
Submitted:
16 October 2023
Posted:
16 October 2023
You are already at the latest version
Abstract
Alheira is a naturally fermented meat sausage traditionally made in the Portuguese region of Trás-os-Montes. Lactic Acid Bacteria (LAB) have been found to be the dominant microorganisms in alheira. This study aimed: 1) to characterize technological features and in vitro antimicrobial activity of LAB isolates; and 2) to reveal associations between such phenotypic characteristics and the isolates genotypically identified to species level through amplification and sequencing of the 16S ribosomal gene. Sixty-two LAB isolates were identified and Enterococcus faecium was the most prevalent (32.3%), followed by Leuconostoc mesenteroides (19.4%) and Latilactobacillus sakei (17.7%), aligning with previous research on traditional Portuguese fermented meat sausages. The phenotypic analysis of LAB isolates indicated diverse acidification capacities, proteolytic activities, and inhibitory effects against foodborne pathogens. Comprising the technological attributes, L. paracasei and L. mesenteroides had higher proteolytic activity (3.46 and 3.43 mm), whereas E. faecium and L. paracasei stood out for their acidifying capacity, with mean pH after 6 h of 5.78 and 6.03, respectively. Overall, lactobacilli presented higher inhibition diameter against the pathogens Staphylococcus aureus, Listeria monocytogenes and Salmonella Typhimurium. Although the mechanisms for inhibition of pathogen growth needs to be further elucidated, these findings enhance our understanding of LAB diversity and functionality in alheira sausages, contributing to product safety and quality assessment.
Keywords:
fermented sausage
; lactic acid bacteria
; food quality
; phylogenetic analysis
1. Introduction
Mediterranean artisanal foods play a key role on the development of rural regions, allowing and stimulating local commercialization, employment of the rural population, and preservation of local heritage. Alheira is a naturally fermented meat sausage traditionally made in the Portuguese region of Trás-os-Montes. The production of alheira uses various meats (most commonly pork and poultry meat) and the final product is a paste that is stuffed in pig intestinal – or cellulose based – casings [1]. In addition, the alheira sausage goes through an intermittent smoking process (for 2–7 days) that reduces water activity (aw) and pH – though this process does not ensure microbiological safety since some pathogenic organisms have the ability to develop under these conditions [1,2,3]. Alheira manufacturing process and final composition can vary considerably between regional producers and even within different batches of a factory. These differences may arise from variations in the initial formulation, fermentation's duration, or drying processing times. [1,2,3]. Lactic Acid Bacteria (LAB) have been found to be the dominant microorganisms in alheira [1,2,3]. LAB are similar in morphology, metabolism and are related phylogenetically, they can be described as Gram-positive, catalase-negative, facultative anaerobic, cocci, coccobacilli or rods; and ferment glucose to lactic acid – through either homofermentative or heterofermentative pathways [4,5,6]. The conventional method for identifying LAB species involves utilizing phenotypic attributes such as morphology, fermentation patterns, and lactic acid isomer profiles; or genetic methods, such as 16S rRNA sequencing – a gene which contains both well-conserved and less-conserved regions used to sort different genera and species; or species-specific genes to discriminate those that are closely related [4,5,6]. LAB associated with food are generally restricted to the genera Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Oenococcus, Pediococcus, Streptococcus, Tetragenococcus, Vagococcus, and Weissella [5]. Importantly, LAB strains utilized as starter cultures or those developing spontaneously during meat fermentation must display specific and discernible metabolic attributes. These encompass the capacity to generate acids and aromatic compounds; curb the propagation of harmful microorganisms; enzymatically degrade proteins; and lack the ability to express virulence factors and toxins. These attributes stand as imperative safeguards, ensuring the elevated quality and safety of the final product [5]. Thus, the objectives of this study were: 1) to conduct a phenotypic characterization of LAB isolates, including technological features such as proteolytic activity, production of L-lactic acid, acidifying potential and antimicrobial activity against Staphylococcus aureus, Salmonella Typhimurium and Listeria monocytogenes; and 2) to identify genotypically the LAB isolates to species level, and reveal any associations between desired phenotypic characteristics and LAB species.
2. Materials and Methods
2.1. Sampling
Fifty-nine alheira sausages were obtained from thirteen artisanal establishments located in the towns of Bragança (BST, TB, BF, QP), Mirandela (CM, FN), Mogadouro (BPM), Vimioso (BV), Vinhais (CSM, AG, AMO), and Valpaços (SM, MP), which belong to the Portuguese northeastern region of Trás-Os-Montes (Figure 1). The sausages were acquired unpacked at traditional markets and underwent physicochemical and microbiological analysis. It is worth mentioning that alheira sausages were purchased from artisanal producers that do not employ starter cultures in their elaboration.
2.2. Physicochemical and microbiological analysis of sausages
The casings enveloping the sausages were removed, and the contents were then partitioned and homogenized. Physicochemical analysis of the sausage included measurement of pH, with a probe (Hanna Instruments, HI5522, Woonsocket, RL, USA); and aw with an Aqualab meter (4TE Decagon, WA, USA). Both measurements were done in triplicate. Additional determinations of moisture, dry matter and ashes, were determined according to ISO 1442:2023 (direct drying method) [7], ISO 937:2023 [8] and ISO 936:1998 [9], respectively, and the results were expressed in dry matter basis (%). Finally, measurements of carbohydrates (CHO), fat and protein contents (%), were carried out in duplicate and expressed in dry matter basis (%) [2].
For the microbiological assays, the samples (10 g) were homogenized in buffered peptone water and ten-fold dilutions were prepared thereof. Total mesophiles were determined in Plate Count Agar at 35 °C for 48 h, S. aureus was counted on Baird-Parker Agar supplemented with Egg Yolk Tellurite incubated at 35 °C for 48 h, following ISO 6888-1:2021 [10]; and Clostridium perfringens was determined using Tryptose Sulphite Cycloserin Agar with egg yolk supplement and incubated at 35 °C for 24 h, in accordance with the compendium methods for microbiological evaluation of foods from the American Public Health Association (APHA) [11]. Additionally, Salmonella spp. search was performed using tetrathionate broth, Rappaport-Vassiliadis broth, Hektoen enteric agar, Bismute-Sulfite agar, and Xylose Lysine Desoxycholate agar, according to the Bacteriological Analytical Manual (BAM) for Salmonella detection in food matrices from Food and Drug Administration (FDA) [12], with an additional step of serological confirmation. Samples were assigned 1 if positive and 0 if negative, for the statistical analysis. Determinations were carried out in duplicate, and further details on the microbiological protocols can be found in [2,13].
2.3. LAB isolation
In order to isolate LAB from the alheira sausages, one-ml volumes from the ten-fold dilutions were incorporated in De Man, Rogosa and Sharpe (MRS) and M17 selective agars, and later overlayed with agar 1.2%, and incubated at 30 °C for 48 h [13]. Five typical colonies on MRS and M17 agar (each) were selected for purification and incubated at 30 °C for another 48 h in the respective media. Finally, isolates were confirmed by catalase (3% hydrogen peroxide) and Gram tests, as well as morphologic observation. Confirmed isolated were cryopreserved in 25% glycerol at -80 °C.
2.4. Phenotypic characterization
Antimicrobial activity of LAB isolates was tested against three foodborne pathogens: Staphylococcus aureus subsp. aureus ATCC 6538, Salmonella enterica subsp. enterica serovar Typhimurium ATCC 43971 and Listeria monocytogenes WDCM 00019 using the agar spot method. Each LAB strain reactivated in MRS or M17 broth after an overnight culture was spotted (3 or 5 µl) on solidified MRS or M17 agar plates, respectively. Then, the plates were covered with 10 mL of BHI broth with 0.75% (w/v) bacteriological agar seeded with 1 mL of each bacterial indicator strain (separately) at approximately 8 log CFU/mL – pathogenic strains were revived in 10 mL BHI broth for 16 h at 37 °C. The bacterial strains of L. monocytogenes went through an additional step of activation in 5 mL of BHI for 16 h at 37 °C, cultures were then successively inoculated until reaching the concentration mentioned above. After solidification, plates were incubated at temperatures of 37 °C for 16 h or 10 °C for 10 days. The inhibition diameter was measured (mm) in duplicate [13]. Sixty-two presumptive LAB with highest antimicrobial capacity at both temperatures were selected [13]. On this subset of strains, the additional phenotypic assays were carried out: proteolytic activity, acidifying capacity and L-lactic acid production.
For the determination of exocellular proteolytic activity, overnight cultures were spotted (3 µl) onto the surface of milk agar (composed of 10% (w/v) skim milk powder and 2.5% (w/v) agar) and incubated at 35 °C for 4 days. Proteolytic activity was measured as the diameter of the clear zones around each LAB colony.
To quantify the acidifying capacity, each isolate was reactivated separately in MRS or M17 broth overnight (30 °C, 24 h). Then, a loop of culture was placed in 10 mL of sterile reconstituted skim milk supplemented with yeast extract (0.3% (w/v)) and glucose (0.2% (w/v)) for two successive subcultures (30 °C for 24 h). Sterile reconstituted skim milk (100 mL) was then inoculated with 1 mL of the 24 h activated culture. For the acidification profiling, pH changes were determined using a pH meter (Hanna Instruments, model HI5522, USA) equipped with a HI1131 glass penetration probe during incubation at 30°C during 8 h (t = 0, 3, 6, 8 h), and after 24 h. For every strain, pH data was fitted to a decay curve to characterize acidification capacity [13]. The following descriptors were extracted from the fitted curves: ΔpH03: pH decrease between t = 0 h and t = 3 h; ΔpH06: pH decrease between t = 0 h and t = 6 h; ΔpH36: pH decrease between t = 3 h and t = 6 h; and pH6: pH at t = 6 h.
To quantify the L-lactic acid (g/L) produced by the LAB, isolates underwent a revival process in 10 mL of MRS or M17 broth, followed by an incubation at 37 °C for 24 hours. Subsequently, the inoculum was transferred to MRS or M17 agar plates, corresponding to the isolation media, with the aim of ensuring purity of colonies. These plates were then placed in an anaerobiosis jar and incubated at 30 °C for 48 hours. Two isolated colonies were carefully selected and combined with 5 mL of saline solution. The measurement of absorbance at 625 nm was done once turbidity reached an estimated 0.5 on the McFarland scale. Samples were adjusted to fall within absorbance values of 0.08-0.13. Following this, 100 µl of the adjusted samples were transferred to 4 mL of MRS broth and incubated at 30 °C for 4 hours. After the incubation period, 1 mL of the culture underwent centrifugation for 5 minutes at 13,000 rpm, and the resulting pellet was discarded. Subsequently, 10 µl of the supernatant was added to 500 µl of deionized water and vortexed. The concentration of L-lactic acid in g/L was determined using the Kit Nzytech L-lactic (NzyTech, Portugal), UV method, following the manufacturer's instructions.
2.5. Genotypic characterization
2.5.1. DNA extraction
Pure genomic DNA was obtained using the GF-1 Bacterial DNA Extraction Kit (Vivantis, Malaysia). LAB isolates were grown in MRS or M17 broth for 24 h at 37 °C, and 3 mL of bacterial culture was centrifuged at 6000 x g for 2 min to obtain a pellet. The pellet was resuspended in 80 µl of R1 Buffer and treated with 20 µl of lysozyme (50 mg/mL; Vivantis, Malaysia) for 30 min at 37 °C. The cells were centrifuged at 10000 x g for 3 min to form the pellet, which was resuspended in 180 µl of R2 Buffer and 3 µl of Proteinase K (10 mg/mL; Vivantis, Malaysia) and incubated in a dry bath at 65 °C for 40 min. Then, 3 µl of RNAse A (20 mg/mL; Vivantis, Malaysia) was added and incubated at 37 °C for 10 min, and 372 µl of BG Buffer homogeneous solution was added to the sample and incubated at 65 °C for 20 min. Washing of DNA was done with a clean glass filter membrane, to where it was transferred absolute ethanol (200 µl) and sample (558 µl); and centrifuged at 10000 x g for 1 min. The membrane was washed with 650 µl of Wash Buffer and centrifuged at 10000 x g 1 min two times for removal of residual ethanol. Pure DNA in the membrane was eluted in 30 µl pre-heated TE buffer for 2 min, centrifuged at 10000 x g for 2 min, and stored at -20 °C.
2.5.2. 16S rRNA amplification
The primers used for amplification of the 16S rRNA gene [14,15] were 27f 5’- AGA GTT TGA TCC TGG CTC AG -3’ and 1492r 5’-CTA CGG CTA CCT TGT TAC GA-3’ at 5µM (IDT, Belgium), 1X PCR-Buffer (Frilabo, Portugal), 200 µM of each dNTP in a mix (Frilabo, Portugal), 1.25 U of DFS-Taq DNA Polymerase (ThermoFisher Scientific, Portugal), 10 ng/µL of template DNA, adjusted to a 50 µL reaction. The PCR cycle was 94 °C for 2 min, followed by 30 cycles of 94 °C for 10 sec, 62 °C for 20 sec and 72 °C for 1 min. An 80 mL agarose gel 1% (w/v) prepared with 1X TAE Buffer and stained with 4.7 µL EtBr was used to load the DNA samples - 4 µL of PCR product and 1 µL 5X bromophenol blue (Frilabo, Portugal); and 1 Kb DNA (Frilabo, Portugal) Ladder (0.1µg/µL) . Electrophoresis was run at 100 V for 45 min, and the fragments (~1.5 Kb) were visualized in ChemiDocTM (BioRad, Portugal). DNA bands were cleaned up using GF-1 PCR Clean-up Kit (Vivantis, Malaysia). The volume of samples was adjusted to 100 µL with nuclease-free water and mixed with 500 µL of PCR Buffer. The sample was loaded to a glass filter membrane and centrifuged at 10000 x g for 1 min; then the membrane was washed with Wash Buffer (750 µL) and centrifuged at 10000 x g for 1 min two times to remove residual ethanol. Pure DNA in the membrane was eluted in 30 µl TE buffer for 2 min, centrifuged at 10000 x g for 2 min, and stored at -20 °C. For subsequent sequencing reactions, the purity of the amplicon (2 µl) was measured with the 260/280 nm absorbance ratio (~1.8).
2.5.3. 16S rRNA sequencing
Sequencing reactions used BigDyeTM Terminator v3.1 ready reaction mix (ThermoFisher Scientific, Portugal) with 27f and 1492r primers at (3.2 µM), 5X Sequencing Buffer and nuclease-free water to a final volume of 7 µL and mixed with 3 µL of purified amplicon. Samples were assessed in duplicate reactions. The parameters for the sequencing reaction were 96 °C for 1 min and 25 cycles of 96 °C for 10s (denature), 62 °C for 5 s (anneal) and 60 °C (extend) for 4 min. For removal of interferences with base calling, samples were purified with 60 µL of SAM/BigDyeXTerminatorTM bead solution (ThermoFisher Scientific, Portugal) and vortexed at 1800 rpm for 20 min. Capillary electrophoresis carried out in the SeqStudio Genetic Analyzer (Applied Biosystems, Portugal) was run at 12000 V for 25 S, which was more appropriate for long sequences, and the final results were analyzed using the Sequencing Analysis Software v7.0 (Applied Biosystems, Portugal).
2.6. Data analysis
2.6.1. Species identification
Sequence results were aligned with reference sequences from the National Center for Biotechnology Information (NCBI, USA) using the rRNA/ITS – 16S ribosomal RNA sequences database [16] run with the Basic Local Alignment Search Tool (BLAST) algorithm optimized for highly similar sequences [17]. Finally, sequences with identity equal or higher than 97% were accepted as the best match for the LAB isolate at the species level [18].
2.6.2. Phylogenetic tree
The phylogenetic tree was plotted with the R software (version 4.3.0, R Foundation for Statistical Computing, Vienna, Austria) [19]. Briefly, multiple sequence alignment was performed through the msa package [20] by using the ClustalOmega algorithm. A distance matrix was obtained with the seqinr package [21] to calculate the phylogenetic tree, which was built by using the Neighbor Joining (NJ) method using the ape package [22], producing an unrooted tree [23], for phylogenetic analysis. Visuals and annotations were made using the ggtree package [24] and the iTools software (version 6.8.0) [25].
2.6.3. Phenotypic characterization of LAB
Data were divided into three subsets, by foodborne pathogen: L. monocytogenes, S. Typhimurium and S. aureus. Principal component analysis (PCA) of each subset was performed to assess the contribution of the antimicrobial, proteolytic and acidifying capacities to the differentiation of isolates. The function principal() from the psych package was used in R software (version 4.3.0, R Foundation for Statistical Computing, Vienna, Austria) [19], where a varimax-rotated solution for two principal components was obtained. Projections of the sample scores onto the span of the principal components were produced by using the function prcomp from the factoextra package. Heatmaps were created using the pheatmap() function [26] to find relationship patterns within the samples.
3. Results and Discussion
3.1. Genetic identification of Lactic Acid Bacteria Isolates
The identification results of 62 isolates of LAB by 16S gene sequencing are shown in Table 1. The results show that Enterococcus faecium emerged as the most common specie, accounting for 32.3% of the total, followed by Leuconostoc mesenteroides (19.4%) and Latilactobacillus sakei (17.7%). This finding aligns with previous research on traditional Portuguese fermented meat sausages of Chouriço preto and Linguiça [27], smoked meat products, where enterococci were also detected in abundance, with counts ranging from 104 to 108 CFU/g. These Enterococcus species, particularly E. faecalis and E. faecium, are known for their enzymatic activities related to lipid and protein hydrolysis, contributing to the complex flavor profile of the alheira [27,28]. Leuconostoc species have been previously reported in meat sausages [29] and were found throughout the fermentation process [27], spanning 21 days. Interestingly, they exhibited the capability to produce bacteriocins with antilisterial activity, suggesting a potential role in enhancing the safety and quality of these products. Despite being present in lower amounts in the alheira microbiome, other research teams [30] have found L. sakei as the dominant species in the microbiome composition of Salame Piemonte, an Italian sausage, which has a short maturation period. The differences in LAB frequency between these two products are expected since Salame Piemonte is made using raw meat; whereas the pork and chicken meats used in alheira are cooked. Moreover, Lactiplantibacillus plantarum, Pediococcus pentosaceus and Weissela uvarum were identified as less frequent LAB species, constituting 6.5, 4.8, and 1.6% of the total population, respectively.
A phylogenetic tree (Figure 2) was built based on 16S gene sequences to assess the evolutionary relationships between the LAB isolates. From the results, it can be observed that L. mesenteroides and P. pentosaceus isolated from different geographical locations clustered differently; this suggests the presence of distinct strains associated with different producers. Small-scale meat processing typically involves using meat that is produced and transformed on-site, leading to greater variability among producers [31,32,33]. Moreover, the phylogenetic relationships among Enterococcus faecium isolates displayed variations dependent on the sausages collected from the CM producer, implying batch-to-batch differences, denoting that conditions within alheira sausages from a particular producer do not lead to a uniform collection of strains [1,34,35,36,37]. In an artisanal food, the manufacturing and further processing have manual aspects, leading to a less standardized final product [32,36]. By analyzing the tree topology, isolates identified as belonging to the same species were separated, suggesting a heterogeneous collection of strains for the different species obtained; similar findings were reported by [32,36]. For a more comprehensive evaluation of individual strains, it is recommended to utilize molecular techniques that differentiate DNA profiles, facilitating the assessment of genetic heterogeneity at the strain level (e.g., Random Amplified Polymorphic DNA, Denaturing Gradient Gel Electrophoresis) [14,31,38].
3.2. Alheira physicochemical and microbiological analysis
Figure 3 provides a detailed insight into the physicochemical and microbiological features of a total of 59 alheira sausages, with a subset of 22 selected samples (Figure 4) chosen based on LAB antimicrobial activity (section 2.4). This subset showcased 62 presumptive LAB. The heatmaps offer a comprehensive visualization of the interrelationships among the various attributes in both dimensions (Figure 4). A cluster formation between alheiras from MP and FN producers presented more similarities in the final composition compared with those from AG, BST, BV, SM, CSM and CM. Previous research on the physicochemical properties of alheira [32,39] has emphasized considerable variability in chemical, physical and sensory attributes from industrial and small scale producers, which can differ across localities [1,2,38]. Notably, distinct clustering occurs even within samples from a single producer, as it was the case for alheiras from the CM producer, where CM1 and CM2 exhibited greater compositional affinity than CM5, or CM3 and CM4 samples – which formed their own cluster. Such differences could result from small variations on the recipe or manufacturing processes [2]. Overall, the samples presented lower values for ashes, pH and aw among the assayed physicochemical parameters, and higher values for moisture and CHO (%), while protein and fat content presented intermediate values. The reduction of aw and pH during processing of the alheira contributes for the inhibition of pathogens development [1,3]. The microbiological counts of C. perfringens (>0.699-1.699 log CFU/g) and S. aureus (1.699-6.021 log CFU/g) were lower than those of mesophiles (7.161-9.679 log CFU/g) and LAB (7.704 -11.00 log CFU/g for presumptive Lactobacillus and Lactococcus) This aligns with previous findings on microbiological characterization of alheiras – LAB counts (5.900-10.50 log CFU/g) and S. aureus (<1.0-4.1 log CFU/g) [1,35]. LAB are the dominant microorganisms in alheira. The variables more closely related were ashes, pH, C. perfringens and S. aureus counts and aw. Additionally, a strong correlation was noted between mesophiles and LAB (presumptive Lactobacillus and Lactococcus), consistent with common attributes in dry fermented sausages [38], highlighting the impact of alterations in meat properties on parallel changes in the microbiological composition.
3.3. Lactic acid bacteria phenotypic and genetic analysis
The relationships pattern among the physicochemical and genetic features of the sixty-two LAB isolates are shown in Figure 5 and the summary statistics are shown in the Supplementary material (Table 1). These isolates exhibited two primary clusters: 1) First, comprised of LABs from the CM producer; and 2) Second, another one encompassing LABs isolated from alheiras of other artisanal producers. All samples within this cluster were genetically identified as Enterococcus faecium species, originating from isolation on M17 medium. The mean pH at 6 h for this species was 5.78, the lowest pH value in the acidification capacity assay. The significant pH drops were between time intervals of 3 and 6 h. Even though E. faecium had a higher pH drop, it did not seem to influence pathogen inhibition. This species was limited in antimicrobial activity (evaluated at 10 and 37 °C) against the foodborne pathogens tested by the agar spot assay, resulting in inhibition of L. monocytogenes (11.29 and 8.19 mm), S. aureus (4.8 and 0.18 mm) and S. Typhimurium (4.26 and 0 mm). Enterocins isolated from E. faecium have been shown to possess antilisterial activity, namely Enterocin A and B, found to be produced by E. faecium isolated from Spanish dry fermented sausages [37,40]. Moreover, this species was found to have a mean proteolytic activity of 0.055 mm, lower compared to those of other LAB, although previous research [39] has shown that by the action of enzymes able to hydrolyze casein, bovine serum albumin and β-lactoglobulin, the presence of enterococci in fermented sausages results in organoleptically unique products which contribute to the local cuisine and heritage of the region [37,39]. A distinct cluster formed by LABs from producers AG, SM and TB stood out for demonstrating: (i) heightened pH drop values between 3 and 6 h; and (ii) overall superior inhibition values against the three tested foodborne pathogens across both temperatures. This cluster was formed by LAB of the L. mesenteroides, L. paracasei and Lactiplantibacillus species, all of them were isolated from the MRS medium. L. paracasei had the highest proteolytic activity value (3.46 mm), followed by L. mesenteroides (3.43 mm). Moreover, L. paracasei had one of the lowest mean pH (pH = 6.03 at time = 6 h). Lactiplantibacillus species were the most producing strains of L-lactic acid, within the first four hours of incubation, the species L. herbarum yielded 0.55 g/L and L. plajomi 0.42 g/L. The inhibition of pathogens could be mainly attributed to Lactiplantibacillus and Lacticaseibacillus strains, the inhibition of L. monocytogenes at 10 °C was higher by L. plajomi (ID=21.62 mm), followed by L. plantarum (ID=19.77 mm). At 37 °C, the LAB with higher antilisterial activity were L. herbarum (ID=11.79 mm) and L. sakei (ID=11.35 mm). These findings suggest that the species with the most effective antilisterial activity vary with temperature, indicating that inhibition could be occurring by the action of bacteriocins, each with a different optimal temperature for catalytic activity. This is exemplified by the reduced efficiency of L. plajomi and L. plantarum at higher temperatures. Similar patterns were observed in LAB's antimicrobial activity against S. aureus: at 10 °C, L. plajomi (ID=11.91 mm) and L. herbarum (ID=11.29 mm) presented the highest mean inhibition diameters compared to the other LAB; whereas at 37 °C, L. paracasei (ID=9.06 mm) and L. plantarum (ID=8.29 mm), the highest mean values of inhibition, were lower when compared to the antimicrobial activity assayed at 10 °C. Antimicrobial activity against S. Typhimurium remains consistent at both temperatures. At 10 °C, L. herbarum (ID=11.21 mm) and L. mesenteroides (ID=10.89 mm) exhibit the highest inhibition, while at 37 °C, L. mesenteroides (ID=12.89 mm) and L. plantarum (ID=11.56 mm) are the ones with highest antimicrobial activity. The mechanism of inhibition could have been mediated by bacteriocin production, in this case, the results imply that optimal bacteriocin activity can be influenced by higher temperatures. Additionally, the proteolytic activity of LAB could influence the generation of antimicrobial peptides (e.g., through casein hydrolysis, especially in class II bacteriocins) [41]. This phenomenon could clarify why L. mesenteroides, the strain with the highest proteolytic activity, showed greater inhibition of this pathogen.
To undertake a more comprehensive investigation into the variability of physicochemical attributes exhibited by LAB, a principal components analysis (PCA) was employed. Isolates were annotated with their species-level genetic identification. The dataset was segregated to account for the presence of the three foodborne pathogens that were the subject of analysis, yielding three distinct PCA maps (Table 2).
For each map, two principal components were retained (based on their eigenvalues surpassing 1), accounting for a substantial portion of the variation (between 78.5% and 82.4%). In the context of Figure 6, an analysis was conducted on a subset of LAB antimicrobial activity targeting S. Typhimurium at 37 °C and 10 °C (PC1: 66.2%; PC2: 16.2%). The results revealed that L. mesenteroides was the LAB specie responsible for the higher inhibition (value) of this pathogen at 37 °C. The variables ΔpH36, ΔpH06, pH6 and the inhibition diameter at both temperatures presented the highest loadings, therefore, this component primarily measured the antimicrobial activity and acidifying capacity of the LAB. Within the second component, there was stronger positive correlations with the variables ΔpH03 and proteolytic activity, while LAC exhibited more negative associations compared to the first component. A similar pattern was observed for S. aureus (Figure 7; PC1: 62.4%; PC2: 16.1%) and L. monocytogenes assessments (Figure 8; PC1: 64.4%; PC2: 15.2%). However, the influence of different temperatures on inhibition diameters was more evident in the latter, as indicated by the second component. The overarching acidifying capacity was considerably attributed to the action of E. faecium.
4. Conclusions
The careful selection of lactic acid bacteria (LAB) as starter cultures or biological food-grade preservatives in meat sausages is pivotal for regulating the fermentation process – ensuring product safety by inhibiting foodborne pathogens while contributing to the desired organoleptic characteristics, and allowing producers to standardize product characteristics, enhancing consistency and quality. This study provides valuable insights into the LABs community of alheira sausages, E. faecium emerged as the most prevalent species of LAB (32.3%), followed by L. mesenteroides (19.4%) and L. sakei (17.7%). These microbial populations significantly influence the flavor and quality of these traditional Portuguese sausages, many of which hold protected designation of origin certification, attesting to their unique quality. Enterococci, Leuconostoc spp., and lactobacilli play a pivotal role in flavor formation through diverse metabolic pathways, including proteolysis. Isolating wild-type strains from traditional products reveals diverse metabolic profiles; this makes them promising candidates for applications in food fermentation. E. faecium showed great acidifying capacity by the pH drops between time intervals 3 and 6 h which culminated in the lowest final pH compared to the other LAB samples. Lactiplantibacillus and Lacticaseibacillus strains presented strong inhibition diameters against all three pathogens tested, and L. mesenteroides presented high proteolytic activity and antimicrobial inhibition especially of S. Typhimurium. The phylogenetic analysis revealed distinct strains of LAB based on geographical locations, suggesting that different producers may yield unique microbial profiles. The observed batch-to-batch variations, indicate that conditions within alheira sausages from a specific producer do not lead to a uniform collection of strains. These variations have implications for product consistency and may warrant further investigation into genetic heterogeneity. The absence of standardized production processes for alheira sausage leads to a microbiologically unstable product, causing variations in quality. Implementing management practices for food production with a standardized approach focused on enhancing quality and safety could potentially elevate the market for artisanal products. These products, integral to Mediterranean culture, biodiversity, and the economy, warrant thorough investigation and protection.
Author Contributions
Conceptualization, V.C. and U.G.-B.; methodology, N.F., A.S.F., C.R., U.G.-B., V.C.; software, N.F., L.C., V.C. and U.G.-B.; validation, N.F., V.C. and U.G.-B..; formal analysis, N.F., A.S.F., L.C., A.C., C.R. and U.G.-B..; investigation, N.F., A.S.F., V.C., U.G.-B.; resources, A.C., C.R., V.C. and U.G.-B.; data curation, N.F., A.S.F., V.C. and U.G.-B.; writing—original draft preparation, N.F.; writing—review and editing, N.F., V.C. and U.G.-B.; visualization, N.F., V.C. and U.G.-B.; supervision, V.C. and U.G.-B.; project administration, V.C. and U.G.-B.; funding acquisition, V.C. and U.G.-B. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the Foundation for Science and Technology (FCT, Portugal) financial support through national funds FCT/MCTES (PIDDAC) to CIMO (UIDB/00690/2020 and UIDP/00690/2020) and SusTEC (LA/P/0007/2021). The authors are grateful to EU PRIMA and the Portuguese Foundation for Science and Technology (FCT) for funding the PAS-AGRO-PAS (PRIMA/0014/2022) project.
Data Availability Statement
Summary data available upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ferreira V, Barbosa J, Vendeiro S, Mota A, Silva F, Monteiro MJ, et al. Chemical and microbiological characterization of alheira: A typical Portuguese fermented sausage with particular reference to factors relating to food safety. Meat Sci. 2006;73(4):570–5. [CrossRef]
- Coelho-Fernandes S, Zefanias O, Rodrigues G, Faria AS, Fernandes Â, Barros L, et al. Microbiological and Physicochemical Assessment of Artisanally Produced “Alheira” Fermented Sausages in Northern Portugal. Proceedings. 2020;70(1):16.
- Esteves A, Patarata L, Saraiva C, Martins C. Assessment of the Microbiological Characteristics of Industrially Produced Alheira, with Particular Reference to Foodborne Pathogens. J Food Saf. 2008 Feb;28(1):88–102. [CrossRef]
- Mokoena MP. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules. 2017 Aug;22(8):1255. [CrossRef]
- 5. Doyle MP, Steenson LR, Meng J. Bacteria in Food and Beverage Production. In: Rosenberg E, DeLong EF, Lory S, Stackebrandt E, Thompson F, editors. The Prokaryotes: Applied Bacteriology and Biotechnology [Internet]. Berlin, Heidelberg: Springer; 2013 [cited 2023 Aug 21]. p. 241–56. [CrossRef]
- Khalid K. An overview of lactic acid bacteria. IJB. 2011;V1(N3):P1-13.
- 14:00-17:00. ISO. [cited 2023 Oct 9]. ISO 1442:2023. Available from: https://www.iso.org/standard/82664.html.
- 14:00-17:00. ISO. [cited 2023 Oct 9]. ISO 937:2023. Available from: https://www.iso.org/standard/82663.html.
- 14:00-17:00. ISO. [cited 2023 Oct 9]. ISO 936:1998. Available from: https://www.iso.org/standard/24783.html.
- 14:00-17:00. ISO. [cited 2023 Oct 9]. ISO 6888-1:2021. Available from: https://www.iso.org/standard/76672.html.
- Dwivedi HP, Smiley RD, Pincus DH. 11. Rapid Methods for the Detection and Identification of Foodborne Pathogens. In: Compendium of Methods for the Microbiological Examination of Foods [Internet]. American Public Health Association; 2013 [cited 2023 Oct 9]. Available from: https://ajph.aphapublications.org/doi/full/10.2105/MBEF.0222.016.
- Nutrition C for FS and A. BAM Chapter 5: Salmonella. FDA [Internet]. 2023 Sep 18 [cited 2023 Oct 9]; Available from: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-5-salmonella.
- Faria AS, Fernandes N, Cadavez V, Gonzales-Barron U. Screening of Lactic Acid Bacteria Isolated from Artisanally Produced Alheira Fermented Sausages as Potential Starter Cultures. 2021.
- Mohania D, Nagpal R, Kumar M, Bhardwaj A, Yadav M, Jain S, et al. Molecular approaches for identification and characterization of lactic acid bacteria. J Dig Dis. 2008;9(4):190–8. [CrossRef]
- Hou Q, Bai X, Li W, Gao X, Zhang F, Sun Z, et al. Design of Primers for Evaluation of Lactic Acid Bacteria Populations in Complex Biological Samples. Front Microbiol [Internet]. 2018 [cited 2022 Mar 28];9. [CrossRef]
- Sayers EW, Bolton EE, Brister JR, Canese K, Chan J, Comeau DC, et al. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022 Jan 7;50(D1):D20–6.
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990 Oct 5;215(3):403–10.
- Wilhelm H. Holzapfel, Brian J.B. Wood. Lactic Acid Bacteria : Biodiversity and Taxonomy. Chichester, West Sussex, UK: Wiley-Blackwell; 2014.
- R Core Team. R: A Language and Environment for Statistical Computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2021. Available from: https://www.R-project.org/.
- Bonatesta E, Horejs-Kainrath C, Bodenhofer U. msa: Multiple Sequence Alignment [Internet]. Bioconductor version: Release (3.17); 2023 [cited 2023 Oct 10]. Available from: https://bioconductor.org/packages/msa/.
- Charif D, Clerc O, Frank C, Lobry JR, Necşulea A, Palmeira L, et al. seqinr: Biological Sequences Retrieval and Analysis [Internet]. 2023 [cited 2023 Oct 10]. Available from: https://cran.r-project.org/web/packages/seqinr/index.html.
- Paradis E, Blomberg S, Bolker [aut B, cph, Brown J, Claramunt S, et al. ape: Analyses of Phylogenetics and Evolution [Internet]. 2023 [cited 2023 Oct 10]. Available from: https://cran.r-project.org/web/packages/ape/index.html.
- Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol. 1987 Jul 1;4(4):406–25. [CrossRef]
- Yu G, Lam TTY, Xu S, Li L, Jones B, Silverman J, et al. ggtree: an R package for visualization of tree and annotation data [Internet]. Bioconductor version: Release (3.17); 2023 [cited 2023 Oct 10]. Available from: https://bioconductor.org/packages/ggtree/.
- Dinov ID, Rubin D, Lorensen W, Dugan J, Ma J, Murphy S, et al. iTools: A Framework for Classification, Categorization and Integration of Computational Biology Resources. PLOS ONE. 2008 May 28;3(5):e2265. [CrossRef]
- Kolde R. pheatmap: Pretty Heatmaps [Internet]. 2019 [cited 2023 Aug 31]. Available from: https://cran.r-project.org/web/packages/pheatmap/index.html.
- Correia Santos S, Fraqueza MJ, Elias M, Salvador Barreto A, Semedo-Lemsaddek T. Traditional dry smoked fermented meat sausages: Characterization of autochthonous enterococci. LWT - Food Sci Technol. 2017 Jun 1;79:410–5. [CrossRef]
- Fraqueza MJ. Antibiotic resistance of lactic acid bacteria isolated from dry-fermented sausages. Int J Food Microbiol. 2015 Nov 6;212:76–88. [CrossRef]
- Moracanin SV, Turubatovic L, Skrinjar M, Obradovic D. Antilisterial Activity of Bacteriocin Isolated from Leuconostoc mesenteroides ssp mesenteroides IMAU:10231 in the Production of Sremska Sausages: Lactic Acid Bacteria Isolation, Bacteriocin Identification and Meat Application Experiments. FOOD Technol Biotechnol. 2013 Jun;51(2):247–56.
- Franciosa I, Ferrocino I, Giordano M, Mounier J, Rantsiou K, Cocolin L. Specific metagenomic asset drives the spontaneous fermentation of Italian sausages. Food Res Int. 2021 Jun 1;144:110379. [CrossRef]
- Albano H, van Reenen CA, Todorov SD, Cruz D, Fraga L, Hogg T, et al. Phenotypic and genetic heterogeneity of lactic acid bacteria isolated from “Alheira”, a traditional fermented sausage produced in Portugal. Meat Sci. 2009;82(3):389–98.
- Patarata L, Judas I, Silva JA, Esteves A, Martins C. A comparison of the physicochemical and sensory characteristics of alheira samples from different-sized producers. Meat Sci. 2008 May 1;79(1):131–8. [CrossRef]
- Joint FAO/WHO Expert Consultation on. 2001;
- Fernandes N, Faria AS, Carvalho L, Choupina A, Rodrigues C, Cadavez V, et al. Molecular Identification of Lactic Acid Producing Bacteria Isolated from Alheira, a Traditional Portuguese Fermented Sausage. Biol Life Sci Forum. 2022;18(1):73.
- Albano H, Henriques I, Correia A, Hogg T, Teixeira P. Characterization of microbial population of ‘Alheira’(a traditional Portuguese fermented sausage) by PCR-DGGE and traditional cultural microbiological methods. J Appl Microbiol. 2008;105(6):2187–94. [CrossRef]
- Albano H, van Reenen CA, Todorov SD, Cruz D, Fraga L, Hogg T, et al. Phenotypic and genetic heterogeneity of lactic acid bacteria isolated from “Alheira”, a traditional fermented sausage produced in Portugal. Meat Sci. 2009 Jul;82(3):389–98.
- Franz CMAP, Stiles ME, Schleifer KH, Holzapfel WH. Enterococci in foods—a conundrum for food safety. Int J Food Microbiol. 2003 Dec 1;88(2):105–22. [CrossRef]
- Liu Y, Wan Z, Yohannes KW, Yu Q, Yang Z, Li H, et al. Functional Characteristics of Lactobacillus and Yeast Single Starter Cultures in the Ripening Process of Dry Fermented Sausage. Front Microbiol [Internet]. 2021 [cited 2023 Sep 1];11. [CrossRef]
- Worsztynowicz P, Schmidt AO, Białas W, Grajek W. Identification and partial characterization of proteolytic activity of Enterococcus faecalis relevant to their application in dairy industry. Acta Biochim Pol. 2019 Feb 3;66(1):61–9. [CrossRef]
- Hugas M. Bacteriocinogenic lactic acid bacteria for the biopreservation of meat and meat products. Meat Sci. 1998 Jan 1;49:S139–50.
- O’Sullivan L, Ross RP, Hill C. Potential of bacteriocin-producing lactic acid bacteria for improvements in food safety and quality. Biochimie. 2002 May 1;84(5):593–604. [CrossRef]
Figure 1.
Map of the geographical location of different alheira collection sites. Source: Instituto Nacional de Estatística (INE).
Figure 1.
Map of the geographical location of different alheira collection sites. Source: Instituto Nacional de Estatística (INE).
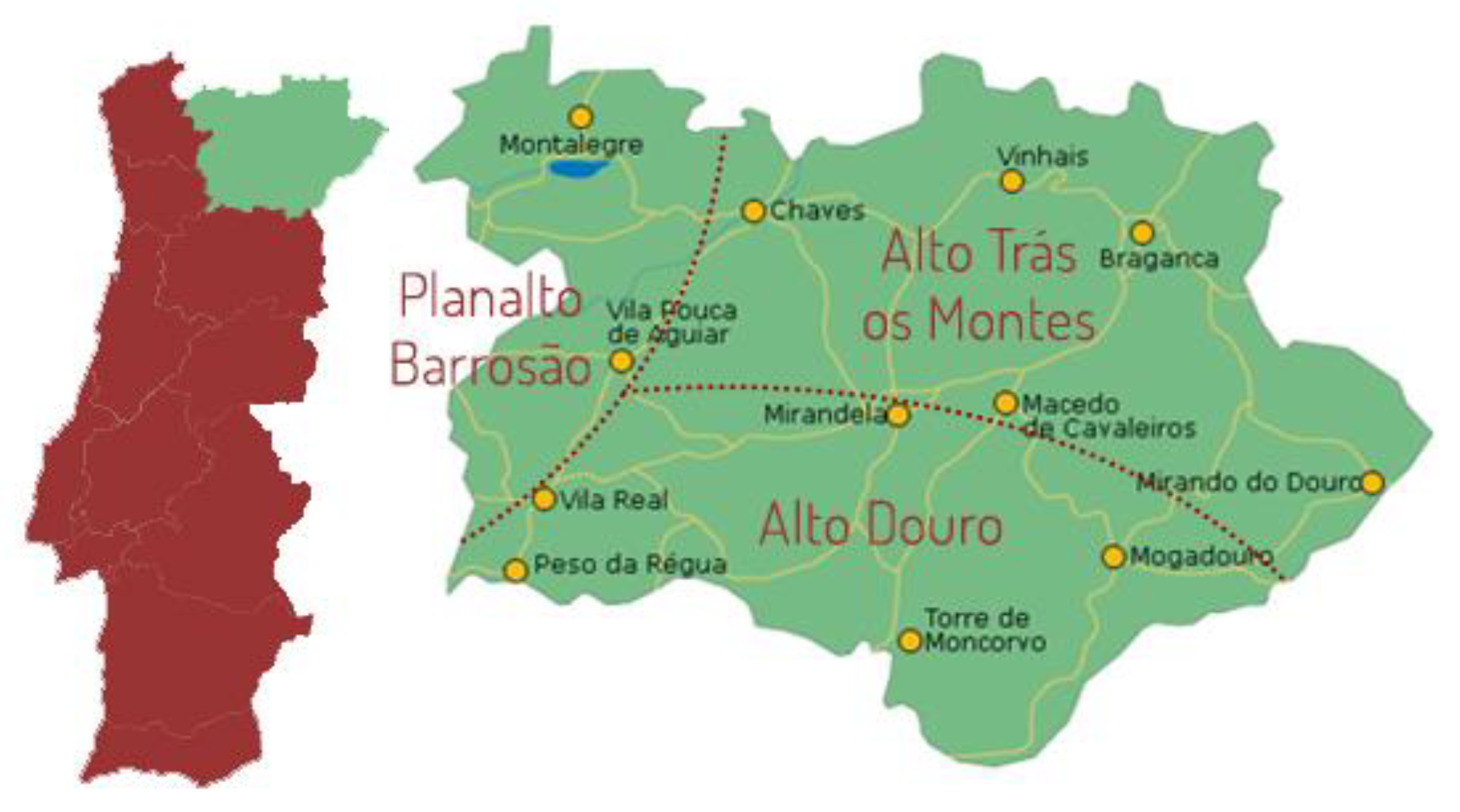
Figure 2.
Phylogenetic tree of 16S rRNA sequences from LAB isolates with information at species (a) and genus (b) level.
Figure 2.
Phylogenetic tree of 16S rRNA sequences from LAB isolates with information at species (a) and genus (b) level.
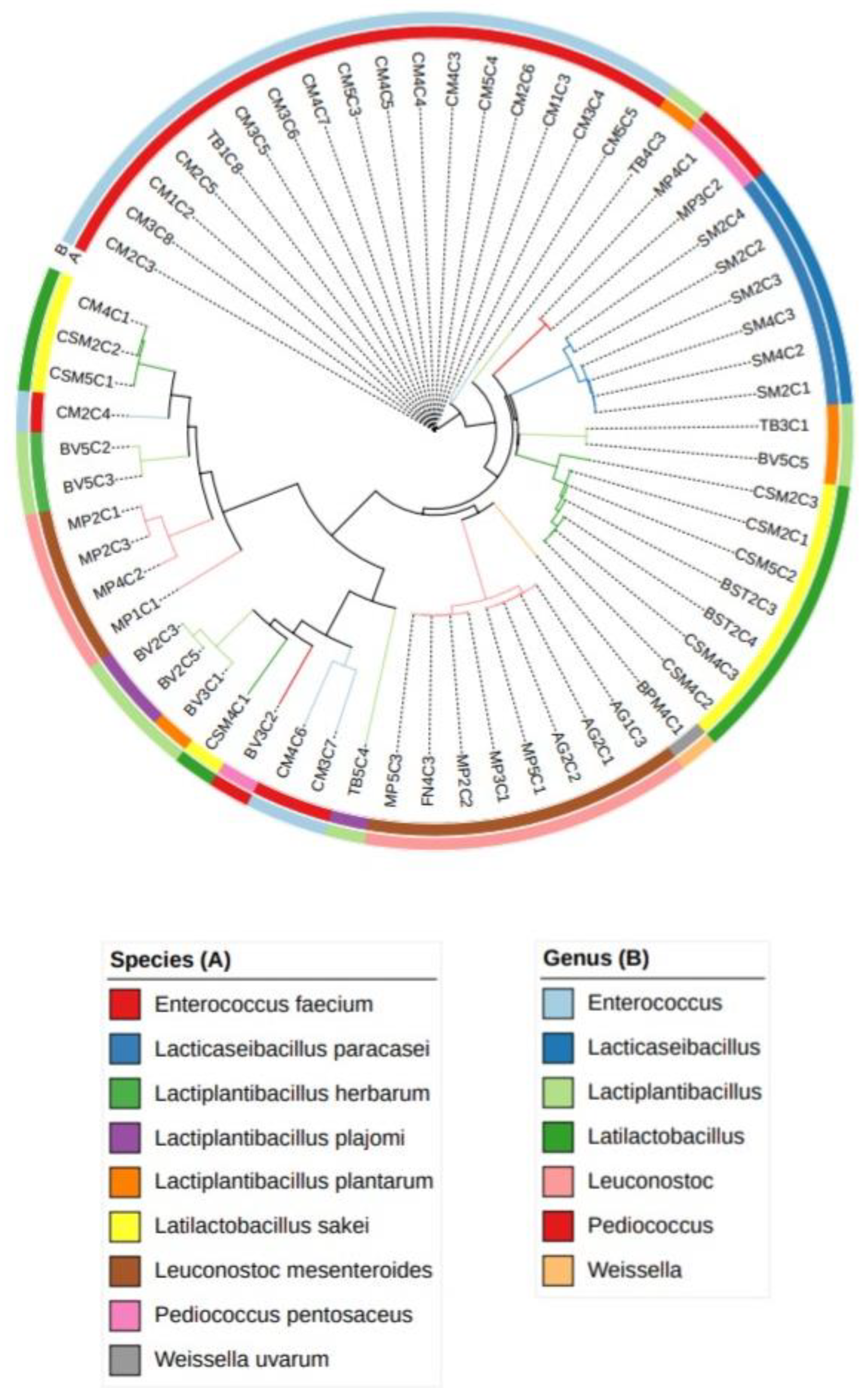
Figure 3.
Heatmap of physicochemical and microbiological data of alheira sausages (n=59). The variables are described in section 2.2.
Figure 3.
Heatmap of physicochemical and microbiological data of alheira sausages (n=59). The variables are described in section 2.2.
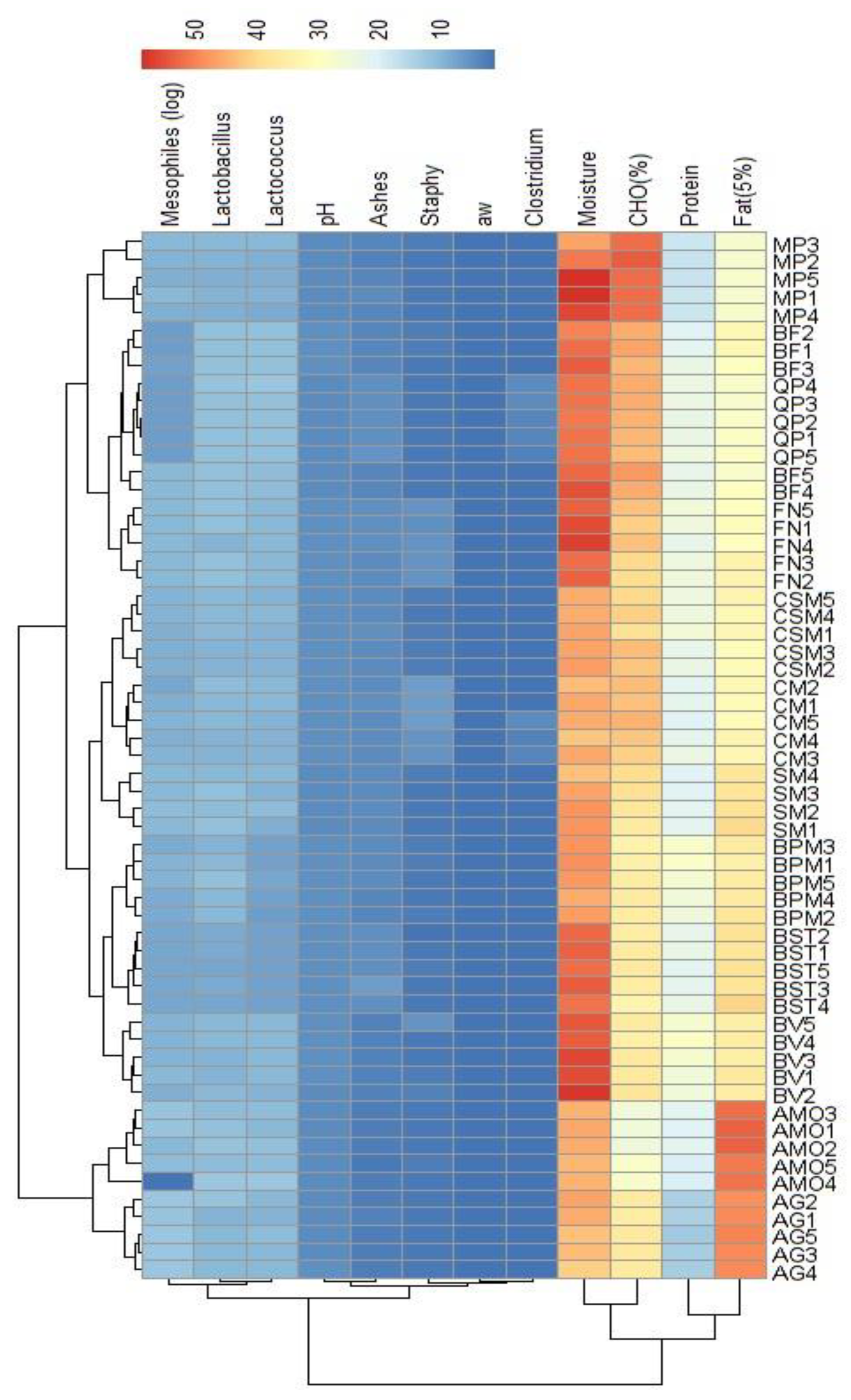
Figure 4.
Heatmap of physicochemical and microbiological data of selected alheira sausages (n=22). The variables are described in section 2.2.
Figure 4.
Heatmap of physicochemical and microbiological data of selected alheira sausages (n=22). The variables are described in section 2.2.
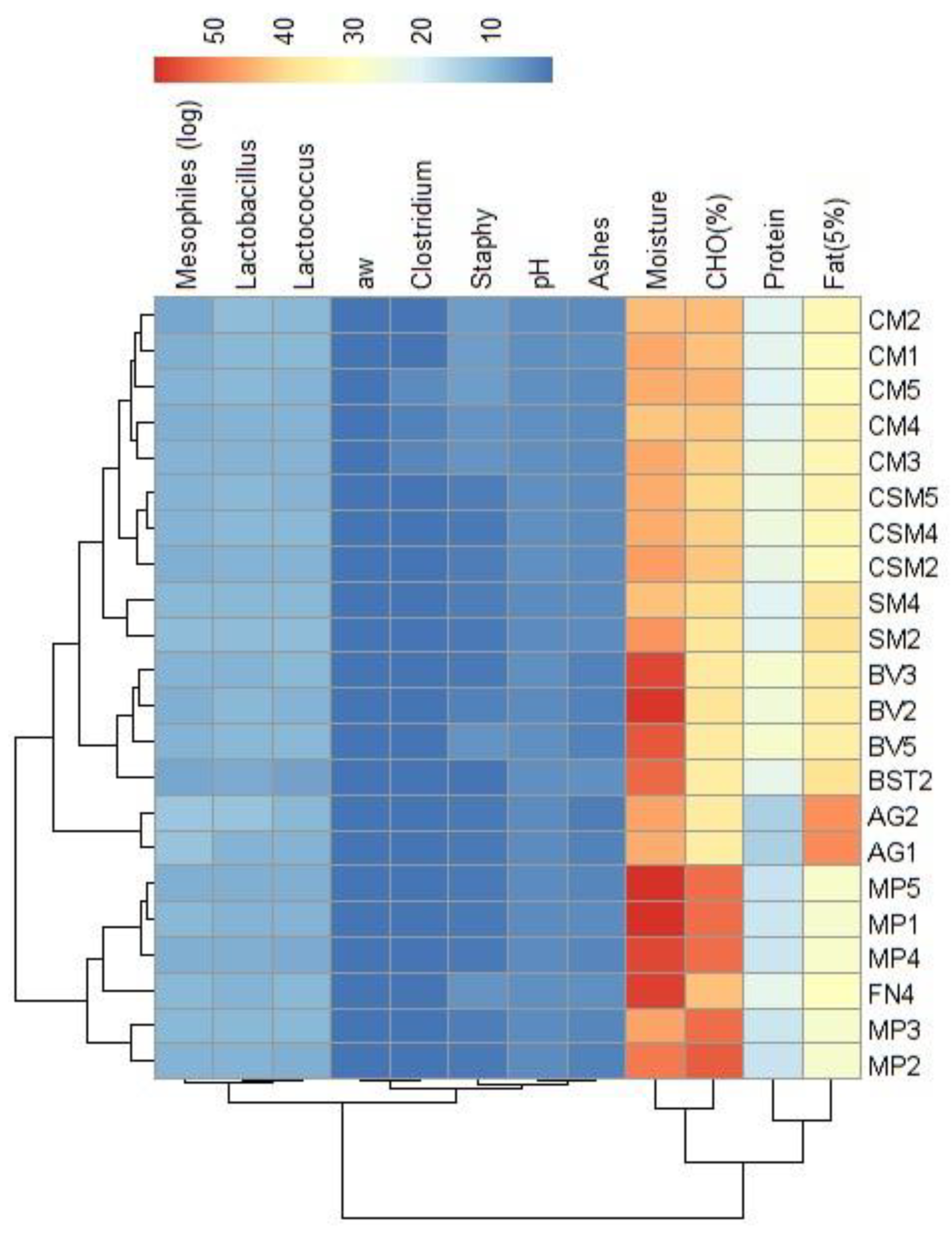
Figure 5.
Heatmap of physicochemical characterization data of lactic acid bacteria (LAB) isolated from alheira and species identification of LAB isolates. Legend: Mean inhibition diameter (ID) in mm at 10 °C for L. monocytogenes (ID_10 °C Listeria), S. Typhimurium (ID_10 °C Salmo), S. aureus (ID_10 °C Staphy) and at 37 °C (ID_37 °C Listeria), (ID_37 °C Staphy) and (ID_37 °C Salmo). pH values: pH6 (after 6 h), Δ03 (pH drop between 0 h and 3 h), Δ06 (pH drop between 0 h and 6 h), Δ36 (pH drop between 3 h and 6 h). Proteolytic activity (ProteolyticAct) in mm and L-lactic acid concentration (LAC) in g/L.
Figure 5.
Heatmap of physicochemical characterization data of lactic acid bacteria (LAB) isolated from alheira and species identification of LAB isolates. Legend: Mean inhibition diameter (ID) in mm at 10 °C for L. monocytogenes (ID_10 °C Listeria), S. Typhimurium (ID_10 °C Salmo), S. aureus (ID_10 °C Staphy) and at 37 °C (ID_37 °C Listeria), (ID_37 °C Staphy) and (ID_37 °C Salmo). pH values: pH6 (after 6 h), Δ03 (pH drop between 0 h and 3 h), Δ06 (pH drop between 0 h and 6 h), Δ36 (pH drop between 3 h and 6 h). Proteolytic activity (ProteolyticAct) in mm and L-lactic acid concentration (LAC) in g/L.
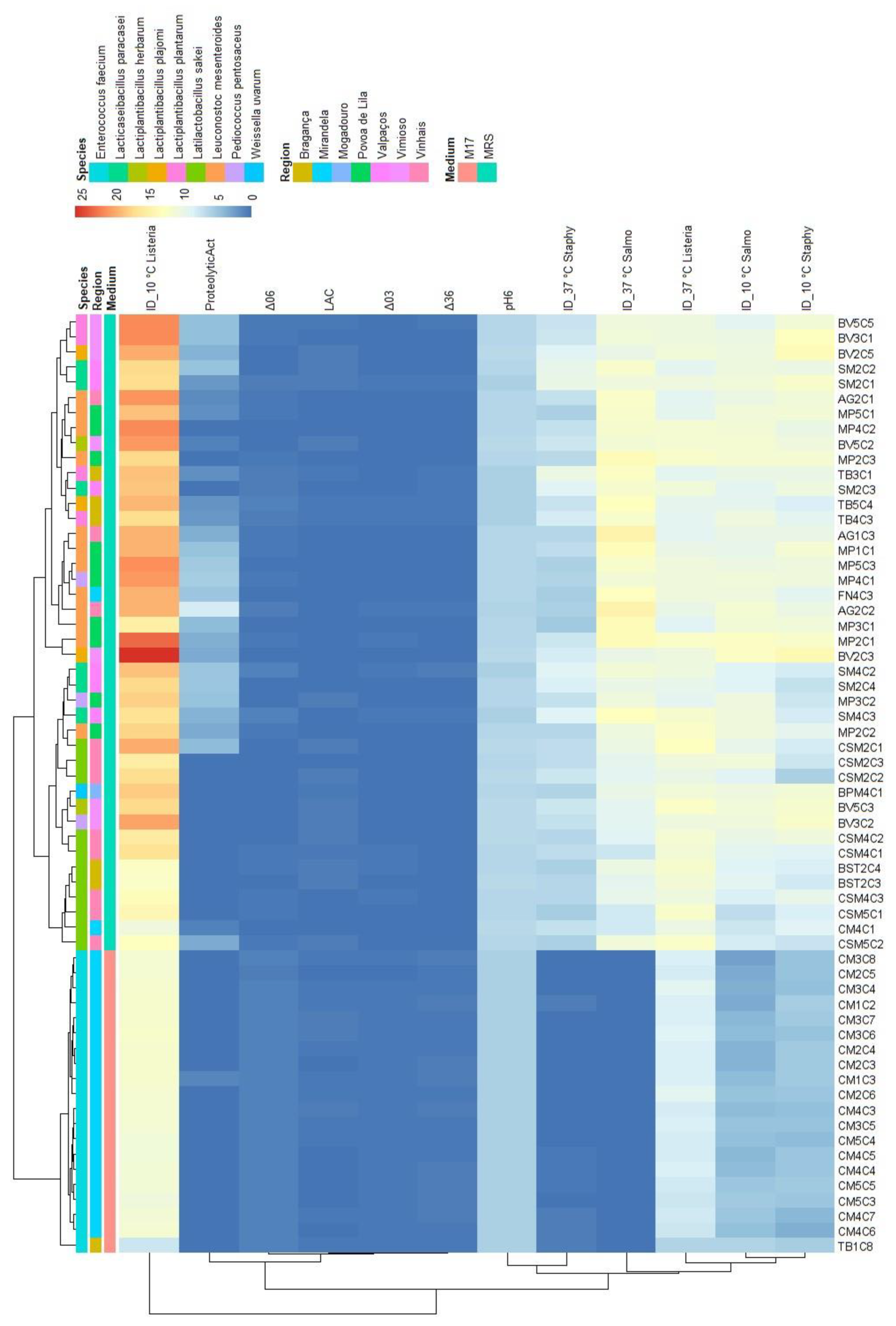
Figure 6.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates. Legend: Delta03, Delta06, Delta36: pH decreases between t= 0 and 3 h, t= 0 and 6 h and t= 3 h and t= 6 h, respectively; pH6: pH value after 6 h at 30 °C; ProteolyticAct: diameter of proteolytic activity (mm); LAC: L-lactic acid (g/L); ID_37C and ID_10C: diameter of inhibition (mm) of S. Typhimurium tested at 37 °C and 10 °C, respectively.
Figure 6.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates. Legend: Delta03, Delta06, Delta36: pH decreases between t= 0 and 3 h, t= 0 and 6 h and t= 3 h and t= 6 h, respectively; pH6: pH value after 6 h at 30 °C; ProteolyticAct: diameter of proteolytic activity (mm); LAC: L-lactic acid (g/L); ID_37C and ID_10C: diameter of inhibition (mm) of S. Typhimurium tested at 37 °C and 10 °C, respectively.
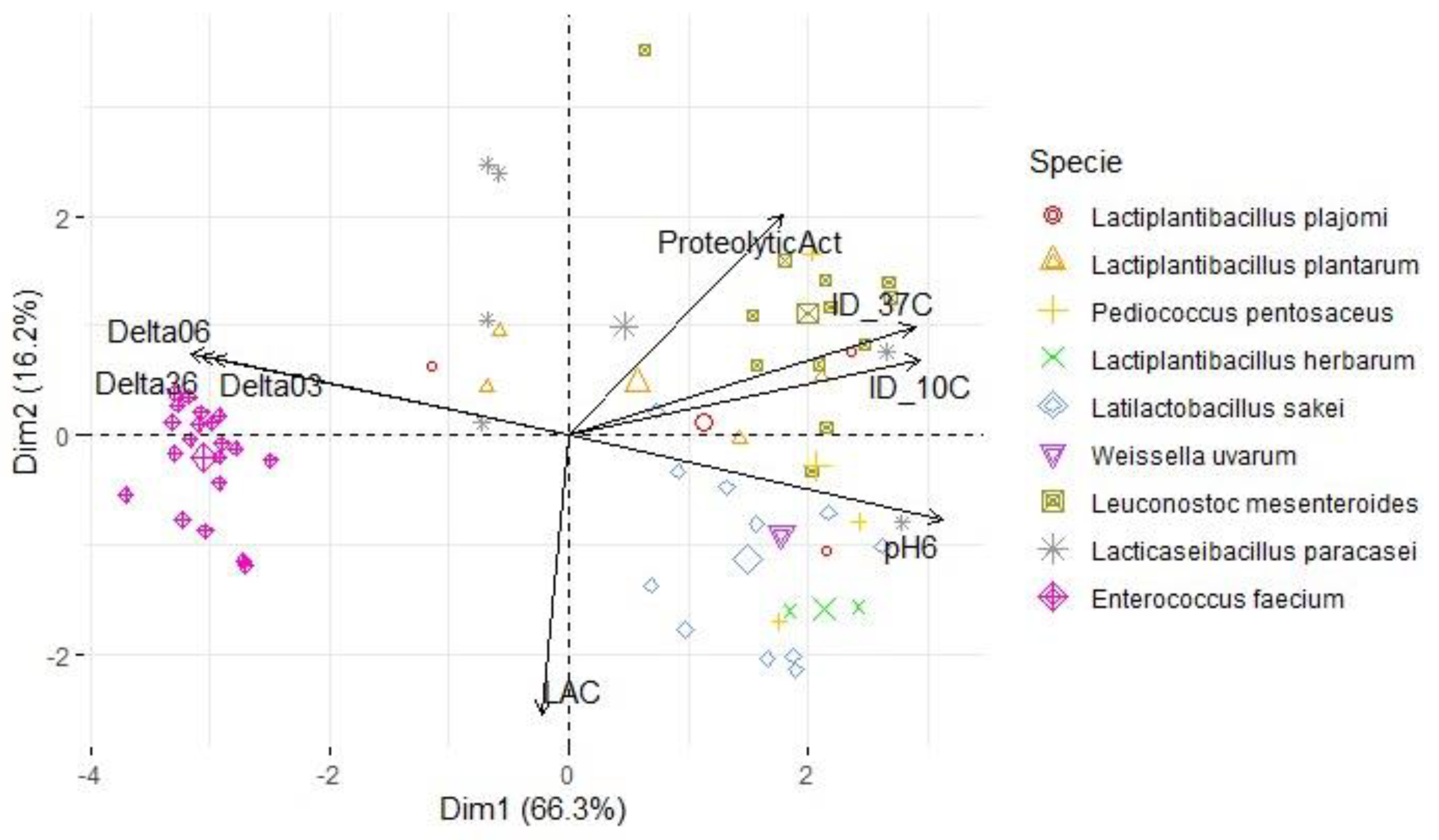
Figure 7.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates. Legend: Delta03, Delta06, Delta36: pH decreases between t= 0 and 3 h, t= 0 and 6 h and t= 3 h and t= 6 h, respectively; pH6: pH value after 6 h at 30 °C; ProteolyticAct: diameter of proteolytic activity (mm); LAC: L-lactic acid (g/L); ID_37C and ID_10C: diameter of inhibition (mm) of L. monocytogenes tested at 37 °C and 10 °C, respectively.
Figure 7.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates. Legend: Delta03, Delta06, Delta36: pH decreases between t= 0 and 3 h, t= 0 and 6 h and t= 3 h and t= 6 h, respectively; pH6: pH value after 6 h at 30 °C; ProteolyticAct: diameter of proteolytic activity (mm); LAC: L-lactic acid (g/L); ID_37C and ID_10C: diameter of inhibition (mm) of L. monocytogenes tested at 37 °C and 10 °C, respectively.
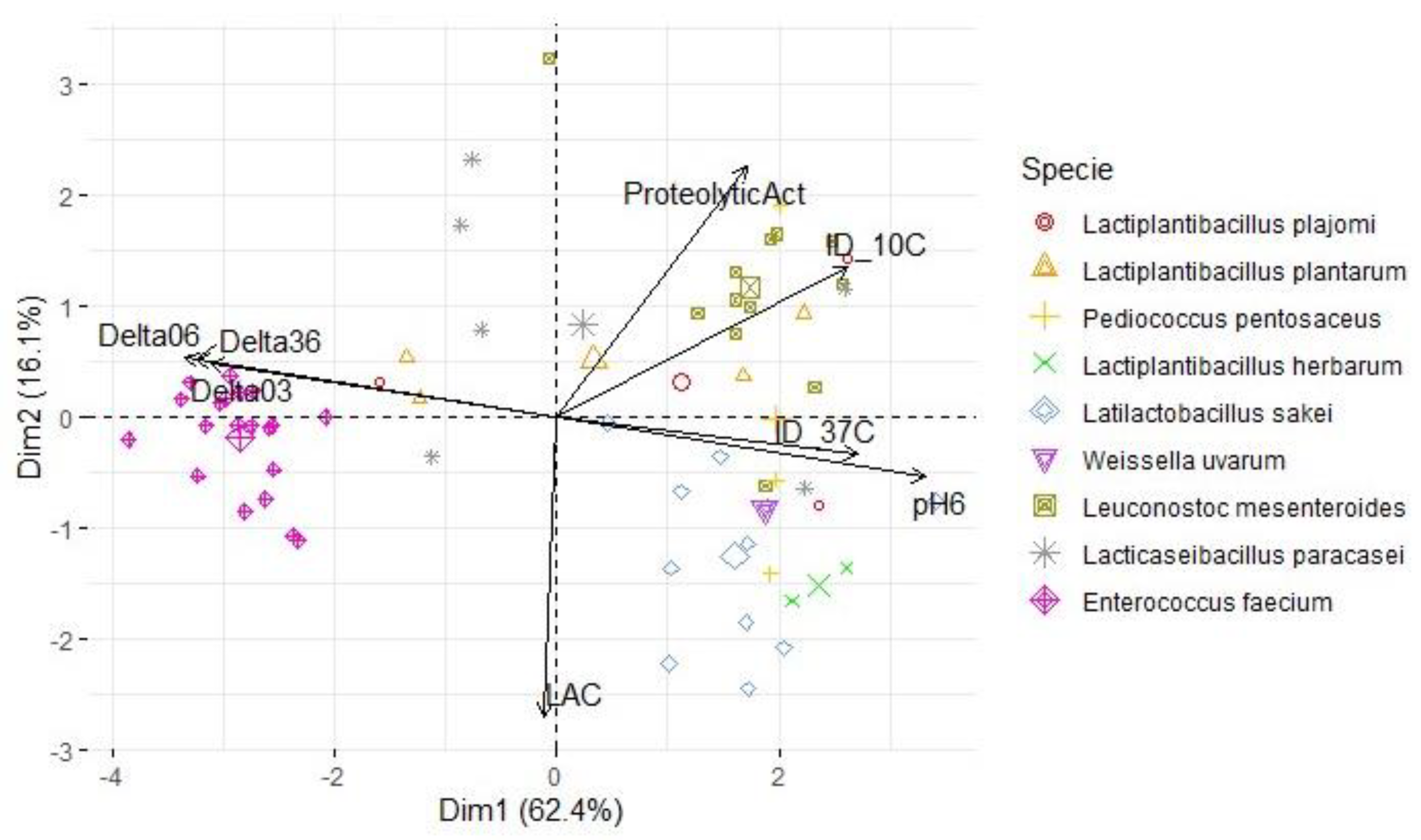
Figure 8.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates. Legend: Delta03, Delta06, Delta36: pH decreases between t= 0 and 3 h, t= 0 and 6 h and t= 3 h and t= 6 h, respectively; pH6: pH value after 6 h at 30 °C; ProteolyticAct: diameter of proteolytic activity (mm); LAC: L-lactic acid (g/L); ID_37C and ID_10C: diameter of inhibition (mm) of S. aureus tested at 37 °C and 10 °C, respectively.
Figure 8.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates. Legend: Delta03, Delta06, Delta36: pH decreases between t= 0 and 3 h, t= 0 and 6 h and t= 3 h and t= 6 h, respectively; pH6: pH value after 6 h at 30 °C; ProteolyticAct: diameter of proteolytic activity (mm); LAC: L-lactic acid (g/L); ID_37C and ID_10C: diameter of inhibition (mm) of S. aureus tested at 37 °C and 10 °C, respectively.

Table 1.
Identification results of 62 isolates of LAB by 16S gene sequencing. The main identity expressed is the mean percentage and in brackets are the [minimum; maximum] individual values.
Table 1.
Identification results of 62 isolates of LAB by 16S gene sequencing. The main identity expressed is the mean percentage and in brackets are the [minimum; maximum] individual values.
| N. isolates | Species | GenBank_ID | Identity (%) |
|---|---|---|---|
| 20 | Enterococcus faecium | NR_115764.1 | 99.2 [98.4; 100.0] |
| 6 | Lacticaseibacillus paracasei | NR_025880.1 | 100.0 [100; 100.0] |
| 2 | Lactiplantibacillus herbarum | NR_145899.1 | 99.5 [99.3; 99.6] |
| 3 | Lactiplantibacillus plajomi | NR_136785.1 | 98.9 [98; 99.8] |
| 4 | Lactiplantibacillus plantarum | NR_042394.1 | 98.5 [97; 100.0] |
| 11 | Latilactobacillus sakei | NR_115172.1 | 99.8 [99.6; 100.0] |
| 4 8 |
Leuconostoc mesenteroides | NR_074957.1 NR_157602.1 |
98.5 [97; 100.0] 99.9 [99.8;100.0] |
| 3 | Pediococcus pentosaceus | NR_042058.1 | 99.5 [99.4; 99.6] |
| 1 | Weissela uvarum | NR_134229.1 | 100.0 |
Table 2.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates.
Table 2.
Map of the first and second principal components of the tested technological properties of LAB isolated from alheira sausage and species identification of LAB isolates.
| Variables | S. Typhimurium | L. monocytogenes | S. aureus | |||
| PC1 | PC2 | PC1 | PC2 | PC1 | PC2 | |
| ProteolyticAct | 0.538 | 0.609 | 0.494 | 0.653 | 0.508 | 0.612 |
| pH6 | 0.942 | -0.232 | 0.960 | -0.159 | 0.950 | -0.175 |
| ΔpH03 | -0.896 | 0.212 | -0.903 | 0.137 | -0.901 | 0.173 |
| ΔpH06 | -0.956 | 0.225 | -0.968 | 0.152 | -0.961 | 0.173 |
| ΔpH36 | -0.927 | 0.218 | -0.940 | 0.149 | -0.931 | 0.162 |
| LAC | -0.068 | -0.771 | -0.032 | -0.783 | -0.032 | -0.812 |
| ID_10C | 0.884 | 0.209 | 0.758 | 0.387 | 0.834 | 0.218 |
| ID_37C | 0.872 | 0.299 | 0.784 | -0.099 | 0.834 | 0.143 |
| % of variance | 66.3 | 16.2 | 62.4 | 16.1 | 64.4 | 15.2 |
| Cumulative % of var. | 66.3 | 82.4 | 62.4 | 78.5 | 64.4 | 79.6 |
Legend: ΔpH03, ΔpH06, ΔpH36: pH decreases between t= 0 and 3 h, t= 0 and 6 h, and t= 3 and 6 h, respectively; pH6: pH value after 6 h at 30 °C; ProteolyticAct: diameter of proteolytic activity (mm); LAC: L-lactic acid concentration (g/L). For each pathogen: ID_10C and ID_37C: diameter of inhibition (mm) tested at 10 °C and 37 °C, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



