
Submitted:
17 October 2023
Posted:
19 October 2023
You are already at the latest version
Abstract
Serine/threonine kinase (AKT) signaling regulates diverse cellular processes and is one of the most important aberrant cell survival mechanisms associated with tumorigenesis, metastasis, and chemoresistance. Targeting AKT has become an effective therapeutic strategy for the treatment of many cancers. AKT3 (PKBγ), the least studied isoform of the AKT family, has emerged as a major contributor to malignancy. AKT3 is frequently overexpressed in human cancers, and many regulatory oncogenic or tumor suppressor small non-coding RNAs (ncRNAs), including microRNAs (miRNAs), have recently been identified to be involved in regulating AKT3 expression. Therefore, a better understanding of regulatory miRNA/AKT3 networks may reveal novel biomarkers for diagnosis of patients with cancer and provide invaluable information for developing more effective therapeutic strategies. The aim of this review is to summarize current research progress in the isoform-specific functions of AKT3 in human cancers and the roles of dysregulated miRNA/AKT3 in specific types of human cancers.
Keywords:
cancer
; microRNA
; AKT3
; AKT signaling
; small non-coding RNAs
; circRNA
; lncRNA
; epigenetic regulation
1. Introduction
AKT, also known as protein kinase (PKB, RAC-PK), is a serine/threonine protein kinase with critical roles in regulating cell signaling in many biological processes. Through phosphorylation of a variety of substrates, AKT regulates the cell cycle and cell proliferation, and inhibits apoptosis via the inactivation of proapoptotic proteins [1]. Aberrant activation or expression of AKT has been reported in many types of human cancers, and found to be involved in tumorigenesis, cancer progression, recurrence, and drug resistance [2]. AKT has become an important target for the treatment of human cancer [3]. AKT is activated by a wide range of growth signals, via phosphoinositide 3-kinase (PI3K)-dependent mechanisms. PI3K, a member of the lipid kinase family, is an important regulator of signaling and intracellular vesicular trafficking [4]. Activated PI3K further activates 3-phosphoinositide-dependent protein kinase 1 (PDK1) and AKT by increasing the production of phosphatidylinositol-3,4,5-trisphosphate (PIP3) from phosphatidylinositol 4,5-bisphosphate (PIP2) [5]. Nonetheless, phosphatase and tensin homologue (PTEN), a well characterized tumor suppressor that inhibits the oncogenic pathway in many cancers, negatively regulates AKT by dephosphorylating PIP3, thereby forming PIP2 [6].
The expression of AKT, in addition to being regulated by proteins, is directly regulated by non-coding RNAs (ncRNAs) [7], which are transcribed from most of the human genome (as much as 80%). Functional ncRNAs, such as microRNAs (miRNAs), circular RNAs (circRNAs), and long ncRNAs (lncRNAs), have garnered research interest because of their versatile roles in fine-tuning signaling pathways involved in diverse biological processes and cancer pathogenesis [8]. MiRNAs are endogenous, highly conserved, and 20–22 nucleotides in length. They block messenger RNA (mRNA) translation or cleavage by binding the 3′-untranslated regions (3’-UTRs) of target mRNAs, including AKT mRNA [7]. The regulatory effects of miRNAs on the PI3K/AKT pathway in human cancers have been reviewed [9]. Currently, miRNAs are considered essential players in almost all carcinogenic processes. CircRNAs are a class of single-stranded covalently closed ncRNAs characterized by their unique structure formed through a 3′ to 5′ end-joining event [10]. CircRNAs are highly stable, and their expression exhibits spatiotemporal patterns according to cell types, tissues, and developmental stages [11]. They function mainly as competitive endogenous RNAs (ceRNAs) that inhibit miRNA activity, thereby preventing miRNA-mediated suppression of downstream target gene expression [12]. CircRNAs play essential roles in regulating cellular biological phenotypes and many different pathophysiological processes [13]. Recent studies have revealed that circRNAs are often involved in the PI3K/AKT signaling pathway [14]. Nonetheless, lncRNAs, defined as transcripts longer than 200 nucleotides, regulate gene expression at both the mRNA and protein levels by forming specific RNA:DNA, RNA:RNA, and RNA:protein structures [15]. LncRNA expression is restricted to low copy numbers in different cells at specific developmental stages [16]. LncRNAs either act as ceRNA molecular sponges for miRNAs or serve as functional scaffolds that recruit regulatory proteins to their target chromosomal regions [17,18].
Three AKT isoforms—AKT1 (PKBα), AKT2 (PKBβ), and AKT3 (PKBγ)—are encoded by three different genes that are conserved in mammalian genomes [19]. Despite sharing similar structures (approximately 80% amino acid similarity) [20], AKT isoforms are activated in an isoform-specific manner [21,22], and exhibit different and potentially opposing biological functions, depending on the cellular context. The ablation of individual AKT isoforms exhibits distinct phenotypes in gene knockout mice: Akt1 knockout mice show growth perinatal lethality and elevated apoptosis [23], Akt2 knockout mice exhibit a diabetes-like phenotype [24], and Akt3 knockout mice have small brains with mild neurologic defects [25]. All these differences may be caused by the distinct tissue distribution and subcellular localization of AKT isoforms. AKT1 localizes primarily to the cytoplasm, AKT2 localizes to the nucleus, and AKT3 localizes to both the nucleus and the nuclear membrane [26].
The AKT3 encoding gene is located on chromosome 1q44 [27]. Like other AKT members, on the structural level, AKT3 contains an N-terminal pleckstrin homology domain, a central kinase domain (catalytic domain), a C-terminal regulatory domain containing a hydrophobic motif site, and a linker region that tethers the pleckstrin homology domain to the catalytic domain [28] (Figure 1). AKT3 has a more restricted tissue distribution than the other two AKT isoforms, and is expressed primarily in the brain, kidneys, testes, and embryonic heart [29]. Aberrant expression of AKT3 is found in many types of cancer, thus indicating an important role of this isoform in tumorigenesis [30]. In contrast to the abundant evidence supporting the involvement of AKT in cancer induction and progression, relatively limited information is currently available regarding the specific role of AKT3 in oncogenesis. Nonetheless, results have revealed that AKT3 is dysregulated in a variety of cancers, and is associated with abnormal proliferation, apoptosis resistance, and poor prognosis of tumors [31]. For example, AKT3 promotes survival of 40–60% non-hereditary melanoma cells and the development of malignant melanomas [32]; in addition, it plays an important oncogenic role in triple negative breast cancer (TNBC), colorectal cancer (CRC), invasion and metastasis of glioma, and lung cancer [2,33,34]. AKT3 may also be involved in tumor angiogenesis by promoting endothelial cell growth, as supported by studies showing that increased endothelial cell proliferation in hemangioma (a benign vascular tumor derived from blood vessel cells) promotes in vitro angiogenesis of HUVECs as well as in vivo tumor vascularization [35,36]. Herein, we systemically summarize the tumorigenesis related functions of AKT3 and the regulation of its expression by ncRNAs in specific cancer types.
2. AKT3-derived circRNAs
Remarkably, recent studies have identified several circRNAs derived from the AKT3 gene and their involvement in the regulation of human cancers (Table 1): hsa_circ_0017250, hsa_circ_0112784, hsa_circ_0112781, hsa_circ_0017252, and hsa_circ_0000199 [37]. In particular, hsa_circ_0000199, originating from exons 8–11 of the AKT3 gene, is overexpressed in cisplatin-resistant gastric cancer [38]. Cisplatin treatment is the main chemotherapeutic strategy for hematologic and solid tumor malignancies. Hsa_circ_0000199 enhances cisplatin resistance and gastric cancer (GC) cell survival through promoting the expression of phosphoinositide-3-kinase regulatory subunit 1 (PIK3R1) by sponging miR-198 [38]. Moreover, hsa_circ_0000199 has been found to be highly expressed in lung cancer and to bind miR-516b-5p, thereby regulating glycolysis and decreasing cisplatin sensitivity through the miR-516b-5p/STAT3 axis [37]. In addition, high hsa_circ_0000199 levels are associated with the clinical pathology of TNBC. This circRNA downregulates expression of the tumor suppressors miR-206 and miR-613, and consequently promotes cell proliferation, migration, and invasion, and facilitates chemo-tolerance in TNBC cells [39]. In contrast, hsa_circ_0017252 is downregulated in renal cell carcinoma (RCC). RCC is one of the most common malignant cancers, and approximately 60–70% of RCC cases are clear cell RCC (ccRCC). Overexpression of hsa_circ_0017252 suppresses ccRCC metastasis via targeting miR-296-3p, and the miR-296-3p/E-cadherin axis mediates the metastasis inhibitory effect of hsa_circ_0017252 [40]. Another AKT3 derived circRNA, hsa_circ_0017250, arises from the circularization of exons 3–7, whose expression is downregulated in highly malignant glioblastoma multiforme (GBM) and glioma-initiating cells. Hsa_circ_0017250 acts as a tumor suppressor by inhibiting the phosphorylation of AKT, thereby decreasing its activity, and ultimately inhibits glioblastoma cell proliferation and invasiveness [41].
3. AKT3 and miRNA/AKT3 axes in human cancers
3.1. Breast cancer
Breast cancer (BC) is among the most common tumors and has the second highest rate of cancer death in women. BC tumorigenesis is a multistage process involving several genetic and epigenetic alterations [42]. The distinct functions of AKT isoforms in BC have been reviewed by Basu and Lambring [22]. Notably, TNBC is an aggressive BC subtype without an available effective targeted therapy. In TNBC, the AKT3 gene is frequently amplified [22,43], and the novel recurrent fusion oncogene MAGI3-AKT3 is enriched [44]. The high level of AKT3 is significantly associated with patient survival duration [45]. Moreover, a splice variant of the AKT3 gene whose product lacks a key regulatory Ser472 phosphorylation site induces apoptosis and suppresses TNBC cell growth through upregulating pro-apoptotic Bim, and activating Bax and caspase-3 processing [45]. Findings from several functional studies also support the oncogenic roles of AKT3 in BC, as follows. 1) AKT3 has been found to be required for TNBC growth, through its downregulation of the cell-cycle inhibitor p27. AKT3 decreases TNBC sensitivity to the pan-Akt inhibitor GSK690693 [46]. 2) AKT3 might stimulate post-irradiation cell survival of K-RAS-mutated cells after irradiation. Toulany et al. have revealed that AKT3 stimulates the repair of DNA double-strand breaks in oncogenic K-RAS-mutated cells and promotes BC tumor growth in vivo [21]. 3) AKT3 may contribute to endocrine therapy resistance of ErbB2(+) BC cells with aggressive behavior [47], because AKT3 expression and activity are elevated in ErbB2(+) TNBC cells and tamoxifen resistant BC cells, and activated AKT3 decreases the sensitivity of ErbB2(+) BC cells to tamoxifen (an endocrine therapy used to treat hormone receptor-positive BC).
In contrast to its oncogenic roles, AKT3 has been found to decrease BC cell migration and bone metastasis, but to have no effects on BC tumorigenesis and metastasis [46,48,49,50,51]. Maroulakou et al. have reported that Akt3 gene ablation has no significant inhibitory effects on the development of mammary adenocarcinomas in mouse mammary tumor virus (MMTV)-ErbB2/neu and MMTV-polyoma middle T (PyMT) transgenic mice [48]. Chung et al. have also shown that knockdown of Akt3 increases cell motility but has no effect on proliferation in mouse BC cells [49]. Beyond these observations in mouse models of BC, Hinz et al. have shown that AKT3 activity is elevated in human bone metastatic MDA-MB-231 cells, and AKT3 may decrease the metastatic potential of these bone-seeking BC cells via activation of HER2 and discoidin domain receptor (DDR) kinases, and downregulation of TGFβ [50]. In addition, Lehman et al. have revealed that AKT3 promotes survival of inflammatory BC (IBC, the deadliest form of BC) cells, but has no effect on the invasion of IBC or non-IBC cell lines [51].
In summary, AKT3 does not appear to have pro-oncogenic effects, and it may exert partly contradictory effects, largely depending on the specific BC cell type (Table 2). Nonetheless, emerging studies indicate that miRNAs have critical roles in BC tumorigenesis through interfering with the PI3K/AKT signaling pathway [52]. The regulation of AKT3 expression by miRNAs and the functions of miRNA/AKT3 axes in BC are summarized in Table 3.
Increasing evidence indicates associations of aberrantly expressed miRNAs with BC development and progression, as well as chemoresistance. Several anti-oncogene miRNAs are downregulated in BCs; AKT3 is one of their direct targets, and its expression level is inversely correlated with these miRNAs’ expression levels. These miRNAs include 1) miR-29b, which simultaneously inhibits tumor angiogenesis and tumorigenesis [36]; 2) miR-145, which is downregulated in BC tissues and in docetaxel-resistant BC cells, thus promoting the docetaxel sensitivity of BC cells [53]; 3) miR-29c, which shows progressive loss of expression during TNBC tumorigenesis and has a critical role in the early development of TNBC [54]; 4) miR-181a, which decreases glycolysis in TNBC cells [55]; 5) miR-326, which inhibits BC cell growth and induces apoptosis [56]; 6) miR-433, which decreases BC cell proliferation and cell survival [57]; 7) miR-489, which increases BC chemosensitivity, and suppresses BC cell growth and invasion [58]; and 8) miR-3614-3p, which decreases BC tumor cell invasion and migration [59].
In addition to being directly regulated by miRNAs, AKT3 expression is indirectly regulated by circRNAs and lncRNAs [60,61]. The oncogenic circWHSC1, which is highly expressed in BC tissues and cells, regulates TNBC cell growth, migration, invasion, and survival by sponging the tumor suppressor miR-212-5p, thus promoting AKT3 expression [61]. In BC cell lines and clinical BC tissues, the pseudogene-derived lncRNA RP11-480I12.5 has been found to be overexpressed [60]. Both RP11-480I12.5 and its transcript, RP11-480I12.5-004, exhibit pro-tumorigenic activity through increasing AKT3 expression by competitively binding miR-29c-3p [60].

Table 3.
The ncRNA and miRNA/AKT3 axes in human cancers.
| Tumor tissue/cell lines | LncRNA or circRNA/functions | MiRNAs in the miRNA/AKT3 axis/related functions | Ref. |
|---|---|---|---|
| Breast cancer (BC) | |||
| BC tissues; MDA-MB-231, HUVECs |
miR-29b/ angiogenesis↓ tumorigenesis↓ |
[36] | |
| Docetaxel resistance of BC MCF7, MDA-MB-231, docetaxel resistant cell lines: MCF7/DTX, MDA-MB-231/DTX | miR-145/ cell viability↓, colony formation↓, docetaxel sensitivity↑ |
[53] | |
| MCF10A, MCF10. AT1, MCF10.neoT, CF10. Ca1d, MCF10. Ca1h, MCF10. DCIS | miR-29c/ preneoplastic TNBC cell proliferation↓, colonization ability↓ |
[54] | |
| MDA-MB-231 | miR-181a-5p/ viability↓, migration↓, survival↓, Warburg effect↓ |
[55] | |
| BC tissues; SKBR3, MDA-MB-231, HS578T, MCF7, BT474 |
miR-326/ cell growth↓, colony formation↓, apoptosis↑, migration↓ |
[56] | |
| BC tissues; BT-549, MCF-7, MDA-MB-453, MDA-MB-231 |
miR-433/ cell proliferation↓, cell viability↓, apoptosis↑ |
[57] | |
| Drug-resistant and drug-sensitive BC tumor tissues; MCF-7, MDA-MB-231, MDA-MB-468, T47D |
miR-489/ chemosensitivity↑, cell proliferation↓, invasion↓ |
[58] | |
| MCF-7, MDA-MB-231 | miR-3614-3p/ invasion↓, migration↓ |
[59] | |
| BC tissues; MDA-MB-231 |
circWHSC1/ cell growth↑, proliferation↑, migration↑, invasion↑, glycolysis↑, apoptosis↓ |
miR-212-5p | [61] |
| HCC1937, BT549, MDA-MB-231, MCF-7, T47D, BT474 | RP11-480I12.5-004/ cell proliferation↑, colony formation↑, apoptosis↓ |
miR-29c-3p | [60] |
| Non-small cell lung cancer (NSCLC) | |||
| NSCLC tissues; cell lines: BEAS-2B, A549, HCC823, NCL-H23, NCL-H358 cells |
miR-217/ cell proliferation↓, apoptosis↑ |
[62] | |
| NSCLC tissues; A549 |
circulating miR-320a/ metastatic potential↓, apoptosis↑ |
[63] | |
| NSCLC tissues; NSCLC cell lines: CALU3, CALU6, A549, H1229, H1975 |
circWHSC1/ colony formation↑, viability↑, metastasis↑, progression↑ |
miR-296-3p | [64] |
| NSCLC tissues; NSCLC cell lines: A549 and H460 |
circ_0016760/proliferation↑, migration↑, apoptosis↓ | miR-646 | [65] |
| NSCLC tissues; Cell lines: NCI-H1299, A549, H460, NCI-H2106, H1975 |
circ_0000520/ cell growth↑, migration↑, invasion↑ |
miR-1258 | [66] |
| Hepatocellular carcinoma (HCC) | |||
| HCC specimens; HCC cell lines: HepG2 | miR-122, miR-124/ function not validated |
[67] | |
| HCC tissue samples and cell lines: Huh-7, SNU-182, SNU-475, Hep3B2, HepG2 | miR-122/ cell growth↓, migration↓, apoptosis↑, |
[68] | |
| HCC-BCLC9 cell | miR-122/ cell proliferation↓, dormancy↑ |
[69] | |
| HCC specimens; HepG2, HuH7, SMMC-7721 |
miR-144/ cell proliferation↓, migration↓, invasion↓ |
[70] | |
| HCC tissues of solitary large, nodular, and small HCC; HCC cell lines: SMMC7721, HepG2, HUH7, MHCC97-L, MHCC97-H, HCCLM3 |
miR-424/ cell proliferation↓ |
[71] | |
| HCC specimens; HCC cell lines: QGY-7703, Huh7, BEL-7402, HepG2, Hep3B |
miR-582-5p/ colony formation↓, cell proliferation↓ |
[72] | |
| HCC tissues; HCC cell lines: SNU-449, SNU-182, Huh7, LM3, Bel-7405, SK-hep1, Hep3B | LINC00680/stemness behavior↑, chemosensitivity↓ | miR-568 | [73] |
| HCC specimens; HCC cell lines: HepG2, Hep3B, Huh-7, SNU398, NU449, SNU182, SNU475 |
miR-519d/AKT3? miR-519d/ cell proliferation↑, migration↑, apoptosis↓ |
[74] | |
| Colorectal cancer (CRC) | |||
| CRC cell lines: RKO, HCT116 | miR-124-3p.1/ proliferation↓, metastasis↓ |
[75] | |
| CRC tissues CRC cell lines: SW480, HCT116, LOVO, SW620 |
miR-384/ proliferation↓ |
[76] | |
| CRC tissues; CRC cell lines: Colo205, SW620, HCT116, HT29, LOVO, SW480 |
LINC02163/proliferation↑, metastasis↑ | miR-511-3p | [77] |
| CRC tissues; CRC cell lines: LOVO, PKO, SW480, HT29 |
lncRNA DSCAM-AS1/proliferation↑, invasion↑, migration↑ | miR-384 | [78] |
| Gastric carcinoma (GC) | |||
| GC tissues; cell lines: SGC-7901, MKN45, BGC823 |
miR-195/ apoptosis↑ |
[79] | |
| Gastric adenocarcinoma serum; cell line: MGC-803 |
MALAT1/ apoptosis↓ |
miR-181a-5p | [80] |
| GC tissues; GC cell lines: MKN28, NCI-N87, AGS, KATOIII, RF1, RF48 |
circNF1/cell proliferation↑ | miR-16 | [81] |
| Cholangiocarcinoma (CCA) cell lines: HCCC-9810, RBE | circRNA CDR1a/proliferation↑, invasion↑ | miR-641 | [82] |
|
Pancreatic cancer (PC) tissues; cell line: PANC-1 |
miR-489/proliferation↓, apoptosis↑ | [83] | |
| Ovarian cancer (OC)/epithelial ovarian cancer (EOC) | |||
| EOC tissues; cell lines: SKOV3, A2780, HO8910, 3AO |
miR-29b/ Warburg effect↓, tumor growth↓ |
[84] | |
| OC cell lines: SKOV3, OVCAR3, cisplatin-resistant SKOV and OVCAR3 cells | miR-489/ survival↓, growth↓, apoptosis↑, sensitivity of cisplatin-resistant OC to cisplatin↑ |
[85] | |
| OC tissues; Cell lines: CaOV3, OVCAR3, SKOV3 |
RHPN1-AS1/proliferation↑, migration↑, invasion↑ | miR-665 | [86] |
| OC tissues; OC cell lines: SKOV-3, ES-2, OVCAR3, A2780, CAOV3 |
lncRNA EMX2OS/proliferation↑, invasion↑, tumor growth ↑ | miR-654 | [87] |
| Endometrial carcinoma (EC) | |||
| EC tissues; EC cell line ECC1 |
miR-582-5p/proliferation↓, apoptosis↑ | [88] | |
| Endometrial adenocarcinoma cell line: Ishikawa (ISK) cells | lncCDKN2B-AS1/proliferation↑, invasion↑ | miR-424-5p | [89] |
| EC tissues; EC cell lines: HEC1A, HEC1B, Ishikawa |
LINC01224/ proliferation↑, apoptosis↓ |
miR-485-5p | [90] |
| Thyroid carcinoma (TC)/papillary thyroid carcinoma (PTC) | |||
| TC tissues; cell lines: TPC-1, FTC-133, 8505C; primary PTC cells; |
miR-145/ growth↓, metastasis↓ |
[91] | |
| PTC tissues; PTC cell line: K1 |
miR-29a/ growth↓, apoptosis↑, metastasis↓ |
[92] | |
| TC tissues; PTC cell lines: 8505C, TPC-1, SW1736 |
miR-217/ proliferation↓, migration↓, invasion↓ |
[93] | |
| PTC tissues; PTC cell lines: B-CPAP, KTC-1 |
lncRNA n384546/progression↑, metastasis↑ | miR-145-5p | [94] |
| TC tissues; TC cell lines: BCPAP, K1, H7H83, TPC-1 |
circ_0000144/proliferation↑, migration↑, invasion↑ | miR-217 | [95] |
| Nasopharyngeal carcinoma (NPC) | |||
| NPC tissues; Human primary NPC cell |
miR-424-5p/ proliferation↓, migration↓, apoptosis↑ |
[96] | |
| NPC tissues; NPC cell lines: C666-1, SUNE1, 5-8 F, HNE1, HNE2 |
circTRAF3/proliferation↑, invasion↑, apoptosis↓ | miR-203a-3p | [97] |
|
Oral squamous cell carcinoma (OSCC) tissues; cell line: SCC-4, SCC-25, HN-6, CAL-27, TCA-83 |
miR-16/ proliferation↓, apoptosis↑ |
[98] | |
| Glioblastoma multiforme (GBM) | |||
| GBM cell lines: T98G, U87, A172, LN229, LN18 | miR-610/ proliferation↓, anchorage independent growth↓ |
[99] | |
| GBM cell lines: LN229, A172, U373, SHG44 | lncRNA, GAS5/proliferation↓, migration↓, invasion↓ | miR-424 | [100] |
| Multiple myeloma (MM) | |||
| MM cell lines: OPM2, RPMI-8226; Endothelial cell: HUVECs |
miR-29b/ endothelial cell proliferation↓, migration↓, tube formation↓ |
[101] | |
| MM tissues; Cell lines: KM3, H929, MM1S, U266 cells |
circ_0000142/proliferation↑, metastasis↑ | miR-610 | [102] |
| MM tissues; cell lines: OPM-2, U266, KM3, U1996, H929 |
lncRNA FEZF1-AS1/ proliferation↑, apoptosis↓ |
miR-610 | [103] |
| Osteosarcoma (OS) | |||
| OS tissues; cell lines: HOS, MG-63, Saos-2, SW1353, U2OS |
miR-1258/ proliferation↓ |
[104] | |
| OS tissues; OS cell lines: HOS, MG-63, SaOS-2, U2OS, |
MALAT1/ glycolysis↑, proliferation↑, metastasis↑ |
miR-485-3p | [105] |
|
Uveal melanoma (UM) tissues; UM cell line: OCM-1A |
miR-224-5p/ proliferation↓, migration↓, invasion↓ |
[106] | |
| UM cell lines: OMM2.5, UM001, Mel285, Mel290; UM xenografts |
miR-181a-5p/ proliferation↓, colony formation↓, apoptosis↑, tumor growth ↓ |
[107] | |
|
NK/T cell lymphoma (NKTL) tissues; cell lines: KHYG-1, NK-92, HANK-1, SNK-1, SNK-6 |
miR-150/ sensitivity of NKTL to radiation treatment↑ |
[108] | |
| Bladder cancer TCGA database | miR-195/ cell proliferation↓ |
[109] | |
|
Wilms’ tumor tissues; cell lines: 17-94, WIT49 |
miR-22-3p/ proliferation↓, invasion↓ |
[110] |
3.2. Lung cancer
Lung cancer is the second most common cancer worldwide and the leading cause of cancer death. Approximately 80–85% of lung cancer is non-small cell lung cancer (NSCLC). AKT3 has been found to have high activity in NSCLC cells, thereby promoting proliferation, survival, and migration [111]. Emerging studies indicate that AKT3 expression is regulated by several miRNAs that are downregulated in NSCLC (Table 3). People with NSCLC with lower levels of miR-217 have shorter overall survival. Overexpression of miR-217 inhibits NSCLC cell proliferation and induces apoptosis [62]. Circulating miR-320a levels are relatively low in the plasma of people with NSCLC; these low levels are correlated with clinicopathological characteristics, such as tumor size, tumor stage, and lymph node metastasis. Circulating miR-320a functions as a tumor-suppressor miRNA that decreases metastatic potential and increases apoptosis of NSCLC cells [63]. Notably, three circRNAs are upregulated in NSCLC and consequently promote NSCLC cell growth and metastasis: circWHSC1, circ_0016760, and circ_0000520. These circRNAs act as ceRNAs, thereby increasing AKT3 expression by directly targeting miR-296-3p, miR-646, and miR-1258, respectively, in NSCLC cells [64,65,66].
3.3. Digestive/gastrointestinal cancers
Hepatocellular carcinoma (HCC), the most frequently occurring liver malignancy, has high rates of fatality, recurrence, and chemotherapeutic resistance. Deregulation and activation of the AKT signaling pathway is common in HCC and is associated with poor patient prognosis. AKT3 expression is upregulated in the HCC cell lines SNU-182, Hep3B2, and SNU-475 [68]. Furthermore, a growing number of HCC-associated genes are being found to be regulated by ncRNAs. For example, a panel of serum miRNAs comprising miR-26a-5p, miR-122-5p, miR-141-3p, miR-192-5p, miR-199a-5p, miR-206, miR-433-3p, and miR-1228-5p has shown clinical value in HCC diagnosis [112]. The oncogenic lncRNA LINC00680 is markedly upregulated in HCC tissues. LINC00680 acts as a ceRNA sponging miR-568, thus activating AKT3, enhancing HCC stemness behavior, and decreasing chemosensitivity to 5-fluorouracil (a highly effective classical chemotherapeutic agent in the treatment of HCC) [73]. Moreover, Zhang et al., in a comprehensive analysis of the miRNA-regulated protein interaction network, have identified a list of miRNAs targeting AKTs, which includes miR-122/AKT3 and miR-124/AKT3 interactions, thus suggesting the critical roles of these miRNAs in HCC malignant progression [67]. Further studies have validated that several tumor suppressor miRNAs that directly suppress AKT3 expression are lost or downregulated in HCC cells (Table 3). MiR-122, the most highly expressed miRNA in the healthy adult liver, is responsible for liver stem cell differentiation toward the hepatocyte lineage. Its expression is frequently lost in HCC tissues and HCC cell lines [68,69]. Through binding the 3′-UTR of AKT3 and controlling AKT3 gene expression, miR-122 inhibits HCC cell proliferation, increases the chemosensitivity of HCC cells, and attenuates HCC tumor growth in vivo [68,69]. Forced overexpression of miR-144, miR-424, or miR-582-5p in HCC cells results in anti-HCC effects, by suppressing cell proliferation, migration, invasion, or survival [70,71,72]. Notably, miR-519d is upregulated in HCC and contributes to hepatocarcinogenesis after anticancer treatments [74]. Although AKT3 is known to be directly targeted by oncogenic miR-519d, the pathophysiological role of AKT3 in HCC remains unclear.
CRC is a common digestive malignancy in females and males worldwide. The initiation and malignant progression of CRC is a long-term multi-stage process involving genetic and epigenetic alterations. Oncogenic GLUT5 expression in tumor tissue is associated with aggressive behavior and invasiveness in cancer cells, and also regulates migratory activity in drug-resistant CRC cells [113]. In CRC, AKT3 contributes to drug resistance by aberrantly downregulating miR-125b-5p expression, thus leading to expression of the glucose transporter GLUT5. In addition, AKT3 expression in CRC cells is regulated by miR-124-3p.1 [75] and miR-384 [76]. Both miR-124-3p.1 and miR-384 exhibit anti-CRC effects by inhibiting cell proliferation [75,76]. Moreover, two lncRNAs, LINC02163 and Down syndrome cell adhesion molecule antisense1 (DSCAM-AS1), have shown oncogenic effects in CRC development and progression [77,78]. Their expression is upregulated in CRC, and is associated with tumor metastasis and poor prognosis in people with CRC. LINC02163 and DSCAM-AS1 function as ceRNAs that target miR-511-3p and miR-384, respectively, and subsequently induce AKT3 expression [77,78] (Table 3).
GC, the third leading cause of cancer deaths worldwide, is usually diagnosed in advanced stages. The miR-195/AKT3 axis has a critical role in GC development. The transcription factor early growth response 1 (EGR1) functions as an oncogene in GC through suppressing apoptosis of GC cells via directly inhibiting the expression of miR-195 and activating AKT3 [79]. Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1), also known as non-coding nuclear-enriched abundant transcript 2 (NEAT2), promotes gastric adenocarcinoma through the MALAT1/miR-181a-5p/AKT3 axis [80]. High levels of MALAT1 are detected in the serum in people with gastric adenocarcinoma. MALAT1 regulates GC cell proliferation and apoptosis by decreasing the expression of miR-181a-5p, which in turn upregulates AKT3 protein levels [80]. On the basis of its high expression in GC tissues and cell lines, circNF1 has been identified as an oncogenic circRNA in GC, which promotes GC cell proliferation, binds miR-16, and consequently derepresses its downstream target AKT3 [81].
Cholangiocarcinoma (CCA, also known as bile duct cancer), is one of the most common hepatic malignancies and accounts for 3% of all gastrointestinal cancers. CCA is difficult to diagnose in early stages, and, because of early metastasis, its 5-year survival rates are only 20–40%. The upregulated circRNA cerebellar degeneration-associated protein 1 antisense (CDR1as, also known as cirs-7) has oncogenic roles in CCA. CDR1as binds miR-641, and subsequently accelerates miR-641 degradation and may possibly lead to the upregulation of AKT3 expression [82].
Pancreatic cancer (PC) is a high-grade malignancy of the digestive system with vague clinical features in early stages. The 5-year overall survival rate of PC is extremely low, at less than 5%. The level of miR-489 is markedly low in PC. Upregulation of miR-489 significantly inhibits cell proliferation and induces cell apoptosis of PC PANC-1 cells through targeting AKT3 [83].
3.4. Gynecologic cancers
Ovarian cancer (OC) is the most lethal gynecological cancer, accounting for 5% of female cancer cases and deaths worldwide. Epithelial ovarian cancer (EOC) accounts for ~90% of all ovarian malignancies. In EOC cells, AKT3 expression is negatively regulated by downregulated miR-29b, thus inducing the rate-limiting glycolytic genes hexokinase 2 and pyruvate kinase M2, and increasing the Warburg effect and ovarian cancer progression [84]. A role of AKT3 in cisplatin resistance of OC cells has been suggested, because MK-2206 2HCl, an AKT3 inhibitor, increases the sensitivity of cisplatin-resistant OC cells to cisplatin [85]. However, miR-489, which regulates AKT3 expression, is downregulated in OC cells. MiR-489 overexpression increases the sensitivity of cisplatin-resistant OC cells to cisplatin by inhibiting cell growth and promoting apoptosis [85]. Expression of the oncogenic lncRNA RHPN1-AS1, a 2030 bp transcript from human chromosome 8q24, is upregulated in OC tissues and cell lines. RHPN1-AS1 promotes OC cell proliferation and migration via the miR-665/AKT3 axis [86]. Another oncogenic lncRNA, EMX2OS, is overexpressed in OC tissues, and directly suppresses miR-654 expression, thus leading to the upregulation of AKT3; therefore, the EMX2OS/miR-654/AKT3 axis may confer aggressiveness in OC [87].
Endometrial carcinoma (EC) is a common cancer of the female genital tract. Invasion and recurrence contribute to the prognosis and survival rate of patients with EC. AKT3 is involved in the regulation of cell proliferation and the invasion of endometrial stromal cells [89]. AKT3 expression is regulated by the tumor inhibitor miR-582-5p, whose expression is significantly diminished in human EC tissues. MiR-582-5p strongly inhibits cell proliferation and promotes apoptosis of EC cells [88]. The oncogenic lncRNA CDKN2B-AS1 has been observed in Ishikawa endometrial adenocarcinoma cells, and found to promote cellular proliferation and invasion by sponging miR-424-5p, thus upregulating the expression of AKT3 [89]. High levels of the lncRNA LINC01224 have been found in both EC tumor tissue and cell lines. LINC01224 promotes EC cell proliferation and inhibits apoptosis by elevating expression of AKT3 via targeting miR-485-5p [90].
3.5. Thyroid carcinoma and head and neck cancers
Papillary thyroid cancer (PTC), the most common histological type of thyroid carcinoma (TC), accounts for more than 85% of TC cases. Environmental exposure and genetic mutation are the risk factors for PTC. MiR-145, miR-29a, and miR-217 are downregulated in TC/TPC. Their tumor suppressor roles have been demonstrated to involve direct inhibition of AKT3 in PTC cells [91,92,93]. In agreement with these findings, miR-145-5p is a key miRNA target of the oncogenic lncRNA n384546, whose expression is elevated and associated with clinicopathological features in patients with PTC. Moreover, n384546 promotes TPC progression and metastasis by acting as a ceRNA of miR-145-5p, thereby regulating AKT3 [94]. In addition, circ_0000144, an oncogenic circRNA, is markedly elevated in TC tissues and is associated with tumor size, TNM stage, and lymph node metastasis in people with TC. Circ_0000144 shows cancer-promoting effects on TC cells via regulating the miR-217/AKT3 pathway [95].
Nasopharyngeal carcinoma (NPC) is a common head and neck cancer with high incidence in southern China, North Africa, and Southeast Asia. As with most tumors, genetic abnormalities are closely associated with the occurrence of NPC. MiR-424-5p is a tumor-associated miRNA encoded at human Xq26.3. In NPC, miR-424-5p expression is downregulated, and is associated with lymph node metastasis and clinical staging. MiR-424-5p exhibits anti-oncogenic activities by inhibiting the proliferation, migration, and invasion of NPC cells through decreasing AKT3 expression [96]. The circRNA circTRAF3 is highly expressed in NPC, and is associated with metastasis and survival in people with NPC. CircTRAF3 promotes NPC cell proliferation and metastasis through eliminating the inhibitory effect of miR-203a-3p on AKT3 expression [97].
Oral squamous cell carcinoma (OSCC) is the most prevalent subgroup of head and neck cancer and the most common type of oral cancer. AKT3 is directly targeted by miR-16, and its expression is negatively associated with that of miR-16 in OSCC. MiR-16 is dysregulated in OSCC [98], and it functions as a tumor suppressor miRNA, thus inhibiting cell proliferation and inducing apoptosis in OSCC through decreasing AKT3 [98].
3.6. Other types of human cancer
GBM is the most common and lethal primary brain malignancy. People with GBM have poor prognosis and survival. AKT activation is found in approximately 80% of human GBMs. AKT3 is overexpressed in glioma cells and has been found to play a critical role in GBM [114]. Studies have shown that siRNA knockdown of AKT3 in GBM T98G cells significantly decreases cell viability, proliferation, invasion, and metastasis [34]. MiR-610 is downregulated in GBM, and it directly suppresses AKT3 expression, thus decreasing the proliferation and anchorage independent growth of GBM cells [99]. Interestingly, lncRNA growth arrest-specific 5 (GAS5) functions as a tumor suppressor in glioma cells by alleviating promoter methylation of miR-424, and consequently increasing the expression of miR-424, and suppressing AKT3 and its targets, cyclinD1, c-Myc, Bax, and Bcl-2 [100].
Multiple myeloma (MM), the second most common hematological malignancy, is characterized by high infiltration and multifocal proliferation of malignant plasma cells in the bone marrow. The miR-29b/AKT3 axis has been found to be involved in the progression of MM by regulating the angiogenic activity of MM-derived exosomes [101]. In endothelial cells, after treatment with exosomes released from MM cells treated with C6-ceramide, expression of the tumor suppressor miR-29b is induced, but AKT3 is decreased; consequently, endothelial cell proliferation, migration, and angiogenesis are suppressed [101]. MiR-610 functions as a tumor suppressor and suppresses AKT3 in several human cancers. Both circ_0000142 and lncRNA FEZF1-AS1 are upregulated and associated with poor prognosis in people with MM, and promote MM cell growth though modulating the miR-610/AKT3 axis [102,103].
OS, an extremely malignant primary bone cancer with rapid progression, affects both children and adolescents. The prognosis of OS is poor, owing to its high tendency toward lung metastasis. AKT3 is upregulated in OS and is associated with OS progression; its expression is negatively associated with the expression of miR-1258 and miR-485-3p in OS tissues [104,105]. MiR-1258 has been widely studied in various cancers including OS. Upregulation of miR-1258 significantly inhibits OS cell growth by binding the 3′-UTR of AKT3 [104]. Nonetheless, the anti-oncogene miR-485-3p, with inhibitory effects on glycolysis and metastasis of OS cells, is downregulated in OS by the lncRNA MALAT1 [105].
Uveal melanoma (UM) is a rare intraocular malignancy in adults, which arises from melanocytes in the iris, ciliary body, or choroid. In UM tissues, miR-224-5p expression is low, whereas AKT3 expression is high. MiR-224-5p is involved in the proliferation, invasion, and migration of UM cells, partially through regulation of the expression of AKT3 [106]. Of note, our research team has recently discovered that targeting AKT3 in UM cells might be a mechanism underlying the inhibitory effect of miR-181a-5p on UM development [107].
NK/T cell lymphoma is a rare non-Hodgkin lymphoma with high invasive malignancy, putative NK-cell origin, and poor prognosis. MiR-150 increases the sensitivity of NK/T cell lymphoma to ionizing radiation through targeting AKT3, but it is significantly diminished in NK/T cell lymphoma tissues and cell lines [108].
Bladder cancer is a relatively rare, highly malignant tumor arising from urinary bladder tissues. AKT3 may participate in the proliferation and apoptosis of bladder cancer cells, and it is an important target of miR-195. MiR-195 functions anti-cancer roles by suppressing glucose uptake and proliferation of bladder cancer cells [109].
Wilms’ tumor, or nephroblastoma, the most frequent renal cancer in children, occurs mainly in the first 5 years after birth. AKT3 is upregulated in Wilms’ tumors, whereas miR-22-3p is downregulated. MiR-22-3p regulates the proliferation and invasion of Wilms’ tumor cells through inhibition of AKT3 [110].
4. Dysregulation of the same miRNA/AKT3 axis in different cancers
As discussed above, in a specific type of human cancer, different dysregulated ncRNA/AKT3 axes are involved in tumorigenesis, progression, or drug resistance (Table 3). However, studies have also identified the same disrupted miRNA/AKT3 axis in different cancer types (Figure 2). For instance, the miR-145/AKT3 axis is found in several human cancers including BC, chemo-resistant BC, and PTC [53,91,94]. Furthermore, the miR-181a-5p/AKT3 axis plays important roles in gastric adenocarcinoma, TNBC, and UM [55,80,107]. In addition, the miR-217/AKT3 axis is dysregulated in TC and NSCLC [62,93,95]; the miR-424-5p/AKT3 axis is dysregulated in HCC, NPC, endometrial adenocarcinoma, and glioma [71,89,96,100]; the miR-489/AKT3 axis is dysregulated in drug-resistant BC, cisplatin-resistant OC cells, and PC [58,83,85]; and the miR-610/AKT3 axis is dysregulated in GBM and MM [99,102,103]. Notably, the miR-29 family consists of miR-29a, miR-29b-1, miR-29b-2, and miR-29c, which are involved in many pathophysiological processes, and are associated with cancer development and metastasis [115]. The tumor suppressor role of miR-29b has also been reviewed [116]. Abnormal miR-29/AKT3 axes are found in certain types of human cancer, e.g., miR-29a/AKT3 in thyroid cancer [92]; miR-29b/AKT3 in BC, EOC, and MM [36,84,101]; miR-29c/AKT3 in TNBC [54]; and miR-29c-3p/AKT3 in BC [60]. Together, these findings suggest that a single miRNA may be used as an efficient anti-cancer therapeutic agent for different types of human cancer [36].
5. Conclusion
Despite substantial efforts to develop effective strategies, the outcomes for patients with highly malignant cancer remain unsatisfactory. Studying the potential mechanisms involved in the occurrence and progression of aggressive cancers is crucial to explore novel targets for disease diagnosis and treatment. Given the central role of AKT signaling in the pathogenesis and development of cancer, discovering the exact mechanisms controlling the AKT signaling activity, and understanding how the inhibition of this pathway influences major cellular processes, is critical. Exciting findings have recently indicated that ncRNAs function through molecular associations with the components of classical signaling pathways including the AKT signaling pathway. Many ncRNAs involved in this pathway have been identified and extensively studied. Dysregulated miRNAs and other ncRNAs are gradually becoming accepted as biomarkers in cancer diagnosis and as potential targets in cancer therapy. In particular, the miRNA/AKT3 axis is attracting increasing attention.
Currently, available prediction tools and databases together with microarray techniques and second-generation sequencing readily enable the examination of groups of ncRNAs with sequence complementarity to the 3′-UTR of AKT3. Through integrating the ncRNA profile and miRNA target prediction tools, links between ncRNAs and AKT3 can be established and validated in cancer cells. Hence, many miRNAs that directly bind the 3′-UTR of AKT3 and participate in the regulation of AKT signaling in human cancers have been revealed, and additional AKT3-associated ncRNAs will be identified in the near future. The findings regarding miRNA/AKT3 regulatory networks may revolutionize views regarding the genesis, progression, and treatment of cancer. Because ncRNAs positively or negatively regulate the biological functions of AKT3 in cancer development and progression, regulating ncRNAs in the miRNA/AKT3 axis to control cancer may be a therapeutic option. For developing drugs that stably regulate ncRNA activity and efficiently transduce effects, in-depth studies of the structures and functions of miRNAs associated with the AKT3 pathway will be essential. In addition, AKT3-derived circRNAs target some miRNAs in certain types of cancers, suggesting that AKT3 may regulate epigenetic modifiers in a post-transcriptional manner. However, limited information is available regarding AKT3-driven miRNA alterations. Therefore, analyzing miRNA reprogrammed AKT3 profiles and AKT3 regulated miRNA profiles to explore cancer pathogenesis in a personalized, or cancer-specific manner may advance the development of effective new therapeutic strategies in the future. Comprehensive investigation will ideally allow approaches to specifically inhibit AKT3 signaling be identified and applied in routine clinical practice.
Author Contributions
Writing—original draft preparation, C.Y.; writing—review and editing, C.Y. and P.H.; supervision, P.H.; funding acquisition, P.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Canadian Institutes of Health Research under an operating grant to P. Hardy (475599).
Informed Consent Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
List of abbreviations
| Abbreviation | Full name |
| 3′-UTRs | 3′-untranslated region |
| 5-Fu | 5-fluorouracil |
| CCA | cholangiocarcinoma |
| ccRCC | clear cell renal cell carcinoma |
| ceRNA | competitive endogenous RNA |
| circRNAs | circular RNAs |
| CRC | colorectal cancer |
| DDR | discoidin domain receptors |
| DNA-PKcs | DNA-dependent protein kinase catalytic subunit |
| DSBs | DNA double-strand breaks |
| EC | endometrial carcinoma |
| EOC | epithelial ovarian cancer |
| ERα | estrogen receptor alpha |
| GAS5 | growth arrest-specific 5 |
| GBM | glioblastoma multiforme |
| GC | gastric cancer |
| HCC | hepatocellular carcinoma |
| HER2 | ErbB2 receptor tyrosine kinase 2 |
| HK2 | hexokinase2 |
| HUVEC | human umbilical vein endothelial cells |
| IBC | inflammatory breast cancer |
| K-RASmut | K-RAS-mutated |
| lncRNAs | long non-coding RNAs |
| MALAT-1 | metastasis-associated lung adenocarcinoma transcript 1 |
| miRNA, miR- | microRNA |
| MM | multiple myeloma |
| ncRNAs | non-coding RNAs |
| NKTL | NK/T cell lymphoma |
| NPC | nasopharyngeal carcinoma |
| NSCLC | non-small cell lung cancer |
| OC | ovarian cancer |
| OS | osteosarcoma |
| OSCC | oral squamous cell carcinoma |
| PC | pancreatic cancer |
| PIK3R3 | Phosphoinositide-3-Kinase Regulatory Subunit 3 |
| PKM2 | pyruvate kinase isozyme M2 |
| PTC | papillary thyroid carcinoma |
| RCC | renal cell carcinoma |
| TGFβ | transforming growth factor β |
| TNBC | triple-negative breast cancer |
| ULM | uterine leiomyomas |
| UM | uveal melanoma |
| UTR | untranslated region |
References
- Cardone MH: Roy N, Stennicke HR, Salvesen GS, Franke TF, Stanbridge E et al. Regulation of cell death protease caspase-9 by phosphorylation. Science 1998; 282: 1318-1321. [CrossRef]
- Hinz N, Jücker M. Distinct functions of AKT isoforms in breast cancer: A comprehensive review. Cell Communication and Signaling 2019; 17: 154. [CrossRef]
- Jeong SH, Yang MJ, Choi S, Kim J, Koh GY. Refractoriness of STING therapy is relieved by AKT inhibitor through effective vascular disruption in tumour. Nat Commun 2021; 12: 4405. [CrossRef]
- Bilanges B, Posor Y, Vanhaesebroeck B. PI3K isoforms in cell signalling and vesicle trafficking. Nat Rev Mol Cell Biol 2019; 20: 515-534. [CrossRef]
- Yang Q, Jiang W, Hou P. Emerging role of PI3K/AKT in tumor-related epigenetic regulation. Semin Cancer Biol 2019; 59: 112-124. [CrossRef]
- Li J, Yen C, Liaw D, Podsypanina K, Bose S, Wang SI et al. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science 1997; 275: 1943-1947. [CrossRef]
- Xu M, Mo YY. The Akt-associated microRNAs. Cell Mol Life Sci 2012; 69: 3601-3612. [CrossRef]
- Costa FF. Non-coding RNAs: New players in eukaryotic biology. Gene 2005; 357: 83-94. [CrossRef]
- Selvakumar SC, Preethi KA, Sekar D. MicroRNAs as important players in regulating cancer through PTEN/PI3K/AKT signalling pathways. Biochim Biophys Acta Rev Cancer 2023; 1878: 188904. [CrossRef]
- Ebbesen KK, Hansen TB, Kjems J. Insights into circular RNA biology. RNA Biol 2017; 14: 1035-1045. [CrossRef]
- Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao L, Zhang Y et al. The Landscape of Circular RNA in Cancer. Cell 2019; 176: 869-881.e813. [CrossRef]
- Hansen TB, Jensen TI, Clausen BH, Bramsen JB, Finsen B, Damgaard CK et al. Natural RNA circles function as efficient microRNA sponges. Nature 2013; 495: 384-388. [CrossRef]
- Han B, Chao J, Yao H. Circular RNA and its mechanisms in disease: From the bench to the clinic. Pharmacol Ther 2018; 187: 31-44. [CrossRef]
- Xue C, Li G, Lu J, Li L. Crosstalk between circRNAs and the PI3K/AKT signaling pathway in cancer progression. Signal Transduction and Targeted Therapy 2021; 6: 400. [CrossRef]
- Statello L, Guo C-J, Chen L-L, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nature Reviews Molecular Cell Biology 2021; 22: 96-118. [CrossRef]
- Mattick JS, Amaral PP, Carninci P, Carpenter S, Chang HY, Chen L-L et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nature Reviews Molecular Cell Biology 2023. [CrossRef]
- Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: Insights into functions. Nat Rev Genet 2009; 10: 155-159. [CrossRef]
- Ghafouri-Fard S, Askari A, Hussen BM, Taheri M, Mokhtari M. A long non-coding RNA with important roles in the carcinogenesis. Front Cell Dev Biol 2022; 10: 1037149. [CrossRef]
- Coffer PJ, Woodgett JR. Molecular cloning and characterisation of a novel putative protein-serine kinase related to the cAMP-dependent and protein kinase C families. Eur J Biochem 1991; 201: 475-481. [CrossRef]
- Nicholson KM, Anderson NG. The protein kinase B/Akt signalling pathway in human malignancy. Cell Signal 2002; 14: 381-395. [CrossRef]
- Toulany M, Maier J, Iida M, Rebholz S, Holler M, Grottke A et al. Akt1 and Akt3 but not Akt2 through interaction with DNA-PKcs stimulate proliferation and post-irradiation cell survival of K-RAS-mutated cancer cells. Cell death discovery 2017; 3: 1-10. [CrossRef]
- Basu A, Lambring CB. Akt Isoforms: A Family Affair in Breast Cancer. Cancers (Basel) 2021; 13. [CrossRef]
- Chen WS, Xu PZ, Gottlob K, Chen ML, Sokol K, Shiyanova T et al. Growth retardation and increased apoptosis in mice with homozygous disruption of the Akt1 gene. Genes Dev 2001; 15: 2203-2208. [CrossRef]
- Cho H, Mu J, Kim JK, Thorvaldsen JL, Chu Q, Crenshaw EB, 3rd et al. Insulin resistance and a diabetes mellitus-like syndrome in mice lacking the protein kinase Akt2 (PKB beta). Science 2001; 292: 1728-1731. [CrossRef]
- Tschopp O, Yang ZZ, Brodbeck D, Dummler BA, Hemmings-Miesk M, Watanabe T et al. Essential role of protein kinase B gamma (PKB gamma/Akt3) in postnatal brain development but not in glucose homeostasis. Development 2005; 132: 2943-2954. [CrossRef]
- Wainstein E, Maik-Rachline G, Blenis J, Seger R. AKTs do not translocate to the nucleus upon stimulation but AKT3 can constitutively signal from the nuclear envelope. Cell Rep 2022; 41: 111733. [CrossRef]
- Nakatani K, Thompson DA, Barthel A, Sakaue H, Liu W, Weigel RJ et al. Up-regulation of Akt3 in estrogen receptor-deficient breast cancers and androgen-independent prostate cancer lines. J Biol Chem 1999; 274: 21528-21532. [CrossRef]
- Risso G, Blaustein M, Pozzi B, Mammi P, Srebrow A. Akt/PKB: One kinase, many modifications. Biochem J 2015; 468: 203-214. [CrossRef]
- Konishi H, Kuroda S, Tanaka M, Matsuzaki H, Ono Y, Kameyama K et al. Molecular cloning and characterization of a new member of the RAC protein kinase family: Association of the pleckstrin homology domain of three types of RAC protein kinase with protein kinase C subspecies and beta gamma subunits of G proteins. Biochem Biophys Res Commun 1995; 216: 526-534. [CrossRef]
- Davies MA, Stemke-Hale K, Tellez C, Calderone TL, Deng W, Prieto VG et al. A novel AKT3 mutation in melanoma tumours and cell lines. Br J Cancer 2008; 99: 1265-1268. [CrossRef]
- Yeganeh PN, Richardson C, Bahrani-Mostafavi Z, Tait DL, Mostafavi MT. Dysregulation of AKT3 along with a small panel of mRNAs stratifies high-grade serous ovarian cancer from both normal epithelia and benign tumor tissues. Genes Cancer 2017; 8: 784-798. [CrossRef]
- Madhunapantula SV, Robertson GP. Targeting protein kinase-b3 (akt3) signaling in melanoma. Expert Opin Ther Targets 2017; 21: 273-290. [CrossRef]
- Lin F-M, Yost SE, Wen W, Frankel PH, Schmolze D, Chu P-G et al. Differential gene expression and AKT targeting in triple negative breast cancer. Oncotarget 2019; 10: 4356. [CrossRef]
- Joy A, Kapoor M, Georges J, Butler L, Chang Y, Li C et al. The role of AKT isoforms in glioblastoma: AKT3 delays tumor progression. J Neurooncol 2016; 130: 43-52. [CrossRef]
- Lu S, Chen L, Tang L. Upregulation of AKT1 and downregulation of AKT3 caused by dysregulation of microRNAs contributes to pathogenesis of hemangioma by promoting proliferation of endothelial cells. J Cell Physiol 2019; 234: 21342-21351. [CrossRef]
- Li Y, Cai B, Shen L, Dong Y, Lu Q, Sun S et al. MiRNA-29b suppresses tumor growth through simultaneously inhibiting angiogenesis and tumorigenesis by targeting Akt3. Cancer Lett 2017; 397: 111-119. [CrossRef]
- Xu Y, Jiang T, Wu C, Zhang Y. CircAKT3 inhibits glycolysis balance in lung cancer cells by regulating miR-516b-5p/STAT3 to inhibit cisplatin sensitivity. Biotechnol Lett 2020; 42: 1123-1135. [CrossRef]
- Huang X, Li Z, Zhang Q, Wang W, Li B, Wang L et al. Circular RNA AKT3 upregulates PIK3R1 to enhance cisplatin resistance in gastric cancer via miR-198 suppression. Mol Cancer 2019; 18: 71. [CrossRef]
- Li H, Xu W, Xia Z, Liu W, Pan G, Ding J et al. Hsa_circ_0000199 facilitates chemo-tolerance of triple-negative breast cancer by interfering with miR-206/613-led PI3K/Akt/mTOR signaling. Aging (Albany NY) 2021; 13: 4522-4551. [CrossRef]
- Xue D, Wang H, Chen Y, Shen D, Lu J, Wang M et al. Circ-AKT3 inhibits clear cell renal cell carcinoma metastasis via altering miR-296-3p/E-cadherin signals. Mol Cancer 2019; 18: 151. [CrossRef]
- Xia X, Li X, Li F, Wu X, Zhang M, Zhou H et al. A novel tumor suppressor protein encoded by circular AKT3 RNA inhibits glioblastoma tumorigenicity by competing with active phosphoinositide-dependent Kinase-1. Mol Cancer 2019; 18: 131. [CrossRef]
- Rahmani F, Ferns GA, Talebian S, Nourbakhsh M, Avan A, Shahidsales S. Role of regulatory miRNAs of the PI3K/AKT signaling pathway in the pathogenesis of breast cancer. Gene 2020; 737: 144459. [CrossRef]
- Turner KM, Sun Y, Ji P, Granberg KJ, Bernard B, Hu L et al. Genomically amplified Akt3 activates DNA repair pathway and promotes glioma progression. Proc Natl Acad Sci U S A 2015; 112: 3421-3426. [CrossRef]
- Banerji S, Cibulskis K, Rangel-Escareno C, Brown KK, Carter SL, Frederick AM et al. Sequence analysis of mutations and translocations across breast cancer subtypes. Nature 2012; 486: 405-409. [CrossRef]
- Suyama K, Yao J, Liang H, Benard O, Loudig OD, Amgalan D et al. An Akt3 Splice Variant Lacking the Serine 472 Phosphorylation Site Promotes Apoptosis and Suppresses Mammary Tumorigenesis. Cancer Res 2018; 78: 103-114. [CrossRef]
- Chin YR, Yoshida T, Marusyk A, Beck AH, Polyak K, Toker A. Targeting Akt3 signaling in triple-negative breast cancer. Cancer Res 2014; 74: 964-973. [CrossRef]
- Grabinski N, Möllmann K, Milde-Langosch K, Müller V, Schumacher U, Brandt B et al. AKT3 regulates ErbB2, ErbB3 and estrogen receptor α expression and contributes to endocrine therapy resistance of ErbB2(+) breast tumor cells from Balb-neuT mice. Cell Signal 2014; 26: 1021-1029. [CrossRef]
- Maroulakou IG, Oemler W, Naber SP, Tsichlis PN. Akt1 ablation inhibits, whereas Akt2 ablation accelerates, the development of mammary adenocarcinomas in mouse mammary tumor virus (MMTV)-ErbB2/neu and MMTV-polyoma middle T transgenic mice. Cancer Res 2007; 67: 167-177. [CrossRef]
- Chung S, Yao J, Suyama K, Bajaj S, Qian X, Loudig OD et al. N-cadherin regulates mammary tumor cell migration through Akt3 suppression. Oncogene 2013; 32: 422-430. [CrossRef]
- Hinz N, Baranowsky A, Horn M, Kriegs M, Sibbertsen F, Smit DJ et al. Knockdown of AKT3 Activates HER2 and DDR Kinases in Bone-Seeking Breast Cancer Cells, Promotes Metastasis In Vivo and Attenuates the TGFβ/CTGF Axis. Cells 2021; 10. [CrossRef]
- Lehman HL, Van Laere SJ, van Golen CM, Vermeulen PB, Dirix LY, van Golen KL. Regulation of inflammatory breast cancer cell invasion through Akt1/PKBα phosphorylation of RhoC GTPase. Mol Cancer Res 2012; 10: 1306-1318. [CrossRef]
- Dabi Y, Bendifallah S, Suisse S, Haury J, Touboul C, Puchar A et al. Overview of non-coding RNAs in breast cancers. Transl Oncol 2022; 25: 101512. [CrossRef]
- Zhou Y, Cai W, Lu H. Overexpression of microRNA-145 enhanced docetaxel sensitivity in breast cancer cells via inactivation of protein kinase B gamma-mediated phosphoinositide 3-kinase -protein kinase B pathway. Bioengineered 2022; 13: 11310-11320. [CrossRef]
- Bhardwaj A, Singh H, Rajapakshe K, Tachibana K, Ganesan N, Pan Y et al. Regulation of miRNA-29c and its downstream pathways in preneoplastic progression of triple-negative breast cancer. Oncotarget 2017; 8: 19645-19660. [CrossRef]
- Wang Y, Tahiri H, Yang C, Gu M, Ruan X, Hardy P. Overexpression of miR-181a regulates the Warburg effect in triple-negative breast cancer. Climacteric 2023; 26: 64-71. [CrossRef]
- Ghaemi Z, Soltani BM, Mowla SJ. MicroRNA-326 Functions as a Tumor Suppressor in Breast Cancer by Targeting ErbB/PI3K Signaling Pathway. Front Oncol 2019; 9: 653. [CrossRef]
- Hu X, Wang J, He W, Zhao P, Ye C. MicroRNA-433 targets AKT3 and inhibits cell proliferation and viability in breast cancer. Oncol Lett 2018; 15: 3998-4004. [CrossRef]
- Chen X, Wang YW, Xing AY, Xiang S, Shi DB, Liu L et al. Suppression of SPIN1-mediated PI3K-Akt pathway by miR-489 increases chemosensitivity in breast cancer. J Pathol 2016; 239: 459-472. [CrossRef]
- Wang Z, Jing X, Li F, Chen Y, Huang C. miR-3614-3p suppresses cell aggressiveness of human breast cancer by targeting AKT3 and HDAC1 expression. Transl Cancer Res 2022; 11: 1565-1575. [CrossRef]
- Lou W, Ding B, Zhong G, Yao J, Fan W, Fu P. RP11-480I12.5-004 Promotes Growth and Tumorigenesis of Breast Cancer by Relieving miR-29c-3p-Mediated AKT3 and CDK6 Degradation. Mol Ther Nucleic Acids 2020; 21: 916-931. [CrossRef]
- Ding L, Xie Z. CircWHSC1 regulates malignancy and glycolysis by the miR-212-5p/AKT3 pathway in triple-negative breast cancer. Exp Mol Pathol 2021; 123: 104704. [CrossRef]
- Qi YJ, Zha WJ, Zhang W. MicroRNA-217 alleviates development of non-small cell lung cancer by inhibiting AKT3 via PI3K pathway. Eur Rev Med Pharmacol Sci 2018; 22: 5972-5979. [CrossRef]
- Khandelwal A, Sharma U, Barwal TS, Seam RK, Gupta M, Rana MK et al. Circulating miR-320a Acts as a Tumor Suppressor and Prognostic Factor in Non-small Cell Lung Cancer. Front Oncol 2021; 11: 645475. [CrossRef]
- Shi F, Yang Q, Shen D, Chen J. CircRNA WHSC1 promotes non-small cell lung cancer progression via sponging microRNA-296-3p and up-regulating expression of AKT serine/threonine kinase 3. J Clin Lab Anal 2021; 35: e23865. [CrossRef]
- Chen S, Zhou L, Ran R, Huang J, Zheng Y, Xing M et al. Circ_0016760 accelerates non-small-cell lung cancer progression through miR-646/AKT3 signaling in vivo and in vitro. Thorac Cancer 2021; 12: 3223-3235. [CrossRef]
- Han L, Yang H, Wei W, Hu F, Yuan L. Hsa_circ_0000520 Promotes Non-Small Cell Lung Cancer Progression through the miR-1258/AKT3 Axis. J Oncol 2022; 2022: 3676685. [CrossRef]
- Zhang Y, Guo X, Xiong L, Yu L, Li Z, Guo Q et al. Comprehensive analysis of microRNA-regulated protein interaction network reveals the tumor suppressive role of microRNA-149 in human hepatocellular carcinoma via targeting AKT-mTOR pathway. Mol Cancer 2014; 13: 253. [CrossRef]
- Xing M, Xie X, Liu Z, Du X. Regulation of Tumorigenesis in Hepatocellular Carcinoma via the AKT3 Pathway in Cell Lines. Comput Math Methods Med 2021; 2021: 3267536. [CrossRef]
- Boix L, López-Oliva JM, Rhodes AC, Bruix J. Restoring miR122 in human stem-like hepatocarcinoma cells, prompts tumor dormancy through Smad-independent TGF-β pathway. Oncotarget 2016; 7: 71309-71329. [CrossRef]
- Ma Y, She XG, Ming YZ, Wan QQ, Ye QF. MicroRNA-144 suppresses tumorigenesis of hepatocellular carcinoma by targeting AKT3. Mol Med Rep 2015; 11: 1378-1383. [CrossRef]
- Yang H, Zheng W, Shuai X, Chang RM, Yu L, Fang F et al. MicroRNA-424 inhibits Akt3/E2F3 axis and tumor growth in hepatocellular carcinoma. Oncotarget 2015; 6: 27736-27750. [CrossRef]
- Zhang Y, Huang W, Ran Y, Xiong Y, Zhong Z, Fan X et al. miR-582-5p inhibits proliferation of hepatocellular carcinoma by targeting CDK1 and AKT3. Tumour Biol 2015; 36: 8309-8316. [CrossRef]
- Shu G, Su H, Wang Z, Lai S, Wang Y, Liu X et al. LINC00680 enhances hepatocellular carcinoma stemness behavior and chemoresistance by sponging miR-568 to upregulate AKT3. J Exp Clin Cancer Res 2021; 40: 45. [CrossRef]
- Fornari F, Milazzo M, Chieco P, Negrini M, Marasco E, Capranico G et al. In hepatocellular carcinoma miR-519d is up-regulated by p53 and DNA hypomethylation and targets CDKN1A/p21, PTEN, AKT3 and TIMP2. J Pathol 2012; 227: 275-285. [CrossRef]
- Li Y, Dong W, Yang H, Xiao G. Propofol suppresses proliferation and metastasis of colorectal cancer cells by regulating miR-124-3p.1/AKT3. Biotechnol Lett 2020; 42: 493-504. [CrossRef]
- Wang YX, Zhu HF, Zhang ZY, Ren F, Hu YH. MiR-384 inhibits the proliferation of colorectal cancer by targeting AKT3. Cancer Cell Int 2018; 18: 124. [CrossRef]
- Ma J, Zhang L, Shang A, Song H, Huo J, Zhang M et al. LINC02163 promotes colorectal cancer progression via miR-511-3p/AKT3 axis. Artif Cells Nanomed Biotechnol 2020; 48: 961-968. [CrossRef]
- Li B, Sun H, Zhang J. LncRNA DSCAM-AS1 promotes colorectal cancer progression by acting as a molecular sponge of miR-384 to modulate AKT3 expression. Aging (Albany NY) 2020; 12: 9781-9792. [CrossRef]
- Yang Y, Wu F, Zhang J, Sun R, Li F, Li Y et al. EGR1 interacts with DNMT3L to inhibit the transcription of miR-195 and plays an anti-apoptotic role in the development of gastric cancer. J Cell Mol Med 2019; 23: 7372-7381. [CrossRef]
- Lu Z, Luo T, Pang T, Du Z, Yin X, Cui H et al. MALAT1 promotes gastric adenocarcinoma through the MALAT1/miR-181a-5p/AKT3 axis. Open Biol 2019; 9: 190095. [CrossRef]
- Wang Z, Ma K, Pitts S, Cheng Y, Liu X, Ke X et al. Novel circular RNA circNF1 acts as a molecular sponge, promoting gastric cancer by absorbing miR-16. Endocr Relat Cancer 2019; 26: 265-277. [CrossRef]
- Li D, Tang Z, Gao Z, Shen P, Liu Z, Dang X. Circular RNA CDR1as Exerts Oncogenic Properties Partially through Regulating MicroRNA 641 in Cholangiocarcinoma. Mol Cell Biol 2020; 40. [CrossRef]
- Zeng M, Zhou Y, Zhang Y, Wang T, Wang J. Role of miR-489 in the proliferation and apoptosis of pancreatic carcinoma. J buon 2019; 24: 1574-1580.
- Teng Y, Zhang Y, Qu K, Yang X, Fu J, Chen W et al. MicroRNA-29B (mir-29b) regulates the Warburg effect in ovarian cancer by targeting AKT2 and AKT3. Oncotarget 2015; 6: 40799-40814. [CrossRef]
- Wu H, Xiao Z, Zhang H, Wang K, Liu W, Hao Q. MiR-489 modulates cisplatin resistance in human ovarian cancer cells by targeting Akt3. Anticancer Drugs 2014; 25: 799-809. [CrossRef]
- Zhao J, Yang T, Ji J, Zhao F, Li C, Han X. RHPN1-AS1 promotes cell proliferation and migration via miR-665/Akt3 in ovarian cancer. Cancer Gene Ther 2021; 28: 33-41. [CrossRef]
- Duan M, Fang M, Wang C, Wang H, Li M. LncRNA EMX2OS Induces Proliferation, Invasion and Sphere Formation of Ovarian Cancer Cells via Regulating the miR-654-3p/AKT3/PD-L1 Axis. Cancer Manag Res 2020; 12: 2141-2154. [CrossRef]
- Li L, Ma L. Upregulation of miR-582-5p regulates cell proliferation and apoptosis by targeting AKT3 in human endometrial carcinoma. Saudi J Biol Sci 2018; 25: 965-970. [CrossRef]
- Wang S, Yi M, Zhang X, Zhang T, Jiang L, Cao L et al. Effects of CDKN2B-AS1 on cellular proliferation, invasion and AKT3 expression are attenuated by miR-424-5p in a model of ovarian endometriosis. Reprod Biomed Online 2021; 42: 1057-1066. [CrossRef]
- Zuo X, Li W, Yan X, Ma T, Ren Y, Hua M et al. Long non-coding RNA LINC01224 promotes cell proliferation and inhibits apoptosis by regulating AKT3 expression via targeting miR-485-5p in endometrial carcinoma. Oncol Rep 2021; 46. [CrossRef]
- Boufraqech M, Zhang L, Jain M, Patel D, Ellis R, Xiong Y et al. miR-145 suppresses thyroid cancer growth and metastasis and targets AKT3. Endocr Relat Cancer 2014; 21: 517-531. [CrossRef]
- Li R, Liu J, Li Q, Chen G, Yu X. miR-29a suppresses growth and metastasis in papillary thyroid carcinoma by targeting AKT3. Tumour Biol 2016; 37: 3987-3996. [CrossRef]
- Lin Y, Cheng K, Wang T, Xie Q, Chen M, Chen Q et al. miR-217 inhibits proliferation, migration, and invasion via targeting AKT3 in thyroid cancer. Biomed Pharmacother 2017; 95: 1718-1724. [CrossRef]
- Feng J, Zhou Q, Yi H, Ma S, Li D, Xu Y et al. A novel lncRNA n384546 promotes thyroid papillary cancer progression and metastasis by acting as a competing endogenous RNA of miR-145-5p to regulate AKT3. Cell Death Dis 2019; 10: 433. [CrossRef]
- Fan YX, Shi HY, Hu YL, Jin XL. Circ_0000144 facilitates the progression of thyroid cancer via the miR-217/AKT3 pathway. J Gene Med 2020; 22: e3269. [CrossRef]
- Zhao C, Zhao F, Chen H, Liu Y, Su J. MicroRNA-424-5p inhibits the proliferation, migration, and invasion of nasopharyngeal carcinoma cells by decreasing AKT3 expression. Braz J Med Biol Res 2020; 53: e9029. [CrossRef]
- Fang X, Huang W, Wu P, Zeng J, Li X. CircRNA circTRAF3 promotes nasopharyngeal carcinoma metastasis through targeting miR-203a-3p/AKT3 axis. Pathol Res Pract 2021; 221: 153438. [CrossRef]
- Wang X, Li GH. MicroRNA-16 functions as a tumor-suppressor gene in oral squamous cell carcinoma by targeting AKT3 and BCL2L2. J Cell Physiol 2018; 233: 9447-9457. [CrossRef]
- Mo X, Cao Q, Liang H, Liu J, Li H, Liu F. MicroRNA-610 suppresses the proliferation of human glioblastoma cells by repressing CCND2 and AKT3. Mol Med Rep 2016; 13: 1961-1966. [CrossRef]
- Jin C, Zhao J, Zhang ZP, Wu M, Li J, Xiao GL et al. Long non-coding RNA GAS5, by up-regulating PRC2 and targeting the promoter methylation of miR-424, suppresses multiple malignant phenotypes of glioma. J Neurooncol 2020; 148: 529-543. [CrossRef]
- Liu L, Ye Q, Liu L, Bihl JC, Chen Y, Liu J et al. C6-ceramide treatment inhibits the proangiogenic activity of multiple myeloma exosomes via the miR-29b/Akt pathway. J Transl Med 2020; 18: 298. [CrossRef]
- Liu F, Wang YL, Wei JM, Huang ZD. Upregulation of circ_0000142 promotes multiple myeloma progression by adsorbing miR-610 and upregulating AKT3 expression. J Biochem 2021; 169: 327-336. [CrossRef]
- Li QY, Chen L, Hu N, Zhao H. Long non-coding RNA FEZF1-AS1 promotes cell growth in multiple myeloma via miR-610/Akt3 axis. Biomed Pharmacother 2018; 103: 1727-1732. [CrossRef]
- Liu W, Zhou Z, Zhang Q, Rong Y, Li L, Luo Y et al. Overexpression of miR-1258 inhibits cell proliferation by targeting AKT3 in osteosarcoma. Biochem Biophys Res Commun 2019; 510: 479-486. [CrossRef]
- Wang Q, Liu MJ, Bu J, Deng JL, Jiang BY, Jiang LD et al. miR-485-3p regulated by MALAT1 inhibits osteosarcoma glycolysis and metastasis by directly suppressing c-MET and AKT3/mTOR signalling. Life Sci 2021; 268: 118925. [CrossRef]
- Li J, Liu X, Li C, Wang W. miR-224-5p inhibits proliferation, migration, and invasion by targeting PIK3R3/AKT3 in uveal melanoma. J Cell Biochem 2019; 120: 12412-12421. [CrossRef]
- Wang R, Tahiri H, Yang C, Landreville S, Callejo S, Hardy P. MiR-181a-5p inhibits uveal melanoma development by targeting GNAQ and AKT3. Am J Cancer Res 2023; 13: 293-306.
- Wu SJ, Chen J, Wu B, Wang YJ, Guo KY. MicroRNA-150 enhances radiosensitivity by inhibiting the AKT pathway in NK/T cell lymphoma. J Exp Clin Cancer Res 2018; 37: 18. [CrossRef]
- Wang X, Ding Y, Wang J, Wu Y. Identification of the Key Factors Related to Bladder Cancer by lncRNA-miRNA-mRNA Three-Layer Network. Front Genet 2019; 10: 1398. [CrossRef]
- Luo B, Ma L, Xing X, Wang ZR, Teng Q, Li SG. MiR-22-3p regulates the proliferation and invasion of Wilms’ tumor cells by targeting AKT3. Eur Rev Med Pharmacol Sci 2020; 24: 5996-6004. [CrossRef]
- Grabinski N, Bartkowiak K, Grupp K, Brandt B, Pantel K, Jücker M. Distinct functional roles of Akt isoforms for proliferation, survival, migration and EGF-mediated signalling in lung cancer derived disseminated tumor cells. Cell Signal 2011; 23: 1952-1960. [CrossRef]
- Tan Y, Ge G, Pan T, Wen D, Chen L, Yu X et al. A serum microRNA panel as potential biomarkers for hepatocellular carcinoma related with hepatitis B virus. PLoS ONE 2014; 9: e107986. [CrossRef]
- Park GB, Jeong JY, Kim D. GLUT5 regulation by AKT1/3-miR-125b-5p downregulation induces migratory activity and drug resistance in TLR-modified colorectal cancer cells. Carcinogenesis 2020; 41: 1329-1340. [CrossRef]
- Mure H, Matsuzaki K, Kitazato KT, Mizobuchi Y, Kuwayama K, Kageji T et al. Akt2 and Akt3 play a pivotal role in malignant gliomas. Neuro Oncol 2010; 12: 221-232. [CrossRef]
- Nguyen TTP, Suman KH, Nguyen TB, Nguyen HT, Do DN. The Role of miR-29s in Human Cancers-An Update. Biomedicines 2022; 10. [CrossRef]
- Yan B, Guo Q, Fu FJ, Wang Z, Yin Z, Wei YB et al. The role of miR-29b in cancer: Regulation, function, and signaling. Onco Targets Ther 2015; 8: 539-548. [CrossRef]
Figure 1.
Structure of human AKT3. PH, pleckstrin homology-domain; CD, catalytic domain; RD, regulatory domain; HM, hydrophobic motif.
Figure 1.
Structure of human AKT3. PH, pleckstrin homology-domain; CD, catalytic domain; RD, regulatory domain; HM, hydrophobic motif.

Figure 2.
Dysregulation of the same miRNA/AKT3 axis in different cancers. BC, breast cancer; ccRCC, clear cell renal cell carcinoma; EC, endometrial carcinoma; EOC, epithelial ovarian cancer; GBM, glioblastoma multiforme; GC, gastric cancer; HCC, hepatocellular carcinoma; MM, multiple myeloma; NPC, nasopharyngeal carcinoma; NSCLC, non-small cell lung cancer; OC, ovarian cancer; OS, osteosarcoma; OSCC, oral squamous cell carcinoma; PC, pancreatic cancer; TC, thyroid carcinoma; UM, uveal melanoma.
Figure 2.
Dysregulation of the same miRNA/AKT3 axis in different cancers. BC, breast cancer; ccRCC, clear cell renal cell carcinoma; EC, endometrial carcinoma; EOC, epithelial ovarian cancer; GBM, glioblastoma multiforme; GC, gastric cancer; HCC, hepatocellular carcinoma; MM, multiple myeloma; NPC, nasopharyngeal carcinoma; NSCLC, non-small cell lung cancer; OC, ovarian cancer; OS, osteosarcoma; OSCC, oral squamous cell carcinoma; PC, pancreatic cancer; TC, thyroid carcinoma; UM, uveal melanoma.
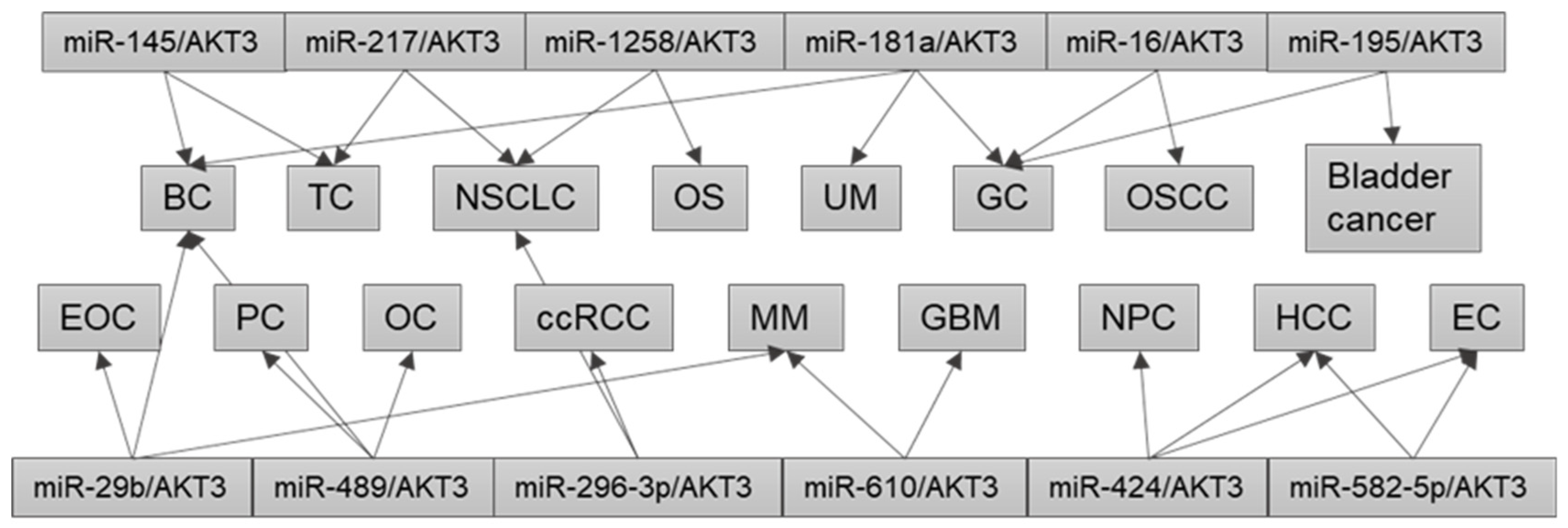
Table 1.
AKT3-derived circRNAs and their effects in human cancers.
| Tumor tissue/cell lines | CircAKT3/functions | Targets | Ref. |
|---|---|---|---|
| Lung cancer tissues; cell lines: A549 and H1299 |
hsa_circ_0000199/ glycolysis↑, cell growth↑, drug sensitivity↓ |
miR-516b-5p, STAT3 | [37] |
| GC tissues; cell lines: SGC7901, BGC823, CDDP-resistant SGC7901, CDDP-resistant BGC823 |
hsa_circ_0000199/ DNA damage repair↑, cell survival↑ |
miR-198 | [38] |
| TNBC tissues; cell lines: MCF-10A, MDA-MB-231, MDA-MB-468, SK-BR-3 |
hsa_circ_0000199/ chemo-tolerance↑, proliferation↑, migration↑, invasion↑ |
miR-206, miR-613 | [39] |
| ccRCC tissues; cell lines: OSRC-2, Caki-1, SN12-PM6, A498, SW839 |
hsa_circ_0017252/ metastasis↓ |
miR-296-3p | [40] |
| GBM tissues; cell lines: U251, HS683, SW1783, U373, glioma-initiating cells |
hsa_circ_0017250/ proliferation↓, invasiveness↓ |
AKT | [41] |
Table 2.
Roles of AKT3/Akt3 in breast cancer.
| BC model system | AKT3 or Akt3/functions | AKT3 targets | Ref. |
|---|---|---|---|
| K-RAS mutated MDA-MB-231 cells; xenografts | AKT3/cell proliferation↑, tumor growth↑, post-irradiation cell survival↑ | [21] | |
| 3475 subline of MDA-MB-231 cells (lung metastasis); MDA-MB-231 | AKT3/tumor growth↑, metastasis↑, apoptosis↓ | ERK, Bim, Bax | [45] |
| MDA-MB-231; MDA-MB-468 and MCF10DCIS xenografts | AKT3/TNBC growth↑ | p27 | [46] |
| ErbB2(+) BC cells, mammary tumor cells | AKT3/cell proliferation↑, tamoxifen sensitivity↓ | pErbB2/pErbB3, Foxo3a, ERα | [47] |
| MMTV-ErbB2, MMTV-PyMT mice (Neu- and PyMT- driven mammary oncogenesis) | Akt3/no effect on tumorigenesis of mouse BC cells | [48] | |
| PyMT mouse BC cells | Akt3/metastasis of mouse BC cells↓ | [49] | |
| MDA-MB-231 BO cells; xenografts | AKT3/migration↓, invasion↓ bone metastasis↓ | HER2, DDR kinase | [50] |
| IBC cells: SUM149 | AKT3/survival of IBC↑, no effect on invasion | [51] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



