
Submitted:
20 October 2023
Posted:
23 October 2023
You are already at the latest version
Abstract
A Bacille Calmette-Guérin (BCG) is still the only licensed vaccine for the prevention of tuberculosis, providing limited protection against Mycobacterium tuberculosis infection in adulthood. New advances in delivery of DNA vaccines by electroporation have been made in the past decade. We evaluated the safety and immunogenicity of DNA-hsp65 vaccine administered by intramuscular electroporation (EP) in cynomolgus macaques. Animals received three doses of DNA-hsp65 at 30-days intervals. We demonstrated that intramuscular electroporated DNA-hsp65 vaccine immunization of cynomolgus macaques was safe, and there were no vaccine-related effects on hematological, renal, or hepatic profiles, compared to the pre-vaccination parameters. No tuberculin skin test conversion nor lung X-ray alteration was identified. Further, low, and transient peripheral cellular immune response and cytokine expression were observed, primarily after the third dose of the DNA-hsp65 vaccine. Electroporated DNA-hsp65 vaccination is safe but provided limited enhancement of peripheral cellular immune responses. Preclinical vaccine trials with DNA-hsp65 delivered via EP may include a combination of plasmid cytokine adjuvant and/or protein prime-boost regimen, to help the induction of a stronger cellular immune response.
Keywords:
tuberculosis
; electroporation
; vaccine
; DNA-hsp65
; nonhuman primate
1. Introduction
Pulmonary tuberculosis (TB) caused by Mycobacterium tuberculosis remains one of the leading worldwide killers among all infectious diseases, despite extensive use of the Mycobacterium bovis bacilli Calmette-Guérin (BCG) vaccine. Ten million infected people developed active disease and 1.6 million died from the disease in 2021 [1]. A new TB vaccine is urgently needed to replace or to be used as a booster for the BCG vaccine. Among the novel vaccine strategies, plasmid DNA-based TB vaccines have received increasing attention because of their safety and induction of both CD4+ and CD8+ T cell responses. One of the main products under development by our group is a DNA vaccine containing the gene that expresses the Mycobacterium leprae hsp65 antigen (DNA-hsp65) with broad immunomodulatory and/or immunoregulatory properties [2]. Our previous studies have demonstrated that DNA-hsp65 confers a high level of protection in mice both prophylactically and therapeutically [2]: it works in prevention [3]; can be used as a BCG booster [4]; acts in the immunotherapy of chronic TB, MDR-TB and LTB [5,6,7], it can be used concomitantly with antimycobacterial drugs to enhance therapeutic and immunotherapeutic action [7,8]; prevents reactivation of infection [5,6], and can act in those animals with compromised immune systems as in helminth coinfections [9,10,11].
Since the discovery that DNA-hsp65 can have immunomodulatory and/or immunoregulatory action against TB several other works have been done by our group with other infectious diseases, autoimmune diseases, allergy, and tumors. Thus, it was demonstrated that DNA-hsp65 presents broad immunotherapeutic properties for leishmaniasis [12], schistosomiasis [9,11], paracoccidioidomycosis [13,14], chromoblastomycosis [15], helminths [10]; autoimmune diseases such as diabetes [16,17,18,19], arthritis [20,21], encephalomyelitis [22,23,24], and atherosclerosis [25], allergic diseases such as asthma and atopic dermatitis [26,27,28] and tumors in experimental models of mice and in phase I/II clinical trial in humans [29,30]. Thus, DNA-hsp65 actively participates in the activation of innate immunity, acting as an endogenous adjuvant and playing a fundamental role in the activation and control of adaptive immunity [29,30,31,32].
Studies have shown that DNA-hsp65 preventive and/or therapeutic activities are associated with in vitro activation of human dendritic cells (DCs) and macrophages [29,31]; stimulation of CD4 and, CD8 [33,34,35,36,37], gamma/delta [38] and B lymphocytes [19,39]; induction of murine Th1, Th17 and Treg pattern of immune response [40,41,42]; production of cytokines and activation molecules necessary to restrict bacterial growth [7,8,29,40]; and activate processes to control fibrosis and granulomatous process [43,44] in mice. The main subpopulations of cells that confer memory and long-term immunity against TB were evaluated in experiments with mice immunized with BCG or DNA-hsp65 and subsequently infected with M. tuberculosis [37]. The main subpopulations linked to the protective role of DNA-hsp65 were CD8+CD44hiIFN-γ+ and CD4+CD44hiIFN-γ+, which were prominent even 8 or 15 months after vaccine administration. On the other hand, in response to BCG vaccination, the primary enriched subpopulation of cells was composed of CD4+CD44loIFN-γ+. In addition, BCG prime and DNAhsp65 booster’s studies improved immunogenicity and efficacy in comparison with the vaccination of BCG or DNA-hsp65 alone [37]. This analysis suggests that the prime-boost strategy is a promising alternative for inducing an effective immune response against TB [37]. Moreover, we have shown that B cells capture plasmid DNA-hsp65 and thereby modulate the formation of CD8+ memory T cells after M. tuberculosis challenge in mice [39,45]. Therefore, B cells functioned as part of the protective immune response after DNA immunization.
Studies of innate immune system activation by DNA-hp65 were performed on cultured dendritic cells (DCs) and macrophages [29,31]. In the immunogenicity studies in human and murine cell cultures the immunostimulatory potential of DNA-hsp65 was evaluated and the results showed that after 4 hours of stimulation, both macrophages (characterized as CD11b+/CD86+/HLA-DR+) and DCs (characterized as CD11c+/CD86+/CD123-/BDCA-4+/IFN-α-) were able to capture the DNA plasmid [29]. Both cell populations expressed mRNA for the hsp65 protein, in addition to exhibiting high constitutive expression of the intrinsic TLR9 receptor. After 48 hours of stimulation, DCs had a positive regulation on the expression of CD80 and CD86 receptors, MHC class I and class II molecules, and produced significant concentrations of IL-12 [29]. In contrast, macrophages did not show changes in the cellular phenotype nor did they secrete IL-12. However, macrophages were able to produce significant concentrations of IL-6, TNF-α and IL-10 [29,31]. Macrophages stimulated with DNA-hsp65 had a higher bactericidal capacity than DCs and were more efficient in restricting the growth of microorganisms in assays following in vitro cell infection [29].
Despite clear activation of the innate and adaptive immune systems by naked DNA-hsp65, denoting their high immunostimulatory potential, the response can be improved using adjuvants constituted by many different compounds. For instance, lipidic emulsions, poly lactic-co-glycolic acid (PLGA) microsphere and liposome formulations as adjuvant delivery systems indeed showed a significant enhancement of the efficacy of the vaccine in response to challenge with M. tuberculosis in a murine model [46,47,48,49,50,51,52,53].
A number of other immunogenic M. tuberculosis antigens, such as rv2190c [54], rv1733c [55], Ag85A [56,57], Ag85B [58,59], Mtb32C [60], and fusion proteins such as HspX-PPE44-EsxV [61] or ESAT6/Ag85A [62] have been expressed in plasmid DNA vectors.
Plasmid DNA TB vaccines can trigger both cell-mediated and humoral immune responses by activating CD4+/CD8+ T and B cells. However, the major disadvantage of plasmid DNA vaccines is low transfection efficiency and the requirement of repeated intramuscular injections. Due of these drawbacks, efforts are being made to increase the immunogenicity of DNA vaccines by improving the DNA delivery techniques. One approach is to increase the efficiency of plasmid-based DNA vaccination by electroporation (EP). In vivo EP is an efficient vaccine strategy that enhances cell permeability, local tissue inflammation and immunogenicity in experimental animals [63,64,65,66,67,68] and in humans [69,70,71,72]. When EP is applied to the target organism, the encoded antigens are expressed and elicit the corresponding immune response, with the potential to induce antibody-mediated, helper T cell-mediated, and cytotoxic T cell-mediated immune responses [73]. TB DNA vaccine administered by EP had been tested in mice [74,75,76], pigs [63] and non-human primates [77,78]. As far as we know, an EP-DNA vaccine against tuberculosis has not yet been tested in humans. Our goal was to determine the safety and immunogenicity induced by the hsp65-DNA vaccine administrated by muscular EP in cynomolgus macaques (Macaca fascicularis), a nonhuman primate model that closely resembles humans in clinical and immune response characteristics of human M. tuberculosis infection.
2. Materials and Methods
2.1. Animal Model and Medical History
Animal selection, quarantine before vaccination and medical history: Twenty-one male adult cynomolgus macaques, ranging from seven to 10 years in age and weighting 6.0 to 7.6 kg were selected from the Animal Breeding Center from the Institute of Science and Technology in Biomodels, Oswaldo Cruz Foundation (Fiocruz), Rio de Janeiro, Brazil. The Oswaldo Cruz Foundation Guidelines of the Animal Care and Use Committee (IACUC/Fiocruz) approved the project (protocol number PO184-03).
Serologic screening was conducted for IgG and IgM anti-cytomegalovirus (CMV), toxoplasmosis, hepatitis B (HBV) and Trypanosoma cruzi (Chagas disease) infection, and clinical chemistry, hematological and immunological profiles were established. Nine of the 21 animals were excluded from the study because they were IgG-positive for CMV and/or toxoplasmosis. Animals were housed in individual cages for a total of twelve months (four before vaccination and six after the first dose of the vaccine). They were housed under controlled conditions of temperature, light (12-h light/12-h dark cycles) and humidity. Food and water were available ad libitum. Animals were monitored at least twice daily and fed commercial monkey chow, and with seasonal fruit once a day by trained personnel. Environmental enrichment consisted of toys and balls filed with treats, visual (TV and mirrors) and audio enrichment.
Behavior histories were assessed daily, three to four months before and after vaccination to ensure absence of any abnormality.
The animals were sedated by intramuscular injection of 5% (10mg/kg) ketamine hydrochloride. Blood samples were collected via femoral venipuncture into Vacutainer tubes.
Animals were submitted to X-ray analysis 30-45 days before the first vaccination and after the last vaccination.
Tuberculin skin test (TST) analyses were performed twice before and four months after the first vaccine dose using 0.1 ml of mammalian tuberculin via intradermal palpebral skin test (Synbiotics, San Diego, California, USA). Palpebral reactions were graded at 48 and 72h with the standard 1 to 5 scoring system [79]. Grades were as follows: 0 - no reaction; 1 - bruise - extravasation of blood in the eyelid associated with the injection of tuberculin; 2 - varying degrees of palpebral erythema with minimal swelling; 3 - moderate swelling with or without erythema; 4 - obvious palpebral swelling with drooping and varying degrees of erythema and 5 - marked swelling with necrosis and eyelid closed or partially closed were used. Grades interpretation: Grades 0, 1 and 2 were considered a TST negative; grades 3, 4 and 5 were considered a TST positive.
2.2. Vaccine Schedule and Electroporation
Vaccination was performed by injection of 1,000μg pVAX-hsp65-DNA in 1mL of plasmid DNA diluted in saline into the quadriceps muscle of eight monkeys. The injection site was subject to EP using equipment (Inovio Biomedical Co. - San Diego/CA - USA) adjusted according to the following parameters: Time=60; Ntrain=1; Nsequence=2; Volt=200. The animals from control group (n=4) were immunized with an empty plasmid DNA (DNA-pVAX) and subjected to the same EP protocol. Animals were boosted at four-week intervals for three vaccinations. The vaccination scheme and laboratory tests are represented in Figure 1.
2.3. Genetic Vaccine Construction
The vaccine DNA-hsp65 was derived from the pVAX vector (Invitrogen®, Carlsbad, CA, USA), previously digested with BamHI and NotI (Gibco BRL, Gaithersburg, MD, USA) by inserting a 3.3-kb fragment corresponding to the M. leprae hsp65 gene and the CMV intron A. The empty DNA-pVAX vector was used as the control. DH5a Escherichia coli transformed with DNA-pVAX plasmid or the plasmid carrying the hsp65 gene (DNA-hsp65) were cultured in LB liquid medium (Gibco BRL) containing 100µg/ml of ampicillin. The plasmids were purified using the Concert High Purity Maxiprep System (Gibco BRL). Plasmid concentrations were determined by spectrophotometry at k=260nm and 280nm by using the Gene Quant II apparatus (Pharmacia Biotech, Buckinghamshire, UK).
2.4. Clinical Signs and Vaccine Safety
The animals were assessed daily to identify changes in behavior, such as appetite loss, refusal to receive snacks, apathy or aggressiveness; and signs as local pain, itching, redness and swelling. Systemic analysis was conducted monthly, including fever, weight loss, bruises, anemia, red and white cell counts, and renal and hepatic enzyme alteration.
2.5. Cellular Immune Profile
Whole blood and specific antibody mixtures were incubated in the dark at room temperature for 30 min. Then, a FACS™ Lysing solution (BD, USA) was added, and the mixture was gentile homogenized. After incubation for 15 minutes, samples were washed with 4mL of cold phosphate-buffered saline (PBS), pH 7.2. At the end of the protocol, 300μL of 2% paraformaldehyde solution in PBS, pH7.2, was added. Final volume was adjusted with 800μl PBS, pH7.2. Data acquisition was performed using a Dako Cyan ADP Flow cytometer, and results were analyzed with Summit V4.3 (Dako Colorado, USA). The following fluorochrome-conjugated anti-human monoclonal antibodies (MAb) that cross-react with cynomolgus monkey cellular antigens were used (Beckson-Dickinson, BD Pharmigem, USA): CD3 APC-Cy7 or FITC (SP34), CD4 APC or PE-Cy7 (L200), CD8 PercP-Cy5 (SK1), CD11c (S-HCL-3), CD14 FITC (M5E2), CD16 ECD or FITC (3G8), CD20 PercP (L27) or FITC (2H7), CD28 APC (CD28.2), CD95 PE (DX2), CD123 PE (7G3) and HLA-DR ECD (Immu 357).
Negative lineage (Lin-) CD3-/CD14-/CD16-/CD20- was gated to characterize myeloid dendritic cells (DCs) - mDCs (Lin-, HLA-DR+/CD11c+/CD123-) and plasmacytoid - pDCs (Lin-, HLA-DR+/CD11c-/CD123+) phenotypes. Central memory (CM, CD3+/CD4+ or CD3+/CD4- and CD28+/CD95+) and effector memory (EM., CD3+/CD4+ or CD3+/CD4- and CD28-/CD95+) T lymphocytes subpopulations were analyzed.
2.6. HSP65 Peptide Specific Cellular Stimulation
Freshly isolated PBMC were incubated with monoclonal anti-human CD28 (clone 28.2; BD) and CD49d (clone 9F10; BD), each at a final concentration of 10µg/ml, in tubes slanted 5° from horizontal at 37°C with 5% CO2, as described by Gauduin et al., 2004 [80].
After this first stimulus, cells (1x106) were washed and stimulated with 10 µg/mL of 105 HSP65 (15- to 20-mer overlapping AA) peptides pool (PEPscreen®, Sigma-Aldrich, Supplemental Table S1). The internal controls of the experiment included stimulation with a combination of 100 ng/mL of both Staphylococcal enterotoxin A and B (SEA and SEB, Sigma, USA) or RPMI medium. Cells were incubated for 16h at 37ºC in a humidity chamber. Supernatants were recovered for Th1 and Th2 cytokine stimulation. The cells were resuspended in 300µl RPMI and 10 µg/mL of Brefeldin A (Sigma). After 6h of incubation, cells were recovered, washed, and incubated at 4ºC for 30 min with the following antibodies for surface markers with anti-human: CD3 APC-Cy7 (SP34), CD4 APC (L200), CD8 PercP-Cy5 (SK1), CD14 FITC (M5E2) and CD69 PE (FN50) from BD (USA) ; CD16 ECD (3G8) and CD20 APC (B9E9) from Beckman-Coulter (USA). Cells were fixed using Cytofix (BD), washed and permeabilized by Permwash (BD). Cells were staining at 4ºC for 30 min with intracellular markers with anti-human: IFN-γ FITC (B27), TNF-α PE (MAb11) and Ki-67 FITC (B56) ); anti-IL-10 PE (JES3-9D7) and Granzyme B APC (GB12) from Caltag-Life Technology (USA), IL-12 FITC (MT618) and Perforin FITC (Pf-344) (Mabtech Inc, USA). Data acquisition was performed using a Dako Cyan ADP Flow cytometer, and results were analyzed with Summit V4.3 (Dako).
2.7. Th1 and Th2 Cytokines Induced by EP-pVAX-hsp65-DNA Vaccine
The cultured cell supernatants collected just before Brefeldin A administration were stored frozen at -80°C. A Non-Human Primate Th1/Th2 Cytokine Kit (Cytometric Bead Array, BD Pharmingen) was used to quantify IFN-γ, TNF-α, IL-2, IL-4, IL-5 and IL-6 cytokines. Positive and negative internal controls were cultured cells stimulated with and without SEA and SEB. The samples were thawed and centrifuged at 5,000g for 5 minutes at 4°C to remove any precipitate. The supernatants were then incubated in darkness with the microspheres for 3h at room temperature and washed. Data acquisition was performed using a Dako Cyan ADP Flow cytometer and results were analyzed with Summit V4.3 (Dako).
2.8. Muscle and Lung Histopathology
Samples of right and left quadriceps muscle, removed from the EP injection site; and right and left upper, medium and lower lung lobe tissues samples; were collected and maintained in 10% neutral buffered formalin or in tissue-tek Optimum Cutting Temperature (OCT, Sakura, CA, USA) at room temperature or at -800C, respectively. Tissues were sectioned and stained by hematoxylin and eosin. Histopathology analysis was performed to identify tissue alterations and infiltration of inflammatory cells, particularly lymphocytes and macrophages.
2.9. Outcomes
The primary safety outcome was determined by changes in laboratory (hematology, renal and hepatic values), behavioral, and physical parameters. The secondary outcome was determined by the increase in peripheral immune cells and their activation and the increase in cytotoxic markers (CD69+, perforin and granzyme) and cytokines (mainly IFN-γ and TNF-α) during the post-vaccination period.
2.10. Statistical Analysis
Statistical analysis was carried out with Prism 9.0 software (Graph-Pad, San Diego, CA). The two-way ANOVA with the Sidak correction was used to compare DNA-hsp65 vaccinated and control group. The Sidak multiple comparisons test was used to compare statistical differences between the groups for each time point evaluated, before and after vaccination. Paired intragroup comparisons were used to analyze statistical differences among time points. A p value of <0.05 was considered significant. Clinical, biochemistry, hematological and immunological data were analyzed as follows: i) data from the DNA-hsp65 vaccinated group was compared with their own baseline data by paired T test analysis, and ii) data from the EP-pVAX-hsp65-DNA vaccinated group was compared with the control group (intergroup unpaired analysis).
3. Results
3.1. Animal Preclinical Parameters
Clinical data were collected over 12 months. All of the 21 selected animals were IgM negative for CMV, toxoplasmosis, Trypanosoma cruzi and HBV infection, and 12 of them were IgG negative. The other nine animals were IgG-positive for CMV and/or toxoplasmosis and were excluded from the study. All laboratory results were within the normal parameters described by the Center for Nonhuman Primates of the Institute of Science and Technology in Biomodels, Oswaldo Cruz Foundation, for male and age-matched cynomolgus macaques. The animals that were identified during the pre-vaccination period to harbor intestinal parasites (such as Blastocystis hominis, Balantidium coli, Iodomoeba butschlii, Entamoeba histolytica and E. coli) were treated with 90 mg/kg of Secnidazol. The TST was negative (grade zero), and no pulmonary abnormalities were identified in the X-ray analysis.
3.2. DNA-Hsp65 Vaccine is Safe with No Adverse Events
Twelve monkeys were vaccinated and followed for six months. None of the vaccinated animals showed cutaneous alterations at the vaccine administration site or at any other body site, nor changes in body weight, temperature, food consumption, behavior, morbidity or mortality during the entire experiment. There were no vaccine-related effects on hematological profiles (Supplemental Table S2) or renal and hepatic profiles (Supplemental Table S3), compared to the pre-vaccination parameters.
3.3. DNA-Hsp65 Vaccine Did Not Induce Tuberculin Skin Test Conversion or Pulmonary X-ray Alteration
Three doses of DNA-hsp65 vaccine did not result in TST conversion. Five (62.5%) of the eight vaccinated animals showed no reaction (TST grade zero, Figure 2A), and three showed only the presence of bruises (TST grade 1, Figure 2B) at 72h. Two of four control animals displayed erythema without edema (TST grade 2), and the other two displayed only bruises (TST grade 1) at the site of injection. Pulmonary X-rays were taken 30 days after the third vaccine dose, and no alterations were identified in vaccinated or control animals (data not shown).
3.4. Peripheral Cell Immune Profile and Activation Markers Induced by DNA-Hsp65 Vaccine
No differences were identified in the frequency of peripheral CD4+ and CD8+ T, B, or NK cells or in mDCs and pDCs (Figure 3). Indeed, in five of eight vaccinated animals, an increase in CM CD4+ T cells at 30 days post-vaccination period was observed compared to baseline data; however, no statistical difference was demonstrated, perhaps because of the large variation in baseline values (19.2 to 74%). No changes were induced in the CM or EM T CD8+ cells (Figure 4) by the DNA-hsp65 vaccine. The expression of CD69+ activation marker in CD8+, CD4+T and B cells increased transitorily in three of the eight vaccinated animals, with no statistical difference at 120 days post vaccination (Figure 5). No changes in the HLA-DR+ expressing CD4+ T and B cells were observed (Figure 5).
3.5. Proliferative, Lytic, and Apoptotic Markers Induced by DNA-Hsp65 Vaccine
There was no alteration in the intracellular expression of Ki-67+ marker in either T or B cells. However, Ki-67+ showed augmented expression on B cells in four of the eight vaccinated animals at 120 days post-vaccination. Granzyme B+ and perforin+ granules, and BCL-2+ markers, showed no statistical difference, after stimulation with a pool of HSP65 peptides in DNA-hsp65 vaccinated animals (Figure 6).
3.6. Cytokines Induced by DNA-Hsp65 Vaccine
De novo cytokine modulation (IFN-γ, TNF-α, IL-10 and IL-12) was assessed by intracellular flow cytometry at 0-, 60-, 120- and 180-days post DNA-hsp65 vaccination. No statistical differences were observed among the time points. Two of the eight animals showed increased CD4+, CD8+ T and NK cells expressing IFN-γ at 60- and 120-days post vaccination. CD4+ T cells expressing TNF-α were increased in four of eight vaccinated animals at 120 days post vaccination; and CD8+ T and NK cells were increased in four of eight vaccinated animals at 180 days post vaccination (Figure 7). This transient increase in cytokine expression occurred after the third dose of the vaccine.
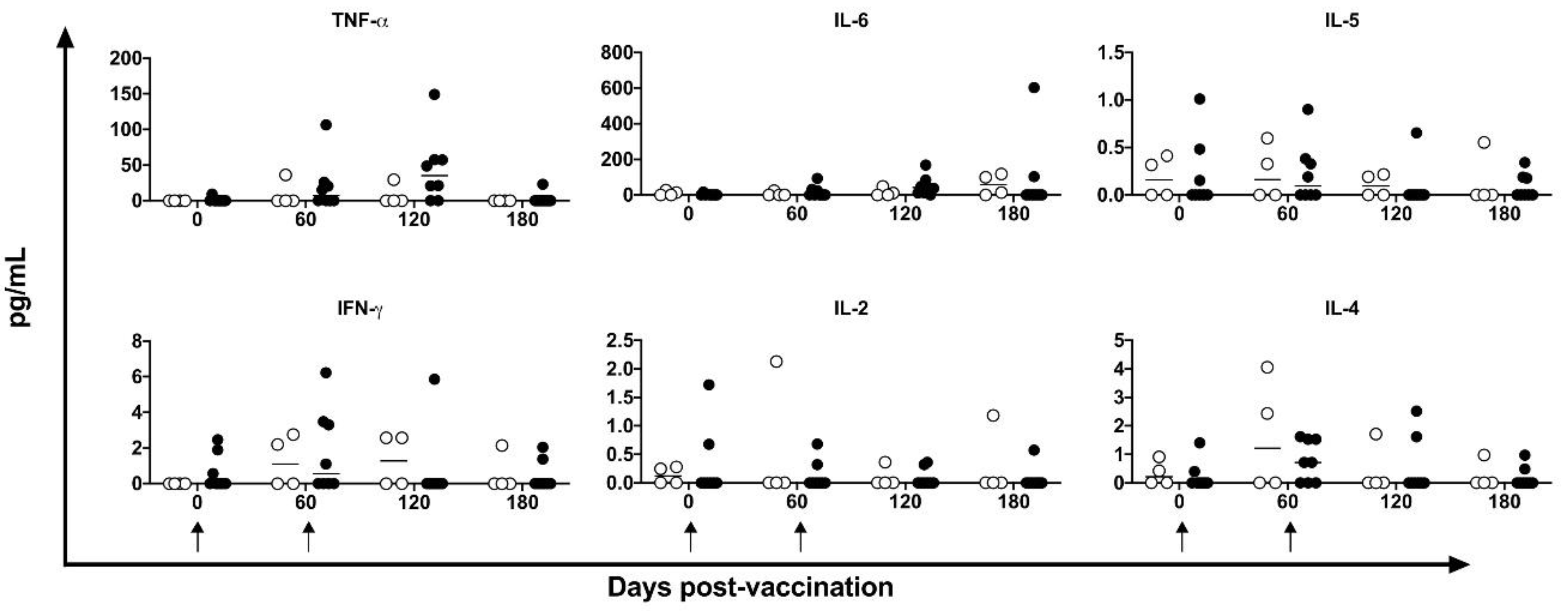
The supernatants of the PBMC cultures were assayed for Th1/Th2 profile using a non-human primate CBA kit. No statistically significant difference was observed after a two-way ANOVA test at any time point for any cytokine evaluated (Figure 8). However, the level of TNF-α was higher in four of the eight vaccinated animals at 120 days post-vaccination. Although the level was 6.01-fold higher in vaccinated animals (44.32±48.47 pg/mL) compared to unvaccinated animals (7.37±14.32 pg/mL), there was a large within-group difference. The IL-6 levels showed a similar profile to the TNF-α profile.
3.7. Muscle and Lung Histopathological Analysis
Histopathology analysis of all lung sections presented no structural alterations, neither relevant inflammatory cells infiltration (data not shown). Cases of anthracosis were reported, without relations with vaccine immunization since it was either present in the healthy animals. A focal and sparse mononuclear inflammatory process was seen in both side of the quadriceps muscle tissue mainly inter muscular fibers and in the conjunctive tissue, without fibrinoid necrosis. This feature was observed in all vaccinated animals (Supplemental Figure S1).
4. Discussion
Previously, we demonstrated that DNA-hsp65 confers a high level of protection in mice both prophylactically and therapeutically [2]: in heavily infected mice, simply by giving DNA-hsp65 immunotherapy, the immune response can be caused to switch from one that is relatively inefficient and gives bacterial stasis (Th2) to one that kills the bacteria (Th1), and persistent bacteria can be eliminated [5,6,7,8]. Here we report the results of the first test of DNA-hsp65 immunogenicity and safety in a primate species using an EP-assisted delivery system DNA vaccine.
Enhancement of naked DNA vaccination through EP has emerged as a new technology to increase the efficacy of DNA vaccines administration and has been shown to be safe, tolerable, and acceptable in most healthy human trial participants [72,81,82,83,84].
Cynomolgus macaques have been extensively used as an animal model to several preclinical vaccine trials, using an EP-assisted delivery system, including DNA targets against hemorrhagic fever virus [85], Lassa virus [86] Ebola [87], Marburg virus [88], SHIV [89], Venezuelan equine encephalitis virus [90] and HIV [91]. The results of this first preclinical trial demonstrate that DNA-hsp65 vaccine administered by muscular injection through EP is safe and well tolerated in cynomolgus macaques. No significant differences in local or systemic parameters were observed between vaccinated and control animals at a dose of 1,000 µg DNA. None of the animals showed reactional lesions or behavioral changes indicative of local discomfort or pain, or abnormal systemic alterations related to vaccination, suggesting that this vaccination scheme is likely to be safe for advancement to a clinical trial. None of the DNA-hsp65 vaccinated animals converted the TST: this result suggests that it will be possible to distinguish between people vaccinated with this vaccine and those with latent or active M. tuberculosis infection.
Naked plasmid DNA vaccination induces poor immune response in large animal models, including nonhuman primates, as well as in humans. Previous results using HIV-1 Gag DNA vaccine with or without IL-12 and/or IL-15 plasmid adjuvant did not induce strong cellular immunity in healthy human subjects [92].
The primary concept for EP-derived vaccination is the uptake of DNA into various cell types, such as subcutaneous, muscle, and dendritic cells, in which it reaches the nucleus and initiates gene transcription, protein production and post-translational modifications. New exogenous proteins formed are presented in the context of HLA class I and II molecules [73], resulting in an expected increase in a specific immune response. In our NHP model, the DNA-hsp65 vaccination system induced transient cellular immunogenicity in peripheral blood. To our knowledge, only one previous report described the immunogenicity or efficacy of an EP-DNA vaccine against tuberculosis in NHP. In that previous report, three doses (0.5 mg) of EP and non-EP Ag85A/ESAT6 DNA vaccine induced equal amounts of specific antibodies in rhesus macaques. After a booster with both Ag85A and ESAT6 proteins, the levels of antibodies increased 7-8 times [77]. The same profile was seen in rhesus macaques vaccinated with a cocktail of EP-Gag/Env-DNA vaccine; where the frequencies of granzyme B+ and CD4+ and CD8+ T cells expressing IFN-γ+ were low (0.02 -0.08%), even after one year post-vaccination [93]. An increment of humoral immune response to this vaccine prototype was achieved when the animals were boosted with both Gag/Env proteins [78]. No differences in the frequency of CD4+ and CD8+ T cells, or in the activation marker CD69+, or even in the CM, EF or naïve cells in the peripheral blood were observed in rhesus macaques vaccinated with a Mtb mutant in SigH (MtbΔsigH) vaccine through the mucosal route [94]. Future studies using BCG vaccine as a prime and HSP65 as a booster may enhance the peripheral immune response of the DNA-hsp65 in NHP model.
In the tuberculosis vaccine pipeline, the candidate M72/AS01E formulation has proceeded to several clinical trials. A phase II, double blind randomized, controlled clinical trial conducted in India proved its immunogenicity for up to three years. The analysis showed no specific-CD8+ T cells response after vaccination, as we observed in our pre-clinical model. Regarding the specific-CD4+ T cells, the frequency dropped from seven months post-vaccination to three years. The levels of cells expressing at least two cytokines were 0.15% [95]. In our study, only CD4+ and CD8+ T cells expressing TNF-α were reported transitorily at 120 days post-vaccination in 50% of the vaccinated animals. IFN-γ is a critical cytokine for control of M. tuberculosis infection; however, its correlation with vaccine protection in NHP is controversial [96].
Although it is widely accepted that CD4+ T cells play an essential role in the resistance to M. tuberculosis infection in experimental animals, their role is still unclear in human latent tuberculosis infection and disease. The absence of a high number of CD4+ T cells in our study does not mean that the animals were not efficiently immunized. Even in the site of infection, CD4+ T cells do not have an abundant profile. In an intravital image approach, reduced specific T cells and low cytokine secretion were not markedly present within the granuloma [97]. Our knowledge of the role of CD4+ T cells in the control of M. tuberculosis infection, in the peripheral blood as well as at the site of infection, is still incomplete. On the other hand, many in vivo and in vitro studies have provided evidence of the significant role of CD8+ T cells in control the disease [98]. In our study, six of eight immunized animals showed a slight increase above the normal range in frequency of CD8+ T cells at 120 days after DNA-hsp65 vaccination. This transient increase might reflect the presence of EF CD8+ T cells in the vaccinated animals. The large variation in levels of these cells in the vaccinated animals prevents the identification of subtle changes in these populations. Therefore, the absence of significant differences prevents us from reaching a clear conclusion about the role of these cells in DNA-hsp65 vaccination. This transient increase might reflect the posterior migration of CD8+ T cells in the vaccinated animals. Differences in activation markers in lung resident and peripheral CD4+ and CD8+ T cells were reported previously by Sallin et al [99] and reviewed by Lewinsohn & Lewinsohn [100] and clearly suggest that tissue location is an important factor to vaccine design.
Pre-clinical studies in NHP have an impediment in obtaining statistically significant results because of the typical small sample sizes that are practical, and the large individual and genetic differences among animals. These disadvantages were identified in our study, where the high level of inter-individual variation in the values of the immunological parameters in the vaccinated animals prevents the identification of subtle alterations related to the control group. Therefore, the absence of significant differences prevents us from reaching a clear conclusion about the role of these cells in DNA-hsp65 vaccination.
Despite the lower magnitude of the immune cell and activation marker frequencies in the peripheral blood, our results suggest that our DNA-hsp65 vaccination regimen is safe, but provided limited ability to enhance peripheral cellular immune responses in the current vaccine format. Future preclinical trials with this vaccine delivered via EP may include a combination of plasmid cytokine adjuvant and/or protein prime-boost regimen, to help the induction of robust cellular immune responses.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, MGBA, CLS, JLV.; methodology, MGBA, MRL, ACL, ALS, MMB, ATGF, JKRC, EDCG.; animal care and veterinary support: JKRC, MRL; formal analysis, MGBA, MRL, ACL, EHR; investigation, MGBA, CLS, JLV; resources, MGBA, CLS, JLV; data curation, ACL and EHR; writing—original draft preparation, MRL and ACL.; writing—review and editing, MGBA, ACL, EHR, GGS, JLV, CLS; supervision, MGBA, CLS, JLV; project administration, MGBA and CLS.; funding acquisition, MGBA, CLS, JLV. All authors have read and agreed to the published version of the manuscript.”
Funding
The research was funded in part by NIH grant R01 AI065697 to JLV; FAPESP and CNPq to CLS; and PAPES/Fiocruz to MGBA. Funding sources had no role in writing the manuscript or in the decision to submit for publication.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of Fundação Oswaldo Cruz (protocol number P0184-03, approved in 11/09/2010).
Data Availability Statement
Data will be made available on request.
Acknowledgments
We gratefully acknowledge the technicians Izaira Tincani Brandão and Wendy Martin Rios from the Department of Biochemistry and Immunology, Ribeirão Preto Medical School, University of São Paulo, for excellent technical support. The animal technicians and veterinarians at the Animal Breading Center from the Instituto de Ciência e Tecnologia em Institute of Science and Technology in Biomodels, Oswaldo Cruz Foundation (Fiocruz), Rio de Janeiro, Brazil are likewise gratefully acknowledge for their excellent assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- WHO Global Tuberculosis Report 2022; 2022.
- Silva, C.L.; Malardo, T.; Tahyra, A.S.C. Immunotherapeutic Activities of a DNA Plasmid Carrying the Mycobacterial Hsp65 Gene (DNAhsp65). Front. Med. Technol. 2020, 2, 603690. [Google Scholar] [CrossRef]
- Lowrie, D.B.; Tascon, R.E.; Colston, M.J.; Silva, C.L. Towards a DNA Vaccine against Tuberculosis. Vaccine 1994, 12, 1537–1540. [Google Scholar] [CrossRef]
- Gonçalves, E.D.C.; Bonato, V.L.D.; da Fonseca, D.M.; Soares, E.G.; Brandão, I.T.; Soares, A.P.M.; Silva, C.L. Improve Protective Efficacy of a TB DNA-HSP65 Vaccine by BCG Priming. Genet. Vaccines Ther. 2007, 5, 7. [Google Scholar] [CrossRef]
- Lowrie, D.B.; Tascon, R.E.; Bonato, V.L.; Lima, V.M.; Faccioli, L.H.; Stavropoulos, E.; Colston, M.J.; Hewinson, R.G.; Moelling, K.; Silva, C.L. Therapy of Tuberculosis in Mice by DNA Vaccination. Nature 1999, 400, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Lowrie, D.B.; Silva, C.L. Enhancement of Immunocompetence in Tuberculosis by DNA Vaccination. Vaccine 2000, 18, 1712–1716. [Google Scholar] [CrossRef]
- Rodrigues, R.F.; Zárate-Bladés, C.R.; Rios, W.M.; Soares, L.S.; Souza, P.R.M.; Brandão, I.T.; Masson, A.P.; Arnoldi, F.G.C.; Ramos, S.G.; Letourneur, F.; et al. Synergy of Chemotherapy and Immunotherapy Revealed by a Genome-Scale Analysis of Murine Tuberculosis. J. Antimicrob. Chemother. 2015, 70, 1774–1783. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.L.; Bonato, V.L.D.; Coelho-Castelo, A. a. M.; De Souza, A.O.; Santos, S.A.; Lima, K.M.; Faccioli, L.H.; Rodrigues, J.M. Immunotherapy with Plasmid DNA Encoding Mycobacterial Hsp65 in Association with Chemotherapy Is a More Rapid and Efficient Form of Treatment for Tuberculosis in Mice. Gene Ther. 2005, 12, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Frantz, F.G.; Ito, T.; Cavassani, K.A.; Hogaboam, C.M.; Lopes Silva, C.; Kunkel, S.L.; Faccioli, L.H. Therapeutic DNA Vaccine Reduces Schistosoma Mansoni-Induced Tissue Damage through Cytokine Balance and Decreased Migration of Myofibroblasts. Am. J. Pathol. 2011, 179, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Frantz, F.G.; Rosada, R.S.; Peres-Buzalaf, C.; Perusso, F.R.T.; Rodrigues, V.; Ramos, S.G.; Kunkel, S.L.; Silva, C.L.; Faccioli, L.H. Helminth Coinfection Does Not Affect Therapeutic Effect of a DNA Vaccine in Mice Harboring Tuberculosis. PLoS Negl. Trop. Dis. 2010, 4, e700. [Google Scholar] [CrossRef] [PubMed]
- Espíndola, M.S.; Frantz, F.G.; Soares, L.S.; Masson, A.P.; Tefé-Silva, C.; Bitencourt, C.S.; Oliveira, S.C.; Rodrigues, V.; Ramos, S.G.; Silva, C.L.; et al. Combined Immunization Using DNA-Sm14 and DNA-Hsp65 Increases CD8+ Memory T Cells, Reduces Chronic Pathology and Decreases Egg Viability during Schistosoma Mansoni Infection. BMC Infect. Dis. 2014, 14, 263. [Google Scholar] [CrossRef] [PubMed]
- Coelho, E.A.F.; Tavares, C.A.P.; Lima, K. de M.; Silva, C.L.; Rodrigues, J.M.; Fernandes, A.P. Mycobacterium Hsp65 DNA Entrapped into TDM-Loaded PLGA Microspheres Induces Protection in Mice against Leishmania (Leishmania) Major Infection. Parasitol. Res. 2006, 98, 568–575. [Google Scholar] [CrossRef]
- Ribeiro, A.M.; Bocca, A.L.; Amaral, A.C.; Faccioli, L.H.; Galetti, F.C.S.; Zárate-Bladés, C.R.; Figueiredo, F.; Silva, C.L.; Felipe, M.S.S. DNAhsp65 Vaccination Induces Protection in Mice against Paracoccidioides Brasiliensis Infection. Vaccine 2009, 27, 606–613. [Google Scholar] [CrossRef]
- Ribeiro, A.M.; Bocca, A.L.; Amaral, A.C.; Souza, A.C.C.O.; Faccioli, L.H.; Coelho-Castelo, A.A.M.; Figueiredo, F.; Silva, C.L.; Felipe, M.S.S. HSP65 DNA as Therapeutic Strategy to Treat Experimental Paracoccidioidomycosis. Vaccine 2010, 28, 1528–1534. [Google Scholar] [CrossRef]
- Siqueira, I.M.; Ribeiro, A.M.; Nóbrega, Y.K. de M.; Simon, K.S.; Souza, A.C.O.; Jerônimo, M.S.; Cavalcante Neto, F.F.; Silva, C.L.; Felipe, M.S.S.; Bocca, A.L. DNA-Hsp65 Vaccine as Therapeutic Strategy to Treat Experimental Chromoblastomycosis Caused by Fonsecaea Pedrosoi. Mycopathologia 2013, 175, 463–475. [Google Scholar] [CrossRef]
- Santos, R.R.; Sartori, A.; Lima, D.S.; Souza, P.R.; Coelho-Castelo, A.A.; Bonato, V.L.; Silva, C.L. DNA Vaccine Containing the Mycobacterial Hsp65 Gene Prevented Insulitis in MLD-STZ Diabetes. J. Immune Based Ther. Vaccines 2009, 7, 4. [Google Scholar] [CrossRef]
- Santos Júnior, R.R. dos; Sartori, A.; Bonato, V.L.D.; Coelho Castelo, A. a. M.; Vilella, C.A.; Zollner, R.L.; Silva, C.L. Immune Modulation Induced by Tuberculosis DNA Vaccine Protects Non-Obese Diabetic Mice from Diabetes Progression. Clin. Exp. Immunol. 2007, 149, 570–578. [Google Scholar] [CrossRef]
- Pileggi, G.S.; Clemencio, A.D.; Malardo, T.; Antonini, S.R.; Bonato, V.L.D.; Rios, W.M.; Silva, C.L. New Strategy for Testing Efficacy of Immunotherapeutic Compounds for Diabetes in Vitro. BMC Biotechnol. 2016, 16, 40. [Google Scholar] [CrossRef] [PubMed]
- da Rosa, L.C.; Chiuso-Minicucci, F.; Zorzella-Pezavento, S.F.G.; França, T.G.D.; Ishikawa, L.L.W.; Colavite, P.M.; Balbino, B.; Tavares, L.C.B.; Silva, C.L.; Marques, C.; et al. Bacille Calmette-Guérin/DNAhsp65 Prime-Boost Is Protective against Diabetes in Non-Obese Diabetic Mice but Not in the Streptozotocin Model of Type 1 Diabetes. Clin. Exp. Immunol. 2013, 173, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Santos-Junior, R.R.; Sartori, A.; De Franco, M.; Filho, O.G.R.; Coelho-Castelo, A.A.M.; Bonato, V.L.D.; Cabrera, W.H.K.; Ibañez, O.M.; Silva, C.L. Immunomodulation and Protection Induced by DNA-Hsp65 Vaccination in an Animal Model of Arthritis. Hum. Gene Ther. 2005, 16, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Narciso, C.; Pérez-Tapia, M.; Rangel-Cano, R.M.; Silva, C.L.; Meckes-Fisher, M.; Salgado-Garciglia, R.; Estrada-Parra, S.; López-Gómez, R.; Estrada-García, I. Expression of Mycobacterium Leprae HSP65 in Tobacco and Its Effectiveness as an Oral Treatment in Adjuvant-Induced Arthritis. Transgenic Res. 2011, 20, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Zorzella-Pezavento, S.F.G.; Guerino, C.P.F.; Chiuso-Minicucci, F.; França, T.G.D.; Ishikawa, L.L.W.; Masson, A.P.; Silva, C.L.; Sartori, A. BCG and BCG/DNAhsp65 Vaccinations Promote Protective Effects without Deleterious Consequences for Experimental Autoimmune Encephalomyelitis. Clin. Dev. Immunol. 2013, 2013, 721383. [Google Scholar] [CrossRef]
- Zorzella-Pezavento, S.F.G.; Chiuso-Minicucci, F.; França, T.G.D.; Ishikawa, L.L.W.; da Rosa, L.C.; Colavite, P.M.; Balbino, B.; Marques, C.; Ikoma, M.R.V.; Masson, A.P.; et al. PVAXhsp65 Vaccination Primes for High IL-10 Production and Decreases Experimental Encephalomyelitis Severity. J. Immunol. Res. 2017, 2017, 6257958. [Google Scholar] [CrossRef]
- Zorzella-Pezavento, S.F.G.; Chiuso-Minicucci, F.; França, T.G.D.; Ishikawa, L.L.W.; da Rosa, L.C.; Colavite, P.M.; Marques, C.; Ikoma, M.R.V.; Silva, C.L.; Sartori, A. Downmodulation of Peripheral MOG-Specific Immunity by PVAXhsp65 Treatment during EAE Does Not Reach the CNS. J. Neuroimmunol. 2014, 268, 35–42. [Google Scholar] [CrossRef]
- Fonseca, D.M.; Bonato, V.L.D.; Silva, C.L.; Sartori, A. Th1 Polarized Response Induced by Intramuscular DNA-HSP65 Immunization Is Preserved in Experimental Atherosclerosis. Braz. J. Med. Biol. Res. 2007, 40, 1495–1504. [Google Scholar] [CrossRef]
- Prado, R.Q.; Bertolini, T.B.; Piñeros, A.R.; Gembre, A.F.; Ramos, S.G.; Silva, C.L.; Borges, M.C.; Bonato, V.L.D. Attenuation of Experimental Asthma by Mycobacterial Protein Combined with CpG Requires a TLR9-Dependent IFN-γ-CCR2 Signalling Circuit. Clin. Exp. Allergy 2015, 45, 1459–1471. [Google Scholar] [CrossRef]
- Fonseca, D.M.; Wowk, P.F.; Paula, M.O.; Gembre, A.F.; Baruffi, M.D.; Fermino, M.L.; Turato, W.M.; Campos, L.W.; Silva, C.L.; Ramos, S.G.; et al. Requirement of MyD88 and Fas Pathways for the Efficacy of Allergen-Free Immunotherapy. Allergy 2015, 70, 275–284. [Google Scholar] [CrossRef]
- Fonseca, D.M.; Wowk, P.F.; Paula, M.O.; Campos, L.W.; Gembre, A.F.; Turato, W.M.; Ramos, S.G.; Dias-Baruffi, M.; Barboza, R.; Gomes, E.; et al. Recombinant DNA Immunotherapy Ameliorate Established Airway Allergy in a IL-10 Dependent Pathway. Clin. Exp. Allergy 2012, 42, 131–143. [Google Scholar] [CrossRef]
- Franco, L.H.; Wowk, P.F.; Silva, C.L.; Trombone, A.P.F.; Coelho-Castelo, A.A.M.; Oliver, C.; Jamur, M.C.; Moretto, E.L.; Bonato, V.L.D. A DNA Vaccine against Tuberculosis Based on the 65 KDa Heat-Shock Protein Differentially Activates Human Macrophages and Dendritic Cells. Genet. Vaccines Ther. 2008, 6, 3. [Google Scholar] [CrossRef]
- Binder, R.J. Functions of Heat Shock Proteins in Pathways of the Innate and Adaptive Immune System. J. Immunol. 2014, 193, 5765–5771. [Google Scholar] [CrossRef]
- Wowk, P.F.; Franco, L.H.; Fonseca, D.M. da; Paula, M.O.; Vianna, É.D.S.O.; Wendling, A.P.; Augusto, V.M.; Elói-Santos, S.M.; Teixeira-Carvalho, A.; Silva, F.D.C.; et al. Mycobacterial Hsp65 Antigen Upregulates the Cellular Immune Response of Healthy Individuals Compared with Tuberculosis Patients. Hum. Vaccin. Immunother. 2017, 13, 1040–1050. [Google Scholar] [CrossRef]
- Srivastava, P. Roles of Heat-Shock Proteins in Innate and Adaptive Immunity. Nat. Rev. Immunol. 2002, 2, 185–194. [Google Scholar] [CrossRef]
- Silva, C.L.; Lowrie, D.B. Identification and Characterization of Murine Cytotoxic T Cells That Kill Mycobacterium Tuberculosis. Infect. Immun. 2000, 68, 3269–3274. [Google Scholar] [CrossRef]
- Bonato, V.L.; Lima, V.M.; Tascon, R.E.; Lowrie, D.B.; Silva, C.L. Identification and Characterization of Protective T Cells in Hsp65 DNA-Vaccinated and Mycobacterium Tuberculosis-Infected Mice. Infect. Immun. 1998, 66, 169–175. [Google Scholar] [CrossRef]
- Bonato, V.L.D.; Gonçalves, E.D.C.; Soares, E.G.; Santos Júnior, R.R.; Sartori, A.; Coelho-Castelo, A. a. M.; Silva, C.L. Immune Regulatory Effect of PHSP65 DNA Therapy in Pulmonary Tuberculosis: Activation of CD8+ Cells, Interferon-Gamma Recovery and Reduction of Lung Injury. Immunology 2004, 113, 130–138. [Google Scholar] [CrossRef]
- Silva, C.L.; Bonato, V.L.; Lima, K.M.; Coelho-Castelo, A.A.; Faccioli, L.H.; Sartori, A.; De Souza, A.O.; Leão, S.C. Cytotoxic T Cells and Mycobacteria. FEMS Microbiol. Lett. 2001, 197, 11–18. [Google Scholar] [CrossRef]
- Silva, C.L.; Bonato, V.L.; Lima, V.M.; Faccioli, L.H.; Leão, S.C. Characterization of the Memory/Activated T Cells That Mediate the Long-Lived Host Response against Tuberculosis after Bacillus Calmette-Guérin or DNA Vaccination. Immunology 1999, 97, 573–581. [Google Scholar] [CrossRef]
- Zárate-Bladés, C.R.; Rodrigues, R.F.; Souza, P.R.M.; Rios, W.M.; Soares, L.S.; Rosada, R.S.; Brandão, I.T.; Masson, A.P.; Floriano, E.M.; Ramos, S.G.; et al. Evaluation of the Overall IFN-γ and IL-17 pro-Inflammatory Responses after DNA Therapy of Tuberculosis. Hum. Vaccin. Immunother. 2013, 9, 1093–1103. [Google Scholar] [CrossRef]
- Fontoura, I.C.; Trombone, A.P.F.; Almeida, L.P.; Lorenzi, J.C.C.; Rossetti, R. a. M.; Malardo, T.; Padilha, E.; Schluchting, W.; Silva, R.L.L.; Gembre, A.F.; et al. B Cells Expressing IL-10 MRNA Modulate Memory T Cells after DNA-Hsp65 Immunization. Braz. J. Med. Biol. Res. 2015, 48, 1095–1100. [Google Scholar] [CrossRef]
- Zárate-Bladés, C.R.; Bonato, V.L.D.; da Silveira, E.L.V.; Oliveira e Paula, M.; Junta, C.M.; Sandrin-Garcia, P.; Fachin, A.L.; Mello, S.S.; Cardoso, R.S.; Galetti, F.C. de S.; et al. Comprehensive Gene Expression Profiling in Lungs of Mice Infected with Mycobacterium Tuberculosis Following DNAhsp65 Immunotherapy. J. Gene Med. 2009, 11, 66–78. [Google Scholar] [CrossRef]
- Zárate-Bladés, C.R.; Silva, C.L.; Passos, G.A. The Impact of Transcriptomics on the Fight against Tuberculosis: Focus on Biomarkers, BCG Vaccination, and Immunotherapy. Clin. Dev. Immunol. 2011, 2011, 192630. [Google Scholar] [CrossRef]
- Fedatto, P.F.; Sérgio, C.A.; Paula, M.O. e; Gembre, A.F.; Franco, L.H.; Wowk, P.F.; Ramos, S.G.; Horn, C.; Marchal, G.; Turato, W.M.; et al. Protection Conferred by Heterologous Vaccination against Tuberculosis Is Dependent on the Ratio of CD4(+) /CD4(+) Foxp3(+) Cells. Immunology 2012, 137, 239–248. [Google Scholar] [CrossRef]
- Padua, A.I. de; Silva, C.L.; Ramos, S.G.; Faccioli, L.H.; Martinez, J.A.B. Influence of a DNA-Hsp65 Vaccine on Bleomycin-Induced Lung Injury. J. Bras. Pneumol. 2008, 34, 891–899. [Google Scholar] [CrossRef]
- Zucchi, F.C.R.; Tsanaclis, A.M.C.; Moura-Dias, Q.; Silva, C.L.; Pelegrini-da-Silva, A.; Neder, L.; Takayanagui, O.M. Modulation of Angiogenic Factor VEGF by DNA-Hsp65 Vaccination in a Murine CNS Tuberculosis Model. Tuberculosis (Edinb) 2013, 93, 373–380. [Google Scholar] [CrossRef]
- Almeida, L.P.; Trombone, A.P.; Lorenzi, J.C.; Rocha, C.D.; Malardo, T.; Fontoura, I.C.; Gembre, A.F.; Silva, R.L.; Silva, C.L.; Castelo, A.P.; et al. B Cells Can Modulate the CD8 Memory T Cell after DNA Vaccination Against Experimental Tuberculosis. Genet. Vaccines Ther. 2011, 9, 5. [Google Scholar] [CrossRef]
- Souza, P.R.M.; Zárate-Bladés, C.R.; Hori, J.I.; Ramos, S.G.; Lima, D.S.; Schneider, T.; Rosada, R.S.; Torre, L.G.L.; Santana, M.H.A.; Brandão, I.T.; et al. Protective Efficacy of Different Strategies Employing Mycobacterium Leprae Heat-Shock Protein 65 against Tuberculosis. Expert. Opin. Biol. Ther. 2008, 8, 1255–1264. [Google Scholar] [CrossRef]
- Silva, C.L.; De Souza, A.O. Vaccines of the Future: From Rational Design to Clinical Development. 17-19 October 2001, Paris, France. Expert Opin Biol Ther 2002, 2, 219–222. [Google Scholar] [CrossRef]
- Lima, K.M.; dos Santos, S.A.; Santos, R.R.; Brandão, I.T.; Rodrigues, J.M.; Silva, C.L. Efficacy of DNA-Hsp65 Vaccination for Tuberculosis Varies with Method of DNA Introduction in Vivo. Vaccine 2003, 22, 49–56. [Google Scholar] [CrossRef]
- Rosada, R.S.; de la Torre, L.G.; Frantz, F.G.; Trombone, A.P.F.; Zárate-Bladés, C.R.; Fonseca, D.M.; Souza, P.R.M.; Brandão, I.T.; Masson, A.P.; Soares, E.G.; et al. Protection against Tuberculosis by a Single Intranasal Administration of DNA-Hsp65 Vaccine Complexed with Cationic Liposomes. BMC Immunol. 2008, 9, 38. [Google Scholar] [CrossRef]
- Lima, K.M.; Santos, S.A.; Lima, V.M.F.; Coelho-Castelo, A. a. M.; Rodrigues, J.M.; Silva, C.L. Single Dose of a Vaccine Based on DNA Encoding Mycobacterial Hsp65 Protein plus TDM-Loaded PLGA Microspheres Protects Mice against a Virulent Strain of Mycobacterium Tuberculosis. Gene Ther. 2003, 10, 678–685. [Google Scholar] [CrossRef]
- Ribeiro, A.M.; Souza, A.C.O.; Amaral, A.C.; Vasconcelos, N.M.; Jeronimo, M.S.; Carneiro, F.P.; Faccioli, L.H.; Felipe, M.S.S.; Silva, C.L.; Bocca, A.L. Nanobiotechnological Approaches to Delivery of DNA Vaccine against Fungal Infection. J. Biomed. Nanotechnol. 2013, 9, 221–230. [Google Scholar] [CrossRef]
- Lima, K.M.; dos Santos, S.A.; Rodrigues, J.M.; Silva, C.L. Vaccine Adjuvant: It Makes the Difference. Vaccine 2004, 22, 2374–2379. [Google Scholar] [CrossRef]
- Ruberti, M.; De Melo, L.K.; Dos Santos, S.A.; Brandao, I.T.; Soares, E.G.; Silva, C.L.; Júnior, J.M.R. Prime-Boost Vaccination Based on DNA and Protein-Loaded Microspheres for Tuberculosis Prevention. J. Drug Target. 2004, 12, 195–203. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, X.; Bai, X.; Xiao, L.; Wang, X.; Zhang, J.; Yang, Y.; Song, J.; Wang, L.; Wu, X. Immunogenicity and Therapeutic Effects of a Mycobacterium Tuberculosis Rv2190c DNA Vaccine in Mice. BMC Immunol. 2017, 18, 11. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, H.; Bai, Y.-L.; Kang, J.; Xu, Z.-K.; Wang, L.-M. Construction and Immunogenicity of the DNA Vaccine of Mycobacterium Tuberculosis Dormancy Antigen Rv1733c. Scand. J. Immunol. 2014, 79, 292–298. [Google Scholar] [CrossRef]
- Li, W.; Li, M.; Deng, G.; Zhao, L.; Liu, X.; Wang, Y. Prime-Boost Vaccination with Bacillus Calmette Guerin and a Recombinant Adenovirus Co-Expressing CFP10, ESAT6, Ag85A and Ag85B of Mycobacterium Tuberculosis Induces Robust Antigen-Specific Immune Responses in Mice. Mol. Med. Rep. 2015, 12, 3073–3080. [Google Scholar] [CrossRef]
- Poecheim, J.; Barnier-Quer, C.; Collin, N.; Borchard, G. Ag85A DNA Vaccine Delivery by Nanoparticles: Influence of the Formulation Characteristics on Immune Responses. Vaccines (Basel) 2016, 4, 32. [Google Scholar] [CrossRef]
- Husain, A.A.; Warke, S.R.; Kalorey, D.R.; Daginawala, H.F.; Taori, G.M.; Kashyap, R.S. Comparative Evaluation of Booster Efficacies of BCG, Ag85B, and Ag85B Peptides Based Vaccines to Boost BCG Induced Immunity in BALB/c Mice: A Pilot Study. Clin. Exp. Vaccine Res. 2015, 4, 83–87. [Google Scholar] [CrossRef]
- Khan, A.; Sayedahmed, E.E.; Singh, V.K.; Mishra, A.; Dorta-Estremera, S.; Nookala, S.; Canaday, D.H.; Chen, M.; Wang, J.; Sastry, K.J.; et al. A Recombinant Bovine Adenoviral Mucosal Vaccine Expressing Mycobacterial Antigen-85B Generates Robust Protection against Tuberculosis in Mice. Cell Rep. Med. 2021, 2, 100372. [Google Scholar] [CrossRef]
- Teimourpour, R.; Zare, H.; Rajabnia, R.; Yahyapour, Y.; Meshkat, Z. Evaluation of the Eukaryotic Expression of Mtb32C-Hbha Fusion Gene of Mycobacterium Tuberculosis in Hepatocarcinoma Cell Line. Iran. J. Microbiol. 2016, 8, 132–138. [Google Scholar]
- Moradi, B.; Sankian, M.; Amini, Y.; Meshkat, Z. Construction of a Novel DNA Vaccine Candidate Encoding an HspX-PPE44-EsxV Fusion Antigen of Mycobacterium Tuberculosis. Rep. Biochem. Mol. Biol. 2016, 4, 89–97. [Google Scholar]
- Liang, Y.; Wu, X.; Zhang, J.; Li, N.; Yu, Q.; Yang, Y.; Bai, X.; Liu, C.; Shi, Y.; Liu, Q.; et al. The Treatment of Mice Infected with Multi-Drug-Resistant Mycobacterium Tuberculosis Using DNA Vaccines or in Combination with Rifampin. Vaccine 2008, 26, 4536–4540. [Google Scholar] [CrossRef]
- Bruffaerts, N.; Pedersen, L.E.; Vandermeulen, G.; Préat, V.; Stockhofe-Zurwieden, N.; Huygen, K.; Romano, M. Increased B and T Cell Responses in M. Bovis Bacille Calmette-Guérin Vaccinated Pigs Co-Immunized with Plasmid DNA Encoding a Prototype Tuberculosis Antigen. PLoS ONE 2015, 10, e0132288. [Google Scholar] [CrossRef]
- Datta, D.; Bansal, G.P.; Grasperge, B.; Martin, D.S.; Philipp, M.; Gerloff, D.; Ellefsen, B.; Hannaman, D.; Kumar, N. Comparative Functional Potency of DNA Vaccines Encoding Plasmodium Falciparum Transmission Blocking Target Antigens Pfs48/45 and Pfs25 Administered Alone or in Combination by in Vivo Electroporation in Rhesus Macaques. Vaccine 2017, 35, 7049–7056. [Google Scholar] [CrossRef]
- Huang, X.; Zhu, Q.; Huang, X.; Yang, L.; Song, Y.; Zhu, P.; Zhou, P. In Vivo Electroporation in DNA-VLP Prime-Boost Preferentially Enhances HIV-1 Envelope-Specific IgG2a, Neutralizing Antibody and CD8 T Cell Responses. Vaccine 2017, 35, 2042–2051. [Google Scholar] [CrossRef]
- Williams, M.; Ewing, D.; Blevins, M.; Sun, P.; Sundaram, A.K.; Raviprakash, K.S.; Porter, K.R.; Sanders, J.W. Enhanced Immunogenicity and Protective Efficacy of a Tetravalent Dengue DNA Vaccine Using Electroporation and Intradermal Delivery. Vaccine 2019, 37, 4444–4453. [Google Scholar] [CrossRef]
- Patel, A.; Walters, J.N.; Reuschel, E.L.; Schultheis, K.; Parzych, E.; Gary, E.N.; Maricic, I.; Purwar, M.; Eblimit, Z.; Walker, S.N.; et al. Intradermal-Delivered DNA Vaccine Induces Durable Immunity Mediating a Reduction in Viral Load in a Rhesus Macaque SARS-CoV-2 Challenge Model. Cell Rep. Med. 2021, 2, 100420. [Google Scholar] [CrossRef]
- Chen, J.; Deng, Y.; Huang, B.; Han, D.; Wang, W.; Huang, M.; Zhai, C.; Zhao, Z.; Yang, R.; Zhao, Y.; et al. DNA Vaccines Expressing the Envelope and Membrane Proteins Provide Partial Protection Against SARS-CoV-2 in Mice. Front. Immunol. 2022, 13, 827605. [Google Scholar] [CrossRef]
- Spearman, P.; Mulligan, M.; Anderson, E.J.; Shane, A.L.; Stephens, K.; Gibson, T.; Hartwell, B.; Hannaman, D.; Watson, N.L.; Singh, K. A Phase 1, Randomized, Controlled Dose-Escalation Study of EP-1300 Polyepitope DNA Vaccine against Plasmodium Falciparum Malaria Administered via Electroporation. Vaccine 2016, 34, 5571–5578. [Google Scholar] [CrossRef]
- Yang, F.-Q.; Rao, G.-R.; Wang, G.-Q.; Li, Y.-Q.; Xie, Y.; Zhang, Z.-Q.; Deng, C.-L.; Mao, Q.; Li, J.; Zhao, W.; et al. Phase IIb Trial of in Vivo Electroporation Mediated Dual-Plasmid Hepatitis B Virus DNA Vaccine in Chronic Hepatitis B Patients under Lamivudine Therapy. World J. Gastroenterol. 2017, 23, 306–317. [Google Scholar] [CrossRef]
- Mpendo, J.; Mutua, G.; Nanvubya, A.; Anzala, O.; Nyombayire, J.; Karita, E.; Dally, L.; Hannaman, D.; Price, M.; Fast, P.E.; et al. Acceptability and Tolerability of Repeated Intramuscular Electroporation of Multi-Antigenic HIV (HIVMAG) DNA Vaccine among Healthy African Participants in a Phase 1 Randomized Controlled Trial. PLoS ONE 2020, 15, e0233151. [Google Scholar] [CrossRef]
- Ahn, J.Y.; Lee, J.; Suh, Y.S.; Song, Y.G.; Choi, Y.-J.; Lee, K.H.; Seo, S.H.; Song, M.; Oh, J.-W.; Kim, M.; et al. Safety and Immunogenicity of Two Recombinant DNA COVID-19 Vaccines Containing the Coding Regions of the Spike or Spike and Nucleocapsid Proteins: An Interim Analysis of Two Open-Label, Non-Randomised, Phase 1 Trials in Healthy Adults. Lancet Microbe 2022, 3, e173–e183. [Google Scholar] [CrossRef]
- Sardesai, N.Y.; Weiner, D.B. Electroporation Delivery of DNA Vaccines: Prospects for Success. Curr. Opin. Immunol. 2011, 23, 421–429. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, L.; Xiao, L.; Liu, X.; Yang, Y.; Ling, Y.; Wang, T.; Wang, L.; Wang, J.; Wu, X. Immunotherapeutic Effects of Different Doses of Mycobacterium Tuberculosis Ag85a/b DNA Vaccine Delivered by Electroporation. Front. Immunol. 2022, 13, 876579. [Google Scholar] [CrossRef]
- Villarreal, D.O.; Walters, J.; Laddy, D.J.; Yan, J.; Weiner, D.B. Multivalent TB Vaccines Targeting the Esx Gene Family Generate Potent and Broad Cell-Mediated Immune Responses Superior to BCG. Hum. Vaccin. Immunother. 2014, 10, 2188–2198. [Google Scholar] [CrossRef]
- Tang, J.; Cai, Y.; Liang, J.; Tan, Z.; Tang, X.; Zhang, C.; Cheng, L.; Zhou, J.; Wang, H.; Yam, W.-C.; et al. In Vivo Electroporation of a Codon-Optimized BERopt DNA Vaccine Protects Mice from Pathogenic Mycobacterium Tuberculosis Aerosol Challenge. Tuberculosis (Edinb) 2018, 113, 65–75. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, H.; Fan, X.; Zhang, Y.; Huang, J.; Liu, Q.; Tjelle, T.E.; Mathiesen, I.; Kjeken, R.; Xiong, S. DNA Electroporation Prime and Protein Boost Strategy Enhances Humoral Immunity of Tuberculosis DNA Vaccines in Mice and Non-Human Primates. Vaccine 2006, 24, 4565–4568. [Google Scholar] [CrossRef]
- Otten, G.; Schaefer, M.; Doe, B.; Liu, H.; Srivastava, I.; zur Megede, J.; O’Hagan, D.; Donnelly, J.; Widera, G.; Rabussay, D.; et al. Enhancement of DNA Vaccine Potency in Rhesus Macaques by Electroporation. Vaccine 2004, 22, 2489–2493. [Google Scholar] [CrossRef]
- Kennard, M.A.; Schroeder, C.R.; Trask, J.D.; Paul, J.R. A CUTANEOUS TEST FOR TUBERCULOSIS IN PRIMATES. Science 1939, 89, 442–443. [Google Scholar] [CrossRef]
- Gauduin, M.-C.; Kaur, A.; Ahmad, S.; Yilma, T.; Lifson, J.D.; Johnson, R.P. Optimization of Intracellular Cytokine Staining for the Quantitation of Antigen-Specific CD4+ T Cell Responses in Rhesus Macaques. J. Immunol. Methods 2004, 288, 61–79. [Google Scholar] [CrossRef]
- Diehl, M.C.; Lee, J.C.; Daniels, S.E.; Tebas, P.; Khan, A.S.; Giffear, M.; Sardesai, N.Y.; Bagarazzi, M.L. Tolerability of Intramuscular and Intradermal Delivery by CELLECTRA(®) Adaptive Constant Current Electroporation Device in Healthy Volunteers. Hum. Vaccin. Immunother. 2013, 9, 2246–2252. [Google Scholar] [CrossRef]
- Kalams, S.A.; Parker, S.D.; Elizaga, M.; Metch, B.; Edupuganti, S.; Hural, J.; De Rosa, S.; Carter, D.K.; Rybczyk, K.; Frank, I.; et al. Safety and Comparative Immunogenicity of an HIV-1 DNA Vaccine in Combination with Plasmid Interleukin 12 and Impact of Intramuscular Electroporation for Delivery. J. Infect. Dis. 2013, 208, 818–829. [Google Scholar] [CrossRef]
- Elizaga, M.L.; Li, S.S.; Kochar, N.K.; Wilson, G.J.; Allen, M.A.; Tieu, H.V.N.; Frank, I.; Sobieszczyk, M.E.; Cohen, K.W.; Sanchez, B.; et al. Safety and Tolerability of HIV-1 Multiantigen PDNA Vaccine given with IL-12 Plasmid DNA via Electroporation, Boosted with a Recombinant Vesicular Stomatitis Virus HIV Gag Vaccine in Healthy Volunteers in a Randomized, Controlled Clinical Trial. PLoS ONE 2018, 13, e0202753. [Google Scholar] [CrossRef]
- Edupuganti, S.; C De Rosa, S.; Elizaga, M.; Lu, Y.; Han, X.; Huang, Y.; Swann, E.; Polakowski, L.; A Kalams, S.; Keefer, M.; et al. Intramuscular and Intradermal Electroporation of HIV-1 PENNVAX-GP® DNA Vaccine and IL-12 Is Safe, Tolerable, Acceptable in Healthy Adults. Vaccines (Basel) 2020, 8, 741. [Google Scholar] [CrossRef]
- Hawman, D.W.; Ahlén, G.; Appelberg, K.S.; Meade-White, K.; Hanley, P.W.; Scott, D.; Monteil, V.; Devignot, S.; Okumura, A.; Weber, F.; et al. A DNA-Based Vaccine Protects against Crimean-Congo Haemorrhagic Fever Virus Disease in a Cynomolgus Macaque Model. Nat. Microbiol. 2021, 6, 187–195. [Google Scholar] [CrossRef]
- Cashman, K.A.; Wilkinson, E.R.; Shaia, C.I.; Facemire, P.R.; Bell, T.M.; Bearss, J.J.; Shamblin, J.D.; Wollen, S.E.; Broderick, K.E.; Sardesai, N.Y.; et al. A DNA Vaccine Delivered by Dermal Electroporation Fully Protects Cynomolgus Macaques against Lassa Fever. Hum. Vaccin. Immunother. 2017, 13, 2902–2911. [Google Scholar] [CrossRef]
- Patel, A.; Reuschel, E.L.; Kraynyak, K.A.; Racine, T.; Park, D.H.; Scott, V.L.; Audet, J.; Amante, D.; Wise, M.C.; Keaton, A.A.; et al. Protective Efficacy and Long-Term Immunogenicity in Cynomolgus Macaques by Ebola Virus Glycoprotein Synthetic DNA Vaccines. J. Infect. Dis. 2019, 219, 544–555. [Google Scholar] [CrossRef]
- Grant-Klein, R.J.; Altamura, L.A.; Badger, C.V.; Bounds, C.E.; Van Deusen, N.M.; Kwilas, S.A.; Vu, H.A.; Warfield, K.L.; Hooper, J.W.; Hannaman, D.; et al. Codon-Optimized Filovirus DNA Vaccines Delivered by Intramuscular Electroporation Protect Cynomolgus Macaques from Lethal Ebola and Marburg Virus Challenges. Hum. Vaccin. Immunother. 2015, 11, 1991–2004. [Google Scholar] [CrossRef]
- Martinon, F.; Kaldma, K.; Sikut, R.; Culina, S.; Romain, G.; Tuomela, M.; Adojaan, M.; Männik, A.; Toots, U.; Kivisild, T.; et al. Persistent Immune Responses Induced by a Human Immunodeficiency Virus DNA Vaccine Delivered in Association with Electroporation in the Skin of Nonhuman Primates. Hum. Gene Ther. 2009, 20, 1291–1307. [Google Scholar] [CrossRef]
- Dupuy, L.C.; Richards, M.J.; Ellefsen, B.; Chau, L.; Luxembourg, A.; Hannaman, D.; Livingston, B.D.; Schmaljohn, C.S. A DNA Vaccine for Venezuelan Equine Encephalitis Virus Delivered by Intramuscular Electroporation Elicits High Levels of Neutralizing Antibodies in Multiple Animal Models and Provides Protective Immunity to Mice and Nonhuman Primates. Clin. Vaccine Immunol. 2011, 18, 707–716. [Google Scholar] [CrossRef]
- Cristillo, A.D.; Weiss, D.; Hudacik, L.; Restrepo, S.; Galmin, L.; Suschak, J.; Draghia-Akli, R.; Markham, P.; Pal, R. Persistent Antibody and T Cell Responses Induced by HIV-1 DNA Vaccine Delivered by Electroporation. Biochem. Biophys. Res. Commun. 2008, 366, 29–35. [Google Scholar] [CrossRef]
- Kalams, S.A.; Parker, S.; Jin, X.; Elizaga, M.; Metch, B.; Wang, M.; Hural, J.; Lubeck, M.; Eldridge, J.; Cardinali, M.; et al. Safety and Immunogenicity of an HIV-1 Gag DNA Vaccine with or without IL-12 and/or IL-15 Plasmid Cytokine Adjuvant in Healthy, HIV-1 Uninfected Adults. PLoS ONE 2012, 7, e29231. [Google Scholar] [CrossRef]
- Kulkarni, V.; Valentin, A.; Rosati, M.; Rolland, M.; Mullins, J.I.; Pavlakis, G.N.; Felber, B.K. HIV-1 Conserved Elements P24CE DNA Vaccine Induces Humoral Immune Responses with Broad Epitope Recognition in Macaques. PLoS ONE 2014, 9, e111085. [Google Scholar] [CrossRef]
- Kaushal, D.; Foreman, T.W.; Gautam, U.S.; Alvarez, X.; Adekambi, T.; Rangel-Moreno, J.; Golden, N.A.; Johnson, A.-M.F.; Phillips, B.L.; Ahsan, M.H.; et al. Mucosal Vaccination with Attenuated Mycobacterium Tuberculosis Induces Strong Central Memory Responses and Protects against Tuberculosis. Nat. Commun. 2015, 6, 8533. [Google Scholar] [CrossRef]
- Kumarasamy, N.; Poongulali, S.; Beulah, F.E.; Akite, E.J.; Ayuk, L.N.; Bollaerts, A.; Demoitié, M.-A.; Jongert, E.; Ofori-Anyinam, O.; Van Der Meeren, O. Long-Term Safety and Immunogenicity of the M72/AS01E Candidate Tuberculosis Vaccine in HIV-Positive and -Negative Indian Adults: Results from a Phase II Randomized Controlled Trial. Medicine (Baltimore) 2018, 97, e13120. [Google Scholar] [CrossRef]
- Larsen, M.H.; Biermann, K.; Chen, B.; Hsu, T.; Sambandamurthy, V.K.; Lackner, A.A.; Aye, P.P.; Didier, P.; Huang, D.; Shao, L.; et al. Efficacy and Safety of Live Attenuated Persistent and Rapidly Cleared Mycobacterium Tuberculosis Vaccine Candidates in Non-Human Primates. Vaccine 2009, 27, 4709–4717. [Google Scholar] [CrossRef]
- Egen, J.G.; Rothfuchs, A.G.; Feng, C.G.; Horwitz, M.A.; Sher, A.; Germain, R.N. Intravital Imaging Reveals Limited Antigen Presentation and T Cell Effector Function in Mycobacterial Granulomas. Immunity 2011, 34, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Kathamuthu, G.R.; Sridhar, R.; Baskaran, D.; Babu, S. Dominant Expansion of CD4+, CD8+ T and NK Cells Expressing Th1/Tc1/Type 1 Cytokines in Culture-Positive Lymph Node Tuberculosis. PLoS ONE 2022, 17, e0269109. [Google Scholar] [CrossRef] [PubMed]
- Sallin, M.A.; Sakai, S.; Kauffman, K.D.; Young, H.A.; Zhu, J.; Barber, D.L. Th1 Differentiation Drives the Accumulation of Intravascular, Non-Protective CD4 T Cells during Tuberculosis. Cell Rep. 2017, 18, 3091–3104. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, D.M.; Lewinsohn, D.A. New Concepts in Tuberculosis Host Defense. Clin. Chest Med. 2019, 40, 703–719. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Vaccination scheme. Cynomolgus macaques were immunized with electropotered DNA-hsp65 vaccine delivered by three doses at 30-days intervals by in vivo muscular electroporation. Clinical, hematological, biochemical, and immunological profiles were evaluated as represented.
Figure 1.
Vaccination scheme. Cynomolgus macaques were immunized with electropotered DNA-hsp65 vaccine delivered by three doses at 30-days intervals by in vivo muscular electroporation. Clinical, hematological, biochemical, and immunological profiles were evaluated as represented.

Figure 2.
Tuberculin skin test. Cynomolgus macaques were immunized with electropotered DNA-hsp65 vaccine delivered by three doses at 30-day intervals by in vivo muscular electroporation. Reaction to the tuberculin skin test was performed 30 days after the last dose of the vaccine. (A) no reaction (grade zero), (B) bruises (grade 1) and (C) number and frequency of reactional tuberculin skin test.
Figure 2.
Tuberculin skin test. Cynomolgus macaques were immunized with electropotered DNA-hsp65 vaccine delivered by three doses at 30-day intervals by in vivo muscular electroporation. Reaction to the tuberculin skin test was performed 30 days after the last dose of the vaccine. (A) no reaction (grade zero), (B) bruises (grade 1) and (C) number and frequency of reactional tuberculin skin test.
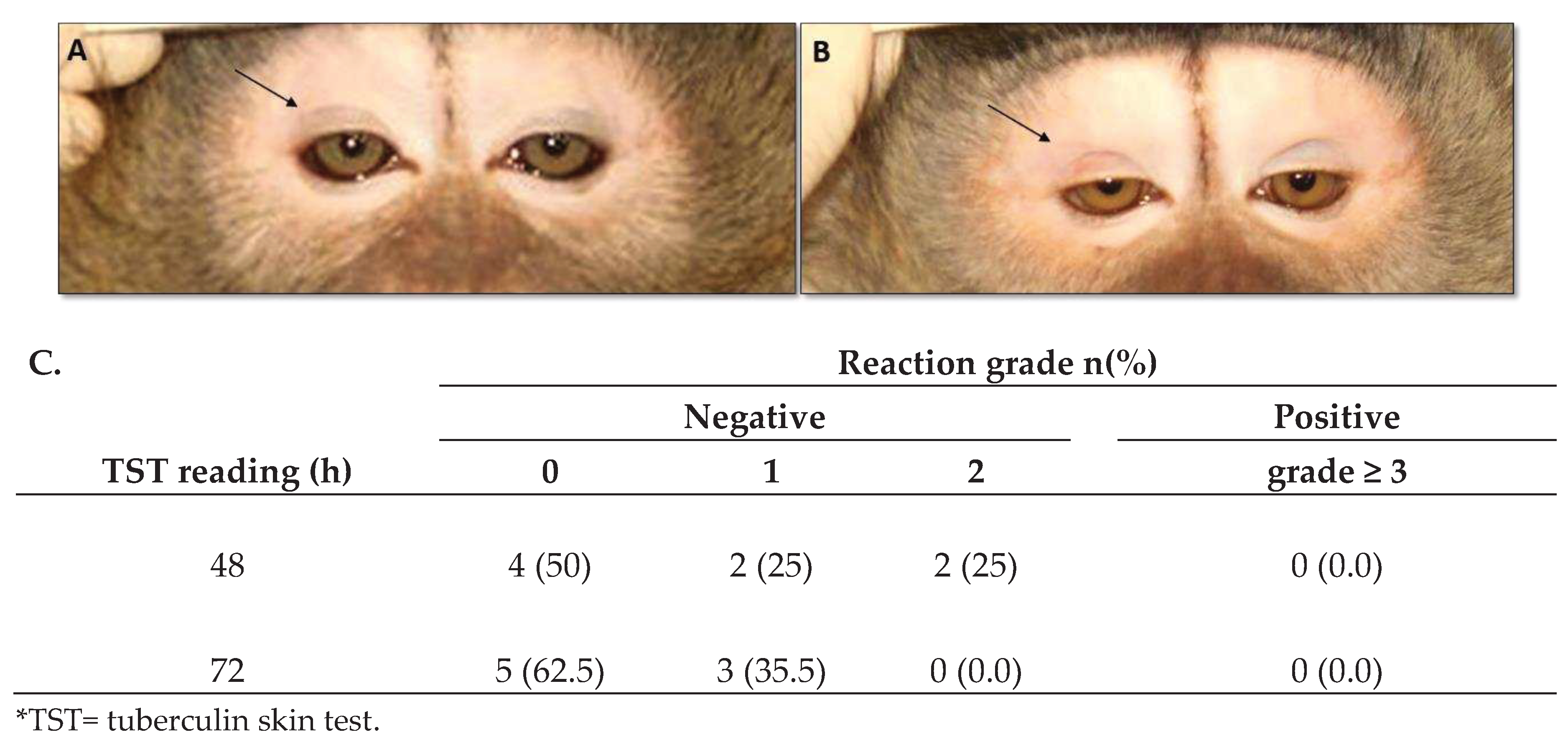
Figure 3.
Frequency of immune cells in the peripheral blood. Frequency of CD4+ and CD8+ T cells; B cells, NK cells; myeloid dendritic and plasmocytoid dendritic cells in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
Figure 3.
Frequency of immune cells in the peripheral blood. Frequency of CD4+ and CD8+ T cells; B cells, NK cells; myeloid dendritic and plasmocytoid dendritic cells in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
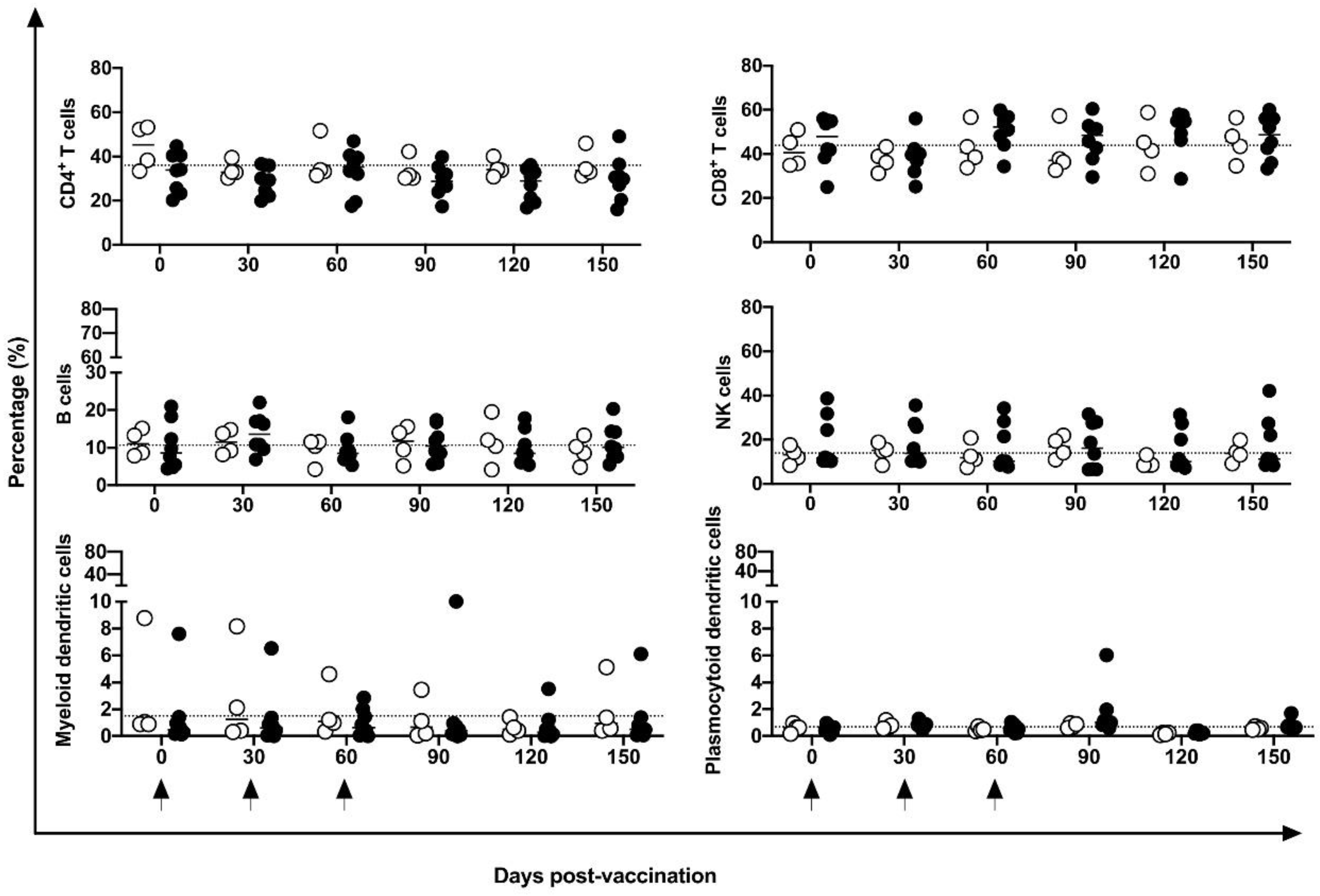
Figure 4.
Frequency of central and effector memory cells in the peripheral blood. Frequency of CD4+ and CD8+ T CM and EF cells in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
Figure 4.
Frequency of central and effector memory cells in the peripheral blood. Frequency of CD4+ and CD8+ T CM and EF cells in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
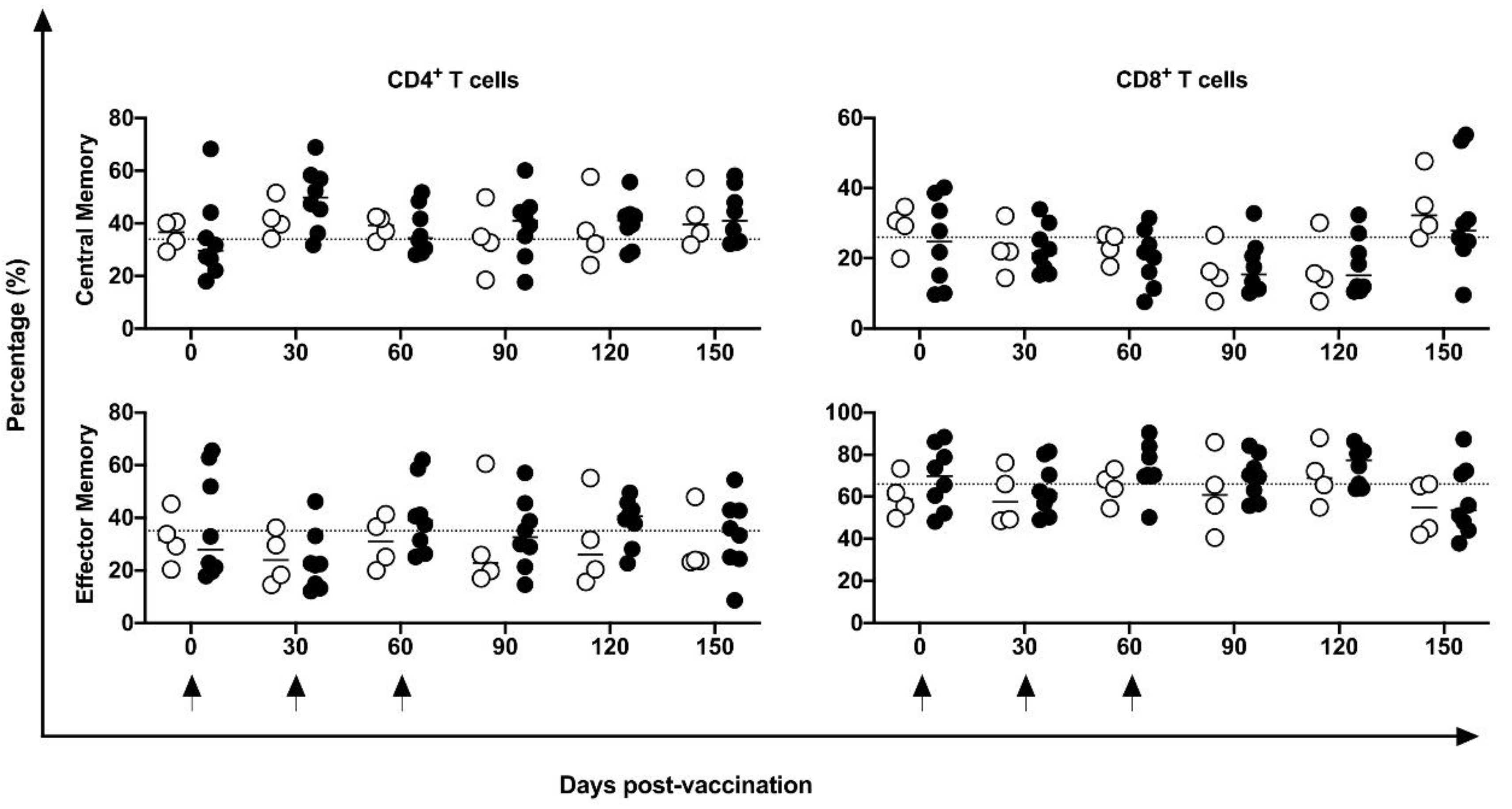
Figure 5.
Frequency of activation marker CD69+ in peripheral blood. Frequency of CD4+/CD69+; CD8+/CD69+ and CD19+/CD69+ cells in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
Figure 5.
Frequency of activation marker CD69+ in peripheral blood. Frequency of CD4+/CD69+; CD8+/CD69+ and CD19+/CD69+ cells in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
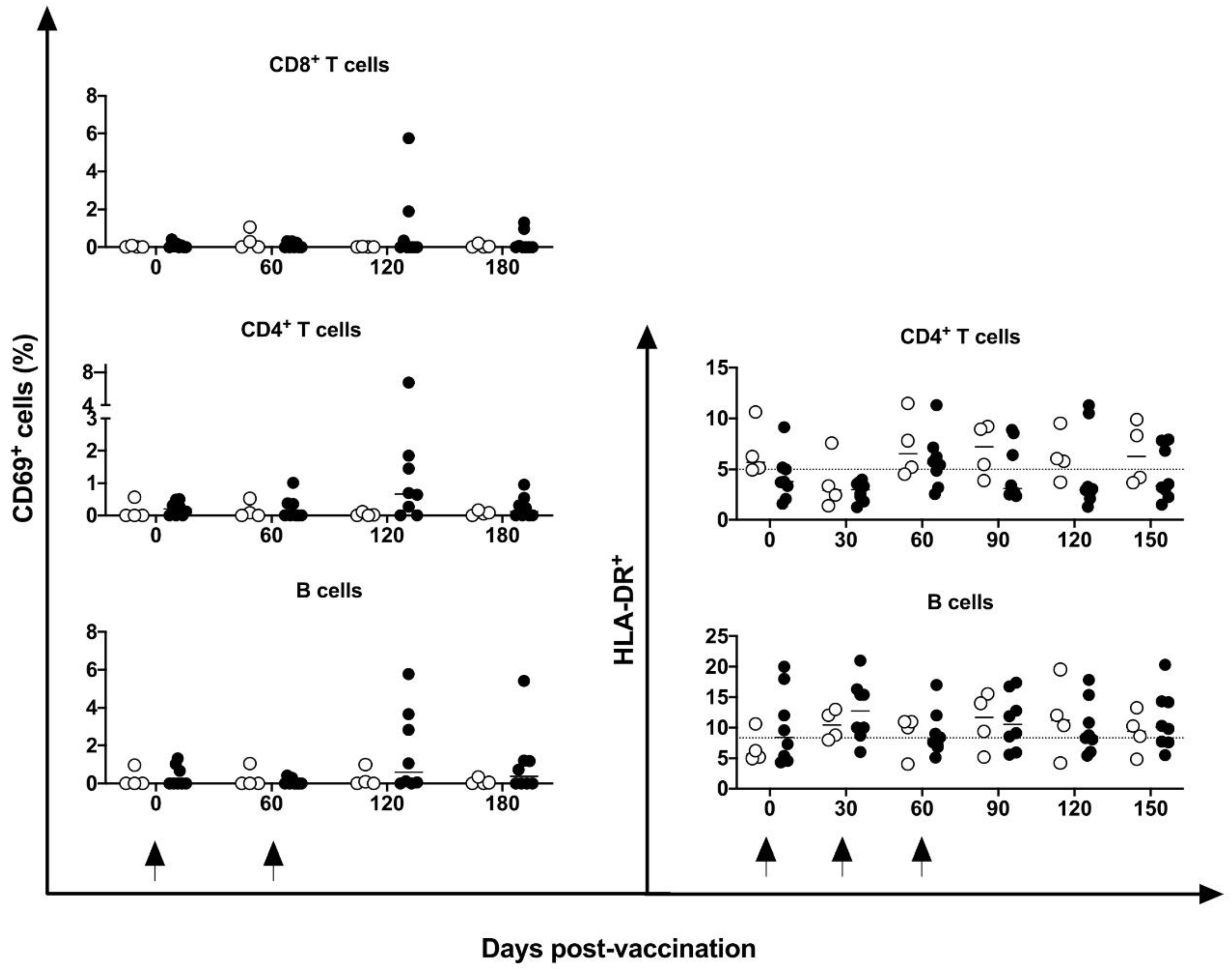
Figure 6.
Frequency of proliferation, lytic and apoptotic markers in peripheral blood. Frequency of CD4+; CD8+ and CD19+ cells expressing Ki67+, BCL2+ and granzyme/perforin markers in unvaccinated (open circles) and electroporated DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
Figure 6.
Frequency of proliferation, lytic and apoptotic markers in peripheral blood. Frequency of CD4+; CD8+ and CD19+ cells expressing Ki67+, BCL2+ and granzyme/perforin markers in unvaccinated (open circles) and electroporated DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
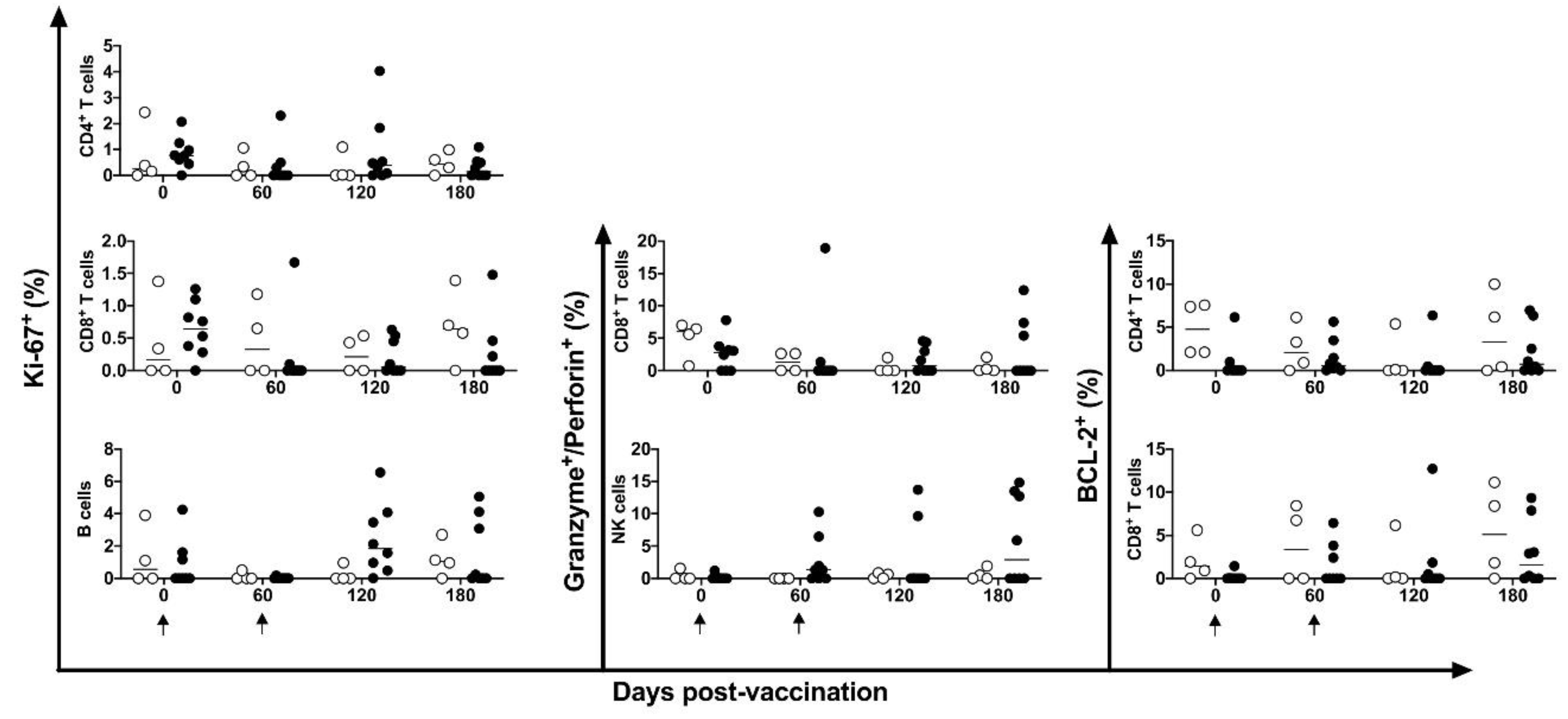
Figure 7.
De novo cytokines expression in immune cells. IFN-γ, TNF-α, IL10 and IL-12 expressing in CD4+ and CD8+ T cells, NK, CD19+ and CD14+ cells were identified after cells stimulus with HSP65 peptide pool in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques . Arrows represent the three doses of vaccine scheme.
Figure 7.
De novo cytokines expression in immune cells. IFN-γ, TNF-α, IL10 and IL-12 expressing in CD4+ and CD8+ T cells, NK, CD19+ and CD14+ cells were identified after cells stimulus with HSP65 peptide pool in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques . Arrows represent the three doses of vaccine scheme.
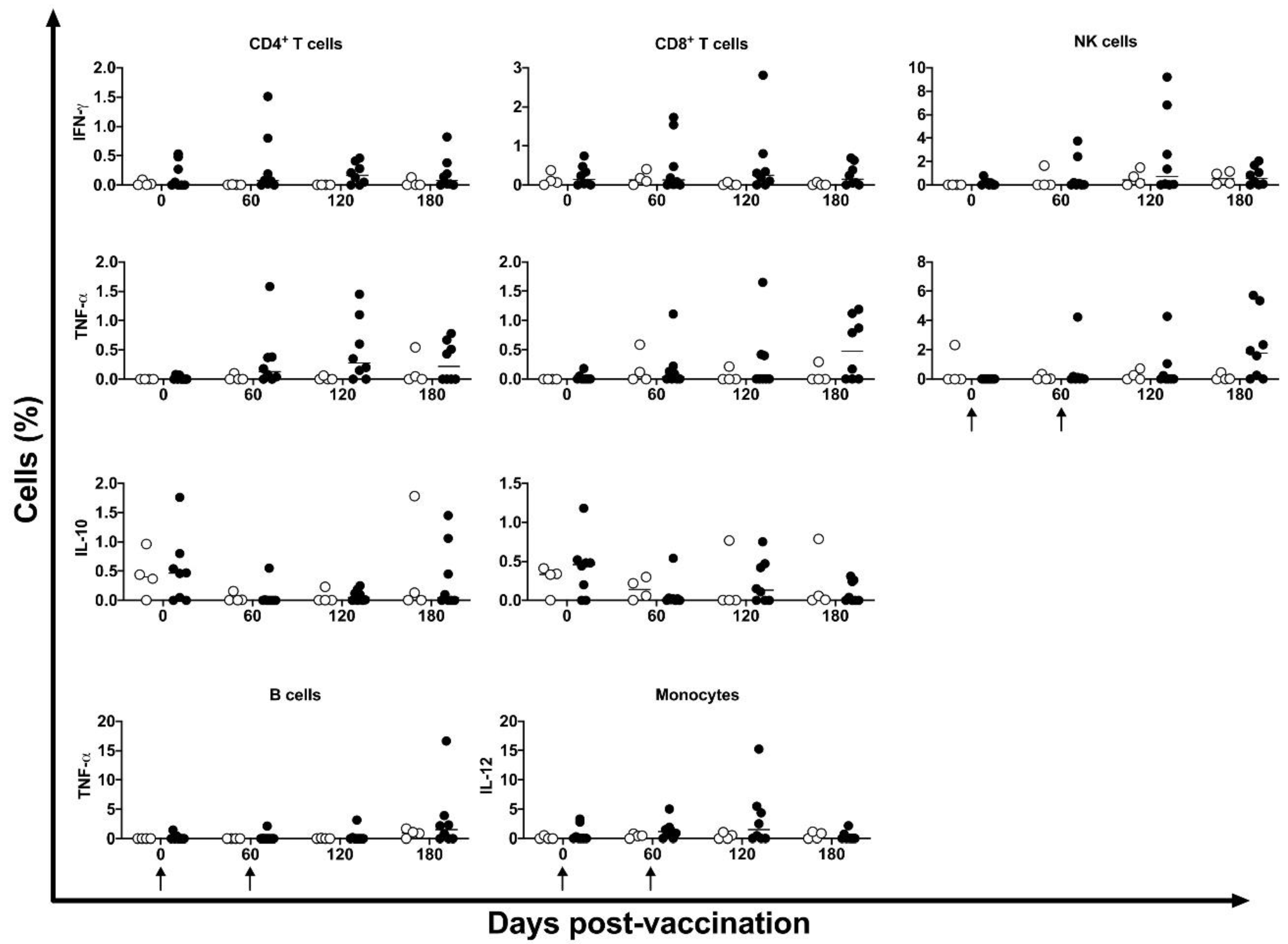
Figure 8.
De novo cytokines quantification in supernatant. TNF-α, IL-6, IL-5, IFN-γ, IL-2 and IL-4 were measured in the supernatant of stimulated cell culture with HSP65 peptides pool by CBA in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
Figure 8.
De novo cytokines quantification in supernatant. TNF-α, IL-6, IL-5, IFN-γ, IL-2 and IL-4 were measured in the supernatant of stimulated cell culture with HSP65 peptides pool by CBA in unvaccinated (open circles) and electropotered DNA-hsp65 vaccinated (black circles) cynomolgus macaques. Arrows represent the three doses of vaccine scheme.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



