
Submitted:
23 October 2023
Posted:
24 October 2023
You are already at the latest version
Abstract
Bacillus subtilis NCD-2 showed a promising biocontrol effect against plant soil-borne diseases, and was developed as a commercial microbial fungicide against cotton verticillium wilt in China. Spores were main ingredient of the fungicide and played the crucial role in biological control of plant diseases. Therefore, the key to reducing the cost of fungicide was to find ways to increase the number of spores of strain NCD-2 during fermentation. In this study, 755 substances were evaluated by Phenotype Microarray technology, and 5 kinds of carbon sources and 1 kind of nitrogen source were found to promote the metabolism of strain NCD-2. Among carbon sources L-arabinose showed strongest ability to promote the bacteria growth and sporulation of strain NCD-2. When L-arabinose was used as a single carbon source, it could increase the bacteria concentration and the sporulation efficiency of strain NCD-2 by 2.04 times and 1.99 times, respectively, compared with D-glucose. Moreover, L-arabinose could significantly decrease the autolysis of strain NCD-2 revealed by microscopy observation. The mechanism for promoting the sporulation by L-arabinose was analysed by RNA-seq sequencing. Results showed that total of 790, 923 and 1270 genes were up-regulated and 639, 850 and 1001 genes were down-regulated under L-arabinose supplies when cultured at 8 h, 12 h and 16 h, respectively. Fourteen differentially expressed genes associated with arabinose transportation and sporulation were selected for qRT-PCR analysis, and the result showed basically consistent expression trend with transcriptome. Notably, genes associated with arabinose metabolism, sporulation, spore resistance to heat and spore coat formation were significantly up-regulated, and genes associated with sporulation-delaying protein were significantly down-regulated under L-arabinose supplies. msmX gene which was involved in arabinose transport in Bacillus genus was deleted, and the mutant decreased the growth and sporulation by 53.71% and 86.46%, respectively, when compared to strain NCD-2 wild type. Complementary of the mutant by importing intact msmX gene could restore the growth and sporulation of the mutant strain. In conclusion, arabinose played an important role in regulating the growth and sporulation of strain NCD-2.
Keywords:
Phenotype Microarrays
; Bacillus subtilis
; L-arabinose
; sporulation
; transcriptome
1. Introduction
Plant soil-borne diseases, such as verticillium wilt and fusarium wilt caused serious loss in plant production, were very difficult to control. Microbial fungicide using living microorganisms as active compound were confirmed as effective and environmental friendly methods to suppress plant soil-borne diseases and reduce the amount of chemical fungicides [1,2]. The control capabilities of biocontrol agents were influenced by the concentration of biocontrol agent in plant rhizosphere. Therefore, it was important to increase the application dose of biocontrol agent.
B. subtilis was an important resource for development of microbial fungicide due to its abilities to produce a variety of antibiotics and form highly resistance spores [3]. Wettable powders and dry powder seed-coating agents were the main formulations of microbial fungicides for suppressing plant soil-borne diseases. However, the processing of the two formulations involved of instantaneous high-temperature as high as 170℃ to dry the bacteria, under such high temperatures only the spores could survive. Therefore, microbial fermentation should consider the bacteria concentration as well as the sporulation [4]. Ideal fermentation system was firstly to increase the fermentation level of the bacterium, and subsequently to make the maximum possible conversion of the bacteria into spores through nutrient regulation and other methods [5].
The carbon and nitrogen sources in the medium were the main factors affecting bacterial growth and sporulation [6,7,8]. Suitable and sufficient carbon and nitrogen sources could promote the growth of the bacterium, but sporulation generally occurred in unfavorable environments such as nutrient starvation [9]. During the stable phase of bacterial growth, residual carbon and nitrogen sources in the medium might inhibit the sporulation of Bacillus. Spp [10]. In microbes, carbohydrates were catabolized into pyruvate, which entered the tricarboxylic acid cycle, mainly through the glycolytic pathway (EMP) and pentose phosphate pathway (PPP). By comparing the effect of the sugars existed in pathway of EMP and PPP on sporulation of B. subtilis, it was found that the sugars existed in PPP pathway but not in EMP pathway could increase the sporulation of Bacillus. So it was concluded that PPP was an important carbohydrates catabolic pathway that affected sporulation [11]. In addition, metal ions such as Ca2+, Mg2+, Zn2+ and Mn2+ also affected the sporulation of B. subtilis, and addition of appropriate concentrations of metal ions in the medium could promote the sporulation [12,13,14]. Different strains had different nutrient requirements suitable for their growth and sporulation, therefore, specific nutrients needed to be explored for a specific strain.
Bacillus subtilis NCD-2 showed a promising biocontrol effect against plant soil-borne diseases, and was developed as a commercial microbial fungicide against cotton verticillium wilt in China. [15,16,17]. The objectives of this study were to screen the nutrients suitable for the growth and sporulation of strain NCD-2, and then explore the mechanism for regulating sporulation by the nutrients. Results of this study will provide important information for large-scale and efficient fermentation of strain NCD-2.
2. Results
2.1. Screening of nutrients that facilitate the metabolism of strain NCD-2
The metabolic activities of strain NCD-2 for carbon, nitrogen, phosphorus, sulfur and trace elements were determined by BIOLOG's Phenotype Microarrays (PMs) technology (Figure S1). For carbon source utilization, strain NCD-2 had higher metabolic activity in L-arabinose, D-arabinose, D-xylose, D-ribose and D-glucosamine supplies, respectively. For nitrogen source utilization, strain NCD-2 had higher metabolic activity in cysteine supply. For phosphorus and sulfur sources utilization, strain NCD-2 had low metabolic activities in 59 phosphorus sources and 35 sulfur sources.
2.2. Effects of L-arabinose, D-ribose, and D-xylose on growth and sporulation efficiency of strain NCD-2
The effects of different carbohydrates on the growth of strain NCD-2 were evaluated (Figure 1a). When D-glucose was used as carbon source, the bacteria concentration of strain NCD-2 reached the maximum of 2.55×107 CFU/mL, while that of L-arabinose was 5.20×107 CFU/mL, which was 2.04 times higher than that of D-glucose. When D-ribose and D-xylose were used as the sole carbon source, the bacteria concentration of strain NCD-2 reached the maximum of 3.90×107 CFU/mL and 3.10×107 CFU/mL, respectively. It was revealed that L-arabinose was the most suitable carbon source to promote the growth of strain NCD-2.
The effects of different carbohydrates on the sporulation of strain NCD-2 were evaluated (Figure 1b). Using D-glucose, L-arabinose, D-ribose and D-xylose as the sole carbon source, the sporulation efficiencies of strain NCD-2 were 2.33%, 27.12%, 11.40%, and 15.39% after 24 h of inoculation, respectively. The sporulation efficiencies of strain NCD-2 were 41.35%, 82.18%, 20.26% and 20.45% after 48 h inoculation, when using D-glucose, L-arabinose, D-ribose and D-xylose as carbon sources, respectively. The sporulation efficiencies of strain NCD-2 increased from 24 h to 48 h inoculation. In addition, the increase of sporulation by L-arabinose was also confirmed by microscopic observation (Figure S2). The results indicated that the L-arabinose could increase the sporulation of strain NCD-2 during the early and later growth stages, but the D-ribose and D-xylose only increased the sporulation of strain NCD-2 at early growth stage, when compared with D-glucose.
2.3. Different proportions of L-arabinose and D-glucose on sporulation of strain NCD-2
The effects of different proportions of L-arabinose and D-glucose on the sporulation of strain NCD-2 were evaluated (Figure 2). Forty-eight hours after inoculation, the sporulation efficiencies of strain NCD-2 were 41.35% and 82.43% when using D-glucose and L-arabinose as sole carbon source, respectively. The sporulation efficiencies of strain NCD-2 gradually decreased with the decrease of L-arabinose proportions in the mixture of L-arabinose and D-glucose. The sporulation efficiencies of strain NCD-2 were 68.82%, 56.62%, and 32.79% when L-arabinose and D-glucose at proportions of 2∶1, 1∶1, and 1∶2, respectively.
2.4. Transcriptome analysis
The effects of L-arabinose and D-glucose on the gene expression of strain NCD-2 were compared by transcriptome sequencing. Compared with the D-glucose total of 1483 differential expression genes (DEGs) including 790 up-regulated genes and 693 down-regulated genes, were identified from L-arabinose treatment 8 h (Ara-8 h) after inoculation (Figure 3a). Total 1773 DEGs including 923 up-regulated genes and 850 down-regulated genes, were identified from L-arabinose treatment 12 h (Ara-12 h) after inoculation (Figure 3b). Total 2271 DEGs including 1270 up-regulated genes and 1001 down-regulated genes, were identified from L-arabinose treatment 12 h (Ara-16 h) after inoculation (Figure 3c). GO annotations revealed that the DEGs associated with sporulation (GO:0043934), spore wall (GO:0031160), endospore-forming forespore (GO:0042601) and asexual sporulation (GO:0030436) were significantly up-regulated at 8 h and 12 h after inoculation. However, only the DEGs associated with spore germination (GO:0009847) were enriched at 16 h after inoculation (Figure 4). KEGG enrichment analysis found that ABC transporters (map02010), polyketide sugar unit biosynthesis (map00523) and ribosome (map03010) were significantly enriched at 8 h and 12 h (Figure 5).
2.5. Confirmation of transcriptional results by qRT-PCR
To verify the results of transcriptome, 14 DEGs associated with arabinose transportation, sporulation, spore resistance to heat, etc. were selected to analyze the expression in L-arabinose treatment by qRT-PCR. Results revealed that all the 14 genes showed basically consistent expression trend with transcriptome, It was indicated that the transcriptome results were reliable and could be used for further experimental analysis (Table S2).
2.6. Analysis of genes associated with sporulation in strain NCD-2
The Venn plot showed that there were 717 genes significantly differentially expressed at all three time points (Figure S3), and most of the up-regulated genes involved in sporulation (Table 1). Among them sigK was a transcriptional regulator encoding spore formation. cotE, cotF, cotG, cotS, cotT, cotV, cotW, cotX, and yheD were responsible for spore coat proteins encoding. dpaA and dpaB were responsible for synthetases encoding pyridine dicarboxylic acid (DPA), a substance within the spore core. gerBA, gerE, gerQ, and gerT were responsible for encoding spore germination proteins. spoIIIAH and spoIIQ encoded polymeric complexes that connected forespore and mother cell [18]; spoIVB and spoIVFB encoded the activating proteins of protease and Sig-K, which catalyzed the formation of the spore cortex, respectively. sspA, sspB, sspD, and sspE encoding small acid-soluble proteins (SASPs) associated with spore resistance to heat. sdpC, encoded a cannibalism factor that delayed the sporulation in B. subtilis, were significantly down-regulated at all three time points.
2.7. Deletion of the msmX gene decreased the sporulation efficiency in strain NCD-2
To confirm that L-arabinose influence sporulation, the msmX gene, which encoded the ATPase, responsible for arabinose uptake, was deleted from strain NCD-2 (ΔmsmX). Additionally, the msmX complementary strain was developed for the ΔmsmX mutant (CPmsmX). The growth and sporulation efficiencies of the WT, ΔmsmX and CPmsmX strains were compared in M9 medium with L-arabinose as carbon source (Figure 6a). Results showed that the bacteria concentrations of WT were 4.60×107 CFU/mL, 5.25×107 CFU/mL and 4.35×107 CFU/mL at 24, 36 and 48 h after inoculation, respectively. Comparatively, strain ΔmsmX decreased the growth, and the bacteria concentrations were 1.18×107 CFU/mL, 2.25×107 CFU/mL and 2.43×107 CFU/mL after 24 h, 36 h and 48 h of inoculation, respectively. However, the complementary strain (CPmsmX) restored the growth, and the bacteria concentrations were 3.53×107 CFU/mL, 6.20×107 CFU/mL and 5.45×107 CFU/mL after 24 h, 36 h and 48 h of inoculation, respectively.
The sporulation efficiencies of strain NCD-2 wild type (WT), msmX-null mutant (ΔmsmX) and its complemented strain (CPmsmX) were also compared in M9 medium with L-arabinose as carbon source (Fingure 6b). The Results showed that the sporulation efficiencies of strain WT were 27.12%, 70.95% and 82.18%, at 24, 36 and 48 h after inoculation, respectively. Comparatively, strain ΔmsmX decreased the sporulation efficiencies by 7.17%, 11.67% and 11.13%, at 24, 36 and 48 h after inoculation, respectively. However, the complementary strain (CPmsmX) restored the sporulation efficiencies by 44.63% and 65.44%, at 36 h and 48 h after inoculation, respectively. The regulation of growth and sporulation of strain NCD-2 by msmX were also confirmed by microscopic observation (Figure S4).
3. Discussion
Spores of Bacillus species with strong resistant to stresses were the key ingredient in the formulation of microbial fungicide. Generally speaking, the control effect of plant soilborne diseases was positively correlated with the population of bacteria colonized in plant rhizosphere [19]. To obtain ideal biocontrol effect, it is expected to increase the amount of spore applied to the soil as much as possible. Therefore, promoting the yield of spores during fermentation process of Bacillus is definitely important for reducing the cost as well as ensuring the wide application of microfungicide. The key factor to reduce the cost of bio-fungicides was how to promote the growth of bacteria and yield of spores during fermentation process of Bacillus. It was known that the sporulation process and final spore yield depended on carbohydrate and amino acid [20]. The combined effects of yeast extract, peptone and glucose enhanced the spore yield of B. megaterium [21]. Likewise, the addition of glucose and ribose in the sporulation medium increased the spore yield of B. subtilis and B. cereus [6,11,22]. In this study, we focused on nutrients that promoted strain NCD-2 growth and spore formation, which required screening for a large number of nutrients due to different strains had different nutrient requirements. Phenotype Microarrays were commercially available microplate assays that could be used to test more than 1,000 phenotypic traits simultaneously by recording the microorganism’s respiration over time on many distinct substrates [23,24]. Therefore, PMs could be used to screen nutrients suitable for the catabolism of a specific organism quickly and in high throughput, which had the advantages of large amount of information and time saving [25]. In this way, the catabolic capability of strain NCD-2 to approximately 200 carbon sources, 400 nitrogen sources, and 100 phosphorous and sulfur sources were determined by PMs. Results showed that strain NCD-2 had higher metabolic capacity to L-arabinose, D-xylose and D-ribose as sole carbon sources, among which L-arabinose could significantly increase the bacteria concentration and sporulation efficiency of strain NCD-2.
B. subtilis was able to grow on medium with L-arabinose as the sole carbon and energy source. In the absence of L-arabinose, AraR protein bounded to a site within the araABDLMNPQ-abfA operon promoter region preventing transcription. In the presence of L-arabinose a conformational change was induced in AraR such that recognition and binding to DNA was no longer possible, and the operon could be expressed [26]. After entering the cell, L-arabinose was sequentially converted to L-ribulose, L-ribulose 5-phosphate, and D-xylulose 5-phosphate by the action of L-arabinose isomerase (encoded by araA), L-ribulokinase (encoded by araB), and L-ribulose-5-phosphate 4-epimerase (encoded by araD), respectively. D-xylulose 5-phosphate was further catabolized through the pentose phosphate pathway [27,28]. Transcriptome analysis of this study showed that L-arabinose strongly up-regulated araABDLMNPQ-abfA operon expression in strain NCD-2 (Accession number: SUB12858722), and genes associated with sporulation were also strongly up-regulated (Table 1). Moreover, compared with L-arabinose as the sole carbon source, the sporulation efficiency of strain NCD-2 significantly decreased when both glucose and arabinose present in M9 medium (Figure 2), which might be attributed to the fact that the presence of glucose repressed the expression of both araE, a gene for L-arabinose transporter, and ara operon at the transcriptional level [29,30]. Thus, L-arabinose might be involved in regulating the expression of genes related to sporulation in strain NCD-2 by regulating ara operon.
Previous studies found that the AraNPQ-MsmX system was involved in the transport of arabinans, and knocking out araNPQ reduced the growth rate of B. subtilis [26,31]. Therefore, the deletion of the msmX gene, which encoded ATPase for providing energy to the AraNPQ transporter, inevitably led to a decrease in the growth rate of B. subtilis. In this study, msmX was deleted from strain NCD-2 wild type, and the mutant did reduce the bacteria growth and sporulation efficiency of strain NCD-2 with L-arabinose as the sole carbon source (Figure 6), which was consistent with the previously study, but not with D-glucose as the sole carbon source (Data not shown). It was clear that L-arabinose somehow entered the metabolic process of strain NCD-2 via the AraNPQ-MsmX system, then affected the growth and sporulation. The results would provide knowledge for effectively improving the growth and spore production during the fermentation of strain NCD-2.
The cell of B. subtilis might autolysed during fermentation, so resulting in a large number of cell deaths and reducing bacterial fermentation concentration [32]. There were many factors that led to autolysis of the bacterium [33,34,35,36]. Among them, a phenomenon of "cannibalism" was described [37,38], in which the master regulator of sporulation Spo0A was active, and released two toxins Skf and SdpC to kill Spo0A inactive sister cells. The nutrients released by the dead cells were used for the growth of cells that were not yet fully committed to sporulate. In this study, it was observed that cell autolysis produced a large amount of cell debris in the medium with D-glucose as sole carbon resource, but not in the medium with L-arabinose. In transcriptome, L-arabinose was significantly down-regulated the transcription of sdpC, compared with D-glucose at 8 h, 12 h and 16 h after inoculation. It was suggested that L-arabinose increasing the bacteria concentration of strain NCD-2 was due to L-arabinose inhibiting the process of "cannibalism" in strain NCD-2.
4. Materials and Methods
4.1. Bacterial strains and growth conditions
The strains used in this study were listed in Table 2, B. subtilis strains were stored at -80℃ in LB medium containing 30% glycerol. Strains were cultured at 30℃ without special instructions and the appropriate concentration of antibiotics was added to the medium as needed.
4.2. Phenotype microarrays analysis
The metabolic phenotype of B. subtilis strain NCD-2 on 755 nutrients was evaluated using the Biolog Phenotype MicroArrays system (Biolog, Hayward, CA, USA). In ninety-six-well PM1-8 microplates, including carbon source (PM1, PM2A), nitrogen source (PM3B, PM6-8), phosphorus source and sulfur source (PM5), were assayed , and the names of nutrients were described in the literature of Bochner et al [23]. The experiment was conducted according to the procedures developed by the manufacturer [25]. Briefly, strain NCD-2 was firstly scribed on BUG+B plates and cultured overnight at 33℃, single colony was selected and inoculated on BUG+B plates again, after cultured overnight at 33℃, Bacteria were dipped into the BUG+B medium with sterile cotton swabs and transferred to 20 mL of 1×IF-0a inoculum, stirred well to obtain a suspension of the bacteria, and the bacteria were continuously added until the turbidity of the suspension reached 81% T (T is the standard concentration unit of Biolog), and the suspension were added to the PM1-8 MicroPlatesTM according to the instructions. The OmniLog software was set up and loaded, and the PM MicroPlatesTM were incubated in the OmniLog incubator at 30℃ for 48 h. The plates were scanned every 15 min, and the results were analyzed and plotted using the OmniLog software at the end of the incubation.
4.3. Determination of cell concentration and sporulation efficiency
A preculture was prepared by incubating a colony of strain NCD-2 in 100 ml Luria-Bertani (LB) broth at 30℃ with agitation (180 rpm) for 12 h, centrifuged to collect the cells, and then resuspended with sterile water and adjusted to OD600=1.0 with sterile water. The bacteria suspension was added to the M9 medium (12.8 g·L-1, Na2HPO4·7H2O, 3 g·L-1 KH2PO4, 0.5 g·L-1 NaCl, 1 g·L-1 NH4Cl, 0.24 g·L-1 MgSO4, 0.011 g·L-1 CaCl2, 4 g·L-1 D-glucose) at 1% inoculation volume. To evaluate the effect of different carbohydrates on sporulation of strain NCD-2, the D-glucose was replaced with same concentration of L-arabinose, D-xylose and D-ribose, respectively. Samples were harvested at 24, 36 and 48 h after inoculation, respectively, to calculate sporulation effeciency according to previous protocol [39].
4.4. RNA extraction and RNA sequencing
Strain NCD-2 was inoculated in M9 medium with D-glucose or L-arabinose as sole carbon source, respectively, strain NCD-2 was cultured at 30℃, 180 rpm, then the cells were collected at at 8 h, 12 h and 16 h after inoculation by centrifugation at 4℃ with 10,000 rpm for 5 min. The bacterium was rapidly frozen with liquid nitrogen and stored at -80℃, three replicates were set for each treatment. The total RNA of the collected bacteria was extracted according to the instructions of the RNAprep Pure Cell/Bactria Kit (TianGen Biotech, Beijing, China), and the quality and concentration of total RNA were measured with the NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA). The cDNA library construction and RNA sequencing (RNA-seq) were performed by Illumina platform at Majorbio Co., Ltd. (Shanghai, China).
4.5. Transcriptome data and differential gene expression analysis
The transcriptome raw data has been uploaded to NCBI-SRA database (Accession number: SUB12858722). Using fastp (https://github.com/OpenGene/fastp) to remove low-quality reads and adapters from the data. Then, these clean reads were mapped to reference genome (Bacillus subtilis NCD-2) using Bowtie (http://bowtie-bio.sourceforge.net/index.shtml). The screening criteria for differentially expressed genes (DEGs) were |log2(Fold Change)| > 1 and adjusted p-value < 0.05. DEGs were used for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis using Goatools (https://github.com/tanghaibao/goatools) and the R package ClusterProfile, respectively, and enrichment results were filtered with the parameters of p-value < 0.05.
4.6. Confirmation of transcriptome analysis results
To validate the transcriptome results, 14 genes (Supplementary Table S2) were selected for expression analysis by qRT-PCR. Primers were designed using primer premier 5.0 software (Applied Biosystems). B. subtilis strain NCD-2 were cultured in M9 medium with L-arabinose or D-glucose as the sole carbon source, and cultured at 30℃ with shaking at 180 rpm, respectively. Strain NCD-2 was collected by centrifugation after 8 h, 12 h and 16 h of incubation, respectively. Total RNA was extracted as described above, and adjusted to 50 ng·μL-1. The extracted total RNA was used as a template to synthesize the cDNA first strand using TransScript® One-Step gDNA Removal and cDNA Synthesis SuperMix (TransGen Biotech, Beijing, China), and the cDNA concentration was then adjusted to 100 ng·μL-1 with double distilled water. The qRT-PCR reaction was conducted according to the instructions of TransStart® Top Green qPCR SuperMix (TransGen Biotech, Beijing, China). Three replicates were set up for each gene detection, and the gyrB gene was used as the internal reference gene, and the relative change in target gene expression was calculated by the formula 2-ΔΔCt [40].
4.7. Function analysis of msmX gene
To delete the msmX gene from strain NCD-2, msmX upstream fragments were amplified using primers msmX-P1 (CGAGCTCTTTCAGCGGTTCGGGTG) and msmX-P2 (GGGGTACCGATCAAAAAAACCGGACATGGGG), msmX downstream fragments were amplified using primers msmX-P3 (GGGGTACCACCCAGCCATCTAACATCCCCC) and msmX-P4 (GCTCTAGATCCCGGTTCGATTGTGTCTG). The upstream and downstream amplification fragments were digested with Kpn I restriction enzyme respectively, and then the two fragments were ligated with T4 DNA ligase. Using the ligation product as a template, PCR amplified with msmX-P1 and msmX-P4 primers. The amplificon was digested with Sac I and Xba I restriction enzymes and then attached to the corresponding digestion site of the pKSV7 plasmid [41]. The recombinant plasmid was transformed into strain NCD-2 via electroporation, and the knockout of msmX was conducted by in-frame deletion, described as Arnaud et al [42]. The deletion of msmX (ΔmsmX) was confirmed by PCR and sequencing with primers msmX-P1 and msmX-P4. To complementation of the ΔmsmX mutant, intact msmX was amplified from strain NCD-2 with primers CPmsmX-F (GGGGTACCTTATCGAATTCTCATTTCTG) and CPmsmX-R (GCAGGTCGACATTGGAAATATGCACGAAAA), which included the Kpn I and Sal I restriction sites, respectively. The amplicon was digested with Kpn I and Sal I and inserted into pHY300PLK, which is a shuttle vector for E. coli and B. subtilis [43]. The recombinant plasmid was transformed into mutant strain ΔmsmX by electroporation to obtain complementary strain (CPmsmX). The wild type strain NCD-2 and mutants were cultured in M9 medium with L-arabinose as sole carbon source, after which the bacteria concentration and sporulation efficiencies were calculated by plate counting as described above.
4.8. Statistical analyses
Statistically significant differences (P<0.05) in NCD-2 CFU, sporulation efficiency, as well as the CFU and sporulation efficiency between wild-type and mutant strains were evaluated by ANOVA using SPSS 18.0 software (SPSS, Chicago, IL, USA) followed by Tukey’s post hoc test. Figures were prepared with Origin Pro 8.6 software (OriginLab Corporation, Hampton, MA, USA).
5. Conclusion
L-arabinose can significantly increase the bacteria concentration and sporulation efficiency of strain NCD-2. In this study, we used PMs technology to screen out several nutrients with high metabolic activity of strain NCD-2, among which L-arabinose can significantly increase the bacteria concentration and sporulation efficiency of strain NCD-2, and repress cells autolysis. Transcriptome results showed that L-arabinose up-regulated the expression of sporulation related genes and down-regulated the expression of cannibalism related genes. After knocking out msmX, which is responsible for transporting arabinose, significantly decreased the bacteria concentration and sporulation efficiency of strain NCD-2 in the medium with L-arabinose as the carbon source. Results will provide assistance in the study of directed fermentation and the mechanism of regulating sporulation of strain NCD-2.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Author Contributions: Conceptualization, Guo Qinggang and Ma Ping; Data curation, Y.F. and X.L. (Xiaomeng Liu); Formal analysis, Z.S.; Funding acquisition, Q.G. and P.M.; Investigation, Y.F.; Methodology, Y.F. and X.L. (Xiaomeng Liu); Software, Z.S. and Y.F.; Supervision, Q.G. and P.M.; Validation, Y.F. and X.L. (Xiaomeng Liu); Visualization, Y.F and Z.S.; Writing—original draft, Y.F.; Writing—review & editing, Z.S., P.W., Q.G. and P.M. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
This work was supported by the Chinese National Natural Science Foundation (32172487), the China Agriculture Research System of MOF and MARA (CARS-15-19), the Natural Science Foundation of Hebei Province (C2021301030), and the HAAFS Science and Technology Innovation Special Project (2022KJCXZX-ZBS-1).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fiers, M.; Edel-Hermann, V.; Chatot, C.; Hingrat, Y.L.; Steinberg, A.C. Potato soil-borne diseases. A review. Agro. Sustain. Dev. 2012, 32, 93-132. [CrossRef]
- Bisutti, I.L.; Pelz, J.; Büttner, C.; Stephan, D. Field assessment on the influence of RhizoVital® 42 fl. and Trichostar® on strawberries in the presence of soil-borne diseases. Crop Prot. 2017, 96, 195-203. [CrossRef]
- Pérez-García, A.; Romero, D.; de Vicente, A. Plant protection and growth stimulation by microorganisms: biotechnological applications of Bacilli in agriculture. Curr. Opin. Biotechnol. 2011, 22 (2), 187-193. [CrossRef]
- Rao, Y.K.; Tsay, K.J.; Wu, W.S.; Tzeng, Y.M. Medium optimization of carbon and nitrogen sources for the production of spores from Bacillus amyloliquefaciens B128 using response surface methodology. Process Biochem. 2007, 42 (4), 535-541. [CrossRef]
- Posada-Uribe, L.F.; Romero-Tabarez, M.; Villegas-Escobar, V. Effect of medium components and culture conditions in Bacillus subtilis EA-CB0575 spore production. Bioproc. Biosyst. Eng. 2015, 38 (10), 1879-1888. [CrossRef]
- de Vries, Y.; Atmadja, R.; Hornstra, L.; de Vos, W.; Abee, T. Influence of glutamate on growth, sporulation, and spore properties of Bacillus cereus ATCC 14579 in defined medium. App. Environ. Microb. 2005, 71 (6), 3248-3254. [CrossRef]
- Donohue, T.; Bernlohr, R. Effect of cultural conditions on the concentrations of metabolic intermediates during growth and sporulation of Bacillus licheniformis. J. Bacteriol. 1978, 135 (2), 363-372. [CrossRef]
- Singh, R.M. Role of carbon and nitrogen sources in bacterial growth and sporulation. Appl. Microbiol. 1971, 22 (1), 131-132. [CrossRef]
- Lopez, J.M.; Marks, C.L.; Freese, E. The decrease of guanine nucleotides initiates sporulation of Bacillus subtilis. Biochim. Biophys. Acta 1979, 587 (2), 238-252. [CrossRef]
- Monteiro, S.M.S.; Clemente, J.J.; Carrondo, M.J.T.; Cunha, A.E. Enhanced Spore Production of Bacillus subtilis Grown in a Chemically Defined Medium. Adv. Microbiol. 2014, 4 (8), 444-454. [CrossRef]
- Warriner, K.; Waites, W.M. Enhanced sporulation in Bacillus subtilis grown on medium containing glucose:ribose. Lett. Appl. Microbiol. 1999, 29 (2), 97-102. [CrossRef]
- Nguyen Thi Minh, H.; Durand, A.; Loison, P.; Perrier-Cornet, J.; Gervais, P. Effect of sporulation conditions on the resistance of Bacillus subtilis spores to heat and high pressure. Appl. Microbiol. Biotechnol. 2011, 90 (4), 1409-1417. [CrossRef]
- Bressuire-Isoard, C.; Broussolle, V.; Carlin, F. Sporulation environment influences spore properties in Bacillus: evidence and insights on underlying molecular and physiological mechanisms. FEMS Microbiol. Rev. 2018, 42 (5), 614-626. [CrossRef]
- Widderich, N.; Rodrigues, C.D.; Commichau, F.M.; Fischer, K.E.; Ramirez-Guadiana, F.H.; Rudner, D.Z.; Bremer, E. Salt-sensitivity of σ(H) and Spo0A prevents sporulation of Bacillus subtilis at high osmolarity avoiding death during cellular differentiation. Mol. Microbiol. 2016, 100 (1), 108-124. [CrossRef]
- Li, S.Z.; Lu, X.Y.; Ma, P.; Gao, S.G.; Liu, G. Evaluation of biocontrol potential of a bacterial strain NCD-2 against cotton verticillium wilt in field trials. Acta Phytopathol. Sin. 2005, 35, 451-455.
- Guo, Q.; Dong, W.; Li, S.; Lu, X.; Wang, P.; Zhang, X.; Wang, Y.; Ma, P. Fengycin produced by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling damping-off disease. Microbio. Res. 2014, 169, 533-540. [CrossRef]
- Zhao, W.S.; Ban, Y.Y..; Su, Z.H.; Li, S.Z.; Liu, X.Y; Guo, Q.G.; Ma, P. Colonization Ability of Bacillus subtilis NCD-2 in Different Crops and Its Effect on Rhizosphere Microorganisms. Microorganisms 2023, 11 (3), 776. [CrossRef]
- Camp, A.H.; Losick, R. A novel pathway of intercellular signalling in Bacillus subtilis involves a protein with similarity to a component of type III secretion channels. Mol. Microbiol. 2008, 69 (2), 402-417. [CrossRef]
- Dong, L.H.; Wang, P.; Zhao, W.S.; Su, Z.H.; Zhang, X.Y.; Lu, X.Y.; Guo, Q.G. Surfactin and fengycin contribute differentially to the biological activity of Bacillus subtilis NCD-2 against cotton verticillium wilt. Biol. Control 2022, 174, 104999. [CrossRef]
- Schaeffer, P.; Millet, J.; Aubert, J.P. Catabolic repression of bacterial sporulation. Proc. Natl. Acad. Sci. U.S.A. 1965, 54 (3), 704-711. [CrossRef]
- Verma, N.; Singh, N.A.; Kumar, N.; Raghu, H.V. Screening of different media for sporulation of Bacillus megaterium. Int. J. Microbiol. Res. Rev 2013, 1, 68-73.
- Monteiro, S.M.; Clemente, J.J.; Henriques, A.O.; Gomes, R.J.; Carrondo, M.J.; Cunha, A.E. A procedure for high-yield spore production by Bacillus subtilis. Biotechnol. Prog. 2005, 21 (4), 1026-1031. [CrossRef]
- Bochner, B.R; Gadzinski, P.; Panomitros, E. Phenotype microarrays for high-throughput phenotypic testing and assay of gene function. Genome Res. 2001, 11 (7), 1246-1255. [CrossRef]
- Bochner, B.R. New technologies to assess genotype-phenotype relationships. Nat. Rev. Genet. 2003, 4, 309-314. [CrossRef]
- Mackie, A.; Hassan, K.; Paulsen, I.; Tetu, S. Biolog Phenotype Microarrays for phenotypic characterization of microbial cells. Methods Mol. Biol. 2014, 1096, 123-130. [CrossRef]
- Sá-Nogueira, I.; Nogueira, T.V.; Soares, S.; de Lencastre, H. The Bacillus subtilis L-arabinose (ara) operon: nucleotide sequence, genetic organization and expression. Microbiol. (Reading) 1997, 143 Pt 3, 957-969. [CrossRef]
- Lepesant, J.A.; Dedonder, R. Metabolism of L-arabinose in Bacillus subtilis Marburg Ind-168. C. R. Acad. Sci. Ser. D. 1967, 264 (23), 2683-2686.
- Sá-Nogueira, I.; de Lencastre, H. Cloning and characterization of araA, araB, and araD, the structural genes for L-arabinose utilization in Bacillus subtilis. J. Bacteriol. 1989, 171 (7), 4088-4091. [CrossRef]
- Sá-Nogueira, I.; Mota, L.J. Negative regulation of L-arabinose metabolism in Bacillus subtilis: characterization of the araR (araC) gene. J. Bacteriol. 1997, 179 (5), 1598-1608. [CrossRef]
- Sá-Nogueira, I.; Ramos, S.S., Cloning, functional analysis, and transcriptional regulation of the Bacillus subtilis araE gene involved in L-arabinose utilization. J. Bacteriol. 1997, 179 (24), 7705-7711. [CrossRef]
- Ferreira, M.; Sá-Nogueira, I. A multitask ATPase serving different ABC-type sugar importers in Bacillus subtilis. J. Bacteriol. 2010, 192 (20), 5312-5318. [CrossRef]
- Iyer, J.L.; Shetty, P.; Pai, J. Evaluation of whole cells of Bacillus subtilis as substrate for measurement of autolysin activity. Process Biochem. 2005, 40 (5), 1593-1597. [CrossRef]
- Krogh, S.; Jørgensen, S.T.; Devine, K.M. Lysis genes of the Bacillus subtilis defective prophage PBSX. J. Bacteriol. 1998, 180 (8), 2110-2117. [CrossRef]
- Nandy, S.K.; Venkatesh, K.V. Effect of Carbon and Nitrogen on the cannibalistic behavior of Bacillus subtilis. Appl. Biochem. Biotechnol. 2008, 151, 424-432. [CrossRef]
- Sahoo, S.; Rao, K.K.; Suraishkumar, G.K. Reactive oxygen species induced by shear stress mediate cell death in Bacillus subtilis. Biotechnol.Bioeng. 2006, 94 (1), 118-127. [CrossRef]
- Jolliffe, L.K.; Langemeier, S.O.; Doyle, R.J. Hydrogen ion control of autolysin-dependent functions in Bacillus subtilis. Microbios 1983, 38 (153-154), 187-194.
- González-Pastor, J.E.; Hobbs, E.C.; Losick, R. Cannibalism by sporulating bacteria. Science (New York, N.Y.) 2003, 301 (5632), 510-513. [CrossRef]
- González-Pastor, J.E. Cannibalism: a social behavior in sporulating Bacillus subtilis. FEMS Microbiol. Rev. 2011, 35 (3), 415-424. [CrossRef]
- Eswaramoorthy, P.; Guo, T.; Fujita, M. In vivo domain-based functional analysis of the major sporulation sensor kinase, KinA, in Bacillus subtilis. J. Bacteriol. 2009, 191 (17), 5358-5368. [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.) 2001, 25 (4), 402-408. [CrossRef]
- Liu, P.; Xia, L.Q.; Hu, S.B.; Yan, L.; Ding, X.Z; Zhang, Y.M; Yu, Z. Site-specific integration of heterologous gene into Bacillus thuringiensis chromosome and its expression. Acta Microbiol. Sin. 2008, 48 (5), 661-666.
- Arnaud, M.; Chastanet, A.; Débarbouillé, M. New vector for efficient allelic replacement in naturally nontransformable, low-GC-content, gram-positive bacteria. Appl. Environ. Microbiol. 2004, 70 (11), 6887-6891. [CrossRef]
- Guo, Q.; Li, S.; Lu, X.Y.; Li, B.Q.; Ma, P. PhoR/PhoP two component regulatory system affects biocontrol capability of Bacillus subtilis NCD-2. Genet. Mol. Biol. 2010, 33 (2), 333-340. [CrossRef]
Figure 1.
Effects of different carbon sources on the bacteria concentration (a) and sporulation efficiency (b) of strain NCD-2. Columns represent the average of four replicates, error bars show standard deviations, different letters indicate significant (P<0.05) difference according to ANOVA with Tukey’s post hoc test.
Figure 1.
Effects of different carbon sources on the bacteria concentration (a) and sporulation efficiency (b) of strain NCD-2. Columns represent the average of four replicates, error bars show standard deviations, different letters indicate significant (P<0.05) difference according to ANOVA with Tukey’s post hoc test.
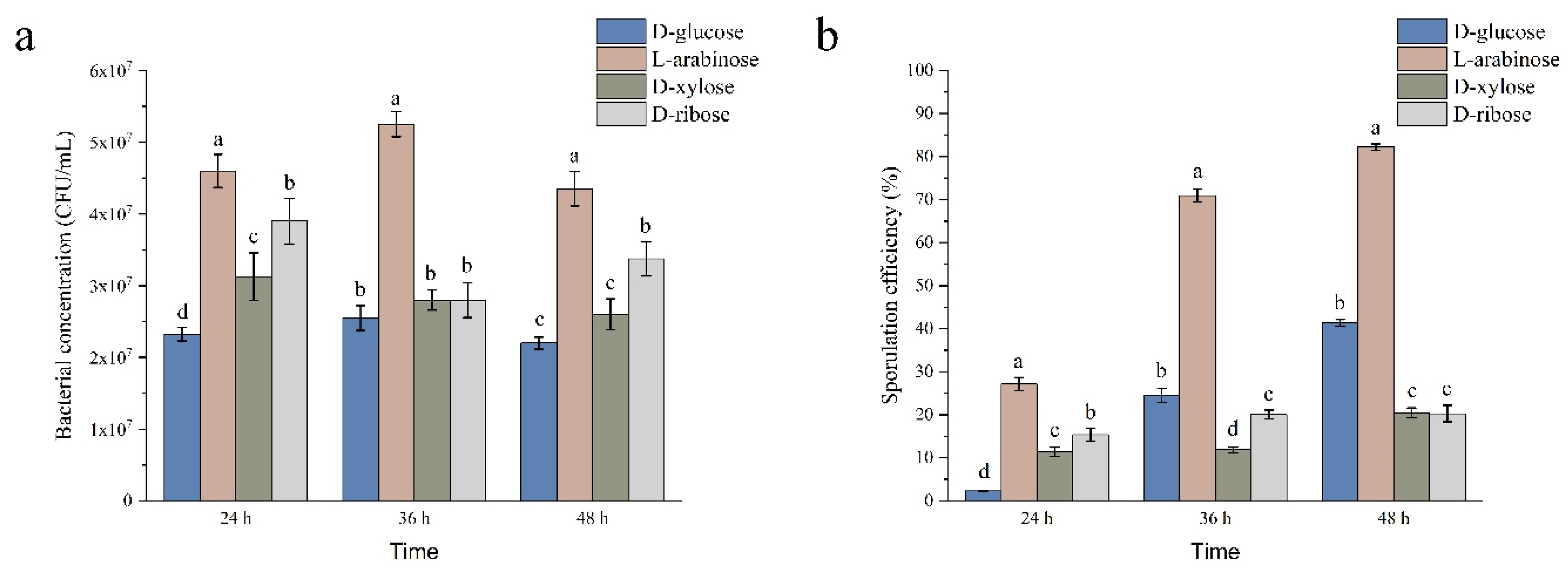
Figure 2.
Different proportions of L-arabinose and D-glucose on the sporulation of strain NCD-2. Strain NCD-2 was cultured in M9 medium containing 4 g·L-1 carbohydrates, and the sporulation efficiency was determined 48 h after inoculation . Columns represent the average of four replicates, error bars show standard deviations, different letters indicate significant (P<0.05) difference according to ANOVA with Tukey’s post hoc test.
Figure 2.
Different proportions of L-arabinose and D-glucose on the sporulation of strain NCD-2. Strain NCD-2 was cultured in M9 medium containing 4 g·L-1 carbohydrates, and the sporulation efficiency was determined 48 h after inoculation . Columns represent the average of four replicates, error bars show standard deviations, different letters indicate significant (P<0.05) difference according to ANOVA with Tukey’s post hoc test.
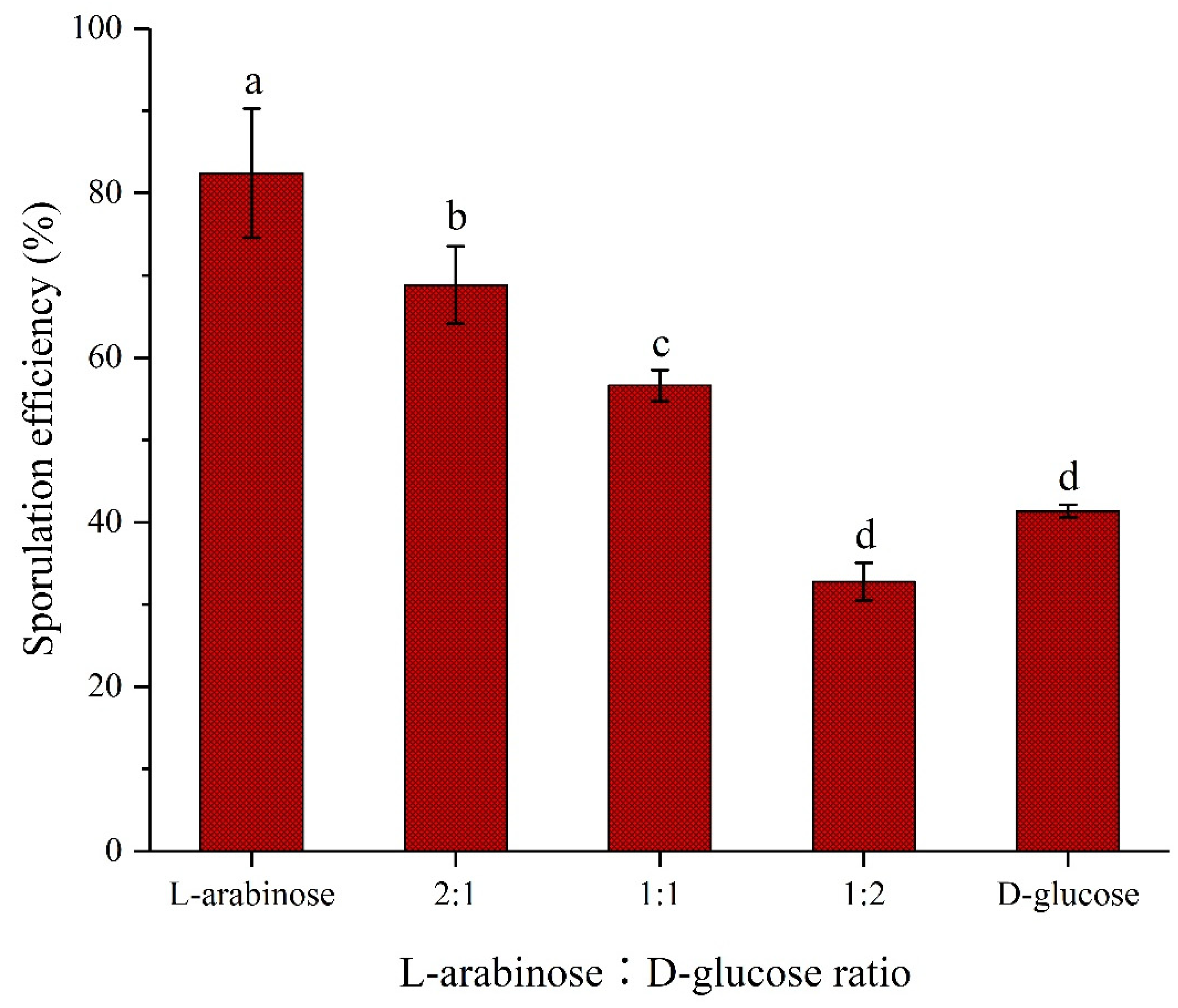
Figure 3.
Volcano plots of transcriptomics comparison of strain NCD-2. X-axis indicates the average of log2 fold change from the replicates. Negative values indicate down-regulation and positive values indicate up-regulation. Y-axis is -log10 padj. Dots in blue or red indicate differentially expressed genes. Dots in black indicate proteins that are not significantly changed in genes expression. (a) 8 hours post inoculation. (b) 12 hours post inoculation. (c) 16 hours post inoculation.
Figure 3.
Volcano plots of transcriptomics comparison of strain NCD-2. X-axis indicates the average of log2 fold change from the replicates. Negative values indicate down-regulation and positive values indicate up-regulation. Y-axis is -log10 padj. Dots in blue or red indicate differentially expressed genes. Dots in black indicate proteins that are not significantly changed in genes expression. (a) 8 hours post inoculation. (b) 12 hours post inoculation. (c) 16 hours post inoculation.
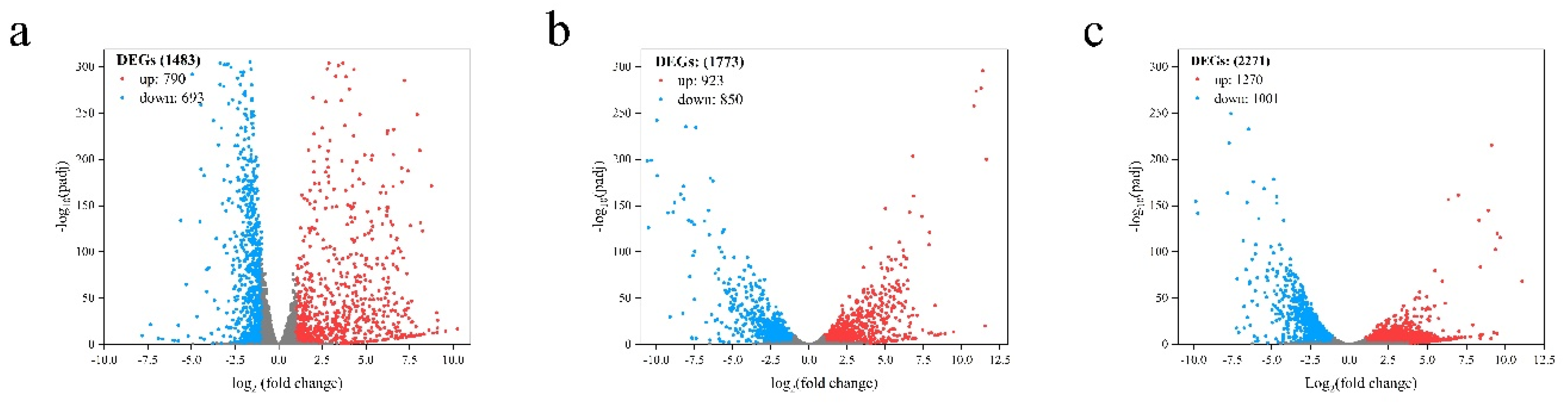
Figure 4.
Go enrichment analysis based on the differential expression genes between L-arabinose and D-glucose-cultured strain NCD-2. The X-axis indicates the number of genes classified into regulatory or functional categories, as depicted on the Y-axis. Columns in blue indicate down-regulation and in red indicate up-regulation. (a) 8 hours post inoculation. (b) 12 hours post inoculation. (c) 16 hours post inoculation.
Figure 4.
Go enrichment analysis based on the differential expression genes between L-arabinose and D-glucose-cultured strain NCD-2. The X-axis indicates the number of genes classified into regulatory or functional categories, as depicted on the Y-axis. Columns in blue indicate down-regulation and in red indicate up-regulation. (a) 8 hours post inoculation. (b) 12 hours post inoculation. (c) 16 hours post inoculation.
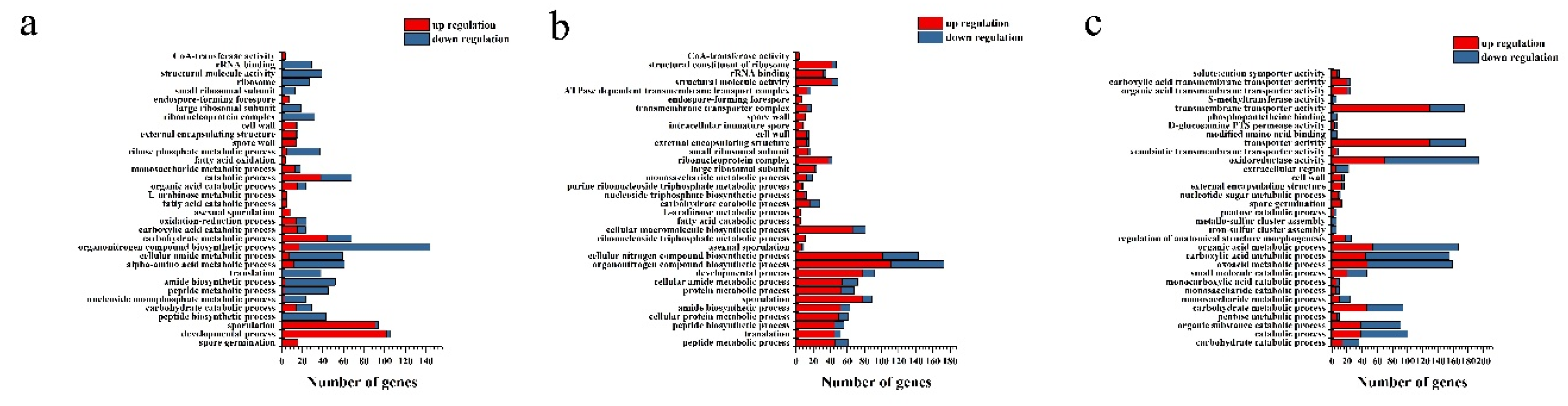
Figure 5.
KEGG enrichment analysis based on the differential expression genes between L-arabinose and D-glucose cultured strain NCD-2. The X-axis indicates the number of genes classified into regulatory or functional categories, as depicted on the Y-axis. Columns in blue indicate down-regulation and in red indicate up-regulation. (a) 8 hours post inoculation. (b) 12 hours post inoculation. (c) 16 hours post inoculation.
Figure 5.
KEGG enrichment analysis based on the differential expression genes between L-arabinose and D-glucose cultured strain NCD-2. The X-axis indicates the number of genes classified into regulatory or functional categories, as depicted on the Y-axis. Columns in blue indicate down-regulation and in red indicate up-regulation. (a) 8 hours post inoculation. (b) 12 hours post inoculation. (c) 16 hours post inoculation.
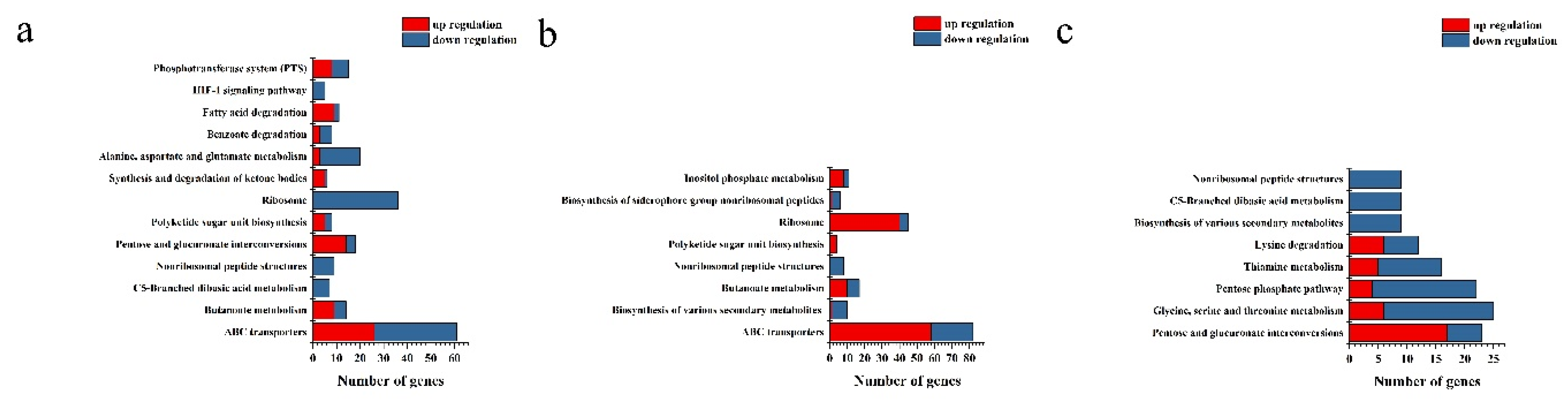
Figure 6.
The bacteria concentration (a) and sporulation efficiency (b) of strain NCD-2 wild type (WT), msmX-null mutant (ΔmsmX) and its complemented strain (CPmsmX) in M9 medium with L-arabinose as carbon source, at 24 , 36 and 48 h after inoculation. Columns represent the average of three replicates, error bars show standard deviations, different letters indicate significant (P<0.05) difference according to ANOVA with Tukey’s post hoc test.
Figure 6.
The bacteria concentration (a) and sporulation efficiency (b) of strain NCD-2 wild type (WT), msmX-null mutant (ΔmsmX) and its complemented strain (CPmsmX) in M9 medium with L-arabinose as carbon source, at 24 , 36 and 48 h after inoculation. Columns represent the average of three replicates, error bars show standard deviations, different letters indicate significant (P<0.05) difference according to ANOVA with Tukey’s post hoc test.
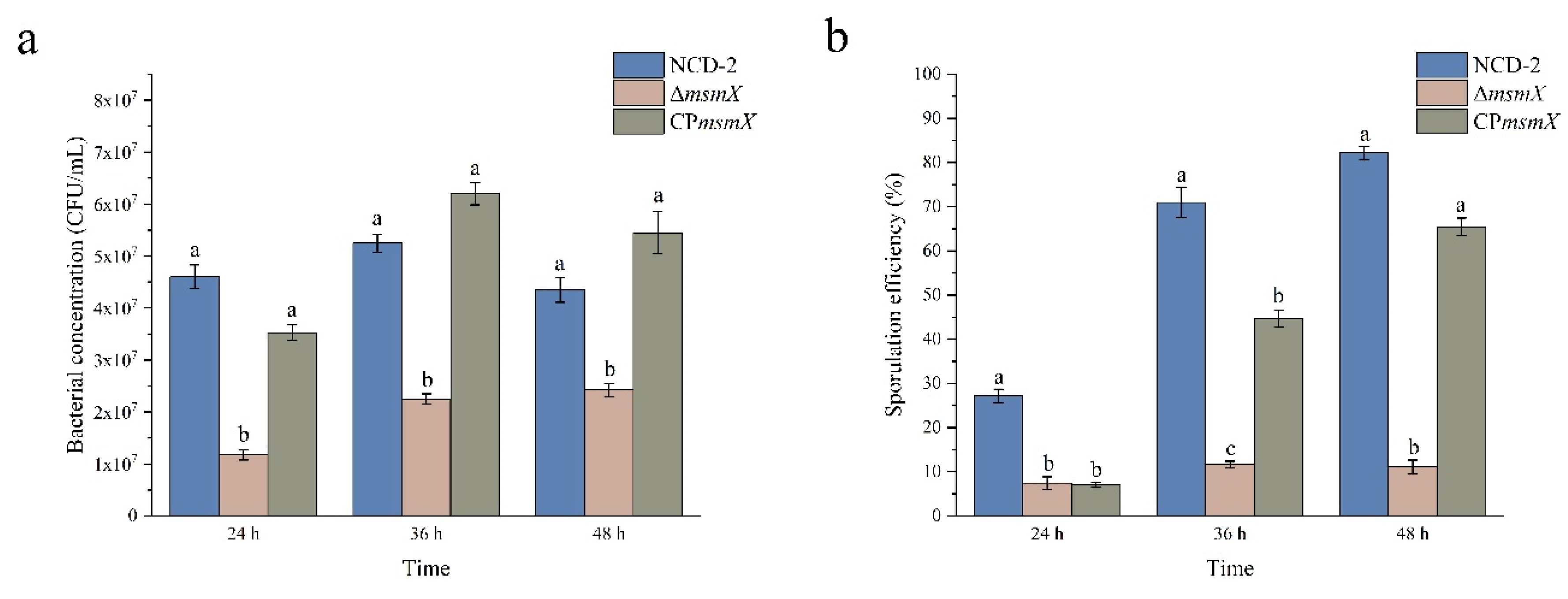

Table 1.
Differentially expressed sporulation related genes in L-arabinose treated strain NCD-2.
| Accession ID | Gene nane | Log2(Ara/Glc) | Production | ||
|---|---|---|---|---|---|
| 8 h | 12 h | 16 h | |||
| WP_003231833.1 | cotE | 7.65 | 5.81 | 2.56 | outer spore coat protein CotE |
| WP_003243364.1 | cotF | 6.59 | 9.47 | 5.54 | spore coat protein CotF |
| WP_080344234.1 | cotG | 3.55 | 5.59 | 3.82 | spore coat protein CotG |
| ADV92699.1 | cotM | 5.01 | 1.53 | 4.21 | spore coat protein (outer) |
| WP_047183078.1 | cotS | 3.98 | 6.01 | 4.53 | spore coat protein CotS |
| PSM02245.1 | cotT | 5.88 | 5.95 | 8.39 | spore coat protein |
| AGE63031.1 | cotV | 4.37 | 6.32 | 5.46 | spore coat protein (insoluble fraction) |
| WP_069486390.1 | cotW | 4.53 | 6.81 | 5.73 | spore coat protein |
| WP_014476454.1 | cotX | 4.28 | 6.83 | 6.13 | spore coat protein |
| WP_003231888.1 | dpaA | 6.29 | 6.84 | 4.81 | dipicolinic acid synthetase subunit A |
| WP_003231884.1 | dpaB | 5.76 | 6.12 | 4.21 | dipicolinate synthase subunit B |
| WP_015383228.1 | yheD | 5.17 | 3.35 | 2.27 | spore coat associated protein YheD |
| WP_063336053.1 | gerBA | 4.55 | 2.20 | 4.73 | spore germination protein GerKA |
| WP_003184172.1 | gerE | 3.94 | 5.01 | 3.91 | spore germination protein GerE |
| WP_014478336.1 | gerQ | 7.95 | 4.77 | 1.32 | spore coat protein GerQ |
| WP_047182746.1 | gerT | 5.10 | 6.02 | 6.03 | spore germination protein GerT |
| AKE24397.1 | sigK | 6.45 | 4.08 | 3.26 | RNA polymerase sporulation-specific sigma factor |
| WP_047182864.1 | spoIIIAE | 3.84 | 2.72 | 1.41 | stage III sporulation protein AE |
| WP_003221804.1 | spoIIID | 10.25 | 6.81 | 4.40 | sporulation transcriptional regulator SpoIIID |
| WP_004398593.1 | spoIIM | 1.78 | 1.84 | 2.35 | stage II sporulation protein M |
| WP_047183325.1 | spoIIQ | 5.69 | 1.78 | -1.82 | stage II sporulation protein SpoIIQ |
| WP_004398697.1 | spoIVB | 6.19 | 1.77 | 2.12 | SpoIVB peptidase |
| WP_015483522.1 | spoIVFB | 1.25 | 1.39 | 1.84 | stage IV sporulation protein SpoIVFB |
| WP_003230465.1 | spoVAD | 6.29 | 1.70 | 4.08 | stage V sporulation protein AD |
| AGE63365.1 | spoVD | 2.67 | 3.20 | 2.32 | penicillin-binding protein |
| WP_047182441.1 | yjcA | 6.69 | 5.32 | 5.67 | sporulation protein YjcA |
| WP_015383520.1 | ykvU | 3.60 | 3.01 | 4.18 | sporulation protein YkvU |
| WP_003223491.1 | sspA | 7.22 | 3.14 | 3.04 | alpha/beta-type small acid-soluble spore protein |
| WP_003233287.1 | sspB | 7.39 | 3.43 | 2.66 | alpha/beta-type small acid-soluble spore protein |
| WP_003218568.1 | sspD | 7.57 | 3.93 | 4.00 | alpha/beta-type small acid-soluble spore protein |
| BAI84385.2 | sspE | 6.59 | 4.34 | 3.30 | gamma-type small acid-soluble spore protein |
| WP_003244950.1 | sdpC | -1.40 | -5.57 | -6.81 | sporulation delaying protein family toxin |
| WP_003228357.1 | sdpI | -4.53 | -3.43 | -4.90 | immunity protein SdpI |
| WP_003243541.1 | sdpR | -4.51 | -3.34 | -2.64 | sporulation delaying system autorepressor SdpR |
Table 2.
Strains used in this study.
| Strain | Genotype | Source |
|---|---|---|
| WT | Bacillus subtilis NCD-2 wild type | Lab stock |
| ΔmsmX | NCD-2 mutant, msmX deletion mutant | This study |
| CPmsmX | Complementary of ΔmsmX by intact msmX, CmR | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



