
Submitted:
16 October 2023
Posted:
19 October 2023
You are already at the latest version
Abstract
The analysis of the land use dynamics of the Lac Télé Community Reserve (RCLT) using Landsat Thematic Mapper (TM) and (Enhance Thematic Mapper) ETM+ images highlight significant changes in the vegetation cover from 1980 to 2000 and 2018. Thus, the rate of forest area decreased by 21.41% for the entire LTCR in favor of savannahs which increased by 15.23%. The conversion of this forest area to savannah due to the practice of slash and burn agriculture facilitates the opening up of the forest area and contributes to greatly degrading the spawning grounds of fish species from the Likouala aux herbes river. For the mapping of fishing activity in general and the ecological characterization of the 151 identified spawning grounds in particular; the respective mean values of the physical and chemical water parameters; temperature (28.13°C), pH (4.23) and depth (3.34) did not vary significantly from one selected village to another between July and September 2019. The fish diversity unregistered during the study, in the 07 pilot villages would be due to the diversity of the microhabitats noted in the villages of the LTCR, especially from the villages; Botongo, Mossengue and Bouanela where the indices of ichthyological diversity were the highest.
Keywords:
remote sensing
; Geographic Information Systems (GIS)
; fish biodiversity
1. Introduction
The Lac Télé Community Reserve (RCLT) with coordinates (0030'-01058'N and 016050-17030'E) is a category VI protected area according to the IUCN classification, whose main manager is the Congolese State. through the Congolese Agency for Wildlife and Protected Areas (ACFAP) which benefits, thanks to the RCLT Support Project installed since 2001, from the support of the American NGO Wildlife Conservation Society (WCS) in the management of the protected area. The RCLT has an area of 438,960 ha. It is located in the department of Likouala whose capital is the city of Impfondo with two districts including the district of Epéna located 85 km from Impfondo and the district of Bouanela as administrative subdivisions. The vegetation of the RCLT is part of the Congolese natural region called lower Sangha, Cameroonian-Congolese sector or the Guinean-Congolese region [6]). In terms of vegetation, the reserve contains a wide diversity of habitats, including swamp floodplain forests which make up 49%; the forests liable to flooding during the rainy season which occupy 17% of the reserve, made up mostly of gallery or riparian forests located along the watercourses of the LTCR; the mixed forests on dry land which occupy about 10% and finally the savannahs which represent about 16% of the surface of the reserve [6]. This diversity of habitats plays a very large role in the abundance, distribution and diversity of fauna in general [1] and fishery resources in particular, which constitute the bulk of the protein supply. animal populations living near the LTCR [5]. However, like fishing catches at the global level, those also recorded within the LTCR are decreasing [5]. What would be the cause of the degradation of fishery resources within the RCLT, particularly in the Likouala aux herbes river s? Is it the degradation of the environment around the watershed of the Likouala aux herbes river over time, including climate change, which is the cause of the fall in catches? Is it the overexploitation of fish stocks resulting from an uncontrolled intensification of catches that would be responsible for the drop-in catches? Or is it the reduction and exploitation of spawning grounds ensuring the restoration and recruitment within the fisheries of the Likouala aux herbes river, which would be the cause of the degradation of fishery resources? The overall objective of this study is to contribute to the sustainable management of fisheries resources within the LTCR. More specifically, through an analysis of the spatio-temporal dynamics of land use in the watershed of the Likouala aux herbes river, we proceeded to a mapping of the spawning grounds of the Likouala aux herbes river in sight, if necessary, to protect them for the future through the conservation.
2. Materials and Methods
According to the environmental context of the RCLT, especially the distances between the 07 pilot villages, 21 sampling stations were established in the villages of the study, at the rate of 3 stations per village.
- -
- 1st station: Upstream of the village (Northern limit with the spawning grounds) (i)
- -
- 2nd station: Village area (near the main port of the village) (ii)
- -
- 3rd station: Downstream of the village (southern limit with spawning grounds) (iii)
The sampling unit being the village, the three values recorded in each locality were cumulated into an average which served as a reference value with regard to the physico-chemical characterization (water height (m), width (m ), speed (m/s), flow (m3/s), transparency (m), color, temperature (°C), pH, conductivity at 25°C, nature of the substrate and the occupation of the ground of the station ) of the waters of each pilot village. These data were recorded monthly throughout the study period according to the recommendations of Zebaze [13] who confirm and conclude that this frequency accounts for the functioning of hydrosystems. With regard to data on physico-chemical parameters namely; temperature, pH and conductivity at 25°C were measured using the HACH IntelliCAL phC101 multi-parameter (b). Water depth (c) and transparency (d) were measured using a homemade water level gauge and Secchi disk, respectively. The width of the sampled wetted section was measured using a tape measure. The water speed measured in m/s, thanks to an empirical method which consisted in recording the distance and the time traveled using a stopwatch by a float immersed and followed over a certain distance. Subsequently the flow rate was deducted from the formula.
| Q = V X S | with | V = Flow velocity in m/s |
| S = wetted area in m2 | ||
| Q = flow in m3/s |
The fish samples were collected from July 2019 to September 2019 using different techniques: gillnets 20 m long and 1 m high and whose meshes between knots were 1.5 cm, 2cm, 2.5cm, 3cm and 6cm; traditional traps about 1 m long and 30 cm in diameter. Individual lines and longlines 25 m long with number 8 hooks; 10; 12; 14; 16; 18 and 20 which were baited with earthworms, molluscs and small fish. Harpoons were also used to capture large specimens.
3. Results
3.1. Descption of differents results
3.1.1. Spatio-temporal dynamics of land use in the Likouala aux Herbes watershed between 1980, 2000 and 2018
- we observe a predominance of dense and swampy forests and savannah dynamics around the catchment area of the Likouala river with herbs, i.e.: 2.29% (10,340.55 ha) of the area of the RCLT in 1980, 9 .37% (7,716.3ha) in 2000 and 13.35% (60,281.64 ha) in 2018 (Figure A1);
- The analysis reveals that the dense floodplain forest and the swamp forest have remained substantially unchanged i.e. respectively (203,404.59 ha) 45% of the area of the LTCR and (184,343.85 ha) 40.8% of the LTCR between 1980 and 2018;
- The water superficies’ have also remained constant regardless of their evolution over time and the different dry and rainy seasons, or 0.7% (3,423.06 ha) of the area of the LTCR. Table A2 and Table A3 show the variation in land use areas between the different periods;
- The occupation classes: seasonally flooded gallery forest and herbaceous savannah were converted respectively by 24.11% and 65% into shrubby savannah and wooded savannah.
- From 2000 to 2018, the occupation classes : water, swamp forest and dense floodplain forest have remained practically unchanged as in the dynamics of the previous period ;
- However, according to the observation of Figure A1 and the analysis of Table A2, it can be said that the savannah class composed in 2,000 of herbaceous savannah 0.58%, 0.7% tree savannah and 1.7% shrubby savannah were completely converted respectively into slash and burn savannah, shrubby, wooded and herbaceous savannah in 2018, as shown in Figure A1.
3.1.2. Characterization of the ichthyological diversity of the Likouala aux herbes river
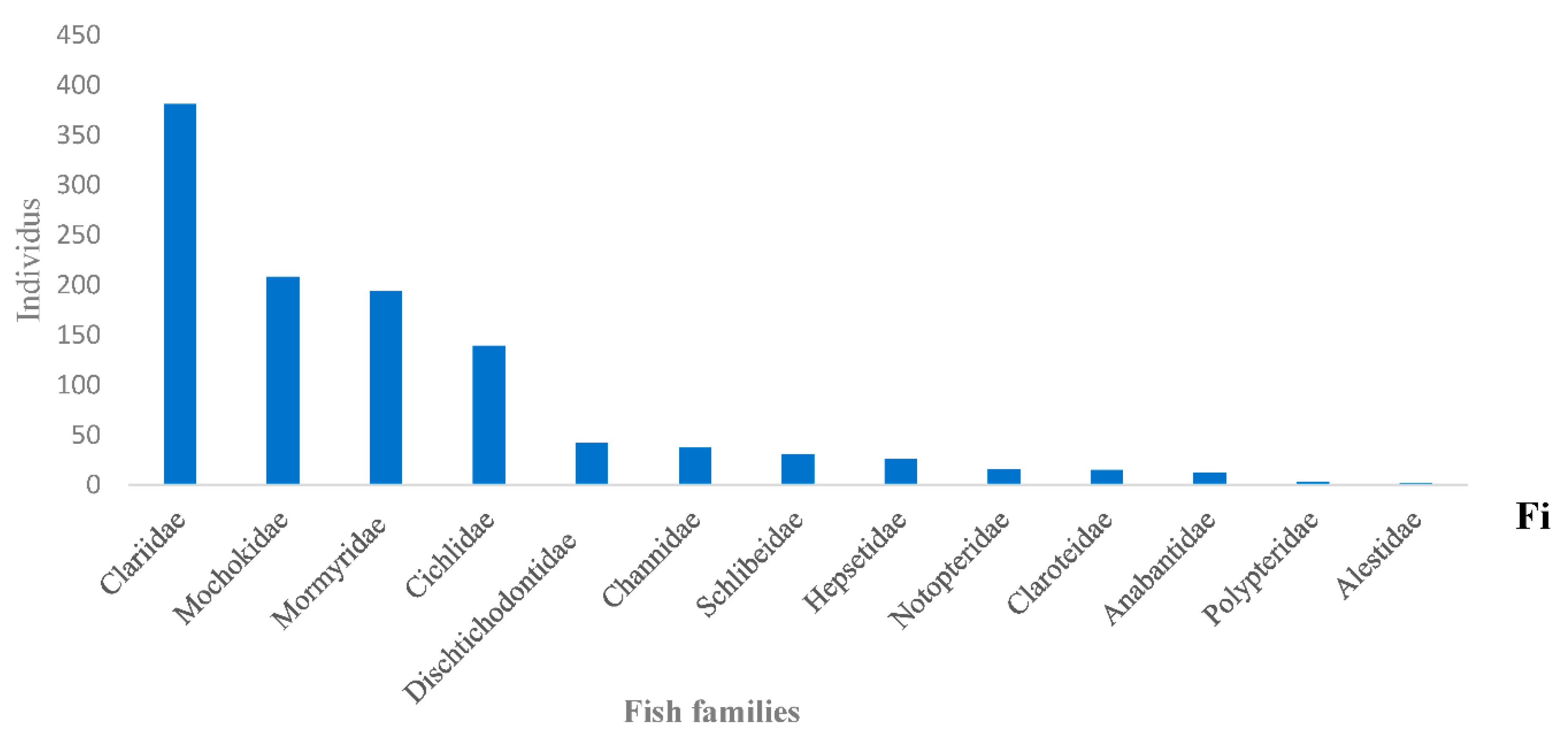
- One thousand one hundred and six (1106) fish in total were harvested, divided into 25 genera belonging to 13 families (Table A4);
- The Clariidae family was the most abundant with 381 specimens (Figure A3) representing 34.07%, followed by the Mochokidae family with 208 individuals, or 19.67%. The Bouanela and Mossengue stations contain the largest numbers of individuals, representing respectively 20.61% and 31.1%, while the Moungouma-Bailly and Dzeke stations recorded the lowest numbers of 8 and 28 individuals, i.e. 0.72% and 2 .53% (Figure A3 and Table A4 and Table A5);
- In axis II of the Principal Composant Aanalysis (Component 2, Figure A2 (a)), the distribution of the families Alestidae and Mormyridae was influenced by the presence of sandy type substrates, foliage and macrophytes coupled with the increase in temperature of the 'water (Table A4 and Table A5). The presence of Alestidae according to (Component 2, Figure A2 (b)), is correlated with temperature peaks, namely 28.79 ± 1.7°C for the Botongo station and 28.13 ± 1.17°C for the Epena station (Table A5). As for the Mormyridae, the highest diversity indices in this case; 0.6229 for Mossengue and 0.5993 for Botongo were recorded at the stations with the highest temperature peaks.
- The families of Dichtichodontidae, Hepsetidae, Notopteridae, Anabantidae, Schilbeidae and Polypteridae show no affinity either with the different substrates recorded or with the physico-chemical parameters (Table A4 and Table A5)of the water in axis III, paradoxically to the distribution of the previous families of fish species presented in axes I and II (Figure A2) and (Table A5).
3.2. Figures, Tables and Schemes
Figure A1.
Land cover dynamics analysis of LTCR in 1980, 2000 and 2018.
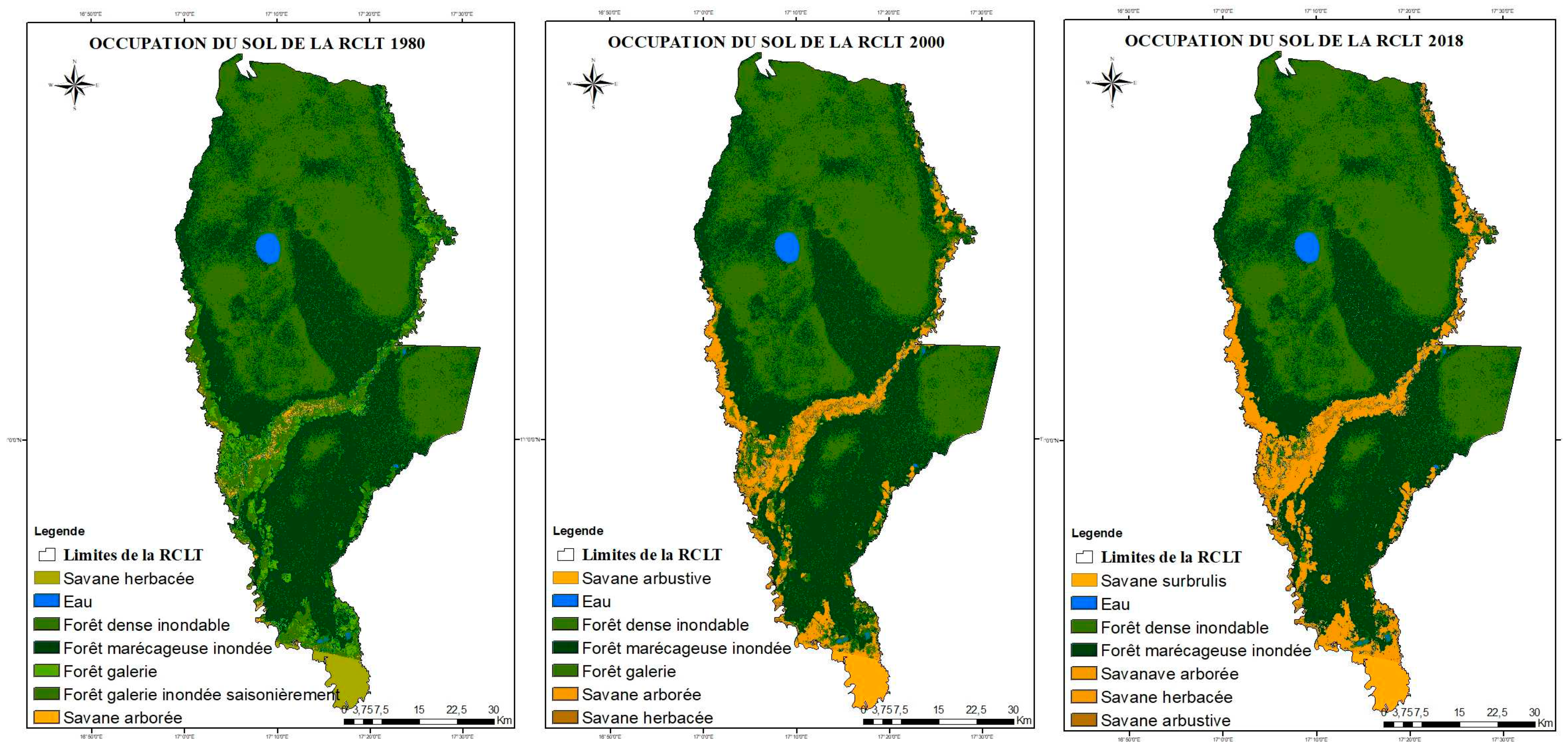

Table A1.
Distribution of LTCR land cover classes between 1980, 2000 and 2018
| Land cover (ha) | 1980 | 2000 | 2018 |
|---|---|---|---|
| Grassy savannah (ha) | 7708,68 | 2631,87 | 31972,86 |
| Wetlands (ha) | 3423,06 | 3423,06 | 3423,06 |
| Flood dense forest (ha) | 203404,59 | 203404,59 | 203404,59 |
| Flooded swamp forest (ha) | 184343,85 | 184343,85 | 184343,85 |
| Gallery forest (ha) | 17968,23 | 17968,23 | 17968,23 |
| Seasonally flooded gallery forest | 31972,86 | 7708,68 | 7708,68 |
| Tree savannah (ha) | 2631,87 | 31972,86 | 2631,87 |
| TOTAL (ha) | 451453,14 | 451453,14 | 451453,14 |
Table A2.
Variation in land use areas of the LTCR from 1980 to 2000.
| Areas (ha) | Variation of areas (ha%) | |||
|---|---|---|---|---|
| Land cover | 1980 | 2000 | (ha) | % |
| Grassy savannah (ha) | 7708,68 | 2631,87 | +5076,81 | + 65,85 |
| Wetlands (ha) | 3423,06 | 3423,00 | 0,00 | 0 |
| Flood dense forest (ha) | 203404,59 | 203404,59 | 0,00 | 0 |
| Flooded swamp forest (ha) | 184343,85 | 184343,85 | 0,00 | 0 |
| Gallery forest (ha) | 17968,23 | 17968,23 | 0,00 | 0 |
| Seasonally flooded gallery forest (ha) | 31972,86 | 7708,68 | -31 904,86 | - 24,11 |
| Tree savannah (ha) | 2631,87 | 31972,86 | -29 340,13 | 9,23 |
| TOTAL | 451453,14 | 451453,14 | 100,00 | |
Table A3.
Variation in land use areas of the LTCR from 2000 to 2018.
| Areas (ha) | Variation of areas (ha et %) | |||
|---|---|---|---|---|
| Land cover units | 2000 | 2018 | (ha) | % |
| Grassy savannah | 7708,68 | 7708,68 | 0,00 | 0 |
| Wetlands | 3423,06 | 3423,06 | 0,00 | 0 |
| Flood dense forest | 203404,59 | 203404,59 | 0,00 | 0 |
| Flooded swamp forest | 184343,85 | 184343,85 | 0,00 | 0 |
| Gallery forest | 17968,23 | 12339,77 | -5623,23 | - 31,29 |
| Tree savannah | 31972,86 | 44917,86 | +12 944,86 | + 15,48 |
| Overburn Savannah | 2631,87 | 1231,87 | -1403,00 | + 53,3 |
| TOTAL | 451453,14 | 451453,14 | 100,00 | |
Figure A2.
Results of Principal Component Analyzes (PCA) between the hydromorphological, physico-chemical parameters and the distribution of fish families from the Likouala to the herbs recorded in the 7 pilot villages of the RCLT between July and September 2019.
Figure A2.
Results of Principal Component Analyzes (PCA) between the hydromorphological, physico-chemical parameters and the distribution of fish families from the Likouala to the herbs recorded in the 7 pilot villages of the RCLT between July and September 2019.
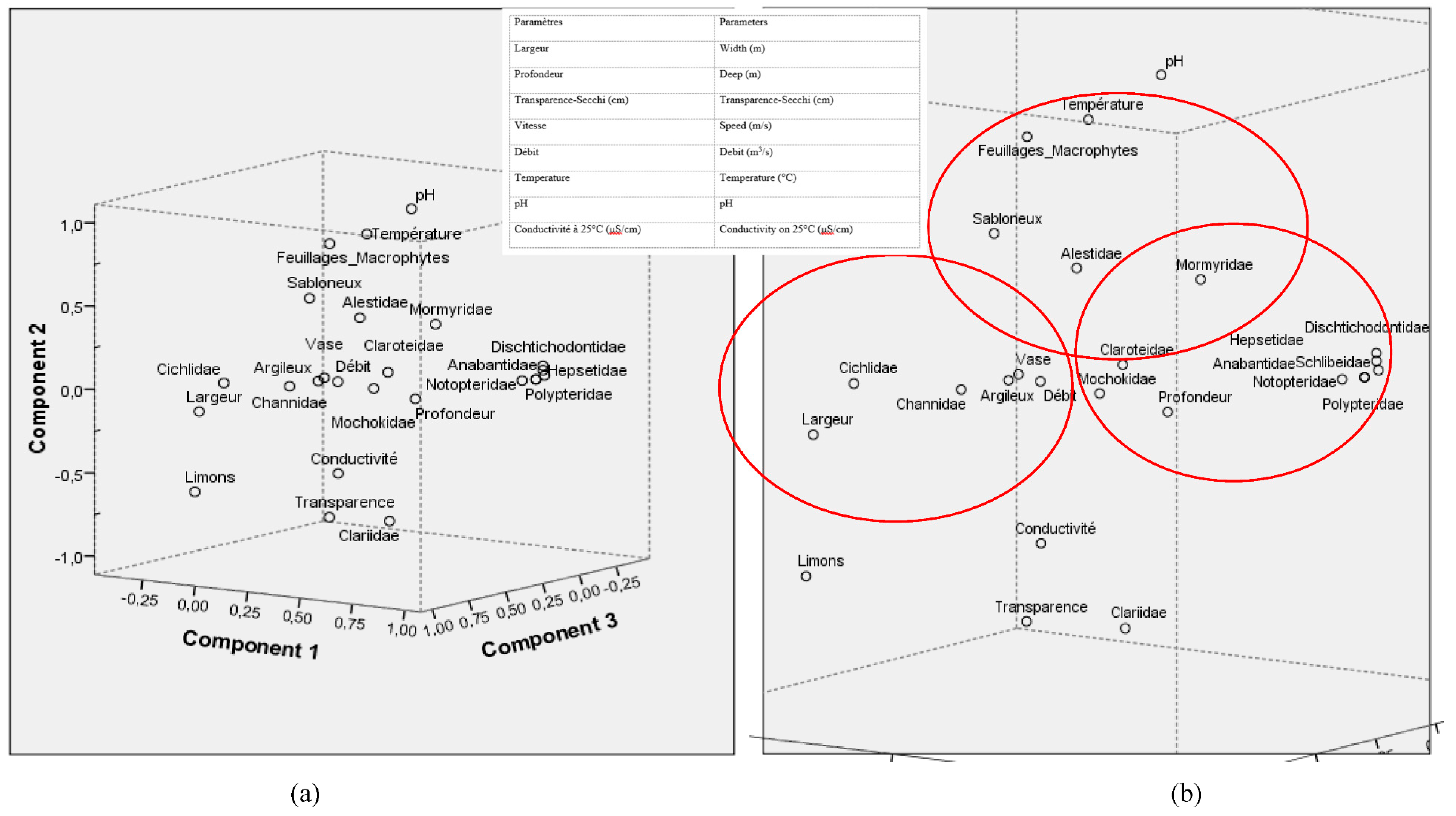
Figure A3.
Fishs families identified in the Likouala aux Herbes river near 7 pilot villages of the RCLT, from July to September 2019.
Figure A3.
Fishs families identified in the Likouala aux Herbes river near 7 pilot villages of the RCLT, from July to September 2019.
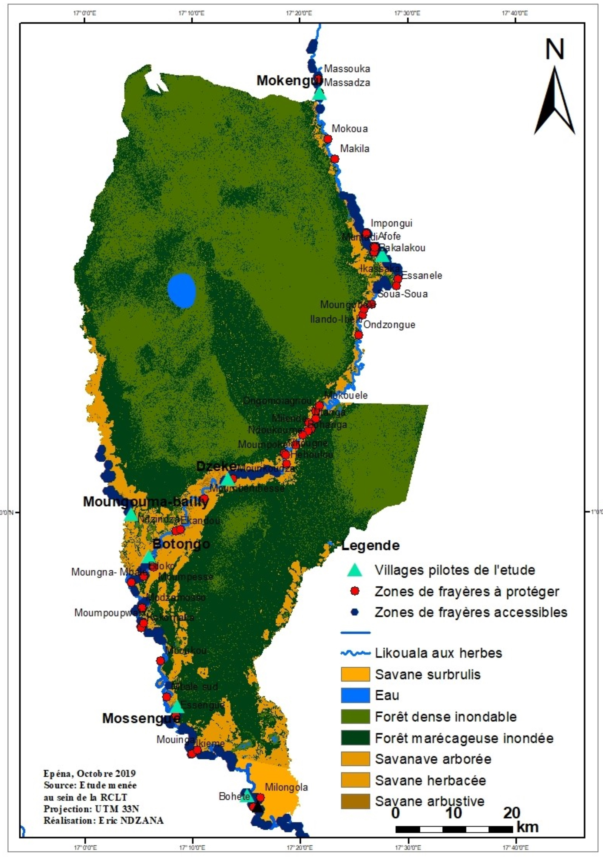
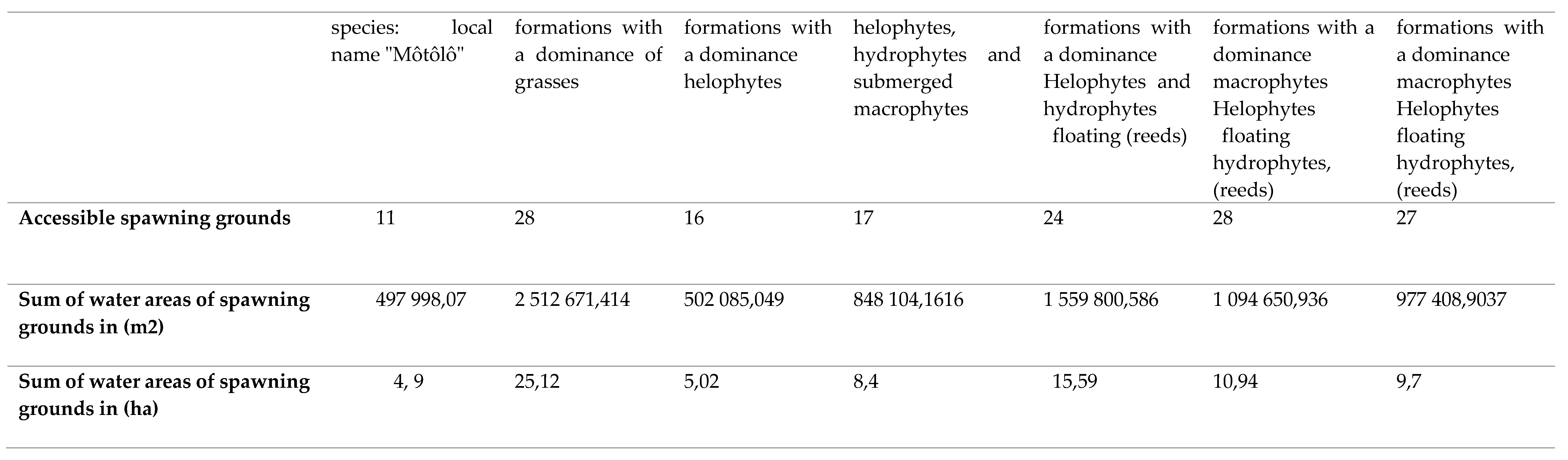
The spawning ground is a space developed by fish species where they reproduce, feed and ensure the development of their fry and/or fry. These sites are generally subservient to the natural environment in which they are built and also serve as shelter during predator attacks. They are generally found along the shore, on the sides of the banks of watercourses, floodplains, substrates; vegetation, rocky, sandy or on the benthos (Figure A4 and Table A5). The spawning grounds identified (Figure A4) between July and September 2019 in the RCLT are mainly present in the gallery forests that run along all of the three sub-basins of the Likouala river with herbs. Certain spawning grounds, by consensus with the fishing communities of the RCLT, as part of the project for the development and validation of the Management Plan of the RCLT, have been declared accessible in order to allow fishermen to obtain permanent supplies. in fishery resources regardless of the season during the year. However, in order to guarantee the sustainability of the fishing activity within the RCLT, certain spawning grounds, in a participatory manner, have also been set aside through a fishermen's charter. (Figure A4) shows the location of these two types of spawning grounds established in a participatory manner with the fishing community in the LTCR.
Figure A4.
Mapping of prohibited and accessible spawning grounds in the Likouala aux Herbes near 07 pilot villages of the 3 fish basins of the RCLT.
Figure A4.
Mapping of prohibited and accessible spawning grounds in the Likouala aux Herbes near 07 pilot villages of the 3 fish basins of the RCLT.
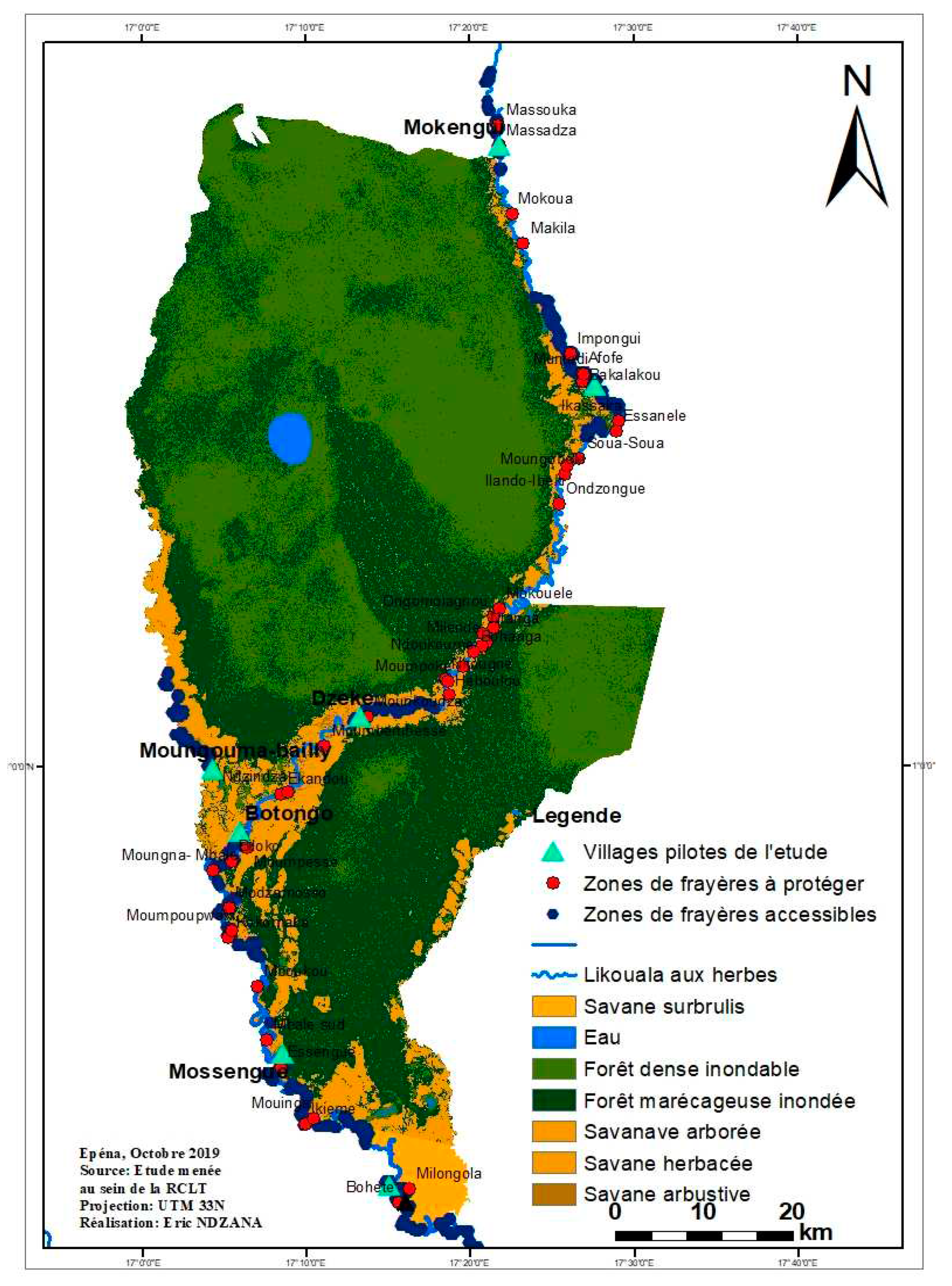
Table A4.
Diversity of fish populations identified in seven stations of the Likouala aux Herbes (RCLT) from July to September 2019.
Table A4.
Diversity of fish populations identified in seven stations of the Likouala aux Herbes (RCLT) from July to September 2019.
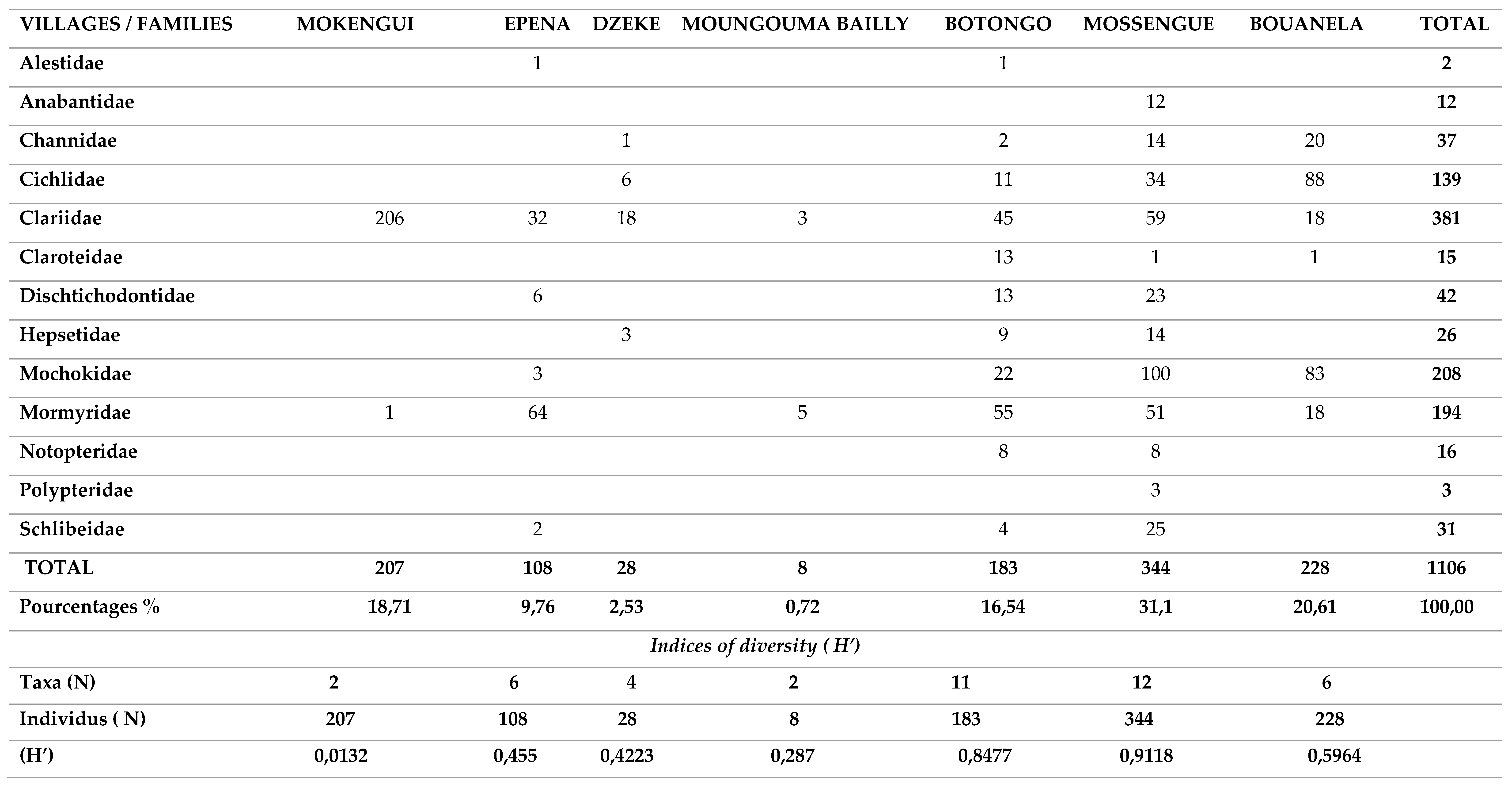
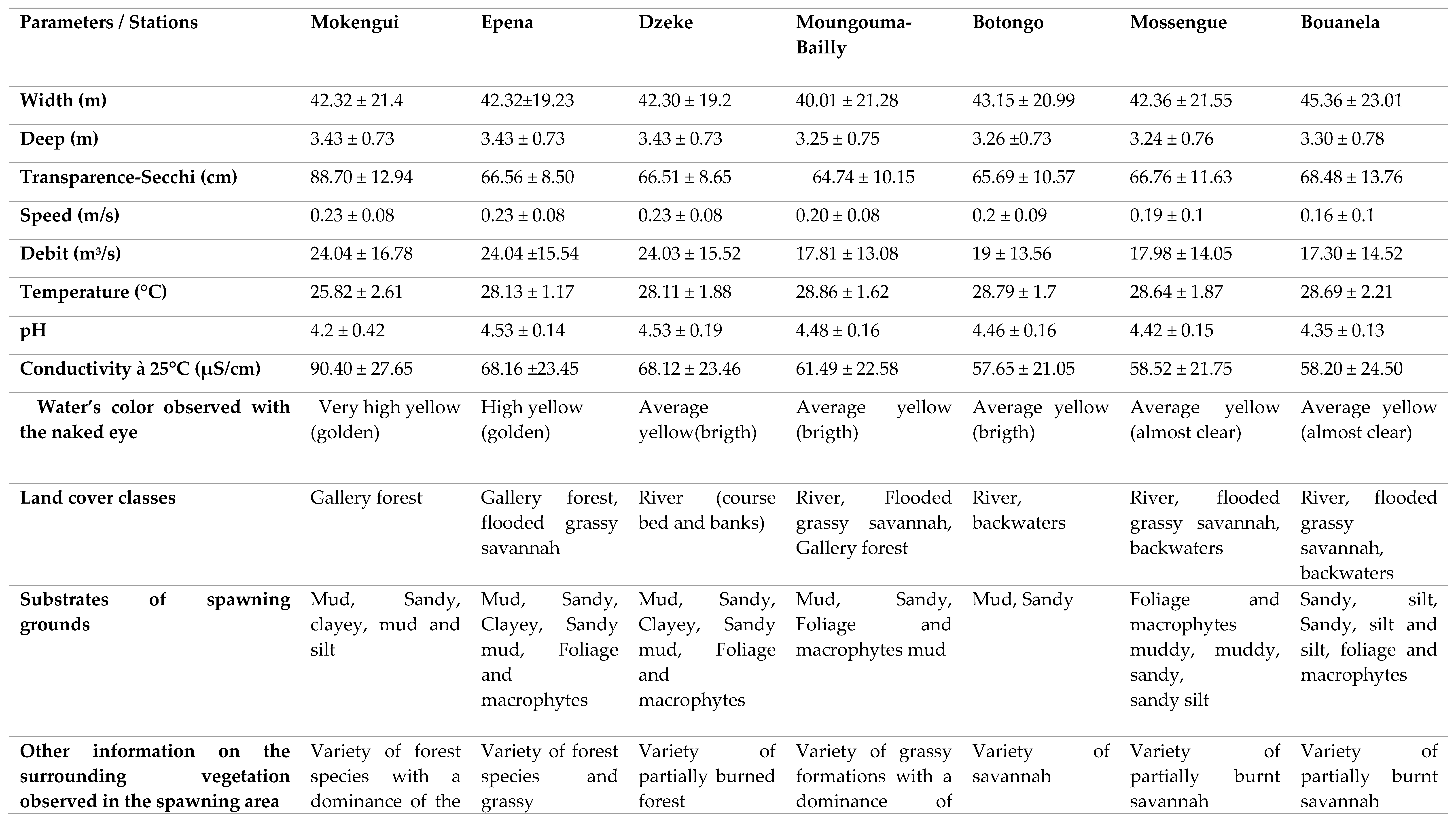
Table A5.
Compared average hydromorphological and physico-chemical characteristics of the spawning grounds of the Likouala river with herbs near 7 pilot villages of the 3 LCRT fish basins from July to September 2019.
Table A5.
Compared average hydromorphological and physico-chemical characteristics of the spawning grounds of the Likouala river with herbs near 7 pilot villages of the 3 LCRT fish basins from July to September 2019.
4. Discussion
The analysis of land cover from 1980 to 2018 showed that the RCLT remained mostly stable at 86% over time. However, in 38 years, the evaluation of the spatio-temporal dynamics of the vegetation of the RCLT has revealed the regression of gallery forests and mosaics of seasonally flooded dense forests, which occupied almost 13% of the RCLT in favor of savannah formations (grassy, tree, shrub and slash and burn). These results are similar to those obtained by Taibou Ba [10]), in the Saloum delta where they present the conversion of open and dry forests to savanna formations between 1979 and 1999. This dynamic of land cover in the center of Togo was also presented [12]. In 24 years, the area of open and dry forests has been degraded and has decreased by 157,988 ha, representing a regression rate of 23.44%. The reasons that explain these changes to the environment are identical regardless of the sites: probably in part climate change but also and above all the intense and permanent exploitation of natural resources by man who systematically sets fire to this area to recover some fish in small permanent ponds or in the process of drying up but difficult to access. In the LTCR, forests provide many ecosystem goods and services that benefit all people in and around the LTCR villages. In addition to Non-Timber Forest Products (NTFPs), forests are spiritual places for communities [5]). Many indigenous communities (pygmies) present in the villages of the LTCR have lifestyles entirely dependent on the forest. Certain forest species used for pharmacopoeia, wood-energy and materials for various activities (construction, crafts, etc.) are regularly extracted in the gallery and dense flood-prone forests encountered in the LTCR. The practice of slash-and-burn agriculture. The conditions of exploitation of forest areas sometimes completely flooded in the rainy season, impose a considerable effort in the dry season on the communities of the RCLT, who are culturally fishermen, to develop them. In order to capitalize on the effort, they proceed to the use of bush fires to recover some fish and create agricultural areas. The extensive slash-and-burn agriculture forces farmers to clear several hectares each year [12]. This causes the degradation of the plant cover and therefore the impoverishment of the soil. The increase in population density could be a driving force of this degradation which does not exclusively affect the terrestrial forest environment, but also the aquatic environment. The fish resources of the RCLT as well as the large terrestrial fauna regulate their lifestyles according to the forest environment. The edge that comes from the decomposition of the organic matter of dead trees or parts of trees (leaves, bark, branches, roots and fruits) contributes to strengthening the aquatic food chain on which biodiversity in general and ichthyology in particular strongly depends. By destroying gallery forests through agricultural practices based on the use of bush fires, certain species of fish whose feeding and reproduction are conditioned by the presence of a ligneous support (tree roots) will hardly ensure the survival of their species. This observation was made during the study period when the most minimal fishing performance both in terms of diversity and quantity of catches were recorded in the villages of the RCLT located in the North axis. where gallery forests have been converted into shrubby savannah by bush fires, notably in Epéna, Matoko Botakola and towards Mokengui. This corroborates the analysis of the spatio-temporal dynamics which revealed that the gallery forests are entirely converted between 1980, 2000 and 2018 into herbaceous, tree, shrub and slash and burn savannahs, to the detriment of the sustainability and durability of the activity. fishing which is the main activity of the communities of the villages of the RCLT. The fisheries of the Likouala River with herbs commonly called "spawning areas" were diagnosed during the period from July to September 2019. A total of 204 spawning areas (i.e. 162 accessible areas and 42 prohibited areas) were identified in the 07 pilot villages of the study in 3 LTCR Fish Basins. Overall, the ecological characteristics of these spawning grounds do not differ significantly from one station to another. The previous studies clearly show that the physico-chemical parameters of the waters of the Likouala aux Herbes are substantially identical from upstream to downstream [3,4,5]. This further reinforces the results recorded during the study period in the various villages of the RCLT. The sampling stations were compared with each other based on each hydromorphological parameter measured, independently of other factors between July and September 2019. The variation in the width of the Likouala aux herbes river changes from upstream to downstream. Thus, the Bouanela and Mossengue stations have greater values of the bed width of the Likouala river with herbs. On the other hand, the depth does not vary significantly from one station to another. the Likouala aux Herbes River is a watercourse with a very low gradient due to the topography of the RCLT [3]. This could justify the practically identical value (3.43 ± 0.73 at Mokengui in the north of the LTCR and 3.30 ± 0.78 in the south at Bouanela) of the depth over the entire extent of the waters of the Likouala aux Herbes in the LTCR. However, width and depth are two hydromorphological parameters that contribute to the diversification of ichthyofauna in ichthyological hydrosystems. Several studies carried out on different hydrosystems have already established relationships between species and environmental variables. Distribution of Mormyridae in the Yoko River at Kisangani in the DRC was strongly correlated with a large width of the stream as well as the depth [2]. According to ([2] et [11]) the average water temperature in a rainforest stream rarely exceeds 24°C’. The average temperature recorded during the water sampling campaigns in the various pilot villages was mostly high in the Reserve with values that fluctuated around 28°C with the exception of the Mokengui station which recorded 25°C. The similar values for the temperature of 27.3°C on Likouala with herbs was obtained by Mamonekene [3]. The high rainfall gallery forest with shading caused by contiguous treetops that run along the river may be the source of this value which supports assertion regarding the impact of forest vegetation cover on temperature variation in the Congo Basin [2]. The waters of the Likouala aux Herbes are acidic with pH values hovering around 4 [3]. The results obtained during the study are identical to those of Mamonekene with values of 4.2 ± 0.42 to Mokengui and 4.35 ± 0.13 in Bouanela [3].The putrefaction of organic matter resulting from the continuous decomposition of drained plants (mudflat, wood debris, tree fruits, helophytes and macrophytes) all along the river contributes through oxidation-reduction reactions to reduce the pH waters of the river). The conductivity at 25°C recorded during the data collection campaign in the pilot villages of the RCLT, show higher values in the North of the reserve (90.40 μS/cm ± 27.65 in Mokengui and relatively low values in Bouanela in the South of the RCLT (58.20 μS/cm ± 24.50).These values are close whose conductivity values at 25°C ranged from 56.5 μS/cm to 84.93 μS/cm [3]. The type of substrate encountered in the different stations of the Likouala river with herbs between July and September 2019 presents a great diversity from upstream to downstream and is mainly dominated by the mud-sand couple. A study carried out in Pool Malebo in the DRC on the Congo River, of which the Likouala River by the Sangha River is one of the important tributaries in the North of the Republic of Congo, specifies that the diversity of substrates encountered in this part of the Congo River is one of the factors responsible for the diversity of fish species present in the Pool Malebo fishery [9]. But in addition to this diversity of station substrates, it is also important to mention that the diversity of vegetation mainly encountered in the southern part of the reserve in the pilot villages Mossengue, Botongo, Bouenala whose aquatic vegetation is essentially made up of a variety of partially burned savannah formations with a dominance of macrophytes helophytes, floating hydrophytes, Eichornia crassipes, and fixed Echinochla crassipes and many others also constitute fundamental elements to be taken into consideration in the description of the sampling stations. In short, the physico-chemical characteristics being identical in all the stations, particularly in the spawning grounds, it clearly appears that the diversity of the fish populations would be more linked to the diversity of the microhabitats encountered in the Likouala river with herbs. The interpretation of the data on the physico-chemical and hydromorphological parameters as well as the study of the substrate of each station (Table A5) showed that the stations of the South axis of the RCLT, have in addition a greater width in the area, a diversified aquatic vegetation that colonizes the banks and the bed of the watercourse, fixed or floating on the waters of the Likouala river with herbs. All this diversity of substrates in the waters of these localities greatly contributes to creating micro-habitats favorable to the development of fish species. The more these microhabitats dependent on parameters such as the width of the river bed and the depth are diversified, the more the ichthyofauna of these localities will be diversified. This remark was confirmed, who asserted that in ecoregion 539, tropical floodplain forest rivers consist of diverse micro-habitats, likely to be the source of high biodiversity and endemism [1]. It is thought that the number of species of Mormyridae inventoried results from this factor. The same observation regarding the distribution of Cyprinidae Labeo has been made in the Malebo pool [9]. Their distribution is more related to the diversity of micro-habitats encountered in the study area and whose main substrates were: mud, sand, stone and silt. In axis II of the PCA (Component 2, Figure A2(a)), the distribution of the families of Alestidae and Mormyridae was influenced by the presence of sandy type substrates, foliage and macrophytes coupled with the increase in temperature some water. The presence of Alestidae is correlated with temperature peaks, namely 28.79 ± 1.7°C for the Botongo station and 28.13 ± 1.17°C for the Epena station (Table A4 and Table A5). As for the Mormyridae, the highest diversity indices in this case; 0.6229 for Mossengue and 0.5993 for Botongo were recorded at the stations with the highest temperature peaks. The families of Dichtichodontidae, Hepsetidae, Notopteridae, Anabantidae, Schlibeidae and Polypteridae, show no affinity either with the different substrates recorded or with the physico-chemical parameters of the water in axis III. However, they have affinities with each other, hence their clustering in axis III. This affinity would be linked to the different trophic relationships (predator-prey) that exist between the ecological niches of these different species. In short, the diversity of fish species encountered in the Likouala results from our study of the different micro habitats existing in the bed and outside the bed of the watercourse.
5. Conclusions
In conclusion, the study on the contribution of remote sensing and GIS in the sustainable management of fisheries resources of the Lac Télé Community Reserve on the Likouala aux Herbes River, has made it possible; To have primary knowledge of the dynamics of land use around the basin of the Likouala river with herbs in the LTCR which is deteriorating under the probable effect of climate change but also and above all the direct action of man who each year burns the floodplain forest and the gallery forest degrading the potential spawning grounds;
- To know the distribution dynamics of fish species populations during the period from July to September 2019 in the LTCR;
- To locate and characterize the 204 spawning grounds of the Likouala aux Herbes fisheries
Regarding the spatio-temporal dynamics in the LTCR, the analysis of the land cover of the LTCR using Landsat MSS, ETM+ and OLI images revealed significant changes in the vegetation cover between 1980, 2000 and 2018. Thus, the rate of forest area decreased by 21.41% for the entire LTCR to the benefit of the savannahs which respectively increased by 15.23%. That is a conversion rate of 13% over the entire analysis period. The ichthyological diversity recorded during the period from July to September 2019, in the 07 pilot villages of the study of the 3 fishing basins of the Likouala in the LTCR would be due to the diversity of the micro-habitats noted in the villages of the axis South of the LTCR as described in the characterization phase of the spawning grounds. For the mapping of fishing activity in general and the ecological characterization of the 204 spawning grounds; the hydromorphological parameters did not vary significantly from one pilot village to another. However, we noticed a diversity of substrates in the spawning grounds of the villages of the southern axis of the LTCR, the main components of which were the mud-sand couple and the influx of aquatic vegetation composed of Helophytes, fixed and floating macrophytes on the waters of the Likouala aux Herbes. By way of perspective, the extent of research opportunities with the aim of strengthening knowledge of biodiversity in general and ichthyological in particular in the LTCR, in order to contribute to the preservation of this RAMSAR site, would be a key asset for conservation. generally, in the Congo Basin and particularly in the Republic of Congo.
Author Contributions
Conceptualization, Eric NDZANA. and Jean-Claude Micha.; methodology, Eric NDZANA and Ben EVANS; software, Eric NDZANA.; validation, Ben Evans, Jean-Claude MICHA. and Victor MAMONEKENE.; formal analysis, Eric NDZANA.; investigation, Eric NDZANA.; resources, Ben EVANS, Eric NDZANA.; data curation, Eric NDZANA.; writing—original draft preparation, Eric NDZANA.; writing—review and editing, Jean-Claude MICHA.; visualization, BEN EVANS.; supervision, Jean -Claude MICHA.; project administration, BEN EVANS.; funding acquisition, BEN EVANS. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Deutsche Gesellschaft für Internationale Zusammenarbeit (GIZ), grant number ERAIFT ECOFAC 6 - RCO/FED/ 039-795 and “The APC was funded by Wildlife Conservation Society (WCS)”by CARPE fund.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Ethics Committee of ERAIFT (protocol code 0034 and 09/11/2019).” for studies involving humans and animals.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Written informed consent has been obtained from the patients to publish this paper.
Data Availability Statement
Suggested Data Availability Statements are available in section “MDPI Research Data Policies” at https://www.mdpi.com/ethics.
Acknowledgments
The authors sincerely thank the organizations, United Nations Educational, Scientific and Cultural Organization (UNESCO), Deutsche Gesellschaft für Internationale Zusammenarbeit (GIZ) and Wildlife Conservation Society (WCS) for the funding granted for the execution of the research work which led to these results. They also thank the University of Kinshasa (UNIKIN) and the Regional Post-University School of Forestry Planning and Management of Tropical Territories (ERAIFT) for their material and technical assistance in carrying out this research. The authors also extend their gratitude to the reference readers who were able to revise the manuscript for improvement. The authors also thank the populations of the localities of the Likouala Department in the Republic of Congo in general and particularly those of the pilot villages that hosted the research work, for their hospitality and their generosity during the collection periods spent in these respective villages.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results”.
References
- Ashley B, Robin A. Ecoregion 539: Upper Congo Rapids in Freshwater ecoregion of the world 2008. Conservation Science Program, WWF-US: Washington, DC, USA., pp. 234-235.
- Byanikiro Richard Mahamba, A.B.; Jacob Ndjaki Nsila, C.D.; Alidor Kankonda Busanga, E.F.; Joseph Ulyelali-patho,G.H.; Jean-Claude MICHA,I.J. Caractérisation et écologie des peuplements des mormyridae dans la réserve de Yoko (Kisangani, RD Congo). International Journal of Biological and Chemical Sciences 2017, pp.13-15.
- Mamonekene Victor, A.B. Les ressources halieutiques de la Réserve communautaire du Lac Télé Likouala aux Herbes. Diversité et exploitation, Rapport de consultance 2006.
- Micha Jean Claude, A. Défis dans la conservation du poisson dans les aires protégées du bassin du Congo, 2019, La Revue Nature & Faune Volume 32, Numéro 2, 69 – 74. http://www.fao.org/3/ca4151fr/ca4151fr.pdf.
- Micha Jean Claude,A.B. ; Shango Mutambue,C.D.; Felin Twagirashyaka, E.F., Roger Mobongo,G.H., Parfait Bakabana, I..J., Thierry Mangonga,K.L. Plan d’aménagement et de gestion des zones humides de la réserve communautaire du lac Télé 2016, République du Congo (PAGZHRCLT),pp. 4-5.
- Moutsambote A.B. ; Rapport de mission botanique effectuée au lac Télé (District d'Epéna, Likouala) 08-27 juin 1992, Laboratoire de Botanique, CERVE, Brazzaville, Congo, pp.6.
- Paugy D A.B. ; Lévêque C.C.D. ; Teugels GG, E.F. Poissons d’Eaux Douces et Saumâtres de l’Afrique de l’Ouest, Edition ORSTORM, (Tome I et II), Institut français de recherche scientifique pour le développement en coopération, Paris, France, 1990, ; pp. 56.
- Poll M, Gosse JP.A.B. Généra des Poissons d’Eau Douce de l’Afrique (eds). Académie Royale de Belgique : Bruxelles. 1995, pp.15.
- Pwema Kamfu A.B. ; Nseu Mbomba Bekeli C.D. ; Lise-marie Pigneur E.F. ; Takoy Lomema, G.H. ; Jean-Claude MICHA, Environmental variables structuring Labeo species (Pisces, Cyprinidae) in Malebo Pool, Congo River ; International Journal of Biological and Chemical Sciences 2011, Democratic Republic of Congo, pp.34-38.
- Taibou Ba, A.B.; Dieynaba Seck. C.D. Dynamique de L’Occupation des sols, cartographie des CLPA, des zones de pêche et mise en place d’un système d’information géographique. Centre de Suivi Ecologique et USAID/COMFISH Project 2012, Senegal, University of Rhode Island, Narragansett RI, 66 pp.
- Tomedi Eyango M., A.B. ; Efole E.T., C.D. ; Blé M.C., E.F. ; Ndzana B.E.B., G.H. ; Songmo B. L., I.J. ; Nyamsi Tchatcho N. L., I.J. ; Mikolasek O., L.M. ; Tchoumbougnang F., N.O. ; Rizipisciculture : voie d’intensification écologique des systèmes piscicoles extensifs F.Tech&Doc. Vul. : 18-23-Volume spécial 2015, CRO : Centre de Recherche Océanographique d’Abidjan. Côte d’Ivoire, pp.30-32.
- Zakariyao Koumoi,A.B. ; Abdourazakou Alassane,C.D. ; Minkilabe Djangbedja,E.F. ; Tchaa Boukpessi, G.H. ; Ama-Edi Kouya I.J.; Dynamique spatio-temporelle de l’occupation du sol dans le centre-togo. ahoho - Revue de Géographie du LARDYMES, Université de Lomé, 2013, 7 (10), pp.163-172. <hal-01564256>.
- Zebaze Togouet S. H., A.B. ; Njine T., C.D. ; Kemka N., E.F. ; Foto Menbohan S., G.H. ; Niyitegeka D., I.J. ; Ngassam P. K.L. ; Boutin C., M.N. Composition et distribution spatio-temporelle des protozoaires ciliés dans un petit lac hyper trophique du Cameroun (Afrique Centrale). Revue Science Eau 2006, Volume 19, pp. 151-162.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



