
Submitted:
11 October 2023
Posted:
13 October 2023
You are already at the latest version
Abstract
Bacillus Calmette-Guérin (BCG) is the only widely used prophylactic tuberculosis (TB) vaccine that can prevent severe TB in infants. However, it provides poor protection in adults, and therefore, there in ongoing research on new TB vaccines and immunization strategies with more durable immune effects. The recombinant BCG and BCG prime-protein booster are two important vaccine strategies that have recently been developed based on BCG and could improve immune responses. In this study, three immune strategies based on four protective antigens, namely, ESAT-6, CFP-10, nPPE18, and nPstS1, were applied to construct recombinant rBCG-EPCP009, EPCP009 subunit protein, and BCG prime-EPCP009 booster vaccine candidates. The short- and long-term immune effects after vaccination in Balb/c mice were evaluated based on humoral immunity, cellular immunity, and the ability of spleen cells to inhibit in vitro mycobacterial growth. At 8 and 12 weeks after the initial immunization, splenocytes from mice inoculated with the BCG prime-EPCP009 protein booster secreted higher levels of PPD- and EPCP009-specific IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, and IL-12 and had a higher IFN-γ+CD4+ TEM:IL-2+CD8+ TCM cell ratio than splenocytes from mice inoculated with the rBCG-EPCP009 and EPCP009 proteins. In addition, the EPCPE009-specific IgG2a/IgG1 ratio was slightly higher in the BCG prime-EPCP009 protein booster group than in the other two groups. The in vitro mycobacterial inhibition assay showed that the splenocytes of mice from the BCG prime-EPCP009 protein booster group exhibited stronger inhibition of Mycobacterium tuberculosis (M. tuberculosis)growth than the splenocytes of mice from the other two groups. These results indicate that the BCG prime-EPCP009 protein booster exhibited superior immunogenicity and M. tuberculosis growth inhibition to the parental BCG, rBCG-EPCP009, and EPCP009 proteins under in vitro conditions. Thus, the BCG prime-EPCP009 protein booster may be important for the development of a more effective adult TB vaccine.
Keywords:
prime-protein booster
; recombinant BCG
; vaccine
; tuberculosis
Introduction
Tuberculosis (TB) is a chronic infectious disease that is mainly caused by M. tuberculosis. According to the World Health Organization, 10.6 million new TB cases were reported worldwide in 2021, and this is 4.5% more than the 10.1 million new cases reported in 2020[1]. In recent years, the spread of multidrug-resistant M. tuberculosis strains has made TB an even bigger threat and challenge to global public health, and has made the development of a more effective vaccine an urgent need [2]. As of September 2022, there were 16 TB vaccine candidates in clinical trials, and they are classified as recombinant Bacillus Calmette-Guérin (BCG) vaccines, M. tuberculosis live attenuated vaccines, protein/adjuvant vaccines, virus vector vaccines, and inactivated M. tuberculosis vaccines [3]. The components of the vaccines are closely related to their immune strategies, and currently, there are two promising strategies for the development of TB vaccines: ① modified recombinant BCG vaccines, which are mainly administered to infants and is a replacement for BCG; ② subunit protein vaccines, which are mainly administered to adults to enhance the protective effect of BCG after primary immunization [4].
As the only vaccine that can prevent TB at present, BCG has been in use since 1921, with a global vaccination rate of more than 90% in infants [5]. It can prevent tuberculous meningitis, miliary tuberculosis, and other serious forms of tuberculosis, and plays an important role in the prevention of TB in infants in countries with a high TB burden [6]. However, the protective effect of BCG decreases from adolescence, and it has almost no protective effect on adults [7, 8]. Further, the effectiveness of revaccination of adults with BCG is controversial [7, 8]. The protective effect of BCG in adults may be reduced because BCG loses some important immunodominant antigen genes during the long-term passage of the strain. These deletion fragments are referred to as the region of differences (RD) (in comparison to M. tuberculosis) and include esxB and esxA, among other fragments [9]. The construction of recombinant BCG vaccines is based on molecular biology technology and involves knocking out the original gene or inducing overexpression of an immunodominant antigen gene based on the parental BCG gene [10]. This strategy overcomes the deficiency in the immune effect and duration of effect of the original BCG (thus extending its protective effect on adults), while retaining the advantages of BCG in terms of stimulating immune protection (especially in children) [10]. One such recombinant vaccine is the VPM1002 vaccine, in which the urease C gene is deleted and transferred into the Hly gene of Listeria monocytogenes to reduce the pH of phagocytic lysosomes. This improves the phagocytosing and antigen cross-presenting abilities of antigen-presenting cells. VPM1002 was proved to induce stronger immune response and protection against M. tuberculosis and was expected to be a viable alternative to BCG [11]. As a result, the construction of safe and effective recombinant BCG vaccines has become a hot topic in the research on TB vaccines.
The immune efficacy of BCG, which is a live attenuated vaccine, is mainly dependent on its effective components and its ability to replicate and persist in the body [12]. Another reason for the reduced protective effect of BCG could be pre-sensitization to non-tuberculosis mycobacterium and cross-immunization with BCG, which may inhibit BCG replication in humans and lead to reduced colonization ability [12]. Unlike BCG and other live vaccines, subunit protein vaccines are non-replicative, and their immune effect is mainly dependent on the antigenic component and adjuvant and is not affected by pre-sensitization to M. tuberculosis and non-tuberculosis mycobacterium [13]. The antigen component is clear and safe, and is especially suitable for people with HIV infection [14]. Since newborns in most countries are vaccinated with BCG after birth, combining the subunit vaccine with BCG can expand the antigen spectrum and induce different types of immune responses, while prolonging the protective effect of BCG and improving the body’s ability to resist TB infection. At present, most TB subunit vaccines are in the clinical phase of research. For example, MVA85A, H4/IC31, and H56/IC31 have been designed for use in adults who have been vaccinated with BCG [15-17]. However, there is still no booster vaccine approved for use. It is speculated that the main limitation of current booster shots may be the cross-immune reaction between the components of the booster vaccine and BCG, as this inhibits the growth of BCG and reduces its immune effect. To avoid the effect of the proteins of the booster immunization on BCG colonization and excessive immune response, the target antigen protein of the booster needs to have high specificity for M. tuberculosis and produce no or little cross-reactivity with BCG [18]. Although vaccines with two different components of recombinant BCG and subunit proteins have become the main research focus of TB vaccines in recent years, there are few comparative studies on the different immune strategies. Therefore, it is of great significance to clarify the differences in the immunogenicity of these immunization strategies and the level of protection they offer.
At present, the immunodominant antigens of TB mainly include early secretory antigens, virulence-associated antigens, cell membrane antigens, growth phase antigens, and dormant phase antigens, which are potential targets for TB vaccine development [3]. Culture filtrate proteins (CFPs) are secreted proteins present in the supernatant of M. tuberculosis culture that can be recognized by T cells and stimulate the production of protective cytokines such as IFN-γ [19]. For example, the RD region proteins ESAT-6 and CFP-10 were widely used in early research on TB vaccines [20]. In addition, the PPE18 protein, which belongs to the PE/PPE gene family, is encoded by the Rv1196 gene, and is enriched with the N-terminal proline (P) and glutamate (E) sequence, has been shown to stimulate rapid proliferation of human T cells in in vitro experiments and can be used as a potential anti-TB vaccine component [21]. Further, the Rv0934 gene encodes phosphate-specific transport substrate binding protein-1 (PstS1), which is involved in the active transport of inorganic phosphate across the membrane and can induce activation of mouse CD8+ T cells and produce Th1 and Th17 immune-protective responses [22]. Among these, PPE18 and PstS1 are expressed at low levels in BCG.
Due to the complex composition of M. tuberculosis, the immunogenicity of individual antigens is low and it is difficult to provide sufficient immune protection with a single antigen. Therefore, combining multiple immunodominant antigens to construct multi-component vaccines is a popular trend in TB vaccine development. In this study, we selected four immunodominant antigens, namely, ESAT-6, CFP-10, nPPE18 (epitope-enriched antigen), and nPstS1 (epitope-enriched antigen), which had been validated in our previous study[23], to construct recombinant BCG and subunit proteins. First, we induced expression of the fusion protein EPCP009 containing ESAT-6, CFP-10, nPPE18, and nPstS1, and used DDA/poly:IC as an adjuvant to create the EPCP009/DP subunit vaccine. In addition, the genes of the four immunodominant antigens were concatenated and introduced into the pMV361 plasmid, and the recombinant BCG vaccine rBCG-EPCP009 was constructed using the BCG Chinese strain as the parent. With Balb/c mice as a model, we examined the short- and long-term immune responses and immune memory levels, in vitro mycobacterial growth inhibition ability, and safety after immunization with the BCG, rBCG-EPCP009, BCG prime-EPCP009/DP booster, and EPCP009/DP vaccines. We explore the optimal vaccine components and immunization strategies for constructing a novel and effective TB vaccine and provide a theoretical basis for the study of prime-boost immunization strategies.
Materials and methods
Bacterial strains and culture conditions
M. tuberculosis H37Rv, M. bovis BCG China, and rBCG strains were cultured on Difco ™ Middlebrook 7H9 (BD, USA) or Difco ™ Middlebrook 7H10 agar (BD, USA) supplemented with 10% oleic-albumin-dextrose-catalase (OADC), 0.5% glycerol, and 0.05% Tween 80. Escherichia coli DH5α and BL21(DE3) were cultured in Luria-Bertani medium or agar and used for cloning and expression. Kanamycin was used at a concentration of 25 μg/mL and ampicillin used at a concentration of 100 μg/mL.
Recombinant rBCG-EPCP009 and the fusion protein EPCP009
The expression plasmid pET43.1a-009 and shuttle plasmid pMV361-009 were constructed by Universal Biosystems Co. Ltd. (Anhui, China). The procedure is briefly described here: EPCP009 (esxA, esxB, nPPE18, and nPstS1 genes) was attached to the pET43.1a vector and pMV361 vector via a linker. As an integral component of fusion protein recombination, the linker is an amino acid chain that connects two fusion genes and has a certain degree of flexibility that allows the proteins on both sides to perform their independent functions [24]. Non-polar hydrophobic amino acids, such as glycine (Gly) and serine (Ser), are commonly used. In the design of our fusion protein vaccines, the linker we chose was Gly-Gly-Ser-Gly-Gly, which is encoded by the GGTGGTTCTGGCGGT sequence.
The pMV361-009 plasmid was transformed into the BCG receptor state by electroshock (voltage, 2.5 kV; capacitance, 25 μF, resistance, 1000 Ω) and screened with the 7H10 resistance medium containing 25 μg/mL kanamycin. A monoclonal colony of rBCG-EPCP009 was cultured in 7H9 medium containing 25 μg/mL kanamycin. The integration of pMV361-009 into BCG was verified by PCR, and the amplified product was sent to the TsingKe Biotech Corp. (Beijing, China) for sequencing to verify the successful construction of the rBCG-EPCP009 strain. The expression level of the target gene was verified by extracting mRNA and using it for RT-qPCR. The primers for verifying the plasmid integration of rBCG-EPCP009 and mRNA expression levels of rBCG-EPCP009 are shown in Table 1.
The transformation of plasmids and the expression and purification of EPCP009 protein are described in previous articles[25]. Briefly, pET43.1a-009 was transformed into E. coli BL21 (DE3). A monoclonal colony of BL21 was cultured in Luria-Bertani liquid medium containing 100 μg/mL ampicillin (37°C, 180 rpm). The bacterial cultures were induced with 1 mM isopropyl β-d-1-thiogalactopyranoside. Then, the proteins were purified from the inclusion bodies by nickel chelate chromatography (GE Healthcare, PA, USA). Endotoxin was removed from the purified proteins using a ToxinEraser™ Endotoxin Removal Kit (GenScript, NJ, USA) and the proteins were filtered through 0.45-μm filters.
Immunization regimens and sample collection
Specific pathogen-free, 4- to 6-week-old female Balb/c mice were obtained from Beijing HFK Bioscience Co. Ltd. (Beijing, China). In order to evaluate the impact of different immunization schemes (shown in Figure 1), the mice were randomly divided into five groups of six mice each for vaccination. Mice of group 1 (PBS) were immunized with 100 μL of phosphate-buffered saline (PBS, pH 7.4); mice of groups 2 (BCG) and 3 (rBCG-EPCP009) were immunized intradermally with 1 × 106 colony-forming units (CFUs) of the BCG/rBCG strain in 100 μL of PBS (pH 7.4); mice of group 4 (BCG+EPCP009) were immunized intradermally with 1 × 106 CFUs of the BCG strain at week 0 and immunized subcutaneously three times (at weeks 0, 2, and 4) with 10 μg EPCP009 protein formulated in DP (composed of 250 μg dimethyldioctadecylammonium [DDA]/50 μg poly I:C) in 100 μL of PBS (pH 7.4); mice of group 5 (EPCP009) were immunized subcutaneously three times (at weeks 0, 2, and 4) with 10 μg EPCP009 protein formulated in DP. Eight or twelve weeks from the first immunization, blood samples were collected from the orbits of the mice and they were sacrificed by cervical dislocation. The coagulated blood was centrifuged at 4000 rpm for 10 min, and the serum was stored at -20℃ for antibody titer analysis. Spleen cells were isolated under aseptic conditions by using mouse lymphocyte separation medium (Dakewe, Beijing, China) according to the manufacturer’s instructions.
Mice in each group were immunized with PBS, BCG, rBCG-EPCP009, BCG+EPCP009/DP, or EPCP009/DP, delivered by either the intradermal or the subcutaneous route. Eight or twelve weeks from the first booster, blood samples (from the orbit) and spleen cells were collected to analyze their immunogenicity and in vitro protection ability.
Luminex cytokine test and enzyme-linked immunospot assay
The nine cytokines representing different cellular immune types were detected by Luminex assay, and Th1-type cytokine IFN-γ and Th2-type cytokine IL-4 were detected Enzyme-linked immunospot (ELISpot) method as previously described[25]. What's different is that cells are stimulated with PPD and EPCP009, respectively. Briefly, the mouse splenocytes were plated in 96-well Luminex Mouse Magnetic Assay (R&D Systems, Minneapolis, MN, USA) culture plates at 2 × 105 cells/well and cultured with 10 μg of the PPD or EPCP009 antigen (i.e., 50 μL of a 200 ng/μL solution) for 24 h. Add biotin antibodies and antibodies separately, wash, and then perform testing. In addition, spleen cells detected by Elispot method were stimulated with 2 μg of PPD or EPCP009 protein, respectively.
ELISA
The IgG, IgG1, and IgG2a antibody concentrations in the serum were measured by ELISA[25]. Briefly, 96-well ELISA plates were coated with 200ng of the PPD or EPCP009 protein and incubated overnight at 4°C, and then washed five times with PBST (PBS with 0.5% Tween 20). The plates were blocked with 3% skimmed milk for 2 h at 37°C and washed five times with PBST. Two-fold dilutions serum was added and incubated for 2 h at 37°C and washed five times, and then added 100 μL/well HRP-conjugated goat anti-mouse IgG, IgG1, or IgG2a (Sigma, St. Louis, MO, USA) diluted to 1:5000 in PBS. The plates were incubated for 1 h at 37°C and washed five times, and 100 μL of tetramethylbenzidine substrate was added for 15 min at 37°C. The reactions were terminated with the addition of 100 μL/well of 2 M H2SO4, and absorbance was measured on an ELISA plate reader at a wavelength of 450 nm. Absorbance values of >2.1 (OD450 [experimental group]/OD450 [PBS control]) were considered to indicate the presence of IgG.
Flow cytometry
In order to further analyze the immune mechanism of each vaccine, the spleen cells of each group were stimulated with PPD and EPCP009 after 8 and 12 weeks of the initial immunization, and the proportions of CD3+CD4+Th cells and CD3+CD8+CTL cells among T cells were analyzed by flow cytometry combined with intracellular cytokine staining. Following this, the expression of CD62L and CD44 was examined as sorting markers for determining the proportion of effector memory T cells (TEM cells: CD62Llo CD44hi) and central memory T cells (TCM cells: CD62Lhi CD44hi) in the CD4+ or CD8+ cell populations, as well as the proportion of cells secreting IFN-γ among TEM cells and the proportion of cells secreting IL-2 among TCM cells.
The mouse splenocytes were plated in 96-well culture plates containing 100-μL Luria-Bertani culture medium at a density of 2 × 107 cells/well. Then, 2 × 106 cells from the 100-μL cell suspension were added to 96-well culture plates, to which 100 μL of the prepared stimulation mixture was also added to obtain final concentrations of 10 μg/mL of specific antigens, 1 μg/mL of the CD28 antibody, 1 μg/mL of the CD49d antibody, and 1000× dilution of the Brefeldin A (BFA) blocker, and the cultured cells were stimulated for 8 h in a 5% CO2 incubator at 37°C after mixing. Two positive control wells were set up, and the non-specific stimulant Cell Stimulation Cocktail (with BFA) was added to the wells. Lymphocytes were collected from the culture well in a 5-mL flow tube, to which 2 mL of PBS was added and mixed well. The solution was then centrifuged at 300 g for 5 min, and the supernatant was discarded. This solution was mixed well with 1 ml of Ghost Dye™ Violet 510 (for labelling dead cells) prepared by diluting 1000 times with PBS, and incubated for 15 min at room temperature in the dark. Next, 2 mL of flow staining wash was added and mixed well. The solution was centrifuged at 300 g for 5 min, and the supernatant was discarded. The cell pellet was resuspended in 100 μL of flow cytometry washing solution, and fluorescence-labeled antibodies (CD3, CD4, CD8, CD62L, and CD44) (BioLegend, USA) of cell surface antigens were added, mixed well, and incubated for 20 min at room temperature in the dark. Next, 2 mL of flow staining wash was added and mixed well, and the supernatant was discarded after centrifugation at 300 g for 5 min. Cell fixative (0.5 mL) was added to each tube, mixed, and incubated for 20 min at room temperature in the dark. The solution was centrifuged at 300 g for 5 min, and the supernatant was discarded. Following this, 2 mL of cytoplasmic membrane breaking lotion was added, mixed well, and centrifuged at 300 g for 5 min. The supernatant was discarded, and the pellet was mixed with a solution containing fluorescence-labeled antibodies (IL-2, TNF-α, and IFN-γ) (BioLegend, USA) against the cytoplasmic antigens, mixed well, and incubated in the dark at room temperature for 30 min. Then, 2 mL of cytoplasmic membrane-breaking lotion was added to each tube and mixed well. The solution was centrifuged at 300 g for 5 min, and the supernatant was discarded. The cells were resuspended in 0.5 mL of cell stain/wash and subjected to flow cytometry analysis (Cytek NL-CLC3000, USA). The data obtained were analyzed with the FACS Diva 8.0 software.
In vitro mycobacterial growth inhibition assay
The mycobacterial growth inhibition assay (MGIA) could evaluate the ability of the splenocytes of mice to inhibit M. tuberculosis growth under in vitro conditions and evaluate the protective effect of TB vaccines, which is in line with the 3R principles in animal experiments: reduction, replacement, refinement. The protocol was adapted from a previously published MGIA protocol for mice splenocytes and further optimized in this laboratory[26]. Briefly, the cells were adjusted to a density of 2 × 106 splenocytes per 300 µL of antibiotic-free RPMI 1640 medium containing 10% heat-inactivated fetal bovine serum, 10 mM HEPES, and 2 mM l-glutamine. M. tuberculosis H37Rv was diluted to a concentration of 50 CFUs/300 µL. Bacterial aliquots (300 µL) were added to the splenocytes to form a splenocyte–mycobacteria co-culture (600 µL), which was incubated in 24-well plates at 37°C for 4 days. Subsequently, the co-cultures were collected and this was followed by centrifugation at 12,000 rpm for 10 min and the supernatant was discarded after centrifugation. Sterile water (500 µL) was added to each tube and mixed, and 50 μL of the solution was inoculated on 7H10 medium containing 5 μg/mL thiophene 2-carboxylic acid hydrazine (TCH) and 10% OADC supplement and incubated at 37°C for 3 weeks. The number of bacteria was counted, and the data were presented as the log10CFU value of the total number per sample.
Statistical analysis
The experimental data were expressed as the mean ± SD using GraphPad Prism 8. One-way analysis of variance (ANOVA) with Tukey’s multiple comparison tests were used for comparison of more than two groups. P-values <0.05 were considered to indicate statistical significance.
Results
Verification of rBCG-EPCP009 and the fusion protein EPCP009
The whole genome of rBCG-EPCP009 was extracted with the cetyltrimethylammonium bromide (CTAB) method and was amplified by PCR. The PCR-amplified bands were verified by agarose gel electrophoresis, and the sequencing results were consistent with expectations (Figure 2B). RT-qPCR confirmed that the levels of the antigen-specific mRNA of nPPE18 and nPstS1 in rBCG-EPCP009 were higher than those in BCG. The mRNA levels of nPPE18 and nPstS1 were five to ten times higher in rBCG-EPCP009 group than in BCG (Figure 2C). The expression plasmid pET43.1a-009 was transformed into E. coli BL21, and the predicted molecular weight of the expressed EPCP009 protein was 51 kD. The recovery yield of EPCP009 proteins was around 0.8 mg/L of medium. SDS-PAGE indicated that the molecular weight of the proteins was as per expectations, and the purity level of the proteins was high (Figure 2D).
Long-term induction of high levels of multiple protective cytokines by BCG+EPCP009
To better compare the short- and long-term cellular immunity levels and the types of immune responses induced in mice after immunization with the different strategies, the spleens of the immunized mice were isolated under aseptic conditions at 8 and 12 weeks from the first immunization and stimulated with 10 μg each of the PPD and EPCP009 antigens. The levels of nine cytokines, such as Th1-, Th2-, and Th17-type cytokines and other types of cytokines, were detected in culture supernatants with the Luminex kit.
After 8 weeks of the initial immunization, mouse spleen lymphocytes were stimulated by PPD (Figure 3A). For all the cytokines compared, the PBS group secreted lower levels of cytokines than the other groups. Compared with the BCG group mice, the rBCG-EPCP009 and BCG+EPCP009 group mice produced significantly higher levels of IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, IL-12, IL-4, and IL-6 (p < 0.05). The BCG+EPCP009 group mice produced significantly higher levels of IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, IL-12, IL-4, and IL-6 than the rBCG-EPCP009 group (p < 0.05) and EPCP009 group mice (p < 0.05). The level of the Th2-type cytokine IL-4 was lower than 100 pg/mL in the BCG, rBCG-EPCP009, BCG+EPCP009, and EPCP009 groups.
After 8 weeks of the initial immunization, mouse spleen lymphocytes were stimulated with the EPCP009 protein (Figure 3B). As observed for PPD, the PBS group mice secreted lower levels of cytokines than mice from the other groups. Compared with the BCG group mice, the rBCG-EPCP009, BCG+EPCP009, and EPCP009 group mice produced higher levels of IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, IL-12, and IL-6 (p < 0.05). Further, the BCG+EPCP009 group mice produced significantly higher levels of IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, and IL-12 than the rBCG-EPCP009 and EPCP009 group mice (p < 0.05).
After 12 weeks of the initial immunization, mouse spleen lymphocytes were stimulated by PPD (Figure 4A). The PBS group mice secreted low levels of cytokines. Compared with the BCG group mice, the rBCG-EPCP009 and BCG+EPCP009 group mice produced significantly higher levels of IFN- γ, IL-2, TNF- α, IL-17, GM-CSF, and IL-12 (p < 0.05); further, the BCG+EPCP009 group mice produced significantly higher levels of IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, and IL-6 than the rBCG:: 009 and EPCP009 group mice (p < 0.05). In conclusion, the BCG+EPCP009 group mice produced higher levels of PPD-specific cytokines, with a bias towards Th1- and Th17-type cytokines, while the BCG group mice did not produce IL-17 or GM-CSF.
After 12 weeks of the initial immunization, mouse spleen lymphocytes were stimulated with the EPCP009 protein (Figure 4B). PBS and BCG group mice secrete low levels of the EPCP009-specific cytokines. The BCG+EPCP009 group mice produced significantly higher levels of IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, and IL-6 than the rBCG-EPCP009 and EPCP009 group mice (p < 0.05). The levels of IL-4 and IL-10 produced by mice from all the groups were lower than 100ng/μl.
Long-term induction of high levels of IFN-γ in the spleen of mice vaccinated with BCG+EPCP009
At 8 and 12 weeks from the first immunization, spleens were isolated under aseptic conditions and stimulated with 2 μg PPD and 2 μg EPCP009, and the culture supernatant was assayed for the Th1-type cytokine IFN-γ and the Th2-type cytokine IL-4 using the Elispot kit.
After 8 weeks of the initial immunization, compared with the PBS group mice, the BCG, rBCG-EPCP009, and BCG+EPCP009 group mice produced higher levels of PPD-specific IFN-γ and IL-4 (p < 0.05), and the rBCG-EPCP009, BCG+EPCP009, and EPCP009 group mice produced higher levels of EPCP009-specific IFN-γ (p < 0.05, Figure 5A). Compared with the BCG group mice, both the rBCG-EPCP009 and BCG+EPCP009 group mice produced higher levels of IFN-γ in response to PPD or EPCP009 stimulation (p < 0.01). However, the BCG+EPCP009 group mice produced significantly higher levels of IFN-γ than the rBCG-EPCP009 and EPCP009 group mice (p < 0.001).
At 12 weeks after immunization, the PBS group still secreted lower levels of IFN-γ and IL-4 cytokines than the four immunization groups. Compared with the PBS group mice, the BCG, rBCG-EPCP009, and BCG+EPCP009 group mice produced higher levels of PPD-specific IFN-γ and IL-4 (p < 0.05) (Figure 5B); the BCG+EPCP009 and EPCP009 group mice produced higher levels of EPCP009-specific IFN-γ (p < 0.05) (Figure 5B). The BCG+EPCP009 group mice produced significantly higher levels of PPD- and EPCP009-specific IFN-γ than the BCG, rBCG-EPCP009, and EPCP009 group mice (p < 0.01) (Figure 5B).
Induction of Th1-specific antibody types by BCG+EPCP009
Total IgG, IgG1, and IgG2a antibody titers and IgG2a/IgG1 ratios were compared between the PBS (control group) and the four immunization groups at 8 (A) and 12 (B) weeks after immunization. The p-values indicating the significance of the differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (“ns” represents no significance).
At 8 and 12 weeks after immunization, the levels of PPD- and EPCP009-specific IgG, IgG1, and IgG2a antibodies in serum samples from the mice were detected by indirect ELISA, and the immune response type (Th1/Th2) was determined by calculating the ratio of IgG2a to IgG1.
With regard to the levels of PPD-specific antibodies (Figure 6), after 8 and 12 weeks of immunization, compared with the PBS group mice, the BCG, rBCG-EPCP009, and BCG+EPCP009 group mice produced higher levels of IgG, IgG1, and IgG2a antibodies, respectively. At 8 weeks, the BCG+EPCP009 group mice produced higher levels of IgG, IgG1, and IgG2a antibodies than the BCG group mice and higher levels of IgG1 and IgG2a antibodies than the rBCG-EPCP009 group mice (p < 0.05). The IgG2a/IgG1 ratio of the BCG, rBCG-EPCP009, and BCG+EPCP009 groups was greater than 1, and there was no statistically significant difference between groups.
With regard to the levels of EPCP009-specific antibodies (Figure 6), after 8 weeks, the BCG+EPCP009 and EPCP009 group mice produced higher levels of IgG, IgG1, and IgG2a antibodies than the rBCG-EPCP009 group mice (p < 0.05). After 12 weeks of immunization, compared with the rBCG-EPCP009 group mice, the BCG+EPCP009 group mice produced higher levels of IgG and IgG2a antibodies (p < 0.05), and the IgG2a/IgG1 ratio of the BCG+EPCP009 group was slightly higher than that of the other three immunization groups.
Consistently high levels of IFN-γ+TEM and IL-2+TCM cells in the spleens of mice vaccinated with BCG+EPCP009
Splenocytes from Balb/c mice immunized with the candidate vaccines were stimulated with the PPD or EPCP009 antigen at 8 and 12 weeks, and the proportions of IFN-γ+CD4+ and IFN-γ+CD8+TEM cells (A, B, E, F) and IL-2+CD4+ and IL-2+CD8+TCM cells (C, D, G, H) were assessed by flow cytometry. The p-values indicating the significance of differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
After 8 weeks of immunization, the proportions of PPD- and EPCP009-specific IFN-γ+CD4+TEM and IFN-γ+CD8+TEM cells were significantly increased in the BCG, rBCG-EPCP009, BCG+EPCP009, and EPCP009 groups compared with the PBS group (p < 0.001) (Figure 7A and 7B). Further, the proportions of PPD- and EPCP009-specific IL-2+CD4+TCM and IL-2+CD8+TCM cells in the rBCG-EPCP009 and BCG+EPCP009 groups were significantly increased in comparison to the PBS group (p < 0.001). In contrast, the BCG and EPCP009 group mice did not produce IL-2+CD4+TCM or IL-2+CD8+TCM cells (Figure 7C and 7D). After 12 weeks of immunization, the BCG group mice did not produce IFN-γ+TEM or IL-2+TCM cells, while the rBCG-EPCP009 and BCG+EPCP009 groups produced PPD- and EPCP009-specific IFN-γ+CD4+TEM, IFN-γ+CD8+TEM, IL-2+CD4+TCM, and IL-2+CD8+TCM cells (p < 0.05).
The BCG+EPCP009 group mice produced significantly higher proportions of PPD- and EPCP009-specific IFN-γ+CD4+TEM and IL-2+CD8+TCM cells than the rBCG-EPCP009 and EPCP009 group mice when the three experimental immunization groups were compared (p < 0.05). In addition, the proportions of PPD- and EPCP009-specific IFN-γ+CD8+TEM and IL-2+CD4+TCM cells in the BCG+EPCP009 group were equivalent to those of the rBCG-EPCP009 group.
Better growth inhibition of H37Rv by the BCG prime-EPCP009 booster than by rBCG-EPCP009 and EPCP009
To determine the ability of immunized mice to inhibit M. tuberculosis growth under in vitro conditions and the duration of this effect, splenocytes were isolated after 8 and 12 weeks of immunization and co-cultured with H37Rv, and the number of colonies formed were counted. After 8 weeks of immunization, the highest colony counts were observed in the PBS-negative control group. Compared with the PBS group, the colony counts in the BCG, rBCG-EPCP009, BCG+EPCP009, and EPCP009 groups were significantly lower (p < 0.01). Compared with the BCG group, the colony counts in the rBCG-EPCP009 and BCG+EPCP009 groups were significantly lower (p < 0.01). Further, the BCG+EPCP009 group had the lowest colony count, and this was significantly lower than that of the rBCG-EPCP009 (p < 0.001) and EPCP009 groups (p < 0.05) (Figure 8).
After 12 weeks of immunization, the PBS-negative control group still had the highest colony count, and the BCG, rBCG-EPCP009, BCG+EPCP009, and EPCP009 groups had significantly lower colony counts than the PBS group (p < 0.01). The number of colonies formed was significantly lower in the BCG+EPCP009 group than in the BCG group (p < 0.01), but there was no significant difference between the rBCG-EPCP009 and EPCP009 groups. Moreover, the colony counts in the BCG+EPCP009 group were significantly lower than those in the rBCG-EPCP009 (p < 0.001) and EPCP009 groups (p < 0.001). In conclusion, after 12 weeks of immunization, the BCG+EPCP009 group showed significantly better inhibition than the BCG, rBCG-EPCP009, and EPCP009 groups (Figure 8).
Discussion
Tuberculosis remains a serious infectious disease that poses a threat to human health. BCG is the only preventive vaccine currently approved for human use. However, while it is effective in preventing severe TB in infants and children, it has severely reduced protection in adolescents and adults [6]. Currently, 16 novel TB vaccine candidates are in clinical trials worldwide, and recombinant BCG and subunit protein vaccines are two important vaccine strategies. Recombinant BCG is intended for neonatal replacement of BCG, and the subunit protein is mainly used for booster vaccines after primary BCG immunization. Both recombinant BCG and the subunit protein are expected to be useful strategies for the development of new vaccines and for the development of immunization protocols that provide stronger and longer-lasting immunity based on the inherited parental BCG [27]. In this study, we constructed recombinant rBCG-EPCP009 and subunit protein EPCP009 vaccine candidates based on the multi-component immunodominant antigen EPCP009, and evaluated the short- and long-term immunogenicity and in vitro protection offered by three immunization strategies, namely, rBCG-EPCP009, BCG+EPCP009, and EPCP009/DP. Our results showed that the BCG prime-EPCP009 booster provided superior and longer-lasting immunogenicity and better in vitro protection than rBCG-EPCP009 and EPCP009/DP, as reflected in the following results. That is, BCG+EPCP009 induced stronger PPD- and EPCP009-specific Th1-type and Th17-type immune responses based on the levels of the cytokines GM-CSF and IL-12, and the IFN-γ+CD4+TEM and IL-2+CD8+TCM cell ratios, at both 8 and 12 weeks after immunization, while BCG+EPCP009 induced EPCP009-specific antibodies that were indicative of a bias toward Th1-type immune responses. In addition, immunization with BCG+EPCP009 resulted in stronger in vitro mycobacterium inhibition by splenocytes and had a longer in vitro protective effect.
The selection of an appropriate immunodominant antigen is an important factor in determining vaccine efficacy, irrespective of whether the strategy involves the recombinant BCG or subunit vaccine. rBCG30, which expresses the Ag85B antigen, was the first vaccine candidate of this type to enter the clinical trial phase and showed superior immune efficacy to the parent BCG in both mice and guinea pigs. However, the phase I clinical trials conducted in the United States in 2004 showed that the immunogenicity of rBCG30 was lower than that of BCG, and therefore, the findings from human trials were not encouraging [28]. Subsequently, the strategy for construction of recombinant BCG vaccines changed from overexpression of a single antigen to simultaneous overexpression of multiple immune dominant fusion antigens, in order to further improve the immune effect of recombinant BCG. For example, rBCG::XB, which is in the preclinical phase, is a novel recombinant BCG that expresses both the Ag85B and HspX antigens and induces a stronger antigen-specific Th1-type immune response in mice than BCG [29]. In addition, rBCG::685A, which expresses both ESAT-6 and Ag85A, also induced higher levels of PPD or r685A-specific IFN-γ in mice [30]. In this study, based on the ESAT-6 and CFP-10 proteins, the fusion gene EPCP009 was constructed by adding the antigens nPPE18 and nPstS1. Recombinant rBCG-EPCP009 overexpressing the EPCP009 protein was constructed by the pMV361 shuttle plasmid. In addition, the ESAT-6, CFP-10, nPPE18, and nPstS1 genes were arranged in tandem, and the fusion protein EPCP009 and DDA/poly:IC adjuvant were mixed to construct the EPCP009/DP vaccine candidate, which was used as a booster vaccine after initial immunization with BCG. Compared with mice immunized with rBCG-EPCP009, mice immunized with the BCG prime-EPCP009 booster strategy secreted higher levels of PPD- and EPCP009-specific IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, and IL-12 after 8 and 12 weeks of immunization. In vitro protection assays showed that splenic lymphocytes from BCG+EPCP009-immunized mice showed superior M. tuberculosis growth inhibition than splenocytes from rBCG-EPCP009-immunized mice at both 8 and 12 weeks. This indicates that although mice from all the immunization groups expressed the fusion protein EPCP009, the immune mechanisms induced differed according to the technology platforms and immunization strategies employed.
Under the stimulation of different antigens, Th0 cells differentiate into different Th1 and Th2 cell subsets, and different subsets of T cells secrete different types of cytokines in response to pathogens [31]. As an intracellular bacterium, M. tuberculosis mainly produces IL-12 by inducing its secretion from antigen-presenting cells. These antigen-presenting cells induce Th1 cells to release IFN-γ, IL-2, and TNF-α and activate macrophages and cytotoxic T lymphocytes, which then fulfill their function of M. tuberculosis clearance [32]. However, IL-4 induces the polarization of Th0 to the Th2 subset, and the production of IL-4 and IL-10 inhibits the anti-infection process and leads to the dissemination of M. tuberculosis [33]. Th17 cells are a unique group of Th cells that mainly secrete IL-17 and GM-CSF, which recruit and activate neutrophils to the site of infection. Th17cells and neutrophils enhance the protective immune response by producing IL-12 and inducing a Th1 response [34]. IL-6 is a pro-inflammatory cytokine that can enhance the immune response by promoting the proliferation of T cells, B cells, and other immune cells [35]. According to the latest evidence, the main effector T-cell subsets involved in M. tuberculosis clearance are Th1 and Th17 cell subsets [36]. However, certain components of the M. tuberculosis cell wall, such as ManLAM, can promote IL-10 production and secretion by inhibiting the production of IFN-γ and TNF-α, thereby affecting antigen presentation or inhibiting the migration of effector T cells to the site of M. tuberculosis infection and resulting in immune escape [37]. Compared with BCG, the recombinant BCG vaccines rBCG-EPCP009 and BCG+EPCP009 resulted in the secretion of increased levels of PPD- and EPCP009-specific IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, and IL-12 from mouse splenocytes after 8 and 12 weeks of immunization. These results indicate that rBCG-EPCP009 and BCG+EPCP009 induced a protective immune response which was superior to that of BCG. In addition, on comparing the three EPCP009-based immunization strategies, the level of PPD- and EPCP009-specific IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, and IL-6 stimulated by BCG+EPCP009 was found to be significantly higher than that stimulated by rBCG-EPCP009 and EPCP009 after both 8 and 12 weeks of immunization. This result indicated that the combined strategy involving BCG+EPCP009 promoted not only the immune effect of BCG itself but also the immune response to the EPCP009 protein. Thus, BCG and EPCP009 may have induced a stronger immune effect that was brought about by a mutual adjuvant effect.
Memory T cells are mainly divided into TCM cells and TEM cells [38], and T cells are highly heterogeneous in terms of their immune response. When exposed to antigen stimulation, TCM cells mainly secrete IL-2, rapidly proliferate and differentiate into effector T cells, and produce IFN-γ. TEM cells mainly secretes IFN-γ, which migrates to the site of inflammation and rapidly exerts its immune effects [39]. It has been shown that IFN-γ+TEM and IL-2+TCM cells are biomarkers of vaccine-induced production of anti-M. tuberculosis antibodies and early and persistent infection, respectively [40]. The main reason for the decrease in the protective properties of BCG with time may be that its induced protective immune response decreases with time so that it induces a weaker CD8+ immune response in adolescents and adults [41]. The results of the present study showed that the proportions of neither IFN-γ+TEM nor IL-2+TCM cells were significantly higher in the BCG group compared with the PBS group after 12 weeks of immunization. This may partly explain why the protective effect of BCG is only maintained for 10–15 years and highlights the need for a TB vaccine with a longer immune memory. After 8 or 12 weeks of immunization, the rBCG-EPCP009 and BCG+EPCP009 vaccines induced significantly higher proportions of PPD- or EPCP009-specific IFN-γ+CD4+TEM, IL-2+CD4+TCM, and IL-2+CD8+TCM cells than BCG. This finding indicates that immune memory was significantly enhanced with the rBCG-EPCP009 and BCG+EPCP009 vaccine candidates. In addition, the proportion of antigen-specific IL-2+CD8+TCM cells induced by PPD or EPCP009 was significantly higher in the BCG+EPCP009 group than in the rBCG-EPCP009 and EPCP009 groups. When CD8+TCM cells were exposed to the antigen again, they rapidly proliferated and differentiated and produced IL-2, while promoting the cytotoxic functions of effector CD8+ T cells, which in turn enhanced their anti-infective effects [42, 43]. Thus, the stronger protective effect of VPM1002 than BCG may be related to its ability to induce high levels of TCM production [11]. In conclusion, several studies have demonstrated the correlation between the proportions of IFN-γ+ TEM and IL-2+ TCM cells and the protective effect of TB vaccines. With regard to the vaccine candidates evaluated in the presents study, BCG+EPCP009 induced higher and longer-lasting levels of IFN-γ+CD4+ TEM and IL-2+CD8+ TCM cells. Thus, this immunization strategy may provide a longer-lasting immune protective effect.
MGIA is an effective method to comprehensively evaluate the protective effect of TB vaccines in vitro [26, 44]. Parra et al. used mice as a model to test the inhibitory effect of splenocytes from vaccine-immunized mice on M. tuberculosis in macrophages, and the results were consistent with the results of challenge protection experiments on animals [45]. The results of this study showed that after 8 weeks of the initial immunization, splenocytes from the rBCG-EPCP009- and BCG+EPCP009-immunized groups showed stronger in vitro mycobacterial inhibition than splenocytes from the BCG group; further, EPCP009 group splenocytes showed comparable inhibition to BCG group splenocytes. After 12 weeks of the initial immunization, the BCG+EPCP009 group splenocytes still exhibited better inhibition than the BCG group splenocytes, while the rate of inhibition of the rBCG-EPCP009 and EPCP009 group splenocytes were not significantly different from that of the BCG group splenocytes. This indicates that rBCG-EPCP009 provided better short-term protection than BCG, but did not provide long-term protection. At both 8 and 12 weeks after immunization, BCG+EPCP009 exhibited durable superior bacterial growth inhibition to rBCG-EPCP009 and EPCP009; this may be related to its ability to induce the production of PPD- and EPCP009-specific TNF-α, IFN-γ, IL-12, IL-17, IFN-γ+CD4+ TEM cells, and IL-2+CD8+ TCM cells.
In recent years, the protective effect of the BCG prime-BCG homologous enhancement strategy has been under debate, and many studies conducted in different countries, such as Japan and Finland, have shown that BCG revaccination does not improve the protective effect of BCG itself and may even cause pathological damage [46, 47]. Currently, the immunization strategy involving primary immunization with BCG and a heterologous boost has become the focus for the development of novel vaccines for TB, and several subunit protein and viral vector vaccines are in the clinical phase of evaluation for their application as booster immunization. The results of this study showed that the BCG+EPCP009 immunization group exhibited a more durable protective immune response and in vitro M. tuberculosis inhibition. Thus, administering the EPCP009 subunit protein vaccine as a booster immunization vaccine for BCG could enhance the immune protective response of the body. Overall, comparison of the different strategies revealed that the BCG prime-EPCP009 protein booster immunization strategy was superior to the rBCG-EPCP009 and EPCP009 strategies and may provide a theoretical basis for future immunization strategies of TB vaccines.
Funding
This work was financially supported by the Major Projects of the Thirteenth Five-Year Plan Special for Infectious Diseases (Grant No. 2018ZX10731301-002 and 2018ZX10731301-002-005). The funders had no role in the study design, data collection and analysis, manuscript preparation, or decision to publish.
Ethics approval
All the experimental procedures and protocols were approved by the Chinese Center for Disease Control and Prevention and the Ethics Committee of the National Institute for Communicable Disease Control and Prevention. Animal experiments were conducted in accordance with the guidelines of the Animal Experiment Ethics Committee of the China Center for Disease Control and Prevention (No. 2022-CCDC-IAC-026).
Disclosure statement
All the authors declare that they have no competing financial interests or personal relationships.
References
- Bagcchi, S. WHO's Global Tuberculosis Report 2022. Lancet Microbe. 2023, 4, e20. [Google Scholar] [CrossRef]
- Lange, C.; Chesov, D.; Heyckendorf, J.; Leung, C.C.; Udwadia, Z.; Dheda, K. Drug-resistant tuberculosis: An update on disease burden, diagnosis and treatment. Respirology. 2018, 23, 656–673. [Google Scholar] [CrossRef]
- Romano, M.; Squeglia, F.; Kramarska, E.; Barra, G.; Choi, H.G.; Kim, H.J.; et al. A Structural View at Vaccine Development against M. tuberculosis. Cells. 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Dalmia, N.; Ramsay, A.J. Prime–boost approaches to tuberculosis vaccine development. Expert Rev. Vaccines 2012, 11, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Tagliabue, A.; Boraschi, D.; Leite, L.C.C.; Kaufmann, S.H.E. 100 Years of BCG Immunization: Past, Present, and Future. Vaccines 2022, 10, 1743. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Aaby, P.; A Behr, M.; Donald, P.R.; E Kaufmann, S.H.; Netea, M.G.; Mandalakas, A.M. 100 years of Mycobacterium bovis bacille Calmette-Guérin. Lancet Infect. Dis. 2022, 22, e2–e12. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.C.; Pereira, S.M.; Cunha, S.S.; Genser, B.; Ichihara, M.Y.; de Brito, S.C.; A Hijjar, M.; A Cruz, A.; Sant'Anna, C.; Bierrenbach, A.L.; et al. Effect of BCG revaccination on incidence of tuberculosis in school-aged children in Brazil: the BCG-REVAC cluster-randomised trial. Lancet 2005, 366, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Barreto, M.L.; Pereira, S.M.; Pilger, D.; Cruz, A.A.; Cunha, S.S.; Sant’anna, C.; Ichihara, M.Y.; Genser, B.; Rodrigues, L.C. Evidence of an effect of BCG revaccination on incidence of tuberculosis in school-aged children in Brazil: Second report of the BCG-REVAC cluster-randomised trial. Vaccine 2011, 29, 4875–4877. [Google Scholar] [CrossRef]
- Kalra, M.; Grover, A.; Mehta, N.; Singh, J.; Kaur, J.; Sable, S.B.; Behera, D.; Sharma, P.; Verma, I.; Khuller, G. Supplementation with RD antigens enhances the protective efficacy of BCG in tuberculous mice. Clin. Immunol. 2007, 125, 173–183. [Google Scholar] [CrossRef]
- Marques-Neto, L.M.; Piwowarska, Z.; Kanno, A.I.; Moraes, L.; Trentini, M.M.; Rodriguez, D.; Silva, J.L.S.C.; Leite, L.C.C. Thirty years of recombinant BCG: new trends for a centenary vaccine. Expert Rev Vaccines 2021, 20, 1001–1011. [Google Scholar] [CrossRef]
- Cotton, M.F.; A Madhi, S.; Luabeya, A.K.; Tameris, M.; Hesseling, A.C.; Shenje, J.; Schoeman, E.; Hatherill, M.; Desai, S.; Kapse, D.; et al. Safety and immunogenicity of VPM1002 versus BCG in South African newborn babies: a randomised, phase 2 non-inferiority double-blind controlled trial. Lancet Infect. Dis. 2022, 22, 1472–1483. [Google Scholar] [CrossRef] [PubMed]
- Brandt, L.; Feino Cunha, J.; Weinreich Olsen, A.; Chilima, B.; Hirsch, P.; Appelberg, R.; Andersen, P. Failure of the Mycobacterium bovis BCG Vaccine: Some Species of Environmental Mycobacteria Block Multiplication of BCG and Induction of Protective Immunity to Tuberculosis. Infect. Immun. 2002, 70, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Poyntz, H.C.; Stylianou, E.; Griffiths, K.L.; Marsay, L.; Checkley, A.M.; McShane, H. Non-tuberculous mycobacteria have diverse effects on BCG efficacy against Mycobacterium tuberculosis. Tuberculosis 2014, 94, 226–237. [Google Scholar] [CrossRef] [PubMed]
- McClean, S. Prospects for subunit vaccines: Technology advances resulting in efficacious antigens requires matching advances in early clinical trial investment. Hum. Vaccines Immunother. 2016, 12, 3103–3106. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, M.; Satti, I.; Minhinnick, A.; Harris, S.; Riste, M.; Ramon, R.L.; Sheehan, S.; Thomas, Z.-R.M.; Wright, D.; Stockdale, L.; et al. A phase I trial evaluating the safety and immunogenicity of a candidate tuberculosis vaccination regimen, ChAdOx1 85A prime – MVA85A boost in healthy UK adults. Vaccine 2020, 38, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Bekker, L.-G.; Dintwe, O.; Fiore-Gartland, A.; Middelkoop, K.; Hutter, J.; Williams, A.; Randhawa, A.K.; Ruhwald, M.; Kromann, I.; Andersen, P.L.; et al. A phase 1b randomized study of the safety and immunological responses to vaccination with H4:IC31, H56:IC31, and BCG revaccination in Mycobacterium tuberculosis-uninfected adolescents in Cape Town, South Africa. EClinicalMedicine 2020, 21, 100313. [Google Scholar] [CrossRef] [PubMed]
- Tameris, M.D.; Hatherill, M.; Landry, B.S.; Scriba, T.J.; Snowden, M.A.; Lockhart, S.; E Shea, J.; McClain, J.B.; Hussey, G.D.; A Hanekom, W.; et al. Safety and efficacy of MVA85A, a new tuberculosis vaccine, in infants previously vaccinated with BCG: a randomised, placebo-controlled phase 2b trial. Lancet 2013, 381, 1021–1028. [Google Scholar] [CrossRef]
- Woodworth, J.S.; Clemmensen, H.S.; Battey, H.; Dijkman, K.; Lindenstrøm, T.; Laureano, R.S.; Taplitz, R.; Morgan, J.; Aagaard, C.; Rosenkrands, I.; et al. A Mycobacterium tuberculosis-specific subunit vaccine that provides synergistic immunity upon co-administration with Bacillus Calmette-Guérin. Nat. Commun. 2021, 12, 6658. [Google Scholar] [CrossRef]
- Tomee, J.; Kauffman, H.F.; Klimp, A.H.; Demonchy, J.; Köeter, G.H.; Dubois, A.E. Immunologic significance of a collagen-derived culture filtrate containing proteolytic activity in Aspergillus-related diseases. J. Allergy Clin. Immunol. 1994, 93, 768–778. [Google Scholar] [CrossRef]
- Valizadeh, A.; Fooladi, A.A.I.; Sedighian, H.; Mahboobi, M.; Parizad, E.G.; Behzadi, E.; Khosravi, A. Evaluating the Performance of PPE44, HSPX, ESAT-6 and CFP-10 Factors in Tuberculosis Subunit Vaccines. Curr. Microbiol. 2022, 79, 260. [Google Scholar] [CrossRef]
- Hakim, J.M.C.; Yang, Z. Predicted Structural Variability of Mycobacterium tuberculosis PPE18 Protein With Immunological Implications Among Clinical Strains. Front. Microbiol. 2020, 11, 595312. [Google Scholar] [CrossRef]
- Palma, C.; Spallek, R.; Piccaro, G.; Pardini, M.; Jonas, F.; Oehlmann, W.; et al. The M. tuberculosis phosphate-binding lipoproteins PstS1 and PstS3 induce Th1 and Th17 responses that are not associated with protection against M. tuberculosis infection. Clin Dev Immunol. 2011, 2011, 690328. [Google Scholar] [CrossRef]
- Fan, X.; Li, X.; Wan, K.; Zhao, X.; Deng, Y.; Chen, Z.; Luan, X.; Lu, S.; Liu, H. Construction and immunogenicity of a T cell epitope-based subunit vaccine candidate against Mycobacterium tuberculosis. Vaccine 2021, 39, 6860–6865. [Google Scholar] [CrossRef]
- Crasto, C.J.; Feng, J.-A. LINKER: a program to generate linker sequences for fusion proteins. Protein Eng. Des. Sel. 2000, 13, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Fan, X.; Jiang, Y.; Li, G.; Li, M.; Zhao, X.; Luan, X.; Deng, Y.; Chen, Z.; Liu, H.; et al. Immunogenicity and efficacy analyses of EPC002, ECA006, and EPCP009 protein subunit combinations as tuberculosis vaccine candidates. Vaccine 2023, 41, 3836–3846. [Google Scholar] [CrossRef]
- Jensen, C.; Holm, L.L.; Svensson, E.; Aagaard, C.; Ruhwald, M. Optimisation of a murine splenocyte mycobacterial growth inhibition assay using virulent Mycobacterium tuberculosis. Sci. Rep. 2017, 7, 2830. [Google Scholar] [CrossRef] [PubMed]
- Scriba, T.J.; Netea, M.G.; Ginsberg, A.M. Key recent advances in TB vaccine development and understanding of protective immune responses against Mycobacterium tuberculosis. Semin. Immunol. 2020, 50, 101431. [Google Scholar] [CrossRef] [PubMed]
- Hoft, D.F.; Blazevic, A.; Abate, G.; Hanekom, W.A.; Kaplan, G.; Soler, J.H.; Weichold, F.; Geiter, L.; Sadoff, J.C.; Horwitz, M.A. A New Recombinant Bacille Calmette-Guérin Vaccine Safely Induces Significantly Enhanced Tuberculosis-Specific Immunity in Human Volunteers. J. Infect. Dis. 2008, 198, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Teng, X.; Jing, Y.; Ma, J.; Tian, M.; Yu, Q.; Zhou, L.; Wang, R.; Wang, W.; Li, L.; et al. A live attenuated BCG vaccine overexpressing multistage antigens Ag85B and HspX provides superior protection against Mycobacterium tuberculosis infection. Appl. Microbiol. Biotechnol. 2015, 99, 10587–10595. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Liang, J.; Teng, X.; Wang, X.; Zhang, J.; Yuan, X.; Fan, X. Comparison of BCG prime-DNA booster and rBCG regimens for protection against tuberculosis. Hum. Vaccines Immunother. 2014, 10, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ren, W.; Yuan, J.; Guo, H.; Shang, Y.; Wang, W.; et al. Significant difference in Th1/Th2 paradigm induced by tuberculosis-specific antigens between IGRA-positive and IGRA-negative patients. Front Immunol. 2022, 13, 904308. [Google Scholar] [CrossRef] [PubMed]
- McNab, F.W.; Ewbank, J.; Howes, A.; Moreira-Teixeira, L.; Martirosyan, A.; Ghilardi, N.; et al. Type I IFN induces IL-10 production in an IL-27-independent manner and blocks responsiveness to IFN-gamma for production of IL-12 and bacterial killing in Mycobacterium tuberculosis-infected macrophages. J Immunol. 2014, 193, 3600–3612. [Google Scholar] [CrossRef]
- Liu, Q.; Li, W.; Chen, Y. Association of IL-4 rs2243250 polymorphism with susceptibility to tuberculosis: A meta-analysis involving 6794 subjects. Microb. Pathog. 2021, 158, 104959. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.G.; Kwon, K.W.; Choi, S.; Back, Y.W.; Park, H.S.; Kang, S.M.; et al. Antigen-Specific IFN-gamma/IL-17-Co-Producing CD4(+) T-Cells Are the Determinants for Protective Efficacy of Tuberculosis Subunit Vaccine. Vaccines 2020, 8. [Google Scholar] [CrossRef]
- Appelberg, R.; Castro, A.G.; Pedrosa, J.; Minóprio, P. Role of interleukin-6 in the induction of protective T cells during mycobacterial infections in mice. . 1994, 82, 361–364. [Google Scholar]
- Lyadova, I.V.; Panteleev, A.V. Th1 and Th17 Cells in Tuberculosis: Protection, Pathology, and Biomarkers. Mediators Inflamm. 2015, 2015, 854507. [Google Scholar] [CrossRef]
- Tang, X.; Hu, F.; Xia, X.; Zhang, H.; Zhou, F.; Huang, Y.; Wu, Y. [Mannose-capped lipoarabinomannan (ManLAM) binding TLR2 activates mast cells to release exosomes and induces M2 polarization of macrophages]. .Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2021, 37, 481–486. [Google Scholar]
- Henao-Tamayo, M.I.; Ordway, D.J.; Irwin, S.M.; Shang, S.; Shanley, C.; Orme, I.M. Phenotypic Definition of Effector and Memory T-Lymphocyte Subsets in Mice Chronically Infected withMycobacterium tuberculosis. Clin. Vaccine Immunol. 2010, 17, 618–625. [Google Scholar] [CrossRef]
- Seder, R.A.; Darrah, P.A.; Roederer, M. T-cell quality in memory and protection: implications for vaccine design. Nat Rev Immunol. 2008, 8, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Tian, M.; Fan, X.; Yu, Q.; Jing, Y.; Wang, W.; Li, L.; Zhou, Z. Mycobacterium tuberculosis multistage antigens confer comprehensive protection against pre- and post-exposure infections by driving Th1-type T cell immunity. Oncotarget 2016, 7, 63804–63815. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.W.; Aceves-Sanchez, M.J.; Segura-Cerda, C.A.; Choi, E.; Bielefeldt-Ohmann, H.; Shin, S.J.; et al. BCGDeltaBCG1419c increased memory CD8(+) T cell-associated immunogenicity and mitigated pulmonary inflammation compared with BCG in a model of chronic tuberculosis. Sci Rep. 2022, 12, 15824. [Google Scholar] [CrossRef] [PubMed]
- Schwendemann, J.; Choi, C.; Schirrmacher, V.; Beckhove, P. Dynamic Differentiation of Activated Human Peripheral Blood CD8+ and CD4+ Effector Memory T Cells. J. Immunol. 2005, 175, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.N.; Gebhardt, T.; Carbone, F.R.; Heath, W.R. Memory T cell subsets, migration patterns, and tissue residence. Annu Rev Immunol. 2013, 31, 137–161. [Google Scholar] [CrossRef] [PubMed]
- Pepponi, I.; Khatri, B.; Tanner, R.; Villarreal-Ramos, B.; Vordermeier, M.; McShane, H. A mycobacterial growth inhibition assay (MGIA) for bovine TB vaccine development. Tuberculosis 2017, 106, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.L.; Schmidt, T.E.; Stibitz, S.; Derrick, S.C.; Morris, S.L.; Parra, M. A simplified mycobacterial growth inhibition assay (MGIA) using direct infection of mouse splenocytes and the MGIT system. J. Microbiol. Methods 2016, 131, 7–9. [Google Scholar] [CrossRef]
- Tala-Heikkilä, M.M.; Tuominen, J.E.; Tala, E.O.J. Bacillus Calmette-Guérin Revaccination Questionable with Low Tuberculosis Incidence. Am. J. Respir. Crit. Care Med. 1998, 157, 1324–1327. [Google Scholar] [CrossRef]
- Rahman, M.; Sekimoto, M.; Hira, K.; Koyama, H.; Imanaka, Y.; Fukui, T. Is Bacillus Calmette-Guerin revaccination necessary for Japanese children? Prev Med. 2002, 35, 70–77. [Google Scholar] [CrossRef]
Figure 1.
Vaccination regimen schedules.
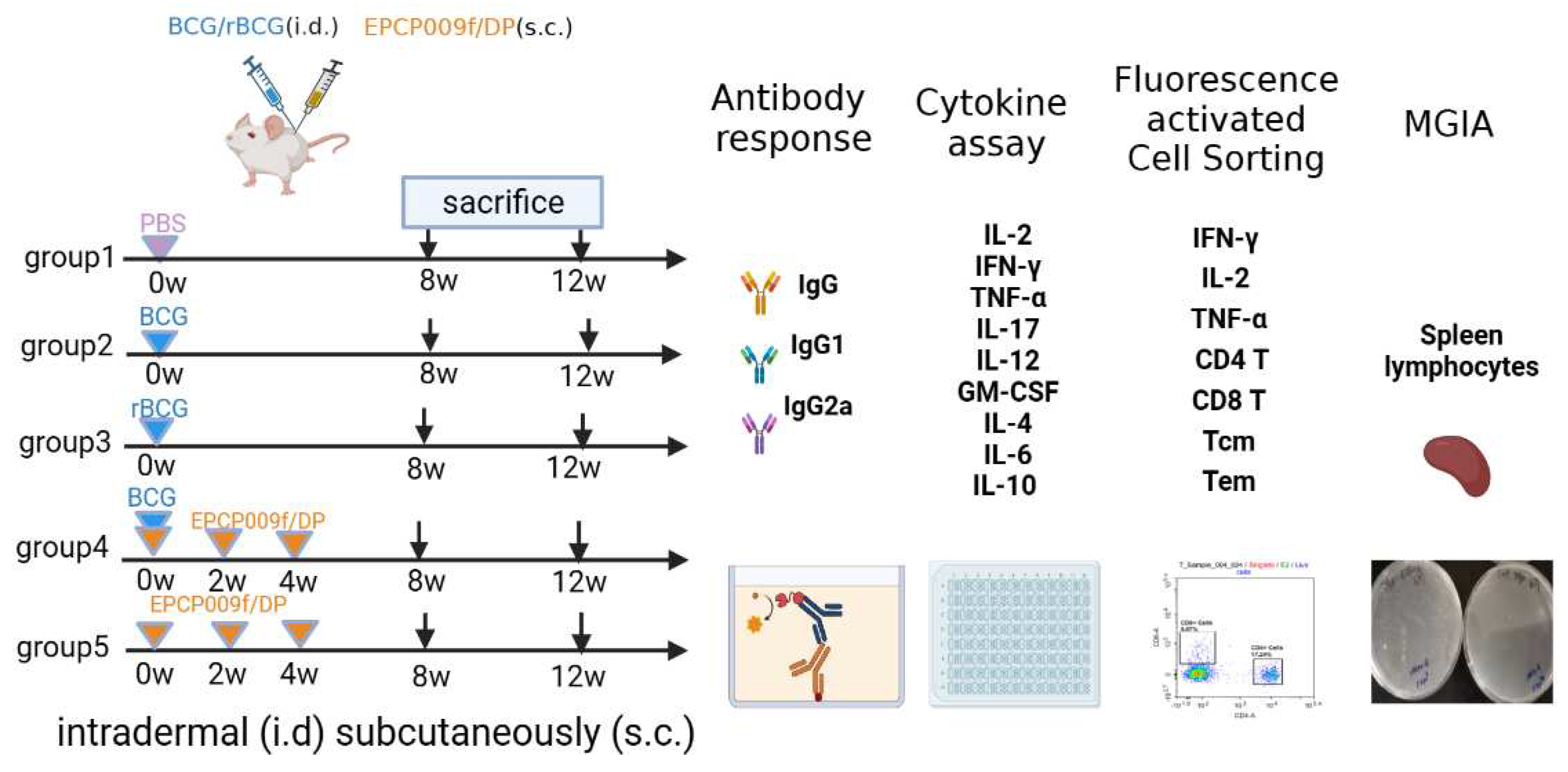
Figure 2.
Verification of rBCG-EPCP009 and the fusion protein EPCP009. (A) Schematic diagram of the recombinant plasmid pMV361-001. (B) Verification of the rBCG-EPCP009 gene. Lane M, standard DNA marker; lane 1, genomic PCR product of BCG; lane 2, genomic PCR product of rBCG-EPCP009. (C) mRNA levels of nPPE18 and nPstS1 in rBCG-EPCP009, as determined by qRT-PCR. cDNA levels were normalized to the endogenous RNA levels of the internal control 16s. The fold increase in gene expression was calculated using the 2−ΔΔCT method. (D) Lane M, molecular weight marker of the pre-stained protein; lane 1, EPCP009.
Figure 2.
Verification of rBCG-EPCP009 and the fusion protein EPCP009. (A) Schematic diagram of the recombinant plasmid pMV361-001. (B) Verification of the rBCG-EPCP009 gene. Lane M, standard DNA marker; lane 1, genomic PCR product of BCG; lane 2, genomic PCR product of rBCG-EPCP009. (C) mRNA levels of nPPE18 and nPstS1 in rBCG-EPCP009, as determined by qRT-PCR. cDNA levels were normalized to the endogenous RNA levels of the internal control 16s. The fold increase in gene expression was calculated using the 2−ΔΔCT method. (D) Lane M, molecular weight marker of the pre-stained protein; lane 1, EPCP009.
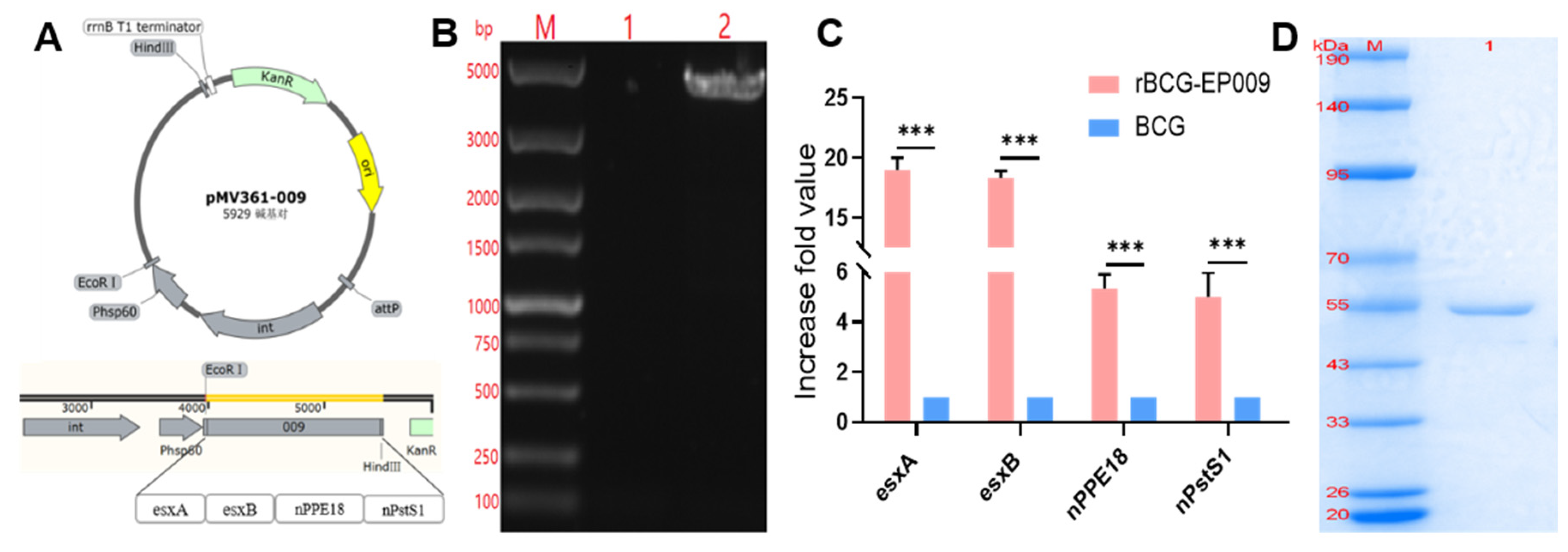
Figure 3.
Levels of cytokines secreted by mouse spleen lymphocytes 8 weeks after immunization with the candidate vaccines. Cytokine levels were examined after stimulation with PPD (A) and EPCP009 (B) after 8 weeks of initial immunization with the different vaccine candidates. The p values indicating the significance of differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
Figure 3.
Levels of cytokines secreted by mouse spleen lymphocytes 8 weeks after immunization with the candidate vaccines. Cytokine levels were examined after stimulation with PPD (A) and EPCP009 (B) after 8 weeks of initial immunization with the different vaccine candidates. The p values indicating the significance of differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).

Figure 4.
Levels of cytokines secreted by mouse spleen lymphocytes 12 weeks after immunization with the vaccine candidates. Cytokine levels secreted on stimulation with PPD-specific proteins (A) or EPCP009-specific proteins (B) were measured after 12 weeks of initial immunization with the different vaccine candidates. The p-values indicating the significance of differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
Figure 4.
Levels of cytokines secreted by mouse spleen lymphocytes 12 weeks after immunization with the vaccine candidates. Cytokine levels secreted on stimulation with PPD-specific proteins (A) or EPCP009-specific proteins (B) were measured after 12 weeks of initial immunization with the different vaccine candidates. The p-values indicating the significance of differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
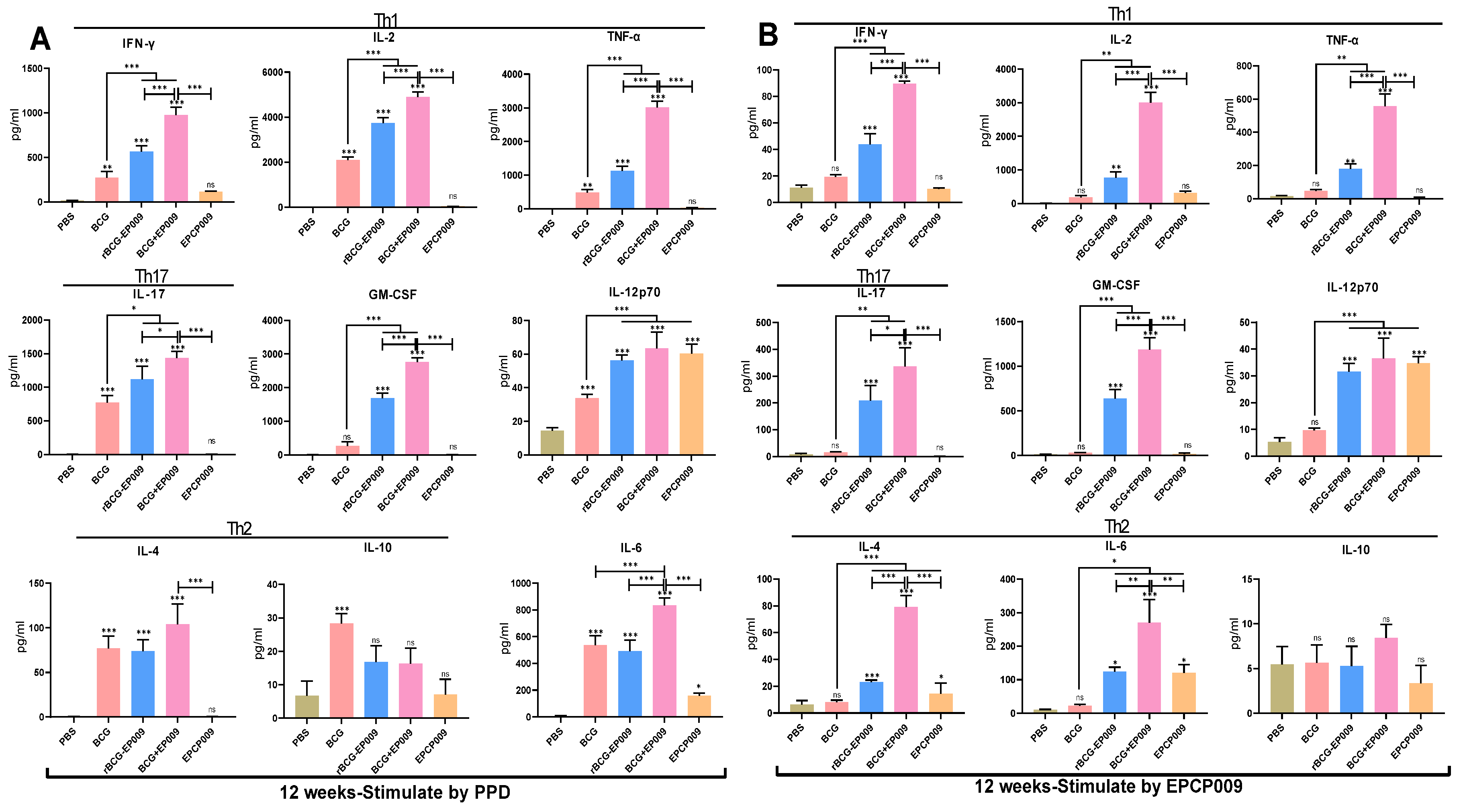
Figure 5.
Antigen-specific IFN-γ and IL-4 levels secreted by mouse splenic lymphocytes. The IFN-γ and IL-4 levels were determined at 8 (A) or 12 (B) weeks after the first immunization. The proportion of T cells secreting IFN-γ and IL-4 was determined with the ELISpot assay. The p values indicating the significance of the differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
Figure 5.
Antigen-specific IFN-γ and IL-4 levels secreted by mouse splenic lymphocytes. The IFN-γ and IL-4 levels were determined at 8 (A) or 12 (B) weeks after the first immunization. The proportion of T cells secreting IFN-γ and IL-4 was determined with the ELISpot assay. The p values indicating the significance of the differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
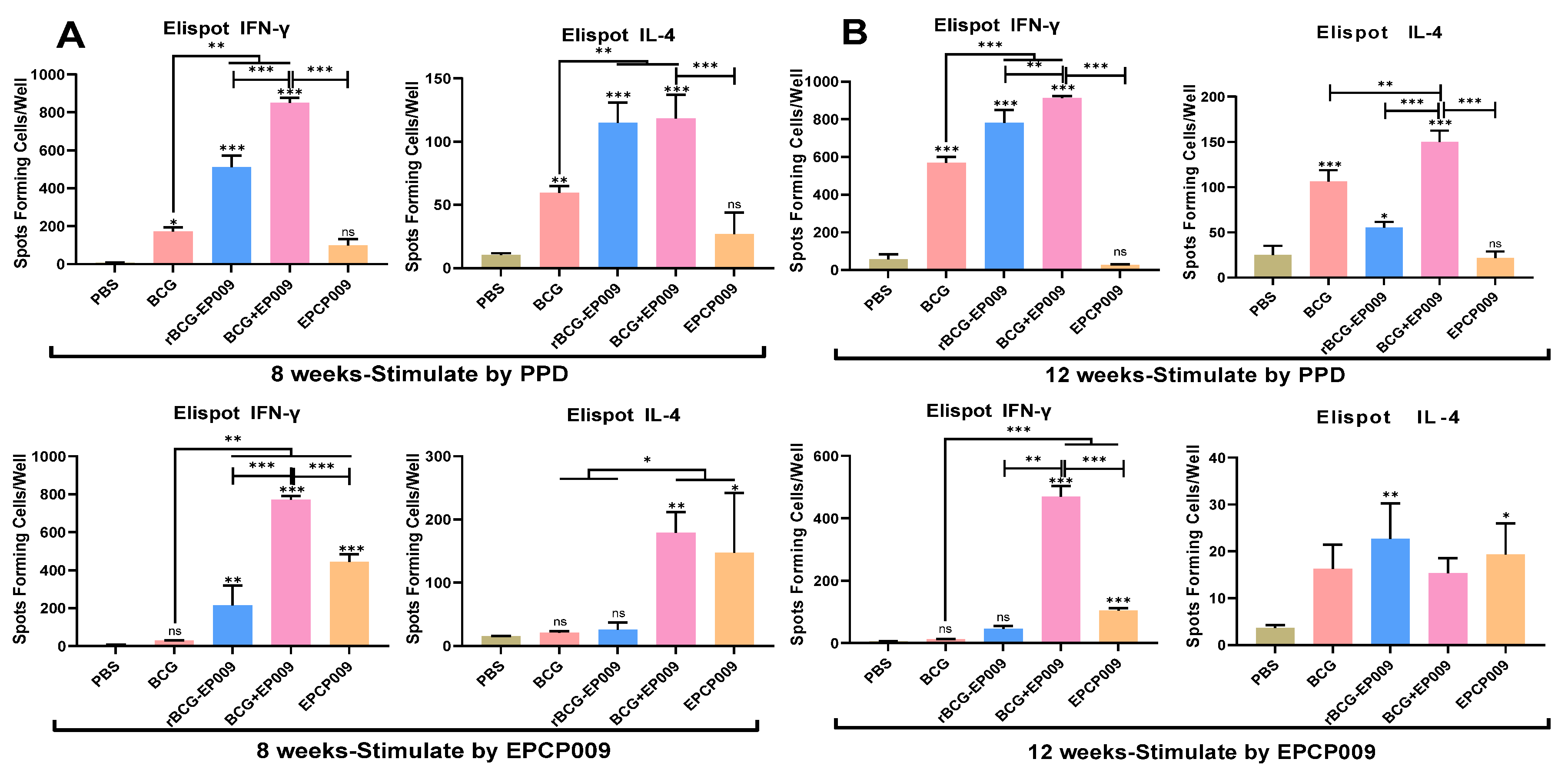
Figure 6.
Antibody response in Balb/c mice immunized with different vaccine candidates at 8 (A) or 12 (B) weeks after the initial immunization.
Figure 6.
Antibody response in Balb/c mice immunized with different vaccine candidates at 8 (A) or 12 (B) weeks after the initial immunization.
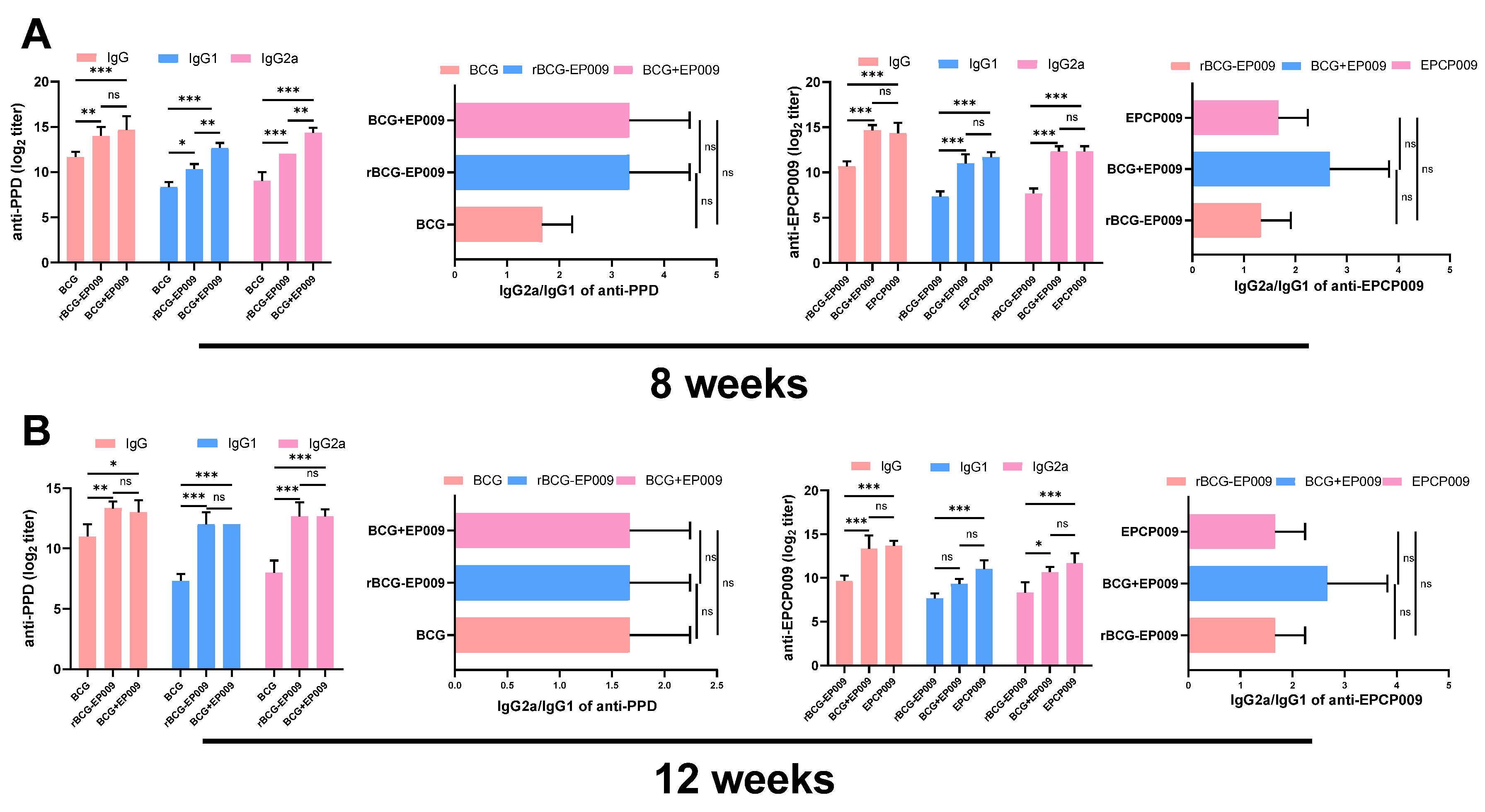
Figure 7.
Proportions of various immune cells in spleen cells from immunized mice.
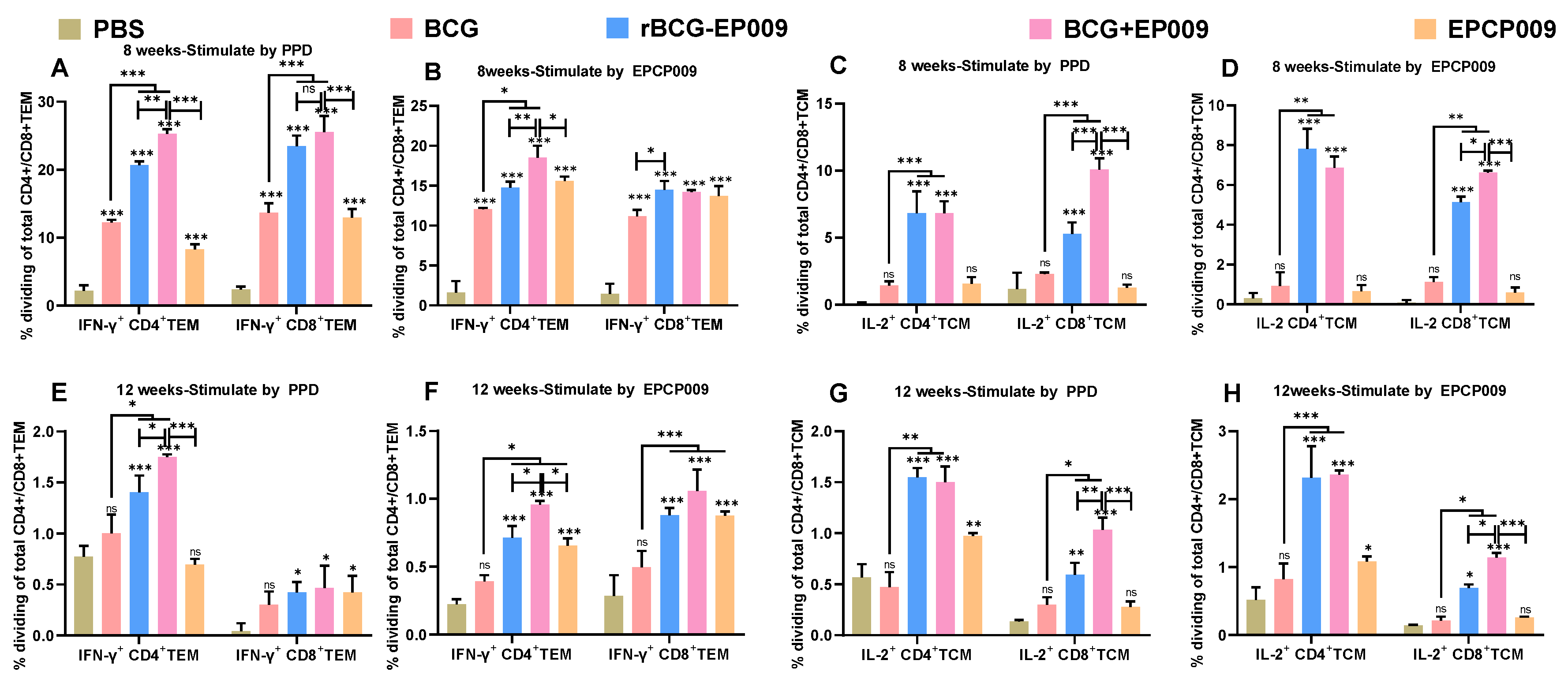
Figure 8.
In vitro growth inhibition of M. tuberculosis H37Rv with splenocytes isolated from Balb/c mice after 8 and 12 weeks of immunization. The p-values of the differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
Figure 8.
In vitro growth inhibition of M. tuberculosis H37Rv with splenocytes isolated from Balb/c mice after 8 and 12 weeks of immunization. The p-values of the differences were determined by one-way ANOVA with Tukey’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001).
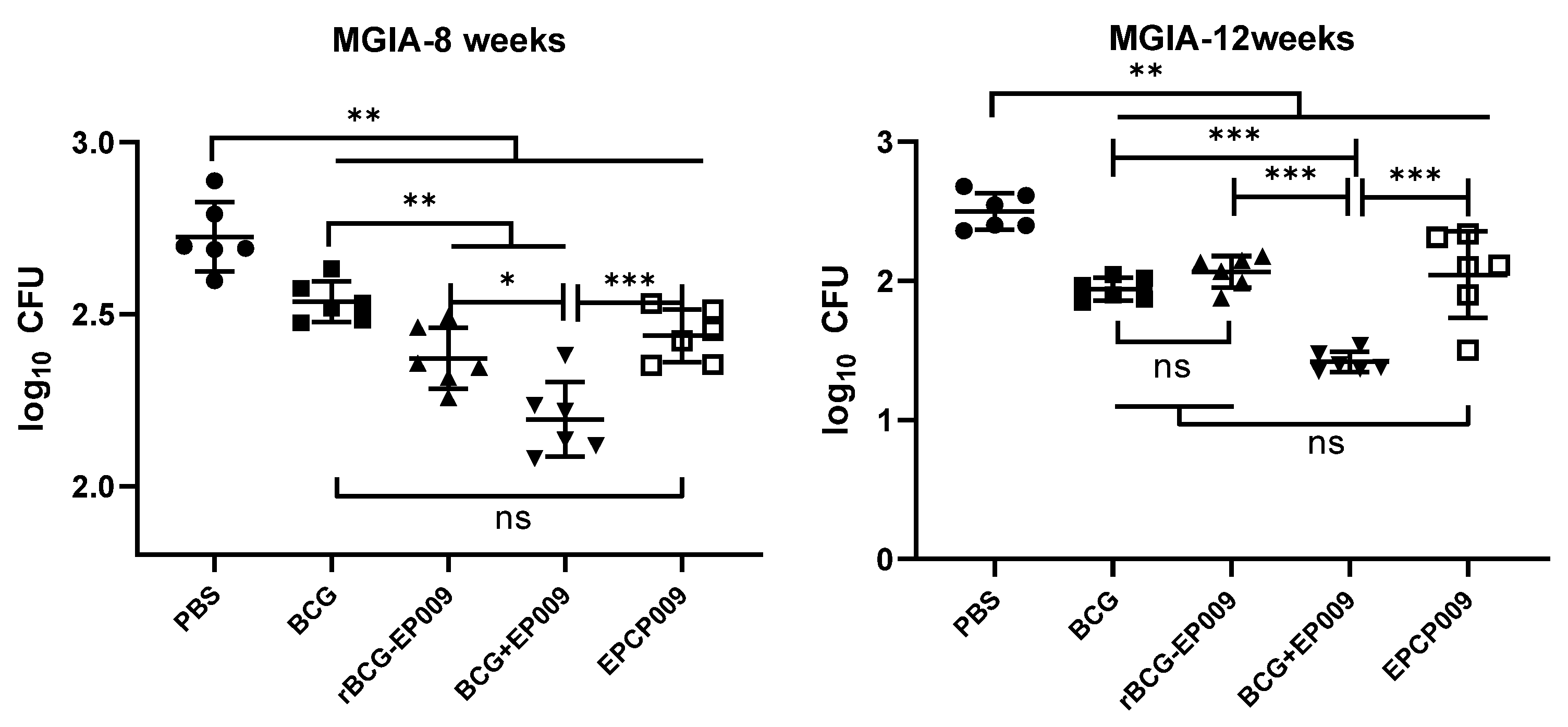

Table 1.
Primers for validation of recombinant BCG.
| Primers | Primer sequences (5′ to 3′) |
|---|---|
| Integration validation primer F | CGGCTTATCAACTAGATCGGCGCAG |
| Integration validation primer R | GACGTCAGGTGGCTAGCTGATCA |
| esxA RT-qPCR primer F | TGACAGAGCAGCAGTGGAATTTCG |
| esxA RT-qPCR primer R | CAAGGAGGGAATGAATGGACGTGAC |
| esxB RT-qPCR primer F | AGCCAATAAGCAGAAGCAGGAACTC |
| esxB RT-qPCR primer R | CTAGAAGCCCATTTGCGAGGACAG |
| nPPE18 RT-qPCR primer F | TGTCGATGACCAACACCTTGAGC |
|
nPPE18 RT-qPCR primer R nPstS1 RT-qPCR primer F nPstS1 RT-qPCR primer R 16s RT-qPCR primer F 16s RT-qPCR primer R |
CCAGAACCACCACCCGAAGAAC CGCCTATCTGTCGGAAGGTGATATG GTTGACCTGCTGAGCGGAGATG CGCACAAGCGGCGGAGCA GCCACAAGGGAACGCCTATCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



