
Submitted:
10 October 2023
Posted:
16 October 2023
You are already at the latest version
Abstract
Hermetia illucens is a very promising insect due to its ability to convert low-value substrates in highly nutrient feed. Its feeding and nutrition are important issues. This work aimed to study the effect of different substrates composed of coffee silverskin, by-product of the roasting process, enriched with different inclusion of microalgae (5%, 10%, 20%, 25%), Schizochytrium limacinum and Isochrysis galbana, coupled with the assessment of environmental sustainability by LCA. In general, the inclusion of microalgae caused an increase in the growth performance of the larvae, although S. limacinum showed the best results. Higher prepupal weight were observed in larvae fed on 20% and 25% of S. limacinum, shorter development times in larvae fed on 25% of both S. limacinum and I. galbana and higher growth rate were observed in larvae fed on 25% of S. limacinum. Anyway, the inclusion of 10% of S. limacinum showed a small difference with the higher inclusions. Moreover, the 10% of S. limacinum achieved the best waste reduction index. Therefore, considering the greater inclusion of microalgae, the higher the environmental impact of larvae production, the addition of 10% of S. limacinum seems to be the best compromise for the larval rearing.
Keywords:
Hermetia illucens
; growth performance
; sustanability
; coffee sirvelskin
; microalgae
1. Introduction
Coffee is one of the most widely consumed beverages in the world and represents the second most valuable commodity produced by developing countries (after petroleum)1. In 2019/2020, around 10 million tons bags of coffee were consumed worldwide2 which caused a large amount of by-product production (pulp, husk, silver skin, and spent coffee) by this industry. Coffee silverskin (CB), the tegument of the outer layer of the coffee bean, is the only by-product of the roasting process. Since the roasting of 10 tons of coffee produces about 83 kg of CB3 this by-product represents a potential pollutant if simply discharged into the environment4. Coffee silverskin is composed by a high amount of dietary fiber (62%), protein (19%), fat (from 1.6 to 3.3%) and minerals (5% ash). Moreover, this by-product has an antioxidant activity due to the presence of melanoidins and phenolic compounds3,5 (e.g., chlorogenic acid). Different valorization alternatives have been explored to both reduce the environmental impact and increase the added value of CB, such as recovering of functional ingredients for potential applications in food, pharmaceutical and cosmetic industries, direct energy generation, composting, bioenergy and biomaterial production3,6–10.Conversion of CB by larvae of Black Soldier Fly (BSF), Hermetia illucens (Diptera: Stratiomydae) constitutes an interesting valorization alternative answering to the principles of sustainability and circular economy. In fact, BSF larvae are able to convert large quantities of organic by products into valuable nutrients (proteins and lipids) that could be used as ingredients for animal feed11–25. Moreover, their frass (excrement from larvae mixed with substrate residues) can be used as an effective organic fertilizer to develop a more sustainable agriculture26.
The European Commission has, recently, allowed the use of insects as ingredient in aquafeed [Reg. (EU) 2017/893] including BSF. BSF larvae composition guarantees a supply of protein and essential amino acids very similar to conventional protein sources used in aquafeed27,28, but, although they have a high lipid content, the amount of polyunsaturated fatty acids is very low17,23,29–34. However, nutritional composition of BSF larvae, in particular lipid content and fatty acid profile, can be modulated by the feeding substrate in accordance with fish nutritional demands23,35–39. Moreover, the quality of feeding substrate also influences the BSF larval performance and their bioconversion efficiency12,20,24,40–43.
To the best of the authors’ knowledge, very few studies have been conducted on the environmental impact associated with BSF larvae production16,19,44–46. Nowadays the most digestible and nutritious aquafeed ingredients are fish meal and fish oil (FAO, 2020). However, these ingredients are no more sustainable, and alternatives are necessary to promote sustainability while promoting fish welfare47. Feed represents the main production cost in aquaculture48. Researchers highlighted the environmental sustainability of different alternative ingredients by Life Cycle Assessment (LCA), reporting that the protein and lipid sources produced from insects are the most environmentally friendly solutions if compared to other alternatives such as microalgae46. It is noted that LCA analyses consider the most important impact categories like global warming, resource depletion, acidification and eutrophication but neglect some important aspects like the declining of wild fish.
In this contest, in the framework of the Project “New nutrients for the production of valuable fish species -NUTRIFISH”, roasting coffee industry by-product (coffee silverskin-CB), locally available, as basal rearing substrate for BSF larvae was used. This basal rearing substrate was enriched with increasing levels of Schizochytrium limacinum or Isochrysis galbana biomass as sources of valuable proteins and lipids (rich in polyunsatureted fatty acids)49–54. In particular, the present study was designed to investigate: i) larval growth performance, ii) total final insect biomass, iii) mature larvae (prepupae) macronutrient composition, iii) bioconversion efficiency and iv) assessment of the environmental sustainability of the insect biomass production by LCA.
2. Materials and Methods
2.1. Ethics
All procedures involving animals were performed in compliance with the Italian legislation on experimental animals. No specific authorization is requested, by the actual legislation, for invertebrates such as insects.
2.2. Diet preparation
By-products derived from roasting mixture of Coffea arabica and Coffea canephora coffee process, the coffee silverskin (CB), were collected at Saccaria Caffe` S.R.L. (Marina di Montemarciano, Ancona, Italy). Coffee silverskin was packed inside airtight plastic bag, transferred to laboratory, and kept at -20°C to prevent decomposition. Prior application, the CB was adapted to room temperature, and grinded by food mill (Ariete, De’Longhi Appliances Srl, Italy) to 2.0 ± 0.4 mm particle size. The material obtained was then thoroughly mixed with increasing levels (5%, 10%, 20%, 25%) of freeze-dried microalgae, Schizochytrium limacinum (S) or Isochrysis galbana (I) (provided by AlghItaly Società Agricola S.R.L., Sommacampagna, VR, Italy), and demineralized water to prepare experimental at 70% moisture diet for BSF larvae. Feed mixtures of CB:S and CB:I were used as experimental diets (N=8) and CB was used as control (Table 1). Samples of CB, S and I were stored at -20 °C for further analysis.
2.2. Experimental design
BSF larvae were provided by Smart Bugs s.s. [Ponzano Veneto (TV), Italy]. For each experimental diet, 6 groups of 100 six-day old larvae were isolated, cleaned from the initial feeding substrate, hand counted, weighted [RADWAG Wagi Elektroniczne, AS 82/220.X2, Poland] and placed, with the rearing substrate (70 g/replicate, N=6), in plastic containers (10 cm x 17.5 cm x 7 cm)17. Every week, feeding substrate was added, until 40% of prepupae occurred, considering a quantity of feeding rate of 100 mg/day/larva11. The containers, previously weighted, were screened with fine mesh cotton gauze (40 x 30 cm), covered with a lid and, given the high tendency to migration of mature larvae, further wrapped in organza. The lid was provided with a single ventilation hole (4.5 cm Ø)14. Each container was inspected every day and, when necessary, the feeding substrate was moved to promote aeration and to prevent the reduction of its quality. Larvae were maintained under constant conditions in a climatic chamber (T: 27±1 °C, RH: 65±5%, 0:24, L:D photoperiod)14. Once in each replicate the 40% of prepupa occurred, prepupae and mature larvae were manually isolated from the diet using forceps and brushes, cleaned from substrate residues, washed in water, dried on a piece of paper and counted. Prepupae were identified by a change in integument color from larval white to black. The total final insect biomass and the residual rearing substrate (excrements from larvae mixed with substrate residues) were weighted. The final biomass was stored at -20 °C for further analysis.
2.3. Growth performance and conversion efficiency of the BSF larvae
The larval development time was calculated as the number of days between the start of the experiment and the observation of the 40% of prepupa in each replicate.
Larval survival rate was calculated as: Survival Rate (%) = number of larvae survived / number of initial larvae * 100.
Larval growth was measured by biomass gained and expressed in Growth Rate (GR) by the following formula56, in which higher GR indicates a faster growth of larvae:
Growth rate (g/day) = (final weight (g) – initial weight (g)) / time needed to reach prepupal phase (d)
Larvae ability to reduce feed was calculated by WRI (Waste Reduction Index), by the following formula in which high WRI values indicate good reduction efficiency of the diet17 :
WRI = [(total feed (g) – residue(g)) /total feed (g)]/ feeding time (d) * 100
Feed conversion efficiency, that is the amount (kg) of feed needed to obtain one kg of animal weight gain, was calculated as Feed conversion ratio (FCR) by the following formula12 in which low FCR values indicate high food conversion efficiency.
FCR = (distributed substrate (g) − residual substrate (g)) / (total final biomass (g) - total initial biomass (g))
All parameters were calculated on a fresh matter basis12,13,17.
2.4. Diet ingredients and larval composition analysis
Coffee silverskin, S. limacinum and I. galbana freeze dried biomasses, insect diets and insects were analyzed for moisture (Method #950.46), crude protein, CP as Kjeldhal nitrogen (Method #976.05), and ash (Method #920.153) contents according to AOAC (2006) and total lipid according to54, at the laboratories of the University of Udine (Italy).
2.5. Life Cycle Assessment
To support the choice of the most sustainable experimental diet to feed larvae on roasted coffee by-products enriched with two different microalgae species in BSF industrial rearing, a simplified Life Cycle Assessment (LCA) of the insect biomass production has been carried out to have a first indication of the less impacting option from an environmental point of view. To this aim LCA methodology has followed the ISO 14040 and 14044 standards requirements.
2.5.1. Goal and Scope
The goal was to quantify the environmental impact of the BSF insect biomass production based on four (5%, 10%, 20%, 25%) freeze-dried microalgae inclusion levels in the basal rearing substrate, a by-product of the roasted coffee industry (coffee silverskin-CB). The alternative functional units of 1 kg protein and 1 kg lipid were added as performed by other authors19 considering that the protein and lipid contents of final BSF biomass reared on the different diets were assessed as reported in subparagraph 2.4. The functional unit was 1 kg of freeze-dried insect biomass produced, in line with scientific literature to ease results comparison. The system boundaries of the analysis are reported in Figure 1.
For the microalgae production, primary data were provided by an Italian producer while for insect production were used the results of the experiments carried out by Entomologists of the Dipartimento di Scienze Agrarie, Alimentari ed Ambientali—Università Politecnica delle Marche and previously reported, integrated with secondary data taken from literature16,44,45,57–59 and internationally accepted LCA databases (Ecoinvent v. 3.5, Agri-footprint v. 4.0).
2.5.2. Life Cycle Inventory
For the microalgae production, site investigations were carried out to acquire all the data related to the production process based on photobioreactors process and the related inputs, outputs and emissions. It was not possible to get specific data for the two different microalgae species, but average values were collected. In details the following information has been collected by specific questionnaires as primary data: water, chemicals and energy consumption, wastewater produced. These primary data cannot be reported due to confidentiality issues.
For the insect biomass production, the data were collected at the Dipartimento di Scienze Agrarie, Alimentari ed Ambientali, Università Politecnica delle Marche – Entomology laboratories. Primary data are related to the diet ingredients used (amount of microalgae and coffee silverskin employed for the formulation of each diet), to the relative output including the amount of insect biomass and residual rearing substrate (excrement from larvae mixed with substrate residues and exuviae) produced and the related FCR values. For water, energy consumption and direct greenhouse gas emissions, secondary data were used45,58.
2.5.3. Life Cycle Impact Assessment
The LCA results were calculated by the SimaPro 9 software application [PRé Sustainability B.V., Amersfoort, The Netherlands, 2021]. The Life Cycle Impact Assessment method ReCiPé 2016 has been used, in line with other similar studies. The following impact categories have been considered: Global Warming, Freshwater Eutrophication, Water Use, Land Use/transformation, Mineral Resource (materials) and Fossil Resource (energy).
2.6. Statistical analysis
Statistical analyses were performed using the R statistical programming. Data recorded on prepupal weight, time of development, survival rate, Growth Rate (GR), Waste Reduction Index (WRI), Feed Conversion Ratio (FCR) and chemical composition of BSF larvae fed on the eight different experimental diets and on the control diet (Table 1) were compared by means of Kruskal-Wallis test or by one-way analysis of variance (ANOVA), according to data distribution. The pairwise Wilcoxon post-hoc test or the Tukey-Kramer’s Honestly Significant Difference (HSD) multiple comparison post-hoc test, respectively, were applied for the mean separation (p < 0.05) between the tested diets. Data normality was assessed by the Shapiro-Wilk test and multiple testing correction was performed using the Benjamini-Hochberg (BH) method.
3. Results
3.1. Growth performance and conversion efficiency of the BSF larvae
The effect of the different tested diets on BSF growth performance and quality attributes is reported in Table 2 and Table 3.
All the experimental diets were found to significantly enhance prepupal weight, developmental time and biomass gained compared to control. At the same inclusion level in the basal substrate (CB), larvae fed on diets enriched with S. limacinum performed better than those with I. galbana in terms of prepupal weight, biomass gained and ability to reduce feeding substrate. Moreover, survival rate of BSF larvae was unaffected by dietary treatments.
In particular, the growth dynamics of BSF larvae fed on the different tested diets, were investigated by considering prepupal weight, survival rate, development time and growth rate (Table 2). In diets enriched with S. limacinum, differences respect to control group were observed in the prepupal weight (Table 2), which increased in a dose-dependent way (the higher the microalga inclusion the higher the insect weight) with a higher weight in larvae fed on 25S (df = 4, p = 0.0001). In diets enriched with I. galbana, prepupal weight increased in larvae fed on all diets respect to control, but 10I and 25I showed the highest prepupal weight (df = 4, p = 5.75x10−5). When comparing all the experimental diets formulated with the addition of different levels of both species of microalgae, authors found S. limacinum to better support insect growth performance than I. galbana, with emphasis on the prepupal weight, especially with regards to larvae fed on 20S and 25S diets (df = 8, p = 2x10−16).
The percentage of microalgae included in the diets significantly influenced the development time of the larvae (Table 2). More specifically, larvae fed on 25S and 25I diets showed shorter development time than those fed on the other tested diets (df = 4, p = 7.82x10−6).
The different concentration of both microalgae significantly influenced the Growth Rate (GR), which indicates high efficiency of larval growth (Table 2). Larvae fed on 25% S. limacinum (25S) showed higher GR values when compared to the other tested diets (df = 4, p =3.65x10−5). Among diets enriched with I. galbana, larvae fed on 10I and 25I showed higher values of GR (df = 4, p = 5.92x10−5).
Both the WRI and the FCR of BSF larvae were affected by different tested diets. Larvae fed on C showed higher values of Food Conversion Ratio (FCR) (df = 8, p = 9.41x10−7) and lower values of Waste Reduction Index (WRI) (df = 8, p = 7.08x10−8) (Table 3). Larvae fed on 10% S. limacinum (10S) showed statistically higher values of WRI when compared to larvae fed on all the other tested diets (df = 4, p = 3.32x10−5). Within the larvae fed on I. galbana group, 20I showed higher values of WRI (df = 4, p = 0.0002). With regard to the FCR, we found that 5S, 20S, 25S, 10I and 25I caused significantly lower values of FCR than all other tested diets (df = 4, p = 0.003).
3.2. Diet ingredient and larva composition analysis
3.2.1. Diet composition analysis
The chemical composition of the experimental diets including coffee silverskin and increasing (5, 10, 20, 25 %) level of S. limacinum and I. galbana is reported in Table 4.
Increasing the addition of S. limacium dry biomass to the coffee silverskin basal diet resulted in an increased protein content that ranged from 20.96 g/100g d.m. of the control diet to 31.07 g/100g d.m. of diet 25S. The dietary lipid content was limited because affected by the addition of S. limacium.
The addition of increasing levels of I. galbana biomass to the coffee by-product as substrate for the growth of H. illucens resulted in an increase of the substrate protein content and a substantial increase in the total lipid level up to 3.56 g/100g corresponding to 25% addition (Diet 25I) relatively to the control diet.
3.2.2. Insect composition analysis
The characterization of the proximal? composition of the BSF larvae reared on the different experimental diets is reported in Table 5.
The inclusion of both microalgae in the basal rearing substrate (coffee silverskin) resulted in a significant modulation of the nutrient composition of BSF larvae.
Compared to the composition of the experimental diets, the larvae of BSF reared on diets containing increasing levels of S. limacinum have shown a proportional increased nutritional value in terms both of protein and lipid content, thus confirming the initial hypothesis that modulating the substrate composition, the effect in nutritional terms is also reflected on the nutritional composition of BSF. The highest protein content was registered in larvae reared on diet 25S (18.3 % CP). On the other hand, the larvae lipid content was significantly affected by the increasing microalgae inclusion level till diet 20S. S. limacinum inclusion level higher than 20% did not significantly changed larvae lipid content (P>0.05)
The proximate composition of the BSF larvae reared on diets containing increasing levels of I. galbana resulted in a significant increase both in terms of protein and lipid content compared to larvae fed on the control diet (P <0.05). However, in larvae there is no corresponding variation in protein levels proportional to increasing levels of inclusion of the microalgal biomass greater than 10%; I. galbana inclusion levels of 25% determined the highest lipid contents in the larvae (4.95%).
3.3. Life Cycle Assessment
The LCA analysis returned the results reported in the Table 6 and Table 7, related to the diets containing S. limacinum and I. galbana respectively. All the values are referred to the same functional unit, 1 kg of dried larvae produced. As a consequence, the values can be directly compared. The variability of the average values resulted always less than 10%.
The results showed how all the considered environmental impacts are strongly affected by the percentage of microalgae inclusion in the diets. The coffee silverskin is a leftover and is not linked to impacts due to production. As a consequence, the lowest impacts were calculated for 5I and 5S diets. The contribution of the larvae drying process is not relevant if compared with the microalgae production, especially for the inclusion of 10% or more in the diet where the microalgae impact contributes more than 80% on the total impacts. The impact of microalgae production is mainly due to the high energy inputs employed and the low level of technology maturity.
The insect residual rearing substrate was not considered as a waste to be managed because it could be used as fertilizer in sustainable agriculture to partially substitute the chemical fertilizer and providing several benefits in terms of plant growth promotion and increased tolerance to abiotic stresses and resistance to pathogens and pests26. The real effect of this specific substitution in terms of the considered impact categories however is difficult to be estimated60 and in this case is limited if compared to the obtained results (e.g., up to a saving of 2 kg CO2 eq / kg of dried larvae produced, for global warming), mainly due to the low nitrogen content of residual rearing substrate (about 4% on dry mass basis). For these reasons this limited and uncertain impact reduction was not included in the calculation.
As an additional indication, the results were expressed also using the alternative functional units related to 1 kg protein (Table 8 and Table 9) and 1 kg lipid, based on the different protein and lipid (Table 10 and Table 11) contents of BSF larvae reared on the different diets reported in Table 5.
The increase in protein and lipid content for BSF larvae fed on the experimental diets containing the highest inclusion of microalgae partly balances the increase of impacts but to a very limited extent.
4. Discussion
Several studies have been carried out on the nutritional needs of BSF and on the evaluation of by-products used as diets15,17,61,62, but much more information is still needed. For this reason, the growth performance of BSF and its environmental impact when raised on potentially eco-sustainable feeding substrates was evaluated. This study provided data on the effect of enriching the coffee silver skin by-product with different microalgae inclusion levels (S. limacinum and I. galbana) on BSF growth performance. These rearing substrates were characterized by a variable chemical composition which influenced the growth performance of the larvae. Considering that BSF stores most of its nutrients during larval development since the adult stage is not able to feed63, it is clear that the quality of the rearing substrate was essential for their fitness41,65. Hence, the nutritional composition of the rearing substrates had a great influence on critical production factors as the survival rate and the weight of the larvae which positively correlates with the growth rate12,61,67,68. Furthermore, because the diet is a critical point for the insect fitness and for the environmental impact, the study of their effect on insects, on the processing conditions and on the properties of the final product is very important16.
In this study, according to Truzzi et al. [54], both microalga species (S. limacinum and I. galbana) showed a crude protein and lipid content higher than the coffee silverskin. Consequently, the best growth performance was achieved at the higher microalgae inclusion levels. Despite both experimental diets significantly increased larval weight and survival ratio, S. limacinum provided greater benefits than I. galbana. In fact, because the composition of the two microalgae was different, S. limacinum resulted in higher amount of protein and lipids respect to I. galbana, with emphasis on unsaturated fatty acids.
A shorter larval developmental time in BSF larvae reared on microalgae-enriched diets (18-20 days), compared to the control diet (coffee silverskin) (39 days) and compared to what is reported by several authors was observed12,41,42,69. The larval developmental time of larvae reared with higher microalgae inclusions (25S and 25I) was 18 days, similar to that reported by Barragan-Fonseca et al. [70]. The development time recorded for the larvae reared on coffee silverskin only was very long, showing that rearing substrates low in fat, rather than protein, negatively affected the larval fitness. This result confirmed what is reported by present literature, namely that to store body lipid needed to complete development, BSF larvae need high-fat diets63,65,71. In fact, if the rearing substrate has a low lipid level, larvae will take longer time to acquire their body fat and complete their development41.
Microalgae-enriched substrates were found to be more effective in comparison to coffee silverskin solely also in terms of increasing the prepupal weight. In fact, the weight of the larvae reared on the experimental diets (average of 0.1363 g) was considerably higher than those reared on the control diet (0.0724 g), and they were comparable to those reported in the literature by different authors41,65,71–73. Accordingly with41,71, we observed that the larval weight increased in relation to both dietary protein and lipid content.
A faster larval growth indicates higher growth rate, thus an advantage for the rearing74. In fact, also the growth rate was better in larvae reared on microalgae-enriched diets. In particular, the highest growth rate was recorded for the S. limacinum enriched diets, followed by the I. galbana enriched ones, while larvae reared on coffee silverskin only showed the lowest value of growth rate. These results confirm that, although BSF needs proteins and lipids in its diet, an excessive percentage of these macromolecules in the substrate (e.g., lipids in I. galbana enriched diets) is often detrimental for the growth rate41,75.
Regarding the survival rate achieved in the present study, for all experimental and control diets, it was in line with the range reported by several authors on a wide variety of rearing substrates12,19,69,71,73.
Nowadays, by-products are becoming increasingly important feed ingredients76, especially as they are considered to have a lower economic value and a lower environmental impact than the main product77. Their advantage also depends on how well these by-products can be converted into body mass by insects12. For this reason, the combination of different substances, including by-products, can be used as efficient diets, and their composition is the main variable to determine Feed Conversion Efficiency (FCR)78. Basically, FCR is the amount of feed needed to obtain one kg of weight increase of the production animal. In order to have better FCRs, which give optimal economic and environmental benefits in the use of BSF as alternative feed, it is necessary that the experimented diets are used directly and effectively by insects. In this work FCR was more favorable for microalgae-enriched diets, thus indicating a better use of feed nutrients. The values recorded in this work (FCR from 4.5±0.5 to 9.5±0.9 for microalgae-enriched diets; 34.2±6.4 for control diet) resulted much higher than FCR recorded by Oonicx et al. [12] (FCR from 1.4±0.1 to 2.6±0.9) that tested by-products derived from food manufacturing diets. Anyway, these values resulted lower than how recorded by Rehman et al. [79] (FCR from 6.3±0.2 to 10.1±0.2) that tested larvae reared on dairy manure and soybean curd residue. These results are in agreement with Oonincx et al. [12] that showed that high protein diets resulted in lower FCRs. Indeed, the composition and the density of the protein are important for insects because they do not use energy to maintain a constant body temperature80,81.
Finally, it was also analyzed the Waste Reduction Index (WRI), a parameter that is used to calculate the ability of the insect to reduce waste and to calculate the conversion efficiency of the diet into biomass. The higher the WRI value, the more effective the larvae are in bio-converting the by-products19. In this work it was observed that the WRI in 10S (2.6±0.2) was higher compared to other diets of the experiment. This WRI values were in agreement with Leong et al. [82], that reared BSF larvae fed on fruit waste collected from a cafeteria (WRI 2.8±0.1), palm decanter cake (WRI 2.8±0.1) but it resulted much lower than how observed by Bava et al. [19] that reared the larvae in okara (WRI 4.9±0.1), maize distiller (WRI 3.2±0.2), hen diet and brewer’s grains diets (WRI 3.0±0.1).
The inclusion of both microalgae biomass in the rearing substrate resulted in a significant modulation of the nutrient composition of BSF larvae. The proximate composition of the BSF larvae reared with diets containing increasing levels of microalgae resulted in a significant increase both in terms of protein and lipid content compared to larvae raised on the only coffee silverskin, thus confirming the hypothesis that modulating the substrate composition, the effect in nutritional terms is also reflected on the nutritional composition of the insect larvae23,35–38. In particular, the BSF larvae reared on diets containing increasing levels of S. limacinum have shown a proportional increased nutritional value in terms both of protein and lipid content.
The protein content of the larvae was significantly affected by the increasing microalgae inclusion level, reaching the highest content in larvae reared on diet 25S and 25I (18.3 % CP). This trend was linear with the increase in protein content observed in the diet proximate composition. The protein content obtained in the present study was lower than that reported by several authors19,30,83, but, in these works, the BSF larvae were reared in much higher proteic diets, which could have influenced the nutritional composition of the larvae.
A different trend was observed for the larval lipid content that showed to be affected by the increasing S. limacinum inclusion level, where the 20I and 25I showed the highest content (7.74 and 7.93 g/100g respectively), and to not be affected by the increasing I. galbana inclusion level. In fact, although diets with inclusions of I. galbana showed a significantly higher lipid content than diets with inclusions of S. limacinum, the biochemical composition of the larvae showed lipid values similar to larvae reared on 5S and 10S diets.
Based on the LCA results, the calculated impact values were high when compared with those reported in literature for BSF production based on different substrates44 for all the impact categories considered. This is mainly due to the high impact of microalgae production, in line with the outcome of other studies84,85. In one research study, microalgae were considered as a less environmentally friendly substitute of insect meal45. Therefore, as a consequence, using microalgae for insect production is very likely to increase the impact of insect production. Anyway, to correctly evaluate the different solutions, the nutritional quality aspects should also be considered simultaneously as reported in the present study. In addition, it should be highlighted that the impact of fishmeal resulted in some specific cases higher than 5S and 5I86 and the negative effect of fishmeal in decreasing the sea fish stocks is not considered in LCA analyses.
To conclude, the data available in the literature do not allow to establish which is the best growth substrate for BSF larvae. Therefore, the results of this work can be considered as an enrichment of the database that collects information on how the performance of BSF larvae can be affected by the substrates. Also, regarding the LCA, as a first indication it could be useful to limit the microalgae percentage or choosing a more sustainable microalgae production. This last option will be probably possible in the future with the improvement of microalgae production technology. It should be noted that significant impact reduction could be obtained in specific scenarios where also the side streams are valorized like spent substrate as compost or where the insect production is considered as an alternative waste management option.
Although this study was conducted under laboratory conditions, it is a good prospect for the use of BSF as a by-product degrader.
5. Conclusions
In the framework of the project Nutrifish, several studies on the rearing chain of BSF larvae and how they were affected by rearing substrates based on coffee silverskin supplemented with increasing percentages of Schizochytrium limacinum or Isochrysis galbana microalgae were carried. Authors analyzed the influence of these feeding substrates looking at fatty acids profile of BSF prepupae (Truzzi et al., 2019 [88]), and the occurrence of antibiotic resistance genes in BSF larvae fed on them (Milanovic et al., 2021 [89]). Moreover, BSF reared on these diets as sustainable terrestrial ingredient for aquafeeds production (Zarantoniello et al., 2019 [90]), studying Zebrafish physiological and behavioral responses to insect-based diets (Zarantoniello et al., 2020 [41]) and the possible dietary effects of these diets across Zebrafish generations (Zarantoniello et al., 2021 [25]). Last, but not the least, this work evaluates the growth performance of BSF and its environmental impact when raised on potentially eco-sustainable feeding substrates in order to propose it as an alternative feed ingredient in aquaculture. This study becomes important, because, to date, several studies have been carried out on the nutritional needs of BSF and on the evaluation of by-products used as diets [Tschirner and Simon, 2015 [63]; Jucker et al., 2017 [15]; Liu et al., 2018 [64]; Meneguz et al., 2018 [17]], but there is still little information about it.
The BSF rearing substrate selection is very important for its growth performance and to have a low environmental impact product. With this work it has been confirmed that the by-product coffee silverskin, a great pollution hazard if discharged into the environment used as diet ingredient in order to be revalued, has low amounts of proteins and lipids to guarantee a good growth performance of BSF. However, enriching this substrate with S. limacinum and I. galbana, two ingredients containing the right amount of nutrients, solved these problems. Anyway, between the two microalgae, S. limacinum was found to provide greater benefits than I. galbana.
Considering that BSF rearing substrate significantly contributes also to the environmental impact of larvae production, it is very important to deeply study it from the sustainability of the substrate components to its effect for the insects. This is necessary to better evaluate the environmental benefits of the insects as alternative feed ingredients. Due to the limited sustainability of the current microalgae production, the inclusion percentage in the substrate should be limited and could be useful to study also the interaction with other residues.
Author Contributions
Conceptualization, P.R. and I.O.; methodology, P.R., S.R., F.T. and D.D.; formal analysis, S.R, D.D., F.T. and D.B..; investigation, S.R., P.R., G.C., F.T. and D.D.; help in investigations: L.C., M.Z.; resources, P.R. and N.I.; data curation, P.R., S.R., F.T. and D.D.; writing—original draft preparation, S.R. and D.D.; and writing—review and editing, S.R., P.R, F.T., G.C., I.O., M.Z. and D.D.; visualization, P.R. and S.R.; supervision, P.R.; project administration, P.R.; funding acquisition, I.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by FONDAZIONE CARIVERONA, Ricerca Scientifica 2017, project NUTRIFISH, code n. 2017.0571.
Acknowledgments
Authors would like to thank Saccaria srl and Alghitaly for providing coffee silverskin and microalgae, rispectively.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tucker, C.M. Coffee Culture: Local Experiences, Global Connections, 2nd ed.; Taylor & Francis: Abingdon, UK, 2017; ISBN 9781317392255. [Google Scholar]
- ICO. Historical data on the global coffee trade. 2020. Available online: http://www.ico.org/new_historical.asp?section= Statistics (accessed on 17 November 2020).
- Alves, R.C.; Rodrigues, F.; Antónia Nunes, M.; Vinha, A.F.; Oliveira, M.B.P.P. State of the art in coffee processing by-products. In Handbook of Coffee Processing By-Products; Elsevier, 2017; pp. 1–26 ISBN 978-0-12-811290-8.
- Mussatto, S.I.; Machado, E.M.S.; Martins, S.; Teixeira, J.A. Production, composition, and application of coffee and its industrial residues. Food Bioprocess Technol. 2011, 4, 661–672. [Google Scholar] [CrossRef]
- Tores De La Cruz, S.; Iriondo-DeHond, A.; Herrera, T.; Lopez-Tofiño, Y.; Galvez-Robleño, C.; Prodanov, M.; Velazquez-Escobar, F.; Abalo, R.; Castillo, M.D.D. An assessment of the bioactivity of coffee silverskin melanoidins. Foods 2019, 8, 68. [Google Scholar] [CrossRef]
- Ballesteros, L.F.; Teixeira, J.A.; Mussatto, S.I. Chemical, functional, and structural properties of spent coffee grounds and coffee silverskin. Food Bioprocess Technol. 2014, 7, 3493–3503. [Google Scholar] [CrossRef]
- Sung, S.H.; Chang, Y.; Han, J. Development of polylactic acid nanocomposite films reinforced with cellulose nanocrystals derived from coffee silverskin. Carbohydrate Polymers 2017, 169, 495–503. [Google Scholar] [CrossRef]
- Hijosa-Valsero, M.; Garita-Cambronero, J.; Paniagua-García, A.I.; Díez-Antolínez, R. Biobutanol production from coffee silverskin. Microb. Cell Fact. 2018, 17, 154. [Google Scholar] [CrossRef]
- Bessada, S.M.F.; Alves, R.C.; Costa, A.S.G.; Nunes, M.A.; Oliveira, M.B.P.P. Coffea canephora silverskin from different geographical origins: a comparative study. Science of The Total Environment 2018, 645, 1021–1028. [Google Scholar] [CrossRef]
- Iriondo-DeHond, A.; Iriondo-DeHond, M.; Del Castillo, M.D. Applications of compounds from coffee processing by-products. Biomolecules 2020, 10, 1219. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: establishing optimal feeding rates. Waste Manag. Res. 2009, 27, 603–610. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; Van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]
- Parra Paz, A.S.; Carrejo, N.S.; Gómez Rodríguez, C.H. Effects of larval density and feeding rates on the bioconversion of vegetable waste using black soldier fly larvae Hermetia Illucens (L.), (Diptera: Stratiomyidae). Waste Biomass Valor. 2015, 6, 1059–1065. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (hermetia illucens) prepupae reared on different organic waste substrates: nutritional composition of black soldier fly. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Jucker, C.; Erba, D.; Leonardi, M.G.; Lupi, D.; Savoldelli, S. Assessment of vegetable and fruit substrates as potential rearing media for Hermetia Illucens (Diptera: Stratiomyidae) larvae. Environmental Entomology 2017, 46, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Salomone, R.; Saija, G.; Mondello, G.; Giannetto, A.; Fasulo, S.; Savastano, D. Environmental impact of food waste bioconversion by insects: Application of life cycle assessment to process Using Hermetia Illucens. Journal of Cleaner Production 2017, 140, 890–905. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia Illucens) Larvae: J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Kawasaki, K.; Hashimoto, Y.; Hori, A.; Kawasaki, T.; Hirayasu, H.; Iwase, S.; Hashizume, A.; Ido, A.; Miura, C.; Miura, T.; et al. Evaluation of black soldier fly (Hermetia illucens) larvae and pre-pupae raised on household organic waste, as potential ingredients for poultry feed. Animals 2019, 9, 98. [Google Scholar] [CrossRef]
- Bava, L.; Jucker, C.; Gislon, G.; Lupi, D.; Savoldelli, S.; Zucali, M.; Colombini, S. Rearing of Hermetia illucens on different organic by-products: influence on growth, waste reduction, and environmental impact. Animals 2019, 9, 289. [Google Scholar] [CrossRef]
- Scala, A.; Cammack, J.A.; Salvia, R.; Scieuzo, C.; Franco, A.; Bufo, S.A.; Tomberlin, J.K.; Falabella, P. Rearing substrate impacts growth and macronutrient composition of Hermetia illucens (L.) (diptera: stratiomyidae) larvae produced at an industrial scale. Sci. Rep. 2020, 10, 19448. [Google Scholar] [CrossRef]
- Hadj Saadoun, J.; Montevecchi, G.; Zanasi, L.; Bortolini, S.; Macavei, L.I.; Masino, F.; Maistrello, L.; Antonelli, A. Lipid profile and growth of black soldier flies (Hermetia illucens, Stratiomyidae) reared on by-products from different food chains. J. Sci. Food Agric. 2020, 100, 3648–3657. [Google Scholar] [CrossRef]
- Van Huis, A.; Oonincx, D.G.A.B.; Rojo, S.; Tomberlin, J.K. Insects as feed: house fly or black soldier fly? Journal of Insects as Food and Feed 2020, 6, 221–229. [Google Scholar] [CrossRef]
- Gasco, L.; Biancarosa, I.; Liland, N.S. From Waste to Feed: A Review of recent knowledge on insects as producers of protein and fat for animal feeds. Current Opinion in Green and Sustainable Chemistry 2020, 23, 67–79. [Google Scholar] [CrossRef]
- Barbi, S.; Macavei, L.I.; Fuso, A.; Luparelli, A.V.; Caligiani, A.; Ferrari, A.M.; Maistrello, L.; Montorsi, M. Valorization of seasonal agri-food leftovers through insects. Science of The Total Environment 2020, 709, 136209. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Nozzi, V.; Truzzi, C.; Giorgini, E.; Cardinaletti, G.; Freddi, L.; Ratti, S.; Girolametti, F.; Osimani, A.; et al. Physiological responses of Siberian sturgeon (Acipenser baerii) juveniles fed on full-fat insect-based diet in an aquaponic system. Sci. Rep. 2021, 11, 1057. [Google Scholar] [CrossRef]
- Poveda, J. Insect frass in the development of sustainable agriculture. A review. Agron. Sustain. Dev. 2021, 41, 5. [Google Scholar] [CrossRef]
- Müller, A.; Wolf, D.; Gutzeit, H.O. The black soldier fly, Hermetia illucens—a promising source for sustainable production of proteins, lipids and bioactive substances. Zeitschrift für Naturforschung C 2017, 72, 351–363. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia illucens and poultry by-product meals as alternatives to plant protein sources in gilthead seabream (Sparus aurata) diet: a multidisciplinary study on fish gut status. Animals 2021, 11, 677. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Animal Feed Science and Technology 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; Van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed – A review. Journal of Insects as Food and Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Vargas, A.; Randazzo, B.; Riolo, P.; Truzzi, C.; Gioacchini, G.; Giorgini, E.; Loreto, N.; Ruschioni, S.; Zarantoniello, M.; Antonucci, M.; et al. Rearing zebrafish on black soldier fly (Hermetia illucens): biometric, histological, spectroscopic, biochemical, and molecular implications. Zebrafish 2018, 15, 404–419. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Bruni, L.; Randazzo, B.; Vargas, A.; Gioacchini, G.; Truzzi, C.; Annibaldi, A.; Riolo, P.; Parisi, G.; Cardinaletti, G.; et al. Partial dietary inclusion of hermetia illucens (black soldier fly) full-fat prepupae in zebrafish feed: biometric, histological, biochemical, and molecular implications. Zebrafish 2018, 15, 519–532. [Google Scholar] [CrossRef]
- Cardinaletti, G.; Randazzo, B.; Messina, M.; Zarantoniello, M.; Giorgini, E.; Zimbelli, A.; Bruni, L.; Parisi, G.; Olivotto, I.; Tulli, F. Effects of graded dietary inclusion level of full-fat Hermetia illucens prepupae meal in practical diets for rainbow trout (Oncorhynchus mykiss). Animals 2019, 9, 251. [Google Scholar] [CrossRef]
- Giannetto, A.; Oliva, S.; Riolo, K.; Savastano, D.; Parrino, V.; Cappello, T.; Maisano, M.; Fasulo, S.; Mauceri, A. Waste valorization via Hermetia illucens to produce protein-rich biomass for feed: insight into the critical nutrient taurine. Animals 2020, 10, 1710. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Cranfill, K.; McGuire, M.A.; Mosley, E.E.; Tomberlin, J.K.; Newton, L.; Sealey, W.; Sheppard, C.; Irving, S. Fish offal recycling by the black soldier fly produces a foodstuff high in omega-3 fatty acids. J. World Aquaculture Soc. 2007, 38, 309–313. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.-J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, e0183188. [Google Scholar] [CrossRef]
- Pinotti, L.; Giromini, C.; Ottoboni, M.; Tretola, M.; Marchis, D. Review: Insects and former foodstuffs for upgrading food waste biomasses/streams to feed ingredients for farm animals. Animals 2019, 13, 1365–1375. [Google Scholar] [CrossRef]
- Hoc, B.; Genva, M.; Fauconnier, M.-L.; Lognay, G.; Francis, F.; Caparros Megido, R. About lipid metabolism in Hermetia illucens (L. 1758): on the origin of fatty acids in prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Zimbelli, A.; Randazzo, B.; Compagni, M.D.; Truzzi, C.; Antonucci, M.; Riolo, P.; Loreto, N.; Osimani, A.; Milanović, V.; et al. Black soldier fly (Hermetia Illucens) reared on roasted coffee by-product and Schizochytrium sp. as a sustainable terrestrial ingredient for aquafeeds production. Aquaculture 2020, 518, 734659. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Influence of resources on Hermetia illucens (Diptera: Stratiomyidae) larval development. jnl. med. entom. 2013, 50, 898–906. [Google Scholar] [CrossRef]
- Cammack, J.; Tomberlin, J. The impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.) (diptera: stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef]
- Gligorescu, A.; Toft, S.; Hauggaard-Nielsen, H.; Axelsen, J.A.; Nielsen, S.A. Development, metabolism and nutrient composition of black soldier fly larvae (Hermetia illucens; Diptera: Stratiomyidae) in relation to temperature and diet. Journal of Insects as Food and Feed 2018, 4, 123–133. [Google Scholar] [CrossRef]
- Smetana, S.; Palanisamy, M.; Mathys, A.; Heinz, V. Sustainability of insect use for feed and food: life cycle assessment perspective. Journal of Cleaner Production 2016, 137, 741–751. [Google Scholar] [CrossRef]
- Smetana, S.; Schmitt, E.; Mathys, A. Sustainable use of Hermetia illucens insect biomass for feed and food: attributional and consequential life cycle assessment. Resources, Conservation and Recycling 2019, 144, 285–296. [Google Scholar] [CrossRef]
- Maiolo, S.; Parisi, G.; Biondi, N.; Lunelli, F.; Tibaldi, E.; Pastres, R. Fishmeal partial substitution within aquafeed formulations: life cycle assessment of four alternative protein sources. Int. J. Life Cycle Assess. 2020, 25, 1455–1471. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Secci, G.; Notarstefano, V.; Giorgini, E.; Lock, E.J.; Parisi, G.; Olivotto, I. Application of laboratory methods for understanding fish responses to black soldier fly (Hermetia illucens) based diets. Journal of Insects as Food and Feed 2022, 8, 1173–1195. [Google Scholar] [CrossRef]
- Gambelli, D.; Naspetti, S.; Zander, K.; Zanoli, R. Organic aquaculture: economic, market and consumer aspects. In Organic Aquaculture; Lembo, G., Mente, E., Eds.; Springer International Publishing: Cham, 2019; ISBN 978-3-030-05602-5. [Google Scholar]
- Brown, M.R.; Farmer, C.L. Riboflavin content of six species of microalgae used in mariculture. J. Appl. Phycol. 1994, 6, 61–65. [Google Scholar] [CrossRef]
- Nakahara, T.; Yokochi, T.; Higashihara, T.; Tanaka, S.; Yaguchi, T.; Honda, D. Production of docosahexaenoic and docosapentaenoic acids by Schizochytrium sp. isolated from yap islands. J. Am. Oil. Chem. Soc. 1996, 73, 1421–1426. [Google Scholar] [CrossRef]
- Ren, L.-J.; Ji, X.-J.; Huang, H.; Qu, L.; Feng, Y.; Tong, Q.-Q.; Ouyang, P.-K. Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Fitzer, S.C.; Plancq, J.; Floyd, C.J.; Kemp, F.M.; Toney, J.L. Increased pCO2 changes the lipid production in important aquacultural feedstock algae Isochrysis galbana, but not in Tetraselmis suecica. Aquaculture and Fisheries 2019, 4, 142–148. [Google Scholar] [CrossRef]
- Bruni, L.; Randazzo, B.; Cardinaletti, G.; Zarantoniello, M.; Mina, F.; Secci, G.; Tulli, F.; Olivotto, I.; Parisi, G. Dietary inclusion of full-fat Hermetia illucens prepupae meal in practical diets for rainbow trout (Oncorhynchus mykiss): lipid metabolism and fillet quality investigations. Aquaculture 2020, 529, 735678. [Google Scholar] [CrossRef]
- Truzzi, C.; Giorgini, E.; Annibaldi, A.; Antonucci, M.; Illuminati, S.; Scarponi, G.; Riolo, P.; Isidoro, N.; Conti, C.; Zarantoniello, M.; et al. Fatty Acids Profile of Black Soldier Fly (Hermetia Illucens): Influence of feeding substrate based on coffee-waste silverskin enriched with microalgae. Animal Feed Science and Technology 2020, 259, 114309. [Google Scholar] [CrossRef]
- May, B. M. The occurrence in New Zealand and the life-history of the soldier fly Hermetia illucens (L.)(Diptera: Stratiomyidae). Nzj. Sci. 1961, 55-65, 55–65. [Google Scholar]
- Waldbauer, G.P. The consumption and utilization of food by insects. In Advances in Insect Physiology; Elsevier, 1968; Vol. 5, pp. 229–288 ISBN 978-0-12-024205-4.
- Le Féon, S.; Thévenot, A.; Maillard, F.; Macombe, C.; Forteau, L.; Aubin, J. Life cycle assessment of fish fed with insect meal: case study of mealworm inclusion in trout feed, in France. Aquaculture 2019, 500, 82–91. [Google Scholar] [CrossRef]
- Ermolaev, E.; Lalander, C.; Vinnerås, B. Greenhouse gas emissions from small-scale fly larvae composting with Hermetia illucens. Waste Management 2019, 96, 65–74. [Google Scholar] [CrossRef]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black soldier fly biowaste treatment—assessment of global warming potential. Waste Management 2019, 84, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Hanserud, O.S.; Cherubini, F.; Øgaard, A.F.; Müller, D.B.; Brattebø, H. Choice of mineral fertilizer substitution principle strongly influences LCA environmental benefits of nutrient cycling in the agri-food system. Science of The Total Environment 2018, 615, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Tschirner, M.; Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. Journal of Insects as Food and Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- Liu, Z.; Minor, M.; Morel, P.C.H.; Najar-Rodriguez, A.J. Bioconversion of three organic wastes by black soldier fly (Diptera: Stratiomyidae) larvae. Environmental Entomology 2018, 47, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D. C.; Newton, G. L.; Thompson, S. A.; Savage, S. A value added manure management system using the black soldier fly. Bioresource technology.
- Craig Sheppard, D.; Larry Newton, G.; Thompson, S.A.; Savage, S. A value added manure management system using the black soldier fly. Bioresource Technology 1994, 50, 275–279. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected life-history traits of black soldier flies (Diptera: Stratiomyidae) reared on three artificial diets. Ann. Entomol. Soc. Am. 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Moreau, J.; Benrey, B.; Thiery, D. Assessing larval food quality for phytophagous insects: are the facts as simple as they appear? Funct. Ecology. 2006, 20, 592–600. [Google Scholar] [CrossRef]
- Newton, G.L.; Sheppard, D.C.; Watson, D.W.; Burtle, G.J.; Dove, C.R.; Tomberlin, J.K.; Thelen, E.E. The black soldier fly, Hermetia illucens, as a manure management/ resource recovery tool. Symposium on the State of the Science of Animal Manure and Waste Management. , San Antonio, TX. (http://www.cals.ncsu.edu/waste_mgt/natlcenter/sanantonio/proceedings. 5–7 January.
- Gobbi, P.; Martínez-Sánchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461–468. [Google Scholar] [CrossRef]
- Myers, H.M.; Tomberlin, J.K.; Lambert, B.D.; Kattes, D. Development of black soldier fly (diptera: stratiomyidae) larvae fed dairy manure. Environ. Entomol. 2008, 37(1), 11–5. [Google Scholar] [CrossRef] [PubMed]
- Barragan-Fonseca, K.B.; Dicke, M.; Van Loon, J.J.A. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia Illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of black soldier fly (Diptera: Stratiomyidae) larvae to recycle food waste. Environmental Entomology 2015, 44, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Tomberlin, J.K.; Adler, P.H.; Myers, H.M. Development of the Black Soldier Fly (Diptera: Stratiomyidae) in Relation to Temperature. Environ. Entomol. 2009, 38, 930–934. [Google Scholar] [CrossRef] [PubMed]
- Chia, S.Y.; Tanga, C.M.; Osuga, I.M.; Mohamed, S.A.; Khamis, F.M.; Salifu, D.; Sevgan, S.; Fiaboe, K.K.M.; Niassy, S.; Van Loon, J.J.A.; et al. Effects of waste stream combinations from brewing industry on performance of black soldier fly, Hermetia Illucens (Diptera: Stratiomyidae). PeerJ 2018, 6, e5885. [Google Scholar] [CrossRef] [PubMed]
- Permana, A.D.; Ramadhani Eka Putra, J.E.N. Growth of black soldier fly (Hermetia Illucens) larvae fed on spent coffee ground. IOP Conf. Ser.: Earth Environ. Sci. 2018, 187, 012070. [Google Scholar] [CrossRef]
- Ujvari, B.; Wallman, J.F.; Madsen, T.; Whelan, M.; Hulbert, A.J. Experimental studies of blowfly (Calliphora stygia) longevity: a little dietary fat is beneficial but too much is detrimental. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 2009, 154, 383–388. [Google Scholar] [CrossRef]
- Wilkinson, J.M. Re-defining Efficiency of feed use by livestock. Animals 2011, 5, 1014–1022. [Google Scholar] [CrossRef]
- de Vries, M.; De Boer, I.J.M. Comparing environmental impacts for livestock products: A review of life cycle assessments. Livestock Science 2010, 128, 1–11. [Google Scholar] [CrossRef]
- Scriber, J.M.; Slansky, F. The nutritional ecology of immature insects. Annu. Rev. Entomol. 1981, 26, 183–211. [Google Scholar] [CrossRef]
- Rehman, K.U.; Rehman, A.; Cai, M.; Zheng, L.; Xiao, X.; Somroo, A.A.; Wang, H.; Li, W.; Yu, Z.; Zhang, J. Conversion of mixtures of dairy manure and soybean curd residue by black soldier fly larvae (Hermetia Illucens L.). Journal of Cleaner Production 2017, 154, 366–373. [Google Scholar] [CrossRef]
- Friend, W.G. Nutritional requirements of phytophagous insects. Annu. Rev. Entomol. 1958, 3, 57–74. [Google Scholar] [CrossRef]
- Lundy, M.E.; Parrella, M.P. Crickets are not a free lunch: protein capture from scalable organic side-streams via high-density populations of Acheta domesticus. PLoS ONE 2015, 10, e0118785. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.Y.; Kutty, S.R.M.; Malakahmad, A.; Tan, C.K. Feasibility study of biodiesel production using lipids of Hermetia illucens larva fed with organic waste. Waste Management 2016, 47, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Muros, M.-J.; Barroso, F.G.; Manzano-Agugliaro, F. Insect meal as renewable source of food for animal feeding: A review. Journal of Cleaner Production 2014, 65, 16–27. [Google Scholar] [CrossRef]
- Pérez-López, P.; De Vree, J.H.; Feijoo, G.; Bosma, R.; Barbosa, M.J.; Moreira, M.T.; Wijffels, R.H.; Van Boxtel, A.J.B.; Kleinegris, D.M.M. Comparative life cycle assessment of real pilot reactors for microalgae cultivation in different seasons. Applied Energy 2017, 205, 1151–1164. [Google Scholar] [CrossRef]
- Ye, J.; Zhou, Q.; Zhang, X.; Hu, Q. Microalgal dewatering using a polyamide thin film composite forward osmosis membrane and fouling mitigation. Algal Research 2018, 31, 421–429. [Google Scholar] [CrossRef]
- Henriksson, P.J.G.; Dickson, M.; Allah, A.N.; Al-Kenawy, D.; Phillips, M. Benchmarking the environmental performance of best management practice and genetic improvements in egyptian aquaculture using life cycle assessment. Aquaculture 2017, 468, 53–59. [Google Scholar] [CrossRef]
Figure 1.
System boundaries considered in the study for Life Cycle Assessment (LCA).
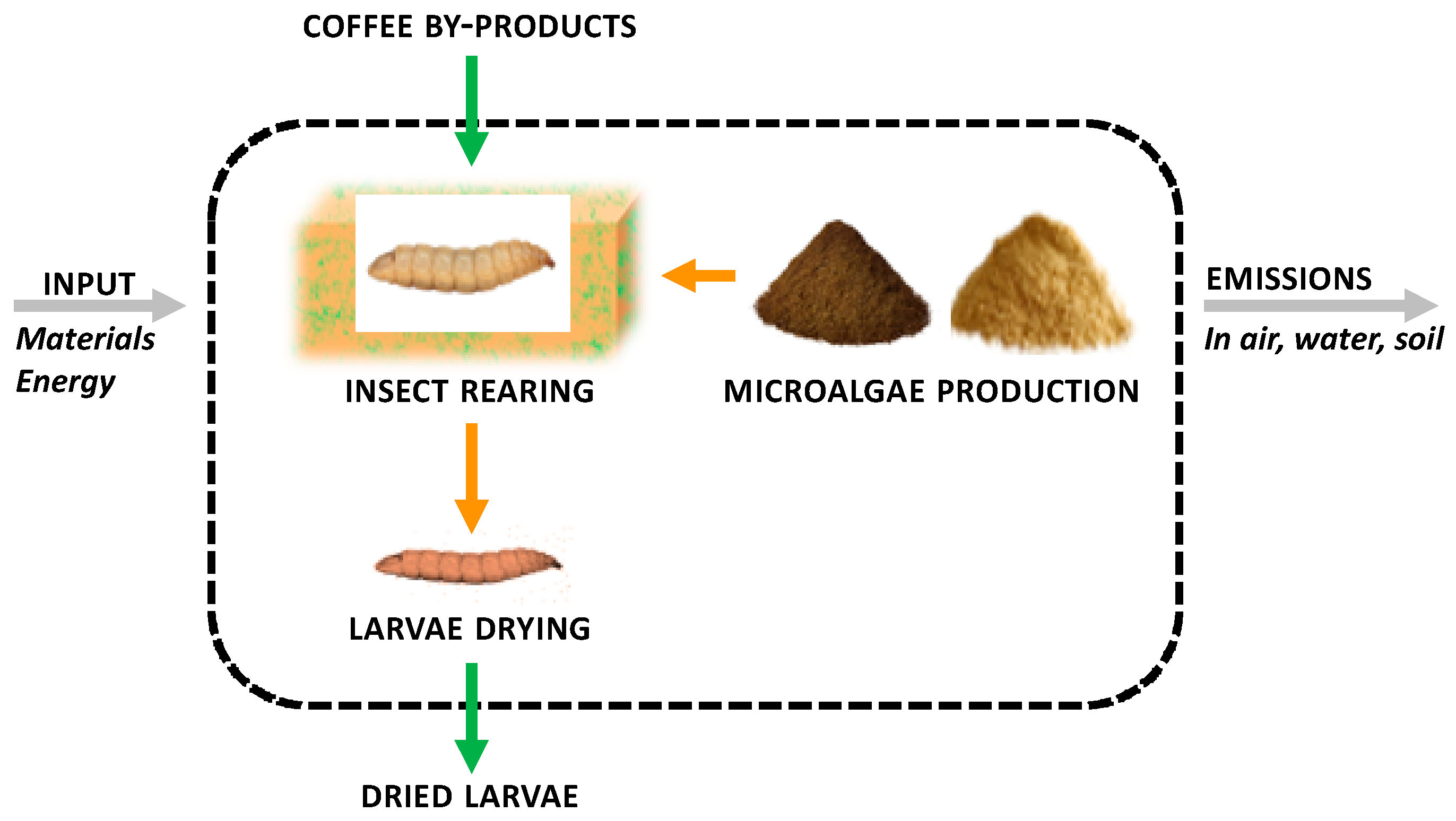

Table 1.
Experimental diet compositions based on coffee by-product (CB) and different level of Schizochytrium limacinum (S) or Isochrysis galbana (I).
Table 1.
Experimental diet compositions based on coffee by-product (CB) and different level of Schizochytrium limacinum (S) or Isochrysis galbana (I).
| Diets | CB (%) | Schizochytrium limacinum (%) | Isochrysis galbana (%) |
|---|---|---|---|
| 5S | 95 | 5 | - |
| 10S | 90 | 10 | - |
| 20S | 80 | 20 | - |
| 25S | 75 | 25 | - |
| 5I | 95 | - | 5 |
| 10I | 90 | - | 10 |
| 20I | 80 | - | 20 |
| 25I | 75 | - | 25 |
| C | 100 | - | - |
Experimental diet abbreviations: 5S = 95% CB + 5% S; 10S = 90% CB + 10% S; 20S = 80% CB + 20% S; 25S = 75% CB + 25% S; 5I = 95% CB + 5% I; 10I = 90% CB + 10% I; 20I = 80% CB + 20% I; 25I = 75% CB + 25% I; C = 100% CB.
Table 2.
Growth performance of BSF larvae reared on the different experimental diets composed of coffee by-products (CB) enriched with Schizochytrium limacinum (S) and Isochrysis galbana (I) dry biomass and control (C) (mean ± SD; n = 6).
Table 2.
Growth performance of BSF larvae reared on the different experimental diets composed of coffee by-products (CB) enriched with Schizochytrium limacinum (S) and Isochrysis galbana (I) dry biomass and control (C) (mean ± SD; n = 6).
| Diets | Prepupal weight (mg) | Survival rate (%) | Development time (days) | Growth Rate (mg/day) |
|---|---|---|---|---|
| 5S | 143.0 ± 12.4cB | 88.2 ± 6.1aA | 20 ± 0cC | 7.0 ± 0.6cC |
| 10S | 161.3 ± 10.7bcAB | 91.0 ± 5.4aA | 19 ± 0bB | 8.4 ± 0.6bB |
| 20S | 173.9 ± 9.6abA | 87.8 ± 3.1aA | 19 ± 0bB | 9.0 ± 0.5bB |
| 25S | 180.2 ± 11.5aA | 84.5 ± 12.6aA | 18 ± 0aA | 9.9 ± 0.7aA |
| 5I | 107.6 ± 5.1dB | 84.3 ± 3.1aA | 20 ± 0cC | 5.3 ± 0.3eC |
| 10I | 143.1 ± 5.2cA | 82.0 ± 7.2aA | 20 ± 0cC | 7.0 ± 0.3cA |
| 20I | 113.1 ± 11.8dB | 85.2 ± 5.6aA | 19 ± 0bB | 5.8 ± 0.6dB |
| 25I | 132.3 ± 11.2cA | 84.3 ± 6.5aA | 18 ± 0aA | 7.2 ± 0.6cA |
| C | 72.4 ± 7.7eCC | 81.0 ± 8.8aAA | 39 ± 0dDD | 1.8 ± 0.2fDD |
Different superscripts in a column denote significant differences (P < 0.05): a,b,c,d,e small letters denote a significant difference among all diets; A,B,C,D,E capital letters denote a significant difference among diets composed of coffee by-products (CB) enriched with different percentage of S. limacinum (S) and control diet (C); A,B,C,D,E capital and bolt letters denote a significant difference among diets composed of coffee by-products (CB) enriched with different percentage of I. galbana (I) and control diet (C). Experimental diet abbreviations: 5S = 95% CB + 5% S; 10S = 90% CB + 10% S; 20S = 80% CB + 20% S; 25S = 75% CB + 25% S; 5I = 95% CB + 5% I; 10I = 90% CB + 10% I; 20I = 80% CB + 20% I; 25I = 75% CB + 25% I; C = 100% CB.
Table 3.
Waste Reduction Index (WRI) and Feed Conversion Ratio (FCR) of BSF larvae reared on the different experimental diets composed of coffee by-products (CB) enriched with different percentage of Schizochytrium limacinum (S) and Isochrysis galbana (I) microalgae and control diet (C) (mean ± SD; n = 6).
Table 3.
Waste Reduction Index (WRI) and Feed Conversion Ratio (FCR) of BSF larvae reared on the different experimental diets composed of coffee by-products (CB) enriched with different percentage of Schizochytrium limacinum (S) and Isochrysis galbana (I) microalgae and control diet (C) (mean ± SD; n = 6).
| Diets | WRI (%/day) | FCR |
|---|---|---|
| 5S | 1.5 ± 0.1dC | 5.2 ± 0.3aA |
| 10S | 2.6 ± 0.2aA | 7.21 ± 0.7cB |
| 20S | 2.1 ± 0.3bcB | 5.6 ± 0.8abA |
| 25S | 2.0 ± 0.2cB | 5.0 ± 0.3aA |
| 5I | 1.2 ± 0.1fCD | 5.8 ± 0.3bB |
| 10I | 1.4 ± 0.1eB | 5.1 ± 0.5aA |
| 20I | 2.2 ± 0.1bA | 9.5 ± 0.9dC |
| 25I | 1.3 ± 0.1efBC | 4.5 ± 0.5aA |
| C | 1.2 ± 0.1gDD | 34.2 ± 6.4eCD |
Different superscripts in a column denote significant differences (P < 0.05): a,b,c,d,e small letters denote a significant difference among all diets; A,B,C,D,E capital letters denote a significant difference among diets composed of coffee by-products (CB) enriched with different percentage of S. limacinum (S) and control diet (C); A,B,C,D,E capital and bolt letters denote a significant difference among diets composed of coffee by-products (CB) enriched with different percentage of I. galbana (I) and control diet (C). Experimental diet abbreviations: 5S = 95% CB + 5% S; 10S = 90% CB + 10% S; 20S = 80% CB + 20% S; 25S = 75% CB + 25% S; 5I = 95% CB + 5% I; 10I = 90% CB + 10% I; 20I = 80% CB + 20% I; 25I = 75% CB + 25% I; C = 100% CB.
Table 4.
Water (g/100g), protein, lipid and ash (g/100 g D.M.) content of the different experimental diets composed of coffee by-products (CB) enriched with Schizochytrium limacinum (S) and Isochrysis galbana (I) dry biomass and control (C) (mean ± SD; n = 6).
Table 4.
Water (g/100g), protein, lipid and ash (g/100 g D.M.) content of the different experimental diets composed of coffee by-products (CB) enriched with Schizochytrium limacinum (S) and Isochrysis galbana (I) dry biomass and control (C) (mean ± SD; n = 6).
| Diet | Crude protein | Lipid | Ash |
|---|---|---|---|
| 5S | 21.42 ± 1.10 | 0.75 ± 0.06 | 8.62 ± 0.13 |
| 10S | 22.42 ± 3.34 | 0.81 ± 0.04 | 8.71 ± 0.05 |
| 20S | 28.06 ± 1.15 | 0.82 ± 0.05 | 8.84 ± 0.01 |
| 25S | 31.07 ± 2.19 | 0.93 ± 0.03 | 8.91 ± 0.75 |
| 5I | 23.52 ± 2.26 | 1.66 ± 0.10 | 8.80 ± 0.25 |
| 10I | 24.79 ± 3.18 | 2.11 ± 0.06 | 8.65 ± 0.16 |
| 20I | 25.63 ± 2.41 | 2.67 ± 0.10 | 9.22 ± 0.10 |
| 25I | 27.00 ± 3.22 | 3.56 ± 0.05 | 9.79 ± 0.48 |
| C | 20.96 ± 1.32 | 0.71 ± 0.07 | 8.71 ± 0.22 |
Experimental diet abbreviations: 5S = 95% CB + 5% S; 10S = 90% CB + 10% S; 20S = 80% CB + 20% S; 25S = 75% CB + 25% S; 5I = 95% CB + 5% I; 10I = 90% CB + 10% I; 20I = 80% CB + 20% I; 25I = 75% CB + 25% I; C = 100% CB.
Table 5.
Chemical composition (g/100g) of the BSF larvae reared on the different experimental diets composed of coffee by-products (CB) enriched with Schizochytrium limacinum (S) and Isochrysis galbana (I) dry biomass and control (C) (mean ± SD; n = 6).
Table 5.
Chemical composition (g/100g) of the BSF larvae reared on the different experimental diets composed of coffee by-products (CB) enriched with Schizochytrium limacinum (S) and Isochrysis galbana (I) dry biomass and control (C) (mean ± SD; n = 6).
| Diet | Crude protein | Lipid | Ash |
|---|---|---|---|
| 5S | 14.86±0.42cDC | 4.05±0.14cC | 7.13±0.13abB |
| 10S | 15.59±0.36bCB | 4.88±0.11bB | 6.34±0.16cbC |
| 20S | 16.85±0.21bBA | 7.74±0.12aA | 4.70±0.26deD |
| 25S | 18.29±0.23aA | 7.93±0.35aA | 4.39±0.13eD |
| 5I | 14.86±0.28cBC | 4.28±0.26cbB | 7.68±0.01abA |
| 10I | 15.59±0.30bAB | 4.00±0.06cB | 6.99±0.77abcAB |
| 20I | 16.85±0.39bA | 3.81±0.03cdB | 5.81±0.29cdCD |
| 25I | 18.29±0.22aA | 4.95±0.07bA | 4.35±0.44eD |
| C | 13.47±0.53dD | 3.18±0.08dD | 8.12±0.20aA |
Different superscripts in a column denote significant differences (P ⩽ 0.05): a,b,c,d,e small letters denote a significant difference among all diets; A,B,C,D,E capital letters denote a significant difference among diets composed of coffee by-products (CB) enriched with different percentage of S. limacinum (S) and control diet (C); A,B,C,D,E capital and bolt letters denote a significant difference among diets composed of coffee by-products (CB) enriched with different percentage of I. galbana (I) and control diet (C). Experimental diet abbreviations: 5S = 95% CB + 5% S; 10S = 90% CB + 10% S; 20S = 80% CB + 20% S; 25S = 75% CB + 25% S; 5I = 95% CB + 5% I; 10I = 90% CB + 10% I; 20I = 80% CB + 20% I; 25I = 75% CB + 25% I; C = 100% CB.
Table 6.
Environmental impact of the production of 1 kg of BSF dried larvae fed on the different experimental diets composed of coffee by-products enriched with different percentage (5, 10, 20 and 25) of Schizochytrium limacinum.
Table 6.
Environmental impact of the production of 1 kg of BSF dried larvae fed on the different experimental diets composed of coffee by-products enriched with different percentage (5, 10, 20 and 25) of Schizochytrium limacinum.
| Impact category | Unit | 5S | 10S | 20S | 25S |
|---|---|---|---|---|---|
| Global warming | kg CO2 eq | 8,03E+00 | 1,85E+01 | 2,73E+01 | 3,11E+01 |
| Freshwater eutrophication | kg P eq | 3,74E-03 | 7,96E-03 | 1,15E-02 | 1,30E-02 |
| Land use | m2a crop eq | 4,09E-01 | 8,89E-01 | 1,29E+00 | 1,46E+00 |
| Mineral resource scarcity | kg Cu eq | 2,72E-02 | 6,58E-02 | 9,83E-02 | 1,12E-01 |
| Fossil resource scarcity | kg oil eq | 1,97E+00 | 4,63E+00 | 6,87E+00 | 7,83E+00 |
| Water consumption | m3 | 1,75E-01 | 4,05E-01 | 5,98E-01 | 6,80E-01 |
Table 7.
Environmental impact of the production of 1 kg dried larvae fed on the different experimental diets composed of coffee by-products enriched with different percentage (5, 10, 20 and 25) of Isochrysis galbana.
Table 7.
Environmental impact of the production of 1 kg dried larvae fed on the different experimental diets composed of coffee by-products enriched with different percentage (5, 10, 20 and 25) of Isochrysis galbana.
| Impact category | Unit | 5I | 10I | 20I | 25I |
|---|---|---|---|---|---|
| Global warming | kg CO2 eq | 8,78E+00 | 1,37E+01 | 4,48E+01 | 2,77E+01 |
| Freshwater eutrophication | kg P eq | 4,04E-03 | 6,00E-03 | 1,85E-02 | 1,16E-02 |
| Land use | m2a crop eq | 4,43E-01 | 6,66E-01 | 2,09E+00 | 1,31E+00 |
| Mineral resource scarcity | kg Cu eq | 2,99E-02 | 4,79E-02 | 1,63E-01 | 9,97E-02 |
| Fossil resource scarcity | kg oil eq | 2,16E+00 | 3,39E+00 | 1,13E+01 | 6,97E+00 |
| Water consumption | m3 | 1,92E-01 | 2,99E-01 | 9,80E-01 | 6,06E-01 |
Table 8.
Environmental impact of the production of 1 kg protein of BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Schizochytrium limacinum.
Table 8.
Environmental impact of the production of 1 kg protein of BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Schizochytrium limacinum.
| Impact category | Unit | 5S | 10S | 20S | 25S |
|---|---|---|---|---|---|
| Global warming | kg CO2 eq | 1,65E+01 | 3,63E+01 | 5,45E+01 | 6,10E+01 |
| Freshwater eutrophication | kg P eq | 7,69E-03 | 1,56E-02 | 2,30E-02 | 2,55E-02 |
| Land use | m2a crop eq | 8,41E-01 | 1,75E+00 | 2,57E+00 | 2,86E+00 |
| Mineral resource scarcity | kg Cu eq | 5,59E-02 | 1,29E-01 | 1,96E-01 | 2,20E-01 |
| Fossil resource scarcity | kg oil eq | 4,05E+00 | 9,09E+00 | 1,37E+01 | 1,54E+01 |
| Water consumption | m3 | 3,60E-01 | 7,95E-01 | 1,19E+00 | 1,33E+00 |
Table 9.
Environmental impact of the production of 1 kg protein of BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Isochrysis galbana.
Table 9.
Environmental impact of the production of 1 kg protein of BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Isochrysis galbana.
| Impact category | Unit | 5I | 10I | 20I | 25I |
|---|---|---|---|---|---|
| Global warming | kg CO2 eq | 1,84E+01 | 2,76E+01 | 8,20E+01 | 4,56E+01 |
| Freshwater eutrophication | kg P eq | 8,46E-03 | 1,21E-02 | 3,39E-02 | 1,91E-02 |
| Land use | m2a crop eq | 9,28E-01 | 1,34E+00 | 3,83E+00 | 2,15E+00 |
| Mineral resource scarcity | kg Cu eq | 6,26E-02 | 9,65E-02 | 2,98E-01 | 1,64E-01 |
| Fossil resource scarcity | kg oil eq | 4,52E+00 | 6,83E+00 | 2,07E+01 | 1,15E+01 |
| Water consumption | m3 | 4,02E-01 | 6,02E-01 | 1,79E+00 | 9,97E-01 |
Table 10.
Environmental impact of the production of 1 kg lipid contained in BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Schizochytrium limacinum.
Table 10.
Environmental impact of the production of 1 kg lipid contained in BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Schizochytrium limacinum.
| Impact category | Unit | 5S | 10S | 20S | 25S |
|---|---|---|---|---|---|
| Global warming | kg CO2 eq | 6,06E+01 | 1,16E+02 | 1,19E+02 | 1,41E+02 |
| Freshwater eutrophication | kg P eq | 2,82E-02 | 4,99E-02 | 5,00E-02 | 5,88E-02 |
| Land use | m2a crop eq | 3,09E+00 | 5,58E+00 | 5,61E+00 | 6,61E+00 |
| Mineral resource scarcity | kg Cu eq | 2,05E-01 | 4,13E-01 | 4,27E-01 | 5,07E-01 |
| Fossil resource scarcity | kg oil eq | 1,49E+01 | 2,91E+01 | 2,98E+01 | 3,54E+01 |
| Water consumption | m3 | 1,32E+00 | 2,54E+00 | 2,60E+00 | 3,08E+00 |
Table 11.
Environmental impact of the production of 1 kg lipid contained BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Isochrysis galbana.
Table 11.
Environmental impact of the production of 1 kg lipid contained BSF dried larvae reared on the experimental diets containing different percentages (5, 10, 20 and 25) of Isochrysis galbana.
| Impact category | Unit | 5I | 10I | 20I | 25I |
|---|---|---|---|---|---|
| Global warming | kg CO2 eq | 6,38E+01 | 1,08E+02 | 3,63E+02 | 1,68E+02 |
| Freshwater eutrophication | kg P eq | 2,94E-02 | 4,71E-02 | 1,50E-01 | 7,05E-02 |
| Land use | m2a crop eq | 3,22E+00 | 5,23E+00 | 1,69E+01 | 7,96E+00 |
| Mineral resource scarcity | kg Cu eq | 2,17E-01 | 3,76E-01 | 1,32E+00 | 6,06E-01 |
| Fossil resource scarcity | kg oil eq | 1,57E+01 | 2,66E+01 | 9,15E+01 | 4,24E+01 |
| Water consumption | m3 | 1,40E+00 | 2,35E+00 | 7,94E+00 | 3,68E+00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



