
Submitted:
19 October 2023
Posted:
20 October 2023
You are already at the latest version
Abstract
Abstract
Background: Gastric cancer, particularly adenocarcinoma, is a significant global health concern. Environmental risk factors, such as Helicobacter pylori infection and diet, play a role in its development.
Purpose: This study aimed to characterize the chemical composition and evaluate the in vitro antibacterial and antitumor activities of AOME. Additionally, morphological changes in gastric cancer cell lines were analyzed.
Study design: AOME was analyzed using HPLC-MS/MS and its antibacterial activity against H. pylori was assessed using the broth microdilution method. MIC and MBC values were determined, and positive and negative controls were included in the evaluation.
Methods: Anticancer effects were assessed through in vitro experiments using AGS, KATO-III, and SNU-1 cancer cell lines. The morphological changes were examined through SEM and TEM analyses.
Results: AOME contained several compounds, including caffeic acid, rutin, and hyperoside. The extract displayed significant antimicrobial effects against H. pylori, with consistent MIC and MBC values of 3.70 ± 0.09 mg/mL. AOME reduced cell viability in all gastric cancer cell in a dose and time-dependent manner.
Morphological analyses revealed significant ultrastructural changes in all tumor cells lines, suggesting the occurrence of cellular apoptosis.
Conclusions: This study demonstrated that AOME possesses antimicrobial activity against H. pylori and potent antineoplastic properties in gastric cancer cell lines. AOME holds promise as a natural resource for innovative nutraceutical approaches in gastric cancer management. Further research and in vivo studies are warranted to validate its potential clinical applications.
Keywords:
gastric cancer prevention
; nutraceuticals
; phytochemicals
; helicobacter pylori
; aristolochia olivieri
; apoptosis
1. Introduction
Gastric cancer includes various subtypes classified by WHO guidelines: adenocarcinoma, ring cell carcinoma, and undifferentiated carcinoma [1].
This pathology represents a leading cause of death globally, with a high incidence in East Asia, Eastern Europe, Central, and South America, and low incidence in other regions, including Italy [2,3].
Most gastric cancer are sporadic, affecting individuals aged 45 and above, with males higher incidence [4].
Environmental factors contributing to gastric cancer risk include H. pylori infection, diet, tobacco smoking, and inflammation [5,6].
Chemotherapy and surgery are essential treatments [7], while exploring new preventive measures, including food and vegetal substances inducing apoptosis, is a promising avanue.
Preclinical studies have shown promising results with various food substances [8]. Several phytocomplexes and isolated compounds have demonstrated significant anticancer activities against gastric cancer cell lines. Extracts from hibiscus displayed cytotoxic effects on gastric adenocarcinoma cells [9] without toxic effects on cardiovascular [10] and central nervous systems [11].
Some phytocomplexes have anticancer properties that, in certain cases, were ascribed to isolated compounds. For instance, an extract from Patrinia heterophylla Bunge roots selectively inhibited carcinoma SGC-7901 cells. The active compounds were identified as sarracenin and caffeic acid methyl ester [12]. Moreover, various food components have shown inhibitory effects on cancer cells. Polyphenols such as flavonoids, phenolic acids, stilbens, and diarylheptanoids have demonstrated inhibitory effects towards gastric cancer cells [13]. These findings align with epidemiological studies suggesting that a polyphenol-rich diet may decrease the risk of gastric cancer, particularly in females [14]. Further investigations on these natural compounds may offer promising avenues to design innovative chemopreventive strategies in gastric cancer.
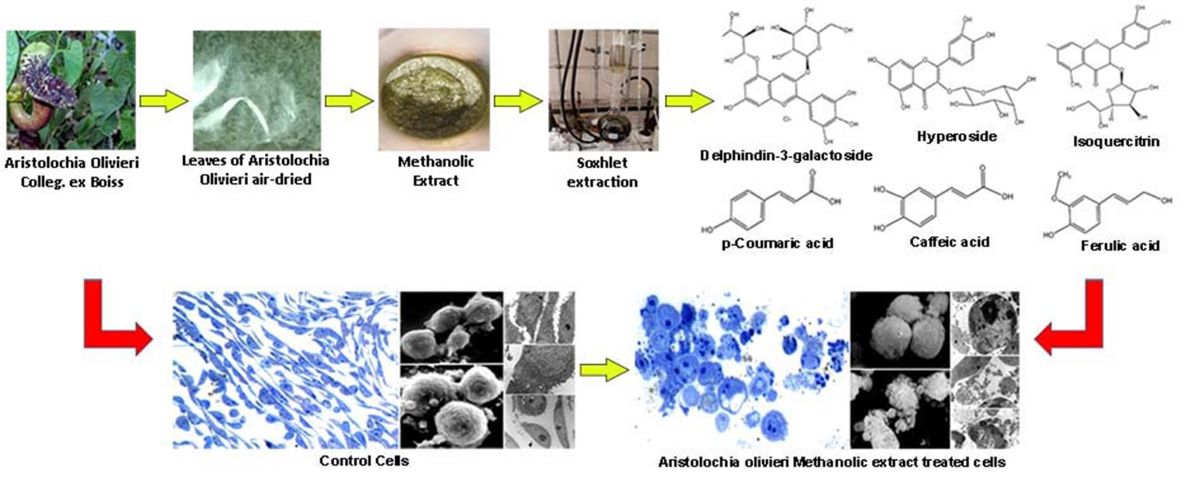
In our study, we conducted a comprehensive evaluation of the chemical composition and in vitro antibacterial and anticancer effects of a methanolic extract obtained from the aerial parts of Aristolochia olivieri Colleg. ex Boiss., a plant used in Kurdish folk medicine for gastro-intestinal ailments. The results revealed the presence of several phenolic acids and flavonoids. The experiments demonstrated significative anticancer activities through apoptosis. These findings highlight the potential of AOME as a novel resource for developing innovative approaches in the management of gastric cancer.
2. Material and Methods
2.1. Plant Material
The plant of Aristolochia olivieri Colleg. ex Boiss. was collected in April from Sakran Mountain, cho-man/Kurdistan region of northern Iraq. The voucher specimens, accession number (7705) were deposited in the Herbarium at Salahaddin University-Erbil, Iraq. The leaves were air-dried separately under shade at room temperature (20 –25 °C).
2.2. Preparation of Extract
The leaves of Aristolochia olivieri Colleg. ex Boiss. were dried for 48 hours in an oven at 40 °C. The dried plant material was then crushed and pulverized. Subsequently, 40 grams of this material were extracted using pure methanol (750 mL) in a Soxhlet apparatus for a period of 24 hours. The extract volume was concentrated using a rotary vacuum evaporator at a temperature of 40 °C until it formed a powder, which was collected and used for further experiments.
2.3. Reagents and Standards
Cyanidin-3-glucoside chloride, delphinidin-3,5-diglucoside chloride, delphinidin-3-galactoside chloride, petunidin-3-glucoside chloride, malvidin-3-galactoside chloride, quercetin-3-glucoside and kaempferol-3-glucoside were purchased from PhytoLab (Vestenbergsgreuth, Germany). The remaining 31 analytical standards of the 38 phenolic compounds were supplied by Sigma-Aldrich (Milan, Italy).
2.4. Analytical Characterization of AOME
Stock solutions (1000 mg L−1) of analytes were prepared in HPLC-grade methanol and stored at 4 °C (or -15 °C for anthocyanins). Daily, working solutions were made by dilution. Formic acid (99%) from Merck, hydrochloric acid (37%) from Carlo Erba Reagents, HPLC-grade methanol from Sigma-Aldrich, and deionized water (> 18 MΩ cm resistivity) purified using a Milli-Q SP Reagent Water System. All solvents and solutions were filtered through a 0.2 μm polyamide filter from Sartorius Stedim. Samples were further filtered with Phenex™ RC 4 mm 0.2 μm syringeless filter, Phenomenex, before HPLC analysis.
2.5. HPLC-ESI-MS/MS
HPLC-MS/MS analysis were conducted as previously reported [15].
2.6. Cell Culture and Treatments
The three commercial ATCC GC cell lines AGS, KATO-III, and SNU-1 were cultured as previously reported [16]. All cell lines were exposed to AOME extract dissolved in DMSO at different concentrations (7.4 mg/ml, 3.7 mg/ml, 1.9 mg/ml, and 0.9 mg/ml). Effects were evaluated after 24h, 48h, and 72h using cell viability, proliferation tests, and ultrastructural investigations.
2.7. Trypan Blue Dye Exclusion Assay
The effect of AOME on cell viability was determined using Trypan blue exclusion and the count of live and dead cells performed through a hemocytometer. Briefly, cells were washed and suspended (1.0 × 105 cells/mL) in a solution of 1× PBS: 0.5 mM EDTA (Life Technologies) containing 0.2% BSA (Sigma-Aldrich). Fifty (50) µL of the cell suspension was taken and mixed with an equal volume of 0.4% Trypan blue. The solution was mixed thoroughly and allowed to stand for 5 min at room temperature. Ten (10) µL of the solution was transferred to a hemocytometer, and both viable (clear) and dead (blue) cells were counted. The number of live cells divided by the total number of counted cells (clear and blue) gave the percent viability [17].
2.8. Cell Proliferation Assay
Cells were seeded in 96-well plates (5000 cells/well, 100 µL medium) and incubated for 24 h for adherence. Kato and AGS gastric cancer cell lines were then cultured with vehicle (DMSO 0.1%) or increasing AOME concentrations (0.47-3mg/ml) for 48 and 72 h. Cell growth was measured using PrestoBlue reagent (Invitrogen, Monza, Italy). After adding PrestoBlue (10 µL) and incubating for 2 h, absorbance was read at 600 nm using Infinite M200 photometer (Tecan Group Ltd., Switzerland). Three replicates per concentration were tested, with at least three independent experiments (bars, ± s.d.).
2.9. SEM
Different gastric cells after washing in 0.1 M phosphate buffer, were fixed in suspension with 2.5% glutaraldehyde. Afterwards, they were deposited on poly-L-lysine-coated coverslips overnight at 4 °C and processed as previously described [18].
2.10. TEM
AGS and KATO III cells were washed and fixed with 2.5% glutaraldehyde in 0.1 MPBS for 30 min, scraped and centrifuged at 300 x g for 10 min. The pellets were fixed in 2.5% glutaraldehyde for an additional 30 min. SNU1 cancer cells pellets were immediately fixed in 2.5% glutaraldehyde in 0.1 M in phosphate buffer (pH 7.3). All cells were post-fixed in 1% OsO4 for 1 h, alcohol dehydrated and embedded in araldite. Thin sections were stained and analyzed using a Philips CM10 transmission electron microscope [18].
2.11. Antibacterial Activity
AOME's antimicrobial activity against Helicobacter pylori was evaluated following CLSI guidelines with slight modifications. MIC and MBC were determined using the broth microdilution method. Extracts were prepared from the powder in sterile distilled water and twofold diluted in a 96-well microtiter plate (final concentrations: 0.19-25 mg/ml). H. pylori liquid culture was added, and plates were incubated for 72h at 37°C under microaerophilic conditions. MIC was the lowest concentration inhibiting visible bacterial growth, while MBC was determined by plating diluted cultures and incubating at 37°C for 48h under microaerophilic conditions. Negative and positive controls were included [19].
3. Results
3.1. AOME Chemical Characterization.
AOME exhibited diverse compounds, categorized into flavonoids and phenolic acids, measured in mg kg-1. Among flavonoids, delphinidin-3-galactoside had the highest concentration (19.85), followed by rutin (483.97), hyperoside (2540.33), isoquercitrin (986.27), delphinidin-3,5-diglucoside (867.66), and kaempferol-3-glucoside (342.52). Notably, other flavonoids like (+)-catechin, procyanidin B2, (-)-epicatechin, cyanidin-3-glucoside, petunidin-3-glucoside, quercitrin, myricetin, naringin, hesperidin, phloretin, kaempferol, and isorhamnetin were not detected.
Among phenolic acids, caffeic acid displayed the highest concentration (9867.16), followed by p-hydroxybenzoic acid (725.76), trans-cinnamic acid (603.99), gallic acid (17.07), neochlorogenic acid (14.49), 3-hydroxybenzoic acid, vanillic acid, resveratrol, syringic acid, and procyanidin A2 were not detected.
The total content of identified polyphenols in AOME was 30956.87 mg kg-1 (Table 1).
3.2. Antibacterial Activity
The AOME antimicrobial activity against Helicobacter pylori was investigated. DMSO was used as negative control. DMSO did not have any significant impact on Helicobacter pylori viability.
On the other hand, AOME showed antimicrobial effects against Helicobacter pylori. AOME MIC and MBC values were 3.70 ± 0.09 mg/mL. The two values were overlapping, indicating the phytocomplex ability to exert a cytotoxic effect at the tested concentrations.
These findings suggest that AOME possesses antimicrobial activity against Helicobacter pylori. The MIC and MBC values demonstrate AOME in vitro efficacy at relatively high concentrations.
3.3. The Antineoplastic Properties
The impact of AOME on AGS and KATO III cell viability was assessed in a dose and time-dependent manner (Figure 1, graphs 1 and 2). The response of SNU1 cells to AOME (graph 3) differed from the other cell lines. SNU1 cells displayed resistance to AOME within the initial 24 hours of treatment; however, their viability declined significantly after 48 hours of treatment, and cell death became evident upon AOME administration at its bactericidal dosage. The dosage of 7.4 mg/ml was excluded due to complete cell susceptibility (data not shown).
To further elucidate the effects of AOME on cell proliferation and survival over extended periods, KATO and AGS cells were exposed to increasing concentrations of AOME for 48 and 72 hours. The treatment significantly affected cell proliferation and viability.
The optimal cell treatment time for the apoptotic effects was 48 hours. KATO III cells showed a lower IC50 at 72 hours. IC50 values for AGS and KATO III cell lines at 48 hours were about 1.34 mg/ml and 1.28 mg/ml, respectively and at 72 hours were 1.45 mg/ml and 0.939 mg/ml, respectively. (Figure 2)
3.4. Ultrastructural Analyses
Morphological analyses confirmed the cytotoxic effect of AOME on gastric cancer cells, exhibiting a dose- and time-dependent response. However, the impact varied among the cell lines, particularly with SNU 1 showing greater resistance to AOME treatment (Figure 3).
AGS control demonstrated well-preserved proliferating cells (A, B, C, D, E, F), as evidenced by the number of metaphases (A). Cells appeared elongated with prominent nucleoli and diffuse chromatin in the nuclei (E, F). SEM images (B) showed intact cellular membranes. TEM analysis revealed well-preserved mitochondrial structures (E, inset F).
At lower and medium doses of AOME, metaphase cells decreased (G, L), but the cells maintained an elongated morphology. SEM showed the formation of blebs (H, I) at both concentrations. In the case of low concentration, the blebs were associated with autophagic vacuoles in the cytoplasm (K), while at medium concentration, micronuclei and chromatin condensation were observed (R, S). At medium doses, a small percentage of cells showed autophagic vacuoles and disrupted mitochondria (inset R).
At the bactericidal dose, rounding cells indicative of cytoskeleton rearrangement were observed (P), along with numerous blebs on the cell surface and a discontinuous membrane (M, Q). TEM images showed chromatin condensation typical of apoptotic cells, as well as micronuclei ejected from the cells, suggesting secondary necrosis (R, S).
Kato III cells (Figure 4) appeared elongated under control conditions (A, D, E, F), forming monolayers joined by junctions (E). The nuclear membrane was well preserved, although rare small blebs were occasionally observed (C, D). Cells gradually exhibited a rounding phenotype (G, M, P), becoming spherical at the bactericidal dose (S). Blebs appeared at all AOME concentrations (H, I, N, O, U) with higher doses showing discontinuity in the cell membrane (T). Numerous autophagic vacuoles were observed at low and medium doses (K, Q), accompanied by mitochondrial (R) damage and chromatin condensation (V). At high doses, many cells exhibited morphological features typical of secondary necrosis (W, Z).
SNU 1 cells (Figure 5) displayed a rounded morphology under control conditions (A, E, L, P) with well-preserved mitochondria (C). At low doses numerous autophagic vacuoles were observed (H, I). Several membrane blebbing appeared at all concentrations (F, M, Q). Bactericidal dose inducing membrane damage (Q) nuclear membrane detachment and chromatin condensation (N, O, R, S).
4. Discussion
Gastric cancer is a global health concern. Its development is influenced by various factors, including genetic predisposition, environmental exposures, diet and microbial infections [1].
The "exposome" encompasses life-long environmental exposures: diet, lifestyle, pollutants, and infectious agents. Helicobacter pylori infection is a major risk factor for gastric cancer, affecting over half the population [5]. Currently, investigating the potential anti H. pylori and anticancer properties of food and vegetal substances, such as AOME, represents a promising avenue for developing preventive strategies for gastric cancer prevention.
AOME chemical composition differs from other plants of the same genus, with varying concentrations of compounds across different species. AOME contains moderate concentrations of compounds like gallic acid, neochlorogenic acid, and delphindin-3-galactoside, ranging from 14.49 to 19.85 mg kg-1.
However, several compounds such as (+)-catechin, procyanidin B2, (-)-epicatechin, cyanidin-3-glucoside, and petunidin-3-glucoside were not detected. On the other hand, AOME exhibits significant concentrations of p-hydroxybenzoic acid (725.76 mg kg-1), 3-hydroxybenzoic acid (1160.66 mg kg-1), caffeic acid (9867.16 mg kg-1), and p-coumaric acid (9976.83 mg kg-1).
In our study, we explored the antibacterial and anticancer activities of AOME. The antibacterial activity against H. pylori occurred at a relatively high concentration (MIC and MBC values of 3.70 ± 0.09 mg/mL). As the high concentration makes it challenging to apply AOME directly as antibacterial agent, we aimed to investigate its in vitro effects against gastric cancer.
We observed significant alterations in cellular morphology, indicative of apoptotic effects. These alterations included prominent vacuolization, changes in mitochondrial structure, as well as evident signs of autophagy and mitophagy.
Observed vacuolization implies intracellular vesicle formation, linked to cellular stress and apoptosis pathways. Changes in mitochondrial morphology align with apoptotic cell death, indicating mitochondrial dysfunction [20]. Furthermore, the presence of autophagic vacuoles and mitophagic structures indicates that AOME treatment may trigger cellular self-degradation processes, suggesting a potential involvement of autophagy in apoptotic effects. These findings align with the growing evidence of crosstalk between apoptosis and autophagy pathways, where autophagy can either promote or suppress apoptosis depending on the cellular context [21]. It's important to highlight that our study focused on morphological changes, rather than investigating the specific molecular mechanisms underlying these effects. While we did not delve into the detailed molecular pathways, the observed alterations in cellular morphology provide valuable preliminary insights into AOME's potential role in inducing apoptosis in gastric cancer cells.
Comparing the IC50 values of individual compounds reported in the literature, with their concentrations at the IC50 value of AOME at 48 hours, caffeic acid, rutin, and quercetin may contribute to the observed effect of AOME.
Ultrastructural changes including blebs formation, chromatin margination and apoptotic bodies, support the apoptotic effects of AOME polyphenols. Indeed, apoptotic blebs are membrane protrusions occurring during apoptosis, induced by caffeic acid, quercetin, and rutin acting through a multitarget mechanism [22,23,24]. These three polyphenols share some key molecular pathways underlying the induction of apoptosis.
Specifically, these compounds promote apoptotic blebs formation, as they affect the expression of pro-apoptotic and anti-apoptotic proteins such as Bad, Bax, Mcl-1, and Bcl-2, leading to a delicate balance shift tipping the scales towards apoptosis in gastric cancer cells.
Additionally, they promote the production of ROS and ΔΨm. Mitochondrial dysfunction and loss of ΔΨm are key events in the apoptotic process, leading to the release of pro-apoptotic factors and the initiation of blebbing [25].
The significant alterations in cellular morphology due to AOME may result from the synergistic interaction among the various phytochemicals.
In summary, apoptotic blebs formation represents a morphological hallmark of the apoptotic process triggered by these compounds.
The other compounds were detected at low concentrations; however, we cannot completely exclude their contribution to AOME apoptotic effects. For instance, ellagic acid [26] p-coumaric acid [27], gallic acid [28] and quercetin [24] may act through complementary mechanisms, targeting different pathways involved in cell survival and proliferation of AGS cells.
Ellagic acid inhibits cell migration and induces apoptosis [26]. Gallic acid triggers apoptosis through the intrinsic pathway, involving the activation of caspases, up-regulation of pro-apoptotic proteins, and down-regulation of anti-apoptotic Bcl-2 family proteins. It also induces the expression of death receptors (Fas, FasL, and DR5) mediated by p53 [28]. Quercetin induces cell apoptosis in AGS cells by increasing ROS production, reducing ΔΨm, and altering apoptosis related-genes expressions [24]. This complementary action of quercetin in inducing apoptosis via ROS-mediated pathways, along with ellagic acid and gallic acid's effects on gene expressions and intrinsic apoptotic pathways, may offer a multifaceted approach to combat gastric cancer by targeting various aspects of the disease.
Ellagic and gallic acid induce apoptosis through different pathways: ellagic acid alters gene expression related to apoptosis, migration, and inflammation, while gallic acid triggers the intrinsic pathway via caspases and Bcl-2 family proteins. Quercetin, on the other hand, inhibits cells growth.
Synergy occurs when the combined effect of multiple compounds is greater than the sum of their individual effects. The presence of diverse bioactive compounds in the AOME extract could potentially create a synergistic network, where the interactions between different compounds amplify their overall cytotoxic and apoptotic effects, thereby increasing the potency of each molecule.
Overall, the observed apoptotic activity of the AOME extract in gastric cancer cells, despite many compounds being present at subactive concentrations, suggests a complex interplay and synergistic effects among the bioactive compounds.
In conclusion, AOME exhibits antimicrobial activity against H. pylori and potent antineoplastic properties in gastric cancer cell lines, showing promise as a natural resource for innovative nutraceutical approaches in gastric cancer management.
Further research is warranted to elucidate the precise mechanisms of action and synergistic interactions within the AOME extract, which could pave the way for the development of novel nutraceutical strategies contributing to gastric cancer prevention.
Author Contributions
MM and MB conceived and designed the experiments. IF, SS, SB, ABS, MC, IV, and MSP performed in vitro. DR performed microbiological assays. DP, GS, GC performed chemical analysis. MM, MB, SS, FOA analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Data Availability Statement
All data were generated in-house, and no paper mill was used. All authors agree to be accountable for all aspects of work ensuring integrity and accuracy.
Acknowledgments
The authors would like to thank Samuela Cappellacci, University of Urbino "Carlo Bo", Italy Authors used generative artificial intelligence (AI) and AI-assisted technologies to improve readability and language.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
Abbreviations
AOME: Aristolochia olivieri; ΔΨm, Mitochondrial potential membrane; MBC, Minimal bactericidal concentration, MIC, Minimal inhibitory concentration; ROS, reactive oxygen species; SEM, Scanning electron microscopy; TEM, Transmission Electron Microscopy.
References
- Nagtegaal, I. D.; Odze, R. D.; Klimstra, D.; Paradis, V.; Rugge, M.; Schirmacher, P.; Washington, K. M.; Carneiro, F.; Cree, I. A. WHO Classification of Tumours Editorial Board. The 2019 WHO classification of tumours of the digestive system. Histopathology. 2020, 76, 182–188. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Donida, B. M.; Tomasello, G.; Ghidini, M.; Buffoli, F.; Grassi, M.; Liguigli, W.; Maglietta, G.; Pergola, L.; Ratti, M. , Sabadini, G.; Toppo, L.; Ungari, M.; Passalacqua, R. Epidemiological, clinical and pathological characteristics of gastric neoplasms in the province of Cremona: the experience of the first population-based specialized gastric cancer registry in Italy. BMC cancer. 2019, 19, 212. [Google Scholar] [CrossRef]
- Cavatorta, O.; Scida, S.; Miraglia, C.; Barchi, A.; Nouvenne, A.; Leandro, G.; Meschi, T.; De' Angelis, G. L.; Di Mario, F. Epidemiology of gastric cancer and risk factors. Acta bio-medica: Atenei Parmensis. 2018, 89, 82–87. [Google Scholar] [PubMed]
- Yang, L.; Ying, X.; Liu, S.; Lyu, G.; Xu, Z.; Zhang, X.; Li, H.; Li, Q.; Wang, N.; Ji, J. Gastric cancer: Epidemiology, risk factors and prevention strategies. Chinese journal of cancer research = Chung-kuo yen cheng yen chiu, 2020, 32, 695–704. [Google Scholar] [CrossRef]
- Farinati, F.; Cardin, R.; Cassaro, M.; Bortolami, M.; Nitti, D.; Tieppo, C.; Zaninotto, G.; Rugge, M. Helicobacter pylori, inflammation, oxidative damage and gastric cancer: a morphological, biological and molecular pathway. European journal of cancer prevention: the official journal of the European Cancer Prevention Organisation (ECP), 2008, 17, 195–200. [Google Scholar] [CrossRef]
- Wagner, A. D.; Syn, N. L.; Moehler, M.; Grothe, W.; Yong, W. P.; Tai, B. C.; Ho, J.; Unverzagt, S. Chemotherapy for advanced gastric cancer. Cochrane Database Syst. Rev. 2017, 8, CD004064. [Google Scholar] [CrossRef]
- Chiaino, E.; Micucci, M.; Cosconati, S.; Novellino, E.; Budriesi, R.; Chiarini, A.; Frosini, M. Olive Leaves and Hibiscus Flowers Extracts-Based Preparation Protect Brain from Oxidative Stress-Induced Injury. Antioxidants (Basel, Switzerland), 2020, 9, 806. [Google Scholar] [CrossRef]
- Lin, H.H.; Huang, H.P.; Huang, C.C.; Chen, J.H.; Wang, CJ. Hibiscus polyphenol-rich extract induces apoptosis in human gastric carcinoma cells via p53 phosphorylation and p38 MAPK/FasL cascade pathway. Mol. Carcinog. 2005, 43, 86–99. [Google Scholar] [CrossRef]
- Micucci, M.; Malaguti, M. , Toschi, T. G., Di Lecce, G., Aldini, R., Angeletti, A., Chiarini, A., Budriesi, R., Hrelia, S., 2015, Cardiac and Vascular Synergic Protective Effect of Olea europea L. Leaves and Hibiscus sabdariffa L. Flower Extracts. Oxidative Med. Cell. Longev. 2015, 318125. [Google Scholar]
- Chiaino, E. , Micucci, M.; Durante, M.; Budriesi, R.; Gotti, R.; Marzetti, C.; Chiarini, A.; Frosini, M., Apoptotic-Induced Effects of Acacia Catechu Willd. Extract in Human Colon Cancer Cells. Int. J. Mol. Sci. 2020, 21, 2102. [Google Scholar] [CrossRef] [PubMed]
- Sheng, L.; Yang, Y.; Zhang, Y.; Li, N. Chemical constituents of Patrinia heterophylla Bunge and selective cytotoxicity against six human tumor cells. J Ethnopharmacol. 2019, 236, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Li, M. , Zheng, Y.; Zhao, J., Liu, M., Shu, X., Li, Q., Wang, Y, Eds.; Zhou, Y. Polyphenol Mechanisms against Gastric Cancer and Their Interactions with Gut Microbiota: A Review. Current oncology 2022. (Toronto, Ont.) 29, 5247–5261. [Google Scholar]
- Fagundes, M.A.; Silva, A.R.C.; Fernandes, G.A.; Curado, M.P. Dietary Polyphenol Intake and Gastric Cancer: A Systematic Review and Meta-Analysis. Cancers (Basel). 2022, 14, 5878. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, A. M.; Angeloni, S.; Abouelenein, D.; Acquaticci, L.; Xiao, J.; Sagratini, G.; Maggi, F.; Vittori, S.; Caprioli, G. A new HPLC-MS/MS method for the simultaneous determination of 36 polyphenols in blueberry, strawberry and their commercial products and determination of antioxidant activity. Food chemistry. 2022, 367, 130743. [Google Scholar] [CrossRef] [PubMed]
- Tedaldi, G.; Molinari, C.; São José, C. , Barbosa-Matos, R.; André, A.; Danesi, R.; Arcangeli, V.; Ravegnani, M.; Saragoni, L.; Morgagni, P.; Rebuzzi, F.; Canale, M.; Pignatta, S.; Ferracci, E.; Martinelli, G.; Ranzani, G.N.; Oliveira, C.<; Calistri, D.; Ulivi, P. 2021. Genetic and Epigenetic Alterations of CDH1 Regulatory Regions in Hereditary and Sporadic Gastric Cancer. Pharmaceuticals (Basel). 2021, 14, 457. [Google Scholar]
- Salucci, S.; Bavelloni, A. ; Stella,; A. B; Fabbri, F.; Vannini, I.; Piazzi, M.; Volkava, K.; Scotlandi, K.K Martinelli, G.; Faenza, I.; Blalock, W.; 2023. The Cytotoxic Effect of Curcumin in Rhabdomyosarcoma Is Associated with the Modulation of AMPK, AKT/mTOR, STAT, and p53 Signaling. (2023). Nutrients 2023, 15, 740. [Google Scholar]
- Battistelli, M.; Salucci, S.; Guescini, M.; Curzi, D., Stocchi. Skeletal Muscle Cell Behavior After Physical Agent Treatments. Curr. Pharm. Des. 2015, 21, 3665–3672. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 2018. 11th ed.; M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA.
- Maltese, W.A.; Overmeyer, J.H. Methuosis: nonapoptotic cell death associated with vacuolization of macropinosome and endosome compartments. Am. J. Pathol. 2014, 184, 1630–42. [Google Scholar] [CrossRef] [PubMed]
- Kma, L.; Baruah, T.J. , . The interplay of ROS and the PI3K/Akt pathway in autophagy regulation. Biotechnol. Appl. Biochem 2021, 69, 248–264. [Google Scholar] [CrossRef]
- Karagoz, L.; Şenturk, M.; Sagir, T.; Karagoz, Y.; Batmax, O.S.; Burban URAS, F., 2023. Investigation of Caffeic Acid Effect on Human Cancer Cell Line and Some Enzymes, Eastern Anatolian. Journal of Science Volume IX, Issue I, 33-42.
- Ghavami, G.; Muhammadnejad, S.; Amanpour, S.; Sardari, S. Bioactivity Screening of Mulberry Leaf Extracts and two Related Flavonoids in Combination with Cisplatin on Human Gastric Adenocarcinoma Cells. Iranian journal of pharmaceutical research: IJPR, 2020, 19, 371–382. [Google Scholar]
- Shang, H. S., Lu, H. F., Lee, C. H., Chiang, H. S., Chu, Y. L., Chen, A., Lin, Y. F., Chung, J. G., 2018. Quercetin induced cell apoptosis and altered gene expression in AGS human gastric cancer cells. Environmental toxicology 33, 1168–1181.
- Zheng, L. C.; Yang, M. D.; Kuo, C. L.; Lin, C. H.; Fan, M. J.; Chou, Y. C. , Lu H. F.; Huang, W. W.; Peng, S. F. Chung, J. G. Norcantharidin-induced Apoptosis of AGS Human Gastric Cancer Cells Through Reactive Oxygen Species Production, and Caspase- and Mitochondria -dependent Signaling Pathways. Anticancer research. 2012, 36, 6031–6042. [Google Scholar] [CrossRef] [PubMed]
- Cheshomi, H.; Bahrami, A. R; Rafatpanah, H.; Matin, M. M. The effects of ellagic acid and other pomegranate (Punica granatum L.) derivatives on human gastric cancer AGS cells. Human & experimental toxicology. 2022, 41, 9603271211064534. [Google Scholar]
- Radziejewska, I.; Supruniuk, K.; Tomczyk, M.; Izdebska, W.; Borzym-Kluczyk, M.; Bielawska, A.; Bielawski, K.; Galicka, A. 2022. p-Coumaric acid, Kaempferol, Astragalin and Tiliroside Influence the Expression of Glycoforms in AGS Gastric Cancer Cells. International journal of molecular sciences. 2022, 23, 8602. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C. L.; Chiu, Y. M.; Ho, T. Y; Hsieh, C. T.; Shieh, D. C.; Lee, Y. J.; Tsay, G. J., Wu. Gallic Acid Induces Apoptosis in Human Gastric Adenocarcinoma Cells. Anticancer research. 2018, 38, 2057–2067. [Google Scholar]
Figure 1.
AOME treatment leads to decreased cell proliferation/viability in gastric cell lines. .
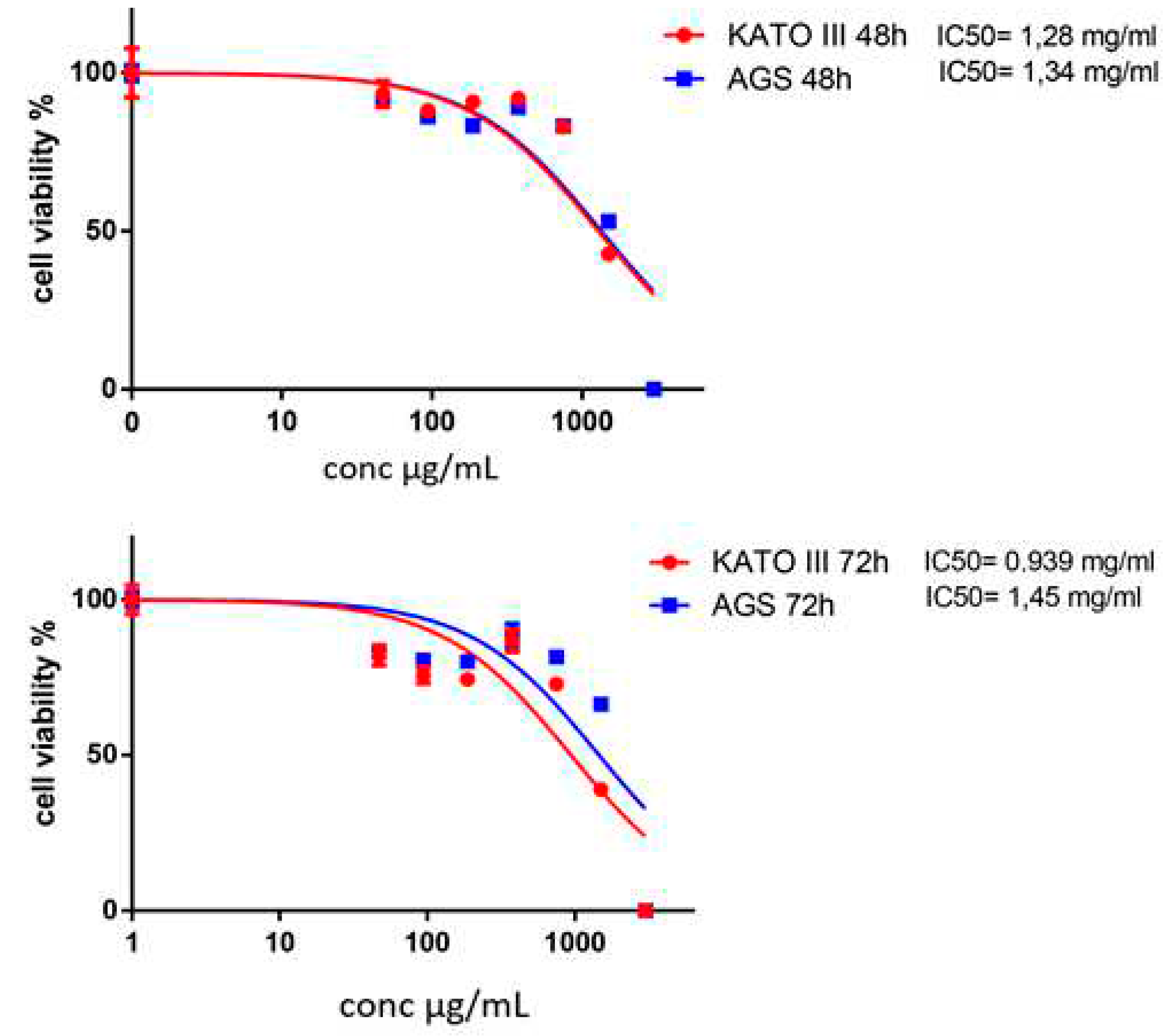
Figure 2.
Effects of AOME on Gastric Cancer Cells: In vitro investigation of AOME's impact on the proliferation and apoptosis of AGS, KATO III, and SNU1 cells. Gastric cancer cells were treated with AOME at concentrations of 0.9, 1.9, and 3.7 mg/ml for 24 or 48 hours. (A) Colony Formation Assays were performed to assess the proliferation of AOME-treated cells. (B and C) The data presented represent the mean ± SD of two independent in vitro experiments.
Figure 2.
Effects of AOME on Gastric Cancer Cells: In vitro investigation of AOME's impact on the proliferation and apoptosis of AGS, KATO III, and SNU1 cells. Gastric cancer cells were treated with AOME at concentrations of 0.9, 1.9, and 3.7 mg/ml for 24 or 48 hours. (A) Colony Formation Assays were performed to assess the proliferation of AOME-treated cells. (B and C) The data presented represent the mean ± SD of two independent in vitro experiments.
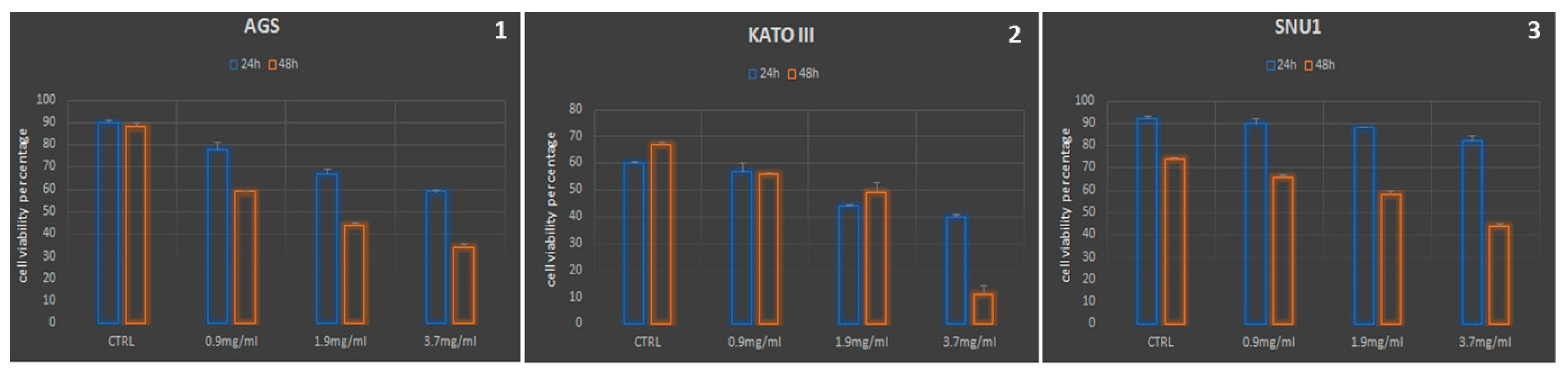
Figure 3.
Optical A, G, L, P, SEM B, C, D, H, I, M, Q, and TEM E, F, J, K, N, O, R, S analyses of AGS cells treated with different AOME doses. A, G, L, P, Bar= 20µm; B, C, D, H, I, M, Q, Bar= 10µm; E, F, J, K, N, O, R, S, Bar=2µm.
Figure 3.
Optical A, G, L, P, SEM B, C, D, H, I, M, Q, and TEM E, F, J, K, N, O, R, S analyses of AGS cells treated with different AOME doses. A, G, L, P, Bar= 20µm; B, C, D, H, I, M, Q, Bar= 10µm; E, F, J, K, N, O, R, S, Bar=2µm.
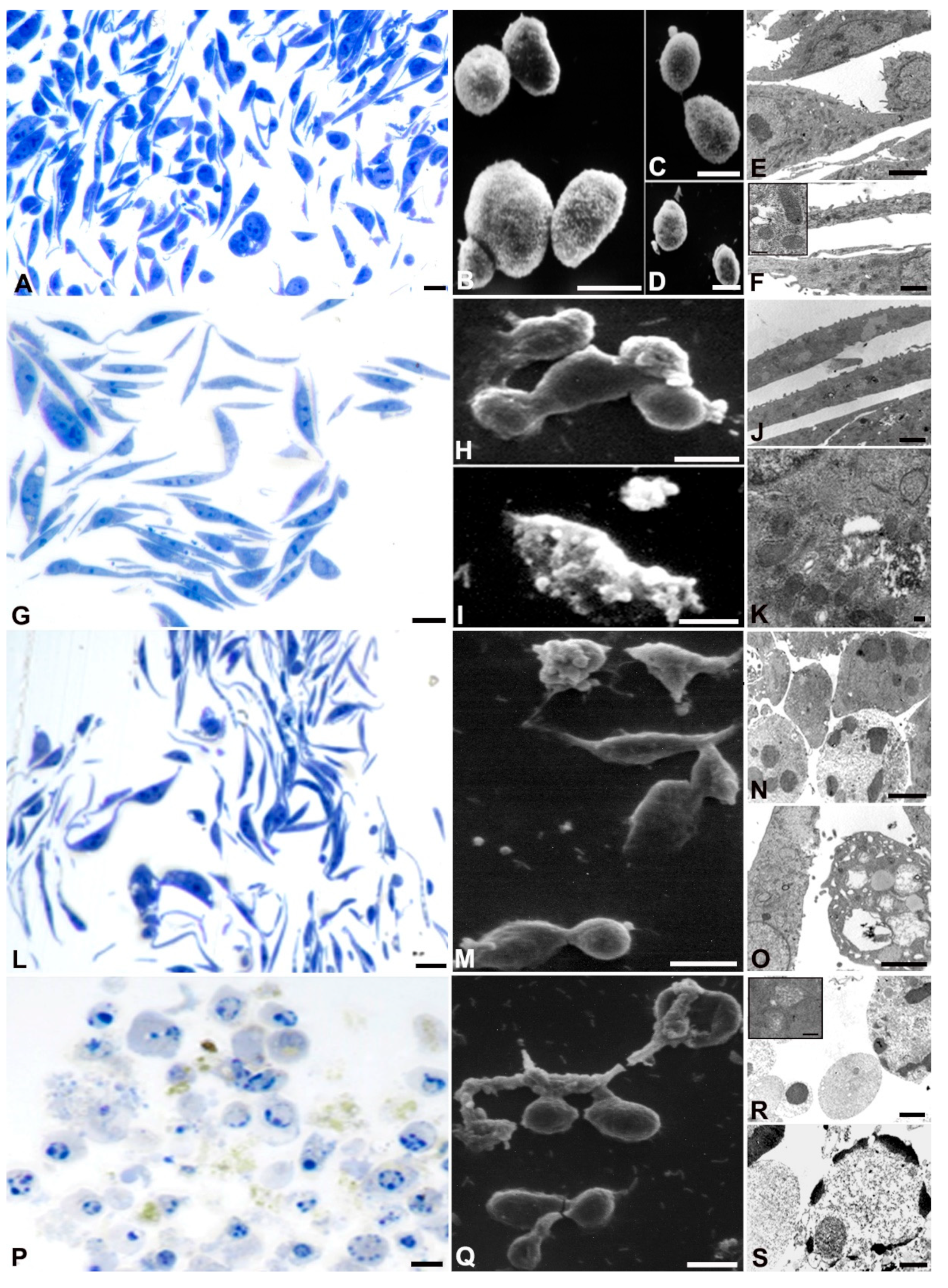
Figure 4.
Optical A, G, M, S, SEM B, C, H, I, N, O, T, U and TEM D, E, F, J, K, L, P, Q, R, V, W, Z analyses of KATO III cells treated with different AOME doses. A, G, M, S, Bar= 20µm; B, C, H, I, N, O, T, U, Bar= 10µm; D, F, J, V, W, Z, Bar=5µm; E, Bar 2µm; K, Q, R, Bar= 1µm.
Figure 4.
Optical A, G, M, S, SEM B, C, H, I, N, O, T, U and TEM D, E, F, J, K, L, P, Q, R, V, W, Z analyses of KATO III cells treated with different AOME doses. A, G, M, S, Bar= 20µm; B, C, H, I, N, O, T, U, Bar= 10µm; D, F, J, V, W, Z, Bar=5µm; E, Bar 2µm; K, Q, R, Bar= 1µm.
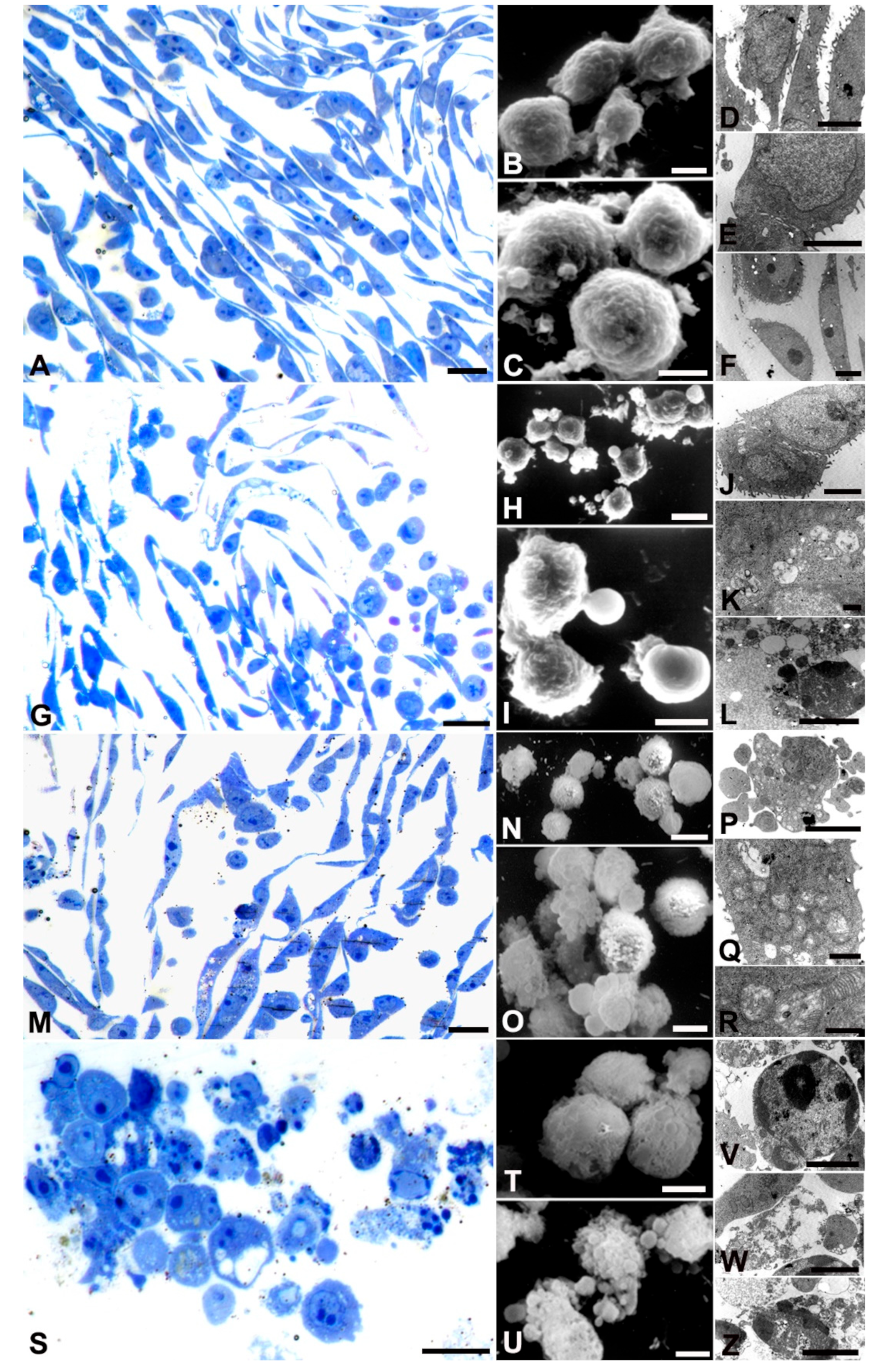
Figure 5.
Optical A, E, L, P, SEM B, F, M, Q and TEM C, D, G, H, I, N, O, R, S analyses of SNU 1 cells treated with different AOME doses. A, E, L, P, Bar= 20µm; B, F, M, Q, Bar= 5µm; C, G, H, I, Bar=1µm; D, N, O, R, Bar= 2µm.
Figure 5.
Optical A, E, L, P, SEM B, F, M, Q and TEM C, D, G, H, I, N, O, R, S analyses of SNU 1 cells treated with different AOME doses. A, E, L, P, Bar= 20µm; B, F, M, Q, Bar= 5µm; C, G, H, I, Bar=1µm; D, N, O, R, Bar= 2µm.
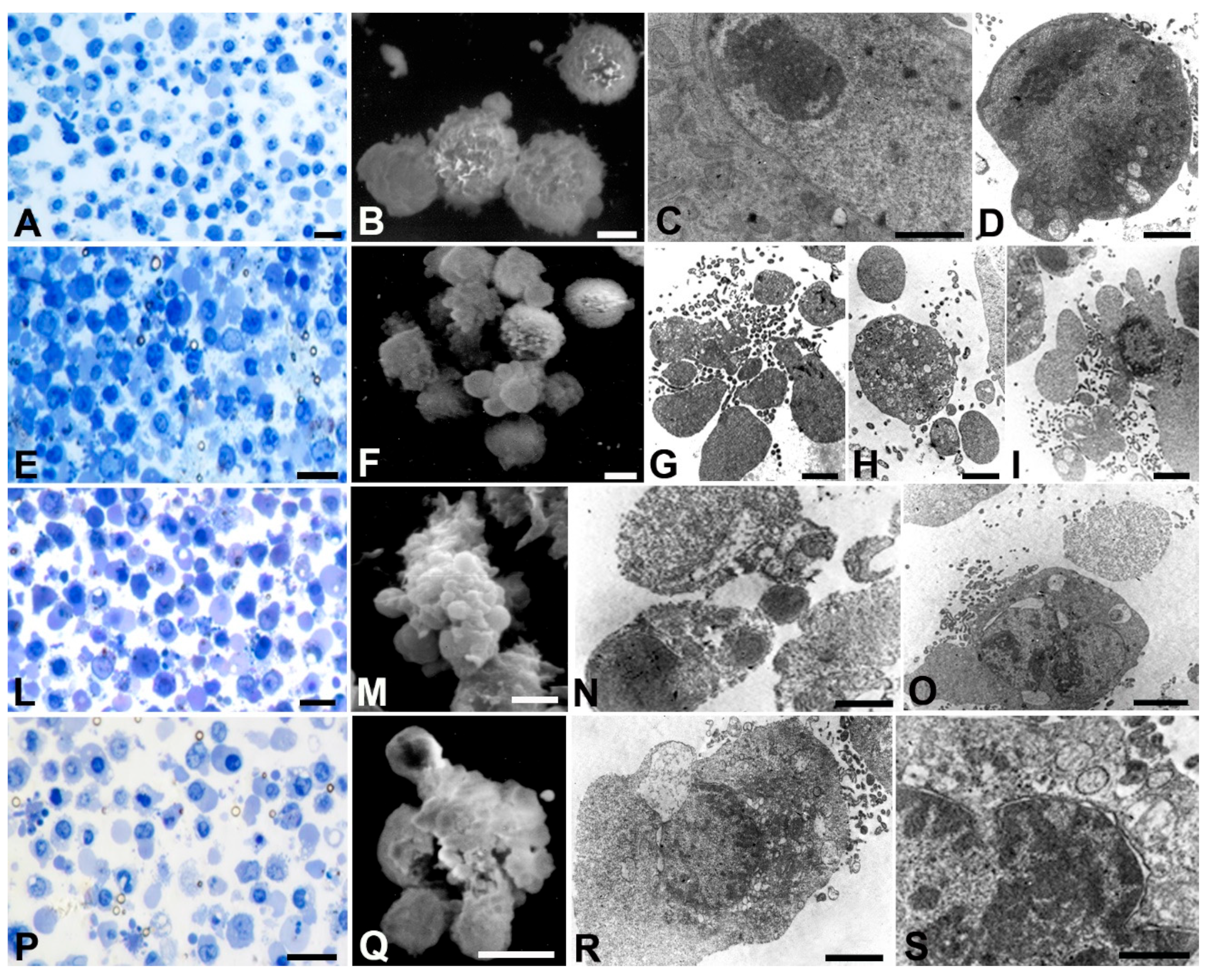

Table 1.
Concentration (mg kg-1 of dried extract) of bioactive compounds in AOME.
| No. | Compounds | Concentration mg kg-1 |
|---|---|---|
| 1 | Gallic acid | 17.07 |
| 2 | Neochlorogenic acid | 14.49 |
| 3 | Delphindin-3-galactoside | 19.85 |
| 4 | (+)-Catechin | n.d. |
| 5 | Procyanidin B2 | n.d. |
| 6 | Chlorogenic acid | 44.52 |
| 7 | p-Hydroxybenzoic acid | 725.76 |
| 8 | (-)-Epicatechin | n.d. |
| 9 | Cyanidin-3-glucoside | n.d. |
| 10 | Petunidin-3-glucoside | n.d. |
| 11 | 3-Hydroxybenzoic acid | 1160,66 |
| 12 | Caffeic acid | 9867,16 |
| 13 | Vanillic acid | n.d. |
| 14 | Resveratrol | n.d. |
| 15 | Pelargonidin-3-glucoside | n.d. |
| 16 | Pelagonidin-3-rutinoside | n.d. |
| 17 | Malvidin-3-galactoside | n.d. |
| 18 | Syringic acid | n.d. |
| 19 | Procyanidin A2 | n.d. |
| 20 | p-Coumaric acid | 9976.83 |
| 21 | Ferulic acid | 2696.19 |
| 22 | 3,5-Dicaffeoylquinic acid | n.d. |
| 23 | Rutin | 483.97 |
| 24 | Hyperoside | 2540.33 |
| 25 | Isoquercitrin | 986.27 |
| 26 | Delphindin-3,5-diglucoside | 867.66 |
| 27 | Phloridzin | 2.64 |
| 28 | Quercitrin | n.d. |
| 29 | Myricetin | n.d. |
| 30 | Naringin | n.d. |
| 31 | Kaempferol-3-glucoside | 342.52 |
| 32 | Hesperidin | n.d. |
| 33 | Ellagic acid | 428.46 |
| 34 | trans-cinnamic acid | 603.99 |
| 35 | Quercetin | 90.59 |
| 36 | Phloretin | n.d. |
| 37 | Kaempferol | 81.17 |
| 38 | Isorhamnetin | 5.73 |
| Total content | 30956.87 | |
a n.d., not detectable. Relative standard deviation (RSD) for all compounds ranged from 3.51 to 9.14%.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



