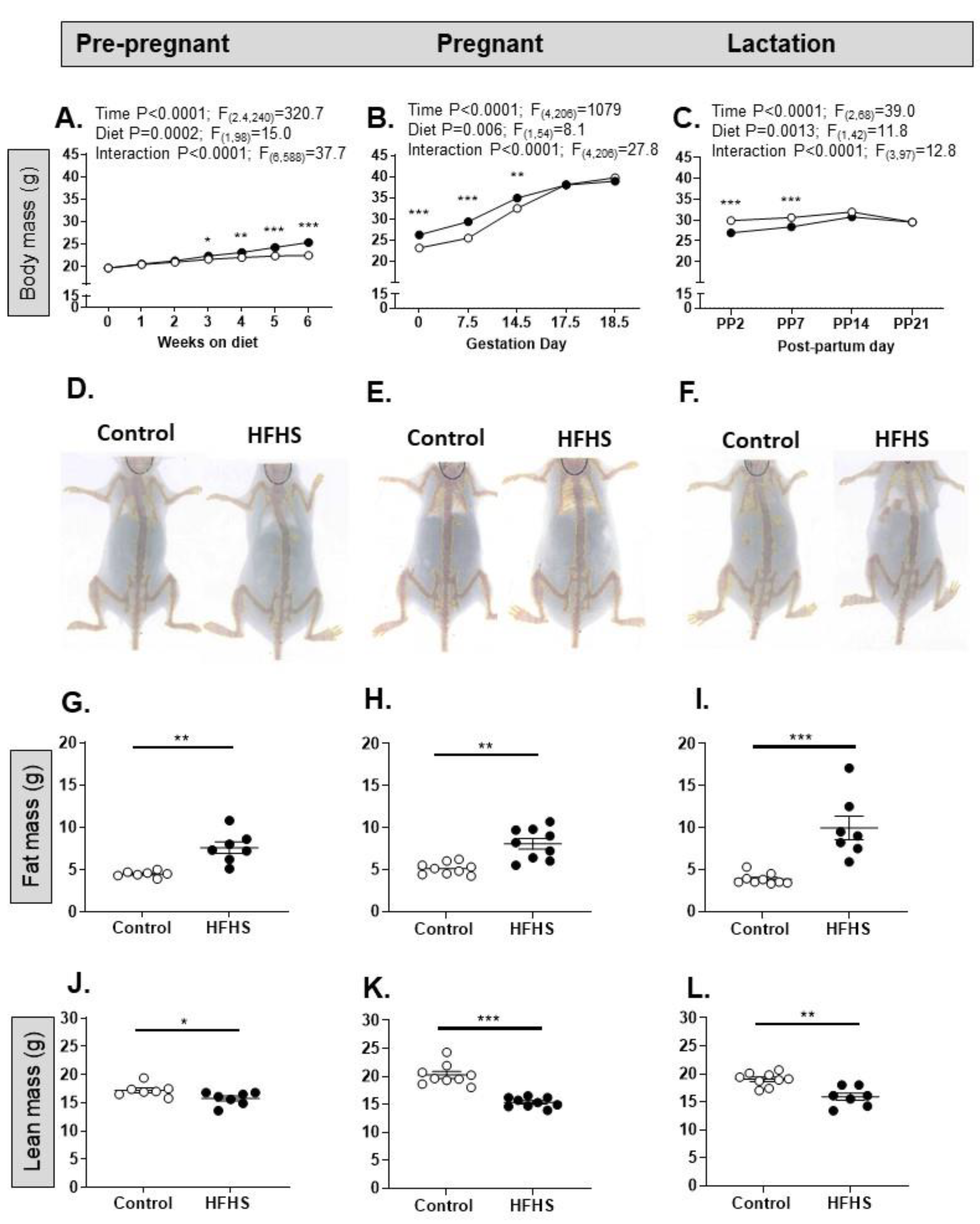
Figure 3.
Pre-weaning growth in offspring of dams on control or high fat, high sugar (HFHS) diets. (A) Total mass during postnatal development (postnatal day 2 to 21) in male offspring of mothers on a control (open symbols, n=19) or HFHS (closed symbols, n=20) diet; (B) Female offspring from mothers on a control (n=19) or HFHS (n=20) diet. Two-way ANOVA results given above graphs; post hoc test of diet effect, *p<0.05, **p<0.01; (C) Fractional growth rate in male offspring of mothers on a control (open symbols and bars, n=19) or HFHS (closed symbols and bars, n=20) diet; (D) Fractional growth rate in female offspring (control, open symbols and bars, n=19; HFHS, closed symbols and bars, n=20). Two-way ANOVA results given above graphs; post hoc test of diet effects, *p<0.05, **p<0.01, ***p<0.001.
Figure 3.
Pre-weaning growth in offspring of dams on control or high fat, high sugar (HFHS) diets. (A) Total mass during postnatal development (postnatal day 2 to 21) in male offspring of mothers on a control (open symbols, n=19) or HFHS (closed symbols, n=20) diet; (B) Female offspring from mothers on a control (n=19) or HFHS (n=20) diet. Two-way ANOVA results given above graphs; post hoc test of diet effect, *p<0.05, **p<0.01; (C) Fractional growth rate in male offspring of mothers on a control (open symbols and bars, n=19) or HFHS (closed symbols and bars, n=20) diet; (D) Fractional growth rate in female offspring (control, open symbols and bars, n=19; HFHS, closed symbols and bars, n=20). Two-way ANOVA results given above graphs; post hoc test of diet effects, *p<0.05, **p<0.01, ***p<0.001.
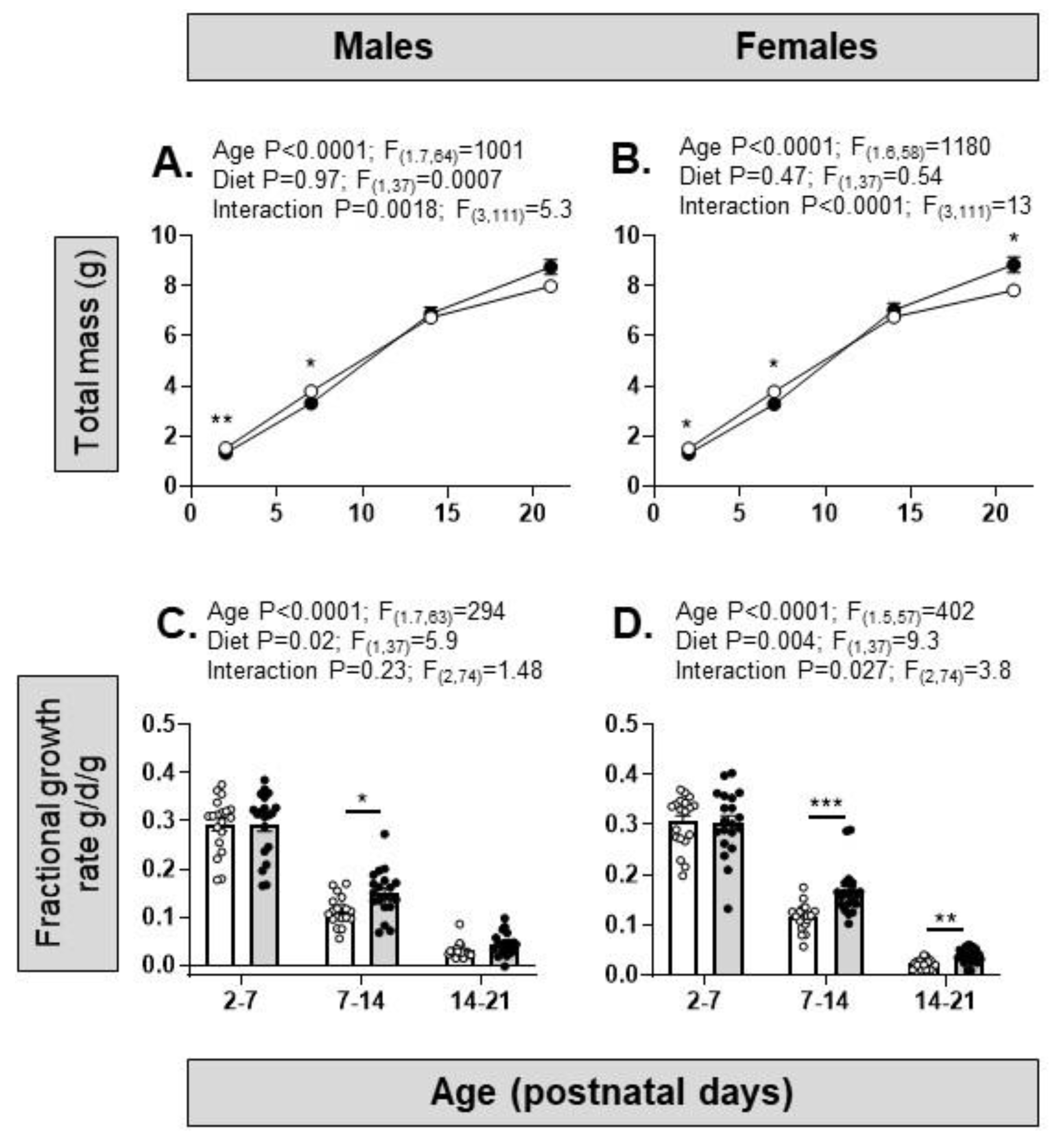
Figure 4.
Post-weaning food intake of offspring from postnatal day 21 to 91. (A) Food intake (gram of food per day) in male mice from mothers on a control (from n=2-5 cages) or high fat, high sugar (HFHS, from n=6-7 cages) diet measured between postnatal days 21 to 91; (B) Food intake (gram of food per day) in female mice from mothers on a control (from n=3-5 cages) or HFHS diet (from n=5 cages) measured between postnatal days 21 to 91; (C, D) Kilocalorie intake (kcal per day) in the same groups of male and female mice from mothers on a control or HFHS diet; (E, F) Protein intake (gram of protein per day) in in the same groups of male and female mice from mothers on a control or HFHS diet. Mixed effects analysis results shown above graphs; post hoc test of diet effect at specific ages, *p<0.05.
Figure 4.
Post-weaning food intake of offspring from postnatal day 21 to 91. (A) Food intake (gram of food per day) in male mice from mothers on a control (from n=2-5 cages) or high fat, high sugar (HFHS, from n=6-7 cages) diet measured between postnatal days 21 to 91; (B) Food intake (gram of food per day) in female mice from mothers on a control (from n=3-5 cages) or HFHS diet (from n=5 cages) measured between postnatal days 21 to 91; (C, D) Kilocalorie intake (kcal per day) in the same groups of male and female mice from mothers on a control or HFHS diet; (E, F) Protein intake (gram of protein per day) in in the same groups of male and female mice from mothers on a control or HFHS diet. Mixed effects analysis results shown above graphs; post hoc test of diet effect at specific ages, *p<0.05.
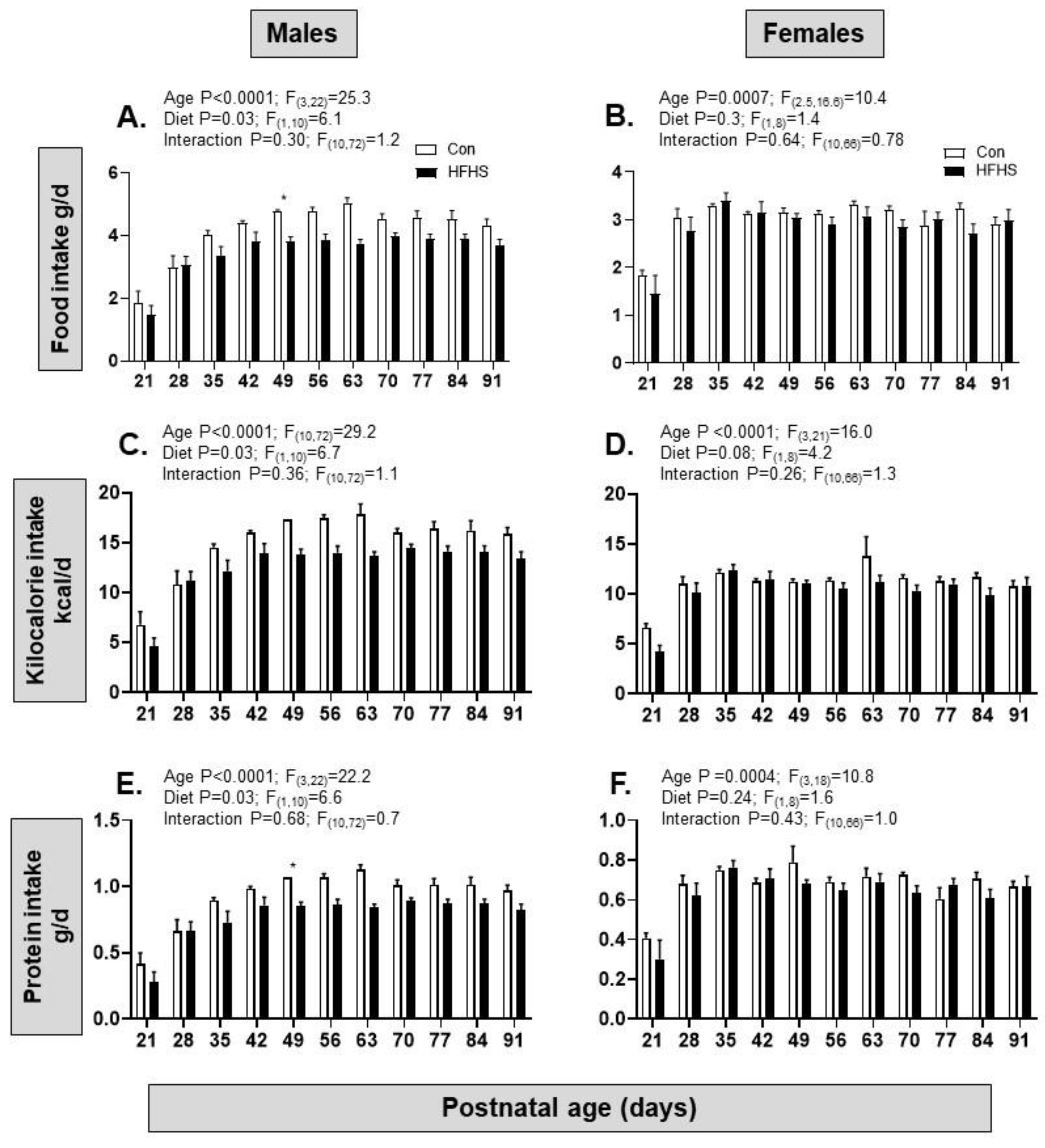
Figure 5.
Post-weaning growth and fat deposition in offspring of mothers on control or high fat, high sugar (HFHS) diets. (A, B) Body mass (g) in male and female offspring from mothers on a control (n=18-19) or HFHS (n=18) diet. Mixed effects analysis (A and B) results are given above graph; post hoc tests show no diet effects at specific ages; (C, D) Fractional growth rate with respect to weekly postnatal periods in male and female offspring from mothers on a control (n=18, open symbols) or HFHS (n=17, closed symbols) diet. Mixed effects analysis (C and D) results given above graph; post hoc tests in males (P21-28: *p<0.01); no effect of diet in females; (E, F) Total fat mass and lean mass (g) in male offspring from mothers on a control (n=8) or HFHS (n=7) diet; (G, H) Fat mass and lean mass (g) in female offspring from mothers on a control (n=6) or HFHS (n=9) diet (*p<0.05, unpaired t-test).
Figure 5.
Post-weaning growth and fat deposition in offspring of mothers on control or high fat, high sugar (HFHS) diets. (A, B) Body mass (g) in male and female offspring from mothers on a control (n=18-19) or HFHS (n=18) diet. Mixed effects analysis (A and B) results are given above graph; post hoc tests show no diet effects at specific ages; (C, D) Fractional growth rate with respect to weekly postnatal periods in male and female offspring from mothers on a control (n=18, open symbols) or HFHS (n=17, closed symbols) diet. Mixed effects analysis (C and D) results given above graph; post hoc tests in males (P21-28: *p<0.01); no effect of diet in females; (E, F) Total fat mass and lean mass (g) in male offspring from mothers on a control (n=8) or HFHS (n=7) diet; (G, H) Fat mass and lean mass (g) in female offspring from mothers on a control (n=6) or HFHS (n=9) diet (*p<0.05, unpaired t-test).
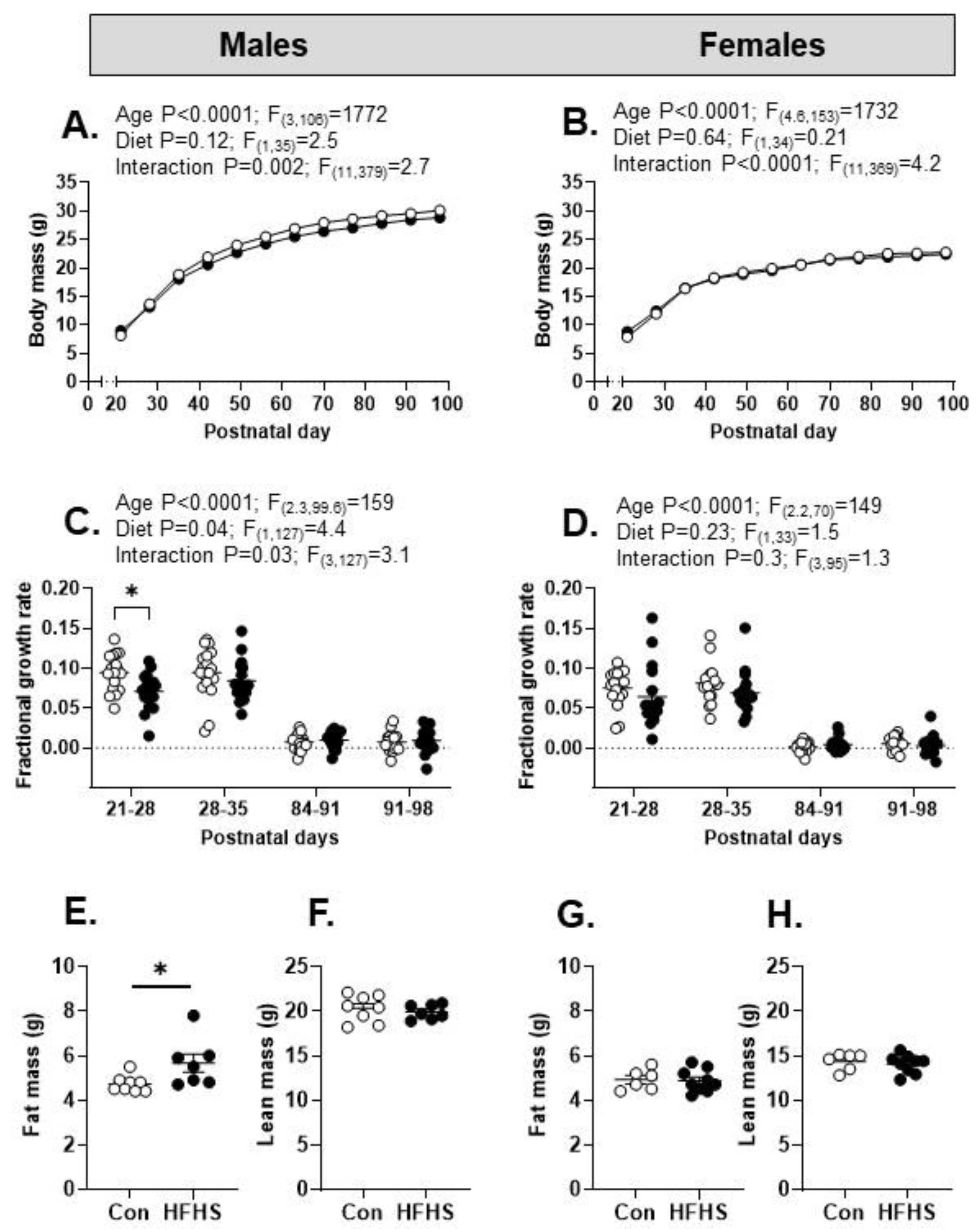
Figure 6.
Pre-weaning Effect of a maternal obesogenic diet on adult offspring behaviour in the Elevated Plus Maze (EPM). (A) The percentage time spent in the open arm of the EPM by male mice from mothers on a control (n=16) or high fat, high sugar (HFHS; n=17) diet (p=0.073, t-test); (B) Percentage time in the open arm for female mice from mothers on a control (n=14) or HFHS diet (n=17; ***p=0.0006, t-test); (C) Number of full entries into the open arm by male mice from mothers on a control (n=15) or HFHS diet (n=17; **p=0.0065, t-test); (D) Full entries into the open arm by female mice (p=0.15, Mann Whitney); (E, F) Number of full entries into the closed arm by male (p=0.91, t-test) and female (p=0.96, t-test) mice from mothers on a control or HFHS diet; (G, H) Number of explorations to the end of the open arm by male (*p=0.022, t-test) and female (**p=0.0015, t-test) mice from mothers on a control or HFHS diet.
Figure 6.
Pre-weaning Effect of a maternal obesogenic diet on adult offspring behaviour in the Elevated Plus Maze (EPM). (A) The percentage time spent in the open arm of the EPM by male mice from mothers on a control (n=16) or high fat, high sugar (HFHS; n=17) diet (p=0.073, t-test); (B) Percentage time in the open arm for female mice from mothers on a control (n=14) or HFHS diet (n=17; ***p=0.0006, t-test); (C) Number of full entries into the open arm by male mice from mothers on a control (n=15) or HFHS diet (n=17; **p=0.0065, t-test); (D) Full entries into the open arm by female mice (p=0.15, Mann Whitney); (E, F) Number of full entries into the closed arm by male (p=0.91, t-test) and female (p=0.96, t-test) mice from mothers on a control or HFHS diet; (G, H) Number of explorations to the end of the open arm by male (*p=0.022, t-test) and female (**p=0.0015, t-test) mice from mothers on a control or HFHS diet.
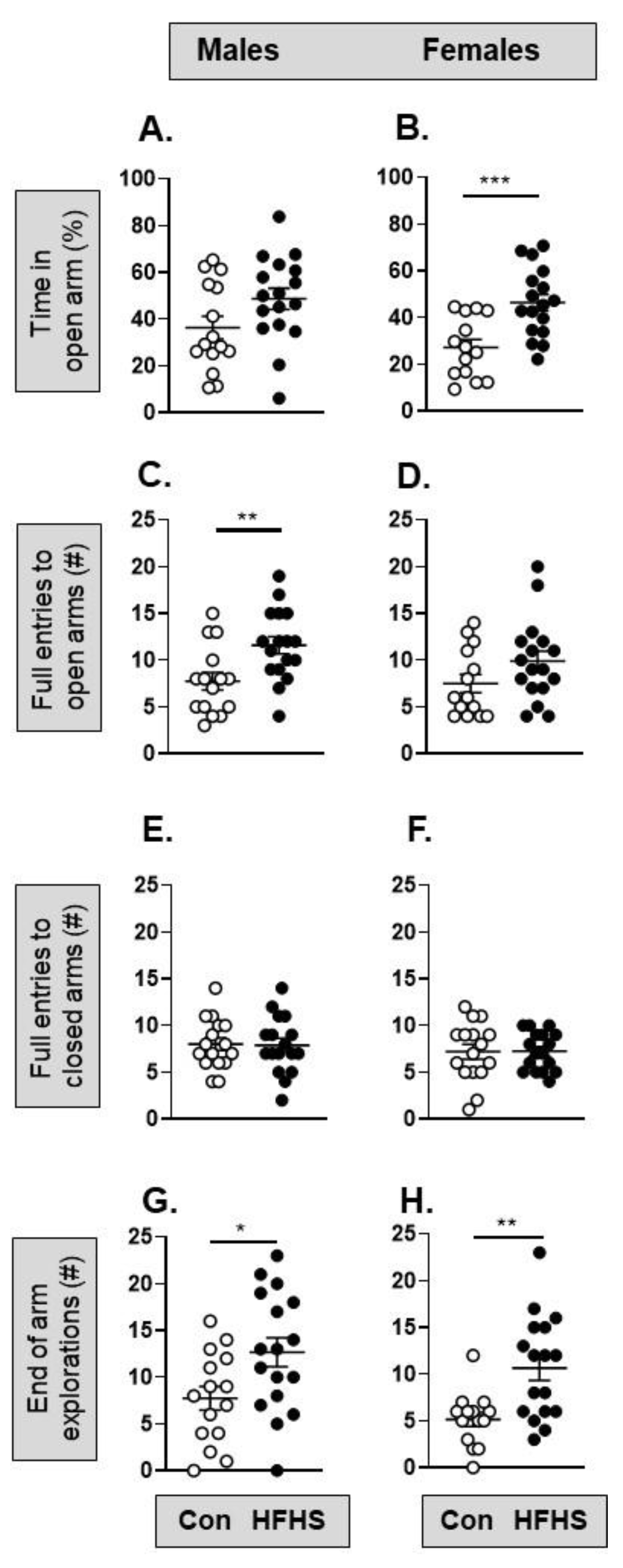
Figure 7.
Effect of a maternal obesogenic diet on adult offspring behaviour in the novel object recognition task. (A-D) Acquisition phase: (A) The time sniffing two identical objects by male mice from mothers on a control (n=16) or high fat, high sugar (HFHS; n=17) diet (p=0.136, t-test); (B) The time sniffing two identical objects by female mice from mothers on a control (n=16) or high fat, high sugar (HFHS; n=16) diet (p=0.78, t-test); (C) Time taken to reach 25s of sniffing the two identical objects by male mice from mothers on a control (n=16) or HFHS diet (n=17; p=0.076, t-test); (D) Time taken to reach 25s of sniffing the two identical objects by female mice from mothers on a control (n=16) or HFHS diet (n=16; p=0.356, Mann Whitney); (E-H) Retention phase: (E, F) Discrimination index score of male (***p=0.0002, t-test; n=16-17) and female (p=0.47, t-test; n=12-16) mice from mothers on a control or HFHS diet in the first minute of object exploration; (G, H) Discrimination index score by male (***p<0.0001, t-test; n=16-17) and female (p=0.104, t-test; n=12-16) mice from mothers on a control or HFHS diet after 5 minutes of object exploration.
Figure 7.
Effect of a maternal obesogenic diet on adult offspring behaviour in the novel object recognition task. (A-D) Acquisition phase: (A) The time sniffing two identical objects by male mice from mothers on a control (n=16) or high fat, high sugar (HFHS; n=17) diet (p=0.136, t-test); (B) The time sniffing two identical objects by female mice from mothers on a control (n=16) or high fat, high sugar (HFHS; n=16) diet (p=0.78, t-test); (C) Time taken to reach 25s of sniffing the two identical objects by male mice from mothers on a control (n=16) or HFHS diet (n=17; p=0.076, t-test); (D) Time taken to reach 25s of sniffing the two identical objects by female mice from mothers on a control (n=16) or HFHS diet (n=16; p=0.356, Mann Whitney); (E-H) Retention phase: (E, F) Discrimination index score of male (***p=0.0002, t-test; n=16-17) and female (p=0.47, t-test; n=12-16) mice from mothers on a control or HFHS diet in the first minute of object exploration; (G, H) Discrimination index score by male (***p<0.0001, t-test; n=16-17) and female (p=0.104, t-test; n=12-16) mice from mothers on a control or HFHS diet after 5 minutes of object exploration.
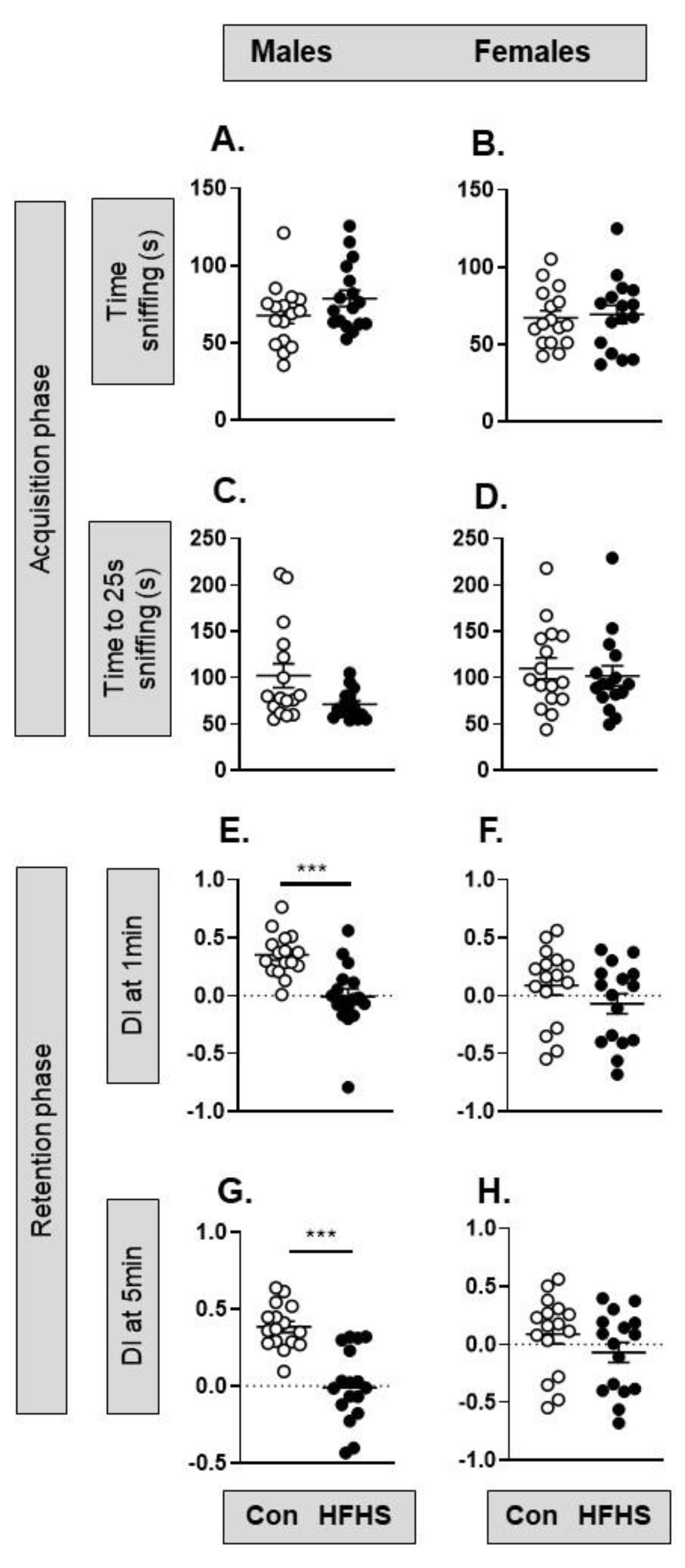

Table 1.
Post-mortem organ weights and fat deposition in pre-pregnant, pregnant and lactating female mice, and of their female and male offspring fed a control diet post weaning. ***p<0.01, **p<0.02, *p<0.05 t-test or Mann Whitney test. NA = Not available.
Table 1.
Post-mortem organ weights and fat deposition in pre-pregnant, pregnant and lactating female mice, and of their female and male offspring fed a control diet post weaning. ***p<0.01, **p<0.02, *p<0.05 t-test or Mann Whitney test. NA = Not available.
| Dams | Offspring | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pre-pregnant | Pregnant | Lactating | Females | Males | ||||||
| Control(n = 7) | HFHS (n = 7) |
Control(n = 9) | HFHS (n = 9) |
Control (n = 14-26) |
HFHS (n = 10-18) |
Control(n = 9-16) | HFHS (n = 13-16) |
Control (n = 11-16) |
HFHS (n = 13) |
|
| Total body weight (g) | 22.4 ± 0.2 | 25.3 ± 0.3*** | 41.4 ± 1.2 | 39.3 ± 0.6 | 29.5 ± 0.4 | 29.5 ± 0.6 | 22.8 ± 0.2 | 22.4 ± 0.3 | 29.9 ± 0.4 | 28.8 ± 0.7 |
| Brain (mg) | 460 ± 7 | 460 ± 4 | 463 ± 8 | 442 ± 8 | 454 ± 6 | 454 ± 19 | 450 ± 2 | 450 ± 4 | 450 ± 4 | 440 ± 7 |
| Liver (g) | 1.20 ± 0.04 | 1.10 ± 0.04 | 1.9 ± 0.07 | 2.30 ± 0.10** | 2.10 ± 0.05 | 2.00 ± 0.08 | 1.11 ± 0.04 | 1.04 ± 0.03 | 1.49 ± 0.06 | 1.36 ± 0.06 |
| Heart (mg) | 120 ± 7 | 110 ± 9 | 152 ± 12 | 146 ± 4 | 0.21 ± 0.01 | 0.21 ± 0.03 | 120 ± 4 | 130 ± 3 | 170 ± 5 | 170 ± 8 |
| Adrenal (mg) | 7.8 ± 0.5 | 9.7 ± 0.5* | 6.8 ± 0.7 | 9.0 ± 1.8 | 7.7 ± 0.4 | 7.4 ± 0.6 | 6.3 ± 0.3 | 6.0 ± 0.3 | 5.0 ± 0.3 | 3.8 ± 0.2*** |
| Gonadal fat (mg) | NA | NA | 294 ± 25 | 484 ± 71** | 341 ± 25 | 1100 ± 142*** | 500 ± 34 | 514 ± 45 | 431 ± 31 | 579 ± 53* |
| Retroperitoneal fat (mg) | 37 ± 3 | 120 ± 21*** | 67 ± 5 | 156 ± 20*** | 55 ± 5 | 208 ± 29*** | 76 ± 4 | 72 ± 6 | 82 ± 8 | 150 ± 26*** |
| Perirenal fat (mg) | 46 ± 5 | 110 ± 27*** | 135 ± 18 | 216 ± 24** | 132 ± 7 | 300 ± 41*** | 109 ± 10 | 143 ± 8** | 49 ± 5 | 80 ± 6*** |
Table 2.
Summary of the effects of a maternal HFHS diet on dietary intake, body mass and composition, and on behaviour and cognitive function of male and female offspring. FGR, fractional growth rate; PN, postnatal day.
Table 2.
Summary of the effects of a maternal HFHS diet on dietary intake, body mass and composition, and on behaviour and cognitive function of male and female offspring. FGR, fractional growth rate; PN, postnatal day.
| Male offspring | Female offspring | |
|---|---|---|
| Dietary intake | ↓ food intake ↓ calorie intake ↓ protein intake |
↔ food intake ↔ calorie intake ↔ protein intake |
| Body mass and growth rate | ↓ PN2-7 ↑ FGR PN7-14 ↓ FGR PN21-28 |
↓ PN2-7 ↑ FGR PN7-21 ↔ FGR PN21-28 |
| Body composition | ↑ fat mass ↔ lean mass ↑ gonadal mass ↑ retroperitoneal mass ↑ perirenal fat mass |
↔ fat mass ↔ lean mass ↔ gonadal mass ↔ retroperitoneal mass ↑ perirenal fat mass |
| Locomotor activity | ↔ | ↔ |
| Anxiety-related behaviours | ↑ open arm exploration ↔ % time in open arm ↑ full entries into open arm |
↑ open arm exploration ↑ % time in open arm ↔ full entries into open arm |
| Social behaviour | ↔ | ↔ |
| Cognition | ↓ object recognition memory | ↔ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



