
Submitted:
22 October 2023
Posted:
23 October 2023
You are already at the latest version
Abstract
(1) Background: The severity of malaria is associated with low bioavailability of antioxidants and high concentration of free radicals that induce oxidative damage in cerebral and pulmonary microcirculation. This can be prevented by the action of consumable antioxidants present in foods. Therefore, we investigated the protective role of lycopene (LYC) on the oxidative changes induced by Plasmodium berghei (Pb); (2) Methods: Mice were infected by intraperitoneal injection of 106 parasitized red blood cells and treated via gavage with LYC (3.11 mg/kg bw/day) or NAC (62 mg/kg bw/day). They were then evaluated for 1, 4, 8 or 12 days after infection. Levels of thiobarbituric acid reactive substances (TBARS), antioxidant capacity by inhibition of ABTS radicals (AC-ABTS) and DPPH (AC-DPPH), uric acid (UA), and nitric oxide (NO) were measured in brain and lung tissues; (3) Results: The infection caused oxidative stress confirmed by increased levels of TBARS, AC-ABTS, AC-DPPH, UA, and NO in the tissues leading to the death of the animals. LYC prevented the increase in TBARS, AU, and NO levels compared to Pb (p< 0.0001) and NAC+Pb groups (p<0.0001), reaching values similar to those of Sham animals; (4) Conclusions: These results are striking evidence of the beneficial effect of lycopene supplementation on oxidative stress in experimental malaria in vivo and emphasize the importance of antioxidant supplementation in the treatment of the disease.
Keywords:
Lycopene
; N-Acetylcysteine
; Malaria
; Oxidative stress
; Antioxidant
1. Introduction
Free radicals are intermediate species with unpaired electrons. The production of these molecules in the human body is inevitable, since many of these molecules originate from the process of physicochemical oxidation of molecular oxygen (O2) and/or molecular nitrogen (N2), leading to the production of reactive oxygen and nitrogen species (RONS) [1]. RONS are highly oxidizing intermediates produced continuously in biological systems from biochemical reactions, including nitric oxide (NO) synthesis and mitochondrial electron transport chain reactions, as well as by metal-catalyzed oxidation and reduction, among others [2]
The main RONS generated in the human body are superoxide anion (O2•-), hydrogen peroxide (H2O2), hydroxyl radical (OH•), singlet oxygen (1O2), NO, nitrogen dioxide (NO2) and peroxynitrite (ONOO−) [3]. There are other biologically important free radicals such as lipid hydroperoxide (ROOH), lipid peroxyl radical (ROO), and lipid alkoxyl radical (RO).
The unpaired electrons give RONS high instability and reactivity. As a result, they have a short half-life of milliseconds or less, but enough time to trigger intermolecular interactions, as well as responses in nearby target cells, leading to oxide-reduction reactions, altering and/or damaging biomolecules, cells and tissues [4].
In response to the production of RONS, the human body induces the synthesis of mobilizable antioxidants that act by preventing, neutralizing or reducing the oxide-reduction reactions of RONS in the body [5,6]. Three enzymes are critical in this process, including superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GSH-Px). These mobilizable antioxidants, respectively, neutralize O2•- and break down H2O2 or ROOH into harmless molecules such as H2O, alcohol, and O2 [7].
However, when these antioxidants are not at adequate and sufficient levels to compensate for the harmful effects of RONS, oxidative stress is installed in the body. Oxidative stress is therefore an imbalance between RONS and antioxidants in favor of RONS [8]. Oxidative stress is involved in: 1- Beneficial effects, such as the body's immune defense system through toxic action on invading pathogens; 2- Harmful effects, leading to an interruption of redox signaling and control and/or molecular damage in chronic, degenerative, neurodegenerative, metabolic, and infectious diseases [4,7].
In this context, studies have discussed the role of RONS and antioxidant defense mechanisms in malaria [9] Malaria is a potentially serious disease, causing more than 200 million malaria episodes and about 500,000 deaths annually (Figure 1), mostly in impoverished communities [10] Among others, Plasmodium falciparum and Plasmodium vivax pose the greatest threat to human health.
Studies have associated the action of RONS with the various pathological manifestations in malaria, which can range from nonspecific symptoms, such as fever and mild anemia, to the complications of severe malaria, which include coma, prostration, respiratory difficulty, metabolic acidosis, renal failure, liver damage, and severe anemia [11,12,13]
According to SUZUKI et al. [14] the pathophysiological changes evident in malaria infection occur because mobilizable antioxidants are drastically reduced or consumed in oxidation-reduction reactions with RONS during infection.
Other factors evidenced in the infection are preponderant for the high production of RONS, including ischemia and reperfusion syndrome (IRS), parasite metabolism, as well as body's immune response against the parasite itself [5,6,12]
During infection, the parasites invade red blood cells and feed on hemoglobin, their main source of amino acids. As a consequence, red blood cells lose their function due to severe hemolysis. This eventually releases heme (Fe2+) into the circulation. Heme can react with O2 and generate O2•−, subsequently increasing H2O2 levels through spontaneous dismutation, as well as increasing HO• levels [15]. The increase in these RONS mediates the oxidation of lipids, proteins, and DNA, resulting in cellular and tissue damage, endothelial dysfunction, and loss of vascular homeostasis [16,17]
RONS can also activate toll-like receptor-4 (TLR4), triggering immune response. According to Ty et al. [18], RONS are a key factor into triggering the immune response during infection. According to the authors, immune cells are activated, including neutrophils and macrophages, which engulf and kill the parasites through the respiratory burst. One of the main responsible for this action is NO.
NO is a gaseous free radical, with an extremely short half-life in tissues, ranging from 3 to 10 seconds, synthesized from L-arginine by the enzyme nitric oxide synthase (NOS). In macrophages NO synthesis is usually low, however, in response to infection, inducible nitric oxide synthase (iNOS) is activated, which can result in large amounts of NO produced over hours or days [19].
In addition to macrophages, endothelial, and neuronal cells synthesize NO, but at a constant rate, regardless of physiological demand. In endothelial cells, NO is synthesized by endothelial NOS (eNOS), acting as an important vasodilator, being able to increase the vascular permeability of smooth muscles [20] Meanwhile, in neuronal cells, neuronal nitric oxide synthase (nNOS), releases NO which can act as a neurotransmitter [21].
However, in malaria the high production of NO favors its reaction with O2•−, even at low concentrations, to produce highly oxidizing ONOO−, which can act locally on the underlying vascular smooth muscle or on monocytes or platelets, promoting apoptosis in the microvascular endothelium and cytoadherence [22]. Localized action occurs because the parasites express cytoadhesion factors on the surface of infected red blood cells, allowing the binding and sequestration of red blood cells in the postcapillary venules, promoting vascular occlusion and tissue hypoxia [23]. Another important point to be considered is the fact that NO can interact reversibly with hemoglobin, which acts as an O2 sensor and regulator of vascular tone in response to local O2 partial pressure, binding to heme, generating nitrate and methemoglobin, which is unable to bind and carry O2 for tissue perfusion, further potentiating ischemia, hypoxia, and anemia [24].
Thus, phagocytosis and the consequent action of RONS including O2•− and NO, as well as other toxic products, can aggravate the condition due to the rupture of parasitized erythrocytes, during which uninfected normal erythrocytes can also be destroyed, stimulating cytoadherence and consequently potentially blocking blood flow and leading to metabolic acidosis, renal failure, liver damage, and severe anemia [5,11,12,19].
Cellular damage produced by oxidative stress can be prevented or reduced by the action of consumable antioxidants present in food. Vitamins, including vitamins E and C, phenolic substances, such as flavonoids and resveratrol, carotenoids, including β-carotene and lycopene (LYC), and drugs such as N-acetylcysteine (NAC), among others, belong to this category [5,6].
Recent interest in carotenoids has focused on the role of LYC in human health. LYC is a natural, monounsaturated, lipophilic carotenoid synthesized by vegetables and fruits, such as tomatoes, watermelons, and carrots [25]. Tomatoes and tomato products are the main source of LYC and are considered an important source of carotenoids in the human diet [26]. It is widely used as a supplement in functional foods, nutraceuticals, and pharmaceuticals, as well as an additive in cosmetics [27,28].
LYC has potent activities, including antioxidant [29,30], anti-inflammatory [31], anticancer [32], neuroprotective [33], cardioprotector [34] and cholesterol reducers, and its concentration in multiple tissues is extremely important for these effects [35]
Consumable carotenoids react with a wide range of radicals, such as O2•−, OH•, NO, and ONOO− by electron transfer, producing radicals with lower oxidizing capacity, or they can lead to hydrogen atom transfer generating neutral radicals [36,37]. This antioxidant action of LYC is favored by the conjugate double bond contained in its structure, which confers high antioxidant capacity, which is about 100 times greater than that of vitamin E [26].
Several studies have indicated an association between LYC use and a decrease in oxidative changes [38,39,40] According to Pan et al. [41] The strong antioxidant activity was able to prevent cyclosporine-induced intestinal injury in mice by increasing antioxidant activity and decreasing the concentration of malondialdehyde (MDA), NO, interleukin (IL)-1β, IL-6, and tumor necrosis factor-alpha (TNF-α). These data reinforce the importance of LYC in the prevention of oxidative stress in malaria [6]
Thus, this study sought to clarify the possible antioxidant mechanisms of LYC, to determine whether LYC is an appropriate antimalarial candidate capable of reducing oxidative stress in male Balb/c mice infected with P. berghei, a strain of murine malaria responsible for inducing in the experimental animal model a syndrome similar to that caused by P. falciparum in humans and that is well characterized regarding the involvement of oxidative mechanisms in its pathophysiology. In this sense, this study provides evidence that, in mice with P. berghei-induced malaria, treatment with the carotenoid lycopene was able to improve several biochemical biomarkers related to oxidative stress. The levels of uric acid (UA), thiobarbituric acid reactive substances (TBARS), and NO in infected mice treated with LYC decreased below the levels found in the untreated infected animals, or those treated with NAC, reaching values similar to those of the non-infected animals. To the best of our knowledge, this study is the first to demonstrate important findings on the benefits of a therapy based on the natural compound lycopene for the management of biochemical changes in experimental malaria in vivo.
2. Results
Our data showed that P. berghei infection induced a significant increase in the level of TBARS compared to the Sham group in brain and lung tissues on days 4, 8, and 12 post-infection (p < 0.0001; Figure 2). Both the LYC+Pb and NAC+Pb groups have significantly lower TBARS levels than the Pb group in both tissues (p < 0.0001). In addition, the LYC treatment provided an apparent normalization in the level of TBARS in the treated animals, which was similar to the values presented by the animals of the Sham group in both brain and lung tissues.
In Figure 3 we observed that P. berghei infection led to a significant increase in AC-ABTS and AC-DPPH in animals of the Pb group compared to the Sham group in brain and lung tissues (p < 0.0001). In the LYC supplemental group, a significant decrease in the level of AC-ABTS and AC-DPPH was observed in relation to the Pb group in both tissues (p < 0.0001), presenting behavior close to that exhibited by the Sham group in both tissues. However, after treatment with NAC, a significant increase in the level of AC-ABTS was observed in the brain tissue compared to the Sham and LYC groups (p < 0.0001).
Figure 4 shows that P. berghei induced a significant increase in NO levels in the Pb group compared to the Sham group in both tissues throughout the study period (p < 0.0001). After treatment with LYC, a significant reduction in the level of NO in brain tissue was demonstrated compared to the Pb and NAC+Pb groups (p < 0.0001). In addition, LYC led to normalization of the NO level in brain tissue. On the other hand, in the lung tissue, LYC induced a significant increase in the level of NO compared to the other groups (p < 0.0001).
Figure 5 shows that P. berghei induced a significant increase in UA levels in the Pb group compared to the Sham group in both tissues (p < 0.0001). On the other hand, LYC supplementation induced a significant reduction (p < 0.0001) of UA levels in relation to the Pb group in both tissues, and to the NAC+Pb group in the brain tissue. Treatment with lycopene provided a normalization of UA levels in both tissues, as well as NAC in lung tissue up to the eighth day of treatment.
3. Discussion
Malaria causes more than 500,000 deaths every year, most of them due to brain and/or lung complications induced by P. falciparum infection. In order to understand the pathophysiology involved in malaria, experimental infection in an mice model with P. berghei infection has been widely used for many studies due to its ability to induce oxidative biochemical changes, including lipid peroxidation and/or decreased antioxidant capacity in vital organs such as lung and brain, with several similarities to the human disease [42,43].
In order to investigate whether substances with antioxidant potential could reduce oxidative biochemical changes during malarial infection, Balb/c mice were induced to malaria by P. berghei inoculation, and treated with LYC or NAC.
Because NAC is a drug widely indicated for the treatment and/or prevention of several respiratory diseases and is involved in the reduction of oxidative stress in diseases including human immunodeficiency virus (HIV) infection, influenza A/H1N1 virus, and malaria, it was used in our study as a standard substance [44,45,46,47]. On the other hand, LYC, as an orally administered antioxidant agent, was tested for its antioxidant activity in mice infected with P. berghei and its action was compared to the effects of NAC. LYC is a carotenoid present in foods such as tomatoes, watermelon, guava, among other foods, which an antioxidant potential approximately twice the activity of β-carotene [48]. Its antioxidant property contributes to optimizing health status and reducing the risk of oxidative stress-based diseases such as cancer and malaria [38,49].
In this study, our data evidenced that LYC and NAC improved several biochemical biomarkers related to oxidative stress. However, LYC presented a more intense effect than NAC. LYC induced a reduction in the levels of UA, TBARS, and NO in mice with malaria, leading to concentrations significantly lower than those found in untreated infected animals, or in those treated with NAC, reaching values similar to those of uninfected animals. In this study, we confirmed important data on the in vivo benefits of a therapy based on this natural compound for the management of oxidative biochemical changes in experimental malaria.
Although there is still no consensus on the precise mechanism responsible for malaria severity, studies indicate that both the host and the parasite are under oxidative stress, due to the increase in circulating RONS, produced during the inflammatory response by cells such as activated monocytes and neutrophils, and mainly due to the degradation of hemoglobin by the parasite [6,12,50,51,52].
During infection, the parasite is able to develop rapidly, because by consuming hemoglobin from the host erythrocyte, degrades it into amino acids, which are used for its nutrition. Fe2+, also released in the cytosol, undergoes Fenton and Haber-Weiss reactions favored by the presence of O2, generating RONS including O2•−, H2O2 and OH•. These RONS can then induce lipid peroxidation, impair microvessel endothelial cells, and cause other important tissue damages [16,17]
In our study, we demonstrated that the experimental malaria induced by P. berghei infection caused a detrimental effect on the oxidative biochemical parameters studied in mice, resulting in an increase in the levels of TBARS, AC-DPPH, AC-ABTS, NO, and UA, which was expected, because, according to BAPTISTA et al. [53] about 60% of mice infected with P. berghei succumb to malaria between days 6 and 8 post-infection with moderate parasitemia between 6 and 11%.
To verify possible oxidative biochemical changes caused by RONS, we measured TBARS in lung and brain tissues of infected BALB/c mice. TBARS are mainly reactive α and β unsaturated aldehydes, such as MDA, 4-hydroxy-2-nonenal, 2-propenal (acrolein) and isoprostanes, and are products of the decomposition of polyunsaturated fatty acid hydroperoxides and laboratory markers of lipid peroxidation, which occurs by the action of RONS on lipids and, therefore, demonstrates the occurrence of oxidative stress [54].
Our data showed that the Pb group had a significantly higher level of TBARS than the Sham group in brain and lung tissues (p < 0.0001; Figure 2), reinforcing the action of RONS as an important mediator of oxidative biochemical changes caused by experimental malaria infection.
Previously, Reis et al. [46] reported increased production of MDA and conjugated dienes in the brains of C57BL/6 mice infected with Pb, indicating the occurrence of oxidative stress. Fernandes et al. [55] correlated increased plasma levels of reactive aldehydes in Pb-infected mice with malaria severity. Scaccabarozzi et al. [56] showed that infection with Pb in C57BL/6J mice induced at the same time biochemical changes in liver and lung tissue, as well as leading to acute respiratory distress syndrome, due to an exacerbated excessive oxidative response, demonstrated by the alteration of mobilizable antioxidant enzymes and the increase in MDA levels. Recently, Chuljerm et al. [57] reported elevated levels of TBARS in the plasma and livers of Pb-infected mice.
Our findings indicate that in P. berghei infected mice an excessive oxidative response seems to predominate, demonstrated by the increase in TBARS levels, which is an index of loss of structure and integrity of the cell membrane of brain and lung tissues, and these changes were related to high parasitemia.
However, treatments with LYC and NAC significantly reversed (p < 0.0001) the increase in TBARS concentration when compared to the Pb group (Figure 2). In addition, LYC treatment brought the TBARS level to similar values exhibited by the animals in the Sham group, which suggests that LYC may be successful in eliminating RONS, inhibiting lipid peroxidation, and protecting membrane lipids from oxidative damage in brain and lung of mice during malarial infection.
These data are in line with previous findings in the literature in which it has been reported that both LYC and NAC are able to reduce endogenous oxidant levels and protect cells against a wide range of pro-oxidative insults [58,59] by reacting directly with RONS [60]. According to Zhang [61], LYC has the ability to inhibit oxidative stress and TBARS, limiting the production of RONS. LYC activity has also been implicated in the elimination of P. falciparum in vitro [49]. However, the direct effect of LYC on the proliferation of the parasite in vivo is unclear. On the other hand, the antioxidant activity of NAC may have involved the direct action of its free thiol (SH) group, which can act as an electron donor, favoring the interaction of NAC with RONS.
By promoting cellular aggression processes, parasites can readily alter the concentration of mobilizable antioxidants in the body, triggering endogenous defense mechanisms against oxidative aggression. The action of these molecules helps protect biological membranes, which are susceptible to lipid peroxidation and oxidative damage in general. Mobilizable antioxidants include enzymes such as SOD, which dismutates O2•− into H2O2, CAT, and GSH-Px, which break down H2O2 and ROOH into poorly reactive molecules, such as H2O, alcohol, and O2, as well as other non-enzymatic molecules such as reduced glutathione (GSH) [5,6]
In this sense, we found a significant increase in AC-ABTS and AC-DPPH in animals in the Pb group compared to the Sham group in brain and lung tissues (p < 0.0001; Figure 3). The significant increase in AC-ABTS and AC-DPPH in the present study may reflect an adaptive response of the animal organism to an increased demand for mobilizable antioxidants, possibly to combat the RONS generated during P. berghei infection.
Reinforcing these results, we found a significant positive correlation between the concentrations of TBARS versus AC-ABTS (r = 0.4; p < 0.0001) and TBARS versus AC-DPPH (r = 0.5; p < 0.0001) in the Pb group. The increase in antioxidant capacity may have occurred due to the action of Pb on mobilizable enzymes or due to the high level of RONS, such as H2O2, O2•−, OH•, ONOO−, which form within cells in response to Pb infection.
On the other hand, LYC supplementation led to a significant decrease in the level of AC-ABTS and AC-DPPH in relation to the Pb group in both tissues (Figure 3). Additionally, the level of AC-ABTS and AC-DPPH was restored by treatment in the LYC+Pb group in brain and lung tissues. Similarly, to what occurred for infected and untreated animals, after treatment with NAC, a significant increase in the level of AC-ABTS in brain tissue was observed compared to the Sham and LYC+Pb groups. In the lung tissue, NAC restored the level of AC-ABTS to the levels presented by the animals of the Sham group.
In view of these findings, it can be suggested that, in mice infected with Pb, the administration of LYC may decrease the activity of mobilizable antioxidants, as a result of cellular redox regulation after the ingestion of exogenous antioxidants. On the other hand, NAC, being an analogue and precursor of GSH, may have induced the replacement of intracellular levels of GSH, also known as the main antioxidant produced by the body and which protects cells from oxidative stress.
In this context, some studies report that, in diseases in which oxidative stress is a pathogenic mediator, such as Alzheimer's, Parkinson's, Chagas, dengue, and malaria, the actions of mobilizable antioxidants are not sufficient to maintain an adequate cellular redox balance [62,63,64,65].
To counteract the deleterious effects of RONS, supplementation with consumable antioxidants, including vitamins E and C, phenolic substances such as flavonoids and resveratrol, carotenoids including β-carotene and LYC, and drugs such as NAC, are essential for maintaining optimal cellular function. LYC is a potent antioxidant that also has anti-inflammatory, anti-atherogenic, antidiabetic, neuroprotective, and anticancer effects [31,32,34,39,66,67,68].
Antioxidants can act by increasing the concentration of other antioxidants in the body, donating electrons to the RONS and neutralizing them, binding directly to RONS and inactivating them, or also by preventing the cascade of formation of highly reactive RONS, such as OH• and ONOO− [69].
These RONS are reported to be a key factor in triggering the devastating inflammatory response that has been associated with disease progression and subsequent fatal outcome [70]. RONS also appear to act as second messengers in a signaling cascade and can activate mononuclear cells, as well as macrophages and dendritic cells that stimulate the release of High Mobility Group Box-1 (HMGB-1) into the intra- and extracellular space [18].
According to Techarang et al. [71] increased expression of HMGB-1 in endothelial cells may stimulate several receptors, including RAGE, TLR-4, and TLR-2, which activate nuclear factor kappa B (NF-κB) leading to the production of TNF-α, interferon-gamma (IFN-γ), IL-1β, and IL-6, involved in the pathogenesis of malaria.
These inflammatory mediators can also stimulate the activation of iNOS in macrophages. In macrophages, iNOS activity is normally low, but the expression of this enzyme is greatly stimulated by the release of inflammatory mediators such as IF N-γ, in response to infection, leading to an increased NO concentration [72].
NO is recognized as a mediator in a wide range of biological systems, plays an important role in the maintenance and regulation of bronchomotor tone by non-adrenergic and non-cholinergic neural mechanisms (NANC), and acts as an important vasodilator in the vascular endothelium [73]. In addition, it is essential for the phagocytic function of macrophages, generating oxidizing molecules such as ONOO− with the ability to destroy invading pathogens.
Despite participating in the body's defense when produced by macrophages, in our study the increase in NO concentration was not correlated with a decrease in parasitemia. According to our results, there was a significant increase (p < 0.0001) in NO levels in the Pb group compared to the Sham group in both tissues throughout the study period (Figure 4). Therefore, it is possible that the exaggerated production of NO may have generated harmful effects to the animal organism, including generalized vasodilation resulting in hypotension and oxidative changes resulting from ONOO−, highly reactive, capable of inducing peroxidation of membrane lipids.
However, after treatment with LYC, a significant reduction in the level of NO in brain tissue was observed in comparison with Pb and NAC+Pb groups (Figure 4). In addition, LYC led to normalization of the NO level in brain tissue. In lung tissue, LYC led to an increase in NO levels in the LYC+Pb group compared to the other groups.
LYC is an extremely hydrophobic molecule with a high capacity to permeate the double layer of phospholipids of the plasma membrane, and can accumulate in different tissues, such as the brain [74]. According to studies, LYC can be absorbed by passive diffusion and active process in which the transporter of the scavenger receptor protein class B type 1 (SR-B1) is involved [38,75]. Additionally, it has a long chain with 11 double conjugate links, which makes it an excellent scavenger for RONS, such as 1O2, O2•−, OH•, NO2 and ONOO− [76,77].
In view of our results, it is possible that lycopene crosses the blood-brain barrier of the central nervous system and exerts neuroprotective effects against oxidative changes induced by malarial infection. Additionally, lycopene serves as an efficient antioxidant, acting on antioxidant defense in brain tissue by binding to RONS, inactivating them and preventing the reaction chains that lead to the formation of 1O2, O2•−, OH•, NO2, and ONOO− highly deleterious to the body. In lung tissue, we believe that LYC can decompose O2•− and thus prevent the production of ONOO− and tissue oxidative nitration, releasing NO to promote vascular vasodilation, contributing to tissue perfusion and the arrival of more defense cells and lycopene molecules in the tissue. Therefore, lycopene may accumulate and exert protective effects on the pulmonary and cerebral vasculature.
Previous studies have shown the protective role of NO in the hemodynamic improvement of cerebral microcirculation and the reduction of vascular pathology in cerebral malaria [78,79]. On the other hand, in malarial infection, ONOO− can induce protein nitration, depletion of consumable antioxidants, peroxidation of the microvascular endothelium [22], resulting in the marked decrease in blood flow leading to ischemia and subsequent hypoxia, vasospasms, and tissue hypoperfusion along with cell-mediated congestion, resulting in pulmonary edema and stroke [80,81]. In addition, tissue ischemia may result from cytoadhesion caused by Plasmodium infection [5,12].
Prolonged hypoxia ceases oxidative phosphorylation in the mitochondria, promoting ATP degradation and accumulation of xanthine oxidase (XO) and hypoxanthine. In an attempt to restore tissue oxygenation, the body stimulates the expression of eNOS to increase the production of NO, aiming at tissue reperfusion. However, when the blood supply is resumed, XO acts on hypoxanthine resulting in the production of O2•−, H2O2, and uric acid, the latter being used in the present study as a marker of the occurrence of IRS. On such IRS, O2•− can still react with NO generating ONOO− [82].
In this context, our data indicated that Pb infection was able to induce a significant increase in UA levels in the Pb group compared to the Sham group in both tissues (p < 0.0001; Figure 5). On the other hand, LYC supplementation induced a significant reduction (p < 0.0001) of UA levels in relation to the Pb group in both tissues, and to the NAC+Pb group in the brain tissue. In addition, treatment with LYC provided a normalization in the levels of UA in both tissues, as well as NAC in the lung tissue, suggesting its role in the prevention of damage caused by IRS.
These results are in agreement with previous studies that showed elevated UA levels in children infected with P. falciparum during acute episodes and with disease severity, suggesting that UA is an important mediator in the pathophysiology of malaria [83].
4. Materials and Methods
A total of 231 adult male mice of the species Mus musculus and Balb/c breed, 7-10 weeks of age, weighing between 25 and 40g (Vivarium of the Evandro Chagas Institute, Ananindeua, Pará-Brazil) were used. The animals were housed in the Experimentation Vivarium of the Oxidative Stress Research Laboratory (LAPEO) of the Institute of Biological Sciences (ICB) of the Federal University of Pará (UFPA), with an ambient temperature of 24±2°C, a 12-hour light/dark cycle (lights from 7 a.m. to 7 p.m.), and free access to food and water. Before any experimental procedure, animals were acclimatized to laboratory conditions for 15 days.
All experimental procedures were performed at LAPEO. The animals were handled and cared for in accordance with the ethical standards of animal experimentation indicated by the Brazilian Society of Laboratory Animal Science. The project, filed under No. 3235130919, was approved by the Ethics Committee on the Use of Animals of UFPA (CEUA/UFPA).
2.1. Protocol for the preparation and administration of lycopene and N-acetylcysteine
The lycopene preparation and administration protocol was chosen based on a dose-response study on the effects of lycopene supplementation on biomarkers of oxidative stress [84], and the dose was calculated by allometric extrapolation [85]. Animals received 3.11mg/kg of body weight/day of lycopene via gavage.
The N-acetylcysteine preparation and administration protocol was chosen based on a randomized, double-blind, placebo-controlled study of chronic obstructive pulmonary disease [86], and the dose was calculated by allometric extrapolation [85]. Animals received 62mg/kg of body weight/day of NAC via gavage.
Mice in the LYC+Pb and NAC+Pb groups were pretreated with a dose of 3.11mg/kg bw/day of LYC or 62mg/kg bw/day of NAC via gavage, respectively, twenty-four hours before infection. Treatments then continued daily until the day before the animals were euthanized. Mice of the Sham group received only vehicle (water) via gavage.
2.2. Plasmodium berghei ANKA -infection protocol
The mice of the groups PB, LYC+Pb and NAC+Pb were performed by intraperitoneal injection (i.p.) of 106 P. Berghei ANKA-parasitized red blood cells (pRBC). On the other hand, the Sham group animals received 106 of non-parazitated red blood cells.
2.3. Protocol for subdivision of the experimental groups
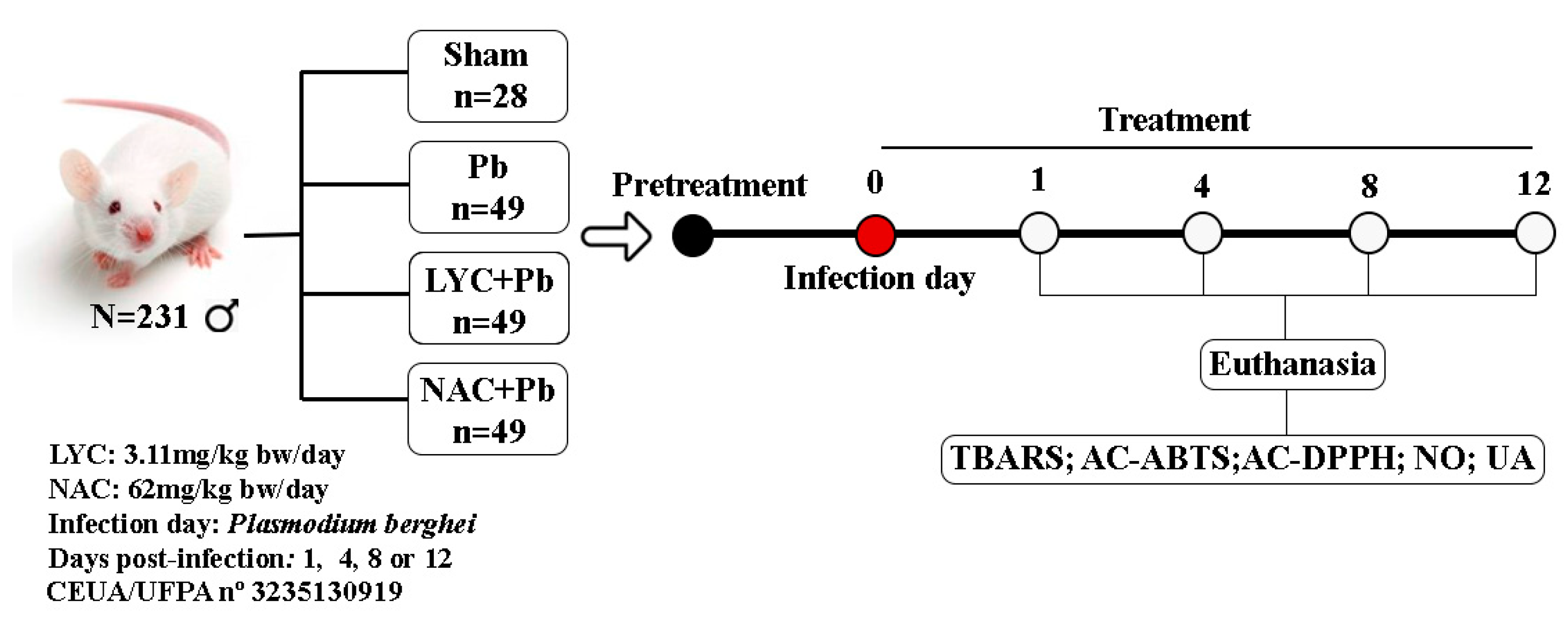
In an experiment of 1, 4, 8 or 12 days of consecutive follow-up, 231 male mice (Balb/c) were randomly assigned to 4 groups (Figure 6), including Sham (n=28): mice received the vehicle (water; gavage) and non-parasitized red blood cells (i.p.); Pb (n=49): mice infected with P. berghei ANKA (i.p.); LYC+Pb (n=49): mice treated with LYC (gavage) and infected with P. berghei (i.p.); NAC+Pb (n=49): mice treated with NAC (gavage) and infected with P. berghei (i.p.).
The subgroups 1- and 4-days were formed by 7 animals each. The 8- and 12-days subgroups were formed by 15 and 20 animals, respectively, due to the higher mortality expected for these subgroups.
2.4. Euthanasia Protocol and Sample Preparation
At the end of each study period, each animal was anesthetized intraperitoneally, using a combination of 0.5 mL of 10% ketamine hydrochloride (9 mg/kg) + 0.25 mL of 2% xylazine hydrochloride (10 mg/kg) + 4.25 mL of water for injection. After confirmation of unconsciousness and loss of corneal reflex, animals were euthanized by exsanguination through intracardiac puncture.
Subsequently, both lungs were extracted, as well as the brain of each animal. The organs were weighed and added phosphate saline-buffer solution (PBS) in a ratio of 1:10 (m:v). Subsequently, the ultrasonic disruption of tissues was performed to obtain a homogenate. After homogenization, the material was centrifuged at 2,500 rpm for 10 minutes and the supernatant was collected, stored in an Eppendorf microtube, and frozen at -20ºC until assayed.
2.5. Biochemical Measurements Protocol
2.5.1. Thiobarbituric acid reactive substances (TBARS)
The method was carried out according to the fundamentals proposed by Kohn and Liversedge [87], with chemical conditions of the reaction adjusted according to Percário et al. [88]. This method evaluates lipid peroxidation and has been used as an indicator of oxidative stress. The test is based on the reaction of thiobarbituric acid (4,6-Dihydroxypyrimidine-2-thiol, TBA; Sigma-Aldrich; T5500; São Paulo/SP) with by-products of lipid peroxidation (e.g. MDA), at acidic pH (2.5) and high temperature (94°C), forming chromogens with absorbance at 535 nm.
Initially, 0.5 mL of the sample or standard was mixed with 1 mL of the TBA solution (10 mM). Then, this solution was placed in a water bath at 94ºC for 60 minutes. Subsequently, 4 mL of n-butyl alcohol were added, the solution was stirred in a vortex-type agitator, then centrifuged at 3000 rpm for 10 minutes. After that, 3 mL of the supernatant was transferred to the cuvette and then spectrophotometry was performed at 535 nm (Spectrophotometer 800XI; Femto; São Paulo/SP).
A standard curve (1,1,3,3, tetrahydroxypropane; standard MDA; 20 μM; Sigma-Aldrich Chemical; 108383; São Paulo/SP) was performed in triplicate and, from the values found, the equation of the line (y = 0.1419x − 0.0037) was calculated, where y represents the absorbance value and x the concentration value, obtaining R2 = 0.9999. From the equation of the line, the concentration of TBARS of the samples was determined.
2.5.2. Antioxidant capacity by radical ABTS•+ inhibition (AC-ABTS)
It was carried out according to the initial foundations proposed by Miller et al. [89], with reaction conditions modified by Re et al. [90]. The method is based on the ability of substances to eliminate the radical cation 2,2'-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS•+), a blue-green chromophore with maximum absorption at 734 nm, resulting in the formation of the stable product ABTS, which is colorless.
Initially, the ABTS•+ solution (2.45 mM) was prepared from the reaction between ABTS (7 mM; Sigma-Aldrich; A1888; São Paulo/SP) and potassium persulfate (140 mM; K2O8S2; Sigma-Aldrich; 216224; São Paulo/SP). Then, the initial reading (T0) of the ABTS•+ solution was performed in an 800XI spectrophotometer (Femto; São Paulo/SP) at 734 nm. Then, 30 μL of sample or standard was added to the solution and, after 5 minutes, the final reading (T5) was performed.
A standard curve (6-hydroxy-2,5,7,8-tetramethylcromono-2-carboxylic acid; Trolox®; 2.5mM; Sigma-Aldrich; 23881-3; São Paulo/SP) was performed in triplicate and, from the absorbance values found, the equation of the line (y = 0.4324x + 0.0049) was obtained, where y represents the absorbance value and x the concentration value, obtaining R2 = 0.9997. From the equation of the line, the AC-ABTS of the samples was determined.
2.5.3. Antioxidant capacity by radical DPPH• inhibition (AC-DPPH)
The test was performed according to the adapted method proposed by Blois [91] This assay evaluates the total antioxidant capacity of synthetic or natural substances to eliminate the DPPH• radical (Sigma-Aldrich; D9132; São Paulo/SP), a violet chromophore with absorption at 517nm, resulting in the formation of the hydrogenated product DPPH, which is yellow or colorless.
First, the DPPH• solution (0.1 mM) was prepared from the reaction between DPPH (394.32 g/mol; Sigma-Aldrich; A1888; São Paulo/SP) and ethyl alcohol (P.A.; C2H6O; Sigma-Aldrich; 216224; São Paulo/SP). Subsequently, the DPPH• solution was read in an 800XI spectrophotometer (Femto; São Paulo/SP) at 517 nm. Then, 50 μL of the sample or standard was mixed in 950 μL of the DPPH• solution and placed in a water bath at 30°C for 30 minutes. After this period, the second reading was held.
A standard curve (Trolox; 2.5mM) was performed in triplicate and, from the absorbance values found, the equation of the line was obtained: y = 0.2041x − 0.0031, where y represents the absorbance value and x, the concentration value, obtaining R2 = 0.9973. From the equation of the line, the AC-DPPH was determined in the samples.
2.5.4. Nitric oxide (NO)
The NO concentration was determined indirectly by the detection of nitrate (NO3−) or nitrite (NO2−) in the samples, using the NO colorimetric assay kit (Elabscience®, Catalog No: E-BC-K035-M). NO is readily oxidized to form nitrite in vivo or in aqueous solution, which can react with the chromogenic reagent nitrate, forming a pale red compound. The concentration of the compound is linearly related to the concentration of NO in the sample.
Initially, 100 μL of sample or standard was mixed with 200 μL of reagent 1 (sulfate solution), and 100 μL of reagent 2 (alkaline reagent). After resting for 15 minutes, the solution was centrifuged at 3000 rpm for 10 minutes. Then, 160 μL of the supernatant was transferred to the microplate, where 80 μL of the chromogenic reagent was added. After 15 minutes of incubation at room temperature, spectrophotometric readings were performed at 550 nm.
A standard curve (sodium nitrite; 100 μM) was performed in triplicate and, from the values found, the equation of the line was obtained (y = 0.0022x − 0.0005), where y represents the absorbance value and x, the concentration value, obtaining R2 = 0.9987. From the equation of the line, the concentration of nitric oxide in the samples was determined.
2.5.5. Uric acid (UA)
The procedure was performed using the Liquiform Uric Acid Kit (Labtest). The technique consists of the oxidation of uric acid by uricase producing allantoin and H2O2. H2O2 in the presence of peroxidase reacts with 3,5-dichloro-2-hydroxybenzene sulfonate acid (DHBS) and 4-aminoantipyrine to form the antipyrylquinonymine chromogen. The intensity of the red color formed is directly proportional to the concentration of uric acid in the sample.
To perform the assay, 0.02 mL of the sample or standard was mixed in 1 mL of uric acid working reagent (4-aminoantipyrine, peroxidase, sodium azide, DHBS, and uricase). The solution was then incubated in a water bath at 37°C for 5 minutes. Then, absorbances were determined using an 800XI spectrophotometer (Femto; São Paulo/SP) at 505 nm.
A standard curve (Uric acid; 20mg/dL) was performed in triplicate and, from the values found, the equation of the line was obtained (y = 0.0166x + 0.0012), where y represents the absorbance value and x, the concentration value, obtaining R2 = 0.9986. From the equation of the line, the concentration of UA in the samples was determined.
2.6. Statistical analysis
For each parameter analyzed, the analysis of possible outliers was performed by calculating the interquartile range, in which the difference between the third quartile (Q3) and the first quartile (Q1) was determined, called dj. Any value lower than Q1-3/2dj or greater than Q3+3/2dj was considered to be discrepancies, and was not considered in the statistical calculations. After the analysis of the discrepant points, normality was assessed using the Levene test. For homoscedastic distribution, the Analysis of Variance (ANOVA) test was applied, and for heteroscedastic dispersion, the Kruskal-Wallis test was applied. Significant differences were compared between the groups using Tukey's post-hoc test.
In the intragroup temporal progression analysis, the unpaired Student's t-test was performed. To verify the possible correlation between parameters, Pearson's correlation test was performed, considering the paired values of two parameters obtained for the same animal, and the calculations were performed with the data obtained from all animals simultaneously, according to the group to which they belong. For the pairs of values in which there was suspicion of a linear relationship, regression analysis was performed, using all animals in both groups simultaneously and each group individually. In all tests, a significance level of 5% (p≤0.05) was considered.
5. Conclusions
Lycopene prevented oxidative damage induced by Plasmodium berghei in brains and lungs of mice, restoring the levels of NO, TBARS, and antioxidant molecules, as well as preventing the occurrence of ischemia and reperfusion syndrome in infected animals. These results are striking evidence of the beneficial effect of lycopene supplementation on oxidative stress in experimental malaria in vivo, and emphasize the importance of antioxidant supplementation in the treatment of the disease.
Thus, lycopene may become an important viable, safe, and innovative strategy for the development of therapeutic alternatives to mitigate the damage caused by malarial infection.
Author Contributions
Conceptualization, E.L.P.V. and S.P.; methodology, E.L.P.V., A.R.Q.G., A.S.B.S., M.S.G., E.P.C.; formal analysis, E.L.P.V.; investigation, E.L.P.V.; data curation, E.L.P.V. and S.P.; writing—original draft preparation, E.L.P.V., A.R.Q.G., A.S.B.S., M.S.G., E.P.C.; O.O.F., M.S.O.; writing—review and editing, E.L.P.V., E.H.A.A., S.P.; visualization, S.P.; supervision, S.P.; project administration, S.P.; funding acquisition, S.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES; Brazil) and Fundação Amazônia de Amparo a Estudos e Pesquisas (FAPESPA; Brazil).
Institutional Review Board Statement
The animal study protocol was approved by Comissão de Ética no Uso de Animais da UFPA (CEUA/UFPA; protocol code no. 3235130919, issued on September 10th, 2021).
Data Availability Statement
Data is freely available upon request.
Acknowledgments
The authors are grateful to the Central Vivarium of the Animal Breeding and Production Section (SCPA) of the Evandro Chagas Institute, which kindly provided us with the animals to carry out the project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Suzen, S.; Gurer-Orhan, H.; Saso, L. Detection of Reactive Oxygen and Nitrogen Species by Electron Paramagnetic Resonance (EPR) Technique. Molecules 2017, 22, 181. [Google Scholar] [CrossRef]
- Hardy, M.; Zielonka, J.; Karoui, H.; Sikora, A.; Michalski, R.; Podsiadły, R.; Lopez, M.; Vasquez-Vivar, J.; Kalyanaraman, B.; Ouari, O. Detection and Characterization of Reactive Oxygen and Nitrogen Species in Biological Systems by Monitoring Species-Specific Products. Antioxid. Redox Signal. 2018, 28, 1416–1432. [Google Scholar] [CrossRef]
- Matuz-Mares, D.; Riveros-Rosas, H.; Vilchis-Landeros, M.M.; Vázquez-Meza, H. Glutathione Participation in the Prevention of Cardiovascular Diseases. Antioxidants 2021, 10, 1220. [Google Scholar] [CrossRef]
- Nyui, M.; Nakanishi, I.; Anzai, K.; Ozawa, T.; Matsumoto, K. Reactivity of Redox Sensitive Paramagnetic Nitroxyl Contrast Agents with Reactive Oxygen Species. J. Clin. Biochem. Nutr. 2019, 64, 13–19. [Google Scholar] [CrossRef]
- Gomes, A.R.Q.; Cunha, N.; Varela, E.L.P.; Brígido, H.P.C.; Vale, V.V.; Dolabela, M.F.; de Carvalho, E.P.; Percário, S. Oxidative Stress in Malaria: Potential Benefits of Antioxidant Therapy. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Varela, E.L.P.; Gomes, A.R.Q.; da Silva Barbosa dos Santos, A.; de Carvalho, E.P.; Vale, V.V.; Percário, S. Potential Benefits of Lycopene Consumption: Rationale for Using It as an Adjuvant Treatment for Malaria Patients and in Several Diseases. Nutrients 2022, 14, 5303. [Google Scholar] [CrossRef]
- Nie, L.; Nusantara, A.C.; Damle, V.G.; Baranov, M. V.; Chipaux, M.; Reyes-San-Martin, C.; Hamoh, T.; Epperla, C.P.; Guricova, M.; Cigler, P.; et al. Quantum Sensing of Free Radicals in Primary Human Dendritic Cells. Nano Lett. 2022, 22, 1818–1825. [Google Scholar] [CrossRef]
- Sies, H. Oxidative Stress: Concept and Some Practical Aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef]
- Herbas, M.S.; Ueta, Y.Y.; Ichikawa, C.; Chiba, M.; Ishibashi, K.; Shichiri, M.; Fukumoto, S.; Yokoyama, N.; Takeya, M.; Xuan, X.; et al. Alpha-Tocopherol Transfer Protein Disruption Confers Resistance to Malarial Infection in Mice. Malar. J. 2010, 9, 101. [Google Scholar] [CrossRef]
- Wu, R.L.; Idris, A.H.; Berkowitz, N.M.; Happe, M.; Gaudinski, M.R.; Buettner, C.; Strom, L.; Awan, S.F.; Holman, L.A.; Mendoza, F.; et al. Low-Dose Subcutaneous or Intravenous Monoclonal Antibody to Prevent Malaria. N. Engl. J. Med. 2022, 387, 397–407. [Google Scholar] [CrossRef]
- Becker, K.; Tilley, L.; Vennerstrom, J.L.; Roberts, D.; Rogerson, S.; Ginsburg, H. Oxidative Stress in Malaria Parasite-Infected Erythrocytes: Host–Parasite Interactions. Int. J. Parasitol. 2004, 34, 163–189. [Google Scholar] [CrossRef]
- Percário, S.; Moreira, D.R.; Gomes, B.A.Q.; Ferreira, M.E.S.; Gonçalves, A.C.M.; Laurindo, P.S.O.C.; Vilhena, T.C.; Dolabela, M.F.; Green, M.D. Oxidative Stress in Malaria. Int. J. Mol. Sci. 2012, 13, 16346. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.P.; Bhatt, R.; Singh, V. Genetic Profiling of the Plasmodium Falciparum Parasite Population in Uncomplicated Malaria from India. Malar. J. 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Suzuki, H.; Kume, A.; Herbas, M. Potential of Vitamin E Deficiency, Induced by Inhibition of α-Tocopherol Efflux, in Murine Malaria Infection. Int. J. Mol. Sci. 2018, 20, 64. [Google Scholar] [CrossRef]
- Schmidt, H.M.; Kelley, E.E.; Straub, A.C. The Impact of Xanthine Oxidase (XO) on Hemolytic Diseases. Redox Biol. 2019, 21, 101072. [Google Scholar] [CrossRef]
- Jortzik, E.; Becker, K. Thioredoxin and Glutathione Systems in Plasmodium Falciparum. Int. J. Med. Microbiol. 2012, 302, 187–194. [Google Scholar] [CrossRef]
- Gupta, M.; Kumar, S.; Kumar, R.; Kumar, A.; Verma, R.; Darokar, M.P.; Rout, P.; Pal, A. Inhibition of Heme Detoxification Pathway in Malaria Parasite by 3-Hydroxy-11-Keto-β-Boswellic Acid Isolated from Boswellia Serrata. Biomed. Pharmacother. 2021, 144, 112302. [Google Scholar] [CrossRef]
- Ty, M.C.; Zuniga, M.; Götz, A.; Kayal, S.; Sahu, P.K.; Mohanty, A.; Mohanty, S.; Wassmer, S.C.; Rodriguez, A. Malaria Inflammation by Xanthine Oxidase-produced Reactive Oxygen Species. EMBO Mol. Med. 2019, 11, e9903. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, K.P.; Bali, P.; Anwar, S.; Kaul, A.; Singh, O.P.; Gupta, B.K.; Kumari, N.; Noor Alam, M.; Raziuddin, M.; et al. INOS Polymorphism Modulates INOS/NO Expression via Impaired Antioxidant and ROS Content in P. Vivax and P. Falciparum Infection. Redox Biol. 2018, 15, 192–206. [Google Scholar] [CrossRef]
- Moreira, A.S.; Estato, V.; Malvar, D.C.; Sanches, G.S.; Gomes, F.; Tibirica, E.; Daniel-Ribeiro, C.T.; Carvalho, L.J.M. L-Arginine Supplementation and Thromboxane Synthase Inhibition Increases Cerebral Blood Flow in Experimental Cerebral Malaria. Sci. Reports 2019 91 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Dhangadamajhi, G.; Mohapatra, B.N.; Kar, S.K.; Ranjit, M. Genetic Variation in Neuronal Nitric Oxide Synthase (NNOS) Gene and Susceptibility to Cerebral Malaria in Indian Adults. Infect. Genet. Evol. 2009, 9, 908–911. [Google Scholar] [CrossRef] [PubMed]
- Kondrikov, D.; Elms, S.; Fulton, D.; Su, Y. ENOS-β-Actin Interaction Contributes to Increased Peroxynitrite Formation during Hyperoxia in Pulmonary Artery Endothelial Cells and Mouse Lungs. J. Biol. Chem. 2010, 285, 35479–35487. [Google Scholar] [CrossRef]
- Knackstedt, S.L.; Georgiadou, A.; Apel, F.; Abu-Abed, U.; Moxon, C.A.; Cunnington, A.J.; Raupach, B.; Cunningham, D.; Langhorne, J.; Krüger, R.; et al. Neutrophil Extracellular Traps Drive Inflammatory Pathogenesis in Malaria. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef] [PubMed]
- Wedgwood, S.; Lakshminrusimha, S.; Farrow, K.N.; Czech, L.; Gugino, S.F.; Soares, F.; Russell, J.A.; Steinhorn, R.H. Apocynin Improves Oxygenation and Increases ENOS in Persistent Pulmonary Hypertension of the Newborn. Am. J. Physiol. Cell. Mol. Physiol. 2012, 302, L616–L626. [Google Scholar] [CrossRef]
- Kang, D.-M.; Kwon, J.-M.; Jeong, W.-J.; Jung, Y.J.; Kang, K.K.; Ahn, M.-J. Antioxidant Constituents and Activities of the Pulp with Skin of Korean Tomato Cultivars. Molecules 2022, 27, 8741. [Google Scholar] [CrossRef]
- Liu, D.; Shi, J.; Colina Ibarra, A.; Kakuda, Y.; Jun Xue, S. The Scavenging Capacity and Synergistic Effects of Lycopene, Vitamin E, Vitamin C, and β-Carotene Mixtures on the DPPH Free Radical. LWT - Food Sci. Technol. 2008, 41, 1344–1349. [Google Scholar] [CrossRef]
- Carvalho, G.C.; Marena, G.D.; Leonardi, G.R.; Sábio, R.M.; Corrêa, I.; Chorilli, M.; Bauab, T.M. Lycopene, Mesoporous Silica Nanoparticles and Their Association: A Possible Alternative against Vulvovaginal Candidiasis? Molecules 2022, 27, 8558. [Google Scholar] [CrossRef]
- Mannino, F.; D’Angelo, T.; Pallio, G.; Ieni, A.; Pirrotta, I.; Giorgi, D.A.; Scarfone, A.; Mazziotti, S.; Booz, C.; Bitto, A.; et al. The Nutraceutical Genistein-Lycopene Combination Improves Bone Damage Induced by Glucocorticoids by Stimulating the Osteoblast Formation Process. Nutrients 2022, 14, 4296. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, H.; Wang, J.; Liu, P.; Tan, X.; Ren, B.; Liu, Z.; Liu, X. Lycopene Supplementation Attenuates Oxidative Stress, Neuroinflammation, and Cognitive Impairment in Aged CD-1 Mice. J. Agric. Food Chem. 2018, 66, 3127–3136. [Google Scholar] [CrossRef]
- Liu, S.; Yang, D.; Yu, L.; Aluo, Z.; Zhang, Z.; Qi, Y.; Li, Y.; Song, Z.; Xu, G.; Zhou, L. Effects of Lycopene on Skeletal Muscle-Fiber Type and High-Fat Diet-Induced Oxidative Stress. J. Nutr. Biochem. 2021, 87, 108523. [Google Scholar] [CrossRef]
- Ni, Y.; Zhuge, F.; Nagashimada, M.; Nagata, N.; Xu, L.; Yamamoto, S.; Fuke, N.; Ushida, Y.; Suganuma, H.; Kaneko, S.; et al. Lycopene Prevents the Progression of Lipotoxicity-Induced Nonalcoholic Steatohepatitis by Decreasing Oxidative Stress in Mice. Free Radic. Biol. Med. 2020, 152, 571–582. [Google Scholar] [CrossRef]
- Cheng, J.; Miller, B.; Balbuena, E.; Eroglu, A. Lycopene Protects against Smoking-Induced Lung Cancer by Inducing Base Excision Repair. Antioxidants 2020, 9, 643. [Google Scholar] [CrossRef]
- LEI, X.; LEI, L.; ZHANG, Z.; CHENG, Y. Neuroprotective Effects of Lycopene Pretreatment on Transient Global Cerebral Ischemia-Reperfusion in Rats: The Role of the Nrf2/HO-1 Signaling Pathway. Mol. Med. Rep. 2016, 13, 412–418. [Google Scholar] [CrossRef]
- Ferreira-Santos, P.; Aparicio, R.; Carrón, R.; Sevilla, M.Á.; Monroy-Ruiz, J.; Montero, M.J. Lycopene-Supplemented Diet Ameliorates Cardiovascular Remodeling and Oxidative Stress in Rats with Hypertension Induced by Angiotensin II. J. Funct. Foods 2018, 47, 279–287. [Google Scholar] [CrossRef]
- Liu, H.; Liu, J.; Liu, Z.; Wang, Q.; Liu, J.; Feng, D.; Zou, J. Lycopene Reduces Cholesterol Absorption and Prevents Atherosclerosis in ApoE –/– Mice by Downregulating HNF-1α and NPC1L1 Expression. J. Agric. Food Chem. 2021, 69, 10114–10120. [Google Scholar] [CrossRef]
- Mortensen, A.; Skibsted, L.H.; Truscott, T.G. The Interaction of Dietary Carotenoids with Radical Species. Arch. Biochem. Biophys. 2001, 385, 13–19. [Google Scholar] [CrossRef]
- Srinivasan, M.; Sudheer, A.R.; Pillai, K.R.; Kumar, P.R.; Sudhakaran, P.R.; Menon, V.P. Lycopene as a Natural Protector against γ-Radiation Induced DNA Damage, Lipid Peroxidation and Antioxidant Status in Primary Culture of Isolated Rat Hepatocytes in Vitro. Biochim. Biophys. Acta - Gen. Subj. 2007, 1770, 659–665. [Google Scholar] [CrossRef]
- Kujawska, M.; Ewertowska, M.; Adamska, T.; Sadowski, C.; Ignatowicz, E.; Jodynis-Liebert, J. Antioxidant Effect of Lycopene-Enriched Tomato Paste on N-Nitrosodiethylamine-Induced Oxidative Stress in Rats. J. Physiol. Biochem. 2014, 70, 981–990. [Google Scholar] [CrossRef]
- Renju, G.L.; Kurup, G.M.; Saritha Kumari, C.H. Effect of Lycopene from Chlorella Marina on High Cholesterol-Induced Oxidative Damage and Inflammation in Rats. Inflammopharmacology 2014, 22, 45–54. [Google Scholar] [CrossRef]
- da Silva Brito, A.K.; de Morais Lima, G.; de Farias, L.M.; Rodrigues, L.A.R.L.; de Carvalho, V.B.L.; de Carvalho Pereira, C.F.; de Macedo Gonçalves Frota, K.; Conde-Júnior, A.M.; Moura, A.M.O.; dos Santos Rizzo, M.; et al. Lycopene-Rich Extract from Red Guava (Psidium Guajava L.) Decreases Plasma Triglycerides and Improves Oxidative Stress Biomarkers on Experimentally-Induced Dyslipidemia in Hamsters. Nutrients 2019, 11, 393. [Google Scholar] [CrossRef]
- Pan, X.; Niu, X.; Li, Y.; Yao, Y.; Han, L. Preventive Mechanism of Lycopene on Intestinal Toxicity Caused by Cyclophosphamide Chemotherapy in Mice by Regulating TLR4-MyD88/TRIF-TRAF6 Signaling Pathway and Gut-Liver Axis. Nutrients 2022, 14, 4467. [Google Scholar] [CrossRef]
- Torre, S.; Langlais, D.; Gros, P. Genetic Analysis of Cerebral Malaria in the Mouse Model Infected with Plasmodium Berghei. Mamm. Genome 2018, 29, 488–506. [Google Scholar] [CrossRef]
- Vandermosten, L.; Pham, T.-T.; Possemiers, H.; Knoops, S.; Van Herck, E.; Deckers, J.; Franke-Fayard, B.; Lamb, T.J.; Janse, C.J.; Opdenakker, G.; et al. Experimental Malaria-Associated Acute Respiratory Distress Syndrome Is Dependent on the Parasite-Host Combination and Coincides with Normocyte Invasion. Malar. J. 2018, 17, 102. [Google Scholar] [CrossRef]
- Aitio, M. N-acetylcysteine – Passe-partout or Much Ado about Nothing? Br. J. Clin. Pharmacol. 2006, 61, 5–15. [Google Scholar] [CrossRef]
- Atkuri, K.R.; Mantovani, J.J.; Herzenberg, L.A.; Herzenberg, L.A. N-Acetylcysteine—a Safe Antidote for Cysteine/Glutathione Deficiency. Curr. Opin. Pharmacol. 2007, 7, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Reis, P.A.; Comim, C.M.; Hermani, F.; Silva, B.; Barichello, T.; Portella, A.C.; Gomes, F.C.A.; Sab, I.M.; Frutuoso, V.S.; Oliveira, M.F.; et al. Cognitive Dysfunction Is Sustained after Rescue Therapy in Experimental Cerebral Malaria, and Is Reduced by Additive Antioxidant Therapy. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef]
- Quadros Gomes, B.A.; da Silva, L.F.D.; Quadros Gomes, A.R.; Moreira, D.R.; Dolabela, M.F.; Santos, R.S.; Green, M.D.; Carvalho, E.P.; Percário, S. N-Acetyl Cysteine and Mushroom Agaricus Sylvaticus Supplementation Decreased Parasitaemia and Pulmonary Oxidative Stress in a Mice Model of Malaria. Malar. J. 2015, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Böhm, V.; Puspitasari-Nienaber, N.L.; Ferruzzi, M.G.; Schwartz, S.J. Trolox Equivalent Antioxidant Capacity of Different Geometrical Isomers of α-Carotene, β-Carotene, Lycopene, and Zeaxanthin. J. Agric. Food Chem. 2002, 50, 221–226. [Google Scholar] [CrossRef]
- Agarwal, S.; Sharma, V.; Kaul, T.; Abdin, M.Z.; Singh, S. Cytotoxic Effect of Carotenoid Phytonutrient Lycopene on P. Falciparum Infected Erythrocytes. Mol. Biochem. Parasitol. 2014, 197, 15–20. [Google Scholar] [CrossRef]
- Hempel, C.; Combes, V.; Hunt, N.H.; Kurtzhals, J.A.L.; Grau, G.E.R. CNS Hypoxia Is More Pronounced in Murine Cerebral than Noncerebral Malaria and Is Reversed by Erythropoietin. Am. J. Pathol. 2011, 179, 1939–1950. [Google Scholar] [CrossRef]
- Sharma, L.; Kaur, J.; Rishi, P.; Shukla, G. Plasmodium Berghei: Influence of Infection on the Oxidant and Antioxidants Levels in Pregnant BALB/c Mice. Exp. Parasitol. 2012, 131, 215–222. [Google Scholar] [CrossRef]
- Zhao, S.; Duan, H.; Yang, Y.; Yan, X.; Fan, K. Fenozyme Protects the Integrity of the Blood–Brain Barrier against Experimental Cerebral Malaria. Nano Lett. 2019, 19, 8887–8895. [Google Scholar] [CrossRef]
- Baptista, F.G.; Pamplona, A.; Pena, A.C.; Mota, M.M.; Pied, S.; Vigário, A.M. Accumulation of Plasmodium Berghei -Infected Red Blood Cells in the Brain Is Crucial for the Development of Cerebral Malaria in Mice. Infect. Immun. 2010, 78, 4033–4039. [Google Scholar] [CrossRef]
- Kurutas, E.B. The Importance of Antioxidants Which Play the Role in Cellular Response against Oxidative/Nitrosative Stress: Current State. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef]
- Fernandes, E.S.; Brito, C.X.L.; Teixeira, S.A.; Barboza, R.; Dos Reis, A.S.; Azevedo-Santos, A.P.S.; Muscará, M.; Costa, S.K.P.; Marinho, C.R.F.; Brain, S.D.; et al. TRPV1 Antagonism by Capsazepine Modulates Innate Immune Response in Mice Infected with Plasmodium Berghei ANKA. Mediators Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- Scaccabarozzi, D.; Deroost, K.; Corbett, Y.; Lays, N.; Corsetto, P.; Salè, F.O.; Van den Steen, P.E.; Taramelli, D. Differential Induction of Malaria Liver Pathology in Mice Infected with Plasmodium Chabaudi AS or Plasmodium Berghei NK65. Malar. J. 2018, 17, 18. [Google Scholar] [CrossRef]
- Chuljerm, H.; Maneekesorn, S.; Somsak, V.; Ma, Y.; Srichairatanakool, S.; Koonyosying, P. Anti-Malarial and Anti-Lipid Peroxidation Activities of Deferiprone-Resveratrol Hybrid in Plasmodium Berghei-Infected Mice. Biology (Basel). 2021, 10, 911. [Google Scholar] [CrossRef]
- Elsayed, A.; Elkomy, A.; Elkammar, R.; Youssef, G.; Abdelhiee, E.Y.; Abdo, W.; Fadl, S.E.; Soliman, A.; Aboubakr, M. Synergistic Protective Effects of Lycopene and N-Acetylcysteine against Cisplatin-Induced Hepatorenal Toxicity in Rats. Sci. Rep. 2021, 11, 13979. [Google Scholar] [CrossRef]
- Ezeriņa, D.; Takano, Y.; Hanaoka, K.; Urano, Y.; Dick, T.P. N-Acetyl Cysteine Functions as a Fast-Acting Antioxidant by Triggering Intracellular H2S and Sulfane Sulfur Production. Cell Chem. Biol. 2018, 25, 447–459. [Google Scholar] [CrossRef]
- Samuni, Y.; Goldstein, S.; Dean, O.M.; Berk, M. The Chemistry and Biological Activities of N-Acetylcysteine. Biochim. Biophys. Acta - Gen. Subj. 2013, 1830, 4117–4129. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Xu, F.; Huang, W.; Ji, Q.; Han, Y.; Shao, B.; Li, Y. Protective Effects of Lycopene against AFB1-Induced Erythrocyte Dysfunction and Oxidative Stress in Mice. Res. Vet. Sci. 2020, 129, 103–108. [Google Scholar] [CrossRef]
- Coles, L.D.; Tuite, P.J.; Öz, G.; Mishra, U.R.; Kartha, R. V.; Sullivan, K.M.; Cloyd, J.C.; Terpstra, M. Repeated-Dose Oral N-Acetylcysteine in Parkinson’s Disease: Pharmacokinetics and Effect on Brain Glutathione and Oxidative Stress. J. Clin. Pharmacol. 2018, 58, 158–167. [Google Scholar] [CrossRef]
- Aqeel, S.; Naheda, A.; Raza, A.; Khan, K.; Khan, W. Differential Status and Significance of Non-Enzymatic Antioxidants (Reactive Oxygen Species Scavengers) in Malaria and Dengue Patients. Acta Trop. 2019, 195, 127–134. [Google Scholar] [CrossRef]
- Sánchez-Villamil, J.P.; Bautista-Niño, P.K.; Serrano, N.C.; Rincon, M.Y.; Garg, N.J. Potential Role of Antioxidants as Adjunctive Therapy in Chagas Disease. Oxid. Med. Cell. Longev. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Ton, A.M.M.; Campagnaro, B.P.; Alves, G.A.; Aires, R.; Côco, L.Z.; Arpini, C.M.; Guerra e Oliveira, T.; Campos-Toimil, M.; Meyrelles, S.S.; Pereira, T.M.C.; et al. Oxidative Stress and Dementia in Alzheimer’s Patients: Effects of Synbiotic Supplementation. Oxid. Med. Cell. Longev. 2020, 2020, 1–14. [Google Scholar] [CrossRef]
- Velmurugan, B.; Bhuvaneswari, V.; Nagini, S. Antiperoxidative Effects of Lycopene during N-Methyl-N′-Nitro-N-Nitrosoguanidine-Induced Gastric Carcinogenesis. Fitoterapia 2002, 73, 604–611. [Google Scholar] [CrossRef]
- Hsiao, G.; Fong, T.H.; Tzu, N.H.; Lin, K.H.; Chou, D.S.; Sheu, J.R. A Potent Antioxidant, Lycopene, Affords Neuroprotection against Microglia Activation and Focal Cerebral Ischemia in Rats. In Vivo 2004, 18, 351–356. [Google Scholar]
- Neyestani, T.R.; Shariatzadeh, N.; Gharavi, A.; Kalayi, A.; Khalaji, N. Physiological Dose of Lycopene Suppressed Oxidative Stress and Enhanced Serum Levels of Immunoglobulin M in Patients with Type 2 Diabetes Mellitus: A Possible Role in the Prevention of Long-Term Complications. J. Endocrinol. Invest. 2007, 30, 833–838. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free Radicals, Antioxidants and Functional Foods: Impact on Human Health. Pharmacogn. Rev. 2010, 4, 118. [Google Scholar] [CrossRef]
- Dieye, Y.; Mbengue, B.; Dagamajalu, S.; Fall, M.M.; Loke, M.F.; Nguer, C.M.; Thiam, A.; Vadivelu, J.; Dieye, A. Cytokine Response during Non-Cerebral and Cerebral Malaria: Evidence of a Failure to Control Inflammation as a Cause of Death in African Adults. PeerJ 2016, 4, e1965. [Google Scholar] [CrossRef]
- Techarang, T.; Jariyapong, P.; Viriyavejakul, P.; Punsawad, C. High Mobility Group Box-1 (HMGB-1) and Its Receptors in the Pathogenesis of Malaria-Associated Acute Lung Injury/Acute Respiratory Distress Syndrome in a Mouse Model. Heliyon 2021, 7, e08589. [Google Scholar] [CrossRef] [PubMed]
- Gramaglia, I.; Sobolewski, P.; Meays, D.; Contreras, R.; Nolan, J.P.; Frangos, J.A.; Intaglietta, M.; van der Heyde, H.C. Low Nitric Oxide Bioavailability Contributes to the Genesis of Experimental Cerebral Malaria. Nat. Med. 2006, 12, 1417–1422. [Google Scholar] [CrossRef]
- Filogonio, R.; Sartori, M.R.; Morgensen, S.; Tavares, D.; Campos, R.; Abe, A.S.; Taylor, E.W.; Rodrigues, G.J.; De Nucci, G.; Simonsen, U.; et al. Cholinergic Regulation along the Pulmonary Arterial Tree of the South American Rattlesnake: Vascular Reactivity, Muscarinic Receptors, and Vagal Innervation. Am. J. Physiol. Integr. Comp. Physiol. 2020, 319, R156–R170. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Mao, X.; Zhang, J.; Sun, P.; Wang, H.; Zhang, Y.; Ma, Y.; Xu, S.; Lv, R.; Liu, X. Oral Delivery of Lycopene-Loaded Microemulsion for Brain-Targeting: Preparation, Characterization, Pharmacokinetic Evaluation and Tissue Distribution. Drug Deliv. 2019, 26, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Ojha, S.; Goyal, S.; Sharma, C.; Arora, S.; Kumari, S.; Arya, D. Cardioprotective Effect of Lycopene against Isoproterenol-Induced Myocardial Infarction in Rats. Hum. Exp. Toxicol. 2013, 32, 492–503. [Google Scholar] [CrossRef]
- Yonar, M.E.; Sakin, F. Ameliorative Effect of Lycopene on Antioxidant Status in Cyprinus Carpio during Pyrethroid Deltamethrin Exposure. Pestic. Biochem. Physiol. 2011, 99, 226–231. [Google Scholar] [CrossRef]
- Dobrzyńska, M.M.; Gajowik, A.; Radzikowska, J. The Effect of Lycopene Supplementation on Radiation-Induced Micronuclei in Mice Reticulocytes in Vivo. Radiat. Environ. Biophys. 2019, 58, 425–432. [Google Scholar] [CrossRef]
- Cabrales, P.; Zanini, G.M.; Meays, D.; Frangos, J.A.; Carvalho, L.J.M. Nitric Oxide Protection Against Murine Cerebral Malaria Is Associated With Improved Cerebral Microcirculatory Physiology. J. Infect. Dis. 2011, 203, 1454–1463. [Google Scholar] [CrossRef]
- Ong, P.K.; Melchior, B.; Martins, Y.C.; Hofer, A.; Orjuela-Sánchez, P.; Cabrales, P.; Zanini, G.M.; Frangos, J.A.; Carvalho, L.J.M. Nitric Oxide Synthase Dysfunction Contributes to Impaired Cerebroarteriolar Reactivity in Experimental Cerebral Malaria. PLoS Pathog. 2013, 9, e1003444. [Google Scholar] [CrossRef]
- Kondrikov, D.; Gross, C.; Black, S.M.; Su, Y. Novel Peptide for Attenuation of Hyperoxia-Induced Disruption of Lung Endothelial Barrier and Pulmonary Edema via Modulating Peroxynitrite Formation. J. Biol. Chem. 2014, 289, 33355–33363. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.F.; Granger, D.N. Blood Cells and Endothelial Barrier Function. Tissue Barriers 2015, 3, e978720. [Google Scholar] [CrossRef]
- Cantu-Medellin, N.; Kelley, E.E. Xanthine Oxidoreductase-Catalyzed Reduction of Nitrite to Nitric Oxide: Insights Regarding Where, When and How. Nitric Oxide 2013, 34, 19–26. [Google Scholar] [CrossRef]
- Das, D.; Guha, S.; Talukdar, A.; Sau, T.; Sanghai, R.; Pant, N.; Jena, A. A Study of Uric Acid Level as a Marker of Severity in Malaria. Int. J. Med. Sci. Diagnosis Res. 2022, 6. [Google Scholar] [CrossRef]
- Devaraj, S.; Mathur, S.; Basu, A.; Aung, H.H.; Vasu, V.T.; Meyers, S.; Jialal, I. A Dose-Response Study on the Effects of Purified Lycopene Supplementation on Biomarkers of Oxidative Stress. J. Am. Coll. Nutr. 2008, 27, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharm. 2016, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.-P.; Wen, F.-Q.; Bai, C.-X.; Wan, H.-Y.; Kang, J.; Chen, P.; Yao, W.-Z.; Ma, L.-J.; Li, X.; Raiteri, L.; et al. Twice Daily N-Acetylcysteine 600 Mg for Exacerbations of Chronic Obstructive Pulmonary Disease (PANTHEON): A Randomised, Double-Blind Placebo-Controlled Trial. Lancet Respir. Med. 2014, 2, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Kohn, H.I.; Liversedge, M. On a New Aerobic Metabolite Whose Production by Brain Is Inhibited by Apomorphine, Emetine, Ergotamine, Epinephrine, and Menadione. J. Pharmacol. Exp. Ther. 1944, 82, 292–300. [Google Scholar]
- Percário, S.; Vital, A.; Jablonka, F. Dosagem Do Malondialdeido. Newslab 1994, 2, 46–50. [Google Scholar]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J.; Gopinathan, V.; Milner, A. A Novel Method for Measuring Antioxidant Capacity and Its Application to Monitoring the Antioxidant Status in Premature Neonates. Clin. Sci. 1993, 84, 407–412. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant Determinations by the Use of a Stable Free Radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
Figure 1.
Timeline of the number of cases (above) and number of deaths (below) of malaria in the world.
Figure 1.
Timeline of the number of cases (above) and number of deaths (below) of malaria in the world.
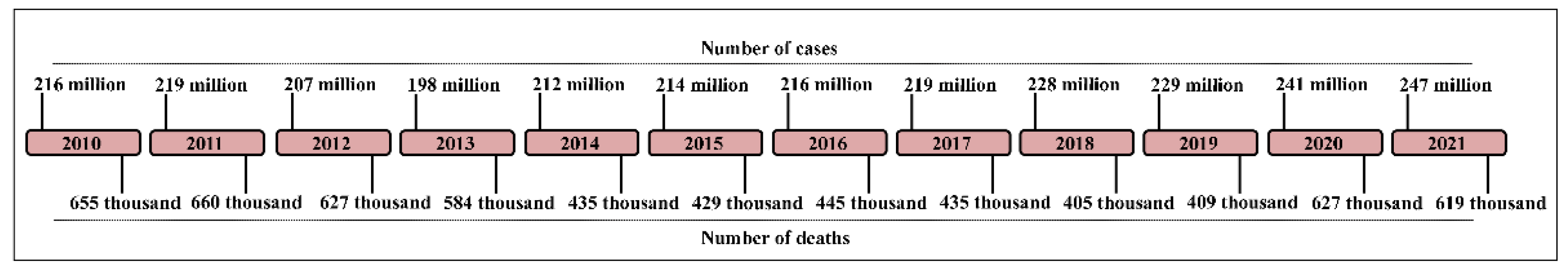
Figure 2.
Concentration of thiobarbituric acid reactive substances (TBARS) in the brain and lungs of mice infected with P. berghei, treated or not with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp ≤ 0.02 versus Sham; op ≤ 0.005 versus Pb. 4 days: *p ≤ 0.001 versus Sham; #p < 0.0001 versus Pb; &p < 0.0001 versus LYC+Pb. 8 days: ap ≤ 0.0002 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p < 0.0001 versus Sham; γp < 0.0001 versus Pb; €p <0.0001 versus LYC+Pb.
Figure 2.
Concentration of thiobarbituric acid reactive substances (TBARS) in the brain and lungs of mice infected with P. berghei, treated or not with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp ≤ 0.02 versus Sham; op ≤ 0.005 versus Pb. 4 days: *p ≤ 0.001 versus Sham; #p < 0.0001 versus Pb; &p < 0.0001 versus LYC+Pb. 8 days: ap ≤ 0.0002 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p < 0.0001 versus Sham; γp < 0.0001 versus Pb; €p <0.0001 versus LYC+Pb.
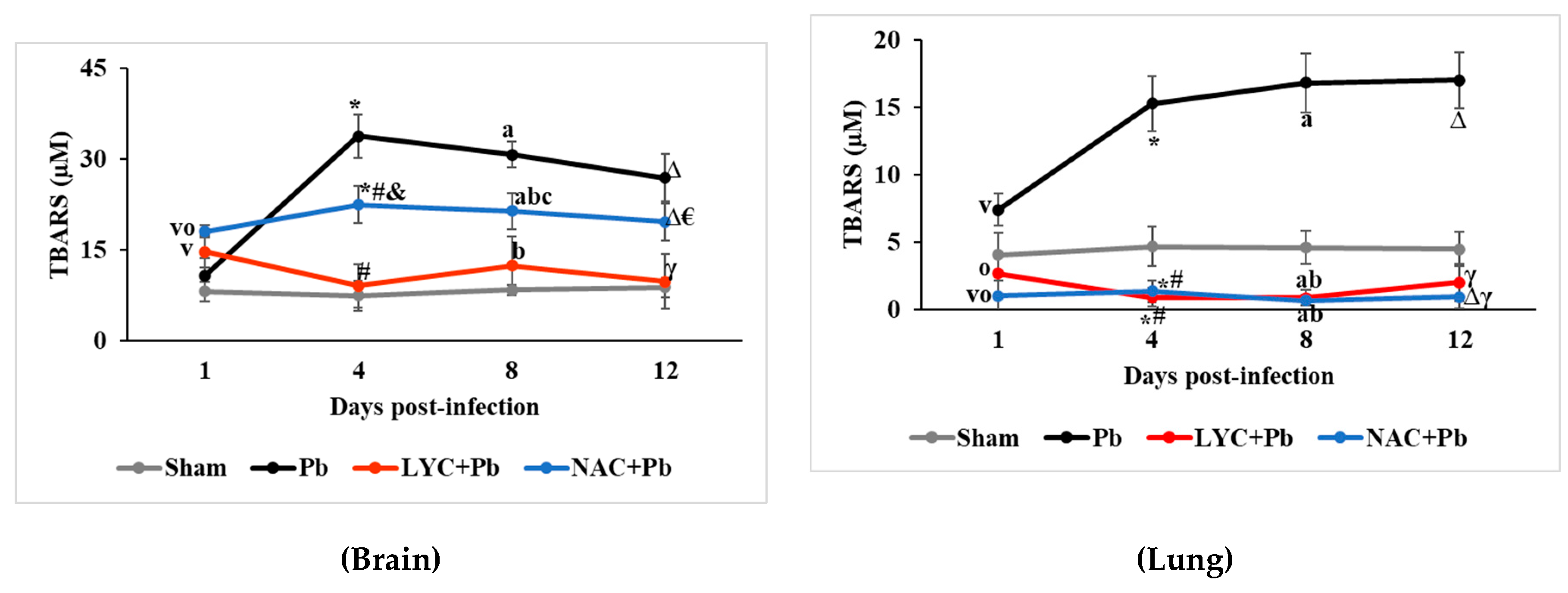
Figure 3.
Antioxidant capacity (AC) by inhibition of ABTS and DPPH radicals in the brain and lungs of mice infected with P. berghei, treated or not with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp < 0.0001 versus Sham; op ≤ 0.005 versus Pb; rp < 0.0001 versus LYC+Pb. 4 days: *p ≤ 0.0001 versus Sham; #p ≤ 0.01 versus Pb; &p ≤ 0.02 versus LYC+Pb. 8 days: ap < 0.0001 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p ≤ 0.005 versus Sham; γp ≤ 0.001 versus Pb; €p <0.0001 versus LYC+Pb.
Figure 3.
Antioxidant capacity (AC) by inhibition of ABTS and DPPH radicals in the brain and lungs of mice infected with P. berghei, treated or not with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp < 0.0001 versus Sham; op ≤ 0.005 versus Pb; rp < 0.0001 versus LYC+Pb. 4 days: *p ≤ 0.0001 versus Sham; #p ≤ 0.01 versus Pb; &p ≤ 0.02 versus LYC+Pb. 8 days: ap < 0.0001 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p ≤ 0.005 versus Sham; γp ≤ 0.001 versus Pb; €p <0.0001 versus LYC+Pb.
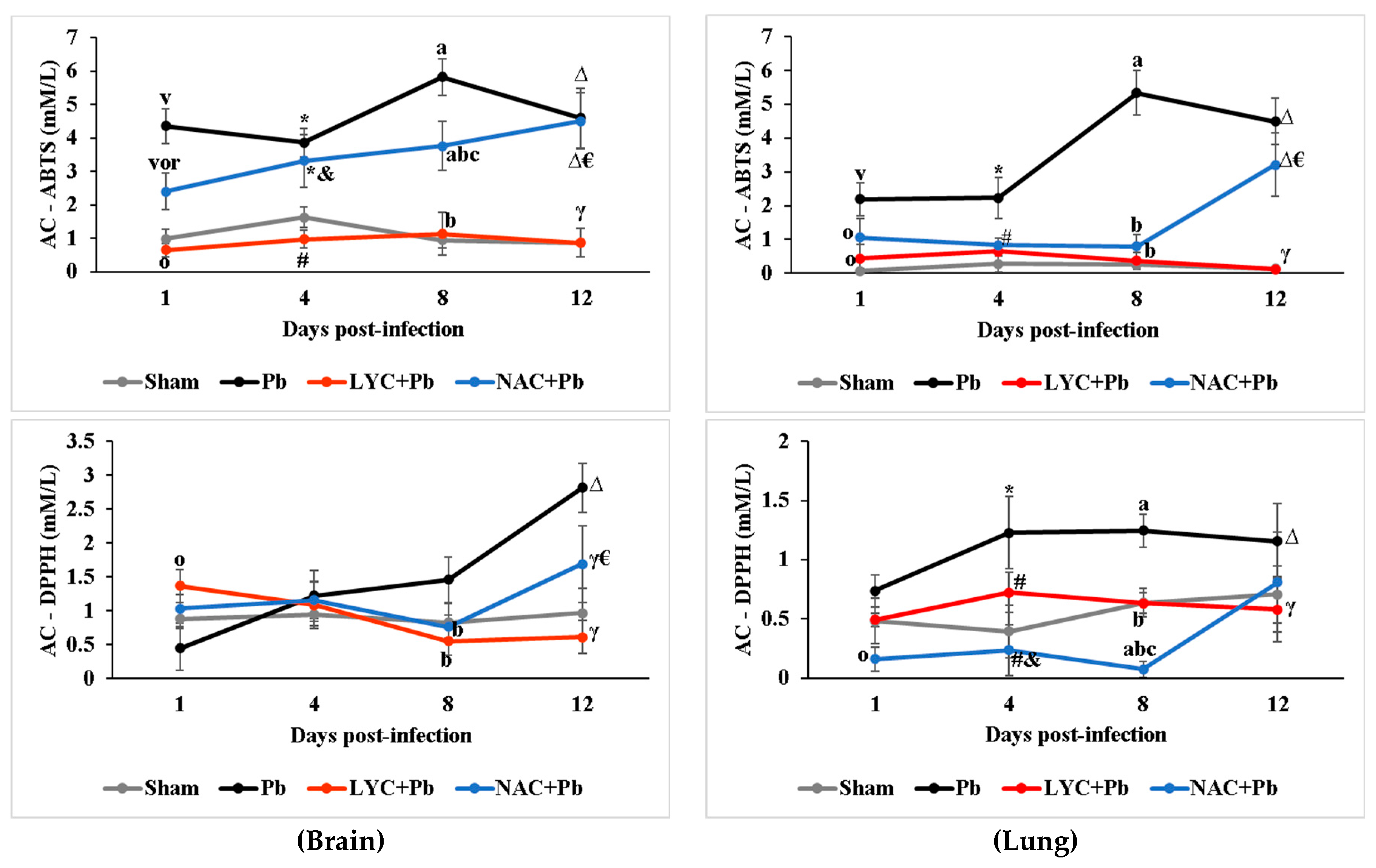
Figure 4.
Nitric oxide concentration in the brain and lungs of mice infected with P. berghei, whether or not treated with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp < 0.0001 versus Sham; op< 0.0001 versus Pb; rp < 0.0001 versus LYC+Pb. 4 days: *p < 0.0001 versus Sham; #p < 0.0001 versus Pb; &p < 0.0001 versus LYC+Pb. 8 days: ap < 0.0001 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p < 0.0001 versus Sham; γp < 0.0001 versus Pb; €p <0.0001 versus LYC+Pb.
Figure 4.
Nitric oxide concentration in the brain and lungs of mice infected with P. berghei, whether or not treated with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp < 0.0001 versus Sham; op< 0.0001 versus Pb; rp < 0.0001 versus LYC+Pb. 4 days: *p < 0.0001 versus Sham; #p < 0.0001 versus Pb; &p < 0.0001 versus LYC+Pb. 8 days: ap < 0.0001 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p < 0.0001 versus Sham; γp < 0.0001 versus Pb; €p <0.0001 versus LYC+Pb.
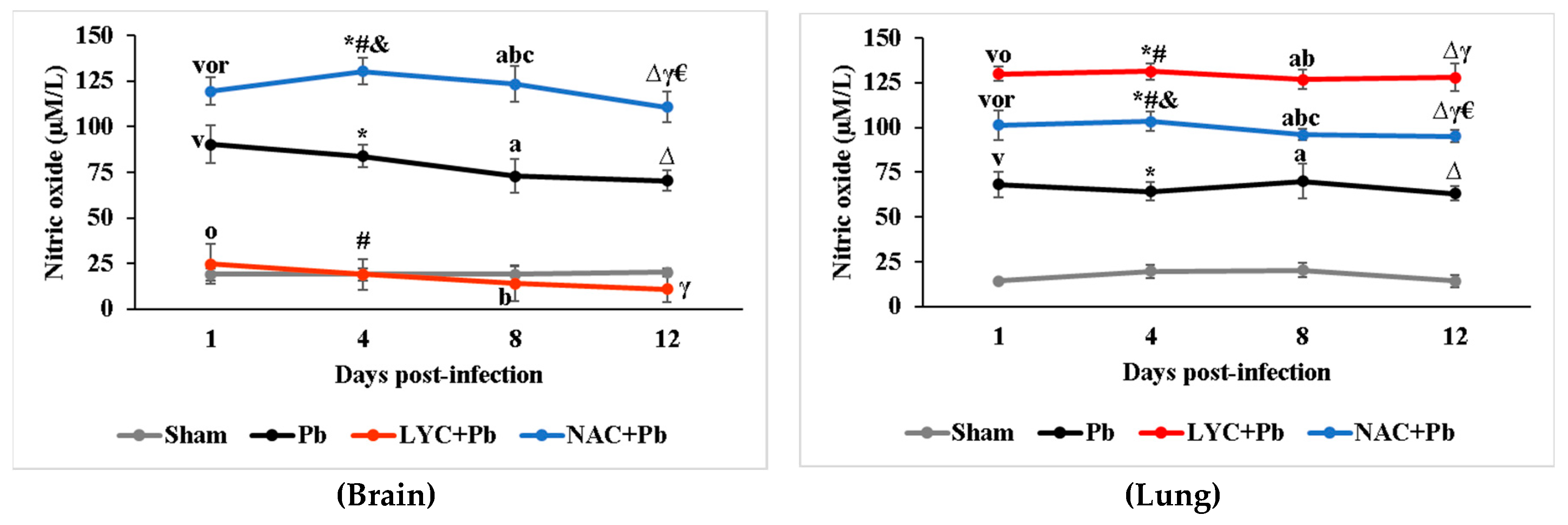
Figure 5.
Uric acid concentration in brain and lungs of animals infected with P. berghei treated or not with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp ≤ 0,0001 versus Sham; Op ≤ 0.005 versus Pb; rp < 0.0001 versus LYC+Pb. 4 days: *p ≤ 0.0007 versus Sham; #p ≤ 0.02 versus Pb; &p < 0.0001 versus LYC+Pb. 8 days: ap < 0.0001 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p < 0.0001 versus Sham; γp < 0.0001 versus Pb; €p < 0.0001 versus LYC+Pb.
Figure 5.
Uric acid concentration in brain and lungs of animals infected with P. berghei treated or not with lycopene or N-acetylcysteine. Data are expressed as means ± standard deviation. 1 day: Vp ≤ 0,0001 versus Sham; Op ≤ 0.005 versus Pb; rp < 0.0001 versus LYC+Pb. 4 days: *p ≤ 0.0007 versus Sham; #p ≤ 0.02 versus Pb; &p < 0.0001 versus LYC+Pb. 8 days: ap < 0.0001 versus Sham; bp < 0.0001 versus Pb; cp < 0.0001 versus LYC+Pb. 12 days: ∆p < 0.0001 versus Sham; γp < 0.0001 versus Pb; €p < 0.0001 versus LYC+Pb.
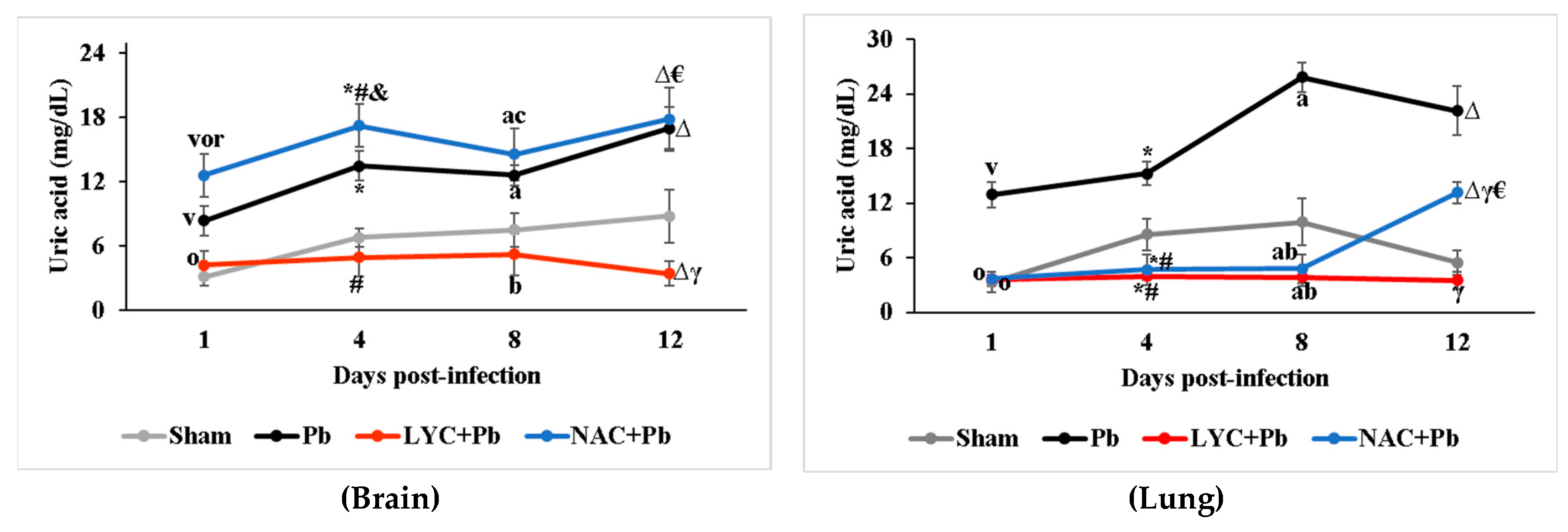
Figure 6.
Schematic representation of the experimental protocol. Balb/c mice were pretreated with lycopene or N-acetylcysteine prior to inoculation with 106 erythrocytes parasitized with Plasmodium Berghei ANKA, and treatment continued daily until the day before euthanasia, for 1, 4, 8 or 12 consecutive days. After euthanasia, brain and lungs were collected for biochemical analyses: Antioxidant Capacity Equivalent to Trolox by inhibition of ABTS (AC-ABTS) and DPPH (AC-DPPH) radicals. TBARS= Thiobarbituric Acid Reactive Substances; UA= Uric Acid; NO= Nitric Oxide.
Figure 6.
Schematic representation of the experimental protocol. Balb/c mice were pretreated with lycopene or N-acetylcysteine prior to inoculation with 106 erythrocytes parasitized with Plasmodium Berghei ANKA, and treatment continued daily until the day before euthanasia, for 1, 4, 8 or 12 consecutive days. After euthanasia, brain and lungs were collected for biochemical analyses: Antioxidant Capacity Equivalent to Trolox by inhibition of ABTS (AC-ABTS) and DPPH (AC-DPPH) radicals. TBARS= Thiobarbituric Acid Reactive Substances; UA= Uric Acid; NO= Nitric Oxide.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



