
Submitted:
17 October 2023
Posted:
17 October 2023
Read the latest preprint version here
Abstract
Clinical work conducted over the last seventeen years at the EEG Institute in Los Angeles and by other neurofeedback providers around the world has demonstrated the utility of extending frequency-based neurofeedback deep into the infra-low frequency (ILF) region, using the method of endogenous neuromodulation described herein. The method is characterized by the absence of any overt reinforcements, which makes it possible to extend the clinical reach to extremely low frequencies. As the training frequency is lowered, the signal becomes more difficult to discriminate, and ultimately it can only be discerned by the brain itself, in the process of endogenous neuromodulation. The method emulates how the brain does skill learning in general: It must observe itself performing the skill, with feedback on its performance.
While the immediate target of ILF neurofeedback is enhanced self-regulatory competence--with symptomatic relief and functional recovery the secondary consequences, progressive lowering of the target frequencies has led to improved outcomes in application to challenging dysfunctions such as episodic suicidality, migraine, seizures, and bipolar mood swings. The work has also yielded insights into how the frequency domain is organized. The training proceeds best at frequencies that are specific to each individual, and these are referred to as optimal response frequencies (ORFs).
These frequencies differ for various placements but stand in two fixed relationships to one another, one that holds over the EEG spectral range, and another that holds over the entire ILF range. Training in the ILF region engages the dynamics of the glial-neuronal networks, which govern tonic, resting state regulation. The collective clinical experience with ILF neuromodulation within a large practitioner network supports the case for making protocol-based, individualized ‘homeodynamic’ regulation a therapeutic priority, particularly for our most impacted clinical populations: addiction, trauma formations, traumatic brain injury, and the dementias. The case is made for further outcome studies and foundational research.
Keywords:
endogenous neuromodulation
; neurofeedback
; Infra-Low frequency
; slow cortical potential
; intrinsic connectivity networks
; infra-slow fluctuations
; EEG biofeedback
; developmental trauma
Introduction
The frequency-based techniques of neuromodulation, in all their variety, have benefited from considerable innovation over the past few years (Evans and Turner, 2017). Nearly all manipulate one or another parameter governing the EEG spectrum with either a reinforcement or stimulation-based technique. There are also methods that modulate the Slow Cortical Potential, both challenge- and stimulation-based, to be discussed further below. We concern ourselves here with the alternative of endogenous neuromodulation, in which the brain engages directly with the spectrally limited EEG or Slow Cortical Potential. More specifically, features are extracted that reflect the mechanisms by which the frequency domain is organized. This more naturalistic way of working with low-frequency signals extends neuromodulation techniques beyond the reach of prescriptive methods such as operant conditioning. It appears to be more effective than prescriptive methods in some generality, in that it merely supports and augments how the brain learns and maintains self-regulation natively. The method offers unique advantages when it is extended deep into the infra-low frequency region, the focus of this paper.
In brief, the method involves having a client watch the time course of the narrow-band filtered Slow Cortical Potential as it unfolds over the course of a 20-to-50-minute session. The signal is derived from a differential montage at selected cortical sites. Since the signal is slowly varying and therefore of limited interest to the trainee, it is typically imbedded in a video game or movie. For example, the signal may be used to modulate the size of the screen display; or it may govern the speed of a rocket. Auditory and tactile feedback complement the visual representation for a more immersive experience. While the trainee is engaged with the movie or the game, the brain tracks the signal of interest. As soon as the brain detects the relevance of the signal to its task of state management, it incorporates the signal into its regulatory schema as an additional control loop. Since there are no cues or stimuli to provoke or direct the brain’s response, the challenge must be entirely self-generated and self-referential. Thus, the response is based on the brain’s ‘detection and correction’ activities, operating on the dynamics of the signal being tracked, rather than on the more conventional algorithmic inputs external to the brain.
To function in real life, the brain must operate on a prediction model. This applies not only to action-perception cycles but to state management as well (Sterling, 2012). Endogenous neuromodulation rests on this operational framework. A plausible model of the process may be delineated as follows: At the outset, the brain detects a correlation between a feature in the sensory signal stream and its internal state, progressively confirming the provisional hypothesis of correlation by virtue of its persistence. The signal is projected forward in time by way of predictive coding, wity successful prediction confirming the correlation. The brain will then act to bring closure between its prediction and the unfolding reality. This constitutes the self-generated challenge. It can be understood as a continuous process of Bayesian inference (Friston, 2010). The operation of all aspects of this continuous process are concurrent rather than sequential, which has been described in terms of circular causality (Freeman, 2006).
Thus, by tracking the surface potential, we have unblinded the brain to a core self-regulatory activity, the dynamic management of tonic cortical activation. We are therefore justified in using language that ‘anthropomorphizes’ the brain, so to speak, because clearly the brain is the agent in this process. Volitional involvement is not required. The method can be used even with infants and with people in diminished states of consciousness.
In the clinical setting, the process becomes an iterative encounter between first- and third-person perspectives, one that allows the clinician to discern the state dependence of dysregulation status. This facilitates the migration to the optimal training parameters in terms of placement and target frequency (Bagdasaryan and Le Van Quyen, 2013). Over the course of sessions, this becomes a scaffolding process in which foundational issues in the hierarchy of regulation are targeted first, setting the stage for addressing subsidiary objectives.
The Historical Context
Before the method is described further, the context for this novel developmental thrust toward low-frequency training is presented. The confluence of three major developments in the 1960’s to 1980’s laid the foundation for the development of ILF neurofeedback. There was the concurrent development 1) of classical biofeedback, with its emphasis on training the regulation of the autonomic nervous system with the aid of measures of peripheral physiology, and 2) of EEG biofeedback (the common terminology at the time) that was mainly promoting brain stability (with a focus on epilepsy) and hyperactivity, with a focus on ADHD. Both objectives were pursued by way of calming motoric excitability. The third major development was the pursuit of Slow Cortical Potential training in Europe during the eighties, again with the objective of calming cortical excitability.
Therapeutic EEG biofeedback was discovered quite accidentally in the late sixties in animal work. Under umbrella of sleep research, cats were operantly conditioned to enhance the incidence of spindle-bursts at the frequency of the sleep spindle, 12-15 Hz. This bursting activity was labeled the sensorimotor rhythm (SMR) (Wyrwicka and Sterman, 1968). Down-training of the bursting activity was demonstrated as well. To prove that operant conditioning had occurred, two groups of cats were trained successively in both directions, in a crossover design. Motoric excitability diminished with the SMR reinforcement training, and sleep parameters were altered, as indexed by sleep spindle density (Sterman et al.,1970). Months later, these same cats were committed to a test of the toxic effects of low levels of monomethylhydrazine, a rocket fuel (Sterman, 1976). This substance depletes GABA stores, which renders cats highly susceptible to seizure. Surprisingly, half of the group demonstrated heightened resistance to seizure onset, and upon inspection it was found that these were the cats that had finished with SMR up-training in the prior experiment. The learned behavior change had not suffered the extinction that commonly attends operant conditioning designs after reinforcement ceases. Here we had an establishing study that quite by happenstance was both blinded and controlled, as well as suitably powered. The researchers had been unaware of the ground-breaking import of the serial experiments that they had just consummated. No respectable scientist would have posited what was found here as a hypothesis a priori.
Human subjects research followed, as interest in the method flourished for some years (Lubar et al, 1981; Lantz and Sterman, 1988). A summary of those studies was published in 2000 (Sterman, 2000). As all subjects in those studies had been treated to medical standards and were medically stable, the further reduction of seizure incidence by some 60% (mean value) renders the method comparable to what can be achieved with a second or third anti-convulsant. Often, medication could be titrated down or even withdrawn entirely. And not infrequently (~5%) the trainees became entirely seizure-free. Nevertheless, Federal funding of this research terminated in 1985, and the method has been kept alive subsequently mainly in clinical work and via self-funded studies. It had been a case of ‘prematurity in science’—a discovery made before its time (Hook, 2002).
The third tributary to the confluence was the work originating in Europe to train the Slow Cortical Potential (SCP) toward diminished cortical excitability by way of a repetitive transient challenge to the trainee to shift the SCP toward increased positivity (Rockstroh et al, 1989; Birbaumer et al, 1990; Birbaumer, 1999). In this learning task, clients are challenged to alter the ambient level of the SCP by several microvolts within nominally eight seconds (Strehl, 2009). For the first time, tonic state regulation—the focus of nearly all of biofeedback—was being trained directly with a neurofeedback technique. This broadened the clinical agenda considerably.
An extensive research history exists for SCP-based neurofeedback, and the clinical footprint largely overlaps with what can be accomplished with frequency-based training in the low beta band. The core claim of moderating cortical excitability has implications for epilepsy (Rockstroh et al, 1993; Kotchoubey et al, 2001), migraines (Siniatchkin et al, 2000), and even schizophrenia (Schneider et al, 1992; Gruzelier et al, 1999). The predominant application has been to the ADHD spectrum, just as in the case of SMR and low beta-band training (Strehl et al, 2006).
In all these applications, the objective is diminished excitability as a learned response, with consequences for tonic state regulation. This objective is explicit in the last of the studies cited above, by Gruzelier et al. Here a shift in the hemispheric asymmetry of negativity (i.e., excitability) is directly targeted by way of the differential signal at {C3 – C4}. The training is challenge-based, within the customary eight-second window. Thus, all the above techniques for training the SCP rely on phasic responding to train tonic activation. Nevertheless, this study can be seen in retrospect as another technological precursor to the technique of endogenous neuromodulation under discussion here.
The entire field of biofeedback collectively made the case for the clinical utility of enhancing autonomic regulation, and of restoring sympathetic/parasympathetic balance. SCP training then enlarged the agenda for tonic state regulation to cover a broader range of state variables, as well as to enhance cerebral stability. And EEG frequency-based training enhanced our capacity to focus on specific functional domains. The stage was thus set for the exploration of the role of infra-low frequencies in state regulation. The transition into ILF NF was more a matter of continuity rather than of discontinuity, however, so our treatment of the topic necessarily takes us back to the origins of therapeutic neurofeedback.
It is helpful, in that regard, to survey much of the spectral range of clinical interest. The results are shown in Figure 1, which presents the frequency-based analysis of a three-hour 19-channel record while the participant is passively watching a movie. We observe a power-law relationship, and in this case a single exponent holds throughout the range of 0.1mHz to 10 Hz. A power-law relationship characterizes dynamically self-organizing systems (Plenz and Niebur, 2014). The observed power-law exponent of ~1.5 is the expected value for self-organizing systems operating at the edge of criticality, conditions under which responsiveness is maximized within the constraints of system stability (Beggs JM and Plenz D, 2003; Kitzbichler MD et al, 2009). In the present instance, this single exponent applies equally to the EEG and SCP regimes. Implicit here is a frequency hierarchy of regulation in which each frequency is responsible for its corresponding niche in the frequency domain, as well as setting the tone, so to speak, for activities organized at higher frequencies. The phase of the low-frequency signal modulates the amplitude of higher bands (He et al, 2010; Buzsáki and Watson, 2013). The relationship holds in the infra-low frequency region (Monto et al, 2008). This constitutes a potent impetus to train tonic state regulation by operating in the infra-low frequency region.
Early Developments in Therapeutic Neurofeedback
Neurofeedback did not evolve in the US in a climate favorable to innovation. After all, it emerged as a clinical discipline well before the concept of brain plasticity was generally accepted, so it was widely dismissed as an expensive placebo (Thibault et al, 2018). In response, clinicians hewed closely to the protocol that had been validated in research by Sterman and Lubar: the operant conditioning of the sensorimotor rhythm (12-15 Hz) and low-beta band (15-18 Hz). Training was typically done on the left sensorimotor strip, at {C3 – T3}, i.e., in bipolar montage. Sterman had carved one road into the jungle, so to speak, and everyone was on it (Sterman and Friar, 1972). In the 1985-to-1988-time frame, our group developed the NeuroCybernetics system as a computerization of Sterman’s laboratory instrument, and it was oriented entirely to this protocol.
For application to ADHD, Lubar moved the training of the SMR band to Cz (in referential montage), following Tansey, where the theta-band excess, the key signature of dysregulation in ADHD, was observed to be at its extremum along the central strip (Tansey, 1985). An inhibit strategy of cueing the brain with respect to episodes of dysregulation was installed by Sterman to assure the integrity of the discrete rewards, and this evolved into a shaping strategy (systematic down-training of excess theta-band activity) at the hands of Joel Lubar (1997). We adopted both changes.
Protocol innovation was first potentiated with the availability of digital EEG analysis to the private practitioner in the early nineties. This permitted targeting dysregulation wherever it was observed in cortex, and that also served more generally to make the climate within the field more open to innovation. The resulting proliferation of methods is encompassed by the term ‘qEEG-based neurofeedback.’ The downside was the elevation of the localization hypothesis of neuropsychology (just as we were entering the realm of modern network theory!), and with it a shift toward single-site training with referential placement. This development also shifted the perspective to a dysfunction-focus, contrary to all the traditions in biofeedback, which focused on functional enhancement. The quantification of EEG behavior brought in train aspirations toward a manualizable, ‘scientific neurofeedback’, with the implicit mandate that matters had to be done in this way.
As the mechanisms-based training of Sterman and Lubar was function-focused, it was protocol-driven, with little or no individualization. Innovation therefore needed to be hypothesis-driven, based on functional neuroanatomy. As theory was in short supply, innovation transpired much more slowly. In our early work with ADHD, we moved from Cz to C4 with SMR-band training in 1992, and reliably established that the two hemispheres needed to be trained differently. For many years, the standard protocol became the combination of C3-beta training (15-18 Hz) with C4-SMR (12-15 Hz) for thousands of clinicians (Kaiser & Othmer, 2000). The right-hemisphere placement had a calming influence, while the left-hemisphere training was more activating. We now had a predictable handle on the regulation of central arousal: lower training frequencies took us to lower arousal levels. The next critical step was a return to bipolar montage for all our protocols. This allowed us to migrate off the sensorimotor strip while keeping one sensor on the familiar sensorimotor or temporal site (C3 or T3). More fundamentally, the training variable once again became the relationship between two sites: we were training connectivity relationships.
The “C3-beta/C4-SMR” protocol found extensive representation in the literature through the work of John Gruzelier and colleagues for over a decade (Egner and Gruzelier, 2001; Gruzelier and Egner, 2005; Gruzelier et al, 2014). The protocol was also the impetus for a large-scale controlled study of neurofeedback-aided addiction treatment (Scott et al, 2005). Also notable was the controlled study by Thomas Fuchs in which the protocol was compared against stimulant medication (Fuchs et al, 2003). The attempt to discriminate between SMR and beta training was published in two papers (Kaiser and Othmer, 1997; Barnea et al, 2004). Our multi-site retrospective appraisal of the protocol in application to ADHD was published in 2000 (Kaiser and Othmer, 2000).
It was mainly families whose children did not do well with medication that were attracted to the training. It was possible to resolve the sleep problems as well as the tics and bruxism that sometimes cropped up with stimulants. Commonly we were engaged with the comorbid disruptive behavior disorders that don’t resolve with stimulants. As these findings were considered anecdotal, they were taken up in a book chapter (Othmer et al, 1999a). The broader reach of the protocol, to the anxiety-depression spectrum, sleep disorders, pain syndromes, brain injury, and the autistic spectrum was likewise consigned to a book chapter (Othmer et al, 1999b).
The singular experience at our clinic of a client becoming manic with SMR-training made it clear that even standard training frequencies entailed significant risks, and that in consequence training frequencies needed to be individualized. The resulting shift in clinical focus to the regulation of central arousal led to the observation that a more optimal response could be obtained over a wider range of training frequencies. This led to the critical observation that potentiated protocol innovation in this approach going forward, the finding that highly dysregulated clients trained best at very specific frequencies. The search for an optimal response frequency (ORF) in each client became the driver that gradually extended the training across the entire EEG spectrum, but mainly moved us progressively lower in the EEG band. The most severely dysregulated clients typically ended up training at the lowest frequency we had available in the software, thus gradually pushing the boundaries ever lower. As practitioners learned how to work with these new lower frequencies and their clientele experienced better outcomes, a corresponding shift occurred in the clientele, in that practitioner started to attract ever more complex presentations.
Working Models
The journey to lower frequency has been underway for a quarter century, with seventeen of those years spent exploring the ILF regime. The centrality of early attachment deficits and developmental trauma in our most challenging clients became apparent, and this has relevance to chronic medical disease as well. Our approach has been organized around several models.
The Trauma Model
“PTSD [Post-Traumatic Stress Disorder] and Traumatic Brain Injury are the leading cause of sustained physical, neurological, cognitive, and behavioral deficits in military personnel and the civilian population.”Mingxiong Huang, PhD (Presentation at Combat Operational Stress Conference, 2010)
The connection between psychophysiological state and chronic medical disease was first brought to scientific notice with the “Adverse Childhood Experiences (ACE) Study” in the late nineties (Felitti et al, 1998). Here the focus was on overt traumatic experiences or contexts of living: psychological, physical, or sexual abuse, etc., during formative years. Evaluation of some 9500 adult questionnaires yielded the finding that “Persons who had experienced four or more categories of childhood exposure had 4- to 12-fold increased health risks for alcoholism, drug abuse, depression, and suicide attempt.” With respect to chronic medical disease, with four or more ACEs the risk of ischemic heart disease, cancer, lung disease, liver disease, and diabetes was found to be elevated by a factor of two.
These findings, although troubling in themselves, under-estimate the medical risk because the most severely impacted individuals most likely had already attritioned out of the pool and were no longer available for the interview. A later study found that those with six or more ACE’s died 20 years earlier than those without (Brown et al, 2009). A prospective study could resolve the issue. As it happens, the Grant study of 256 Harvard students of the late forties and early fifties had already filled that niche. When the 160 remaining participants were re-evaluated in the 1990s, it was found that if the person had grown up in a positive emotional environment, the incidence of the usual diseases of aging was 25%. If they had grown up in an adverse emotional climate, the incidence was 89%. The risk multiplier was an astounding 3.6. In other words, matters did not have to rise to the level of discrete, countable adverse events to create medical risk. The Grant study results informed our thinking from that time forward.
Results were similar for a Scandinavian study that evaluated the somatic health impact of psychological stress. In tracking the increased incidence of disability-related pensions over a five-year period starting at age 65, a dramatic correlation with early psychological stressors was noted. 3-7 stress factors sufficed to yield a doubling of the incidence of disability-related pension over the period. 9-12 stress factors yielded a risk multiplier of four. Overall, “over a quarter of… disability pensions granted for somatic diagnoses could be attributed to psychological distress.” Even more concerning, “…even mild psychological distress was associated with later onset of long-term disability” (Rai et al, 2011).
The implication is that our general health status (mental and physical well-being) is closely correlated with early emotional upbringing. Harry Harlow, the psychologist who conducted the study in which rhesus monkeys were taken from their mothers to be nourished by wire surrogates, encapsulated his results cogently: “Learning to love, like learning to walk and talk, can’t be put off too long without crippling effects.” The infant learns to love by being loved—unconditionally. When that does not occur, or it occurs inconsistently and unreliably, the infant’s orientation to safety and threat is altered. This threat-consciousness is physiologically encoded, with impacts on arousal regulation and the autonomic nervous system (Porges, 2011; van der Kolk, 1994). Tonic regulation of state is fundamentally altered, in consequence of a shift toward over-arousal and even hyper-arousal. Any tendency toward hyper-excitability—genetically rooted or trauma-induced—is exacerbated. Altered tonic regulation of state thus predisposes to the emergence or compounding of cerebral dysregulation.
The Dysregulation Cascade
Consider the finding that even a single change in domicile in a 14-year-old teenager doubles the cumulative risk (to middle age) of attempted suicide, substance abuse, and violent offending. What appears to be a minor traumatic episode may in fact be quite significant in some children’s lives. The only reasonable explanation is that this apparently ‘minor’ trauma has outsize consequences for a subset of the teenage population that is already at risk by virtue of their prior history (Webb et al, 2016).
Traumatic experiences therefore cannot be appraised in isolation. They are connected by what we have termed the dysregulation cascade, in which the thread of continuity is provided by way of neural network—and visceral/autonomic—encoding of the dysfunction. Vulnerability compounds by virtue of repeat exposure before full recovery has been effectuated. A representative model for the dysregulation cascade is the boxing match. Boxing is in essence an assault upon the functional organization of the brain, one that allows little time for recovery between rounds. But substantial recovery typically ensues between bouts, or else the boxer has a short career.
Post-traumatic stress disorder (PTSD) is also subject to the dysregulation cascade. Most cases of Vietnam era PTSD resolved without resort to professional care. (PTSD was not even formally recognized until the eighties.) Those cases that did not resolve were largely refractory to conventional treatment. These cases were likely rooted in early trauma. This pattern became very clear with the application of ILF neurofeedback to combat-related PTSD during the recent wars. The population divided into ‘fast responders’ who recovered quite quickly in the training (twenty sessions or less), a larger category that recovered in forty sessions, and a substantial cohort who failed to recover fully, or required many more sessions, or both. Members of the latter group were much more likely to report early life crises in their history. They are recognized in the European ICD formalism as cases of Complex PTSD (WHO, 2022).
This discussion must also include the topic of minor traumatic brain injury (mTBI) and its chronic sequelae, an issue that has been as much neglected in medicine as emotional trauma. There is nothing minor about the chronicity of mTBI (Herrera-Escobar and Schneider, 2022). The label was introduced in wartime for any head injury that did not involve skull fracture or a penetrating wound. One major thrust in the early days of EEG neurofeedback was working with the persistent deficits of mTBI, which were commonplace in the population. The deficits are largely remediable with neurofeedback, even years after injury. The capacity for recovery is not lost. Left unremediated, however, mTBI is yet another contributor to the dysregulation cascade. Significantly, it is being recognized that a succession of minor brain insults (such as headers in soccer) can eventuate in diagnosable mTBI if the incidence is sustained at a sufficiently high level (Lipton et al, 2013). Head hits are a predominant risk factor as well for Chronic Toxic Encephalopathy (Daneshvar et al, 2023). In every brain at risk, there exists an ongoing contention between such episodic functional decrements and the available resources of recovery.
As in the case of minor brain insults, physical and emotional, there are also cumulative effects from the burdens of low socio-economic status (SES). A recent study quantified the impact. Low SES was shown to involve a hazard ratio of 2 for cerebral hemorrhage, and of about 1.7 for obesity, self-harm, poisoning (i.e., substance abuse), and psychotic disorders. The authors then showed that starting with the psychiatric conditions of self-harm, substance abuse and psychotic disorder a cascade of some sixteen medical conditions of increasing mortality risk could be identified: liver and renal disease, ischemic heart disease, cerebral infarction, chronic obstructive bronchitis, lung cancer, and dementia (Kivimäki et al, 2020). A thread of continuity exists that begins with deficiencies in early attachment or developmental trauma, compounds with adverse life events, and eventuates in chronic medical disease and foreshortened life span. The increased mortality occasioned by poverty has been tracked prospective study that is now in its fifth decade. Recently published results revealed an 80% elevation of mortality risk by age 53 among those raised in poverty and in circumstances of either crowded housing or of parental separation (Yu et al, 2022).
The Causal Chain
That brings us, then, to the question of mechanisms. This topic has been extensively explored by Martin Teicher and colleagues at Harvard. The term ‘maltreatment trauma,’ which includes both overt mistreatment and abject neglect, physical as well as sexual and emotional abuse, is applied to this population. The impact is so substantial that in the characterization of mental disorders, those with a maltreatment history may well constitute a distinct ecophenotype (Teicher and Samson, 2013).
Evidence for altered brain development already exists (Teicher et al, 2016). Of particular interest to us is evidence for altered functional connectivity, as illustrated in Figure 2. Here principal connectivity relationships are discerned with respect to key hubs of interest. In unexposed brains one sees a healthy network under the aegis of the left anterior cingulate, directing a confident interaction with the outside world. By contrast, in the exposed brain the well-elaborated network informs the right anterior insula, which has primary responsibility for securing personal safety, in collaboration with the right precuneus. At the same time, the left pre-frontal circuitry is relatively impoverished in comparison with unexposed individuals. The trauma history has fundamentally altered both how the brain engages with the outside world and how it manages its internal regulatory regime to assure its own safety (Teicher et al, 2014).
The differences shown in Figure 2 lie in the functional domain, and are therefore accessible to a training strategy, at least in principle. However, in both cases we are seeing patterns of connectivity that are congruent with the person’s life experience. What is the impetus for change? We know for example from cases of chronic anxiety that, despite the associated discomforts, the state of anxiety may come to confer a certain sense of security to the patient and therefore may not be so readily relinquished. The same may hold true for over-arousal and hyper-vigilance in a person with combat-related PTSD. These may well continue to be experienced as the life preservers they once were. An effective therapeutic strategy would need to offer the trainee the visceral experience of calmness in a safe context to effect a global physiological ‘reset.’ The approach of endogenous neuromodulation, which goes to the foundations of the regulatory regime that took shape originally in an environment of threat, appears to accomplish this, based on tracked symptom changes over the course of ILF treatment (Othmer et al, 2011; Kirk and Dahl, 2022; Spreyermann, 2022)
Based on the above, we can reasonably argue that mental dysfunctions lie in the causal chain of chronic medical disease, and that early childhood trauma accounts for the relatively intractable portion of the healthcare burden. One might invoke Pareto’s Law on this point, conjecturing that some 75% of medical costs may be attributable to nominally 25% of the population, and that this cohort consists largely of those who endured early childhood trauma or physical brain trauma. Quite literally, there is no health without mental health. It follows that a comprehensive strategy to address the risk of chronic medical illness in a prevention model involves utilizing effective methods of resolving these early emotional or physical brain traumas.
The Regulatory Hierarchy as Therapeutic Hierarchy
Working at extremely low frequencies enables us to engage the foundations of the regulatory hierarchy in three aspects: 1) the developmental hierarchy (arousal and affect regulation); 2) the functional hierarchy (from the more general to the more specific; from the more distributed to the more localized; from the more contextual to the more specifically task-oriented); and 3) the hierarchy in the frequency domain. In the latter, the lower frequencies establish the conditions for the dynamics unfolding at higher frequencies. This hierarchy extends into the gamma range of frequencies, and as low as we can reliably measure into the low-frequency domain. The implication of our clinical success is that the intrinsic connectivity networks are sufficiently plastic so that re-normalization of function can be mediated by way of ILF Neurofeedback, despite their being rooted in early development. The above insights have been consolidated through clinical work over the past sixteen years. The relevant citations are covered below in the section on Results.
The Method
The training strategy involves sequentially addressing the core regulatory domains of 1) cerebral stability, 2) arousal regulation, 3) autonomic regulation, 4) affect regulation, and 5) executive function. A complementary target is the left parietal heteromodal cortex. We review each in turn.
Cerebral Stability
The fundamental burden of any self-organizing control system is the maintenance of unconditional stability, sufficient to sustain basic functionality. Brain instabilities violating this criterion include seizures, migraines, panic attacks, asthmatic episodes, syncope, vertigo, sleep apnea, narcoleptic events, and suicidal episodes. The ability to address this disparate range of instabilities with ILF NF supports the view that it constitutes a comprehensive approach to the improvement of cerebral self-regulatory competence. Since the presence of such instabilities interferes with the subtlety that characterizes good regulation, addressing them must constitute the primary objective of a training strategy toward functional normalization.
The management of seizure susceptibility with neurofeedback has more solid literature support than prevails for any other condition (Sterman and Egner, 2006; Egner and Sterman, 2006). Excellent results have also been reported for the remediation of migraine risk in a clinical cohort that had been treated to medical standards (Walker, 2011). With a quantitative EEG (qEEG )-based protocol, more than 50% of trainees ended up migraine-free for the one-year follow-up, versus none among those who relied on medication alone. Another 40% improved by more than 50% in incidence, versus 9% for the medication-only group. Only 2% failed to improve incidence significantly.
These excellent results notwithstanding, a significant step forward was taken with the realization that the classic brain instabilities tend to respond best to inter-hemispheric placements, as shown in Figure 3a. This discovery was made initially in working with migraines. With lateralized placement, a lateralized migraine may well just escape to the other hemisphere; targeting it there just brings it back. With inter-hemispheric placement, no such escape is available, so the migraine vanishes. Typically, this occurs within session, and often within just a few minutes. In yet other cases, the migraine is set on a path to resolution that takes a little longer. With extended training with the same protocol, migraine incidence typically subsides substantially, paralleling Walker’s findings. An outcome study on migraine headache using this method has just been published (Legarda et al, 2022)
Reliance on inter-hemispheric placement goes back to the early seventies, when Douglas Quirk used SMR reinforcement in bipolar montage at {C3 – C4} with violent offenders at the Ontario Correctional Institute in Canada. In combination with galvanic skin response biofeedback, he was able to reduce recidivism in selected violent offenders by a factor of more than three (from 65% to 20%, as documented in three-year follow-up) with 30 sessions of training. His work was the first application of inter-hemispheric training to a cerebral instability, as well as the first large-scale clinical utilization of NF (Quirk, 1995). Quirk claimed to have worked successfully with more than 2700 inmates.
Once the impact of inter-hemispheric placements was recognized for migraines, it did not take long to establish that the same held true for brain instabilities in considerable generality. A key cerebral vulnerability appears to lie in the coordination between the two hemispheres. This coordination is managed at such a subtle level that a prescriptive training strategy is likely unavailing. Here the approach of endogenous neuromodulation is obligatory—both for its capacity for refined individualization and for its preferential access to the deep infra-low frequency domain. To re-establish control, the brain merely needs information about its own functioning; it is not in need of instruction.
Consider, in this regard, the attempt by Gruzelier et al to shift hemispheric negativity in schizophrenia that was referred to previously (Gruzelier et al, 1999). If this procedure were adopted for bipolar disorder, the brain would be driven into mania in one case, and into depression in the other. The balance point would remain elusive to any prescriptive type of training. With endogenous neuromodulation, the balance point can be located by fine adjustment of the training frequency. Training too high takes the brain in a manic direction; training too low takes it toward the depressive state. This can take place in a matter of minutes in rapid responders. Training at the ORF serves to reinforce the network status that maintains stability against excursions into either mania or depression. In practice, the search of the optimal response frequency (ORF) might take a few sessions.
The process is illustrated in Figure 4. Positive attributes can be combined to yield an Index of Functionality; adverse observables can be combined to yield an Index of Dysregulation. An optimum in both parameters is achievable at the optimal response frequency, and the therapeutic journey typically begins with the search for the ORF, particularly in highly symptomatic, and thus highly reactive/responsive clients.
When the brain is exposed to information relative to its state, just acting upon that information accelerates its journey through state space. The clinician adjusts the available parameters—placement and target frequency—to move the process toward its most propitious outcome. In a highly symptomatic individual, the journey to identify the ORF may well encounter adverse attractors in the attractor landscape (Freeman, 2006), which would call for rapid accommodation on the part of the therapist. There are compensations: the highly symptomatic individual is likely to be very sensitive to parameter change, thus facilitating the optimization procedure by way of good reporting. Conversely, when the trainee is less parametrically sensitive, then training at the ORF is also less critical.
In the search for a client’s optimal training parameters throughout the training process, we are compelled to operate clinically with what is termed “ipsative trend analysis,” i.e., the discernment of change induced via the training process in our clinical observables (Ulrich, 2020). In this process, the client’s report on subjective experience of change is heavily relied upon—if the client is capable of reporting, and particularly if the client is symptomatic. Symptom changes within session are the primary drivers of the optimization procedure. In the absence of symptoms that can be tracked within session, the process relies on state shifts with respect to issues on which the trainee can report: alertness, calmness, emotional ambience, etc. The objective is to reach the state at which the client feels maximally alert, calm, and euthymic. That specifies the conditions under which improved self-regulatory competence is achieved most efficiently in terms of both target frequency and placement. The conventional measures used in biofeedback can also be helpful here: galvanic skin response, finger temperature, heart rate and its variability, as well as scalp muscle tension. Over the course of sessions, additional information is recruited regarding the quality of sleep, and status with respect to drives such a hunger, pain, and cravings, depending on the client’s issues.
The above approach was adopted in the late nineties, when we were still working exclusively in the EEG spectral range. Here the ORF may have to be fine-tuned at the 1 – 3 % level. It may migrate modestly—and slowly—over the course of sessions, or when other protocols are introduced, but we regard it as a basically stable characteristic of a particular nervous system, at least over the time scales relevant in therapy.
Given the centrality of brain stability in neuroregulation, the hypothesis that all our clinical objectives might be met with inter-hemispheric training at various homotopic sites was evaluated in the early 2000s, concurrent with the migration down to the lowest EEG frequencies (Othmer and Othmer, 2005). Three placements turned out to be of greatest utility: {T3 - T4}, {P3 - P4,} and {Fp1 - Fp2}. Frontal placement {F3 - F4} played a role mainly in the anxiety-depression spectrum. Continuous performance test data, which we had been acquiring systematically on our clients since 1990, continued to yield satisfactory results (Putman et al, 2005). Nevertheless, over the course of two years the addition of lateralized placements was indicated, as shown in Figure 3b. All our lateralized montages had either T3 or T4 in common. Very quickly, {T4-P4} and {T4-Fp2} came to dominate in the practice, indicating a right-side priority in the training—a complete reversal from the early days of the field, when the focus of cognitive neuroscience was on left-hemisphere function almost exclusively. At that time, right hemisphere function was still terra incognita.
What has survived the test of time in terms of principal training sites is shown in Figure 3c. Observe that with the addition of lateralized protocols the inter-hemispheric training defaulted largely to {T3 - T4}. This schema turns out to be independent of whether one is training in the EEG band or at infra-low frequencies. The same networks are being targeted in essentially the same manner; we are merely appealing to different regions of the frequency hierarchy. However, the frequency range selected for training turns out to be determinative for outcomes.
Arousal Regulation
Beyond the primary challenge of assuring cerebral stability, the secondary burden of a self-regulatory control system is to maintain the appropriate setpoint of activation for optimal functionality. The foundational variable in this category is central arousal. This concept traces back to the origins of the neurosciences. As far back as 1895, Freud and Breuer asserted that “a certain measure of arousal exists in the conductive pathways of the resting, waking, engagement-capable brain.” Since that time, the term arousal has typically referred to phasic arousal, the response to a challenge, whereas our present usage concerns tonic arousal level, in line with Freud and Breuer. Arousal is treated as a global state variable, one that characterizes brain state as a unitary entity. The term activation then remains available to refer to more localized excitation, either within regions or within functional domains. A contemporary perspective on arousal is given by Ross and Van Bockstaele (2021). The intimate connection of arousal level with infra-slow fluctuations is reinforced in a recent report by Sihn and Kim (2022).
The whole-brain character of arousal level could serve only a heuristic function for Freud and Breuer, and the same holds true now. We don’t have a good measure. Nevertheless, within the frame of the regulatory hierarchy in which we operate, the training of arousal regulation is the second-highest priority after cerebral stability. One placement has dominated in this task throughout much of our history: {T4 - P4}. A profound and global sense of calm—possibly even unique in the trainee’s life experience—can descend upon a person even within a single session with this protocol. In most cases, of course, we labor for such gains over several sessions. Quite commonly, both cerebral stability and arousal regulation must be addressed within the very first session.
Autonomic Regulation
The next priority in the regulation of core states is the autonomic nervous system (ANS). Many instabilities involve autonomic dysregulation as the primary issue: asthma, panic, narcolepsy, sleep apnea, and Postural Orthostatic Tachycardia Syndrome (POTS). Much autonomic dysregulation is involved in migraine, particularly cluster headaches. But we have already noted that all these instabilities respond to essentially the same protocol, {T3 - T4}, so this does not further complicate the agenda. As for the critical issue of the tonic relationship of the sympathetic and parasympathetic arms of the ANS, {T4 - P4} training plays the dual role of calming over-arousal and of promoting parasympathetic activation. Thus, with the two protocols discussed, which jointly serve as our standard starting protocols, the issue of autonomic regulation is well covered. We have here an approach that can be broadly helpful with issues of dysautonomia. (It remains true, however, that the traditional biofeedback techniques retain their utility in the refinement of regulatory competence.)
Affect Regulation
The next protocol priority is affect regulation, for which we draw on right-prefrontal training at {T4 - Fp2}. This is never a starting point, even if affect regulation is the primary clinical objective. The clinician is obligated to ascertain how a client responds to the two starting protocols before venturing to train the affective domain. It is difficult to generalize about this placement because ‘affect regulation’ covers a lot of ground. The new placement is integrated into the training protocol, in which every placement that continues to be useful is retained.
Executive Function
Executive function is targeted with {T3 - Fp1}, and as with affect regulation, a foundation must first be laid with the starting protocols. Training is never begun with this placement, and in many cases optimal training parameters are difficult to establish in the ILF. Practitioners may then resort to legacy protocols in the EEG range.
Left Parietal Heteromodal Cortex
Left parietal training at {T3 - P3} can be helpful in the processing of detail. It is therefore relevant to the resolution of learning disabilities (dyslexia, dyscalculia, and dysgraphia). It is usually the last to be introduced into the training schema, as the groundwork needs to be laid for best outcomes. Collectively, these five constitute the primary protocols utilized in the ILF regime. They are illustrated in Figure 3c. They respect the principal cortical divisions, hemispheric and front-back.
Synchrony Training in the ILF Regime
Paradoxically, the midline has been left out of the picture. There is one additional ILF protocol that must be mentioned. We often find it beneficial—and sometimes essential—to also train the coordination of the frontal and parietal hubs of the Default Mode Network with an ILF Synchrony protocol that promotes comodulation of the key frequencies in the 0.01 to 0.1 Hz spectral range. This calls for midline placement (referential) at frontal and parietal sites, in sum-of-channels configuration, {AFz} + {Pz}, to promote the common-mode component. The target frequency can also be critical in this protocol. Most clients never move beyond these primary placements in their ILF training. A fuller complement of protocols is covered in the Protocol Guide, which is currently in its Seventh Edition (Othmer, 2019).
The Frequency Domain
Extension of Training into the Infra-Low Frequency Domain
The evolution of frequency-based training through its critical transition into the infra-low frequency domain has had little representation in the literature to date. A case study on anxiety and fear in a cancer patient demonstrates that training could be done productively even in the delta and theta bands via endogenous neuromodulation (Benioudakis et al, 2016). A study on Complex Regional Pain Syndrome captured the state of the art just prior to entry into the ILF regime (Jensen MP et at, 2007). The larger scope of NF in application to chronic pain is covered in a book chapter (Othmer and Othmer, 2006).
With availability of suitable software at our clinic in 2006, the training frequency could be extended to a bandpass of 0.1 Hz, down from its prior limit of 1.5 Hz (0-3Hz bandpass) of the NeuroCybernetics system. Within the first six months, some two-thirds of our clients ended up training at that lowest frequency, a compelling argument to extend the frequency range further. That, however, required new instrumentation design (Cygnet, Bee Group, Switzerland,). 10 mHz in corner frequency was reached in 2008 with the new instrument. However, the pattern of cases piling up at the lowest frequency just repeated. Within just six months, the training band was extended down to 1mHz.
Every step down in frequency presented a somewhat greater challenge to signal processing, and a certain novelty to the clinician. Over time, however, the pattern repeated, with some two-thirds training at the lowest frequency, so the range was extended to 0.1mHz in 2010. This once again pushed the limits of the available architecture, which called for yet another upgrade of the signal acquisition system, both hardware and software. This became available in 2013. By 2015, a repeat of the above pattern forced another extension of the range to 10 µHz (microHz). By 2017 we were working down to 1 µHz, and in 2019 the range was extended to 0.1 µHz. At the beginning of 2022, training in the 0.01 µHz range became available—and almost immediately became indispensable.
With every downward step, our clinical reach extended to some people we had not been able to help adequately before. Despite the increasing technical challenges as we went lower, the essential experience of the training—in the perspective of the trainee—was very similar across the entire range. In particular, the response to alteration of the training frequency was just as prompt at low frequencies as at higher ones. The brain is of course keying on the dynamics prevailing at the target frequency; it is not aware of the target frequency itself, which disappears entirely from view. Even signal extraction deep in the ILF spectrum must reflect real-time dynamics or it would be irrelevant to our project. The observable dynamics at issue lie in the spectral regime covered in fMRI measurements, 0.1-200 mHz, and that holds for all target frequencies that lie below 0.1mHz.
Optimal Response Frequency: A Resonance Phenomenon
With each highly responsive client, an optimal response frequency can be found at which the training is clearly ‘better’ than at neighboring frequencies, both above and below. We are still far from an understanding the neurophysiological basis of what we are dealing with here, but the clinical realities are difficult to dispute. This behavior exhibits the characteristic of a resonance phenomenon, as illustrated in Figure 5.
To explain our observations, resonance is best viewed in the complex plane (Feynman, 1972). The real axis yields the magnitude of the response, which peaks sharply at the resonance frequency. It is described by the parameter Q, the ratio of center frequency to width of the curve. Q falls in the range of 10-20, which serves to confirm the resonance model. The imaginary axis yields the phase response. The resulting system response differs above and below the ORF in a phase-sensitive manner. While the phase response is complex and may be difficult for the client to describe, it is generally perceived as adverse in both cases (Othmer, 2009). That further supports the resonance model.
The strong frequency dependence becomes observable by virtue of the dysregulation status of the trainee. The variables at issue here are multiple, consisting of all that are being tracked through the training as indices to the physiological state of each individual. At the resonance frequency, the functional status is optimized, the dysregulation status is minimized (given the prevailing constraints), and the responsivity, the apparent rate of progress in training, appears to be optimized. Clients have sometimes referred to this as the “sweet spot.” The more severe the dysregulation status, the greater is the necessity to train at the ORF. Fortunately, this covaries with the ability to discriminate it, as indicated in Figure 5c.
For example, it was observed early on that a migraine aura that emerged in a particular client with training at 3.9 Hz could be disrupted by moving to 4.0 Hz. However, with return to 3.9 Hz the aura would be rekindled. Movement back to 4.0 would cause the aura to subside once again. This phenomenon was reproducible, all in the course of a single session. Such a sharp frequency dependence is characteristic of the resonance response, in contrast to the gentler dependence on frequency observed elsewhere across the spectrum. This kind of evidence established the case for the resonance model for the ORF originally in the EEG spectral range some 25 years ago.
The stability of the ORF over the course of sessions—it tends to migrate only incrementally and slowly with a given protocol—argues for the hypothesis that the brain organizes the frequency domain around these key frequencies, which are dispersed throughout the ILF and EEG ranges. Indeed, such a frequency hierarchy has already been conjectured (Gollo et al, 2015). Whereas this is a very congenial hypothesis, one is nevertheless struck by just how deep into the ILF these characteristic frequencies go. But then we also know that bipolar and depressive cycles can extend over many months in a quasi-periodic manner, and that could be reflective of one of these deeper rhythms.
The Frequency Rules
The frequency rules that apply to ORFs in lateralized placements are shown in Figure 6 (Othmer et al, 2013; Othmer and Othmer, 2016; Othmer and Othmer, 2017). They divide into a high-frequency and a low-frequency domain and are observed to be consistent within their respective regions. Both a linear and a logarithmic plot illustrate the frequency relationships, which are arithmetic in the EEG range and geometric in the ILF range. The left hemisphere optimizes at 2 Hz higher than the right and optimizes at a factor of two higher in the ILF range. The Delta band is the transition region in the two domains, as the two criteria coincide at a right-hemisphere frequency of 2 Hz and a left-hemisphere frequency of 4 Hz (i.e., 2 x 2 = 2 + 2).
Different rules apply to inter-hemispheric placements. This is easily said, but it was a long process to tease them out. That was not the case for the lateralized placements, where the legacy protocols of “C3-beta and C4-SMR” differed by 3 Hz, and it was merely a matter of systematic trial to establish that the optimal difference was really 2 Hz rather than 3. For the inter-hemispheric placements, it was noted that the cortical resting rhythms differed by nominally 4 Hz between the posterior sensory regime (i.e., 10 Hz) and the central somatomotor regime (~14 Hz). Thus, it was found that posterior placements optimize at 4 Hz lower than central (and a factor of four lower in the ILF), except for posterior temporal sites (T5/T6), which train identically to T3/T4. Frontal placements, in turn, optimize at 2 Hz lower than central placements (a factor of two lower in the ILF). In actual practice, with most clients optimizing at the lowest available frequency in the ILF so much of the time, and with T3-T4 having such a broad clinical footprint, the opportunity to probe other inter-hemispheric placements at yet lower frequencies has been limited until recently.
It is appropriate at this point for the authors to acknowledge that some of what has been presented no doubt strains credulity on first exposure. We have had the benefit of years to do reality-testing, progressing incrementally by way of Bayesian inference, cumulatively confirming/disconfirming provisional hypotheses. The entire construct rests substantially on ‘soft data,’ namely the subjective reporting of highly symptomatic clients with respect to categories that are not readily quantifiable—and inherently variable. Particular observations are often non-repeatable, as the state vector is itinerant. And yet what has emerged is the hardest of testable hypotheses: the frequency rules. They are constantly under test within the global practitioner network. The existence of the frequency rules validates the ORF principle, and the existence of the ORF in turn validates the approach for elucidating it. There is no alternative to the within-subject design for proving out these hypotheses. Every clinical case is its own A/B design, with an ongoing protocol contingency at every juncture.
Mechanisms Implications
The most striking feature of this entire development is the long-term trend toward ever lower target frequencies. This pattern is driven by the issue of arousal regulation. Those whose training begins with the {T4 - P4} placement exclusively tend to train at the lowest frequencies. By contrast, those whose training begins with the inter-hemispheric protocol exclusively have their target frequencies much more broadly distributed. Over the course of training, most clients (>90%) are likely to experience both starting protocols. And it is found that in such cases the inter-hemispheric placement is more tightly constrained in terms of ORF, as is to be expected for cerebral instabilities. In generality, all right-lateralized placements train at the same ORF as inter-hemispheric placement at {T3 - T4}. This fact alone serves to establish the primacy of {T3 - T4} among alternative inter-hemispheric placements.
The implication of the above divergence in spectral distributions is that our two principal failure modes arise from different sources. Arousal dysregulation tends to be the result of environmental insult or persistent duress, whereas brain instabilities tend to have a genetic foundation, one that promotes the vulnerability to hyper-excitability. The training of both failure modes is favored at the ORF, as the frequency domain organization imposes its own constraints.
Another striking consistency is the right-side dominance that has emerged with training in the ILF regime, with the combination of T4-P4 and T4-Fp2. This has all been empirically driven, with the brain effectively in a controlling role with respect to protocol priorities. However, it is consistent with the focus on early developmental priorities. Right-side function is the first to mature in infancy and early childhood (Chiron, 1997; Gao, 2014). More broadly, we observe that the two principal failure modes that have been identified are not just relevant to those afflicted with early trauma, or are otherwise highly deficited, but to our clinical population in general. Our two starting protocols are essentially universally applicable, which implies that we are all subject to the identified key vulnerabilities. Right-hemisphere function is therefore our primary clinical target across almost the entire range of mental-health related dysfunctions that we encounter, and across our entire clinical population, with only rare exceptions, if any.
The Neurofeedback Therapist’s ‘Systems Perspective’ and its Foundations
The ‘systems perspective’, from the standpoint of a neurofeedback therapist, is shown in Figure 7. We assert the primacy of the regulatory arc that begins with interoception, informs autonomic regulation, and affect regulation, and ultimately governs the tonic ambient of central arousal. All these core functions reside in a state of intimate co-regulation. They are under the primary management of the right hemisphere, and this coordination is organized in the ILF regime. Personal history stamps the proceedings via hippocampal, afferent vagal and cranial nerve pathways. The history of traditional biofeedback has long testified to the intimate association of affect regulation with the autonomic nervous system. The latter is trained in order to tame the former, as for example in the management of anxiety.
The foundation for the above conception was laid years ago in the animal work of Nina Aladjalova, whose book became available in the English language in 1964 (Aladjalova, 1964). Aladjalova studied the infra-slow rhythmic potential oscillations (ISPOs) at great length. “A single stimulation of the reticular formation immediately elicits an arousal reaction in the EEG of the cortex, but has no effect on infraslow activity,” she writes. “This reaction is apparently regulated by the rapid regulatory system. Stimulation of the ventromedial part of the hypothalamus…intensifies infraslow cortical activity within 30-40 minutes. This reaction is presumably regulated by the slow regulatory system.”
“…infraslow activity is intensified by certain actions after a long latency period, 30-100 and 120-200 minutes later. We conjectured that this phenomenon reflects the activity of the slow control system of the brain…not only to automatically adjust the system to keeping internal environment constant but actively to establish a new level of activity” (Aladjalova, 1964)
Even in an optimal functioning context, this remains the priority in training. We have been engaged with the slow control system that Aladjalova first identified, and it has been more a matter of homeodynamics than of homeostasis. This system is centrally regulated by the hypothalamus, which governs our internal milieu—autonomic function, sleep-wake cycle, ultradian rhythms, etc. The brainstem is of course central to arousal regulation via the ascending reticular activating system, and as such constitutes the top of the cerebral regulatory hierarchy. Phasic arousal is subject to input from the thalamocortical networks. The tonic level of arousal is in turn informed by a second regulatory hierarchy, under hypothalamic control.
It is appropriate at this point to make the connection, as best we can, between the principal protocols and the targeted core state variables. These protocols were already well-established before our acquaintance with intrinsic connectivity networks (ICNs). Our working model in the late nineties was that we were engaging the multi-modal association areas, the highest level of integration of our sensorium. These are the last areas to mature in neural development, being associated with the highest level of plasticity. According to a more recent study, these regions also exhibit the highest connectivity gradient (Margulies et al, 2014). That is the equivalent of saying that these sites are the most highly integrative in character.
In the perspective of the ICN model, we are training the sites where the Default Mode Network is accessible to us at the cortical surface at lateralized sites. The two criteria are convergent. Further, the multi-modal association areas serve as input to the salience network, so we are training the nexus of the Sensorium, the Default Mode and the Salience network. Our early work in the ILF regime was influenced by Buckner et al. (2008), which examined the connectivity relationships among the hubs of the DMN. We’ve created our own graphical representation of the data presented there, and this is shown in Figure 8. In addition to the general argument that it is most efficient to train the relationships between the hubs, there is the subsidiary argument that one would like to train those linkages that exhibit the highest connectivity. Those network linkages that the brain keeps under the tightest control make for the most discriminating sources of information back to the brain. These considerations further underpin the primacy of T3, T4, P3, and P4 in our protocols. Buckner’s data also supports the case for right-side priority in the training. Observe that the connectivities are generally larger on the right side, and that T4 is more intimately inter-connected than T3. This biases us toward right hemisphere training.
In order to illuminate the relationship between the two hemispheres, we draw on a seminal paper that yields information flow among the principal hubs of the Default Mode Network. These hubs were originally identified in the study of microstates by the Lehman group in Switzerland (Lehmann et al, 1987). Each of the microstates is identified with one of four hubs of the DMN, three in the posterior region and one with an anterior locus. The two principal hubs, anterior and posterior, lie along the midline, and two posterior hubs are lateralized.
Information flow among these hubs has been determined by means of a measure of directed coherence in the alpha and low beta bands, leading to the finding that information flow was dominant from the left hemisphere to the right, as well as from the left to the midline hub, relative to the flows the other way (Lehmann et al, 2014). The imbalance can be substantial. The clear implication is that with respect to the regulatory role of the lower EEG bands, the left hemisphere is in a commanding position with respect to the right hemisphere.
A division of responsibilities is indicated. We may infer from the above that a reciprocal relationship exists in which the right hemisphere bears the primary burden of organizing our resting states, and in that capacity also governs the left hemisphere. The left hemisphere, in turn, supervises our engagement with the outside world, and in that role also governs right hemisphere function. The ILF regime plays a primary role in organizing the resting state configuration, whereas the EEG regime handles the complexity, the coordination, the immediacy, and the temporal precision required for our interface with the outside world. The upper delta band falls in the middle ground, with right hemisphere primacy extending up to 2 Hz, and left hemisphere primacy extending down to 4 Hz, according to the frequency rules.
Vinod Menon proposed the triple network model of psychopathology in 2010 (preprint), postulating that “aberrant organization and functioning of the Central Executive Network, the Salience Network, and the Default Mode Network are prominent features of several major psychiatric and neurological conditions (Menon, 2011). With our limited focus on remediation—as opposed to phenomenology—it now appears that psychopathology is much more explicitly rooted in the Default Mode Network and the Salience Network, and particularly in their tonic regulation within the ILF regime. The Central Executive is essentially missing from the conversation. A right-hemisphere bias has also been identified (Schore, 1997). This, then, defines our agenda with respect to the primary regulatory arc in Figure 7.
Foundational Research
Basic research into ILF neurofeedback via endogenous neuromodulation has taken place to date solely within Russia, where high-level interest in biofeedback has existed for decades. One of the startling clinical observations has been the common experience of robust first-session effects in the ILF training, although single-session effects on excitability have previously been reported in EEG-band training as well (Ros et al, 2010). It was therefore of critical interest to establish whether such first-session effects could be corroborated in physiological measurements. This challenge was undertaken by Dr. Olga Dobrushina of the International Institute of Psychosomatic Health in Moscow, with the collaboration of the Treatment and Rehabilitation Center of the Russian Federation, also located in Moscow.
The specific objective was to identify the networks engaged in the process of covert neurofeedback by distinguishing them from what transpires in sham neurofeedback. 52 healthy volunteers were recruited to a single session of ILF NF under uniform conditions, and resting state fMRI data were acquired immediately prior to and again following the session (Dobrushina et al, 2020). It is important to note that this study was in no way intended to serve as an efficacy study. First of all, efficacy was no longer in question, and secondly, one would not let the question of efficacy turn on a single session of training. The experiment was process-focused more than outcome-focused: What changes are induced by virtue of the act of training on (possibly transient) network organization?
Significant changes were observed in both groups, and there were systematic findings among the members of each group, despite their heterogeneity. In the veridical training group, “increased connectivity was observed through a network consisting of the right and left inferior lateral occipital cortex, right dorsolateral prefrontal cortex and striatum nuclei.” The sham training group also demonstrated systematic change; it differed significantly both from the pre-training condition as well as from the veridical training cohort.
As a corollary finding, this major study contributes to the cumulative body of evidence that sham neurofeedback is not to be considered a neutral control condition in neurofeedback. Under the given blinded conditions, every participant entered the study with the assumption of undergoing an active process. In the search for persistent correlations that will inevitably be mobilized during the session, the brains in the veridical group experience closure and get to settle down to an actual feedback process, whereas the brains in the sham group remain in perpetual search status, one that is never graced with lasting success. This difference is sufficient to yield divergent results in imaging, which testifies both to the salience of the feedback on ILF activity and to the consequential nature of the sham training challenge.
At the Institute of the Human Brain in St. Petersburg, interest in neurofeedback has existed since the nineties. Several studies have been done in recent years to characterize ILF NF. The first of these involved the training of three cases of depression with 20 sessions of ILF NF. Based on three different rating scales, an average improvement of nominally 80% in symptom severity was found, and this finding held up upon one-year follow-up. Substantial changes were also observed in EEG spectral parameters in each of the participants (Grin-Yatsenko et al, 2018).
The second initiative was a controlled study in which ILF NF was compared to Heart Rate Variability (HRV) training as an active control condition (Grin-Yatsenko et al., 2020). The study was performed on a non-clinical population, so the main objective was to track changes in the spectral characteristics in the ILF regime. After completion of 20 training sessions, all nine trainees in the NF group indicated general improvement in their perceived health status—decreased reactivity to stressors; improved body and spatial awareness; and improved stability of mood. They also noticed improvements in energy level and in cognitive performance. In the HRV group, six of eight reported heightened stress tolerance and a greater ability to relax, while the remaining two could not identify any notable change in their state.
The average changes observed in spectral power in the 0.01 to 0.5 Hz band for the two groups were substantial for all standard 19 sites. All 9 trainees in the ILF NF group demonstrated significant increase in spectral power. The outcomes for the HRV group were more variable, but on average showed a decline in spectral power at all sites. The decline did not reach statistical significance. Additionally, rhythmic oscillations in the 0.06 to 0.12 Hz band became more prominent in 8 of 9 NF trainees. Power in that band increased in only 3 of 8 among the HRV controls, with either no change or a decline in the others. Extended spectral analysis of the data has just been published, revealing a systematic increase in a 0.02 Hz region along with the increased rhythmicity in the 0.08 Hz region (Grin-Yatsenko et al., 2023).
Limited though they are, these data are suggestive of the proposition that ILF NF is effecting readily observable—indeed macroscopic—changes in the frequency-based organization within the upper ILF range, the range of interest in fMRI research. Even a modest recruitment of neuronal populations from distributed activity into correlated activity within transient rhythmic oscillations—infra-slow fluctuations—suffices to account for the substantial increase in spectral power. Tendencies toward the expected power-law distribution are also discernible.
The theoretical aspects of neurofeedback have also attracted academic attention (Sitaram et al, 2017). Among the several models for neurofeedback, covert, closed-loop neurofeedback as well as the skill learning model are discussed. The category of covert, closed-loop neurofeedback encompasses endogenous neuromodulation. The feedback is continuous, and the training process lies beneath awareness. The method at issue here can also be modeled as the enhancement of the brain’s core skill of self-regulation. In fact, the method emulates how the brain does skill learning in generality: It must observe itself performing the skill. There must be feedback on its performance.
Results
ILF neurofeedback has seen little representation in the clinical literature to date. There were three early publications in the neurology literature. One paper focused on side effects of anti-epileptic drugs (Legarda et al, 2011). Another focused on the method, with PTSD as a case example (Othmer et al, 2011). A broader perspective on the clinical reach of ILF neurofeedback was offered in 2013 (Othmer et al. 2013). Here the use of the term endogenous neuromodulation to refer to the absence of overt reinforcers was introduced—likely for the first time. A systematic mixed studies review has just been published (Bazzana et al, 2022). It reports on 18 studies of ILF NF and the alternative of reward-based training on Infra-Slow Fluctuations.
With respect to the issue of emotional trauma specifically, early results for combat-related PTSD were reported by Othmer (2009). More recent publications of note include dramatic recoveries from combat-related PTSD and TBI observed in a small pilot study (Carlson and Ross, 2021) and rapid recovery from a case of complex PTSD (Gerge, 2021). Just published are four additional reports. One is a controlled study of 36 patients suffering from an eating disorder and comorbid PTSD (Winkeler et al, 2022). A second documents the impact of ILF NF on children under the care of the State, most of whom have a trauma history (Fleischman, 2022). A third paper reflects on a multi-year history of working with PTSD using ILF NF in combination with conventional therapies (Spreyermann, 2022). A fourth reports on extended ILF NF training of a veteran with PTSD (Kirk and Dahl, 2022). The broader clinical perspective on ILF NF—including PTSD—has been covered in the book titled Restoring the Brain, which is presently in its second edition (Kirk, 2015, 2020).
Since ILF neurofeedback matured entirely in clinical practice, no formal efficacy studies have been published to date. It’s a little late now, and furthermore, conventional group studies are problematic for various reasons. The ‘special competence’ that may be claimed for endogenous neuromodulation in the ILF regime is the exploitation of latent neuronal/glial plasticity in highly impaired systems, particularly those in which the deficits are rooted in early development. Such special competence is difficult to quantify, given the range of organicity prevailing in such highly impaired neural systems, the individualization of training protocols, and the variance contributed by clinician proficiency. Within the domain of clinical practice, ILF neurofeedback should not be researched as a procedure at all, but rather as a therapy—by way of outcome studies. Foundational mechanisms research is different matter.
The immediate target of ILF neurofeedback is enhanced self-regulatory competence (with symptomatic relief and functional recovery the secondary consequence). We draw upon the Continuous Performance Test, a pressured choice-reaction time test, to demonstrate that self-regulatory competence can be systematically enhanced. This test is routinely given to clients seeking ILF neurofeedback. It has been in continuous use in our practice since 1990. Whereas the original purpose was to assess ADHD children, the test is now used to appraise performance relative to our two core concerns, arousal regulation and cerebral stability. One may regard omission errors, for example, as indexing subtle discontinuities of state (“attentional lapses”), one end of the instability continuum. Commission errors, on the other hand, likely have an arousal level dependence.
For the above purposes we rely on the QIKtest (Bee Medic, Switzerland), which slightly adapted the design of the Test of Variables of Attention (T.O.V.A. ®) (Leark et al., 2007) to a handheld device that gave us 0.1 msec timing accuracy. Central features of the TOVA include an invariant inter-stimulus interval of 2 sec to minimize novelty, and a split between a low-demand and high-demand
phase, in which the ratios of target to non-target are 2:7 and 7:2, respectively. The QIKtest arranged for a return to a low-demand phase for a more complete picture. It also facilitated data collection on a central server, which made it possible to derive population-based norms. These are particularly necessary for the discrete errors, where the distributions are distinctly non-Gaussian. Non-parametric norms were first established in 2013 based on some 50,000 records, which yielded more than 500 samples for each of forty age and gender bins. These norms have subsequently been refined.
Cumulative clinical results for the practitioner network at large are evaluated periodically, and the results for errors of commission are shown in Figure 9 for the period 2018-2022. Plotted is the incidence of cases for a given number of errors per test. In the highly deficited region, incidence of cases is reduced by a factor of 2.5 after nominally twenty sessions of training. The plot demonstrates that anyone sufficiently functional to take the test is quite likely also capable of improving their scores with training. For errors of omission, the corresponding improvement factor was 1.9, and for reaction time outliers, it was 1.4. Surveying the reaction time distribution with the advantage of large numbers had made it clear that reaction time outliers had to be treated as a distinct entity, extending beyond the exponential tail of the expected ex-Gaussian distribution (Leth-Steensen et al, 2000).
The data are plotted as cumulative distributions in Figure 10. The change in median score can be expressed in clinically meaningful terms by way of the norming data. The pre-training median value of 10.4 commission errors translates to a normative score for 13.7-year-olds, and 4.8 errors to a normative score for 20-year-olds. Such data are even more meaningful when related to age-segregated cohorts. A convenient division is into a 6-9-year (inclusive), and a 10-19-year cohort.
With respect to impulsivity, the median mental age for the younger cohort is less than six years of age at intake. As norms don’t extend to younger ages, a value of 6 years is assigned. After nominally twenty sessions of training, the median mental age is 10 years. For the older cohort (10-19), the median age is 9 years at intake, and after 20 sessions we obtain a score of 17 years. Observe that after twenty sessions the younger cohort outperforms the older cohort at intake. Results for all four primary measures of the CPT are shown in Table 1. Impulsivity is the quickest to respond, with the median score rising to above normative values. This is followed by inattention (as indexed by omission errors), for which the median reaches normative values. Mean reaction time and variability in reaction time take longer to plateau. Those whose level of function is not normalized in twenty sessions are well advised to continue the training.
Surprisingly, it was observed some years ago at the EEG Institute that outcomes for omission and commission errors were not predictable on the basis of intake data. Shown in Figure 11 are the results obtained on a subset of 87 trainees whose starting standard score was 85 or less (<16th percentile) out of a total cohort of 350 successive data sets. Data are plotted rank-ordered by starting value. When non-responders are excluded from the plot (i.e., those showing less than 5 pt. improvement in standard score), it is found that the likelihood of reaching a score of 100 (i.e., zero omission errors) is independent of the starting value, as shown in split-half comparison—at about 50%. It appears that initial test results tell us very little about the potential for functional recovery. Only the training itself can disclose what is possible. Results for impulsivity show a similar tendency, along with fewer non-responders (5% of those scoring < 85). These data show that remediation of the cardinal symptoms of ADHD are achievable with a primary reliance on the two principal targets—cerebral stability and arousal regulation—that typically dominate in the first 20 sessions of training.
Conclusion
A method is described for the exploitation of endogenous neuromodulation to target the foundations of our regulatory and developmental hierarchies, with a particular view to the remediation of developmental deficits as well as the consequences of early emotional and physical trauma. The objective is to forestall any subsequent dysregulation cascade early in the state of formation, and thus lay the basis for constituting a resilient self. More generally, it is proposed that the foundation of a better-regulated physiology reduces the severity and improves the manageability of mental dysfunctions in considerable generality. For the population at large, this work should be understood in the frame of optimal functioning.
These objectives drive a focus on the infra-low frequency domain, the foundation of the frequency hierarchy, with a particular emphasis on right-hemisphere function and on inter-hemispheric coordination. The training is ideally conducted in observance of the principle of optimal response frequency, and in cognizance of established frequency rules that relate different protocols. The remedy provided by way of endogenous neuromodulation is available at any age.
The substantial functional improvements that are available even for highly deficited individuals, with modest investments of professional time, present us with the ethical mandate to make such training available to children in need early in their development, as well as to adults enduring the handicaps bequeathed to them from their early upbringing.
Author Contributions
Conceptualization, Susan Othmer; Investigation, Susan Othmer; Methodology, Susan Othmer; Writing – original draft, Siegfried Othmer; Writing – review & editing, Siegfried Othmer.
Funding
Funding for the preparation and the publication of the manuscript is provided by the Anthony and Jeanne Pritzker Family Foundation under a grant of funds to the Brian Othmer Foundation.
Institutional Review Board Statement
All of the authors’ findings, as reported here, were acquired in a clinical setting over the course of many years, and none involved formal research studies that called for the involvement of an IRB.
Informed Consent Statement
Informed consent was obtained on all our clients at intake, giving permission for the utilization of their clinical data in an anonymized fashion.
Data Availability Statement
All the data presented here remain accessible on the original server.
Acknowledgments
Since this manuscript recapitulates a 38-year development, it is appropriate to acknowledge the engineers responsible for our three generations of neurofeedback instrumentation: Edward Dillingham (NeuroCybernetics), Howard Lightstone (EEGer), and Bernhard Wandernoth (Cygnet). We acknowledge Marco Versace for his development of the CPT analysis program, EEG Expert, and thank John Putman for his analyses of CPT data. We also gratefully acknowledge funding support from the Anthony and Jeanne Pritzker Family Foundation for this publication project. I thank Martha Herbert for her critical review of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Aladjalova, N.A. (1964). Slow Electrical Processes in the Brain. Elsevier.
- Bagdasaryan, J. and Le Van Quyen, M. (2013). Experiencing your brain: Neurofeedback as a new bridge between neuroscience and phenomenology. Frontiers in Human Neuroscience, 7 (680), 1-10. [CrossRef]
- Barnea, A., Rassis, A., Raz, A., Othmer, S., Zaidel, E. (2004). Effects of Neurofeedback on Hemispheric Attention Networks, Brain and Cognition, (November 2004). [CrossRef]
- Bazzana F, Finzi S, Di Fini G, Veglia F (2022). Infra-Low Frequency Neurofeedback: A systematic mixed studies review. Front. Hum. Neurosci. 16:920659. [CrossRef]
- Beggs, J.M. and Plenz, D. (2003) Neuronal avalanches in neocortical circuits. J Neurosci, 23(35):11167-77. [CrossRef]
- Benioudakis, E.S., Kountzaki, S., Batzou, K., Markogiannaki, K., Seliniotaki, T., Darakis, E., Saridaki, M., Vergoti, A., and Nestoros, J.N. (2016). Can Neurofeedback Decrease Anxiety and Fear in Cancer Patients, Postepy Psychiatrii i Neurologii 25(1), 59-65. [CrossRef]
- Birbaumer, N., Elbert, T., Canavan, A.G.M., Rockstroh, B. (1990) Slow Potentials of the Cerebral Cortex and Behavior, Physiological Reviews, 70(1), 1-41. [CrossRef]
- Birbaumer, N. (1999). Slow cortical potentials: Plasticity, operant control, and behavioral effects. The Neuroscientist, 5, 74–78. [CrossRef]
- Brown, B.B. (1974). New Mind, New Body: Bio-Feedback: New Directions for the Mind, Harper and Row, New York.
- Brown, D.W., Anda, R.F., Tiemeier, H., Felitti, V.J., & Edwards, V.J., Croft, J.B. Giles, W.H. (2009). Adverse Childhood Experiences and the Risk of Premature Mortality, Am. J. Preventive Med. 37(5), 389-396. [CrossRef]
- Buckner, R.L., Andrews-Hanna, J.R., Schacter, D.L. (2008). The brain's default network: Anatomy, function, and relevance to disease. Ann N Y Acad Sci. 1124(1), 1-38. [CrossRef]
- Buzsáki, G. and Watson, B.O. (2012). Brain rhythms and neural syntax: Implications for efficient coding of cognitive content and neuropsychiatric disease. Dialogues in Clinical Neuroscience, 14(4): 345-367. [CrossRef]
- Carlson, J., and Ross, G. W. (2021). Neurofeedback impact on chronic headache, sleep, and attention disorders experienced by veterans with mild traumatic brain injury: A pilot study. Biofeedback 49, 2–9. [CrossRef]
- Chiron, C., Jambaque, I., Nabbout, R., Lounes, R., Syrota, A., & Duklac, O. (1997). The right brain hemisphere is dominant in human infants. Brain, 120, 1057-1065. [CrossRef]
- Daneshvar, D.H., Nair, E.S., Baucom, Z.H. et al. (2023). Leveraging football accelerometer data to quantify associations between repetitive head impacts and chronic traumatic encephalopathy in males. Nat Commun 14, 3470 (2023). [CrossRef]
- Dobrushina, O. R., Vlasova, R. M., Rumshiskaya, A. D., Litvinova, L. D., Mershina, E. A., Sinitsyn, V. E.; et al. (2020b). Modulation of Intrinsic Brain Connectivity by Implicit Electroencephalographic Neurofeedback. Frontiers in Human Neuroscience 14. [CrossRef]
- Egner, T. and Gruzelier, J.H. (2001). Learned self-regulation of EEG frequency components affects attention and event-related brain potentials in humans. Neuroreport 12(18): 4155-4159. [CrossRef]
- Egner, T., Sterman, M.B. (2006). Neurofeedback treatment of epilepsy: From basic rationale to practical application, Expert Rev. Neurotherapeutics 6(2), 247-257. [CrossRef]
- Evans J.R. and Turner R., editors (2017). Rhythmic Stimulation Procedures in Neuromodulation, Elsevier, London.
- Felitti VJ, Anda RJ, Nordenberg D, Williamson DF, Spitz AM, Edwards V, Marks JS (1998) Relationship of childhood abuse and household dysfunction to many of the leading causes of death in adults. The adverse childhood experiences (ACE) study. Am J Prevent Med 14:245-58. [CrossRef]
- Feynman, R.P. (1972). The Feynman Lectures on Physics, Vol. 1, Addison Wesley.
- Internet-accessible: https://www.feynmanlectures.caltech.edu/I_23.html (accessed 12/27/22).
- Fisher, S.F. (2014). Neurofeedback in the Treatment of Developmental Trauma: Calming the Fear-Driven Brain, WW Norton, New York.
- Fleischman M (2022). Documenting the Impact of ILF Neurofeedback on Underserved Populations with Complex Clinical Presentations. Front. Hum. Neurosci. 16:921491. [CrossRef]
- Freeman, W.J. (2006). Definitions of state variables and state space for brain-computer interfaces, Cognitive Neurodynamics 1(1): 3-14. [CrossRef]
- Friston, K.J. (2010). The free-energy principle: A unified brain theory? Nat. Rev. Neurosci., 11, pp. 127-138. [CrossRef]
- Fuchs, T., Birbaumer, N., Lutzenberger, W., Gruzelier, J.H. and Kaiser, J. (2003) Neurofeedback treatment for attention-deficit/hyperactivity disorder in children: A comparison with methylphenidate. Appl. Pychophysiol. Biofeedback, 28: 1–12. [CrossRef]
- Gao, W., Alcauter, S., Smith, J.K., Gilmore, J.H., Lin, W. (2014) Development of human brain cortical network architecture during infancy. Brain Structure and Function, pp. 1-14. [CrossRef]
- Gerge, A. (2020). A multifaceted case-vignette integrating neurofeedback and EMDR in the treatment of complex PTSD. European Journal of Trauma & Dissociation, 4(3), 100157. [CrossRef]
- Gevirtz, R. (2013). The promise of heart rate variability biofeedback: Evidence-based applications. Biofeedback 41, 110–120. [CrossRef]
- Gollo, L.L., Zalesky, A., Hutchinson, R.M., van den Heuvel, M., Breakspear, M. (2015). Dwelling quietly in the rich club: Brain network determinants of slow cortical fluctuations. Phil. Trans. R. Soc. B 370:20140165. [CrossRef]
- Green, E., and Green, A. (1977) Beyond Biofeedback, Delacorte Press/Seymour Lawrence.
- Grin-Yatsenko, V., Othmer, S., Ponomarev, V.A., Evdokimov, S., Konoplev, Y., and Kropotov, J. D. (2018). Infra-Low Frequency Neurofeedback in Depression: Three case studies. NeuroRegulation 5, 30–42. [CrossRef]
- Grin-Yatsenko, V.A., Kara, O., Evdolimov, S.A., Gregory, M., Othmer, S., Kropotov, J.D. (2020), Infra-Low Frequency Neurofeedback Modulates Infra-Slow Oscillations of Brain Potentials: A Controlled Study, J Biomed Eng Res 4(104): 1-10. [CrossRef]
- Grin-Yatsenko, V.A., Ponomarev, V.A., Kropotov, J. D. (2023). Changes of the Infra-Slow EEG Fluctuations of the Brain Potentials under Influence of Infra-Low Frequency Neurofeedback, Journal of Evolutionary Biochemistry and Physiology, 59(3), pp. 831–840. [CrossRef]
- Gruzelier J, Hardman E, Wild J, et al (1999). Learned control of interhemispheric slow potential negativity in schizophrenia. International Journal of Psychophysiology 1999;34:341-8. [CrossRef]
- Gruzelier, J.H., Egner, T. (2005). Critical validation studies of neurofeedback, Child Adolesc Psychiatric Clin N Am 14, 83-104. [CrossRef]
- Gruzelier, J.H., Foks, M., Steffert, T., Chen, M.J.-L, Ros. T (2014). Beneficial outcome from EEG-neurofeedback on creative music performance, attention and well-being in school children, Biological Psychology, 95: 86-95. [CrossRef]
- He, B.J., Zempel, J.M., Snyder, A.Z., and Raichle, M.E. (2010). The temporal structures and functional significance of scale-free brain activity, Neuron, 66(3): 353–369. [CrossRef]
- Herrera-Escobar, J.P. and Schneider, J.C., (2022). From Survival to Survivorship—Framing Traumatic Injury as a Chronic Condition, N Enlg J Med 387: 581-583. [CrossRef]
- Hook, E.B., editor (2002) Prematurity in Scientific Discovery: On Resistance and Neglect, U. California Press.
- Jensen, M. P., Grierson, C., Tracy-Smith, V., Bacigalupi, S. C., & Othmer, S. (2007). Neurofeedback treatment for pain associated with complex regional pain syndrome type I. Journal of Neurotherapy, 11(1), 45-53. [CrossRef]
- Kaiser, D.A. and Othmer, S. (1997) Efficacy of SMR-Beta Neurofeedback on Attentional Processes, Biofeedback and Self-Regulation (4):299-312. [CrossRef]
- Kaiser, D.A., Othmer, S. (2000). Effect of Neurofeedback on Variables of Attention in a Large Multi-Center Trial, Journal of Neurotherapy, 4(1), (2000), pp.5-15. [CrossRef]
- Kirk HW (2015, 2020). Restoring the Brain, Neurofeedback as an Integrative Approach to Health. Taylor and Francis, Boca Raton FL.
- Kirk HW and Dahl MG (2022) Infra Low Frequency Neurofeedback Training for Trauma Recovery: A Case Report. Front. Hum. Neurosci. 16:905823. [CrossRef]
- Kitzbichler, M.G., Smith, M.L., Christiansen, S.R., et al (2009) Broadband Criticality of Human Brain Network Synchronization. PLOS Comput Biol, 5(3):e1000314. [CrossRef]
- Kivimäki M, Batty GD, Pentti J, Shipley MJ, Sipilä PN, Nybert SJ, Suominen SB, Oksanen T, Stenholm S, Virtanen M, Marmot MG, Singh-Manoux A., Brunner EJ , Lindbohm JV, Ferrie JE, Vahtera J (2020). Association between socioeconomic status and the development of mental and physical health conditions in adulthood: A multi-cohort study. The Lancet Public Health 5(3) E140-E149. [CrossRef]
- Kotchoubey B, Strehl U, Uhlmann C, et al (2001). Modification of slow cortical potentials in patients with refractory epilepsy: A controlled outcome study. Epilepsia 42:406-16. [CrossRef]
- Lantz D and Sterman MB (1988). Neuropsychological assessment of subjects with uncontrolled epilepsy: Effects of EEG biofeedback training. Epilepsia 29(2), 163-171. [CrossRef]
- Leark, R. A., Dupuy, M.S., Greenberg, L.M., Corman, C.L. and Kindschi, C.L. (2007). TOVA: Test of Variables of Attention, Professional Guide. Los Alamitos, CA: Universal Attention Disorders Inc.
- Lehmann, D., Ozaki, H., & Pal, I. (1987). EEG alpha map series: Brain micro-states by space-oriented adaptive segmentation. Electroencephalography and Clinical Neurophysiology, 67(3), 271–288. [CrossRef]
- Lehmann, D., Pascual-Marqui, R.D., Milz, P., Kochi, K., Faber, P., Yoshimura, M., Kinoshita, T. (2014). The resting microstate networks (RMN): Cortical distributions, dynamics, and frequency specific information flow. Available from: http://arxiv.org/abs/1411.1949. [CrossRef]
- Legarda, S. B., McMahon, D., Othmer, S., & Othmer, S. (2011). Clinical neurofeedback: Case studies, proposed mechanism, and implications for pediatric neurology practice. Journal of Child Neurology, 26(8), 1045-1051. [CrossRef]
- Leth-Steensen, C., Elbaz, Z.K., and Douglas, V.I. (2000). Mean response times, variability, and skew in the responding of ADHD children: A response time distributional approach, Acta Psychologica, 104, 167-190. [CrossRef]
- Lipton, M. L., Kim, N., Zimmerman, M. E., Kim, M., Stewart, W. F., Branch, C. A., & Lipton, R. B. (2013). Soccer heading is associated with white matter microstructural and cognitive abnormalities. Radiology, 268(3), 850–857. [CrossRef]
- Lubar J. F., Shabsin, H. S., Natelson, S. E., Holder, G. S., Woodson, S. F., Pamplin, W. E., & Krulikowski, D. I. (1981). EEG operant conditioning in intractable epileptics. Archives of Neurology, 38, 700-704. [CrossRef]
- Lubar, J. F. (1997). Neocortical Dynamics: Implications for Understanding the Role of Neurofeedback and Related Techniques for the Enhancement of Attention. Applied Psychophysiology and Biofeedback, 22(2), 111-126. [CrossRef]
- Margulies, D.S., Ghosh, S.S., Goulas, A., Falkiewicz, M., Huntenburg, J.M., Langs, G., Bezgin, G., Eickhoff, S.B., Castellanos, F.X., Petrides, M., Jefferies, E., & Smallwood, J. (2016). Situating the default-mode network along a principal gradient of macroscale cortical organization. Proceedings of the National Academy of Sciences, 113 (44) 12574-12579. [CrossRef]
- Menon, V. (2011). Large-scale brain network and psychopathology: A unifying triple network model. Trends Cogn Sci, 10:483-506. [CrossRef]
- Monto, S., Palva, S., Voipio, J., Palva J.M. (2008). Very Slow EEG Fluctuations Predict the Dynamics of Stimulus Detection and Oscillation Amplitudes in Humans, J. Neuroscience, 28(33): 8268-8272. [CrossRef]
- Othmer, S., Othmer, S.F., and Kaiser, D.A. (1999a). EEG Biofeedback: Training for AD/HD and Related Disruptive Behavior Disorders, In Understanding, Diagnosing, and Treating AD/HD in Children and Adolescents, An Integrative Approach, James A. Incorvaia, Bonnie S. Mark-Goldstein, and Donald Tessmer, editors, Aronson Press, Northvale, NJ, pp.235-296.
- Othmer, S., Othmer, S.F., and Kaiser, D.A. (1999b). EEG Biofeedback: An Emerging Model for Its Global Efficacy, In Introduction to Quantitative EEG and Neurofeedback, James R. Evans and Andrew Abarbanel, editors, Academic Press, San Diego, pp. 243-310.
- Othmer, S., Othmer, S. (2006). Efficacy of neurofeedback for pain management. In: Boswell M, Cole BE, eds. Weiner’s Pain Management: A Practical Guide for Clinicians. 7th ed. CRC Press; 719-739.
- Othmer, S.F. and Othmer, S. (2007) Interhemispheric EEG Training: Clinical Experience and Conceptual Models, Chapter 5 in Handbook of Neurofeedback, Dynamics and Clinical Applications, James R. Evans, Ed., The Haworth Medical Press, New York, p.109-136.
- Othmer, S. (2009). Neuromodulation Technologies: An Attempt at Classification, Chapter 1 in Introduction to QEEG and Neurofeedback: Advanced Theory and Applications (Second Edition), Thomas Budzynski, T., Evans, J.R., and Andrew Abarbanel, A., Eds, Elsevier, pp. 3-26.
- Othmer, S., & Othmer, S. F. (2009). Post-Traumatic Stress Disorder – The Neurofeedback Remedy. Biofeedback, 37(1), 24–31. [CrossRef]
- Othmer, S., Othmer, S., Legarda, S., (2011). Clinical Neurofeedback: Training Brain Behavior, Treatment Strategies - Pediatric Neurology and Psychiatry, 2(1):67-73.
- Othmer, S., Othmer S.F., Kaiser, D.A., and Putman, J. (2013). Endogenous Neuromodulation at Infra-Low Frequencies, Seminars in Pediatric Neurology, 20(4): 246-260 (2013). [CrossRef]
- Othmer, S. and Othmer, S.F. (2016). Infra-Low-Frequency Neurofeedback for Optimum Performance. Biofeedback, 44, 81-89. [CrossRef]
- Othmer, S. and Othmer, S.F (2017). Development of the Othmer Method of Neurofeedback (PDF). Available from: https://www.researchgate.net/publication/317868978_Development_of_the_Othmer_Method_of_Neurofeedback [accessed Dec 26 2022].
- Othmer, S.F. (2019). Protocol Guide for Neurofeedback Clinicians, 7th Edition (2019), EEG Info, Los Angeles.
- Plenz D, and Niebur E (2014). Criticality in Neural Systems, Wiley-VCH, Weinheim, Germany.
- Porges, S.W. (2011). The Polyvagal Theory; Neurophysiological Foundations of Emotions, Attachment, Communication, and Self-Regulation, WW Norton, New York.
- Putman, J.A., Othmer, S.F., Othmer, S., and Pollock, V.E. (2005), TOVA Results Following Inter-Hemispheric Bipolar Training, Journal of Neurotherapy, 9(1), 27-36 (2005). [CrossRef]
- Quirk, D. A. (1995). Composite biofeedback conditioning and dangerous offenders: III. Journal of Neurotherapy. 1 (2), 44-54. [CrossRef]
- Rai, D., Kosidou, K., Lundberg, M., et al (2011). J. Epidemiological Health. [CrossRef]
- Rockstroh, B., Elbert, T., Birbaumer, N., Wolf, P., Duchting-Roth, A., Reker, M.; et al. (1993). Cortical self-regulation in patients with epilepsies. Epilepsy Research, 14, 63–72. [CrossRef]
- Rockstroh B, Elbert T, Canavan AG, Lutzenberger W, Birbaumer N. 1989. Slow Cortical Potentials and Behavior, Second ed. Baltimore: Urban & Schwarzenberg.
- Ros, T., Munneke, M.A.M., Ruge, D., Gruzelier, J.H., Rothwell, J.C. (2010) Endogenous control of waking brain rhythms induces neuroplasticity in humans, European Journal of Neuroscience, 31 (4), 770-778. [CrossRef]
- Ros, T., Moseley, M.J., Bloom, P.A., Benjamin, L., Parkinson, L.A., Gruzelier, J. H. (2009). Optimizing surgical skills with EEG neurofeedback, BMC Neuroscience, 10:87.
- Ross, J.A. and Van Bockstaele, E.J. (2021) The Locus Coeruleus- Norepinephrine System in Stress and Arousal: Unraveling Historical, Current, and Future Perspectives. Front. Psychiatry 11:601519. [CrossRef]
- Schore, A.N. (1997). Early organization of the nonlinear right brain and development of a predisposition to psychiatric disorders. Development and Psychopathology, 9 (1997), 595–631. [CrossRef]
- Schneider, F., Rockstroh, B., Heimann, H., et al (1992): Self-regulation of slow cortical potentials in psychiatric patients: Schizophrenia. Biofeedback & Self-Regulation 17:277-92. [CrossRef]
- Scott, W.C., Kaiser, D.A., Othmer, S., and Sideroff, S.I. (2005). Effects of an EEG Biofeedback Protocol on a Mixed Substance Abusing Population, American Journal of Drug and Alcohol Abuse, 31(3), 455-469. [CrossRef]
- Sihn, D., and Kim S-P. (2022). Brain Infraslow Activity Correlates with Arousal Levels, Front. Neurosci., 16:765585. [CrossRef]
- Siniatchkin, M., Hierundar, A., Kropp, P., Kuhnert, R., Gerber,W. D., & Stephani, U. (2000). Self-regulation of slow cortical potentials in children with migraine: An exploratory study. Applied Psychophysiology & Biofeedback, 25, 13–32. [CrossRef]
- Sitaram, R., Ros, T., Stoeckel, L., Haller, S., Scharnowski, F., Lewis-Peacock, J.; et al. (2017). Closed-loop brain training: The science of neurofeedback. Nature Reviews Neuroscience 18, 86– 100. [CrossRef]
- Spreyermann, R. (2022). Infra Low Frequency Neurofeedback for PTSD: A Therapist's Perspective . Front. Hum. Neurosci. 16:893830. [CrossRef]
- Sterling, P. (2012). Allostasis: A model of predictive regulation. Physiology and Behavior, 106, 5-15. [CrossRef]
- Sterman, M.B., Howe, R.C., and MacDonald, L.R. (1970). Facilitation of spindle-burst sleep by conditioning of electroencephalographic activity while awake. Science, 167: 1146-1148. [CrossRef]
- Sterman, M.B. (1976). Effects of Brain Surgery and EEG Operant Conditioning on Seizure Latency following Monomethylhydrazine Intoxication of the Cat. Experimental Neurology, 50, 757-765. [CrossRef]
- Sterman ,M.B. (2000). Basic concepts and clinical findings in the treatment of seizure disorders with operant conditioning. Clin. Electroenceph. 31(1), 45-55. [CrossRef]
- Sterman, M.B., Egner, T. (2006). Foundation and Practice of Neurofeedback for the Treatment of Epilepsy, Applied Psychophysiology and Biofeedback, 31(1), 21-35. [CrossRef]
- Sterman, M.B. and Friar, L. (1972). Suppression of seizures in an epileptic following sensorimotor EEG feedback training. Electroencephalogr. Clin Neurophysiol. 33(1), 89-95. [CrossRef]
- Strehl, U., Leins, U., Goth, G., Klinger, Ch., Hinterberger, Th., & Birbaumer, N. (2006). Self-regulation of slow cortical potentials—A new treatment for children with ADHD. Pediatrics, 118, 1530–1540. [CrossRef]
- Strehl, U. (2009) Slow Cortical Potentials Neurofeedback, Journal of Neurotherapy: 13(2): 117-126. [CrossRef]
- Tansey, M. A. (1984). EEG sensorimotor rhythm biofeedback training: Some effects on the neurological precursors of learning disabilities. International J of Psychophys 3, 85-99. [CrossRef]
- Teicher, M. H. & Samson, J. A. (2013) Childhood maltreatment and psychopathology: A case for ecophenotypic variants as clinically and neurobiologically distinct subtypes. Am. J. Psychiatry 170, 1114–1133 (2013). [CrossRef]
- Teicher, M. H., Anderson, C. M., Ohashi, K. & Polcari, A. (2014). Childhood maltreatment: Altered network centrality of cingulate, precuneus, temporal pole and insula. Biol. Psychiatry 76, 297–305. [CrossRef]
- Teicher, M.H. Samson, J.A., Anderson, Carl M., Ohashi, K. (2016). The effects of childhood maltreatment on brain structure, function, and connectivity. Nature Reviews/Neuroscience 17, 652-666.
- Thibault, R. T., Lifshitz, M., & Raz, A. (2018). The climate of neurofeedback: Scientific rigour and the perils of ideology. Brain, 141(2), e11–e11. [CrossRef]
- Ulrich, G. (2020). Ipsative Trend Assessment (ITA), An EEG-based Approach for Semi-quantitative Evaluation of Disease Courses, unpublished monograph.
- Van der Kolk, B. (1994) The Body Keeps the Score: Memory and the Evolving Psychobiology of Posttraumatic Stress, Harvard Review of Psychiatry, 1:5, 253-265. [CrossRef]
- Walker, J.E. (2011). QEEG-Guided Neurofeedback for Recurrent Migraine Headaches, Clin EEG and Neuroscience, 42(1), 59-61. [CrossRef]
- Webb, R.T., Pedersen, C.B., Mok, P.L.H. (2016) Adverse Outcomes to Early Middle Age Linked with Childhood Residential Mobility. Am. J. Prev. Med., 51(3), 291-300. [CrossRef]
- Winkeler A, Markus Winkeler M, Hartmut Imgart H (2022). Infra-Low Frequency Neurofeedback in the treatment of patients with chronic eating disorder and comorbid post-traumatic stress disorder . Front. Hum. Neurosci. 16:890682. [CrossRef]
- World Health Organization (2022). ICD 11 for Mortality and Morbidity Statistics: 6B41 Complex posttraumatic stress disorder. Available online at: http://id.who.int/icd/entity/585833559 (accessed August, 2022).
- Wyrwicka, W., and Sterman, M.B. (1968). Instrumental conditioning of sensorimotor EEG spindles in the waking cat. Physiol. Behav. 3: 703-707. [CrossRef]
- Yu, J, Patel, R.A., Haynie, DL, Vidal-Ribas, P., Govender, T., Sundaram, R., and Gilman, S.E. (2022), Adverse Childhood Experiences and premature mortality through mid-adulthood: A five-decade prospective study, Lancet, published online: https://www.thelancet.com/action/showPdf?pii=S2667-193X%2822%2900166-1. [CrossRef]
Figure 1.
Shown is the spectral power (i.e., [voltage]2) over six orders of magnitude in frequency for a 19-site montage. Individual channels are shown in red, and the average value is shown in purple. An eye-ball fit is shown in blue. A power-law relationship is indicated over the entire range. The power-law coefficient is 1.55. The deviation at higher EEG frequencies may well be explained by scalp muscle activity, or by excitement attributable to the movie being watched by the volunteer over the course of three hours. (Data courtesy of Bee Medic, Switzerland.).
Figure 1.
Shown is the spectral power (i.e., [voltage]2) over six orders of magnitude in frequency for a 19-site montage. Individual channels are shown in red, and the average value is shown in purple. An eye-ball fit is shown in blue. A power-law relationship is indicated over the entire range. The power-law coefficient is 1.55. The deviation at higher EEG frequencies may well be explained by scalp muscle activity, or by excitement attributable to the movie being watched by the volunteer over the course of three hours. (Data courtesy of Bee Medic, Switzerland.).
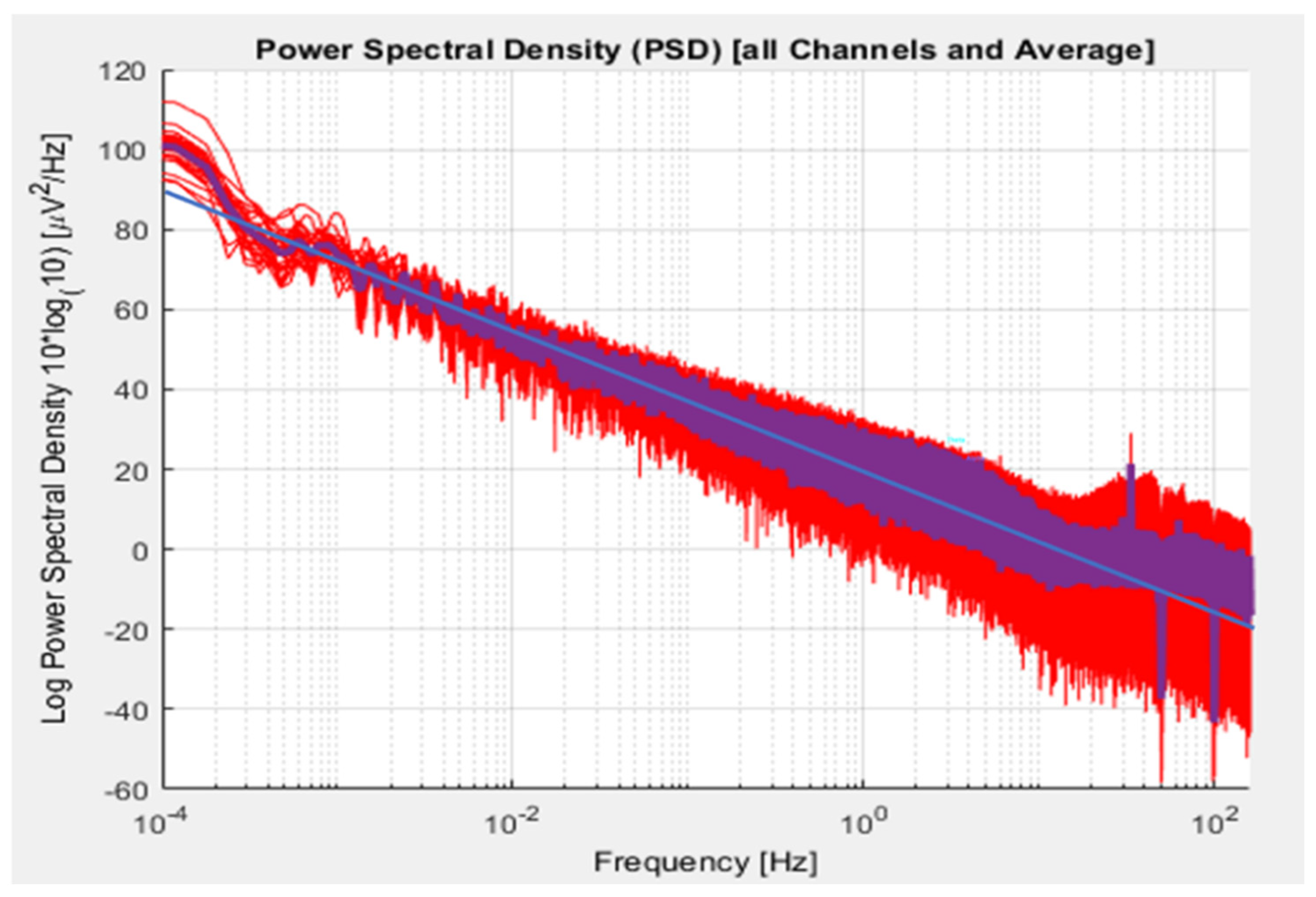
Figure 2.
Network differences are identified for maltreated versus unexposed young adults. The circled dots indicate key regions of interest: the left anterior cingulate cortex, the right anterior insula, and the right precuneus. Connectivity analysis yielded the primary linked nodes that were found to be common within each cohort. These are nodes with direct connections to the region of interest Jointly these linkages determine the relative importance of the three regions of interest for the two cohorts. (Source: Teicher et al, 2016; Original source: Teicher et al, 2014.).
Figure 2.
Network differences are identified for maltreated versus unexposed young adults. The circled dots indicate key regions of interest: the left anterior cingulate cortex, the right anterior insula, and the right precuneus. Connectivity analysis yielded the primary linked nodes that were found to be common within each cohort. These are nodes with direct connections to the region of interest Jointly these linkages determine the relative importance of the three regions of interest for the two cohorts. (Source: Teicher et al, 2016; Original source: Teicher et al, 2014.).
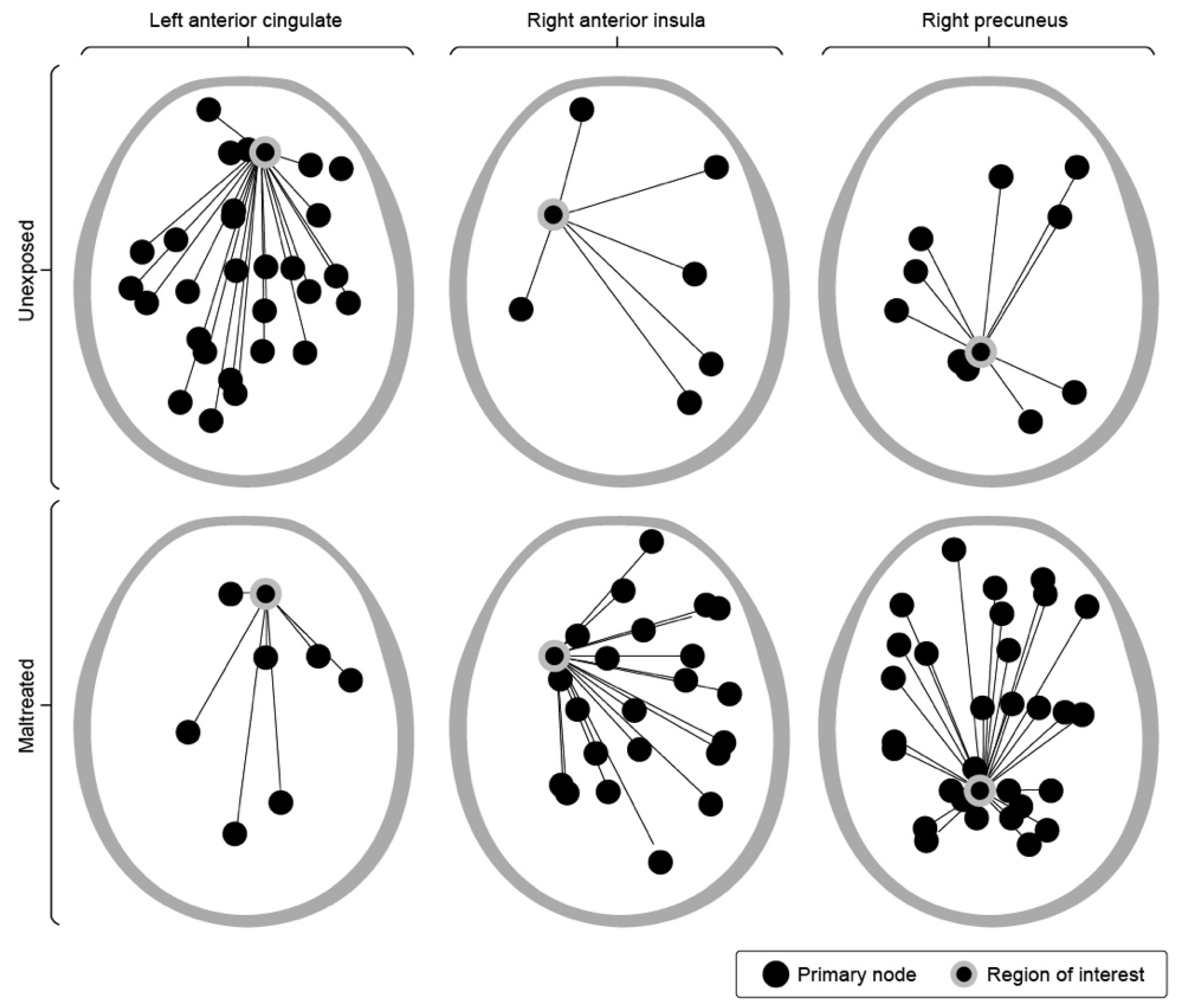
Figure 3.
Principal protocols explored over the course of development. 2a) Inter-hemispheric placements at homotopic sites; 2b) Lateralized placements with T3 or T4 in common; 2c) Principal protocols employed in ILF NF.
Figure 3.
Principal protocols explored over the course of development. 2a) Inter-hemispheric placements at homotopic sites; 2b) Lateralized placements with T3 or T4 in common; 2c) Principal protocols employed in ILF NF.
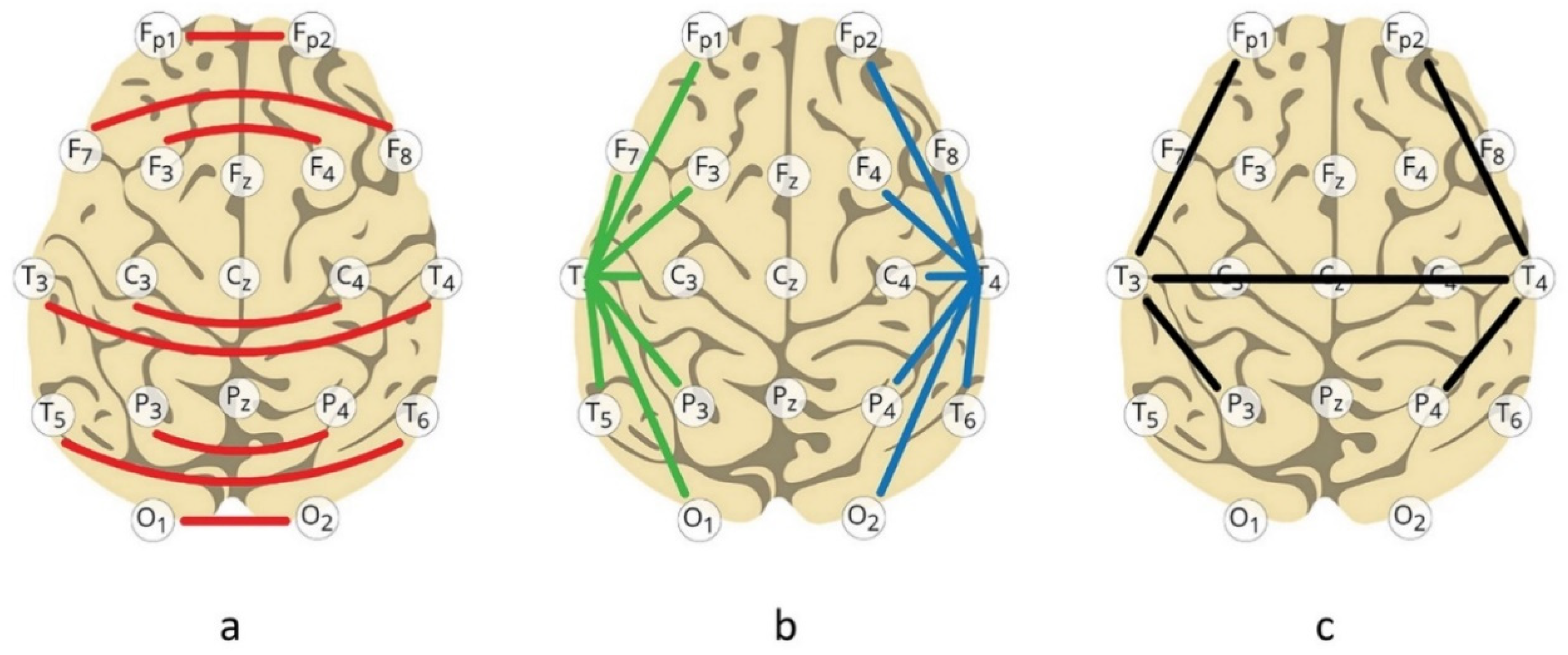
Figure 4.
Both positive and negative features contribute to identifying the Optimal Response Frequency in a trainee. These can be grouped into an Index of Dysregulation and an Index of Functionality. Typically, the process is less formalized: a fluid and dynamic interactive process that moves the client over the behavior surface, gradually converging on the ORF.
Figure 4.
Both positive and negative features contribute to identifying the Optimal Response Frequency in a trainee. These can be grouped into an Index of Dysregulation and an Index of Functionality. Typically, the process is less formalized: a fluid and dynamic interactive process that moves the client over the behavior surface, gradually converging on the ORF.
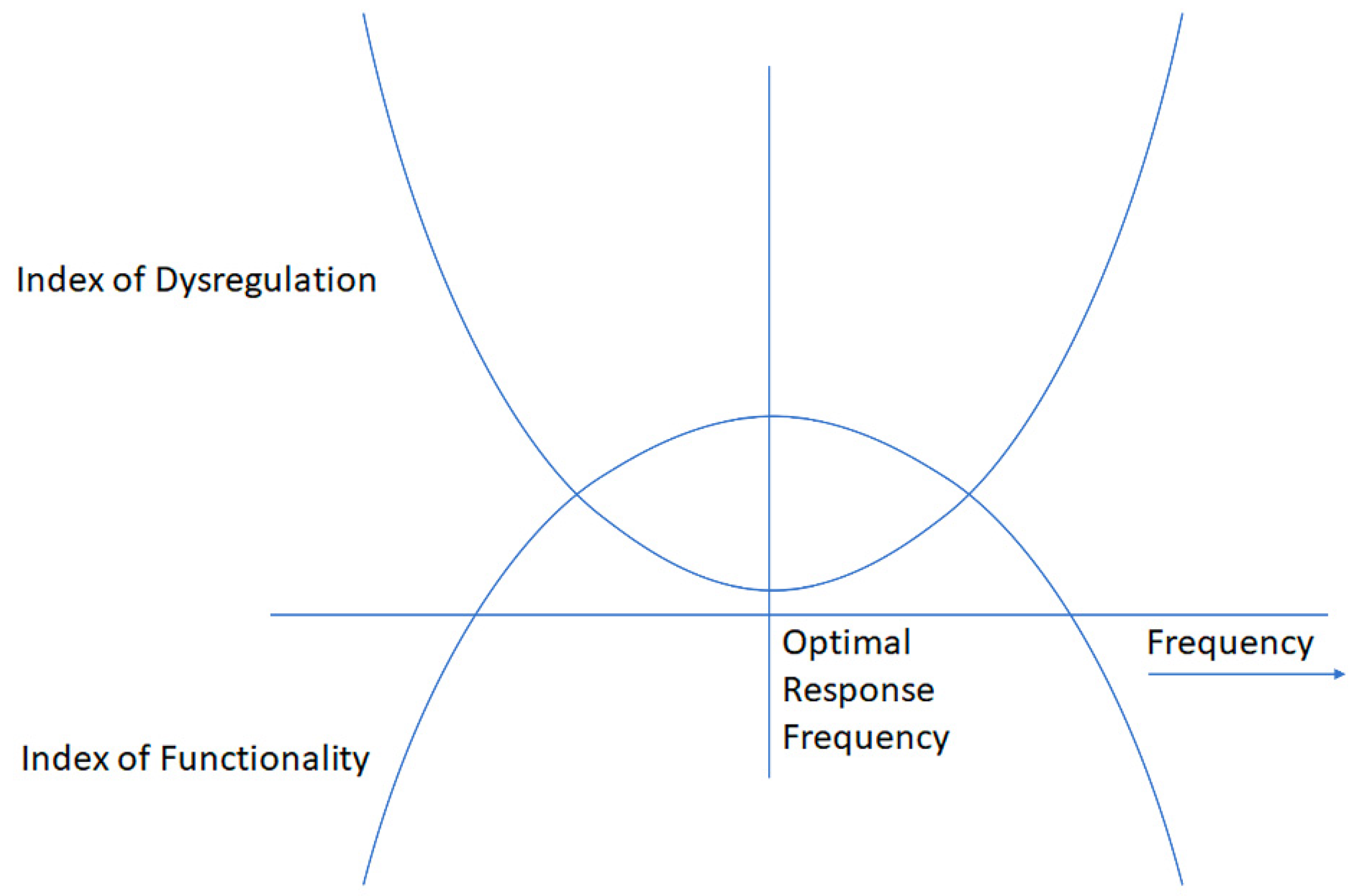
Figure 5.
The physiological response of a dysregulated system near the optimal response frequency (ORF) is illustrated. a) The magnitude of the response, the responsivity, manifests as a narrow peak at the ORF with respect to neighboring frequencies, the core characteristic of resonance. b) The phase response of a resonant system is illustrated for reference (A). The physiological response exhibits a phase-sensitive aspect that differs above and below the resonance frequency, and is restricted to the immediate neighborhood of the ORF, where phase depends strongly on frequency, as shown in (B). c) The net physiological response as training frequency is migrated over the narrow range is illustrated. The optimum response is bracketed above and below by adverse tendencies. Shown are three levels of responsivity (positive) and reactivity (negative), which correlate with dysregulation status.
Figure 5.
The physiological response of a dysregulated system near the optimal response frequency (ORF) is illustrated. a) The magnitude of the response, the responsivity, manifests as a narrow peak at the ORF with respect to neighboring frequencies, the core characteristic of resonance. b) The phase response of a resonant system is illustrated for reference (A). The physiological response exhibits a phase-sensitive aspect that differs above and below the resonance frequency, and is restricted to the immediate neighborhood of the ORF, where phase depends strongly on frequency, as shown in (B). c) The net physiological response as training frequency is migrated over the narrow range is illustrated. The optimum response is bracketed above and below by adverse tendencies. Shown are three levels of responsivity (positive) and reactivity (negative), which correlate with dysregulation status.
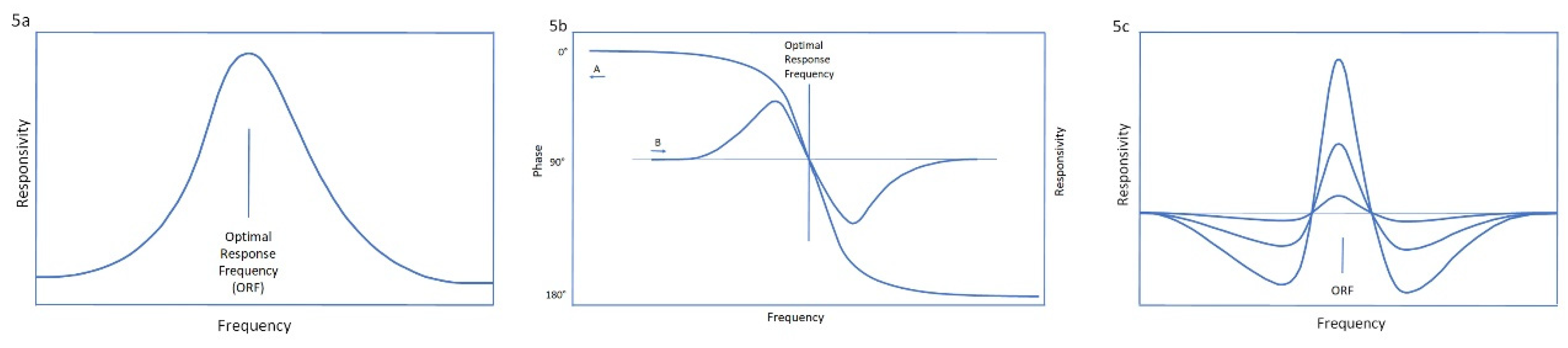
Figure 6.
The frequency rules that apply to ORFs in lateralized placements are illustrated. They divide between the EEG range and the ILF but remain consistent within each. The transition region is the upper delta band. The relationship is arithmetic in the EEG range, and geometric in the ILF—as indeed must be the case.
Figure 6.
The frequency rules that apply to ORFs in lateralized placements are illustrated. They divide between the EEG range and the ILF but remain consistent within each. The transition region is the upper delta band. The relationship is arithmetic in the EEG range, and geometric in the ILF—as indeed must be the case.
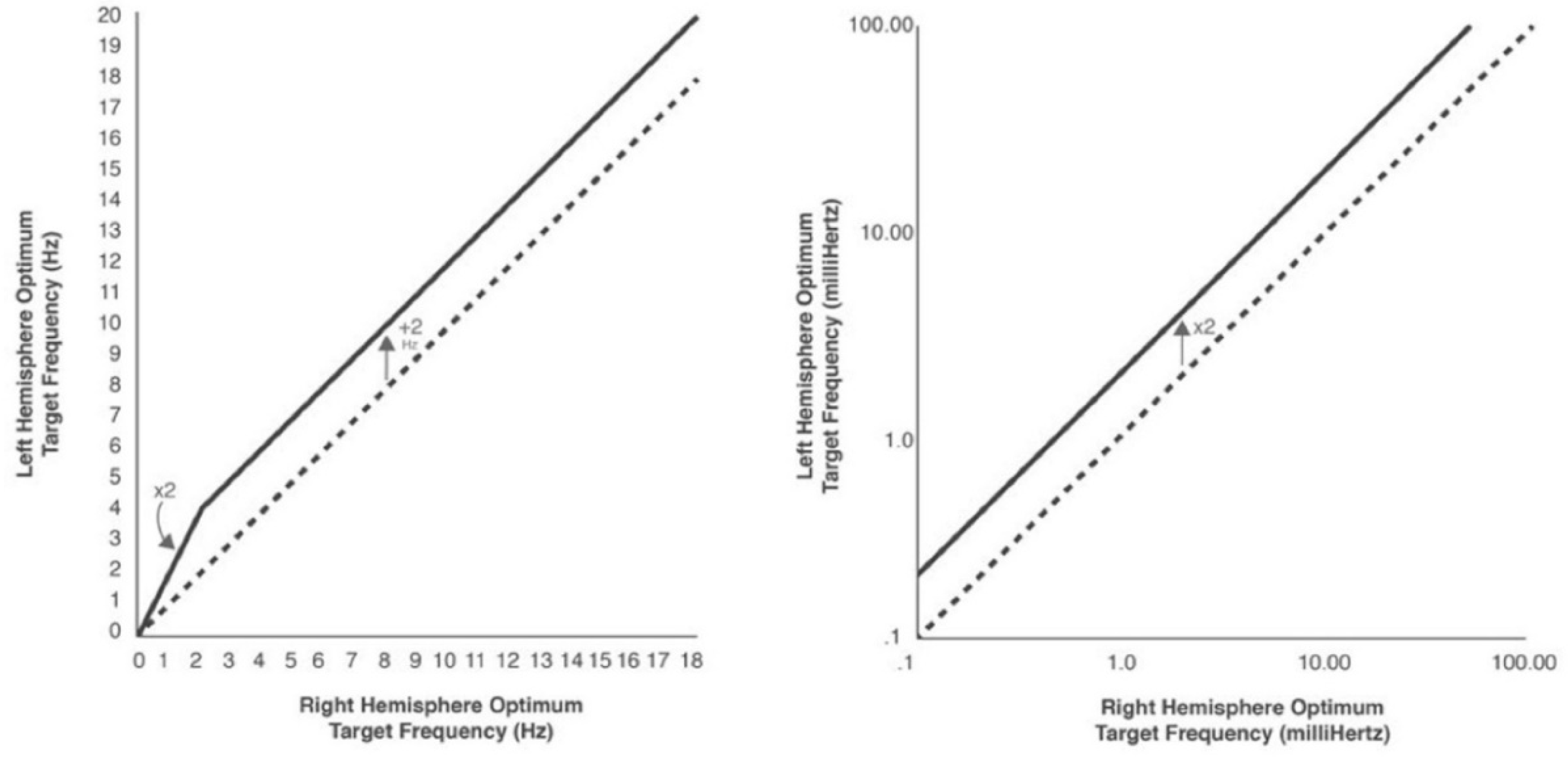
Figure 7.
The systems perspective of the ILF neurofeedback practitioner is illustrated. The therapeutic priority lies with the regulatory arc of interoception, autonomic regulation, affect regulation, and tonic central arousal. .
Figure 7.
The systems perspective of the ILF neurofeedback practitioner is illustrated. The therapeutic priority lies with the regulatory arc of interoception, autonomic regulation, affect regulation, and tonic central arousal. .
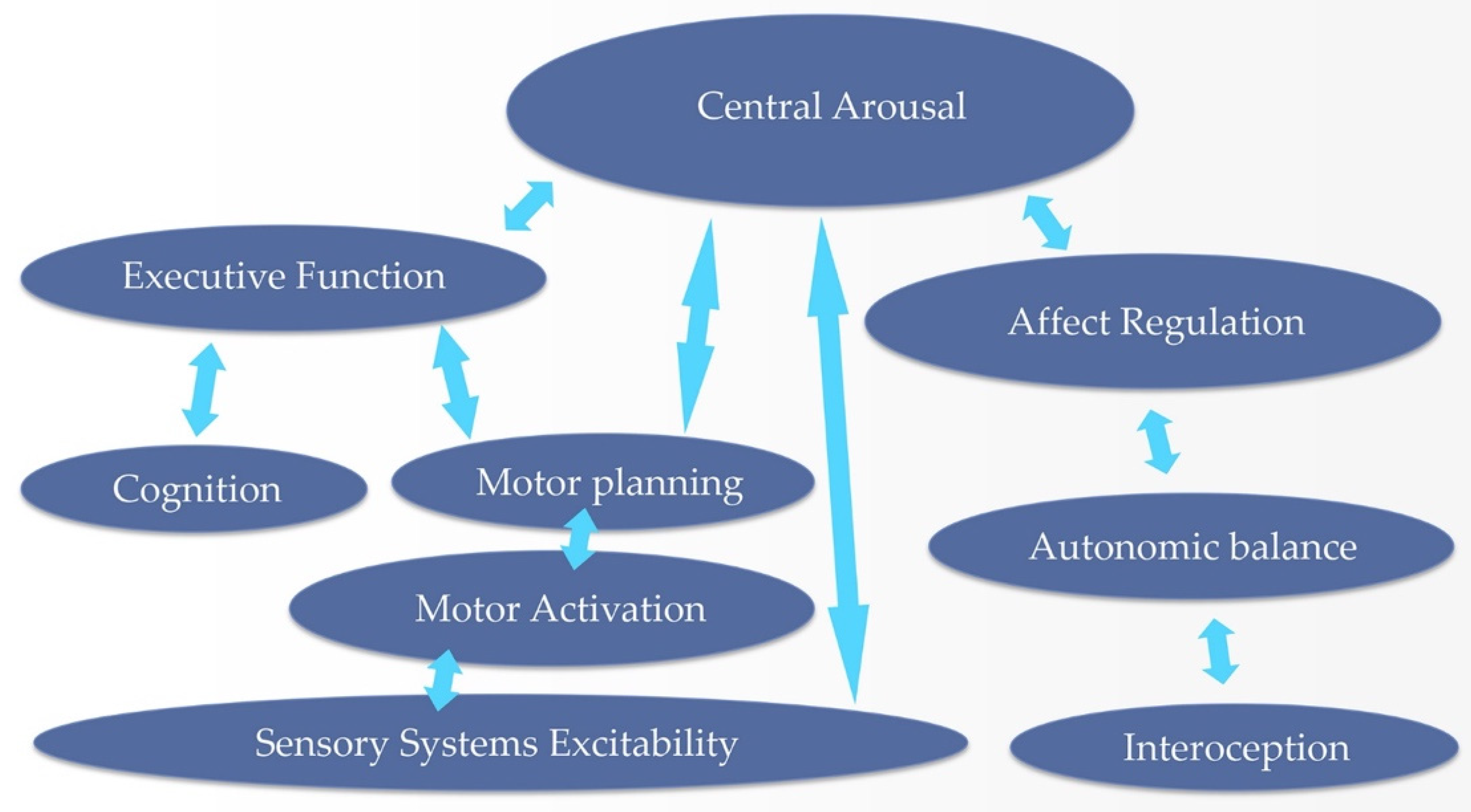
Figure 8.
Connectivity relationships are indicated for the hubs of the Default Mode Network that are directly relevant to neurofeedback protocol decision-making (according to Buckner et al, 2008). A right-hemisphere bias toward greater connectivities is observed. LTC: Lateral Temporal Cortex (T3 and T4); PCC: Posterior Cingulate Cortex; vMPFC: ventromedial pre-frontal cortex; dMPFC: dorsomedial pre-frontal cortex; IPL: Inferior Parietal Lobule (P3 and P4). .
Figure 8.
Connectivity relationships are indicated for the hubs of the Default Mode Network that are directly relevant to neurofeedback protocol decision-making (according to Buckner et al, 2008). A right-hemisphere bias toward greater connectivities is observed. LTC: Lateral Temporal Cortex (T3 and T4); PCC: Posterior Cingulate Cortex; vMPFC: ventromedial pre-frontal cortex; dMPFC: dorsomedial pre-frontal cortex; IPL: Inferior Parietal Lobule (P3 and P4). .
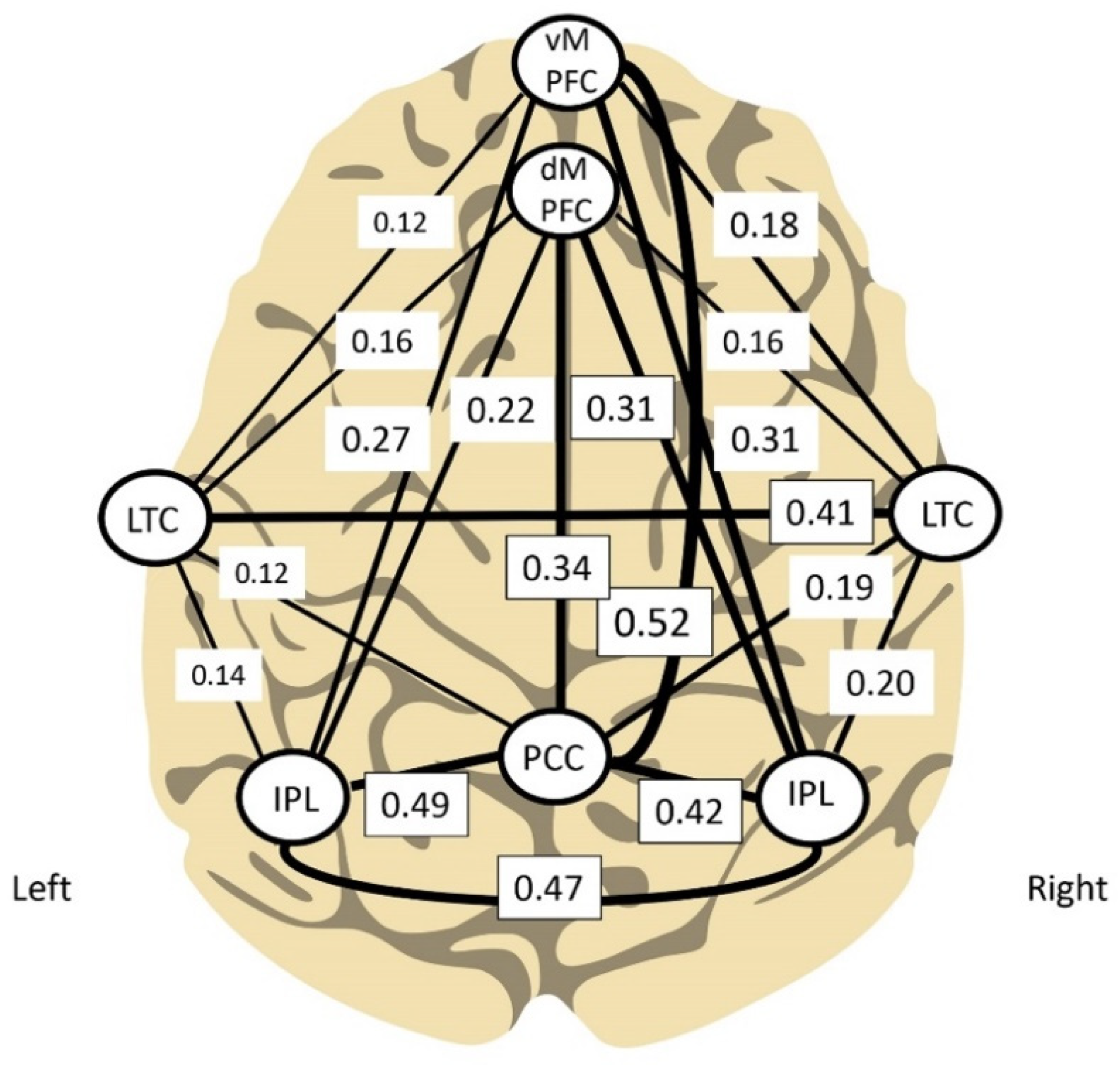
Figure 9.
Pre-post distributions of commission errors are shown here for a non-selective sample of 15,260 cases (data from 2018-2022). Incidence is plotted for the entire range of number of errors per test, out to the pathological extreme of 200 errors. Because of scatter, the data have been smoothed to reveal the underlying trends, and the superimposed curves were by visual inspection. With nominally twenty sessions of training, a reduction in incidence by a factor of 2.5 is indicated for the highly deficited region.
Figure 9.
Pre-post distributions of commission errors are shown here for a non-selective sample of 15,260 cases (data from 2018-2022). Incidence is plotted for the entire range of number of errors per test, out to the pathological extreme of 200 errors. Because of scatter, the data have been smoothed to reveal the underlying trends, and the superimposed curves were by visual inspection. With nominally twenty sessions of training, a reduction in incidence by a factor of 2.5 is indicated for the highly deficited region.

Figure 10.
Cumulative plot of commission error incidence from Figure 9. The median score before training is 10.4 errors, versus 4.8 errors after nominally 20 sessions. Based on our norming data, this translates to an improvement in effective mental maturity from 13.7 to 20 years.
Figure 10.
Cumulative plot of commission error incidence from Figure 9. The median score before training is 10.4 errors, versus 4.8 errors after nominally 20 sessions. Based on our norming data, this translates to an improvement in effective mental maturity from 13.7 to 20 years.
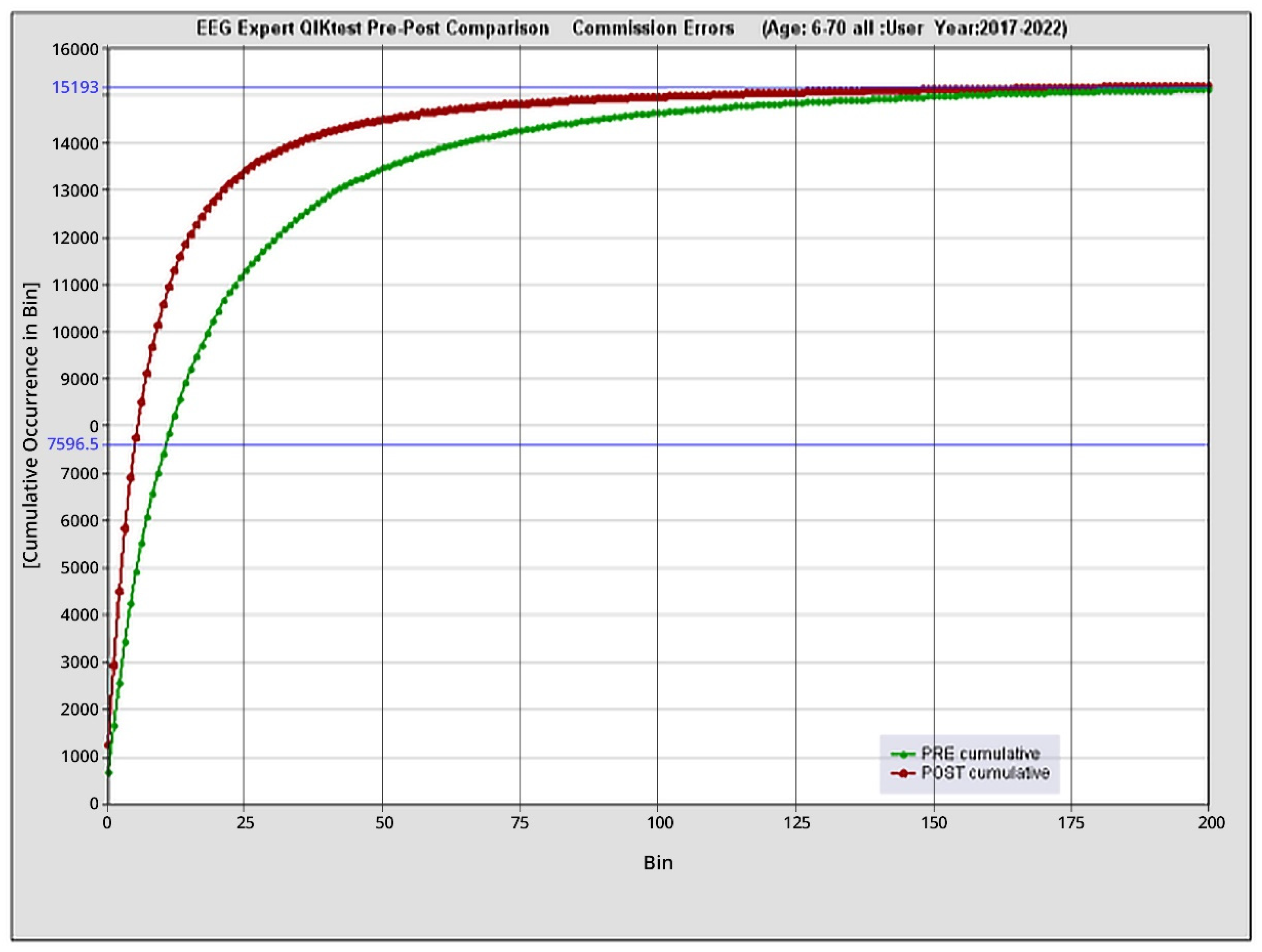
Figure 11.
Pre- and post-data are shown for omission errors for the 87 clients scoring 85 or less in standard score at intake, out of a sample of 350 successive cases seen at the EEG Institute in Los Angeles. The likelihood of scoring zero omission errors (i.e., standard score of 100) after nominally 20 sessions of training is independent of starting value. Non-responders were removed for clarity, as they tend to cluster at the bottom. They numbered 27 in the cohort of 114 scoring < 85 (i.e., 24%, amounting to 8% of the total sample of 350).
Figure 11.
Pre- and post-data are shown for omission errors for the 87 clients scoring 85 or less in standard score at intake, out of a sample of 350 successive cases seen at the EEG Institute in Los Angeles. The likelihood of scoring zero omission errors (i.e., standard score of 100) after nominally 20 sessions of training is independent of starting value. Non-responders were removed for clarity, as they tend to cluster at the bottom. They numbered 27 in the cohort of 114 scoring < 85 (i.e., 24%, amounting to 8% of the total sample of 350).
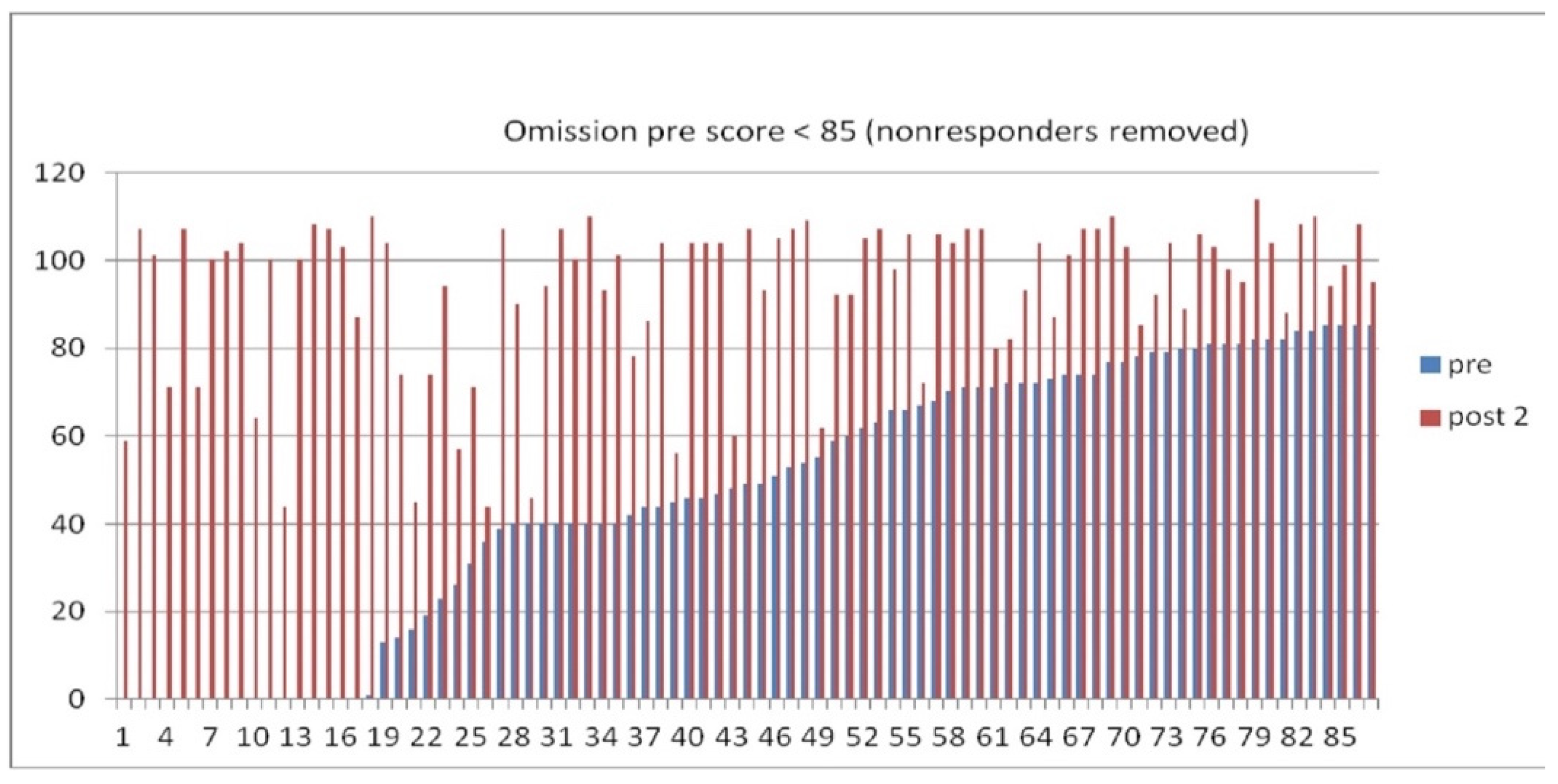

Table 1.
Changes in median mental age with nominally twenty sessions of ILF NF are shown for our two age ranges. Data refer to the entire practitioner network that contributes to the database.
Table 1.
Changes in median mental age with nominally twenty sessions of ILF NF are shown for our two age ranges. Data refer to the entire practitioner network that contributes to the database.
| Age Range | Category | Pre- | Post | Delta |
| 6-9 years, inclusive | Impulse control | 6 | 10 | 4.0 |
| Inattention | 6.2 | 8.5 | 2.3 | |
| Mean reaction time | 7.3 | 8.0 | 0.7 | |
| Variability in reaction time | 6.6 | 7.6 | 1.0 | |
| 10-19 years, inclusive | Impulse control | 9.0 | 17.0 | 8.0 |
| Inattention | 10.3 | 13.8 | 3.5 | |
| Mean reaction time | 12.1 | 12.6 | 0.5 | |
| Variability in reaction time | 10.4 | 11.8 | 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



