
Submitted:
19 October 2023
Posted:
20 October 2023
You are already at the latest version
Abstract
Advanced cutaneous melanoma is considered the most aggressive type of skin cancer with variable rates of treatment response. Nowadays, there are available some classes of immunotherapy and target therapy for its treatment. Immunotherapy can inhibit tumor growth and its recurrence by triggering host's immune system, while targeted therapy acts inhibiting specific molecules or signaling pathways. However, melanoma response to these treatments are highly heterogenous, and frequently become resistant.
Epigenome (DNA/histone modification) contribute to cancer initiation and progression. Epigenetic alterations are divided into four levels of gene expression regulation: DNA methylation, histone modification, chromatin remodeling and non-coding RNAs regulation. The deregulation of lysine methyltransferase enzymes is associated with tumor initiation, invasion, development of metastases, changes in immune microenvironment and drug resistance.
The study of lysine histone methyltransferase (KMT) inhibitors is important to cancer epigenetic mechanisms understanding and to biological processes. In addition to immunotherapy and target therapy, the research and development of KMT inhibitors is ongoing. Many studies are exploring the therapeutic implications and possible side effects of these compounds, besides their adjuvant potential to the approved current therapies. Importantly, as with any drug development, safety, efficacy, and specificity are crucial considerations when developing KMT inhibitors for clinical applications.
Keywords:
advanced melanoma
; immunotherapy
; target therapy
; lysine histone methyl transferase inhibitors
; UNC0642
1. Introduction
Cutaneous melanoma is the skin cancer that originates from melanocytes or their precursors, melanoblast or melanocyte stem cells. Melanoma is considered the most aggressive type of skin cancer due to its ability to fast spread through the body [1]. Endogenous risk factors such as genetic susceptibility, are accountable for 8 to 10% of the cases, being the cyclin-dependent kinase inhibitor 2A (CDKN2A) the main high-risk gene for melanoma [2]. Alternatively, excessive exposure to the sun ultraviolet radiation (modifiable factor) is its main exogenous risk factor for cutaneous melanoma development. This includes prolonged exposure to the sun without adequate protection, as well as, the use of tanning beds, which emits sun-like radiation [1]. Yet, random errors in DNA replication (unmodifiable factor) will contribute as an intrinsic risk factor for the disease development [2].
Here, we will review therapies and their mechanisms for advanced cutaneous melanoma, from immunotherapies to lysine histone methyl transferase inhibitors. Disease early detection is essential for the successful treatment of cutaneous melanoma independent of the treatment type.
2. Advanced Melanoma Treatments
2.1. Immunotherapy
Immunotherapy can inhibit tumor growth and its recurrence by triggering host’s immune system or improving the anti-tumor response. Thus far, several immunotherapy drugs, mainly the blockade of negative immune checkpoints, have been approved by the FDA (Food and Drug Administration) for metastatic melanoma [3]. Currently, immunotherapy has achieved remarkable results in the treatment of advanced cutaneous melanoma. Some patients experience a lasting response, with tumors significantly decreasing in size or even disappearing completely [4,5,6,7].
Three classes of immune checkpoint inhibitors are available for clinical use: the inhibitors of the CTLA4 (Cytotoxic T-Lymphocyte Associated Protein 4) receptors (Ipilimumab), inhibitors of the PD-1 (Programmed Cell Death-1) co-receptor (Nivolumab, Spartalizumab, Pembrolizumab, Toripalimab) or inhibitors of their ligand PD-L1 (Atezolizumab), and inhibitors of the LAG3 (Lymphocyte-Activation Gene 3) transmembrane protein (Relatlimab) or the combination of anti-PD-1 with anti-LAG3 (Opdualag). These proteins are mainly expressed by T cells, PDL1 can also be expressed by tumor cells or other cells in the tumor environment (Figure 1a).
Clinical studies in other types of immunotherapies as therapeutic mRNA vaccines, wich induces the production of tumoral immunogenic proteins by antigen presenting cells and leads to T cells stimulation (Figure 1b), or tumor-infiltrating lymphocytes (TILs), that are removed from inside tumors and “in vitro” expanded, to return to pacients activated and in very higher numbers (Figure 1c). Both treatments point to be promising modalities for advanced cutaneous melanoma, alone or in combination with other therapies [3,8,9,10,11].
Other forms of treatment, such as interleukins and oncolytic viruses, also work by stimulating the immune system. Interleukins are proteins that can boost the immune system by inducing T lymphocytes activation. Interleukin-2 (IL-2, Aldesleukin, Proleukin) are sometimes used in patients with melanoma, and target the T lymphocytes activation enhancing the activation-induced cell death (AICD)[12,13].
Some viruses can be modified in the laboratory to infect and destroy cancer cells. Talimogene laherparepvec, also known as T-VEC, is an example of an oncolytic virus that can be used to treat melanomas and is injected directly into tumors [14].
The response to immunotherapy varies from person to person, and not all patients experience the same results. Around 60% of patients develop resistance to immunotherapy treatment and their disease return to progresses [15]. In addition, it is important to highlight that immunotherapy can also be associated with significant side effects, as it activates the immune system more intensely. These side effects can range from mild symptoms such as fatigue and nausea to more serious immune reactions that can affect specific organs. Therefore, proper administration and monitoring are essential during these type of treatment [16,17,18].
2.2. Target therapy
Targeted therapy is another important approach for the treatment of advanced cutaneous melanoma. This therapy inhibits specific molecules or signaling pathways, the mitogen-activated protein kinase (MAPK) cascade is an intracellular signaling pathway which is implicated in the cell proliferation and survival regulation (Figure 2a) [19,20,21]. Several different mutations, involving BRAF or NRAS, display an oncogenic effect by activating the MAPK pathway (Figure 2b), resulting in an increased tumor cell proliferation [22].
Target therapy is frequently used in melanoma that display mutations in the NRAS and BRAF gene. The BRAF mutations are mainly in BRAFV600E (~80%) and BRAFV600K (~5 to 30%), which cause alteration in the protein, that participates in the MAPK/ERK signaling pathway (Figure 2b-c) [22]. Nevertheless, there is no specific targeted therapy for NRAS-mutant melanomas. Selective MEK inhibitors denote a new therapeutic alternative, having been shown to inhibit growth and induce cell death in both BRAF and NRAS mutant melanoma cell lines (Figure 2c) [23].
There are two mains targeted therapy in cutaneous melanoma, and they are most frequently used in combination: BRAF inhibitors (Vemurafenib, Dabrafenib and Encorafenib) and MEK inhibitors (Trametinib, Cobimetinib and Binimetinib). Unfortunately, most patients respond well initially, but later the response is abrogated due to the development of resistance mechanisms by the tumor. The association of immunotherapy with targeted therapy is widely used to overcome resistance [24].
In addition, studies revealed the correlation of melanoma with several somatic driver mutations, including alterations in KIT, a proto-oncogene encoding for a receptor tyrosine kinase (RTK). Although accounting for only 3% of all melanomas, mutations in c-KIT are mostly derived from acral, mucosal, and chronically sun-damaged melanomas [25]. Target therapy drugs such as Imatinib and Nilotinib can block the proliferation of these tumor cells.
2.3. Lysine histone methyl transferase inhibitors
2.3.1. Methyltransferases
Methyltransferases are essential enzymes for several biological processes, including regulation of gene expression, modulation of chromatin structure, and cell signaling. These enzymes catalyze the transfer of methyl groups (-CH3) from methyl donors to specific targets such as proteins, nucleic acid, or other molecules. The two most important classes are DNA and protein methyltransferases.
2.3.2. DNA methyltransferases
DNA methylation is catalyzed by a group of proteins known as DNA methyltransferases (DNMTs). DMNTs s are enzymes that add methyl groups to nitrogenous bases (Figure 3a), which has a significant impact on the regulation of gene expression, since DNA methylation can silence gene activity. Hypermethylation of gene promoter regions can lead to inhibition of gene transcription and is associated with pathological processes, including cancer [26].
2.3.3. Protein methyltransferases
Protein methyltransferases (PMT) are enzymes that add methyl groups into amino acid residues,, which is principally observed in lysine and arginine residues (Figure 3b). This can affect the structure and function of proteins by regulating enzymatic activity and protein-protein interactions. A well-known example is the methylation of arginine in histones, proteins that help organize DNA into chromatin [26].
2.3.4. Histones and Histone methylation
Histones are proteins that provide structural support to a chromosome and can undergo a series of post-translational modifications, including acetylation, methylation, phosphorylation, ubiquitination, and sumoylation.
Histone methylation, analogous to DNA methylation, has been associated to gene repression [28]. However, several lysine methylation patterns can indicate expressed genes, such as tri-methylated H3K4, or H3K9 mono-methylation [29].
Histones are methylated by histone methyltransferases (HMT) and methylation is removed by histone demethylases (HDM).
Methylation of histone take place either at lysine or arginine residues, on histones H3 and H4, [30]. Methylation of histones H3K4, H3K36, and H3K79 is related with gene activation [30], while methylation on H3K9 or H3K27 is correlated to transcriptional repression. On histone H4, K20 methylation is a known mark of gene silencing [30]. Similarly, to lysine methylation, arginine methylation has been linked to both gene activation (H3R17) and repression (H3R2, H4R3) [31]. Lysine can be mono-, di- and trimethylated, whereas arginine can only be mono- or demethylated [32]. These modifications regulate the ability of transcription factors to access underlying DNA and impact transcription, replication, and chromatin stability [33,34,35]. Histone modifications are critical epigenetic drivers that can alter the chromatin state and are implicated in cancer progression [36].
2.3.5. Histone Methyltransferases (HMT)
Histone methyltransferases (HMT) are a group of enzymes that catalyze the addition of methyl groups (-CH3) to specific amino acid residues on histones (Figure 3c).
HMT are essential for the maintenance of chromatin structure and the regulation of gene expression. Dysfunctions in these enzymes can lead to epigenetic changes that are associated with several diseases, including cancer. For example, aberrant histone methylation can result in inadequate activation or silencing of genes, contributing to the development of tumors [37].
2.3.6. Lysine Histone Methyltransferases (KMT)
Lysine histone methyltransferase (KMT), also known as histone lysine methyltransferases (HKMT), are the group of enzymes that catalyze the addition of methyl groups (-CH3) to lysine residues on histones. KMT transfers methyl group from a methyl donor (usually a coenzyme S-adenosylmethionine, SAM) to a lysine residue present on histones. SAM is assembled with methionine and ATP, a process that produces phosphate and pyrophosphate. After transfer of the methyl group, SAM is converted into S-adenonsyl-homocysteine (SHA) (Figure 4a) [38].
The canonical histone lysine methylation found in humans methylates histone 3 and 4, and specific lysenes sites as H3K4, H3K9, H3K27, H3K36, H3K79 e H4K20. These modifications are generated by a total of 24 different enzymes: 23-SET proteins and one 7βS protein. In general, histone KMTs are highly selective, as an enzymes that methylates H3K36 do not methylates a different lysine if K36 is mutated. Beyond the canonical sites, many other methylation events have been identified by different methods, including mass spectrometry, some only are found during diseases or) [38].
The canonical histone lysine methylation is associated with different chromatin states and have specific implications in gene regulation. As H3K4 is often associated with more accessible and active chromatin regions where gene transcription is more likely to occur. This methylation is generally seen as a marker of active genes enhancer and promoter. H3K9 is typically associated with regions of compact chromatin and gene silencing. This occurs because H3K9 methylation recruits proteins that promote the formation of heterochromatin, a densely packed form of chromatin that tends to silence transcription. H3K27 is also associated with gene silencing. It is often found in regions of chromatin that regulate cell development and differentiation [39].
G9a is a nuclear histone KMT belonging to the Su(var)3-9 family, which mostly catalyzes H3K9 mono- and dimethylation. G9a SET domain is accountable for the addition of methyl groups on H3, whereas the ankyrin repeats have been described to denote mono- and dimethyl lysine binding regions [40]. G9a is thus not only able to methylate histone tails but also able to identify this modification, operating as a platform for the recruitment of other target molecules on the chromatin [41].
A G9a-like protein (GLP) has also been recognized, this protein interacts with G9a, forming a heterodimeric complex. This heteromeric structure (G9a-GLP) is the main form and represents the active stat, of this methyltransferase in vivo [42]. Although the heterodimer appears to be essential for G9a–GLP methyltransferase activity, the enzymatic activity of G9a is more important for the in vivo function of the complex [28].
The H3K9 is initially methylated by G9a-GLP to form active H3K9 monomethylation, which is subsequently methylated by G9a to form repressive H3K9 dimethylation (H3K9me2). G9a directly contributes to the dimethylation and subsequent trimethylation at H3K9 across the genome (Figure 4b). Depending on the modified residue position, histone methylation can either suppress (H3K9, H3K27) or enhance (H3K4) gene expression [39].
During hypoxia, the histone methyltransferase G9a activity increases, leading to an increase in global histone H3K9 methylation (Figure 4c), this higher methylation inhibits the expression of cell adhesion molecules such as E-cadherin. The correlation between G9a-mediated repression of cell adhesion molecules and the increase in their activity during hypoxia strongly supports the direct involvement of G9a in the metastatic pathway [39].
In addition, G9a enhanced expression is also linked to malignant behaviors of cancer cells as aberrant proliferation, metastasis and drug resistance by silencing tumor suppressors or by activating epithelio-mesenchymal transition programs [43,44]. Its overexpression is associated with a poor prognosis in different types of cancer, including melanoma, by positively regulating the Nocth1 signaling pathway [40,45,46]. Positive signaling from the Notch1 pathway specifically contributes to the development of melanoma, allowing the survival and proliferation of these cells in stressful, hypoxic environments [47,48,49,50].
Consequently, lysine methyltransferase (KMT) can be a target molecules for cancer and metastasis treatment, but differently from the therapies discussed before, these therapies are under development. The first FDA approval for KMT inhibitor (Tazemetostat for epithelioid sarcoma4 and subsequently follicular lymphoma) occurred in 2020, and until now, no current approvals is known [51].
2.3.7. KMT inhibitors
KMT inhibitors are a class of compounds that target KMT enzymes. These inhibitors were developed aiming to modulate the activity of KMT and, consequently, epigenetic modifications on histones. This may have implications for the regulation of gene expression and associated cellular processes, including development, cell differentiation, and disease [52].
KMT inhibitors can be classified into two main types.
Selective inhibitors: These inhibitors are designed to target a specific KMT enzyme. This allows a precise approach by targeting specific epigenetic modifications at specific histone sites. Examples of selective inhibitors include those that target KMT for H3K9 or H3K27 methylations [51].
2.3.8. Broad-spectrum inhibitors
These inhibitors target multiple KMTs in a less specific manner. They affect broader spectrum of epigenetic modifications and may have widespread effects in cells. This can be beneficial for manipulating gene expression generally, but may also result on off targeting side effects [51]. Research into KMT inhibitors is an expanding area with significant implications in basic research, as well as, in the development of potential new drugs. However, the complexity of epigenetic regulation and the interconnection in cell signaling pathways, is a challenge. Furthermore, specificity is a critical aspect, since inappropriate inhibition of a KMT can lead to off target side effects.
G9a-GLP inhibitors
G9a and GLP have approximately 80% sequence identity in their conserved catalytic SET domains, which imposes a challenge for developing dual selective inhibitors [53]. Several G9a-GLP dual inhibitors have been developed by targeting either the SAM binding site (methyl donor) or the substrate binding pocket [54].
Currently, three compounds were idenfied as G9a-GLP inhibitors. The compound BIX01294 and UNC0638 are being tested in vitro, and UNC0642 alread being tested in vivo in preclinical studies with animal models.
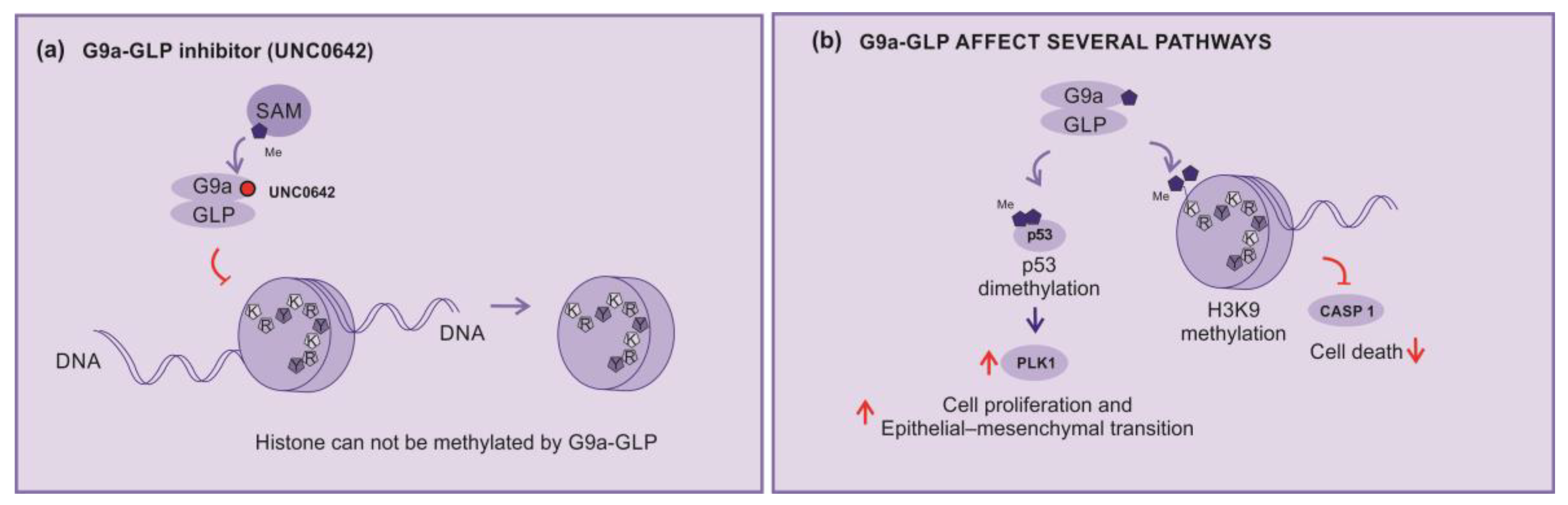
In this sense, the compound UNC0642 acts as a competitive inhibitor of G9a activity, connecting to its binding poket turning it inaccessible (Figure 5a). This compound is described as a potent inhibitor of G9a-GLP, with low toxicity and high selectivity [55,56].
UNC0642 studies are more advanced due to its favorable pharmacokinetics, better half-life, high selectivity and low cellular toxicity, making it more suitable for preclinical studies [56,57].
G9a-GLP is implicated in several mechanisms and pathways as inhibition of the expression of the caspase 1 gene (CASP1) by enhancing H3K9me2 of the promoter region. In addition, G9a is able to dimethylate p53, inhibting their activity, and enhancing the expression of polo-like kinase 1 (Plk1), which has being implicated in cell proliferation and epithelial–mesenchymal transition (Figure 5b) [58].
Consequently, G9a-GLP inhibitors can impede tumor growth by blocking the p53 dimethylation. Active p53 will block cell cycle progression of cells with DNA damages and dimish Plk1 expression. Yet, the unblock of caspase 1 expression, will support the cells inflammasomes formation and cell death [44,58].
Concerning to melanoma, as discussed before the Noch-1 signaling is enhanced in the disease, and also by the H3K9 di-trimethylation, which leads to MEK inhibitors resistence [59], and microphthalmia-associated transcription factor (MITF) repression, via a competition-based mechanism, thereby triggering the critical transit into the invasive melanoma stage [60]. Thus, the G9a inhibitors, in addition to impair commom cancer mechanisms, also impari specific mechanisms related to melanoma [45].
G9a plays significant role in cancer processes, as heterochromatin formation, DNA methylation, transcriptional silencing, proliferation, cell death, differentiation, and mobility of tumor cells [61]. Thus, G9a is held as an outstanding and novel therapeutic target for anti-cancer agents; therefore, the development of novel G9a inhibitors may provide a new option for invasive melanoma treatment. Even though, the knowledge about the mechanisms involved in KMT inhibtors are being improved, much remain to be answered, and in this sense, experimental research and preclinical studies still having plenty to add to the current knowledge.
A recent preclinical study combining G9a inhibitor (UNC0642) with anti-PD1 showed increased immunotherapy efficacy, with increased survival and lower incidence of acquired resistance to checkpoint inhibitor in murine melanoma [62]. Clinical studies using KMT inhibitors for advanced cutaneous melanoma are still in the early stages. They are being tested alone or in combination with immune checkpoint blockage(anti-CTLA4, anti-PD1) or targeted therapy agents (BRAF/MEK inhibitors). It is believed that these drugs can reverse resistance to immunotherapeutics and targeted therapy agents making treatment more effective.
Important to note that and many studies are exploring the therapeutic implicationsof G9a inhibitors, but also their possible side effects. Importantly, as with any pharmacological drug development, safety, efficacy, and specificity are crucial considerations when developing new grugs for aiming clinical aplications.
4. Conclusions
Most patients with advanced cutaneous melanoma usually die from the disease. New treatments for these patients are necessary. One of the lines of this search for new treatments are epigenetic medicines. Lysine methyl transferase inhibitors are being tested alone or in combination with immune checkpoint inhibitors or target therapy and are a promise in reducing acquired resistance to these treatments.
Author Contributions
RSOF: conceptualization and review the manuscript ; DAO: data curation and review the menuscript; MMN: bibliographic survey and review the manuscript; LCM: supervision, illustration and final review. All authors have read and agreed to the published version of the manuscript
Funding
“Jorge de Moura Andrews” Study Center, UNIFESP Plastic Surgery Discipline
Conflicts of Interest
The authors declare no conflict of interest
References
- Shain AH, Bastian BC. From melanocytes to melanomas. Nat Rev Cancer. 2016; 16(6):345-58. [CrossRef]
- Potrony M, Badenas C, Aguilera P, Puig-Butille JA, Carrera C, Malvehy J, Puig S. Update in genetic susceptibility in melanoma. Ann Transl Med. 2015; 3(15):210. [CrossRef]
- Qin Z, Zheng M. Advances in targeted therapy and immunotherapy for melanoma (Review). Exp Ther Med. 2023; 13;26(3):416. [CrossRef]
- Hodi FS, O’Day SJ, McDermott DF, Weber RW, Sosman JA, Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel JC, Akerley W, van den Eertwegh AJ, Lutzky J, Lorigan P, Vaubel JM, Linette GP, Hogg D, Ottensmeier CH, Lebbé C, Peschel C, Quirt I, Clark JI, Wolchok JD, Weber JS, Tian J, Yellin MJ, Nichol GM, Hoos A, Urba WJ. Improved survival with ipilimumab in patients with metastatic melanoma. N Engl J Med. 2010; 363(8):711-23. [CrossRef]
- Eggermont AM, Chiarion-Sileni V, Grob JJ, Dummer R, Wolchok JD, Schmidt H, Hamid O, Robert C, Ascierto PA, Richards JM, Lebbé C, Ferraresi V, Smylie M, Weber JS, Maio M, Bastholt L, Mortier L, Thomas L, Tahir S, Hauschild A, Hassel JC, Hodi FS, Taitt C, de Pril V, de Schaetzen G, Suciu S, Testori A. Prolonged Survival in Stage III Melanoma with Ipilimumab Adjuvant Therapy. N Engl J Med. 2016;375(19):1845-1855. [CrossRef]
- Weber J, Mandala M, Del Vecchio M, Gogas HJ, Arance AM, Cowey CL, Dalle S, Schenker M, Chiarion-Sileni V, Marquez-Rodas I, Grob JJ, Butler MO, Middleton MR, Maio M, Atkinson V, Queirolo P, Gonzalez R, Kudchadkar RR, Smylie M, Meyer N, Mortier L, Atkins MB, Long GV, Bhatia S, Lebbé C, Rutkowski P, Yokota K, Yamazaki N, Kim TM, de Pril V, Sabater J, Qureshi A, Larkin J, Ascierto PA; CheckMate 238 Collaborators. Adjuvant Nivolumab versus Ipilimumab in Resected Stage III or IV Melanoma. [CrossRef]
- Tawbi HA, Schadendorf D, Lipson EJ, Ascierto PA, Matamala L, Castillo Gutiérrez E, Rutkowski P, Gogas HJ, Lao CD, De Menezes JJ, Dalle S, Arance A, Grob JJ, Srivastava S, Abaskharoun M, Hamilton M, Keidel S, Simonsen KL, Sobiesk AM, Li B, Hodi FS, Long GV; RELATIVITY-047 Investigators. Relatlimab and Nivolumab versus Nivolumab in Untreated Advanced Melanoma. N Engl J Med. 2022;386(1):24-34. [CrossRef]
- Rosenberg SA, Yang JC, Sherry RM, Kammula US, Hughes MS, Phan GQ, Citrin DE, Restifo NP, Robbins PF, Wunderlich JR, Morton KE, Laurencot CM, Steinberg SM, White DE, Dudley ME. Durable complete responses in heavily pretreated patients with metastatic melanoma using T-cell transfer immunotherapy. Clin Cancer Res. 2011;17(13):4550-7. [CrossRef]
- Sarnaik AA, Hamid O, Khushalani NI, Lewis KD, Medina T, Kluger HM, Thomas SS, Domingo-Musibay E, Pavlick AC, Whitman ED, Martin-Algarra S, Corrie P, Curti BD, Oláh J, Lutzky J, Weber JS, Larkin JMG, Shi W, Takamura T, Jagasia M, Qin H, Wu X, Chartier C, Graf Finckenstein F, Fardis M, Kirkwood JM, Chesney JA. Lifileucel, a Tumor-Infiltrating Lymphocyte Therapy, in Metastatic Melanoma. J Clin Oncol. 2021;39(24):2656-2666. [CrossRef]
- Seitter SJ, Sherry RM, Yang JC, Robbins PF, Shindorf ML, Copeland AR, McGowan CT, Epstein M, Shelton TE, Langhan MM, Franco Z, Danforth DN, White DE, Rosenberg SA, Goff SL. Impact of Prior Treatment on the Efficacy of Adoptive Transfer of Tumor-Infiltrating Lymphocytes in Patients with Metastatic Melanoma. Clin Cancer Res. 2021;27(19):5289-5298. [CrossRef]
- Qin S, Tang X, Chen Y, Chen K, Fan N, Xiao W, Zheng Q, Li G, Teng Y, Wu M, Song X. mRNA-based therapeutics: powerful and versatile tools to combat diseases. Signal Transduct Target Ther. 2022; 7(1):166. [CrossRef]
- Rosenberg SA, Lotze MT, Muul LM, Leitman S, Chang AE, Ettinghausen SE, Matory YL, Skibber JM, Shiloni E, Vetto JT, et al. Observations on the systemic administration of autologous lymphokine-activated killer cells and recombinant interleukin-2 to patients with metastatic cancer. N Engl J Med. 1985;313(23):1485-92. [CrossRef]
- Muhammad S, Fan T, Hai Y, Gao Y, He J. Reigniting hope in cancer treatment: the promise and pitfalls of IL-2 and IL-2R targeting strategies. Mol Cancer. 2023;22(1):121. [CrossRef]
- Andtbacka RH, Colichio F, Harrington KJ et al. Final analyses of OPTiM: a randomized phase III trial of talimogene laherparepvec versus granulocyte-macrophage colony-stimulating factor in unresectable stage III–IV melanoma. J Immunother Cancer. 2019; 7:145-56.
- Bai R, Chen N, Li L, Du N, Bai L, Lv Z, Tian H, Cui J. Mechanisms of Cancer Resistance to Immunotherapy. Front Oncol. 2020 ;10:1290. . [CrossRef]
- Ribas A, Camacho LH, Lopez-Berestein G, Pavlov D, Bulanhagui CA, Millham R, Comin-Anduix B, Reuben JM, Seja E, Parker CA, Sharma A, Glaspy JA, Gomez-Navarro J. Antitumor activity in melanoma and anti-self responses in a phase I trial with the anti-cytotoxic T lymphocyte-associated antigen 4 monoclonal antibody CP-675,206. J Clin Oncol. 2005;23(35):8968-77. [CrossRef]
- Liao B, Shroff S, Kamiya-Matsuoka C, Tummala S. Atypical neurological complications of ipilimumab therapy in patients with metastatic melanoma. Neuro Oncol. 2014;16(4):589-93. [CrossRef]
- Bertrand A, Kostine M, Barnetche T, Truchetet ME, Schaeverbeke T. Immune related adverse events associated with anti-CTLA-4 antibodies: systematic review and meta-analysis. BMC Med. 2015; 13:211. [CrossRef]
- Chang L, Karin M. Mammalian MAP kinase signalling cascades. Nature. 2001;410(6824):37-40. [CrossRef]
- Liebmann C. Regulation of MAP kinase activity by peptide receptor signalling pathway: paradigms of multiplicity. Cell Signal. 2001;13(11):777-85. [CrossRef]
- Grimaldi AM, Simeone E, Ascierto PA. The role of MEK inhibitors in the treatment of metastatic melanoma. Curr Opin Oncol. 2014;26(2):196-203. [CrossRef]
- Thompson N, Lyons J. Recent progress in targeting the Raf/MEK/ERK pathway with inhibitors in cancer drug discovery. Curr Opin Pharmacol. 2005;5(4):350-6. [CrossRef]
- Grimaldi AM, Simeone E, Festino L, Vanella V, Strudel M, Ascierto PA. MEK Inhibitors in the Treatment of Metastatic Melanoma and Solid Tumors. Am J Clin Dermatol. 2017;18(6):745-754. 7: Dermatol. 2017;18(6). [CrossRef]
- Lopes J, Rodrigues CMP, Gaspar MM et al. Melanoma Management: From Epidemiology to Treatment and Latest Advances. Cancers (Basel). 2022; 14(19): 4652-76. [CrossRef]
- Pham DDM, Guhan S, Tsao H. KIT and Melanoma: Biological Insights and Clinical Implications. Yonsei Med J. 2020;61(7):562-571. [CrossRef]
- Del Castillo Falconi VM, Torres-Arciga K, Matus-Ortega G, Díaz-Chávez J, Herrera LA. DNA Methyltransferases: From Evolution to Clinical Applications. Int J Mol Sci. 2022 ;23(16):8994. [CrossRef]
- Kaniskan HÜ, Martini ML, Jin J. Inhibitors of Protein Methyltransferases and Demethylases. Chem Rev. 2018;118(3):989-1068. [CrossRef]
- Tachibana M, Matsumura Y, Fukuda M, Kimura H, Shinkai Y. G9a/GLP complexes independently mediate H3K9 and DNA methylation to silence transcription. EMBO J. 2008;27(20):2681-90. [CrossRef]
- Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129(4):823-37. [CrossRef]
- Black JC, Van Rechem C, Whetstine JR. Histone lysine methylation dynamics: establishment, regulation, and biological impact. Mol Cell. 2012;48(4):491-507. [CrossRef]
- Zhao Q, Rank G, Tan YT, Li H, Moritz RL, Simpson RJ, Cerruti L, Curtis DJ, Patel DJ, Allis CD, Cunningham JM, Jane SM. PRMT5-mediated methylation of histone H4R3 recruits DNMT3A, coupling histone and DNA methylation in gene silencing. Nat Struct Mol Biol. 2009;16(3):304-311. [CrossRef]
- Rea S, Eisenhaber F, O’Carroll D, Strahl BD, Sun ZW, Schmid M, Opravil S, Mechtler K, Ponting CP, Allis CD, Jenuwein T. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nature. 2000;406(6796):593-9. [CrossRef]
- Bates S. Epigenetic Therapies for Cancer. N Engl J Med. 2021; 383:650-63. [CrossRef]
- Moran B, Silva R, Perry AS, Gallagher W. Epigenetics of malignant melanoma. Semin Cancer Biol 2021;51:80-88. [CrossRef]
- Harel T, Lupski JR. Genomic disorders 20 years on-mechanisms for clinical manifestations. Clin Genet 2021; 93:439-49. [CrossRef]
- Sang Y, Deng Y. Current insights into the epigenetic mechanisms of skin cancer. Dermatol Ther. 2019;32(4):e12964. [CrossRef]
- Nacev BA, Feng L, Bagert JD, Lemiesz AE, Gao J, Soshnev AA, Kundra R, Schultz N, Muir TW, Allis CD. The expanding landscape of ‘oncohistone’ mutations in human cancers. Nature. 2019;567(7749):473-478. [CrossRef]
- Husmann D, Gozani O. Histone lysine methyltransferases in biology and disease. Nat Struct Mol Biol. 2019;26(10):880-889. [CrossRef]
- Karami Fath M, Azargoonjahromi A, Soofi A, Almasi F, Hosseinzadeh S, Khalili S, Sheikhi K, Ferdousmakan S, Owrangi S, Fahimi M, Zalpoor H, Nabi Afjadi M, Payandeh Z, Pourzardosht N. Current understanding of epigenetics role in melanoma treatment and resistance. Cancer Cell Int. 2022;22(1):313. [CrossRef]
- Casciello F, Windloch K, Gannon F, Lee JS. Functional Role of G9a Histone Methyltransferase in Cancer. Front Immunol. 2015; 6:487. [CrossRef]
- Shahbazian MD, Zhang K, Grunstein M. Histone H2B ubiquitylation controls processive methylation but not monomethylation by Dot1 and Set1. Mol Cell. 2005;19(2):271-7. [CrossRef]
- Tachibana M, Ueda J, Fukuda M, Takeda N, Ohta T, Iwanari H, Sakihama T, Kodama T, Hamakubo T, Shinkai Y. Histone methyltransferases G9a and GLP form heteromeric complexes and are both crucial for methylation of euchromatin at H3-K9. Genes Dev. 2005;19(7):815-26. [CrossRef]
- Fan J, Xing Y, Wen X, Jia R, Ni H, He J, Ding X, Pan H, Qian G, Ge S, Hoffman AR, Zhang H, Fan X. Long non-coding RNA ROR decoys gene-specific histone methylation to promote tumorigenesis. Genome Biol. 2015;16(1):139. [CrossRef]
- Liao Q, Yang J, Ge S, Chai P, Fan J, Jia R. Novel insights into histone lysine methyltransferases in cancer therapy: From epigenetic regulation to selective drugs. J Pharm Anal. 2023;13(2):127-141. [CrossRef]
- Dang NN, Jiao J, Meng X, An Y, Han C, Huang S. Abnormal overexpression of G9a in melanoma cells promotes cancer progression via upregulation of the Notch1 signaling pathway. Aging. 2020;12(3):2393-2407. [CrossRef]
- Filho RSO, Peixoto GR, Sangiuliano LDC, Oliveira DA. Notch receptors as a therapeutic target in melanoma: a narrative bibliographic review. Braz. J. Nat. Sci 2021; 4:614-28. [CrossRef]
- Ayaz F, Osborne BA. Non-canonical notch signaling in cancer and immunity. Front Oncol. 2014; 4:345. [CrossRef]
- Bedogni B. Notch signaling in melanoma: interacting pathways and stromal influences that enhance Notch targeting. Pigment Cell Melanoma Res. 2014;27(2):162-8. [CrossRef]
- Zhang K, Wong P, Salvaggio C, Salhi A, Osman I, Bedogni B. Synchronized Targeting of Notch and ERBB Signaling Suppresses Melanoma Tumor Growth through Inhibition of Notch1 and ERBB3. J Invest Dermatol. 2016;136(2):464-472. [CrossRef]
- Tang H, Xiao WR, Liao YY, Li L, Xiao X, Xu XP, Feng H. EGFL7 silencing inactivates the Notch signaling pathway; enhancing cell apoptosis and suppressing cell proliferation in human cutaneous melanoma. Neoplasma. 2019;66(2):187-196. [CrossRef]
- Bhat KP, Ümit Kaniskan H, Jin J, Gozani O. Epigenetics and beyond: targeting writers of protein lysine methylation to treat disease. Nat Rev Drug Discov. 2021;20(4):265-286. [CrossRef]
- Rugo HS, Jacobs I, Sharma S, Scappaticci F, Paul TA, Jensen-Pergakes K, Malouf GG. The Promise for Histone Methyltransferase Inhibitors for Epigenetic Therapy in Clinical Oncology: A Narrative Review. Adv Ther. 2020;37(7):3059-3082. [CrossRef]
- Link PA, Gangisetty O, James SR, Woloszynska-Read A, Tachibana M, Shinkai Y, Karpf AR. Distinct roles for histone methyltransferases G9a and GLP in cancer germ-line antigen gene regulation in human cancer cells and murine embryonic stem cells. Mol Cancer Res. 2009 ;7(6):851-62. [CrossRef]
- Sweis RF, Pliushchev M, Brown PJ, Guo J, Li F, Maag D, Petros AM, Soni NB, Tse C, Vedadi M, Michaelides MR, Chiang GG, Pappano WN. Discovery and development of potent and selective inhibitors of histone methyltransferase g9a. ACS Med Chem Lett. 2014; 2;5(2):205-9. [CrossRef]
- Park KS, Xiong Y, Yim H, Velez J, Babault N, Kumar P, Liu J, Jin J. Discovery of the First-in-Class G9a/GLP Covalent Inhibitors. J Med Chem. 2022;65(15):10506-10522. [CrossRef]
- Liu F, Barsyte-Lovejoy D, Li F, Xiong Y, Korboukh V, Huang XP, Allali-Hassani A, Janzen WP, Roth BL, Frye SV, Arrowsmith CH, Brown PJ, Vedadi M, Jin J. Discovery of an in vivo chemical probe of the lysine methyltransferases G9a and GLP. J Med Chem. 2013; 56(21):8931-42. [CrossRef]
- Flesher JL, Fisher DE. G9a: An Emerging Epigenetic Target for Melanoma Therapy. Epigenomes. 2021;5(4):23. [CrossRef]
- Cao H, Li L, Yang D, Zeng L, Yewei X, Yu B, Liao G, Chen J. Recent progress in histone methyltransferase (G9a) inhibitors as anticancer agents. Eur J Med Chem. 2019;179:537-546. [CrossRef]
- Porcelli L, Mazzotta A, Garofoli M, Di Fonte R, Guida G, Guida M, Tommasi S, Azzariti A. Active notch protects MAPK activated melanoma cell lines from MEK inhibitor cobimetinib. Biomed Pharmacother. 2021;133:111006. [CrossRef]
- Golan T, Levy C. Negative Regulatory Loop between Microphthalmia-Associated Transcription Factor (MITF) and Notch Signaling. Int J Mol Sci. 2019 ;20(3):576. [CrossRef]
- Kato S, Weng QY, Insco ML, Chen KY, Muralidhar S, Pozniak J, Diaz JMS, Drier Y, Nguyen N, Lo JA, van Rooijen E, Kemeny LV, Zhan Y, Feng Y, Silkworth W, Powell CT, Liau BB, Xiong Y, Jin J, Newton-Bishop J, Zon LI, Bernstein BE, Fisher DE. Gain-of-Function Genetic Alterations of G9a Drive Oncogenesis. Cancer Discov. 2020;10(7):980-997. [CrossRef]
- Kelly GM, Al-Ejeh F, McCuaig R, Casciello F, Ahmad Kamal N, Ferguson B, Pritchard AL, Ali S, Silva IP, Wilmott JS, Long GV, Scolyer RA, Rao S, Hayward NK, Gannon F, Lee JS. G9a Inhibition Enhances Checkpoint Inhibitor Blockade Response in Melanoma. Clin Cancer Res. 2021;27(9):2624-2635. [CrossRef]
Figure 1.
How immunotherapy works (a) immune checkpoint inhibitors: Molecules designed to block immune checkpoints (Ex: CTLA4, PD-1, PD-L1 or LAG3) interactions with the specific ligands, (b) mRNA Vaccines: Encapsulated mRNA that target immune cells, leading to protein translation intending their processing and presentation to T cell via MHC class I and II aiming a subsequent immune response, (c) Tumor infiltrating Lymphocytes: A tumor fragment is removed from patient and cultured in Laboratory to a logarithmic expansion of the infiltrating T cell, intending to return them to the patient.
Figure 1.
How immunotherapy works (a) immune checkpoint inhibitors: Molecules designed to block immune checkpoints (Ex: CTLA4, PD-1, PD-L1 or LAG3) interactions with the specific ligands, (b) mRNA Vaccines: Encapsulated mRNA that target immune cells, leading to protein translation intending their processing and presentation to T cell via MHC class I and II aiming a subsequent immune response, (c) Tumor infiltrating Lymphocytes: A tumor fragment is removed from patient and cultured in Laboratory to a logarithmic expansion of the infiltrating T cell, intending to return them to the patient.
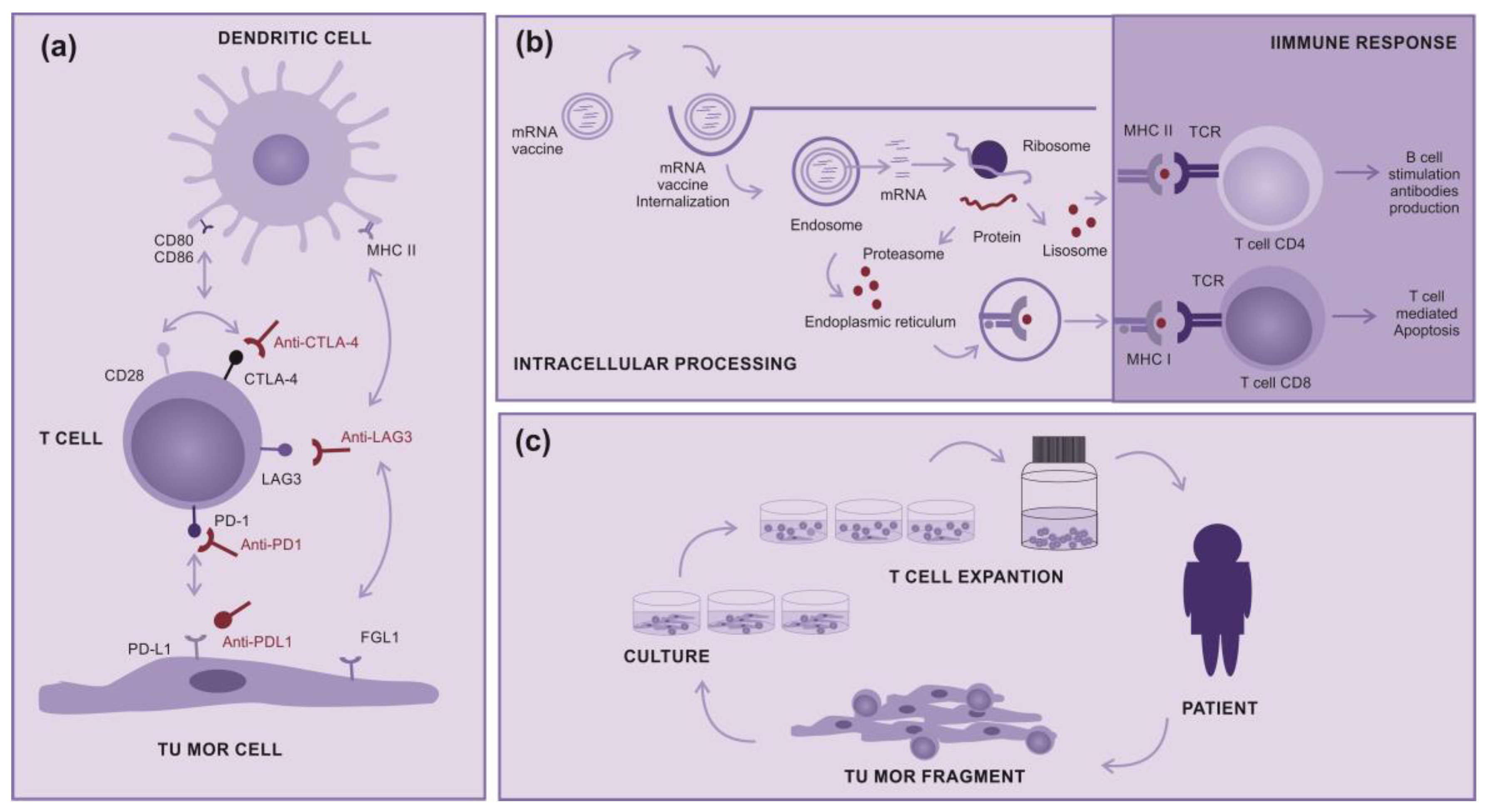
Figure 2.
How Target Therapy works (a) MAPK pathway normal signaling. The BRAF and MEK pathway initiates signaling through extracellular RTK domain. RAS family members are activated through a RTK ligand domain, and activation of RAS proteins bind BRAF isoforms leading to activation of BRAF, followed by MEK, and the final step of phosphorylation of ERK. The net effect leads to increased cell survival and decreased apoptosis, (b) BRAF or N-RAS mutant pathway. In the presence of an activating BRAFV600E mutation, BRAF no longer requires dimerization with RAS, and therefore remains constitutively active, similarly to NRAS mutations, maintaining this pathway constitutively activated leading an enhanced tumor cells proliferation with apoptosis abrogation (c) Inhibition of BRAF or N-RAS mutant pathway. The target therapy inhibits mutant BRAF or MEK, thereby stopping downstream activation of the MAPK pathway, decreasing cellular proliferation, and inducing apoptosis. Abbreviations: ERK, extracellular signal regulated kinase; MAPK, mitogen-activated protein kinase; BRAF, rapidly accelerated fibrosarcoma B type; RAS, rat sarcoma virus monologue; RTK, receptor tyrosine kinases.
Figure 2.
How Target Therapy works (a) MAPK pathway normal signaling. The BRAF and MEK pathway initiates signaling through extracellular RTK domain. RAS family members are activated through a RTK ligand domain, and activation of RAS proteins bind BRAF isoforms leading to activation of BRAF, followed by MEK, and the final step of phosphorylation of ERK. The net effect leads to increased cell survival and decreased apoptosis, (b) BRAF or N-RAS mutant pathway. In the presence of an activating BRAFV600E mutation, BRAF no longer requires dimerization with RAS, and therefore remains constitutively active, similarly to NRAS mutations, maintaining this pathway constitutively activated leading an enhanced tumor cells proliferation with apoptosis abrogation (c) Inhibition of BRAF or N-RAS mutant pathway. The target therapy inhibits mutant BRAF or MEK, thereby stopping downstream activation of the MAPK pathway, decreasing cellular proliferation, and inducing apoptosis. Abbreviations: ERK, extracellular signal regulated kinase; MAPK, mitogen-activated protein kinase; BRAF, rapidly accelerated fibrosarcoma B type; RAS, rat sarcoma virus monologue; RTK, receptor tyrosine kinases.
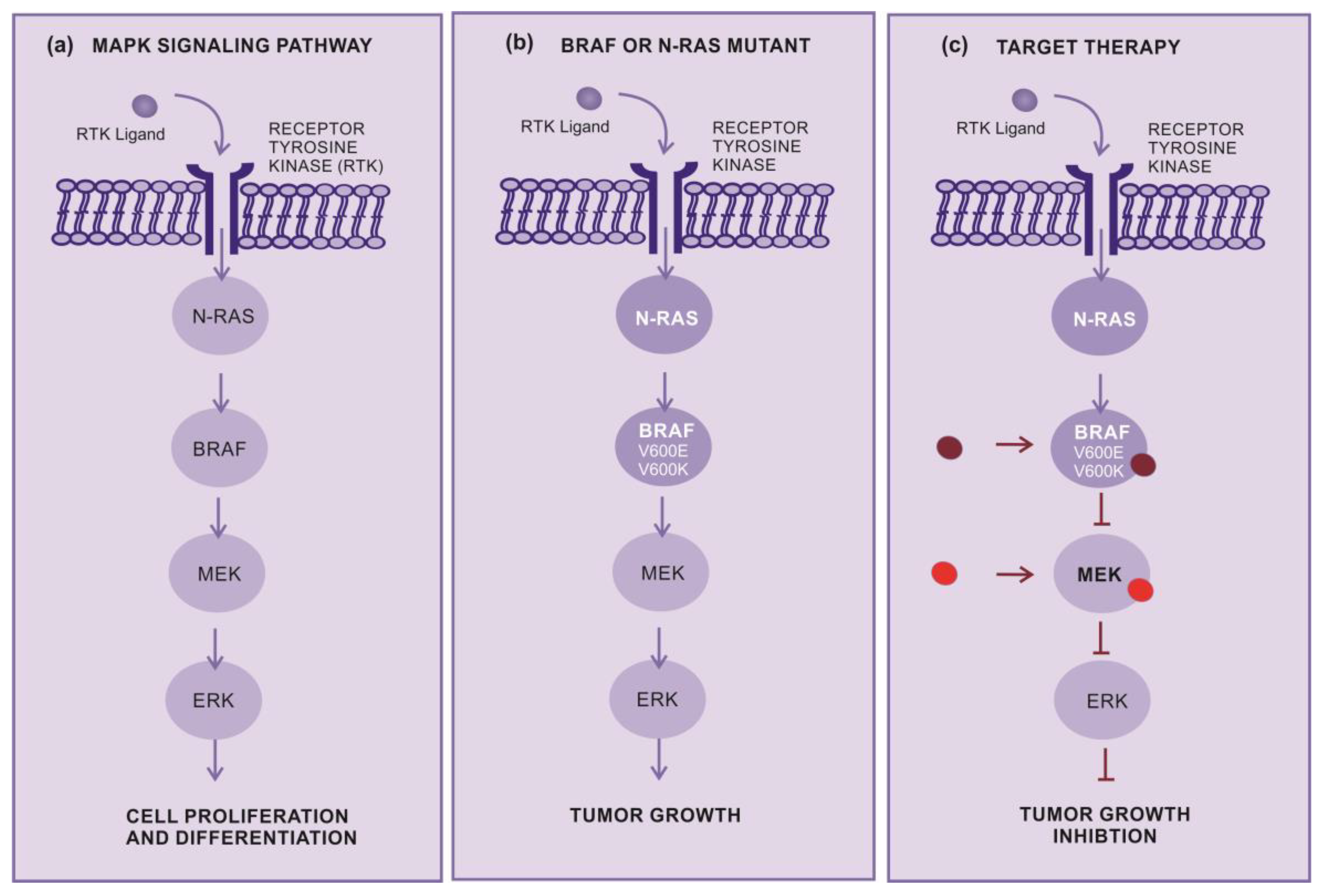
Figure 3.
Methyltransferases (a) DNA methyltransferase (DNMT) – An enzyme that transfer radical methyl to DNA nucleotides, (b) Protein methyltransferase (PMT) - An enzyme that transfer radical methyl to protein aminoacids. (c) Histone methyltransferase (HMT) - An enzyme that transfer radical methyl to histones. (d) G9a – G9a is histone methyltransferase responsible for histone H3 lysine 9 (H3K9) mono- and dimethylation.
Figure 3.
Methyltransferases (a) DNA methyltransferase (DNMT) – An enzyme that transfer radical methyl to DNA nucleotides, (b) Protein methyltransferase (PMT) - An enzyme that transfer radical methyl to protein aminoacids. (c) Histone methyltransferase (HMT) - An enzyme that transfer radical methyl to histones. (d) G9a – G9a is histone methyltransferase responsible for histone H3 lysine 9 (H3K9) mono- and dimethylation.
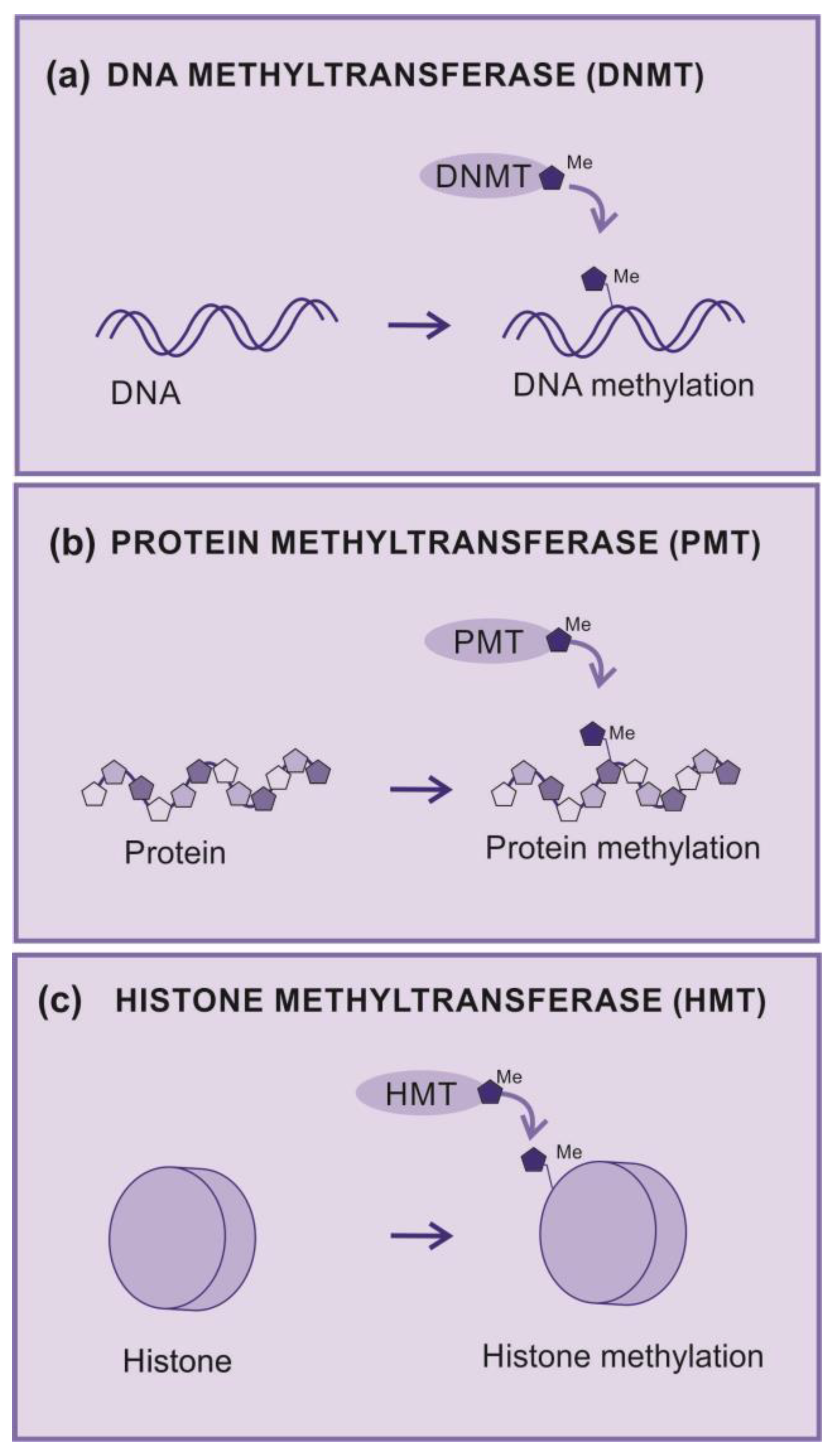
Figure 4.
Lysine Histone Methyltransferases (KMT) (a) KMT – Transfer of Methyl from a donor (SAM) to the Histone, (b) G9a-GLP complex – The complex G9a-GLP responsible for monomethylation of H3K9 histone, but G9a responsible for di and tri methylation of H3K9, (c) G9a activity– G9a under normoxia contributes for punctual dimethylation of H3K9 but under hypoxia increases the global dimethylation of H3K9 silencing the gene responsible for E-Cadherin expression, affecting the adhesion between cells, contributing for metastasis and, also activating Notch-1 signaling pathway contributing for cancer cell survival and proliferation.
Figure 4.
Lysine Histone Methyltransferases (KMT) (a) KMT – Transfer of Methyl from a donor (SAM) to the Histone, (b) G9a-GLP complex – The complex G9a-GLP responsible for monomethylation of H3K9 histone, but G9a responsible for di and tri methylation of H3K9, (c) G9a activity– G9a under normoxia contributes for punctual dimethylation of H3K9 but under hypoxia increases the global dimethylation of H3K9 silencing the gene responsible for E-Cadherin expression, affecting the adhesion between cells, contributing for metastasis and, also activating Notch-1 signaling pathway contributing for cancer cell survival and proliferation.
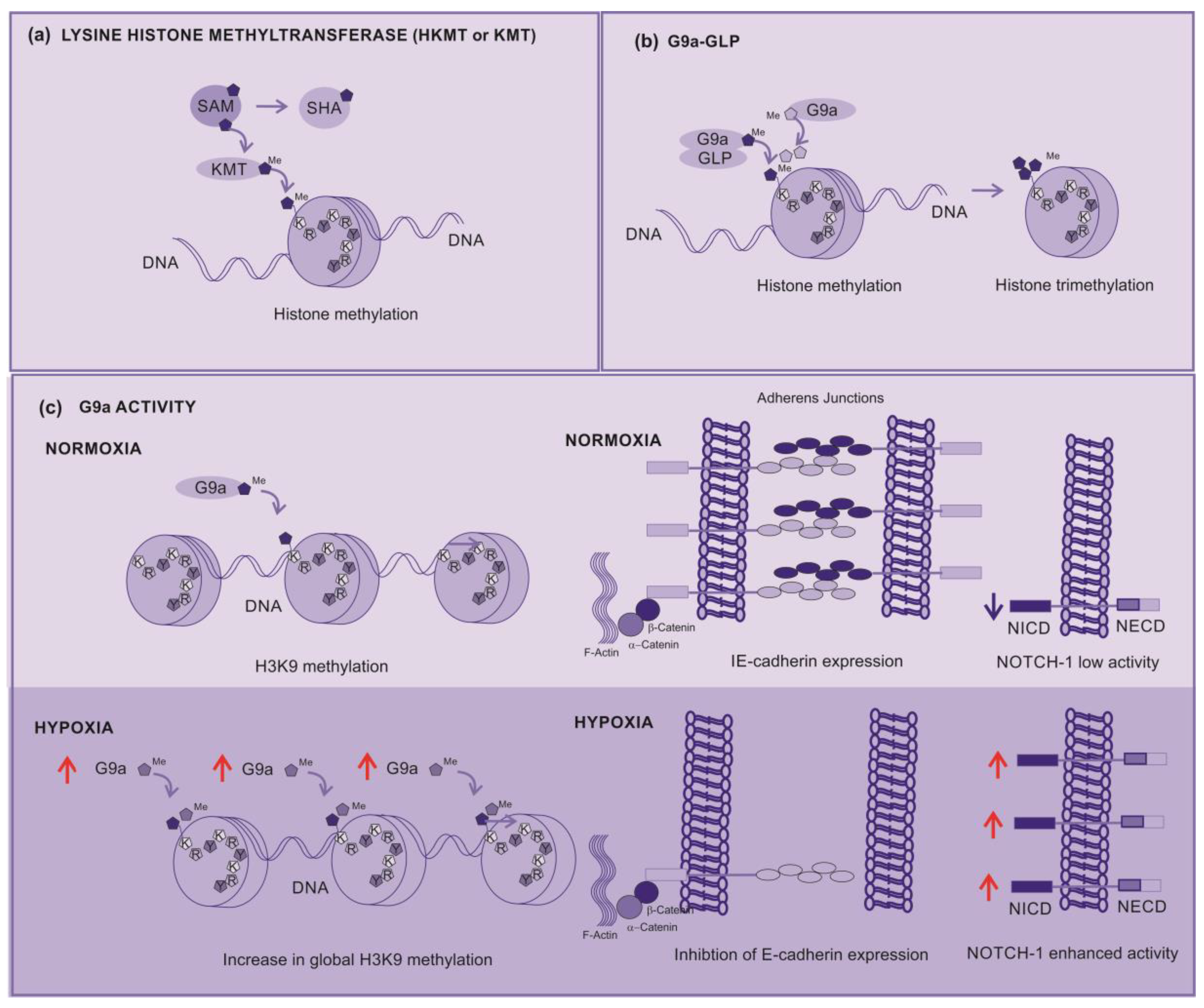
Figure 5.
KMT inhibitors (a) UNC0642 – An example of methylation inhibition by competitive block of G9a biding pocket, (b) G9a-GLP affected pathways – H3K9 dimethylation inhibit caspase 1 expression, G9a demethylates p53, inhibiting their activity, and enhancing the expression of polo-like kinase 1 (PLK1) implicated in cell proliferation and epithelial mesenchymal transition (EMT).
Figure 5.
KMT inhibitors (a) UNC0642 – An example of methylation inhibition by competitive block of G9a biding pocket, (b) G9a-GLP affected pathways – H3K9 dimethylation inhibit caspase 1 expression, G9a demethylates p53, inhibiting their activity, and enhancing the expression of polo-like kinase 1 (PLK1) implicated in cell proliferation and epithelial mesenchymal transition (EMT).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



