
Submitted:
12 October 2023
Posted:
12 October 2023
You are already at the latest version
Abstract
The continued exploration of the diversity of lactic acid bacteria in little-studied ecological niches represents a fundamental activity to understand the diffusion and biotechnological significance of this heterogeneous class of prokaryotes. In this study, Lactiplantibacillus plantarum (Lpb. plantarum) strains have been isolated from Tunisian vegetable sources, including fermented olive, fermented pepper and from dead locust intestines, subsequently evaluated for their antimicrobial activity against foodborne pathogenic bacteria, including Escherichia coli O157:H7 CECT 4267 and Listeria monocytogenes CECT 4031, as well as against some fungi, including Penicillium expansum, Aspergilus niger and Botrytis cinerea. In addition, their resistance to the oro-gastro-intestinal transit, aggregation capabilities, biofilm production capacity, adhesion to human enterocyte-like cells and cytotoxicity to colorectal adenocarcinoma cell line were determined. Further, the adhesion to tomatoes and the biocontrol potential on this model food matrix were analyzed. It has been found that all the strains were able to inhibit the indicator growth, mostly through organic acids production. Besides, these strains showed promising probiotic traits, including in vitro tolerance to oro-gastrointestinal conditions, adhesion to abiotic surface and to Caco-2 cells. Moreover, all tested Lpb. plantarum strains were able to adhere to tomatoes with similar rate (4.0-6.0 LogCFU/g tomato). The co-culture of LAB strains with pathogens on tomatoes showed that Lpb. plantarum could be a good candidate to control pathogen growth. Nonetheless, further studies are needed to guarantee their use as probiotic strains for biocontrol on food matrices.
Keywords:
Lactic acid bacteria
; probiotic
; biocontrol
; niches
; Tunisian
; tomato
1. Introduction
Lactic acid bacteria (LAB) are a group of Gram-positive, non-spore-forming, cocci or rods, catalase-negative microorganisms [1] widely used in fermentation, probiotics, and food/beverage manufacturing industries. LAB are Generally Regarded as Safe (GRAS) by the United States Food and Drug Administration (FDA), and boast the Qualified Presumption of Safety (QPS), as assessed by the European Food Safety Authority (EFSA) [2]. Moreover, LAB have been widely used to improve the taste, texture, and nutritional properties of a wide variety of foods, including vegetables, meat, dairy and cereal substrates [3], and can extend their shelf life by producing organic acids, carbon dioxide, and antimicrobial peptides [4].
In the food industry, LAB have become increasingly important, and many studies have been focused on the possible use of bacteriocin-producing species as alternatives to chemical preservatives in foods [5]. Indeed, certain LAB strains exhibit antimicrobial activity against foodborne pathogens, including bacteria, yeast and filamentous fungi. Furthermore, in recent years, many authors proved that LAB have the ability to neutralize several undesired microbes, including Clostridium spp., Enterococcus faecalis, Listeria monocytogenes [6].
Thanks to their fermentation ability, LAB are employed in manufacturing many foods, such as dairy products, sausages, cucumber pickles and olives [7]. Indeed, LAB can be divided into starter cultures, used to drive biochemical changes of primary relevance for food fermentation, and non-starter cultures, usually deriving from the autochthonous microbiota of the food matrix [8] and contributing in secondary aspects to the organoleptic characteristics of the final product [9]. The antimicrobial potential of non-starter LAB isolated from fermented foods has been substantiated by scientific literature and involves the production of organic acids, hydrogen peroxide and bacteriocins.
Besides antimicrobial and fermentation capacities, some LAB also exhibit probiotic features. According to the United Nations of Food and Agriculture Organization and the World Health Organization (FAO/WHO), probiotics are “live microorganisms which, when administered in adequate amounts, confer health benefits on the host” [10]. It has been confirmed that the consumption of probiotics can reduce cholesterol serum levels, prevent diarrhoea through the enhancement of the intestinal barrier, and decrease irritable bowel syndrome symptoms [11]. To be applied as a probiotic and to function appropriately, a microbial strain must exhibit several characteristics, such as gastrointestinal tolerance and the ability to colonise the human host. Furthermore, the probiotic candidate should be safe for humans, e.g., with an antibiotic sensitivity phenotype [12].
Lactiplantibacillus plantarum (Lpb. plantarum ) is a LAB with a long history of protechnological use in the food sector, in food fermentations and for the design of protective cultures [13,14]. This species can adapt to a variety of niches and is widely distributed in the environment, i.e., it can be found in dairy products, in the gastrointestinal tracts of humans and animals, meat, fish, and fermented vegetables [15]. Increasing evidence also corroborates the probiotic properties of Lpb. plantarum strains [16,17], thus broadening the range of its applications. To date, most studies underline the safety attributes of Lpb. plantarum [18,19], supporting the industrial interest in this species. Recent papers also highlight some Lpb. plantarum properties that make it intriguing for biomedical purposes, e.g., its capacity to regulate the enteric microbiota, alleviate liver disease [20], and its anticancer potential [20]. Moreover, some publications hint at the use of Lpb. plantarum for the bio-suppression of pathogens in food models [21,22].
Fruit and vegetables are important components of a healthy diet, and their consumption helps prevent a wide range of diseases [23]. Thus, both WHO and FAO recommend the intake of a specific dose of vegetables and fruits per day. Tomato is highly consumed worldwide; it is economically considered the second-most important fruit or crop [24]. In 2020, Tunisia ranked tenth in the world in producing tomatoes. The production of tomatoes was estimated at around 1.4 million metric tons in the last four years by Statista Research Department [25]. Tomato contains a high amount of fiber, oligosaccharides and polysaccharides, which act as prebiotics in the gut [26]. However, tomatoes can be easily contaminated and get spoilt very fast through contaminated irrigation water during transporting and storage, causing economic losses and serious health issues related to foodborne diseases [27]. Consequently, there is a need for intervention technologies and techniques to reduce/prevent tomato contamination [28]. To face food contamination/spoilage, several methodologies have been investigated and applied over the years; however, these methods are not applicable to all foods and can alter the sensory properties of the final product [29,30]. Therefore, bio-antimicrobial agents, such as antagonistic bacteria, have emerged as alternatives recently.
Overall, the aims of the present study were to isolate new LAB strains from unexplored Tunisian sources, to evaluate their capability to colonize tomato surfaces and to inhibit the adhesion of pathogenic Escherichia coli O157:H7 CECT 4267 and Listeria monocytogenes CECT 4031 through co-culture on the food matrix. Further, their potential use for the design of probiotic cultures was investigated.
2. Materials and Methods
2.1. Isolation of LAB and preliminary screening for antimicrobial activity
LAB strains used in this study were isolated in Tunisia from a wide variety of niches, including different types of home-made traditional food, fermented olive, fermented pepper and intestines of dead locust Solitaria (Tettigonia viridissima) (see below (Table 1)). The samples were serially diluted in physiologic solution and plated on De Man–Rogosa–Sharpe (MRS) incubated at 37 °C. Gram staining, catalase tests and microscope examination of the cell morphology achieved phenotypic identification. Growth in MRS broth and pH variation were also evaluated. The purified LAB isolates were stored in glycerol 20 % (v/v) at -80 °C.
The LAB isolates were screened for their antibacterial activity against foodborne pathogens, such as Escherichia coli O157:H7 CECT 4267 and Listeria monocytogenes CECT 4031, using the modified agar well method [31]. Briefly, wells were created in Luria-Bertani medium (LB) and 24 h bacterial cultures from the isolated microbes were poured into each well. After 24 h of incubation at 37 °C, LAB strains were discriminated on the basis of inhibition halos around the wells. Six selected strains were further investigated for their biochemical features and stress response in various conditions (e.g acidic condition, osmotic stress [32]), and for some probiotic features as follows.
Enterococcus faecalis CECT 795 [33], Lactobacillus reuteri DSM 17938 [34], Enterococcus faecalis V583 [35,36] and Lpb. plantarum 299V [37] were used as control strains for the hemolytic assay, oro-gastro-intestinal transit tolerance assay, biofilm formation assay and for the adhesion to human enterocyte-like cells, respectively. Lpb. plantarum 299V and L. reuteri DSM 17938 were routinely cultivated in MRS medium at 37°C, while E. faecalis CECT 795 and E. faecalis V583 were cultured in Brain heat infusion (BHI) at 37°C.
2.2. Molecular identification of selected isolates
The six isolates with the best antibacterial performances and from unusual sources were selected for further characterisation and identified using sequencing of the 16S rRNA gene. The genomic DNA of the strains was purified using a genomic DNA extraction kit (Mericon, Hilden, Germany), following the manufacturer’s instructions. The genomic DNA was used as a template to amplify and sequence 16S rDNA, resulting in species identification (OR431596-OR431601).
2.3. Antimicrobial activity and partial-characterisation of the antimicrobial agent
The selected LAB strains were screened for antagonistic activity by agar diffusion method (well method)[38] as described above (part 2.1) using L. monocytogenes and E. coli as indicators. To determine the antifungal spectrum of the selected LAB isolates, the overlay method was performed, according to Russo et al. [39], using Penicillium expansum CECT 2278, Aspergilus niger CECT 2805, Fusarium culmorum CECT 2148, Saccharomyces cerevisiae and Botrytis cinerea CECT 20973 as targets. At the late exponential phase, 5 µL of each LAB culture were spotted on MRS agar plates and incubated at 30 °C for 24 h. Then, plates were overlaid with 10 mL of Malt Extract (Oxoid) Soft Agar (0.75% agar) inoculated (1:100 v/v) with a suspension containing approximately 1 × 106 spores/mL of each fungal species.
Regarding the partial-characterisation of the antibacterial agents: LAB culture, crude cell-free supernatant (CFS), neutralized CFS (pH=6.0), heat-treated CFS (121°C 10min), crude cells and heated cells were prepared as follows. CFS obtained by filtration (0.24 µm) of 24 h LAB cultures were neutralised with 1 M NaOH to pH=6.0 (to eliminate the antibacterial effect of organic acids) and subsequently heat-treated (121 °C, 10 min) (neutralize thermosensitive proteins)[40]. LAB cells obtained after centrifugation (10,000 × g, 5 min) were washed twice with sterile PBS pH=7 and resuspended in the same volume of previous LAB culture in PBS pH=7, later cells were treated by heat (121 °C, 10 min). After 24h of incubation at 37 °C, the halos of inhibition around the wells were measured. The organic acids of the CFSs were quantified by HPLC Spectra System P1000 XR (Thermo Electron Corporation, Madison, WI, US) [41].
2.4. Safety features: Antibiotic resistance and hemolytic activity
The antibiotic susceptibility of lactobacilli strains was determined by the disc diffusion assay according to CLSI guidelines [42]. In brief, antibiotics, i.e., ampicillin (10 μg), oxacilline (1 μg), amikacin (30 μg), gentamicin (10 μg), tetracycline (30 μg), chloramphenicol (30 μg), and clindamycin (2 μg), were placed on the surface of plates and incubated at 37 °C for 24 h. The inhibition zone diameters were measured, and susceptibility was expressed in terms of Resistant (R), Susceptible (S) and Intermediate (I) mentioned [43].
The hemolytic activity was assessed by streaking fresh culture of LAB on Columbia 5% blood agar plates, after incubation for 24 to 48 h at 37 °C [7]. Strains showing a transparent halo or a green-hued halo were considered hemolytic and classified as β-hemolytic and α-hemolytic, respectively, whereas those presenting no halo were considered non-hemolytic and were classified as γ-hemolytic. Enterococcus faecalis CECT 795 was used as a negative control (γ-hemolytic).
2.5. Autoaggregation and Co-aggregation assays
To determine auto-aggregation, we followed previously described procedures with some modifications [44]. Briefly, 5 mL of bacterial suspension were vortexed for 10 s and then, the suspension was incubated for 4 h at 37 °C. The absorbance of the supernatant after 2 h of incubation was subsequently measured (ODt) starting from t0, and the auto-aggregation percentage was calculated as
A= [1 – (ODt/ODt0)] × 100.
For the co-aggregation assay [45], 4 mL of each LAB strain and 4 mL of each pathogen culture (E.coli or L. monocytogenes) were mixed, vortexed for 10 s, and incubated for 4 h at 37 °C. Each control tube contained 4 mL of each single bacterial suspension (i.e., the LAB strain or the pathogen). The absorbance of each mixed suspension was then measured at 600 nm (ODmix) and compared with those of the control tubes containing the LAB strain (ODstrain) and the specific pathogen (ODpathogen) at 2 h of incubation.
Co-aggregation (%) was calculated as C= [1 − ODmix/ (ODstrain + ODpathogen)/2]× 100.
2.6. In vitro oro-gastro-intestinal transit tolerance assay
The oro-gastrointestinal (OGI) transit tolerance assay was performed according to Gheziel et al. [46], with some modifications (Figure 1). Briefly, LAB strains were grown until mid-exponential phase(OD600nm=1), then centrifuged (10,000 × g, 5 min) and resuspended into sterile electrolyte solution (6.2 g/L NaCl, 2.2 g/L KCl, 0.22 g/L CaCl2, 1.2 g/L NaHCO3). To simulate the saliva condition in vitro, bacterial suspensions were incubated at 37 °C for 5 min, with 150 mg/L lysozyme at pH=6.0. Then, the culture was centrifuged in order to remove the first solution and to simulate the gastric stress by adding the second solution (pH=3) containing pepsin 3g/L; later, the second solution was removed by a second centrifugation (10,000 × g, 5 min) and the pH were decreased to 2.0 by a third solution and pepsin 3g/L was added and incubated for 30 min at 37°C. Finally, the intestinal stress was simulated by the fourth solution (pH=6.5) containing 3g/L bile salts and 1g/L pancreatin. Finally, samples were diluted (1:1 v/v) with a sterile electrolyte solution to mimic the large intestine and incubated for 1 h at 37 °C. The relative viability was calculated by spreading on MRS agar serial dilutions from bacterial samples at the different steps of the simulated transit. The survival rate was calculated by comparing the colony-forming units (CFU) from control and OGI-stressed bacterial samples. The survival rate was expressed as LOG (CFUt(1–5)/CFUt0). Results were further compared to that of Lactobacillus reuteri DSM 17938, a probiotic strain used as a positive control for this experiment.
2.7. Biofilm formation assay
The production of biofilms was evaluated on 96-well polystyrene microtiter plates [47]. First, the wells were filled with 200 µl of MRS medium, the absorbance of bacterial suspensions in Maximum Recovery Diluent (MRD) (Liofilchem) was adjusted to 0.5 MacFarland in order to standardise the number of bacteria (107–108 CFU/ml), and 1% of overnight culture was added to each well. The plates were incubated for 24, 48 and 72 h at 37°C. To quantify the biofilm formation, the wells content was carefully removed, and 100 µl of a 0.1% (v/v) crystal violet solution was added to each well and held at ambient temperature for 15 min; then, the wells were gently washed three times with MRD. Then, the dye bound to adherent cells was removed with 100 µl acetone/ethanol mixture (80:20), and absorbance was measured at 595 nm.
In order to detect/estimate the viability of LAB in the wells, Resazurin was used [48]. Briefly, after 3 washes with MDR of 96-well plate cultures, the plates were dried under the laminar flow chamber for 1 hour, and then 100 µl of Resazurin solution freshly prepared (8 µg/ml) were added. The optical density (OD) of each well was measured at 595 nm using a microplate reader (SUNRISE, Serial number 1708003498, XFLUOR4 Version V 4.51). Enterococcus faecalis V583, growing in Brain heart infusion medium (BHI) was used as positive control for biofilm production. Each assay was performed in three replicates and conducted three individual times on different days under the same conditions, and the negative control was performed in uninoculated MRS broth and BHI. The cut-off (ODC) was defined as the mean OD value of the negative control. Based on the OD, strains were classified as non-biofilm producers (OD ≤ ODC), weak (ODC < OD ≤ 2 × ODC), moderate (2 × ODC < OD ≤ 4 × ODC) or strong biofilm producers (4 × ODC < OD) [49].
2.8. Cytotoxic effects of Lactiplantibacillus plantarum strains
All LAB strains were grown in 100 ml MRS broth for 24 h (reaching stationary phase) at 37 °C. In order to obtain CFS (Cell-Free Supernatant), we followed the same protocol indicated in (part 2.3) and then lyophilised. The lyophilised samples were dissolved in sterile distilled water to attain the desired concentrations, from 0.5 mg/ml up to 10 mg/ml.
For cytotoxicity assay, the human colorectal adenocarcinoma LS cells were used [50]. 50 µl of containing ten thousand cells were seeded per well in 96-well plates containing RPMI (Gibco™, Cergy-Pontoise, France; Sigma, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS), 1% L-glutamine and 100 IU/mL penicillin/streptomycin and incubated for 24 h for cells to adhere properly.
After 24 h, 100 μl of fresh medium with different concentrations of lyophilised supernatant was added and incubated for 24 h and for 72h. The medium was removed and 50 µl of MTT (0.5 mg/ml) was added in all wells. After 2 h of incubation at 37 °C, 100 µl of dimethylsulfoside (DMSO) was added, and absorbance was read. The optical density was measured at 560 nm. The assay was repeated at least in three independent experiments in triplicate. The percentage of cell viability was calculated using the following equation: Asample - Ab / Acontrol *100 where the Asample represents the absorbance of treated cells with CFS, Ab represents the absorbance of cells medium (DMEM) with CFS, and Acontrol represents the absorbance of the cells with LAB medium (MRS). The concentration for 50% of growth (IC50) was determined.
2.9. Adhesion to human enterocyte-like cells
The adhesion capacity was investigated using human Caco-2 cell lines as previously described [51]. Cells were grown in a controlled atmosphere of 5% CO2 at 37 °C in Dulbecco’s Modified Eagle’s Minimal Essential Medium (DMEM) (Gibco. Carlsbad, CA, USA), supplemented with 2 mM L-glutamine, 50 U mL−1 penicillin, 50 U mL−1 streptomycin, and 10% (v/v) heat-inactivated fetal bovine serum (FBS). Caco-2 (2 × 105 cells per well) were seeded in 96-well cell culture plates, grown for 2 weeks, changing the medium three times a week, up to get steady monolayers. Twenthy-four hours prior to the adhesion assay, the growth medium was replaced with absolute DMEM. From each bacterial strain, 100uL of mid-exponential phase cultures (OD600nm = 0.6–0.8, corresponding to 5 × 108 CFU mL−1) were centrifuged, after that washed with sterile PBS, resuspended in the same volume of absolute DMEM and finally incubated with Caco-2 cells in a ratio of 1000:1 (bacteria:Caco-2 cells). After 1 h of incubation at 37 °C (5% CO2), Caco-2 monolayers were washed 3 times with PBS to remove non-adherent bacteria and then detached by incubation at 37 °C for 10 min in the presence of trypsin/EDTA (0,05%, Sigma). Trypsin-treated samples were resuspended in PBS, serially diluted and plated onto MRS agar to enumerate the adherent bacteria. CFU obtained from washed wells (cell-adhering bacteria only) were compared with those obtained by trypsinisation from control unwashed wells (total bacteria including adhering and non-adhering ones), in order to calculate the adhesion percentages [(CFU)washed well/(CFU)unwashed well] × 100. The commercial Lpb. plantarum 299V strain was used as a positive control. Adhesion assays were Lpb. plantarum 299V conducted at least in three independent experiments in triplicate.
2.10. Application in food model
2.10.1. Adhesion on tomatoes surface
The adhesion assay was performed on whole grape tomatoes (Solanum lycopersicum L.) as represented in the figure below (Figure 2) [52]; fresh tomatoes were purchased from a local supermarket in Italy the day before the experiment and kept in the refrigerator overnight at 4 °C. On the day of the experiment, pericarp-free tomatoes without visible defects, such as bruises and injuries, were selected, washed with distilled water, placed inside the laminar flow cabinet, and irradiated with UV light for 15 min before being inoculated.
Fresh LAB cultures were washed twice with PBS buffer pH=7 and were adjusted to a final concentration of 1x108 cells/mL by resuspending the pellets in PBS to an OD of 0.5 at 600 nm. Fifty microliters of culture suspension were carefully spotted on the stem scar sites of grape tomatoes at ambient air temperature. The inoculated tomatoes were kept in the laminar flow cabinet for 2 h to allow bacterial attachment to the tomato scar. For microbial enumeration [52], tomato stem scars were aseptically excised from the fruit and combined with physiological water 0.85% in 1/8 ratio (weight/volume), and carefully mixed using vortex for 2 min. After that, serial decimal dilutions of the homogenate were prepared, inoculated in MRS plates (in triplicate) and incubated for 24h at 37°C. For the control, PBS buffer pH=7.0 was used for tomato inoculation, as mentioned above.
2.10.2. Biocontrol/pathogen antagonism on tomatoes
The antagonism assay was performed as described above for LAB adhesion to tomato surface by co-inoculating LAB strains and Escherichia coli O157:H7 CECT 4267 or Listeria monocytogenes CECT 4031. Each LAB was mixed with the corresponding pathogen and inoculated simultaneously on the tomatoes pericarp by spotting them in sterile conditions. The mix of LAB culture and pathogen was prepared previously, as mentioned [53]: each culture was adjusted to a final cell concentration of 109-108 /ml, then washed twice by suspending the pellet in PBS to an optical density of 0.5 for LAB and 0.2 for the pathogens at 600nm. After inoculation, the fruits were kept in the laminar flow cabinet for 2 h to allow the bacterial attachment to the tomato stem scar and, then, stored in sterile bags at room temperature.
For microbial quantification MRS agar, Sorbitol McConkey Agar (SMAC) and Polymyxin Acriflavine Aithium Chloride ceftazidime Aesculin Mannitol agar (PALCAM) were used. Controls inoculated with a single bacterial culture in the corresponding medium were also analyzed.
2.11. Statistical Analysis
All experiments were performed in duplicate or triplicate and carried on in a completely randomized design. Statistical analysis was performed using GraphPad Prism software. One-way ANOVA was used for comparing data with one factor between groups; two-way ANOVA was used for comparing more than one factor between groups, and differences were adjudicated using the post hoc analysis recommended by Prism.
3. Results
3.1. Isolation and characterization of LAB
After enrichment of microbes from diverse matrices, a total of 40 isolates grown on MRS agar, were randomly selected and screened for antibacterial activity against E. coli O157:H7 CECT 4267 and L. monocytogenes CECT 4031. All isolates showed typical morphological characteristics, being Gram-positive bacilli and cocci; all the isolates were catalase-negative and non-motile. All of them showed the capacity to decrease the pH of the medium to 3.0, with tolerance to acidic and basic conditions (2.5 to 9). Out of the isolated microorganisms, only 24 isolates showed various levels of inhibition, from modest (ranging from 7 mm to 10 mm) to good (more than 10 mm) anti-Listeria and anti-E. coli activity (Table 1). Of these, only six strains with the best antibacterial activity were selected for further studies. Molecular analysis revealed that all the selected strains belonged to Lactiplantibacillus plantarum with a percentage of homology higher than 98% (Table 2).
The growth of the six selected strains was also evaluated in different conditions. The tolerance to acidic (pH=4.2) and alkali (pH=9.2) pH was evaluated and at different concentrations of salts NaCl (2%, 4% and 8%, w/v). The ability to grow in MRS medium with different types and different proportions of sugar was also evaluated, e.g., glucose, fructose, and sucrose in 2% and 4%, respectively. All the strains were viable under all of the conditions tested and can survive at different pH values, as well as tolerate up to 8% NaCl (Table 3).
3.2. Antimicrobial activity and some insights into the antibacterial agents
All the selected strains showed antimicrobial activity against the bacterial indicators, with various inhibition zones ranging between 11 mm and 15 mm (Table 1). Four of them, namely 1nm, S4, pepp1, and pepp2, demonstrated a higher antimicrobial activity against E. coli. In order to determine the nature of the antibacterial substance, the CFS of each LAB strain was collected after 24h, and the antibacterial activity of the crude CFS was compared with that of the corresponding crude cells, evidencing that the inhibitory activity depended on secreted substances (Supplementary Figure S1). For the 6 strains, neutralising the CFSs with NaOH abolished the antimicrobial activity, with little or no impact on the growth of indicator bacteria, thus suggesting that the inhibitory effects were due to organic acids (Table 4). Moreover, heat treatment of CFS did not suppress their antimicrobial effect, indicating the presence of thermostable antimicrobial substances. Crude cells and heat-treated cells did not exhibit any antimicrobial effect, confirming the external secretion of the antibacterial agent and not attached to cell membrane.
Quantification of the organic acids contents in the CFS from the selected Lpb. plantarum strains is reported in Table 5. All the strains were able to produce various organic acids, and lactic acid was the most abundant, with a concentration above 13g/L. All the strains, except RSOLV and pepp2, were able to produce ascorbic acid. Only the strain 3nm was able to produce fumaric acid with a quantity of 15.47±6.71 mg/L. These findings confirm the heterofermentative aspect of Lpb. plantarum strains and suggest that the combination of organic acids could be the reason for the observed antimicrobial activity.
Antifungal activities of the six LAB strains against P. expansum CECT 2278, A. niger CECT 2805, F. culmorum CECT 2148, S. cerevisiae and B. cinerea CECT 20973 were assessed, and all the strains were active against the 4 indicator fungal species. All the strains were able to inhibit P. expansum and F. culmorum, with inhibition halos ranging between 4 mm to 8 mm (Table 6). Only two strains (RSOLV and pepp2) showed the best antagonistic effect toward B. cinerea. Only one strain, namely RSOLV was weakly active against A. niger compared to the antifungal activity of the others (Supplementary Figure S2). None of the LAB strains tested in the study showed inhibitory activity against S. cerevisiae.
3.3. Auto-aggregation and co-aggregation assays
Auto-aggregation and co-aggregation abilities are related to the adherence capability to intestinal epithelial cells. Moreover, it is an important phenotypic trait for screening potential probiotic strains. Results obtained showed that all the selected LAB strains exhibited varied auto-aggregation index, which was time-dependent (Table 7). Indeed, the auto-aggregation of the LAB strains ranged from 28.87%±3.68% to 16.70%±1.12% and from 70.30%±4.73% to 34.67%±0.99% after 4 h and 24 h, respectively. Zooming on auto-aggregation abilities after 4 h, the highest auto-aggregation capacities were observed for Lpb. plantarum strain isolated from fermented olive (RSOLV: 28.87%±3.68%), and the lowest autoaggregation index was obtained by Lpb. plantarum strain isolated from locust (1nm) with a value of 16.70%±1.12%. After 24 h, the highest values of auto-aggregation capability were obtained by fermented olives strain S4 as well as fermented pepper strain pepp2 with a value of 70.30%±4.73% and 69.82%±2.20%, respectively. It is noteworthy that Lpb. plantarum pepp2 showed the highest Δ Auto-aggregation index with a value 53.12 in 20 h of incubation.
Co-aggregation ability with Listeria was showed by all LAB, which increased over tested times (4h and 24h) at pH=7.0. The co-aggregation capacity ranged from 7.94%±0.94% to 4.99%±0.55% and from 8.76%±1.16% to 4.61%±0.79% after 4 h and 24 h, respectively (Table 8). Zooming on co-aggregation abilities after 24h, the highest co-aggregation capacities were observed for Lpb. plantarum RSOLV (8.76%±1.16%) and S4 (7.37%±0.46%), both strains isolated from fermented olives.
The co-aggregation capacity to E.coli ranged from 28.30%±10.7% to 15.05%±0.89% and from 54.88%±6.98% to 36.96%±5.72% after 4 h and 24h, respectively as showed in Table 8. Zooming on co-aggregation abilities after 24h, the highest co-aggregation capacities were observed for Lpb. plantarum 1nm strain (54,88%±6,98%), followed by Lpb. plantarum strains isolated from fermented vegetables RSOLV (51.15%±4.69%). These results showed that the co-aggregation ability of Lpb. plantarum strains were strain-specific, with the highest co-aggregation capacity toward E. coli in comparison with L. monocytogenes.
3.4. Safety features: Antibiotic resistance and hemolytic activity
The antibiotic (ATB) susceptibility is one of the required properties by which specific strains can be considered as a potential probiotic bacterium. Indeed, probiotics should not harbor acquired antibiotic resistances. The six selected Lbp. plantarum strains were phenotypically analysed toward various types of ATBs, as reported below (Table 9). Most of the strains were resistant toward kanamycin, streptomycin and tetracyclin while sensitive and intermediate to ampicillin and gentamycin. All strains were considered non-hemolytic (γ-hemolysis).
3.5. In vitro oro-gastro-intestinal transit tolerance assay
The selected Lpb. plantarum strains were investigated for their capability to tolerate OGI stress in vitro (Figure 3). To mimic the OGI transit condition, bacterial cultures were exposed sequentially to lysozyme, pepsin and acidic pH, then bile salts and pancreatic enzymes. As shown in Figure 4, all the strains were able to survive until the end of the experiment and with different levels of viability. There was no significant reduction in the survival rate in the first 2 steps (i.e., saliva stress and gastric stress pH 3) compared to the control; however, a significant decrease in the viability was observed for all the strains by incubating at pH 2, in presence of pepsin. In fact, the viability index decreased four times in comparison with the first two steps. All the strains showed a similar tolerance level of the positive control L. reuteri DSM 17938 with a value 3.80%±0.75%. Nevertheless, the survival rate decreased slightly under simulated intestinal conditions (bile salts and pancreatic enzymes). The highest cell viability was detected for Lpb. plantarum isolated from fermented olive S4 (2.33±0.4), followed by locust strains 1nm (1.90±0.5).
3.6. Biofilm formation
The biofilm formation abilities on the abiotic surface were investigated on 96-well polystyrene microliter plates using Resazurin and Crystal Violet after 24 h, 48 h and 72 h (Figure 4).
Regarding Resazurin detection (Figure 4A), the six Lbp. plantarum strains were found weak to moderate biofilm producers. In addition, their absorbance was similar to the positive control E. faecalis V583. Based on the absorbance, after 24 h Lpb. plantarum S4, isolated from fermented olive, showed the highest rate of viable cells (0.22±0.02); otherwise, after 72 h, the highest rate of viable cells was recorded for Lpb. plantarum isolated from locust, namely 1nm, with a value 0.29±0.00. No significant difference was observed between 24 h and 48 h of incubation for all the strains.
The biofilm formation was also investigated using Crystal Violet (Figure 4B). Similar to what was found with Resazurin, all tested strains were able to form biofilm. Based on absorbance values, Lpb. plantarum isolated from fermented olive S4 was found to be the best biofilm producer candidate after 24h.a significant difference was confirmed(p<0.001) between 24h and 72h of incubation for all the strains. On the other hand, pepp2 showed a good ability to form biofilm on abiotic surfaces after 48h at 37°C. The highest rate of biofilm production after 72h was recorded for the same strain, Lpb. plantarum S4, with a value 1.55±0.01.
3.7. Cytotoxic effects
The cytotoxicity of CFSs from the analysed Lbp. plantarum strain was evaluated on cancer cells LS line upon incubation (24h and 72h) at different concentrations (Figure 5); the bacterial medium (MRS) did not show any cytotoxic effect.
All the strains showed an inhibitory effect on LS cell line after 24h, with IC50 ranging between 9.91mg/ml and 18.27mg/ml (Table 10). Results showed that the best candidate is S4 isolated from fermented olive with IC50 = 9.9mg/ml. Regarding the anti-proliferative activity of CFSs on LS cells, data showed toxicity depends on the incubation duration. Furthermore, after 72h IC50 was around 2.4 mg/ml to 3.7 mg/ml, corresponding, respectively, to 1nm and pepp2 strains. At a concentration 8 mg/ml of CFSs and after 72 hours of incubation, the viability of LS cells decreases with values ranging between 33.64% and 18.68% corresponding, respectively, to 1nm CFS and pepp1 CFS. After 24h and 72h, starting from the concentration of 1mg/ml of CFS, all the strains exhibited a significant inhibition (p<0.0001) of LS proliferation and at a concentration 10mg/ml the pourcentage of survival cells was reduced by 50%. The strains 1nm and S4, isolated from locust intestines and fermented olive, respectively, showed a higher significant toxicity effect compared to the other strains in all tested concentration.
3.8. Adhesion to human enterocyte-like cells
In this study, the adhesive capacities of the selected Lpb. plantarum strains were assessed and compared to that of commercial Lpb. plantarum 299V, a probiotic model. The obtained data confirm the adhesive potential of all the strains, which were higher or equal to the adhesion value of Lpb. plantarum 299V (2.5±0.3%). ranging between 3.4%±2.5% and 6.2%±2.5% (Figure 6). The strains Lpb. plantarum 3nm (6.2±2.4, p<0.05) and S4 (5.5±1.4%) exhibited the highest adhesion percentage with a value of more than 5%, although the difference between S4 and Lpb. plantarum 299V was not significant. No significant difference was found between tested Lpb. plantarum.
3.9. Application in food model
3.9.1. Adhesion on tomatoes surface
The adhesion of the investigated Lpb. plantarum strains to tomatoes was performed using grape tomatoes (Solanum lycopersicum L.), purchased from a local market; the survival and adhesion rates were measured after 2, 3 and 5 days of incubation at ambient temperature, respectively (Figure 7). All the strains could survive and attach to tomato surface after 5 days of incubation at ambient temperature with different levels. The strain Lpb. plantarum 1nm showed the best performance of adhesion to tomatoes after 5 days, contrary to strain RSOLV, which showed a decreased level of attachment during the incubation period. After 3 days of incubation, all the strains showed no significant difference (p<0.05) with a range of attachment ranging between 4.02 and 5.96 (LogCFU/g Tomato). Comparing adhesion capacity on tomatoes in function of time incubation, the strain Lpb. plantarum pepp2 showed a lower rise level with delta time Δt=0.728 between the 3rd and 5th days of incubation.
3.9.2. Biocontrol/pathogen antagonism on tomatoes
In order to elucidate the interaction between LAB and pathogens on a model food matrix, each of the investigated Lpb. plantarum strains was mixed with a bacterial pathogen and then inoculated on tomatoes (Table 11). Tomatoes were kept for 5 days at room temperature to mimic ordinary conditions; later, the attached and alive microorganisms were counted on the selective medium MRS agar, SMAC agar and PALCAM agar. Bacteria (LAB, E.coli and L. monocytogenes) inoculated in pure cultures were used as a control. It seems that the adhesion ability on tomatoes is affected by time (Table 10). Regarding the co-cultures, all LAB strains showed a significant attachment decrease (p<0.0001) after 3 days of incubation, ranging from 1.78±1.53 Log CFU/g tomato to 1.42±1.26 Log CFU/g tomato followed by an increse in the fifth day, only 1nm remain stable compared to the other strains. In fact, all Lpb. plantarum strains co-cultured with E.coli or L. monocyogenes were able to attach and to survive on tomato after 5 days and with a similar rate to the control (Supplementary Table S1). Results indicated that Lpb. plantraum strains were able to significantly decrease/inhibit the viability of E.coli in the fifth day with a range of 1.38±0.31-1.13±0.45 Log CFU/g tomato after the first day of incubation. The inhibitory effect on E.coli was more pronounced in the presence of the strain Lpb. plantarum S4, while the strain 3nm showed the best survival rate in co-culture. In the co-cultures (Lpb. plantarum-Listeria), an inhibitory effect on the growth of the pathogen was observed after 3 days in presence of each of the six Lpb. plantarum strains, with L. monocytogenes count ranging between 1,00±0,67 and 1,54±1,41 Log CFU/g tomato. On the fifth day, the CFU number of Listeria in co-culture with Lpb. plantarum was significantly reduced compared to the control. Interestingly, strains 1nm and S4 inhibited completely (p<0.05) the growth of L. monocytogenes on tomatoes after 5 days.
4. Discussion
Probiotics have been traditionally isolated from dairy products, though, in recent years, there has been an increasing trend in exploring novel and alternative sources [54]. Unconventional niches that are being investigated for discovering new probiotics include traditional fermented foods and beverages, vegetables, fruit juices [55,56] and the intestine of insects [57,58]. According to scientific research, fermented vegetables are interesting sources of potentially probiotic LAB strains [7,59,60]. Many Tunisian publications reported that fermented vegetables host LAB strains with an antimicrobial and bio-protective effect against germs and moulds [61,62,63], but only a few articles mentioned the possible probiotic properties of strains from Tunisian fermented vegetables.
The present study aimed to isolate LAB strains from unconventional sources and to characterise them for further usage, such as probiotic cultures and/or for food biocontrol. After screening presumptive LAB from very diverse sources, including infant faeces, breast milk, rabbit intestine, fermented olive etc., we focused on six strains exhibiting the highest antagonistic action toward foodborne pathogens. Such selected strains, derived from fermented vegetables (fermented olive and pepper) and locust intestine, were identified as Lactiplantibacillus plantarum and were further investigated for their antimicrobial, probiotic and biocontrolling properties. Our findings are supported by similar works which reported Lpb. plantarum as the dominant group in fermented vegetables, e.g. tomatoes, and table olives [64,65]. According to recent studies, Clostridium (Firmicutes) is one of the most abundant genera found in insects [58]. Likewise, Garofalo et al. [66] confirmed the dominance of Clostridium in locust, mainly represented by Enterobacteriaceae and Weissella spp.. Another study reported the presence of Lactobacillus metriopterae sp. nov. in locust gut [67]. Unexpectedly, in the current study, LAB isolated from locust, were identified as Lpb. plantarum with similarity over 98%, and we can mention that these are the first Lpb. plantarum found and characterized in locust. LAB isolated from insects are being considered as promising probiotics for the benefit of human and animal health due to the survive/persist of their host in hard environments [58]. Indeed, some very recent researches have looked into the probiotic potential of LAB obtained from insect gut [57,68,69]. A complete genome sequence of Weissella confusa LM1, found in the gut of the migratory locust, indicated the ability to adapt to different ecological niches [70]. While there are studies on the microbiological communities (including LAB) in grasshoppers (Locusta migratoria migratorioides) sold for human consumption [71], to our knowledge, this is the first report studying the probiotic potentialities of locust-derived LAB.
Nowadays, probiotics with antimicrobial activity are becoming an alternative to traditional drugs due to antibiotics resistance diffusion [72]. Interestingly, it has been found that all the strains were able to inhibit the indicators growth; a similar study [73], showed that Lpb. plantarum exhibits an inhibitory effect against L. monocytogenes and E. coli similar to our tested LAB. Moreover, a recent study confirmed the antibacterial potential of Lpb. plantarum insect strains with a similar range against E. coli [57].
The CFSs from the investigated Lpb. plantarum strains had pH values in the range of 3.5, as reported by previous publications [74]. After pH neutralisation to 6.0, all CFSs showed minimal to no activity against all the pathogens tested, proving the role of organic acids for antimicrobial effects. It was reported that the increased production of organic acid through carbohydrates fermentation decreases the pH of the medium, which is the major factor suppressing pathogen growth [75]. In a similar study [76], CFSs from LAB, including Lpb. plantarum, showed anti-E. coli activity with inhibition zone of ranging between 12.89 ± 0.21 to 15.32 ± 0.28 mm. The authors confirmed that the antimicrobial activity was due to the combination of various metabolites, including organic acids. In fact, CFSs of LAB are a complex mixture of metabolic enzymes, secreted proteins, short-chain fatty acids, vitamins, amino acids, peptides, organic acids, and cell components [77]. It was recently reported that Lpb. plantarum is able to synthesize various beneficial extracellular metabolites, known as postbiotics [20], i.e. bioactive soluble compounds or peptides that are produced during LAB growth [78] that confer health benefits such as infection prevention [79], antitumor and immunomodulatory effects [80]. Likewise, previous works have demonstrated the importance of organic acids as bio-preserving agents [81,82]. In addition, Mirzaei et al. [83] reported that the antimicrobial activity of LAB strains disappeared when their CFSs were adjusted to pH 6.5 and treated with catalase. Nevertheless, the bacteriocin production by LAB is highly affected by several factors, including temperature, pH, and incubation time. It was also reported that the optimum secretion of bacteriocin is when the pH ranges between 5.0 and 6.0 [84]. In the present study, all the tested strains showed antibacterial activity against the target pathogens, most probably due to the organic acids secreted in their CFSs.
The antifungal potential of LAB strains has also been estimated; our results showed that LAB inhibit the growth of P. expansum, A. niger, F. culmorum and B. cinerea. These findings are similar to the work carried out by other investigators [85,86]. A recent Turkish publication [87] showed that Lpb. plantarum was active against P. expansum and A. niger. Indeed, several studies reported that LAB isolated from vegetables and plants possess a better antifungal activity [88,89], while LAB derived from dairy products exhibit antibacterial activity against foodborne pathogens through bacteriocin production capabilities [90,91].
Thereafter, the probiotic traits were investigated. Starting with OGI transit tolerance, all the tested Lpb. plantarum strains were incubated in successive solutions to mimic the human OGI transit, and their viability was evaluated. In fact, to be applied in the food industry, LAB should possess a good resistance to acidic environment, especially in the preparation of high-acid foods [92]. All selected Lpb. plantarum strains were able to survive the simulated OGI transit, with LAB strain of fermented olive S4 exhibiting the highest resistance. Moreover, all the strains showed good resistance to acidic pH through OGI transit, which is congruent with several findings in the literature. Indeed, Lpb. plantarum were proven to be able to survive in pH varies between 2.5 and 4 [93,94]. Besides, in agreement with our findings, several authors demonstrated the ability of Lpb. plantarum to survive to OGI transit with a rate of 103 CFU·mL−1 [41,95].
We further examined the potential probiotics for cell-binding properties. In the current study, all tested LAB exhibited time-dependent auto-aggregation ability, particularly, the fermented olives-derived strains (RSOLV and S4) showed the best auto-aggregation performance. In fact, the auto-aggregation of bacteria has been associated with adherence ability to the intestinal cells, a prerequisite for the colonization of the gastrointestinal tract [96]. In a similar earlier study [97], Lpb. plantarum isolated from fermented vegetables showed auto-aggregation rates very close to our results.
In co-aggregation assays, using the same foodborne pathogenic bacteria as for antibacterial assay, the co-aggregation ability of all tested LAB increased over time and it was high for Lbp. plantarum strains from fermented green pepper, indicating a potential in preventing and/or excluding colonization of pathogens in the gastrointestinal tract. The adhesion ability to cells was strain-specific as it varied even within the same species. Our findings are supported by recent work on LAB strains obtaining similar results [44,53], and are in agreement with the studies of Ben Taheur et al [61], reporting a lower rate of co-aggregation of probiotic LAB with E.coli.
Regarding the adhesion capability, the Lpb. plantarum strains were first evaluated for biofilm production ability on polystyrenes surface, then for adhesion to human enterocyte-like cells and, finally, to tomatoes as a food model. In fact, bacterial adhesion to epithelial cell is considered one of the most accurate features for selection criteria for probiotic strains [98]. Results showed that all selected strains were able to form biofilm on abiotic surface, especially S4 and 1nm in a longer time, with a value similar to previous studies [99], demonstrating that Lpb. plantarum strains provide various levels of biofilm production capability from “no biofilm” to “good biofilm producer”, possibly depending on the amount of exopolysaccharides produced [100]. Furthermore, knowledge about the surface conditions and the bacterial properties influencing adhesion is still insufficient [101]. Concerning the adhesion to human cells, our findings demonstrated that all the tested Lpb. plantarum strains survived and attached to Caco-2 cells better than the probiotic model strain used as control; our results are comparable to values previously obtained for Lpb. plantarum [102]. We also found that the CFSs of the tested Lpb. plantarum strains were able to decrease colorectal adenocarcinoma cells viability, proving their anti-cancer and anti-proliferative potential; these findings are in line with current studies reporting the cytotoxicity of Lpb. plantarum metabolites on cancer cells [20,50].
In the present study, we suggest the use of Lpb. plantarum strains as probiotics and bio-controlling agents on tomatoes. Non-dairy, plant-based food matrices such as fruits, vegetables and legumes have been used successfully in producing probiotic products [103]. The investigated Lpb. plantarum strains showed strong attachment in vivo to tomatoes. Arellano-Ayala et al. [53] reported similar adhesion percentages of LAB to tomato fruits. The selection of tomatoes as a model to elucidate the adhesion ability of the probiotic strains reflect the importance of this vegetable in the human diet. Latest researches investigated LAB's efficiency as protective cultures tool to control L. monocytogenes in ready-to-eat and dairy-ripened products [104]. Moreover, Yin et al., confirmed the possible use of LAB as biocontrol agents for limiting/inhibiting pathogens contamination on leafy greens [105]. Here we investigated the ability of Lpb. plantarum strains to attach to tomatoes in co-culture with pathogens. The enumeration of microorganisms was done in different selective mediums; the biocontrol assay on tomatoes confirmed the ability of all Lpb. plantarum strains, principally 1 nm and S4, to antagonise the tested pathogens, with a maintained count of CFU after 5 days, and with a significant difference compared to the control.
In conclusion, this is one of the few studies that have investigated the probiotic potential of Tunisian vegetables- and locust intestine- derived LAB and their biocontrol capacities on a food model matrix. Six selected Lpb. plantarum strains were analysed for their antagonism potential, demonstrating that they could be good candidates as food-protective cultures and with an interesting probiotic profile. Nevertheless, further studies are needed to deepen their characterisation and to guarantee their use as probiotic strains in biocontrol.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Antibacterial activity of some Lpb. plantarum strains and their CFSs (crude CFS, CFS pH=6.0 and treated cells) against L. monocytogenes (A) and E.coli (B). ; Figure S2: Antifungal activity of Lpb. plantarum strains against some indicator fungi species: Botrytis cinerea CECT 20973 (A) and Penicillium expansum (B). ; Table S1. Pathogen antagonism by Lpb. plantarum on tomatoes. CFU counts of Lpb. plantarum inoculated in MRS as pure culture and inoculated as co-culture with pathogen (L. monocytogenes CECT 4031 or E.coli O157:H7) . Mean ± standard deviation from three independent CFU counts from three different experiments. Each experiment was done in duplicate. Log CFU per tomato compared to single inoculated pathogen used as control. Two-way ANOVA, Dunnett’s multiple comparisons test. § p<0.05 vs 5 days 3nm-Listeria, 1nm-Listeria, RSOLV-Listeria and pepp1-Listeria; & p<0.05 vs 5 days pepp2-Listeria.
Author Contributions
G.S, V.C and F.A.; methodology, H.S.; writing, editing, analysis, draft preparation, V.C, M.T.R, D.F, T.S.L, H.S; review and editing, G.S, T.S.L, F.A., D.F; supervision, G.S., F.A; project administration. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by FCT—Fundação para a Ciência e Tecnologia IP Portugal, throught projects PTDC/OCE-ETA/1785/2020 [EMOTION], UIDB/00276/2020 (CIISA – Centre for Interdisciplinary Research in Animal Health, Faculty of Veterinary Medicine, University of Lisbon) and LA/P/0059/2020-AL4ANIMALS (AL4AnimalS). This work was partially supported by PON project “Conservabilità, qualità e sicurezza dei prodotti ortofrutticoli ad alto contenuto di servizio”—POFACS—CUP B74I20000120005. Teresa Semedo-Lemsaddek is financially supported by national funds through FCT under the Transitional Standard—DL57/2016/CP1438/CT0004. Maria Teresa Rocchetti is the recipient of a research contract for the project no. UNIFG171 – CUP D74I19003340002, as part of the initiative “Research for Innovation (REFIN)—POR PUGLIA FESR FSE 2014- 2020- Azione 10.4”. Hiba Selmi is financed by the Tunisian Ministry of Higher Education and Scientific Research-University of Carthage (Tunisia) through a research grant.
Data Availability Statement
The datasets generated for this study are available on request to the corresponding author.
Acknowledgments
The authors would like to acknowledge the enthusiastic support of the MicroBugs research team, Faculty of Veterinary Medicine, Lisbon, Portugal.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22. [Google Scholar] [CrossRef]
- Hazards, E.P.o.B.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 11: suitability of taxonomic units notified to EFSA until September 2019. EFSA Journal 2020, 18, e05965. [Google Scholar]
- Huang, W.; et al. Evaluation of the fermentation potential of lactic acid bacteria isolated from herbs, fruits and vegetables as starter cultures in nut-based milk alternatives. Food Microbiol 2023, 112, 104243. [Google Scholar] [CrossRef] [PubMed]
- Terefe, N.S. Food Fermentation. In Reference Module in Food Science; Elsevier, 2016. [Google Scholar]
- Mokoena, M.P.; Omatola, C.A.; Olaniran, A.O. Applications of Lactic Acid Bacteria and Their Bacteriocins against Food Spoilage Microorganisms and Foodborne Pathogens. Molecules 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, P.K.; Nehra, K. Bacteriocin-nanoconjugates as emerging compounds for enhancing antimicrobial activity of bacteriocins. Journal of King Saud University-Science 2019, 31, 758–767. [Google Scholar] [CrossRef]
- Touret, T.; Oliveira, M.; Semedo-Lemsaddek, T. Putative probiotic lactic acid bacteria isolated from sauerkraut fermentations. PLoS One 2018, 13, e0203501. [Google Scholar] [CrossRef] [PubMed]
- Medina-Pradas, E.; et al. Chapter 9 - Review of Vegetable Fermentations With Particular Emphasis on Processing Modifications, Microbial Ecology, and Spoilage. In The Microbiological Quality of Food; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing, 2017; pp. 211–236. [Google Scholar]
- Saidi, Y.; et al. Polyphasic Characterisation of Non-Starter Lactic Acid Bacteria from Algerian Raw Camel's Milk and Their Technological Aptitudes. Food Technol Biotechnol 2020, 58, 260–272. [Google Scholar] [CrossRef]
- Hill, C.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature Reviews Gastroenterology & Hepatology 2014, 11, 506–514. [Google Scholar]
- Seddik, H.A.; et al. Lactobacillus plantarum and Its Probiotic and Food Potentialities. Probiotics Antimicrob Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Karnwal, A.; Malik, T. Characterization and selection of probiotic lactic acid bacteria from different dietary sources for development of functional foods. Front Microbiol 2023, 14, 1170725. [Google Scholar]
- Cruz, P.O.D.; et al. Efficacy of Potentially Probiotic Fruit-Derived Lactobacillus fermentum, L. paracasei and L. plantarum to Remove Aflatoxin M(1) In Vitro. Toxins 2020, 13. [Google Scholar] [CrossRef]
- Čanak, I.; et al. Isolation and Characterisation of L. plantarum O1 Producer of Plantaricin as Potential Starter Culture for the Biopreservation of Aquatic Food Products. Food Technol Biotechnol 2018, 56, 581–589. [Google Scholar]
- Mao, B.; et al. Comparative Genomic Analysis of Lactiplantibacillus plantarum Isolated from Different Niches. Genes 2021, 12. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; et al. Health-promoting role of Lactiplantibacillus plantarum isolated from fermented foods. Microorganisms 2021, 9, 349. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, M.T.; et al. Bioprospecting Antimicrobials from Lactiplantibacillus plantarum: Key Factors Underlying Its Probiotic Action. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Kwon, Y.J.; et al. Safety Assessment of Lactiplantibacillus (formerly Lactobacillus) plantarum Q180. J Microbiol Biotechnol 2021, 31, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Ismael, M.; et al. Probiotic of Lactiplantibacillus plantarum NWAFU-BIO-BS29 Isolated from Chinese Traditional Fermented Milk and Its Potential Therapeutic Applications Based on Gut Microbiota Regulation. Foods 2022, 11, 3766. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; et al. Lactobacillus plantarum Metabolites Elicit Anticancer Effects by Inhibiting Autophagy-Related Responses. Molecules 2023, 28. [Google Scholar] [CrossRef]
- Du, G.; et al. Influence of encapsulated Lactobacillus plantarum and eugenol on the physicochemical properties and microbial community of fresh-cut apples. Food Chem X 2023, 17, 100563. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; et al. Inactivation of Foodborne Pathogens by Lactiplantibacillus Strains during Meat Fermentation: Kinetics and Mathematical Modelling. Foods 2023, 12. [Google Scholar] [CrossRef]
- Bvenura, C.; Sivakumar, D. The role of wild fruits and vegetables in delivering a balanced and healthy diet. Food Research International 2017, 99, 15–30. [Google Scholar] [CrossRef]
- Quinet, M.; et al. Tomato fruit development and metabolism. Frontiers in plant science 2019, 10, 1554. [Google Scholar] [CrossRef]
- www.statista.com. Production of tomatoes in Tunisia from 2016 to 2021. 2023.
- Hwang, D.; et al. Cherry tomato supplementation increases the area of the intestinal mucosa and the number of muscle layers in rats. Food Res Int 2014, 64, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Swain, M.R.; et al. Fermented fruits and vegetables of Asia: a potential source of probiotics. Biotechnol Res Int 2014, 2014, 250424. [Google Scholar] [CrossRef]
- Jin, T.Z.; Fan, X.; Mukhopadhyay, S. Antimicrobial coating with organic acids and essential oil for the enhancement of safety and shelf life of grape tomatoes. Int J Food Microbiol 2022, 378, 109827. [Google Scholar] [CrossRef]
- Gray, J.A.; et al. Novel Biocontrol Methods for Listeria monocytogenes Biofilms in Food Production Facilities. Front Microbiol 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Lavilla, M.; et al. Natural Killers: Opportunities and Challenges for the Use of Bacteriophages in Microbial Food Safety from the One Health Perspective. Foods 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Özer, F.; et al. Comparison of antibacterial activity of two dentin bonding systems using agar well technique and tooth cavity model. Journal of Dentistry 2003, 31, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Angmo, K.; Kumari, A.; Bhalla, T.C. Probiotic characterization of lactic acid bacteria isolated from fermented foods and beverage of Ladakh. LWT-food Science and Technology 2016, 66, 428–435. [Google Scholar] [CrossRef]
- Semedo, T.; et al. Comparative study using type strains and clinical and food isolates to examine hemolytic activity and occurrence of the cyl operon in enterococci. Journal of Clinical Microbiology 2003, 41, 2569–2576. [Google Scholar] [CrossRef]
- Dommels, Y.E.; et al. Survival of Lactobacillus reuteri DSM 17938 and Lactobacillus rhamnosus GG in the human gastrointestinal tract with daily consumption of a low-fat probiotic spread. Applied and environmental microbiology 2009, 75, 6198–6204. [Google Scholar] [CrossRef] [PubMed]
- Hancock, L.E.; Perego, M. The Enterococcus faecalis fsr two-component system controls biofilm development through production of gelatinase. Journal of bacteriology 2004, 186, 5629–5639. [Google Scholar] [CrossRef] [PubMed]
- Hancock, L.; Perego, M. Two-component signal transduction in Enterococcus faecalis. J Bacteriol 2002, 184, 5819–25. [Google Scholar] [CrossRef] [PubMed]
- Ahrne, S.; Hagslatt, M.-L.J. Effect of lactobacilli on paracellular permeability in the gut. Nutrients 2011, 3, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Prasad, D. Application of in vitro methods for selection of Lactobacillus casei strains as potential probiotics. International journal of food microbiology 2005, 103, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; et al. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int J Food Microbiol 2017, 247, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; et al. Purification and characterization of plantaricin SLG1, a novel bacteriocin produced by Lb. plantarum isolated from yak cheese. Food Control 2018, 84, 111–117. [Google Scholar]
- De Simone, N.; et al. Antimicrobial Properties, Functional Characterisation and Application of Fructobacillus fructosus and Lactiplantibacillus plantarum Isolated from Artisanal Honey. Probiotics Antimicrob Proteins 2023, 15, 1406–1423. [Google Scholar] [CrossRef]
- CLSI, Performance Standards for Antimicrobial Susceptibility Testing. 31st edition ed. 2021, CLSI supplement M100: Clinical and Laboratory Standards Insitute.
- Charteris, W.P.; et al. Antibiotic susceptibility of potentially probiotic Lactobacillus species. Journal of food protection 1998, 61, 1636–1643. [Google Scholar] [CrossRef]
- Reuben, R.; et al. Characterization and evaluation of lactic acid bacteria from indigenous raw milk for potential probiotic properties. Journal of dairy science 2020, 103, 1223–1237. [Google Scholar] [CrossRef]
- Armas, F.; Camperio, C.; Marianelli, C. In vitro assessment of the probiotic potential of Lactococcus lactis LMG 7930 against ruminant mastitis-causing pathogens. PLoS One 2017, 12, e0169543. [Google Scholar] [CrossRef] [PubMed]
- Gheziel, C.; et al. Evaluating the Probiotic Potential of Lactobacillus plantarum Strains from Algerian Infant Feces: Towards the Design of Probiotic Starter Cultures Tailored for Developing Countries. Probiotics Antimicrob Proteins 2019, 11, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Gómez, N.C.; et al. Use of Potential Probiotic Lactic Acid Bacteria (LAB) Biofilms for the Control of Listeria monocytogenes, Salmonella Typhimurium, and Escherichia coli O157:H7 Biofilms Formation. Front Microbiol 2016, 7, 863. [Google Scholar] [CrossRef] [PubMed]
- Cruz, C. D.; Shah, S.; Tammela, P. Defining conditions for biofilm inhibition and eradication assays for Gram-positive clinical reference strains. BMC Microbiol 2018, 18, 173. [Google Scholar] [CrossRef] [PubMed]
- Borges, S.; Silva, J.; Teixeira, P. Survival and biofilm formation by Group B streptococci in simulated vaginal fluid at different pHs. Antonie Van Leeuwenhoek 2012, 101, 677–82. [Google Scholar] [CrossRef]
- Chuah, L.O.; et al. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement Altern Med 2019, 19, 114. [Google Scholar] [CrossRef]
- Fernández de Palencia, P.; et al. Probiotic strains: survival under simulated gastrointestinal conditions, in vitro adhesion to Caco-2 cells and effect on cytokine secretion. European Food Research and Technology 2008, 227, 1475–1484. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; et al. Inactivation of Salmonella in grape tomato stem scars by organic acid wash and chitosan-allyl isothiocyanate coating. Int J Food Microbiol 2018, 266, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Arellano-Ayala, K.; et al. Hydrophobic and adhesive patterns of lactic acid bacteria and their antagonism against foodborne pathogens on tomato surface (Solanum lycopersicum L.). J Appl Microbiol 2020, 129, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic isolates from unconventional sources: a review. J Anim Sci Technol 2016, 58, 26. [Google Scholar] [CrossRef]
- Meradji, M.; et al. Characterization of Lactic Acid Bacteria Strains Isolated from Algerian Honeybee and Honey and Exploration of Their Potential Probiotic and Functional Features for Human Use. Foods 2023, 12. [Google Scholar] [CrossRef]
- Linares-Morales, J.R.; et al. Selection of Lactic Acid Bacteria Isolated from Fresh Fruits and Vegetables Based on Their Antimicrobial and Enzymatic Activities. Foods 2020, 9. [Google Scholar] [CrossRef]
- SIREGAR, D.J.S.; et al. Selection of probiotic candidate of lactic acid bacteria from Hermetia illucens larvae fed with different feeding substrates. Biodiversitas Journal of Biological Diversity 2022, 23. [Google Scholar] [CrossRef]
- Siddiqui, R.; Elmashak, Y.; Khan, N. A. Cockroaches: a potential source of novel bioactive molecule(s) for the benefit of human health. Appl Entomol Zool 2023, 58, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Alameri, F.; et al. Lactic Acid Bacteria Isolated from Fresh Vegetable Products: Potential Probiotic and Postbiotic Characteristics Including Immunomodulatory Effects. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Hurtado, A.; et al. Lactic acid bacteria from fermented table olives. Food Microbiol 2012, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ben Taheur, F.; et al. Anti-bacterial and anti-biofilm activity of probiotic bacteria against oral pathogens. Microb Pathog 2016, 97, 213–20. [Google Scholar] [CrossRef] [PubMed]
- Fhoula, I.; et al. Diversity and antimicrobial properties of lactic acid bacteria isolated from rhizosphere of olive trees and desert truffles of Tunisia. Biomed Res Int 2013, 2013, 405708. [Google Scholar] [CrossRef] [PubMed]
- Klibi, N.; et al. Genotypic diversity, antibiotic resistance and bacteriocin production of enterococci isolated from rhizospheres. Microbes Environ 2012, 27, 533–7. [Google Scholar] [CrossRef] [PubMed]
- Abouloifa, H.; et al. Characterization of probiotic properties of antifungal Lactobacillus strains isolated from traditional fermenting green olives. Probiotics and antimicrobial proteins 2020, 12, 683–696. [Google Scholar] [CrossRef]
- Kachouri, F.; Hamdi, M. Use Lactobacillus plantarum in olive oil process and improvement of phenolic compounds content. Journal of Food Engineering 2006, 77, 746–752. [Google Scholar] [CrossRef]
- Garofalo, C.; et al. The microbiota of marketed processed edible insects as revealed by high-throughput sequencing. Food Microbiol 2017, 62, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Chiba, M.; et al. Lactobacillus metriopterae sp. nov., a novel lactic acid bacterium isolated from the gut of grasshopper Metrioptera engelhardti. Int J Syst Evol Microbiol 2018, 68, 1484–1489. [Google Scholar] [CrossRef] [PubMed]
- Leska, A.; et al. Binding and Detoxification of Insecticides by Potentially Probiotic Lactic Acid Bacteria Isolated from Honeybee (Apis mellifera L.) Environment-An In Vitro Study. Cells 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Vergalito, F.; et al. Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties. Foods 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; et al. Complete Genome Sequence of Weissella confusa LM1 and Comparative Genomic Analysis. Front Microbiol 2021, 12, 749218. [Google Scholar] [CrossRef] [PubMed]
- Stoops, J.; et al. Microbial community assessment of mealworm larvae (Tenebrio molitor) and grasshoppers (Locusta migratoria migratorioides) sold for human consumption. Food Microbiology 2016, 53, 122–127. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; et al. Benefits and Inputs From Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Front Microbiol 2019, 10, 57. [Google Scholar] [CrossRef]
- Dai, J.; et al. Isolation and identification of new source of bacteriocin-producing Lactobacillus plantarum C010 and growth kinetics of its batch fermentation. World Journal of Microbiology and Biotechnology 2022, 38, 1–14. [Google Scholar] [CrossRef]
- Liu, C.; et al. Probiotic Potential of Lactobacillus Strains Isolated From Fermented Vegetables in Shaanxi, China. Front Microbiol 2021, 12, 774903. [Google Scholar] [CrossRef]
- Kivanc, M.; Yilmaz, M.; Çakir, E. Isolation and identification of lactic acid bacteria from boza, and their microbial activity against several reporter strains. Turkish Journal of Biology 2011, 35, 313–324. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, X.; Xu, Z. Identification of antibacterial substances of Lactobacillus plantarum DY-6 for bacteriostatic action. Food Science & Nutrition 2020, 8, 2854–2863. [Google Scholar]
- Spaggiari, L.; et al. Lactobacillus acidophilus, L. plantarum, L. rhamnosus, and L. reuteri Cell-Free Supernatants Inhibit Candida parapsilosis Pathogenic Potential upon Infection of Vaginal Epithelial Cells Monolayer and in a Transwell Coculture System In Vitro. Microbiol Spectr 2022, 10, e0269621. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; et al. Comparative Studies of Inhibitory and Antioxidant Activities, and Organic Acids Compositions of Postbiotics Produced by Probiotic Lactiplantibacillus plantarum Strains Isolated From Malaysian Foods. Front Vet Sci 2020, 7, 602280. [Google Scholar] [CrossRef]
- Khani, N.; et al. Assessing the growth-inhibitory activity of postbiotics of Lactobacillus spp. against Staphylococcus aureus under in vitro circumstances and food model. Lett Appl Microbiol 2023, 76. [Google Scholar]
- Thorakkattu, P.; et al. Postbiotics: Current Trends in Food and Pharmaceutical Industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef] [PubMed]
- Divyashree, S.; et al. Probiotic properties of Lactobacillus casei–MYSRD 108 and Lactobacillus plantarum-MYSRD 71 with potential antimicrobial activity against Salmonella paratyphi. Biotechnology Reports 2021, 32, e00672. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; et al. Characterization of antimicrobial activity of three Lactobacillus plantarum strains isolated from Chinese traditional dairy food. Food science & nutrition 2019, 7, 1997–2005. [Google Scholar]
- Mirzaei, E.Z.; Lashani, E.; Davoodabadi, A. Antimicrobial properties of lactic acid bacteria isolated from traditional yogurt and milk against Shigella strains. GMS hygiene and infection control 2018, 13. [Google Scholar]
- Sirisopapong, M.; et al. Assessment of lactic acid bacteria isolated from the chicken digestive tract for potential use as poultry probiotics. Asian-Australas J Anim Sci 2023. [Google Scholar] [CrossRef]
- Abd Ellatif, S.A.; et al. Assessment of probiotic efficacy and anticancer activities of Lactiplantibacillus plantarum ESSG1 (MZ683194.1) and Lactiplantibacillus pentosus ESSG2 (MZ683195.1) isolated from dairy products. Environ Sci Pollut Res Int 2022, 29, 39684–39701. [Google Scholar] [CrossRef] [PubMed]
- De Simone, N.; et al. Screening of Lactic Acid Bacteria for the Bio-Control of Botrytis cinerea and the Potential of Lactiplantibacillus plantarum for Eco-Friendly Preservation of Fresh-Cut Kiwifruit. Microorganisms 2021, 9. [Google Scholar] [CrossRef]
- Arsoy, E. S.; Gül, L. B.; Çon, A. H. Characterization and Selection of Potential Antifungal Lactic Acid Bacteria Isolated From Turkish Spontaneous Sourdough. Curr Microbiol 2022, 79, 148. [Google Scholar] [CrossRef] [PubMed]
- Simões, L.; et al. Brazilian Table Olives: A Source of Lactic Acid Bacteria with Antimycotoxigenic and Antifungal Activity. Toxins 2023, 15. [Google Scholar] [CrossRef]
- Zhao, S.; et al. Antifungal Activity of Lactobacillus plantarum ZZUA493 and Its Application to Extend the Shelf Life of Chinese Steamed Buns. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Grujović, M.; et al. Advantages and disadvantages of non-starter lactic acid bacteria from traditional fermented foods: Potential use as starters or probiotics. Compr Rev Food Sci Food Saf 2022, 21, 1537–1567. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; et al. Screening and Identification of Goat-Milk-Derived Lactic Acid Bacteria with Bacteriocin-like Activity and Probiotic Potentials. Microorganisms 2023, 11, 849. [Google Scholar] [CrossRef]
- Rabaoui, G.; et al. Potential Probiotic Lactic Acid Bacteria with Anti-Penicillium expansum Activity from Different Species of Tunisian Edible Snails. Probiotics Antimicrob Proteins 2022. [Google Scholar] [CrossRef]
- Chouikhi, A.; et al. A novel probiotic strain, Lactiplantibacillus plantarum LC38, isolated from Tunisian camel milk promoting wound healing in Wistar diabetic rats. Arch Microbiol 2021, 204, 24. [Google Scholar] [CrossRef]
- Ben Salah, R.; et al. A new Lactobacillus plantarum strain, TN8, from the gastro intestinal tract of poultry induces high cytokine production. Anaerobe 2012, 18, 436–44. [Google Scholar] [CrossRef]
- Rocchetti, M.T.; et al. Immunomodulatory Activity on Human Macrophages by Cell-Free Supernatants to Explore the Probiotic and Postbiotic Potential of Lactiplantibacillus plantarum Strains of Plant Origin. Probiotics and Antimicrobial Proteins 2023, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Del Re, B.; et al. Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Letters in applied microbiology 2000, 31, 438–442. [Google Scholar] [CrossRef] [PubMed]
- das Neves Selis, N.; et al. Lactiplantibacillus plantarum strains isolated from spontaneously fermented cocoa exhibit potential probiotic properties against Gardnerella vaginalis and Neisseria gonorrhoeae. BMC Microbiol 2021, 21, 198. [Google Scholar] [CrossRef] [PubMed]
- Gorreja, F.; Walker, W. A. The potential role of adherence factors in probiotic function in the gastrointestinal tract of adults and pediatrics: a narrative review of experimental and human studies. Gut Microbes 2022, 14, 2149214. [Google Scholar] [CrossRef] [PubMed]
- Mekky, A.F.; et al. Anti-biofilm potential of Lactobacillus plantarum Y3 culture and its cell-free supernatant against multidrug-resistant uropathogen Escherichia coli U12. Saudi J Biol Sci 2022, 29, 2989–2997. [Google Scholar] [CrossRef]
- Maunders, E.; Welch, M. Matrix exopolysaccharides; the sticky side of biofilm formation. FEMS Microbiol Lett 2017, 364. [Google Scholar] [CrossRef]
- Wallis, J. K.; Krömker, V.; Paduch, J. H. Biofilm formation and adhesion to bovine udder epithelium of potentially probiotic lactic acid bacteria. AIMS Microbiol 2018, 4, 209–224. [Google Scholar] [CrossRef]
- De Simone, N.; et al. Antimicrobial Properties, Functional Characterisation and Application of Fructobacillus fructosus and Lactiplantibacillus plantarum Isolated from Artisanal Honey. Probiotics Antimicrob Proteins 2022. [Google Scholar] [CrossRef]
- Rasika, D.; et al. Probiotic delivery through non-dairy plant-based food matrices. Agriculture 2021, 11, 599. [Google Scholar] [CrossRef]
- Martín, I.; et al. Strategies for Biocontrol of Listeria monocytogenes Using Lactic Acid Bacteria and Their Metabolites in Ready-to-Eat Meat- and Dairy-Ripened Products. Foods 2022, 11. [Google Scholar] [CrossRef]
- Yin, H.B.; et al. Pre-harvest biocontrol of Listeria and Escherichia coli O157 on lettuce and spinach by lactic acid bacteria. Int J Food Microbiol 2023, 387, 110051. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram of the oro-gastrointestinal transit (OGI) assay used to test the tolerance of Lpb. plantarum strains in vitro. Bacterial cultures were first washed twice with sterile buffer, then incubated in lysozyme solution to mimic the saliva stress for 5 min; later, the pellet was incubated in a solution containing pepsin with different pH values, arriving at the intestinal stress in which cells were inoculated in bile salts and pancreatin for 60 min and then, incubated for 1h at 37 °C in a sterile electrolyte solution.
Figure 1.
Schematic diagram of the oro-gastrointestinal transit (OGI) assay used to test the tolerance of Lpb. plantarum strains in vitro. Bacterial cultures were first washed twice with sterile buffer, then incubated in lysozyme solution to mimic the saliva stress for 5 min; later, the pellet was incubated in a solution containing pepsin with different pH values, arriving at the intestinal stress in which cells were inoculated in bile salts and pancreatin for 60 min and then, incubated for 1h at 37 °C in a sterile electrolyte solution.
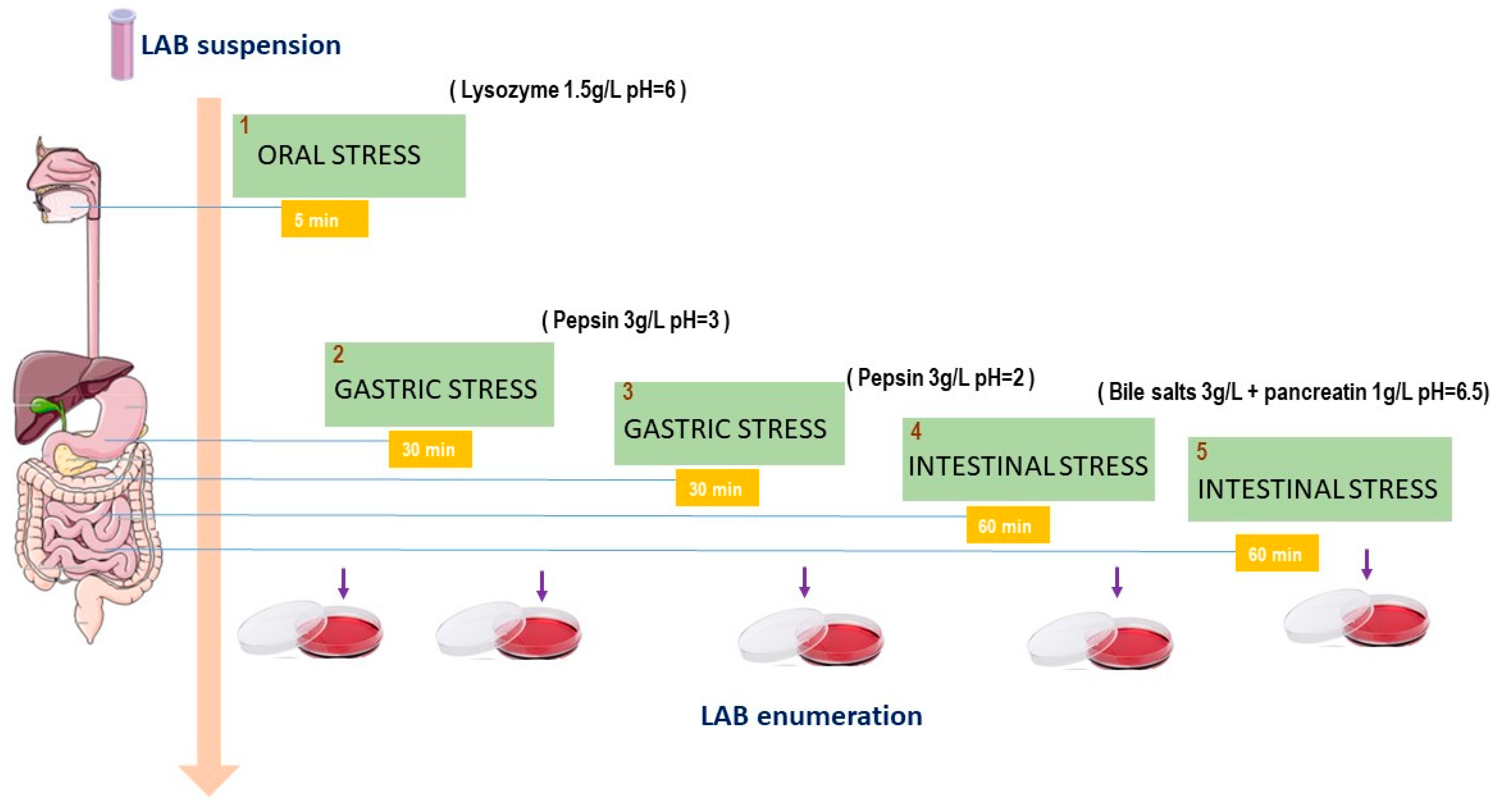
Figure 2.
Schematic representation of adhesion in food model assay. First, tomatoes were washed with sterile distilled water and decontaminated with UV light for 15 mins; then, the scar was created using a sterile syringe and Lpb. plantarum cultures were inoculated in spots. Inoculated tomatoes were kept for 2 h to dry under the laminar flow hood and incubated at room temperature. Sterile PBS buffer pH=7.0 was used as a control.
Figure 2.
Schematic representation of adhesion in food model assay. First, tomatoes were washed with sterile distilled water and decontaminated with UV light for 15 mins; then, the scar was created using a sterile syringe and Lpb. plantarum cultures were inoculated in spots. Inoculated tomatoes were kept for 2 h to dry under the laminar flow hood and incubated at room temperature. Sterile PBS buffer pH=7.0 was used as a control.

Figure 3.
Oro-gastrointestinal tolerance in vitro showed by Lpb. plantarum strains. Strains were incubated in MRS (control), then exposed successively to lysozyme (OG1), pepsin pH 3 (OG2), pepsin pH 2 (OG3) and bile salts and pancreatic enzymes (OG4 ). Finally, bacterial cultures were incubated in an intestinal electrolyte solution pH 6.5 (OG5). The assay was performed in triplicate. ANOVA was used for significant differences at each time point (control, OG1, OG2, OG3, OG4) of the OGI transit between strains. ù p < 0.0001 vs all strains; m p<0.05 vs OG1 of strains 3nm, S4, RSOLV, pepp1 and pepp2; § p<0.05 vs OG1 of strains 1nm, S4, RSOLV and pepp1; $ p<0.0001 vs OG1 of strains S4, RSOLV, pepp1; £p<0.0001 vs OG1 of strains RSOLV, pepp2; !p<0.0001 vs OG1 of strains pepp1 and pepp2; l p<0.0001 vs OG1 pepp2; B p<0.05 vs OG2 of strains S4 and RSOLV; à p<0.05 vs OG2 of strains 3nm, 1nm, RSOLV and pepp2.
Figure 3.
Oro-gastrointestinal tolerance in vitro showed by Lpb. plantarum strains. Strains were incubated in MRS (control), then exposed successively to lysozyme (OG1), pepsin pH 3 (OG2), pepsin pH 2 (OG3) and bile salts and pancreatic enzymes (OG4 ). Finally, bacterial cultures were incubated in an intestinal electrolyte solution pH 6.5 (OG5). The assay was performed in triplicate. ANOVA was used for significant differences at each time point (control, OG1, OG2, OG3, OG4) of the OGI transit between strains. ù p < 0.0001 vs all strains; m p<0.05 vs OG1 of strains 3nm, S4, RSOLV, pepp1 and pepp2; § p<0.05 vs OG1 of strains 1nm, S4, RSOLV and pepp1; $ p<0.0001 vs OG1 of strains S4, RSOLV, pepp1; £p<0.0001 vs OG1 of strains RSOLV, pepp2; !p<0.0001 vs OG1 of strains pepp1 and pepp2; l p<0.0001 vs OG1 pepp2; B p<0.05 vs OG2 of strains S4 and RSOLV; à p<0.05 vs OG2 of strains 3nm, 1nm, RSOLV and pepp2.
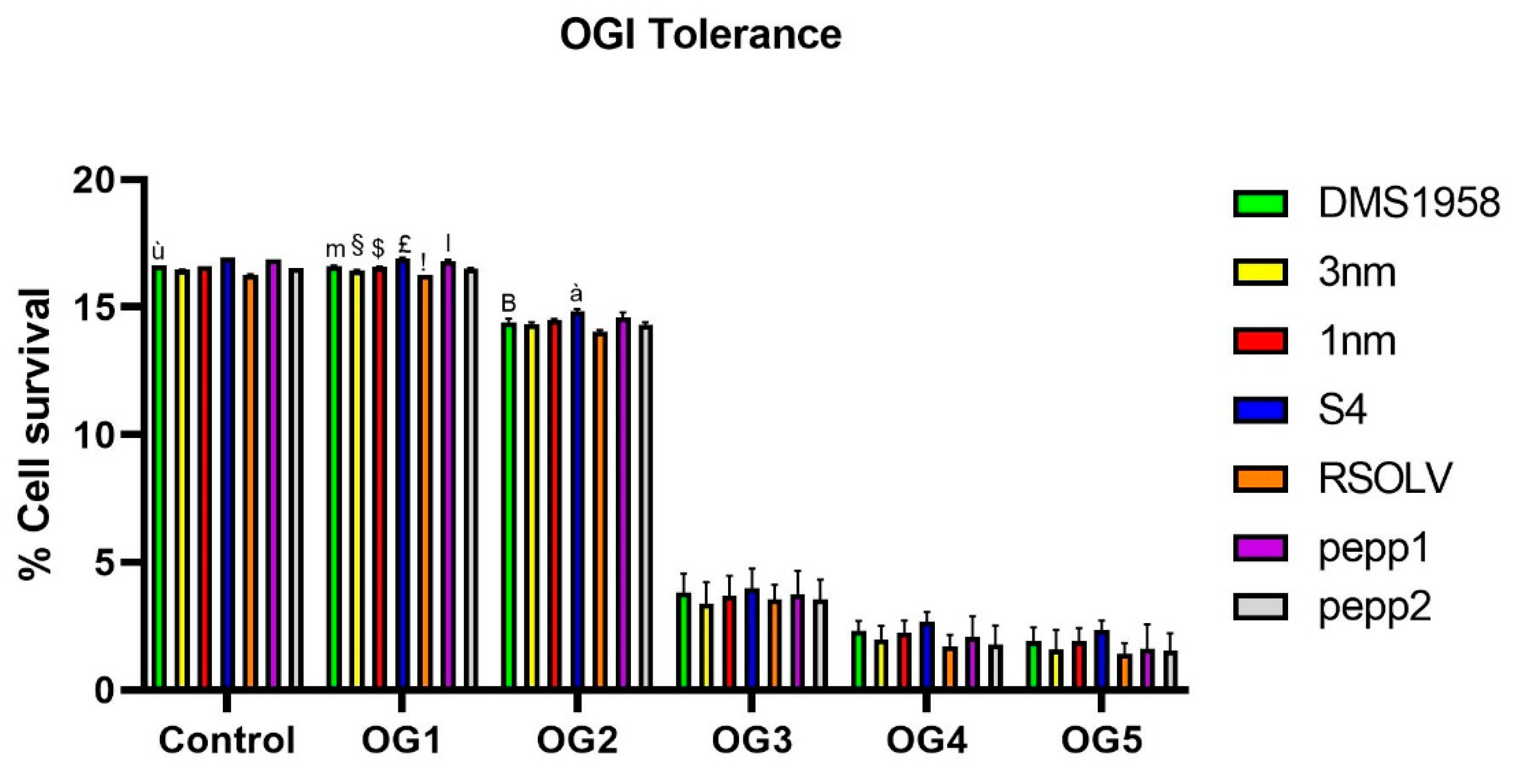
Figure 4.
Biofilm formation (expressed as absorbance at 595 nm) of the Lpb. plantarum strains after 24h and 72h of incubation at 37°C using Resazurin (RZ) (A) or Crystal Violet (CV) (B). Error bars indicate the standard deviation of triplicate experiments.ANOVA was used between strains and to control at each time (24h, 48h and 72h).b p<0.05 vs RZ 24h of all strains except pepp2; T p<0.05 vs RZ 24 of strains 3nm, 1nm and pepp2; H p<0.05 vs RZ 24h of strains V583, RSOLV and pepp1; G p<0.05 vs RZ 72h of strains V583, RSOLV and pepp1; & p < 0.0001 vs CV 24h of all strains; é p<0.0001 vs CV 24h of strains S4 and pepp2; à p<0.05 vs CV 24h of strains S4, RSOLV and pepp2; è p<0.0001 vs CV 24h of strains RSOLV and pepp1; $ p<0.001 vs CV 24h of strain pepp2; * p<0.0001 vs CV 24h of pepp1; @ p<0.0001 vs CV 48h of all strains except S4; “ p<0.0001 vs CV 48h all strains except pepp1; µ p<0.0001 vs CV 48h of all strains; = p<0.0001 vs CV 48h of all strains; L p<0.0001 vs CV 48h of all strains; m p<0.0001 vs CV 48h of all strains; O p<0.0001 vs CV 48h of all strains; p p<0.0001 vs CV 72h of all strains; q p<0.0001 vs CV 72h of all strains except pepp1 and pepp2; n p<0.05 vs CV 72h of all strains; v p<0.0001 vs CV 72h of all strains; I p<0.0001 vs CV 72h of all strains; w p<0.0001 vs CV 72h of 1nm, S4 and RSOLV; Z p<0.05 vs CV 72h of strains 1nm, S4 and RSOLV.
Figure 4.
Biofilm formation (expressed as absorbance at 595 nm) of the Lpb. plantarum strains after 24h and 72h of incubation at 37°C using Resazurin (RZ) (A) or Crystal Violet (CV) (B). Error bars indicate the standard deviation of triplicate experiments.ANOVA was used between strains and to control at each time (24h, 48h and 72h).b p<0.05 vs RZ 24h of all strains except pepp2; T p<0.05 vs RZ 24 of strains 3nm, 1nm and pepp2; H p<0.05 vs RZ 24h of strains V583, RSOLV and pepp1; G p<0.05 vs RZ 72h of strains V583, RSOLV and pepp1; & p < 0.0001 vs CV 24h of all strains; é p<0.0001 vs CV 24h of strains S4 and pepp2; à p<0.05 vs CV 24h of strains S4, RSOLV and pepp2; è p<0.0001 vs CV 24h of strains RSOLV and pepp1; $ p<0.001 vs CV 24h of strain pepp2; * p<0.0001 vs CV 24h of pepp1; @ p<0.0001 vs CV 48h of all strains except S4; “ p<0.0001 vs CV 48h all strains except pepp1; µ p<0.0001 vs CV 48h of all strains; = p<0.0001 vs CV 48h of all strains; L p<0.0001 vs CV 48h of all strains; m p<0.0001 vs CV 48h of all strains; O p<0.0001 vs CV 48h of all strains; p p<0.0001 vs CV 72h of all strains; q p<0.0001 vs CV 72h of all strains except pepp1 and pepp2; n p<0.05 vs CV 72h of all strains; v p<0.0001 vs CV 72h of all strains; I p<0.0001 vs CV 72h of all strains; w p<0.0001 vs CV 72h of 1nm, S4 and RSOLV; Z p<0.05 vs CV 72h of strains 1nm, S4 and RSOLV.
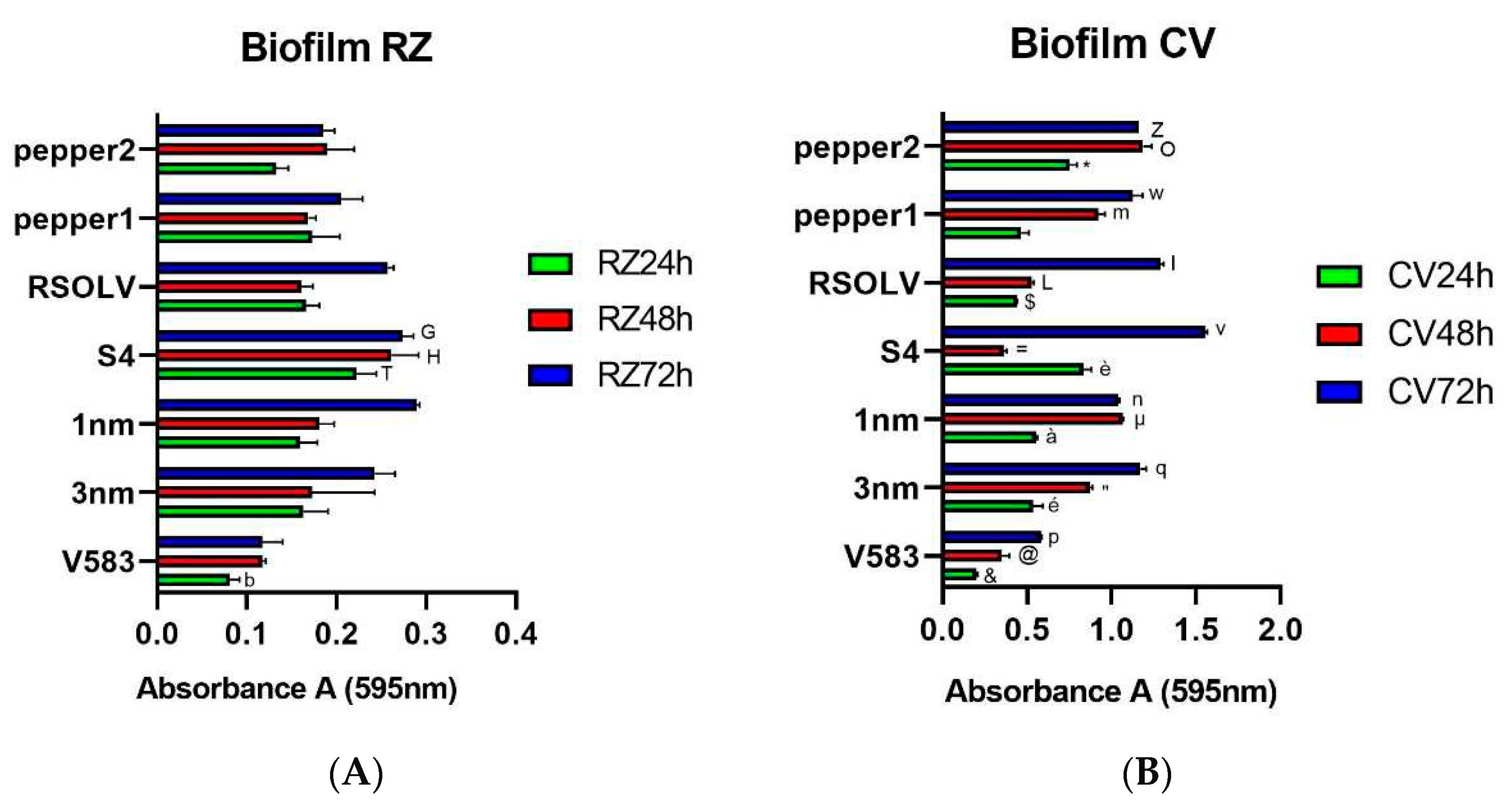
Figure 5.
The effect of different concentration of CFS of the Lpb. plantarum strains on the proliferation of LS cells after 24h (A) and 72h (B) incubation at 37°C. Error bars indicate the standard deviation of triplicate experiments. One-way ANOVA test at each concentration comparing strains among them and to the control (MRS). * p < 0.05 vs all 3nm and all MRS doses; ù p<0.05 vs all 1nm and all MRS doses; µ p<0.05 vs all S4 and all doses; $ p<0.05 vs all RSOLV and all MRS doses; à p<0.005 vs all pepp1 and all MRS doses; ç p<0.05 vs all pepp2 and all MRS doses. + p<0.0001 vs 1mg/ml MRS;& p<0.0001 vs 2mg/ml MRS; “ p<0.0001 vs 4mg/ml of all strains; £ p<0.05 vs 4mg/ml of the strains 3nm, S4, RSOLV, pepp1; / p<0.05 vs 4mg/ml of the strains 1nm;< p<0.05 vs 4mg/ml of S4, RSOLV, pepp1; ; p<0.0001 vs 8mg/ml of all strains; // p<0.05 vs 8mg/ml of strains 3nm, S4, RSOLV and pepp1; ^ p<0.0001 vs 10mg/ml of all strains; ) p<0.05 vs 10 mg/ml of the strains 1nm, S4, RSOLV; ( p<0.05 vs 10mg/ml of the strains pepp1 and pepp2; mp<0.05 vs 10mg/ml of the strains RSOLV and pepp1; np<0.05 vs 10mg/ml of the strains pepp1 and pepp2; l p<0.05 vs 2mg/ml of all strains; - p<0.05 vs 2mg/ml of the strains S4 and pepp1; v p<0.05 vs 4mg/ml of all strains; p p<0.05 vs 4mg/ml of the strains 1nm, S4, RSOLV; h p<0.05 vs 4mg/ml of the strains RSOLV, pepp1 and pepp2; XX p<0.05 vs 4mg/ml of all strains; Y p<0.05 vs 4mg/ml of the strain pepp1; U p<0.001 vs 8mg/ml of all strains; F p<0.05 vs 8mg/ml of all strains; R p<0.05 vs 8mg/ml of all strains; O p<0.05 vs 4mg/ml of the strains 3nm,1nm,pepp1 and pepp2; G p<0.05 vs 8mg/ml of all strains; Q p<0.05 vs 8mg/ml of all strains; Wp<0.001 vs 10mg/ml of all strains; X p<0.05 vs 10mg/ml pepp2.
Figure 5.
The effect of different concentration of CFS of the Lpb. plantarum strains on the proliferation of LS cells after 24h (A) and 72h (B) incubation at 37°C. Error bars indicate the standard deviation of triplicate experiments. One-way ANOVA test at each concentration comparing strains among them and to the control (MRS). * p < 0.05 vs all 3nm and all MRS doses; ù p<0.05 vs all 1nm and all MRS doses; µ p<0.05 vs all S4 and all doses; $ p<0.05 vs all RSOLV and all MRS doses; à p<0.005 vs all pepp1 and all MRS doses; ç p<0.05 vs all pepp2 and all MRS doses. + p<0.0001 vs 1mg/ml MRS;& p<0.0001 vs 2mg/ml MRS; “ p<0.0001 vs 4mg/ml of all strains; £ p<0.05 vs 4mg/ml of the strains 3nm, S4, RSOLV, pepp1; / p<0.05 vs 4mg/ml of the strains 1nm;< p<0.05 vs 4mg/ml of S4, RSOLV, pepp1; ; p<0.0001 vs 8mg/ml of all strains; // p<0.05 vs 8mg/ml of strains 3nm, S4, RSOLV and pepp1; ^ p<0.0001 vs 10mg/ml of all strains; ) p<0.05 vs 10 mg/ml of the strains 1nm, S4, RSOLV; ( p<0.05 vs 10mg/ml of the strains pepp1 and pepp2; mp<0.05 vs 10mg/ml of the strains RSOLV and pepp1; np<0.05 vs 10mg/ml of the strains pepp1 and pepp2; l p<0.05 vs 2mg/ml of all strains; - p<0.05 vs 2mg/ml of the strains S4 and pepp1; v p<0.05 vs 4mg/ml of all strains; p p<0.05 vs 4mg/ml of the strains 1nm, S4, RSOLV; h p<0.05 vs 4mg/ml of the strains RSOLV, pepp1 and pepp2; XX p<0.05 vs 4mg/ml of all strains; Y p<0.05 vs 4mg/ml of the strain pepp1; U p<0.001 vs 8mg/ml of all strains; F p<0.05 vs 8mg/ml of all strains; R p<0.05 vs 8mg/ml of all strains; O p<0.05 vs 4mg/ml of the strains 3nm,1nm,pepp1 and pepp2; G p<0.05 vs 8mg/ml of all strains; Q p<0.05 vs 8mg/ml of all strains; Wp<0.001 vs 10mg/ml of all strains; X p<0.05 vs 10mg/ml pepp2.
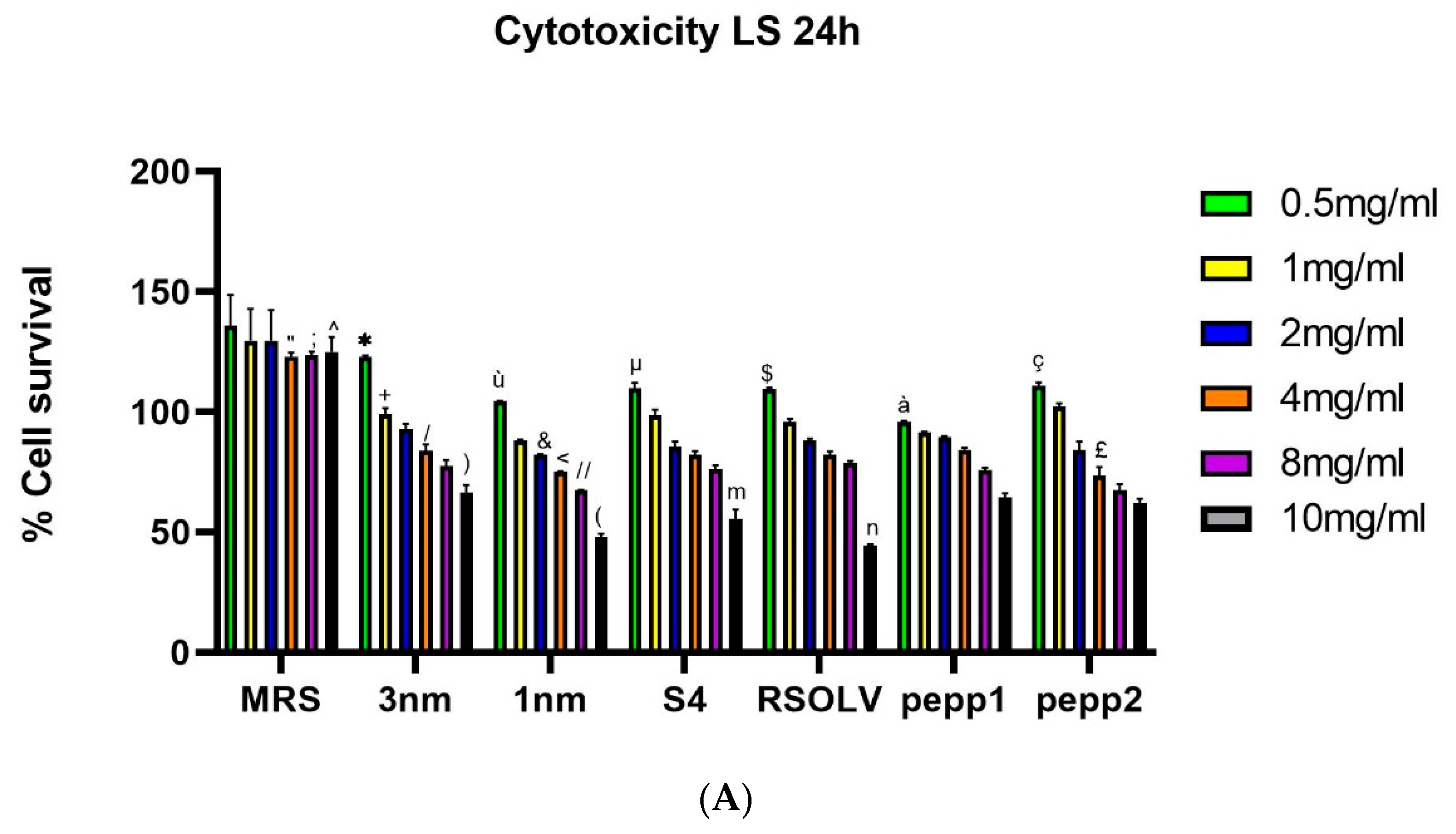
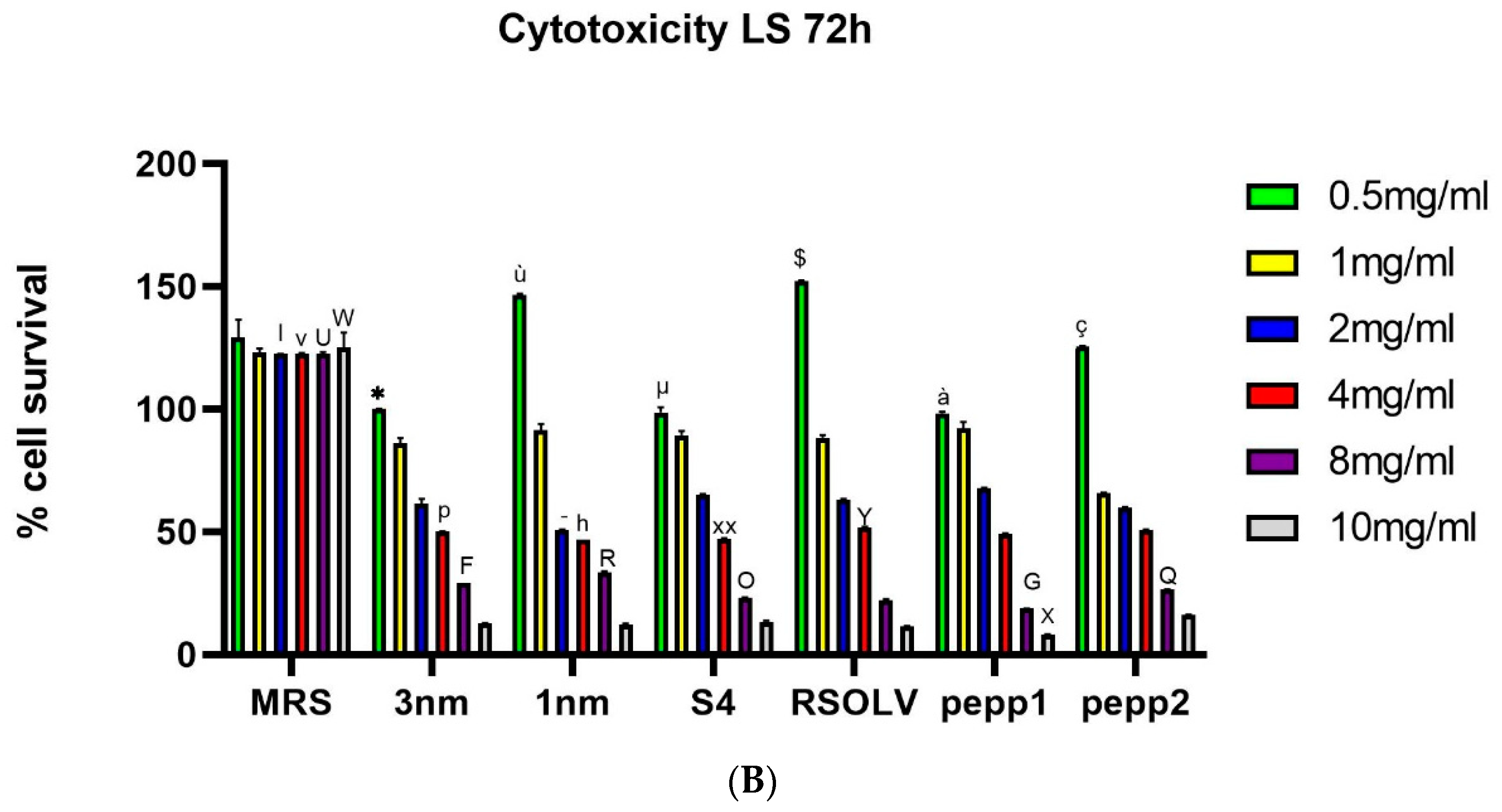
Figure 6.
Adhesion percentage of Lpb. plantarum strains to caco-2 monolayers after 1 h of co-incubation at 37 °C. Lpb. plantarum V299 was used as control. Error bars indicate the standard deviation of triplicate experiments.*: p < 0.05 vs Lpb. plantarum 299V.
Figure 6.
Adhesion percentage of Lpb. plantarum strains to caco-2 monolayers after 1 h of co-incubation at 37 °C. Lpb. plantarum V299 was used as control. Error bars indicate the standard deviation of triplicate experiments.*: p < 0.05 vs Lpb. plantarum 299V.
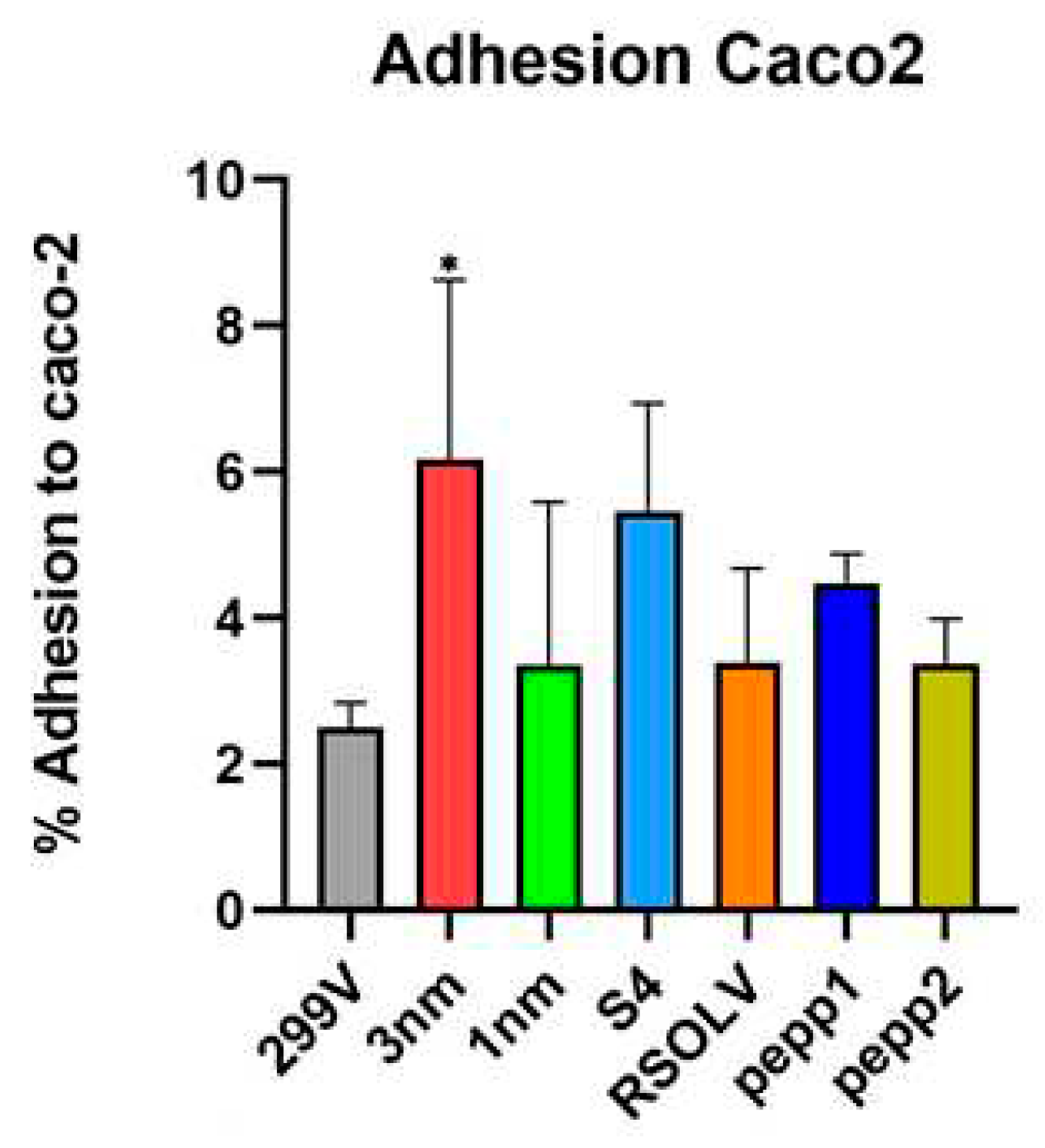
Figure 7.
Adhesion percentage/capacity of Lpb. plantarum strains on Tomato after 2, 3 and 5 days. Error bars indicate the standard deviation of Triplicate experiments. One-way ANOVA test for strains at each incubation time (2days, 3days and 5 days), Tukey’s multiple comparisons test, * p < 0.05; ** p<0.0001.
Figure 7.
Adhesion percentage/capacity of Lpb. plantarum strains on Tomato after 2, 3 and 5 days. Error bars indicate the standard deviation of Triplicate experiments. One-way ANOVA test for strains at each incubation time (2days, 3days and 5 days), Tukey’s multiple comparisons test, * p < 0.05; ** p<0.0001.
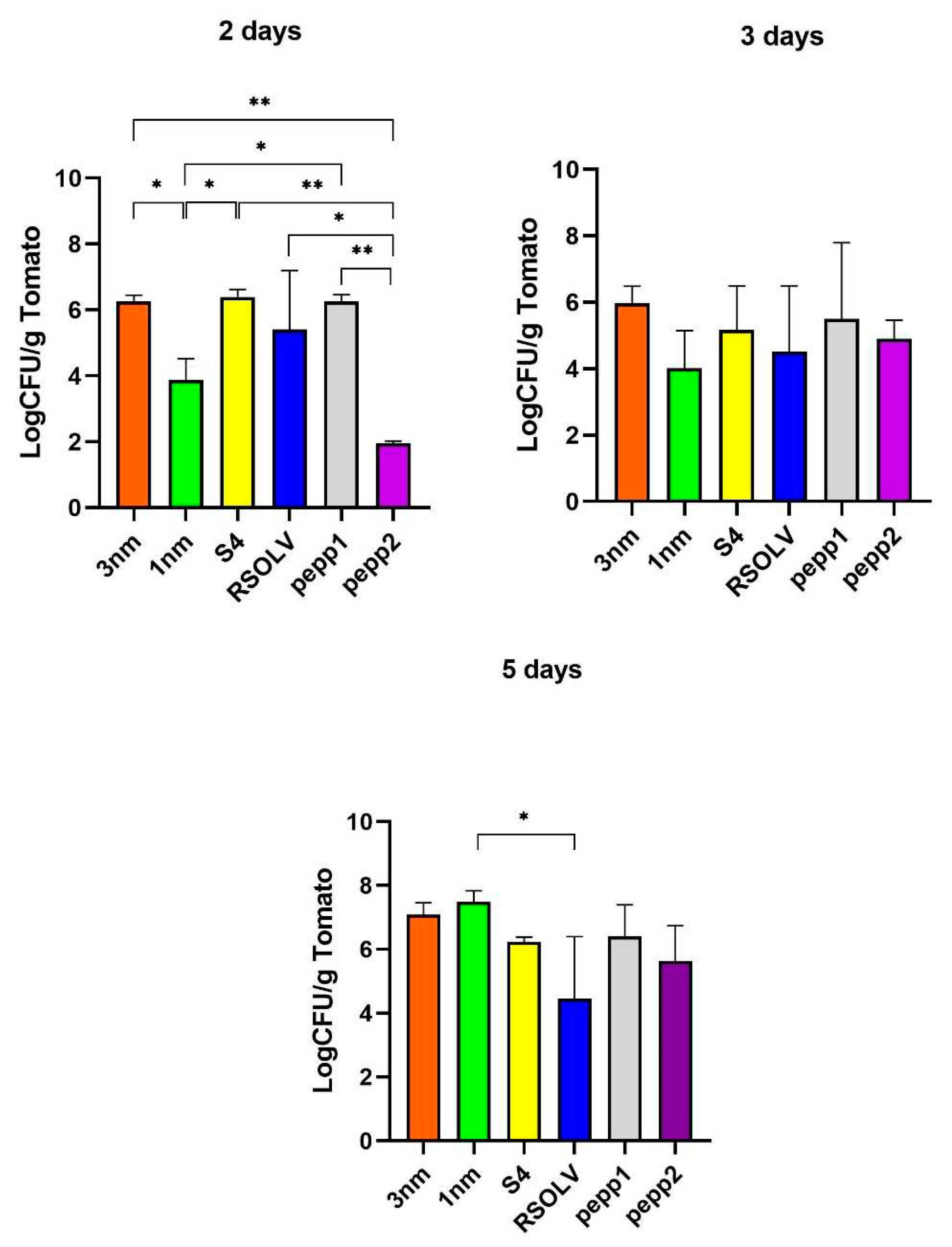

Table 1.
Antibacterial activity of LAB isolated from various matrices against Escherichia coli O157:H7 CECT 4267 and Listeria monocytogenes CECT 4031, as determined by the agar diffusion method. Diameter halos inhibition expressed in mm. Mean values and standard deviations of three replicates are indicated. The six strains selected for further studies are underlined.
Table 1.
Antibacterial activity of LAB isolated from various matrices against Escherichia coli O157:H7 CECT 4267 and Listeria monocytogenes CECT 4031, as determined by the agar diffusion method. Diameter halos inhibition expressed in mm. Mean values and standard deviations of three replicates are indicated. The six strains selected for further studies are underlined.
| Isolated LAB strain | Source/matrix | E. coli (mm) | L. monocytogenes (mm) |
|---|---|---|---|
| 3nm | Intestines of died Locust | 14.17±0.29 | 13.17±0.57 |
| 1nm | Intestines of died Locust | 15.00±0.00 | 14.57±2.32 |
| S4 | Fermented green olive brine | 15.00±1.00 | 11.23±0.68 |
| RSOLV | Fermented olive | 14.17±0.76 | 13.43±0.93 |
| pepp1 | Fermented pepper brine | 15.00±0.00 | 15.10±0.26 |
| pepp2 | Fermented green pepper brine | 15.00±0.00 | 14.50±0.50 |
| S5 | Horse sausage | 10.00±1.00 | 12.07±0.40 |
| S6 | Horse sausage | 0.00±0.00 | 0.00±0.00 |
| N8 | Dried anchovy | 11.00±1.00 | 0.00±0.00 |
| N4c | Dried fermented anchovy | 12.00±1.00 | 8.03±0.25 |
| F1c | Infant feces | 9.13±0.71 | 0.00±0.00 |
| F5a | Infant feces | 12.90±0.17 | 9.97±0.15 |
| LM | Breast milk | 9.83±0.29 | 9.92±0.14 |
| F1 | Infant feces | 10.17±0.29 | 11.00±0.00 |
| Rg4a | Artisanal Tunisian Ricotta cheese | 10.17±0.29 | 11.17±1.26 |
| RL4 | Tunisian fermented milk Leben | 8.83±0.29 | 12.93±0.12 |
| K10 | Tunisian Artisanal Gueddid | 0.00±0.00 | 10.60±0.36 |
| LC4 | Goat milk | 0.00±0.00 | 10.50±0.50 |
| S1 | Fermented olive brine | 10.80±0.36 | 10.47±0.50 |
| AIB | Intestines of rabbit | 0.00±0.00 | 11.00±0.00 |
| S5 | Horse sausage | 11.00±1.00 | 11.00±0.87 |
| N8 | Dried anchovy | 9.90±0.10 | 9.00±0.00 |
| K10 | Tunisian Gueddid | 0.00±0.00 | 0.00±0.00 |
| O3 | Brine fermented olive | 0.00±0.00 | 0.00±0.00 |
Table 2.
The closest species/strain to the identified isolates according to the 16S rRNA sequences.
| Isolate | Source | Closest species/strain | Percentage of identity | Accession number* |
|---|---|---|---|---|
| 3nm | Died locust (intestines) | Lactiplantibacillus plantarum | 100% | OR431596 |
| 1nm | Died locust (intestines) | Lactiplantibacillus plantarum | 99.86% | OR431597 |
| S4 | Fermented Olive | Lactiplantibacillus plantarum | 99.91% | OR431698 |
| RSOLV | Fermented olive | Lactiplantibacillus plantarum | 99.93% | OR431599 |
| pepp1 | Fermented pepper | Lactiplantibacillus plantarum | 99.82% | OR431600 |
| pepp2 | Fermented pepper | Lactiplantibacillus plantarum | 100% | OR431601 |
* Accession number of the sequences of the bacterial isolates in this study, deposited in the NCBI database.
Table 3.
Physiological and Biochemical Characteristics of the selected Lpb. plantarum strains after growth in different conditions: osmotic stress, acidic stress and growth in different sources of carbon e.g glucose (Glc), fructose (Fruc) and Sucrose (Sucr).
Table 3.
Physiological and Biochemical Characteristics of the selected Lpb. plantarum strains after growth in different conditions: osmotic stress, acidic stress and growth in different sources of carbon e.g glucose (Glc), fructose (Fruc) and Sucrose (Sucr).
| Experimental assay | 3nm | 1nm | S4 | RSOLV | pepp1 | Pepp2 |
|---|---|---|---|---|---|---|
| Mobility | - | - | - | - | - | - |
| 2% NaCl growth | + | + | + | + | + | + |
| 4% NaCl growth | + | + | + | + | + | + |
| 8% NaCl growth | + | + | + | + | + | + |
| MRS.Glc 2% | + | + | + | + | + | + |
| MRS.Glc 4% | + | + | + | + | + | + |
| MRS.Fruc 2% | + | + | + | + | + | + |
| MRS.Fruc 4% | + | + | + | + | + | + |
| MRS.Sucr 2% | + | + | + | + | + | + |
| MRS.Sucr 4% | + | + | + | + | + | + |
| MRS pH 3 | + | + | + | + | + | + |
| MRS pH 4 | + | + | + | + | + | + |
| MRS pH 5 | + | + | + | + | + | + |
| MRS pH 9.2 | + | + | + | + | + | + |
Table 4.
(1). Antibacterial activity of whole cultures, CFS and cells from the selected Lpb. plantarum strains; the CFS were used crude, or after neutralization with NaOH, Lpb. plantarum cells were tested before and after heat treatment. The indicator bacteria was Escherichia coli CECT 4267. No(-), mild(+), or strong (++) inhibition, showing zone lower than 7 mm, ranging from 7 to 10 mm, or more than 10 mm, respectively. Assays were performed in duplicate. (2) Antibacterial activity of whole cultures, CFS and cells from the selected Lpb. plantarum strains; the CFS were used crude, or after neutralization with NaOH; Lpb. plantarum cells were tested before and after heat treatment. The indicator bacteria was Listeria monocytogenes CECT 4031. No(-), mild(+), or strong (++) inhibition, showing zone lower than 7 mm, ranging from 7 to 10 mm, or more than 10 mm, respectively. Assays were performed in duplicate.
Table 4.
(1). Antibacterial activity of whole cultures, CFS and cells from the selected Lpb. plantarum strains; the CFS were used crude, or after neutralization with NaOH, Lpb. plantarum cells were tested before and after heat treatment. The indicator bacteria was Escherichia coli CECT 4267. No(-), mild(+), or strong (++) inhibition, showing zone lower than 7 mm, ranging from 7 to 10 mm, or more than 10 mm, respectively. Assays were performed in duplicate. (2) Antibacterial activity of whole cultures, CFS and cells from the selected Lpb. plantarum strains; the CFS were used crude, or after neutralization with NaOH; Lpb. plantarum cells were tested before and after heat treatment. The indicator bacteria was Listeria monocytogenes CECT 4031. No(-), mild(+), or strong (++) inhibition, showing zone lower than 7 mm, ranging from 7 to 10 mm, or more than 10 mm, respectively. Assays were performed in duplicate.
| E. coli | |||||||
|---|---|---|---|---|---|---|---|
| Strain | LAB culture |
Crude CFS |
CFS pH neutralized | CFS treated Temperature | Crude cells | Cells treated Temperature | |
| 3nm | ++ | ++ | - | ++ | - | - | |
| 1nm | ++ | ++ | - | ++ | - | - | |
| RSOLV | ++ | ++ | - | ++ | - | - | |
| S4 | ++ | ++ | - | ++ | - | - | |
| pepp1 | ++ | ++ | - | ++ | - | - | |
| pepp2 | ++ | ++ | - | ++ | - | - | |
| L. monocytogenes | |||||||
| Strain | LAB culture |
Crude CFS |
CFS pH neutralized | CFS treated Temperature | Crude cells | Cells treated Temperature | |
| 3nm | ++ | ++ | - | ++ | - | - | |
| 1nm | ++ | ++ | - | ++ | - | - | |
| RSOLV | ++ | ++ | - | ++ | - | - | |
| S4 | ++ | ++ | - | ++ | - | - | |
| pepp1 | ++ | ++ | - | ++ | - | - | |
| pepp2 | ++ | ++ | - | ++ | - | - | |
Table 5.
Concentration of the main organic acids in CFSs from 24 h cultures of the selected Lpb. plantarum strains grown in MRS broth, as determined by HPLC. Results (mg L−1) are presented as the mean and SD of the two measures.
Table 5.
Concentration of the main organic acids in CFSs from 24 h cultures of the selected Lpb. plantarum strains grown in MRS broth, as determined by HPLC. Results (mg L−1) are presented as the mean and SD of the two measures.
| Tartaric acid | Malic acid | Ascorbic acid | Lactic acid | Acetic acid | |
|---|---|---|---|---|---|
| MRS | 503.27±20.85 | 3 786.64±95.81 | 51.80±1.30 | 0.00±0.00 | 17 906.53±162.88 |
| 3nm | 1 202.55±30.22 | 1 326.48±176.26 | 100.09±42.11 | 14 931.50±133.44 | 16 421.03±2 236.89 |
| 1nm | 1 347.92±25.81 | 1 808.24±42.34 | 102.69±5.66 | 14 620±214.70 | 17 033.25±877.62 |
| S4 | 1 218.73±42.21 | 1 294.83±58.28 | 117.81±1.86 | 15 419.91±89.09 | 16 312.59±1 242.27 |
| RSOLV | 677.69±128.32 | 1 735.10±171.48 | 0.00±0.00 | 14 985.98±195.53 | 19 853.59±541.76 |
| pepp1 | 1 201.36±89.76 | 1 309.51±9.18 | 135.44±4.85 | 13 829.46±132.96 | 19 888.39±809.60 |
| pepp2 | 1 092.72±116.94 | 1 335.78±99.67 | 0.00±0.00 | 15 439.67±140.01 | 42 409.27±4 762.60 |
| Succinic acid | Fumaric acid | Citric acid | |||
| MRS | 1 066.96±27.55 | 0.00±0.00 | 1 206.78 | ||
| 3nm | 5 758.93±729.88 | 15.47±6.71 | 2 662.19±403.92 | ||
| 1nm | 11 693.70±708.58 | 0.00±0.00 | 1 833.64±134.04 | ||
| S4 | 9 009.26±1 374.46 | 0.00±0.00 | 2 224.48±19.15 | ||
| RSOLV | 10 193.35±562.11 | 0.00±0.00 | 1 050.29±200.52 | ||
| pepp1 | 6 897.52±781.84 | 0.00±0.00 | 446.71±45.63 | ||
| pepp2 | 7 336.16±473.52 | 0.00±0.00 | 894.15±25.05 | ||
Table 6.
Antifungal activity of Lpb. plantarum strains against P. expansum, A. niger, F. culmorum CECT 2148, S. cerevisiae and B. cinerea CECT 20973, as determined by the overlay method. No (-), mild (+), or strong (++) inhibition, showing zone lower than 1 mm, ranging from 1 to 5 mm, or more than 5 mm, respectively. Assays were performed in duplicate.
Table 6.
Antifungal activity of Lpb. plantarum strains against P. expansum, A. niger, F. culmorum CECT 2148, S. cerevisiae and B. cinerea CECT 20973, as determined by the overlay method. No (-), mild (+), or strong (++) inhibition, showing zone lower than 1 mm, ranging from 1 to 5 mm, or more than 5 mm, respectively. Assays were performed in duplicate.
| Strain | P. expansum | A. niger | B. cinerea CECT 20973 | F. culmorum | S. cerevisiae |
|---|---|---|---|---|---|
| 3nm | + | ++ | + | ++ | - |
| 1nm | + | ++ | + | + | - |
| RSOLV | ++ | + | ++ | ++ | - |
| S4 | ++ | ++ | + | + | - |
| pepp1 | ++ | ++ | + | + | - |
| pepp2 | ++ | ++ | ++ | ++ | - |
Table 7.
Auto-aggregation ability of Lpb. plantarum strains after 4 h and 24 h incubation in 37 °C in PBS at pH 7. The assay was performed in duplicate.
Table 7.
Auto-aggregation ability of Lpb. plantarum strains after 4 h and 24 h incubation in 37 °C in PBS at pH 7. The assay was performed in duplicate.
| T4 | T24 | |
|---|---|---|
| 3nm | 24.31%±6.84% | 53.46%±2.01% |
| 1nm | 16.82%±2.33% | 53.59%±12.61% |
| S4 | 18.19%±2.54% | 70.30%±4.73% |
| RSLOV | 28.87%±3.68% | 60.97%±5.67% |
| pepp1 | 21.50%±1.75% | 34.67%±0.99% |
| pepp2 | 16.70%±1.12% | 69.82%±2.20% |
Table 8.
Co-aggregation of Lpb. plantarum index with the pathogen Listeria monocytogenes CECT 4031 and E. coli after 4 h and 24 h incubation at 37°C in PBS pH7. Assays were performed in duplicate.
Table 8.
Co-aggregation of Lpb. plantarum index with the pathogen Listeria monocytogenes CECT 4031 and E. coli after 4 h and 24 h incubation at 37°C in PBS pH7. Assays were performed in duplicate.
| L. monocytogenes | E. coli | |||
|---|---|---|---|---|
| Strain | T4 | T24 | T4 | T24 |
| 3nm | 7.94%±0.94% | 7.28%±0.47% | 17.41%±1.54% | 43.42%±6.08% |
| 1nm | 6.38%±1.21% | 5.52%±0.18% | 17.73% ±7.75% | 54.88%±6.98% |
| S4 | 7.47%±0.14% | 7.37%±0.46% | 15.05%±0.89% | 36.96%±5.72% |
| RSOLV | 7.169%±2.25% | 8.76%±1.16% | 18.27%±4.32% | 51.15%±4.69% |
| pepp1 | 5.38%±0.00% | 5.38%±1.34% | 20.72%±2.67% | 43.53%±0.00% |
| pepp2 | 4.99%±0.55% | 4.61%±0.79% | 28.30%±10.70% | 43.16%±1.65% |
Table 9.
Antibiotic resistance assay. The experiment was done in duplicate. The inhibition zone diameters were measured. Susceptibility was expressed in terms of Resistant (R), Susceptible (S) and Intermediate (I) as mentioned.
Table 9.
Antibiotic resistance assay. The experiment was done in duplicate. The inhibition zone diameters were measured. Susceptibility was expressed in terms of Resistant (R), Susceptible (S) and Intermediate (I) as mentioned.
| ATB/strain | 3nm | 1nm | S4 | RSOLV | pepp1 | pepp2 |
|---|---|---|---|---|---|---|
| Ampicillin | I | S | I | I | S | I |
| Vancomycin | R | R | S | R | R | R |
| Gentamycin | I | I | R | S | S | I |
| Kanamycin | R | R | R | R | R | R |
| Streptomycin | R | S | R | R | R | R |
| Tetracyclin | R | R | R | R | R | R |
| Erythromycin | I | R | R | I | R | I |
| Clindamycin | I | R | R | I | I | I |
| Claromycin | I | R | R | R | S | I |
Table 10.
Half-maximal inhibitory concentration (IC50) of different CFSs on the proliferation of LS cell line after 24h and 72h at 37°C.
Table 10.
Half-maximal inhibitory concentration (IC50) of different CFSs on the proliferation of LS cell line after 24h and 72h at 37°C.
| Strain | IC50 (after 24h) mg/mL | IC50 ( after 72h) mg/mL |
|---|---|---|
| 3nm | 12.59 | 4.00 |
| 1nm | 10.21 | 2.40 |
| S4 | 9.91 | 3.21 |
| RSOLV | 11.11 | 3.66 |
| pepp1 | 18.26 | 3.19 |
| pepp2 | 14.88 | 3.69 |
Table 11.
Pathogen antagonism by Lpb. plantarum on tomatoes. CFU counts of pathogens inoculated alone (control) or in presence of each of the indicated Lpb. plantarum strain are reported. Mean ± standard deviation from from three different experiments. Each experiment was done in duplicate. Two-way ANOVA, Dunnett’s multiple comparisons test comparing strains at different time. $ p<0.05 vs 5 days 1nm-Listeria and 5 days pepp1-Listeria; & p<0.05 vs 1 day RSOLV-E.coli and 1 day pepp1-E.coli.
Table 11.
Pathogen antagonism by Lpb. plantarum on tomatoes. CFU counts of pathogens inoculated alone (control) or in presence of each of the indicated Lpb. plantarum strain are reported. Mean ± standard deviation from from three different experiments. Each experiment was done in duplicate. Two-way ANOVA, Dunnett’s multiple comparisons test comparing strains at different time. $ p<0.05 vs 5 days 1nm-Listeria and 5 days pepp1-Listeria; & p<0.05 vs 1 day RSOLV-E.coli and 1 day pepp1-E.coli.
| Number of tomato-attached bacteria (Log CFU/g tomato) | ||||
|---|---|---|---|---|
| Medium agar | 1 day | 3 days | 5 days | |
| Listeria (control) | PALCAM | 1.68±1.39 | 1.67±1.10 | 1.74±0.70$ |
| 3nm-Listeria | PALCAM | 1.73±0.87 | 1.44±0.74 | 1.52±1.48 |
| 1nm-Listeria | PALCAM | 1.70±1.23 | 1.02±0.62 | 0.00 |
| S4-Listeria | PALCAM | 1.46±0.22 | 1.00±0.67 | 0.00 |
| RSOLV-Listeria | PALCAM | 1.77±0.80 | 1.614±1.38 | 1.18 |
| pepp1-Listeria | PALCAM | 1.49±0.997 | 1.54±1.41 | 1.15±0.45 |
| pepp2-Listeria | PALCAM | 1.70±1.14 | 1.16±0.84 | 1.63±1.16 |
| E.coli (control) | SMAC | 1.92±1.61& | 1.72±0.99 | 1.63±1.54 |
| 3nm-E.coli | SMAC | 1.82±1.13 | 1.55±1.43 | 1.34±0.94 |
| 1nm-E.coli | SMAC | 1.87±1.24 | 1.70±0.97 | 1.38±0.31 |
| S4-E.coli | SMAC | 1.88±1.05 | 1.73±1.15 | 1.13±0.45 |
| RSOLV-E.coli | SMAC | 1.87±1.04 | 1.51±1.33 | 1.56±1.24 |
| pepper1-E.coli | SMAC | 1.81±1.22 | 1.45±1.11 | 1.24±0.45 |
| pepper2-E.coli | SMAC | 1.84±1.15 | 1.53±0.96 | 1.22±0.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



