Submitted:
26 October 2023
Posted:
27 October 2023
You are already at the latest version
Abstract
Organic Medium Enclosed Trough (OMET) is a new non-drainable growing technique. The study envisioned to investigate the interactive effects of OMET and Amaranth species vari-ation on some growth attributes, bioactive compounds and nutrients. Three Amaranth species: A. caudatus, A. cruentus and A. tricolor were grown in an OMET and non-OMET growing conditions. Growth attributes, irrigation water, bioactive compounds and nutri-ents were determined. Non-OMET and A. cruentus showed the highest total phenolics and flavonols followed by A. caudatus and A. tricolor, whilst tannins were predominant in A. caudatus followed by A. cruentus and A. tricolor. The use of unsupervised PCA showed major metabolic variation based on Amaranth species and the use of supervised OPLS-DA showed clear metabolic variation based on OMET and non-OMET. OMET and A. cruentus showed the highest plant height, stem diameter, biomass, and some nutrients (%protein, Ca, P, Cu, Se, Zn), followed by A. caudatus based only on growth and biomass, and A. tricol-or based only on some nutrients (Mg, K, Mn, and Fe). The use OMET system on A. cruentus and A. tricolor can be recommended to mitigate climate change effects on nutritional com-position using low irrigation water.
Keywords:
amaranth
; OMET
; bioactive compounds
; amino acids
; minerals
; PCA
1. Introduction
Amaranthus spp. is one of the commonly consumed leafy vegetables from the Amaranthaceae botanical family. In South Africa, tender young leaves are cooked to be part of daily meals as relish [1]. Given its nutritional status predominated by macro and micro nutrients, it falls under the recommended indigenized leafy vegetables with the potential to combat food and nutritional insecurity [1,2]. Amaranths consists of 60 species including A. cruentus, A.caudatus and A. tricolor which differ based on their morphological attributes such as type of leaf venation or blades, count of stamens in the inflorescence as well as the presence of red spots on their leaf surfaces [3]. This further corroborate that there are nutritional composition and secondary metabolites variation in different Amaranth species. For example, Sarker and Oba [4] reported leaves of different A. cruentus accessions to be rich in dietary fiber, protein, fat, carbohydrates, energy and ash. In fact, Amaranth has sulphur-containing amino acids, which are normally limited in other protein rich crops such as grain legumes, thus making Amaranth ideal for supporting human dietary needs. Moreover, Mateos-Maces et al. [5] reported that phytochemical composition in A. cruentus were predominated by total phenolic compounds equipped with phenolic acids (hydoxycinnamic and hydroxybenzoic acid) and flavonoids. However, Amaranths is still one of the forgotten leafy vegetables often produced in a small scale farming which limits their commercial distribution. Different cultivation practices have been reported as part of the strategies to enhance cultivation of different Amaranth species. These include the use of organic fertilizers, biostimulants, regulating the planting date, and irrigation schedules [6,7,8].
The Organic Medium Enclosed Trough (OMET) system is a non-drainable growing technique characterized by sandwiching growth medium to form an enclosed trough using a polyethylene plastic [9]. The OMET system design offers a micro-climate system to the growing seedling which utilizes less water, eliminate water or nutrient seepage and requires no weeding. Therefore, OMET can be regarded a green-smart growth technique to reduce the effect of climate change. The OMET system has been reported to improve growth and yield in okra [10]. Yet there is still a gap of information on its impact on growth, yield, nutritional composition and bioactive compounds in Amaranth species (A. caudatus, A. cruentus and A. tricolor).
2. Materials and Methods
2.1. Experimental site, and treatments of the OMET system
This trial was established in a greenhouse setting in 2021 and was repeated in 2022 between November and December at the Green Biotechnologies Research Centre of Excellence (GBRCE), University of Limpopo, in South Africa (23⁰53′10″S, 29⁰44′15″E). The site is located 2,126 m above the sea level. The temperatures and relative humidity in the greenhouse ranged from 18 °C to 24 °C and 40% and 50%, respectively. A randomized complete block design was used to layout a 2x3 factorial trial consisting of two growing conditions (OMET and non-OMET) and three Amaranth species (A. caudatus, A. cruentus and A. tricolor) in triplicates. The OMET system was prepared on greenhouse benches whereby, a layer of growth medium (25 cm thick) was sandwiched using high density polyethylene (HDPE) plastic (flexible, translucent/waxy, weatherproof, good low temperature toughness (-60°C), easy to process by most methods, low cost and good chemical resistance) to form an enclosed trough [10]. The growth medium comprised of steam pasteurized loam soil, sand and Hygromix growth medium (Hygrotech, Pretoria North, in South Africa) at a ratio of 2:1:1. Holes for planting the seedlings under OMET were made on the top enclosing sheet with row spacing of 20 cm x 20 cm between seedlings. In contrary, non-OMET system (control) was prepared the same way but without plastic.
2.2. Establishment of seedling on the OMET system
Seeds of the three Amaranth species namely A. caudatus, A. cruentus, and A. tricolor, were obtained from the Agricultural Research Council, Vegetables, Industrial, Medicinal Plants (ARC-VIMP), Pretoria, South Africa. Seedlings for each Amaranth species were germinated on a seedling trays filled with Hygromix® growth medium up to four weeks when the seedlings were at four leaf stage. Then, uniform seedlings were transplanted on the holes for OMET and non-OMET treatment per specie. Irrigation of seedling were performed only when the status of ‘dry’ was shown on a Xylux moisture tester and the volume of water per irrigation was 250 ml tap water as described by Mpai et al. [11]. Fertilisation with Multi feeder (Multisol ‘P’) (active ingredients being nitrogen, phosphorus, potassium, magnesium, zinc, iron, copper, manganese and boron) was applied at week 4, and 8 for each Amaranth species. From the preliminary experiments, Amaranth maximases its leaf production before the flowering stage that comes after eight weeks of transplanting, therefore the seedling were grown up to eight weeks (56 days) of harvesting to good quality leaves.
2.3. Growth and yield data collection
Growth parameters: Stem diameter and plant height of seedling were collected once per week during growth period. The stem diameter was measured five centimetres above the soil surface using a digital Vernier calliper; plant height and leaf length were measured using a tape measurer and meter ruler respectively; time of flowering was recorded as per observation. Fresh mass was measured using laboratory weighing balance (YC001, RSA). Thereafter, the leaf samples from each Amaranth species were packaged separately in brown bags and oven dried at 40 °C for 72 hours. After drying them, they were then grinded into fine powder using a coffee grinder (29105A, RSA).
2.4. Determination of total irrigation water
Irrigation water was recorded as weekly sum of irrigation water. The amount used for irrigation was constant (250 ml). Details of calculation of irrigation water were similar to those described by Mpai et al. [11]. The only variation was the day interval for irrigation/week over eight weeks as the plant grows. The Xylux moisture tester was used to guide when to irrigate (when the moisture status was ‘dry’).
2.5. Determination of bioactive compounds and HPLC-MS-QTOF untargeted metabolites.
Extraction of the Amaranth samples for bioactive compounds (total phenols, total flavonoids, and tannins) and HPLC-MS-QTOF untargeted metabolites profiling were performed. A total of 200 mg of dried amaranth sample were homogenized with 2 mL methanol: HCL: distilled water (80:0.5:19.5 v/v/v) to allow a 1:10 ratio of sample and a solvent in a thermostatic shaking water bath at 70 °C [3] for 15 min following the method described by Mpai et al. [12] without any modifications. Prior to the biochemical analysis, the extract were centrifuged for 15 min dried under the N2 gas and re-suspended and filtered as described by Mpai and Sivakumar [13]. Total phenol content was determined following the Folin-Ciocalteu method as the described by Mpai et al. [12]. The results were calculated and reported as mg/100 g using Gallic acid standard curve. Total flavonoids were carried out following an aluminium chloride method [14]. The results were calculated using a catechin standard curve. The Folin-Ciocalteu method [15] was used to determine the tannin content of the three Amaranth species leaves. Tannin content was expressed as mg GAE/100g DW.
Chlorophyll a and b was performed using fresh leaves of Amaranth species (0.2 g) and the procedure adopted was similar to that described by Managa et al. [16]. Chlorophyll were extracted using 1.5 ml of acetone-hexane mixture (4:6 volume/volume, v/v). After shaking and centrifugation at 10 000 rpm for 15 min, 250 μl of the supernatant was transferred into a 96 well micro plate and the absorbance was read at 470, 646 and 662 nm wavelength. The content of Chl a+b gives the total chlorophyll content, and it was expressed in mg per 100 g on a fresh weight basis [16].
2.6. Determination of protein
The percentage protein was estimated using the micro Kjeldahl method as described by the Helrich [17]. In each Amaranth species, two grams (2 g) for each sample was mixed with 10 ml of concentrated sulphuric acid H2SO4, in a heating tube. One tablet of selenium catalyst was added to the tube and the mixture was heated inside a fume hood. The digest material were transferred into a 100 ml volumetric flask and made up with distilled water. Ten millilitre (ml) portion of the digest was mixed with equal volume of 45% NaOH solution and poured into a Kjeldahl distillation apparatus. The mixture was distilled, and the distillate was collected into a 4% boric acid solution, containing 3 drops of indicator. A total of 50 ml distillate was collected and titrated as well. The sample was duplicated three times, and the average value was taken. The nitrogen content was calculated and converted to percentage protein by using a protein conversion factor of 6.25. This was given as:
% nitrogen = (100 x W x N x 14 x Vf) T 100 x Va
Where; W= Weight of the sample, N= Normality of the titrate (0.1N), Vf= Total volume of the digest = 100 ml, T= Titre value and Va= Aliquot volume distilled.
2.7. Determination of minerals or trace elements
Mineral estimation was performed using oven dried leaf samples of the Amaranth species following the method described by Mathipa et al. [18]. A total 10 g dried materials were digested in 40 ml of 4% nitric acid (HNO3), followed by placing the container on a vortex to allow for complete wetting of the mixture. The materials were magnetically stirred, thereafter incubated in a 95 °C water-bath for 90 minutes, allowed to cool down at room temperature, filtered, decanted into 50 ml tubes which were covered with a foil and then selected nutrient elements were analysed using the inductively coupled plasma optical emission spectrometry (ICPE-9000).
2.10. Determination of amino acids
Amino acid analysis was performed using dried leaves of three Amaranth species, according to Mpai et al. [12] using a fresh leaf samples (100 mg) which was vortexed with 6 N HCl 0.5 ml. The outcome mixture was held in an oven at 110 °C for 18 hours and left to cool. After cooling, the mixture was centrifuged and filtered. The resulting filtrate was dried using a speed vac and reconstituted in borate buffer (70 µl) for derivatisation following the method by Mpai et al. [12].
2.11. Data analysis
The study adopted a two factorial analysis (OMET and Amaranth species). Mean separation for significant treatments was achieved through Duncan multiple range test (DMRT) at the significance level of 5% (p≤0.05) using computer statistical software, Statistix 10.0.
3. Results
3.1. Effect between growth conditions and species on growth, flowering, irrigation water and yield attributes of three Amaranth species
The results of the effects of the OMET system on growth of stem diameter and plant height of the three Amaranth species are shown in Figure 1 and Figure 2. There were significant (p≤0.05) difference in the growth of stem diameter and plant height of the three Amaranth species grown under OMET and non-OMET. The stem diameter and plant height in A. cruentus grown in an OMET and non-OMET system outperformed all other studied treatments over the growing period. However, stem diameter and plant height in A. caudatus grown in an OMET system were higher than that of non-OMET and A. tricolor species grown in OMET and non-OMET.
Table 1 shows the flowering time of the three Amaranth species grown on OMET and non-OMET growing conditions. The A. caudatus and A. tricolor grown under the non-OMET flowered early at week four and five respectively. In contrary, the same species grown under OMET and A. cruentus (OMET and non-OMET) flowered a week later at week six.
The amount of water used was constant (250 ml per irrigation day) and increased in the intervals of irrigation per week as observed in Figure 3. Samples grown in OMET system utilised 2500 ml of irrigation water whereas non-OMET system utilized 3000 ml irrespective of the species. The OMET system has utilized 500 ml lesser than the non-OMET system.
The OMET system and species interaction significantly (p≤0.01) affected the plant fresh biomass. Overall, OMET grown A. cruentus (ACr.) measured the highest biomass (103 g), followed by OMET grown A. caudatus (Aca.) (69 g). In contrary, non-OMET grown A. tricolor (ATr.) measured the lowest biomass (34 g). There was no significant difference between the OMET grown A. tricolor and non-OMET grown A. cruentus (36 g) (Figure 4).
3.2. Interactive effect between growth condition and species on bioactive compounds and HPLC-MS-QTOF metabolites
3.2.1. Bioactive compounds
Growing of Amaranth species on a non-OMET were found to increase the contents of total phenols, and total flavonoids, such that samples of A. cruentus (262.45 mg GAE/100g DW and 473.28 mg CA/100g DW ) outperformed all other studied species, followed by A. caudatus (259.68 mg GAE/100g DW and 453.34 mg CA/100g DW) and lastly A. tricolor (257.24 mg GAE/100g DW and 447.59 mg CA/100g DW) (Table 2). The non-OMET further enhanced the accumulation of tannins and samples of A. caudatus were found to be the highest (9.60 mg GAE/100g DW), followed by those of A. cruentus (8.78 mg GAE/100g DW) and A. tricolor (6.89 mg GAE/100g DW). These total tannins found in non-OMET treatment (control) were two-fold higher than those found in OMET treated samples as shown in Table 2. The results in Table 3 illustrates that the OMET system maintained chlorophyll content in all the three Amaranth species. In the present study, the highest chlorophyll (Chl a+b) was recorded in OMET grown A. caudatus (48.085 mg/kg), followed by the non-OMET and OMET grown A. caudatus and A. cruentus respectively. The lowest Chl a+b concentration was obtained in non-OMET grown A. tricolor (30.946 mg/kg) (Table 3). Total chlorophyll content under OMET grown Amaranth species was higher compared to that of the same Amaranth species grown under non-OMET system.
3.2.2. HPLC-MS-TOF metabolites
To summarise the obtained results from the HPLC-MS-Q-TOF of untargeted metabolites profiling, the chemometric analysis approach was applied to observe metabolite variations and similarities within samples treated with OMET and non-OMET in three Amaranth species. With the use of unsupervised principal component analysis (PCA) (Figure 5A), two main clusters were observed and separated based on the Amaranth species. In fact, samples of A. tricolor grown on OMET (TR1) system showed a holistic distinctive metabolome profile from samples of A. caudatus (CA1) and A. cruentus (CR1) irrespective of their growth condition (OMET system). However, there was no clear clustering between samples of A. caudatus and A. cruentus grown on either OMET or non-OMET. Therefore, a supervised OPLS-DA plot was generated, and results obtained therein showed good model statistics with predictive ability (Q2 cum value: 62%) that was above 50% (Figure 5B). Two clear clusters were now observed to separate 1: A. tricolor (TR1) and A. caudatus (CA1) grown on OMET, and 2: A. cruentus grown on OMET (CR1) and the other three species grown on non-OMET. The OPLS-DA model (Figure 8) was generated to demonstrate metabolites biomarker responsible to the OMET and non-OMET clusters irrespective of the studied Amaranth species. The tentative identification based on molecular mass (mz), fragmentation, chemical formula and retention time (rt) were used to identify compounds that contributed significantly to variation between OMET and non OMET (Table 4). The compounds responsible for metabolome variation between OMET and non OMET were detected as 6-Feruloylglucose 2,3,4-trihydroxy-3-methylbutylglycoside, a fatty acyl glycosides of mono and di-saccharides (rt: 15.658; mz: 474.08758, and MSE: 474.08758:246 475), and Apigenin 7-O-glucoside’ (rt: 17.63; mz: 421.28; MSE: 268.03810:269.04341), respectively. Other compounds that showed significant contribution variations were showed at rt: 8.093; mz: 218.10211, MSE: 218.10655:4843 219, which were identified to be pantothenic acid. The ‘Apigenin 7-O-glucoside’ is flavonoid compound which plays a role on improving the adaptation of crops to abiotic stress such as drought. Further studies on its quantification are necessary.
3.3. Interactive effect between growth condition and species on nutritional composition
3.3.1. Protein and nutritive minerals
The % protein content of the three Amaranth species is shown in Table 5. The highest protein content was recorded in OMET grown A. cruentus (28.6%), followed by OMET grown A. caudatus and non-OMET grown A. cruentus (24.1%). The lowest protein content was recorded in non-OMET grown A. tricolor (20.3%), not significantly different from the % protein recorded in non-OMET grown A. caudatus (21.4%).
Table 6A represents the contents of macro-elements (Ca, Mg, K and P) and Table 6B shows micro-elements (Cu, Mn, Fe, Se and Zn) in three Amaranth species grown under the OMET and non-OMET growing system. The OMET grown A. cruentus had the highest Ca (130 mg/kg) and P (34.7 mg/kg) contents, whilst the highest Mg (82.5 mg/kg) and K (276 mg/kg) contents were found in the OMET grown A. tricolor. Non-OMET grown A. cruentus recorded the lowest concentrations of Ca (88.3 mg/kg), Mg (56.6 mg/kg) and K (217 mg/kg) in comparison to all other studied Amaranth species grown under both OMET and non-OMET. The same trend was observed in OMET grown A. tricolor that had the highest concentration of P. In contrast, the non-OMET grown A. tricolor had the lowest concentration of P (21.4 mg/kg) than the other species under both treatments. There was no significant difference Ca (104 mg/kg DW) measured in A. caudatus in both treatment and OMET grown A. tricolor.
The following microelements: Cu, Mn, Fe, Se, and Zn were quantified (Table 6B). It was revealed that the OMET grown A. cruentus had the highest concentration of Cu (1.04 mg/kg), Se (8.13 mg/kg) and Zn (1.66 mg/kg), whereas the A. tricolor grown under OMET system was found to contain the highest concentration of Mn (2.16 mg/kg) and Fe (3.41 mg/kg) than the rest of the species grown both under OMET and non-OMET system. The OMET grown A. caudatus contained the second highest concentration of Fe (2.62 mg/kg) and Se (8.03 mg/kg). The non-OMET grown. A. tricolor measured the lowest Cu (0.78 mg/kg DW), Se (6.36 mg/kg DW) and Zn (0.95 mg/kg DW) than the other Amaranth species grown under both treatments. There was no significant difference Fe measured in all Amaranth species grown under both treatments except for OMET grown A. tricolor which had an outstanding Fe concentration (3.41 mg/kg DW). The highest Fe concentration of A. tricolor (3.41 mg/kg DW) was higher than the Fe concentration reported by Yahaya et al. [19] in A. caudatus (1.06 mg/kg DW), Roselle (1.2 mg/kg DW) and Kenaf (1.17 mg/kg DW).
3.3.2. Amino acid composition
The total of 16 free amino acids were quantified in the three Amaranth species grown under OMET and non-OMET system (Table 7). The results revealed that Amaranth species contained both essential and non-essential amino acids. There was a significant difference (p≤0.05) between amino acid concentration among the same Amaranth species grown under OMET and non-OMET. Essential amino acids threonine, lysine, valine, isoleucine, leucine, and phenylalanine were detected in all Amaranth species grown under both OMET and non-OMET system. The levels of phenylalanine in OMET grown A. cruentus (2.54 mg/kg) (Table 7) were higher than the amount reported in the leaves of A. cruentus (0.66 mg/kg) [20]. The OMET grown A. cruentus also showed a slightly higher amount of leucine (2.21 mg/kg) than the amount reported in A. cruentus (1.55 mg/kg) [20]. The highest concentration of valine (1.45 mg/kg) detected in OMET grown A. cruentus showed to be lower than the amount reported in A. cruentus (1.51 mg/kg) [20]. Moreover, the highest quantity of isoleucine (1.29 mg/kg), and threonine (1.15 mg/kg) detected in OMET grown A. cruentus showed to be both higher than the amount reported in leaves of A. cruentus (isoleucine (0.83 mg/kg) and threonine (0.85 mg/kg) [20]. The lysine concentration was 2.37 and 2.25 mg/kg in OMET grown A. cruentus and OMET grown A. caudatus respectively (Table 7), higher than the concentration reported in the leaves of A. cruentus (1.73 mg/kg) [20].
Nonessential amino acids, serine, arginine, glycine, aspartate, and glutamate were detected in three Amaranth species. The glycine, aspartate and glutamate were identified as the predominant non-essential amino acids. The highest concentration of glycine in non-OMET grown A. cruentus (1.46 mg/kg) was much higher than the amount reported by Manyelo et al. [20] in A. cruentus (0.94 mg/kg). Aspartate content was the highest in non-OMET grown A. cruentus (2.02 mg/kg). The non-OMET grown A. cruentus was predominated by glutamate (2.55 mg/kg). Serine was highest in non-OMET grown A. cruentus (1.16 mg/kg) (Table 7), higher than the amount reported in A. cruentus (0.90 mg/kg) [20]. Moreover, the lowest concentration of arginine (0.53 mg/kg) was detected in non-OMET grown A. cruentus (Table 7), lower than the concentration reported in A. cruentus (0.90 mg/kg) [20].
4. Discussion
The quest for improving food security in South Africa entails scouting for strategies that will be adapted to the changing climate and further contains adequate nutrients for human health nourishment. In this study, the interactive effect between Amaranth species and growth condition (OMET and non-OMET) were showed as crucial factors to manipulate growth, yield, nutrients and secondary metabolites contents.
The obtained results on the growth and yield section may be attributed by variation of species and OMET system influence. The results can be assimilated with those mentioned in a mulching studied due to the similarities of the concepts. Saeed and Ahmad [21] reported that mulch is effective for vegetable growth and yield by improving moisture content of soil, heat energy. Furthermore, reasons related to the potential of OMET to conserve water and nutrients, which causes a moderate availability of water resulting in good nutrient dissolution and absorption by the plants could also attribute to the results obtained [22,23]. Recently, Mokgalabone et al. [10] reported the efficacy of OMET on improving growth and yield attributes in okra seedlings. Plant height and stem diameter have major impact on biomass. Early flowering in A. caudatus and A. tricolor grown under the non-OMET system could probably be due to abiotic stress such as water and nutrient stress. The exposure of the plant to shortened drought period induces early flowering and halts the production of leaves [24]. In this case, the non-OMET system had high rates of water and nutrients loss through evaporation, drainage and leaching.
Irrigation water used for OMET system were lower than those used for non-OMET system, irrespective of the studied species. This could be due to the underlying plastic on the OMET system which is water impermeable, therefore water loss through seepage/ deep drainage were completely eliminated unlike non-OMET system. The high evaporation rate in the OMET system is eliminated by the top plastic covering the growing medium [9].
Furthermore, this can be due to species type in response to different conditions and growing condition factors such as moisture and nutrient availability associated with plant growth and development under each treatment. Although Amaranth is drought resistant, it performs optimally under irrigation [24] and sufficient supply of nutrients especially Nitrogen (N). One of the essential elements, and one which participates directly as an indispensable requirement for normal plant growth, is N. Application of the multi-feeder made N and other vital nutrients become readily available for uptake by the plants. Probably due to the potential of OMET system to conserve irrigation water and nutrients, it prolonged the availability and uptake of water, N and other nutrients by the plants, thus prompting vigorous vegetative growth and high yielding in Amaranth species. The increased yield of Amaranth was probably due to the ability and potential of OMET to supply frequent and enough water, as Amaranth is said to yield optimally under sufficient irrigation. Scharenbroch and Lloyd [25] reported that organic materials increase soil organic matter by directly improving soil properties and ultimately better plant growth and biomass.
Given the response of Amaranth species to OMET and non-OMET, bioactive compounds including total phenols, total flavonoids, and tannins showed a decrease in OMET conditions. These results suggest the efficacy of OMET on keeping water and nutrition (fertilisers) at the plant rooting zones. According to the known carbon-nitrogen theory [26], nitrogen fertiliser enhances the accumulation of nitrogen containing metabolites such as amino acids and protein, whilst reducing the production of compounds such polyphenols (phenols and flavonoids) [27]. On this basis, the OMET grown Amaranth had less bioactive compounds and higher contents of amino acids and proteins. Minor drought stress is associated with major causes for crop loss and poor performance. In this case, Amaranth species grown under non-OMET system developed minor drought, leading to a significant reduction of photosynthesis [28]. These fact are authenticated by the chlorophyll contents on the leaves, which were reduced in non-OMET samples. Some in vivo study demonstrated that water deficit results in damages of the photosynthesis II (PSII) oxygen-evolving complex [29].
Kwenin and Dzomeku [30] reported 4.46% protein content in A. cruentus, much lower than all the protein content of all the Amaranth species in the present study, ranging between 20.3 and 28.6%. The protein content of A. cruentus (28.6%) is considerably higher than common vegetables such as spinach, cabbage and kale consumed in RSA. Hanif et al. [31] measured % protein content ranging from 0.9 to 2.1% in cauliflower, carrot, cabbage, lettuce, spinach. There are numerous factors such as nutrient and water conservation potential by the OMET system that influenced the accumulation of protein in all the three Amaranth species. Wijewardana et al. [32] revealed that maintaining a high level of soil moisture especially during the reproductive stage was beneficial to acquiring higher protein content. In this study, OMET system compared to non-OMET system was moisture conserving growing technique which maintained a higher level of soil moisture content, which might have influenced highest accumulation of protein in OMET grown Amaranth species. Differences in species and treatment probably attributed to different protein contents. Amaranth vegetables grown under OMET system can be an important source of dietary protein.
In the study of Yahaya et al. [19], Se concentrations for A. caudatus was not quantified. Yahaya et al. [19] reported the Zn concentrations of A. caudatus (0.048 mg/kg DW which was lower than the lowest Zn concentration of non-OMET grown A. cruentus (1.66 mg/kg DW) in the present study. The low concentrations of all the microelements in the present study were found in non-OMET grown Amaranth species. A. tricolor grown on non-OMET system had the lowest concentration of Cu (0.78 mg/kg), Fe (2.47 mg/kg), Se (6.36 mg/kg) and Zn (0.95 mg/kg) than the rest of the Amaranth species grown on both OMET and non-OMET system. In addition, the lowest concentration of Mn was found in non-OMET grown A. cruentus (1.17 mg/kg), higher than the concentrations reported by Yahaya et al. [19] for A. caudatus (0.079 mg/kg DW). The A. caudatus grown on both OMET and non-OMET system had average concentrations of the microelements as compared to other species. The OMET system enhanced the concentration of microelements (Table 3).
Minerals are essential for plant growth, development, reproduction, and seed quality. Mineral deficiency and poor minerals uptake and transport due to abiotic stress such as drought especially at the reproductive stage result in yield loss and poor seed quality [33]. Lowered absorption of the minerals could be due to reduced transpiration flow, limited availability of energy for assimilation, and interference in the unloading mechanism [34,35]. OMET system has successfully increased the concentration of the mineral elements for Amaranth species than the non-OMET system. This could probably be due to its potential to eliminate leaching of nutrients, conserving and reserving them for uptake by the plant roots. The less to non-fluctuating temperatures of the OMET system growing medium is conducive for better nutrient absorption. The root exudates become useful for the plant since they are not leached. The non-OMET system on the other hand had little influence on the accumulation of mineral elements, probably due to its inability to conserve nutrients. The nutrients are leached during irrigation through deep drainage or seepage since there’s no underlying plastic to inhibit drainage, the nutrients become out of reach for plant roots absorption. The mineral accumulation is also influenced by the degree of soil moisture stress. Both macro and micro-elements showed to be higher under OMET grown Amaranth species, subjected to highest soil moisture compared to non-OMET grown Amaranth species. It can be concluded that the optimum high moisture content positively enhances elemental composition.
The results clearly showed that both essential and non-essential amino acids are highly available in the OMET grown A. cruentus than any other Amaranth species grown under both treatments. Lysine, leucine, and phenylalanine were the three dominating essential amino acids whereas glycine, aspartate and glutamate were the dominating nonessential amino acids in all the three Amaranth species. Furthermore, both non-OMET grown A. cruentus and OMET grown A. tricolor shows to be the second potential sources of amino acids compared to other Amaranth species with distinct treatments. Based on the amino acid analysis, it can be concluded that OMET grown A. cruentus can be regarded as a reliable growing technique for Amaranth species to be greater sources of essential amino acids (Table 4). Therefore, for the enhancement of a balanced diet, the daily consumption of A. cruentus needs to be encouraged. The trend or pattern in amino acid composition could relate to possible inherent differences between genotypes, species and/or treatment. Similar trends were observed regarding the concentrations of essential amino acids in A. caudatus and A. cruentus probably confirm that these species respond almost the same to the same treatment as compared to A. tricolor.
5. Conclusion and recommendations
The climate change lower crop yields and nutritional quality due to drought, heat waves and flooding as well as increases in pests and plant diseases, leading to high rates of malnutrition and food insecurity. The good news is that there are tools in the form of science-based farming practices that buffer farmers from climate damage and help make their operations more resilient and sustainable for the long term, such as OMET system.
In this study, the aim was to investigate the effects of the OMET system on the growth, yield, nutritional and phytochemical composition of Amaranth species. The findings of the study suggest maintaining optimum soil moisture, soil temperature and nutrient conservation (OMET system) is very significant for the accumulation of improved % protein content, amino acids, and mineral content. We found that OMET system reduces the cases of plant stress, which prohibits early flowering of Amaranth species, positively influencing better plant growth and ultimately, higher yields. This study reveals complementary effects between the OMET system and Amaranth species with regards to the accumulation of primary metabolites. It is important to investigate other growing techniques that will improve the yield and quality of indigenous vegetables. Furthermore, investigations in Amaranth species for the development of nutraceutical products will be essential. Inclusion of the Amaranth species in the food database would be a source of income for the government, Department of Agriculture, Forestry and Fishery.
Author Contributions
M.G.M.: Experimentation, data gathering and data analysis and initial draft write-up; S.M: Conceptualization, revisions. A R.N: Funding acquisition, Conceptualization.
Funding
Authors would like to thank the financial support from the National Research foundation grant number (Grant number 12612) and Department of Science and Innovations (DSI), South African Government (Grant number DSI/CON C2235/2021).
Acknowledgments
The authors would like to acknowledge Mr Ndivhuwo Mutshekwa and Ms Lerato Raphoko for technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bvenura, C., and Afolayan, A. J., 2015. The role of wild vegetables in household food security in South Africa: A review. Food Research International, 76(P4), 1001–1011. [CrossRef]
- South-East Asia Region and WHO, 2021. Regional Nutrition Strategy : Addressing malnutrition and micronutrient deficiencies.
- Taia23TP0F, W.K., 2021. Vegetative morphological variations within some Egyptian Amaranthus L. species. Jordan Journal of Biological Sciences, 14(1). [CrossRef]
- Sarker, U. and Oba, S., 2019. Salinity stress enhances colour parameters, bioactiv leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. Journal of the Science of Food and Agriculture, 99(5), 2275-2284. [CrossRef]
- Mateos-Maces, L., Chávez-Servia, J.L., Vera-Guzmán, A.M., Aquino-Bolaños, E.N., Alba-Jiménez, J.E. and Villagómez-González, B.B., 2020. Edible leafy plants from Mexico as sources of antioxidant compounds, and their nutritional, nutraceutical and antimicrobial potential: A review. Antioxidants, 9(6), 541. [CrossRef]
- Jimoh, M. O., Afolayan, A. J., and Lewu, F. B., 2018. Suitability of Amaranthus species for alleviating human dietary deficiencies. South African Journal of Botany, 115, 65–73. [CrossRef]
- Ngoroyemoto, N., Gupta, S., Kulkarni, M. G., Finnie, J. F., and Van Staden, J., 2019. Effect of organic biostimulants on the growth and biochemical composition of Amaranthus hybridus L. South African Journal of Botany, 124, 87–93. [CrossRef]
- Sarmadi, B., Rouzbehan, Y., and Rezaei, J., 2016. Influences of growth stage and nitrogen fertilizer on chemical composition, phenolics, in situ degradability and in vitro ruminal variables in Amaranth forage. Animal Feed Science and Technology, 215,73–84. [CrossRef]
- Ferreira, J., 2013. OMET farming better than hydroponics? Farmer’s weekly.
- Mokgalabone, T.T., Mpai, S. and Ndhlala, A.R., 2023. Organic Medium Enclosed Trough Growing Technique Improves Abelmoschus esculentus (Okra) Growth, Yield and Some Nutritional Components. Applied Sciences, 13(9), p.5645. [CrossRef]
- Mpai, S., Mokganya, L.M., Raphoko, L., Masoko, P. and Ndhlala, A.R., 2022. Untargeted metabolites and chemometric approach to elucidate the response of growth and yield attributes on different concentrations of an amino acid based biostimulant in two lettuce cultivars. Scientia Horticulturae, 306, p.111478. [CrossRef]
- Mpai, S., Du Preez, R., Sultanbawa, Y. and Sivakumar, D., 2018. Phytochemicals and nutritional composition in accessions of Kei-apple (Dovyalis caffra): Southern African indigenous fruit. Food chemistry, 253, 37-45. [CrossRef]
- Mpai, S. and Sivakumar, D., 2020. Influence of growing seasons on metabolic composition, and fruit quality of avocado cultivars at ‘ready-to-eat stage’. Scientia Horticulturae, 265, p.109159. [CrossRef]
- Makkar, H.P., Siddhuraju, P. and Becker, K., 2007. Plant secondary metabolites (Vol. 393, pp. 1-122). Totowa, NJ, USA: Humana Press.
- Tambe, V.D. and Bhambar, R.S., 2014. Estimation of total phenol, tannin, alkaloid, and flavonoid in Hibiscus tiliaceus Linn. Wood extracts. Journal of Pharmacognosy and Phytochemistry 2(4): 2321-6182.
- Managa, M.G., Sultanbawa, Y. and Sivakumar, D., 2020. Effects of different drying methods on untargeted phenolic metabolites, and antioxidant activity in Chinese cabbage (Brassica rapa L. subsp. chinensis) and nightshade (Solanum retroflexum Dun.). Molecules, 25(6), 1326. [CrossRef]
- Helrich, K., 1990. Official methods of analysis of the Association of Official Analytical Chemists. Association of official analytical chemists.
- Mathipa, M.M., Mphosi, M.S. and Masoko, P., 2022. Phytochemical Profile, Antioxidant Potential, Proximate and Trace Elements Composition of Leaves, Stems and Ashes from 12 Combretum spp. Used as Food Additives. International Journal of Plant Biology, 13(4), pp.561-578. [CrossRef]
- Yahaya, Y., Birnin-Yauri, U.A., Bagudo, B.U. and Noma, S.S., 2012. Quantification of macro and micro elements in selected green vegetables and their soils from Aliero agricultural fields in Aliero, Kebbi State, Nigeria. Journal of Soil science and Environmental Management, 3(8): 207-215. [CrossRef]
- Manyelo, T.G., Sebola, N.A. and Mabelebele, M., 2020. Nutritional and phenolic profile of early and late harvested Amaranth leaves (Amaranthus cruentus) grown under cultivated conditions, 432(10): 05. [CrossRef]
- Saeed, R. and Ahmad, R., 2009. Vegetative growth and yield of tomato as affected by the application of organic mulch and gypsum under saline rhizosphere. Pakistan Journal of Botany, 41(6), 3093-3105.
- Ferrini, F., Fini, A., Frangi, P. and Amoroso, G., 2008. Mulching of ornamental trees: Effects on growth and physiology. Arboric Urban Forum, 34: 157. [CrossRef]
- Ngala, J.M., Ndiso, J.B. and Mundi, E.M., 2019. Effects of selected organic mulches on growth and yield of Amaranth in Kilifi country. International Journal of Agriculture, Environment and Bioresearch. Volume 4, No. 06. [CrossRef]
- Department of Agriculture, Forestry and Fisheries (DAFF), 2010. Amaranthus production guideline. Pretoria, RSA.
- Scharenbroch, B.C. and Lloyd, J.E., 2006. Particulate organic matter and soil N availability in urban landscapes. Arboric Urban Forum. 32: 180. [CrossRef]
- Kazimierczak, R., Hallmann, E. and Rembiałkowska, E., 2015. Effects of organic and conventional production systems on the content of bioactive substances in four species of medicinal plants. Biological Agriculture & Horticulture, 31(2), pp.118-127. [CrossRef]
- Coley, P.D., Bryant, J.P. and Chapin III, F.S., 1985. Resource availability and plant anti-herbivore defense. Science, 230(4728), pp.895-899.
- Da Silva Ferreira, V. and Sant’Anna, C., 2017. Impact of culture conditions on the chlorophyll content of microalgae for biotechnological applications. World Journal of Microbiology and Biotechnology, 33(1): 1-8. [CrossRef]
- Skotnica, J., Matoušková, M., Nauš, J., Lazár, D. and Dvořák, L., 2000. Thermoluminescence and fluorescence study of changes in Photosystem II phytochemistry in desiccating barley leaves. Photosynthesis Research, 65(1): 29-40. [CrossRef]
- Kwenin, W.K.J., Wolli, M. and Dzomeku, B.M., 2011. Assessing the nutritional value of some African indigenous green leafy vegetables in Ghana.
- Hanif, R., Iqbal, Z., Iqbal, M., Hanif, S. and Rasheed, M., 2006. Use of vegetables as nutritional food: role in human health. Journal of Agricultural and Biological Science, 1(1): 18-22.
- Wijewardana, C., Reddy, K.R. and Bellaloui, N., 2019. Soybean seed physiology, quality, and chemical composition under soil moisture stress. Food Chemistry, 278: 92-100. [CrossRef]
- Bellaloui, N., Mengistu, A., Fisher, D.K. and Abel, C.A., 2012. Soybean seed composition constituents as affected by drought and Phomopsis in phomopsis susceptible and resistant genotypes. Journal of Crop Improvement, 26(3), 428-453. [CrossRef]
- Farooq, M., Wahid, A., Kobayashi, N.S.M.A., Fujita, D.B.S.M.A. and Basra, S.M.A., 2009. Plant drought stress: effects, mechanisms and management. In Sustainable agriculture (153-188). Springer, Dordrecht. [CrossRef]
- Rouphael, Y., Cardarelli, M., Schwarz, D., Franken, P. and Colla, G., 2012. Effects of drought on nutrient uptake and assimilation in vegetable crops. In Plant responses to drought stress (171-195). Springer, Berlin, Heidelberg. [CrossRef]
Figure 1.
The effects of OMET system on stem diameter in (A) Amaranthus caudatus, (B) Amaranthus cruentus and (C) Amaranthus tricolor from week 0 to 8 after transplanting. Results are expressed as the mean values ± standard error (n=12).
Figure 1.
The effects of OMET system on stem diameter in (A) Amaranthus caudatus, (B) Amaranthus cruentus and (C) Amaranthus tricolor from week 0 to 8 after transplanting. Results are expressed as the mean values ± standard error (n=12).
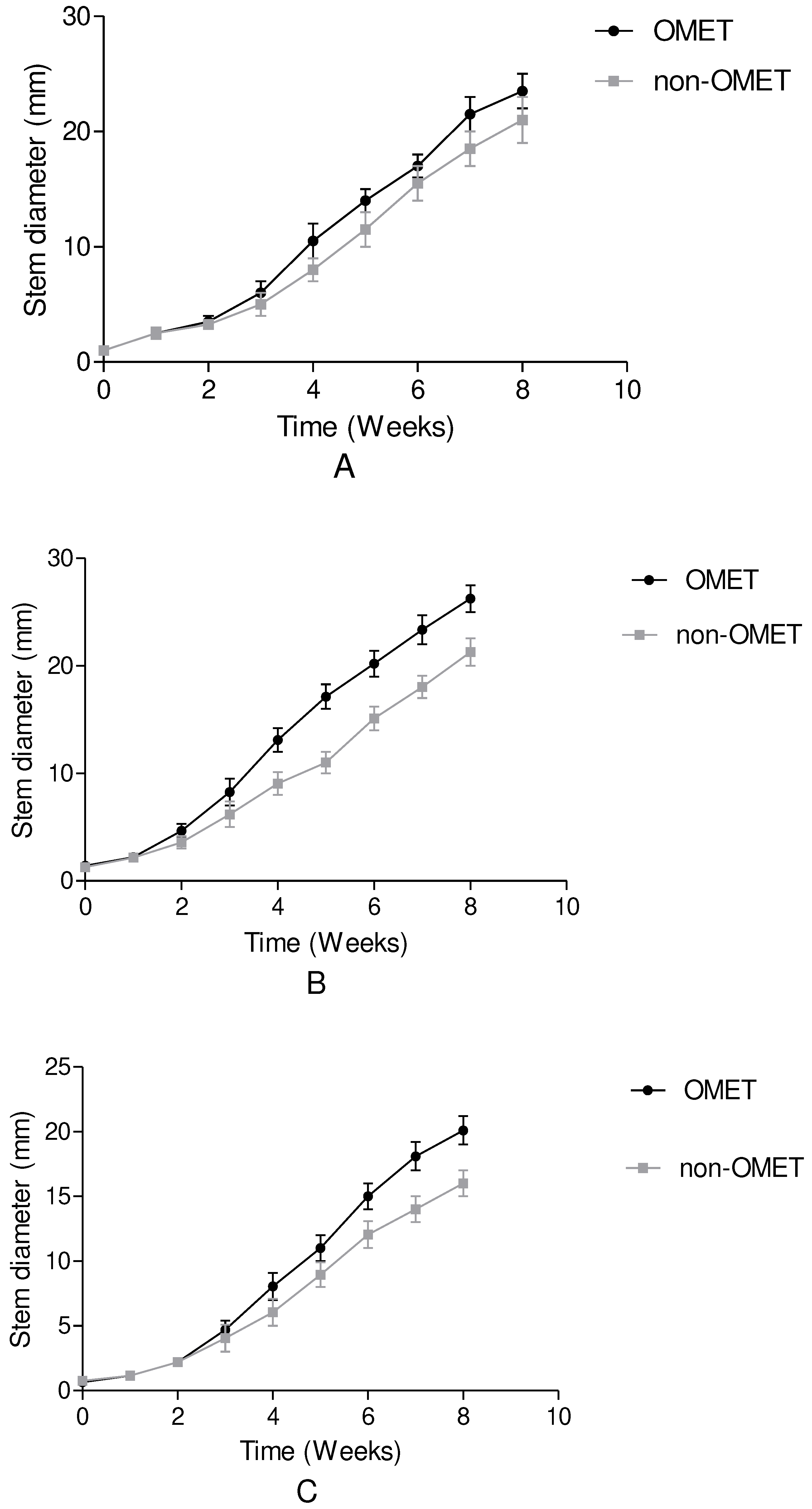
Figure 2.
The effects of OMET system on plant height of (A) Amaranthus caudatus, (B) Amaranthus cruentus and (C) Amaranthus tricolor over 8 weeks after transplanting. Results are expressed as the mean values ±standard error (n=12).
Figure 2.
The effects of OMET system on plant height of (A) Amaranthus caudatus, (B) Amaranthus cruentus and (C) Amaranthus tricolor over 8 weeks after transplanting. Results are expressed as the mean values ±standard error (n=12).
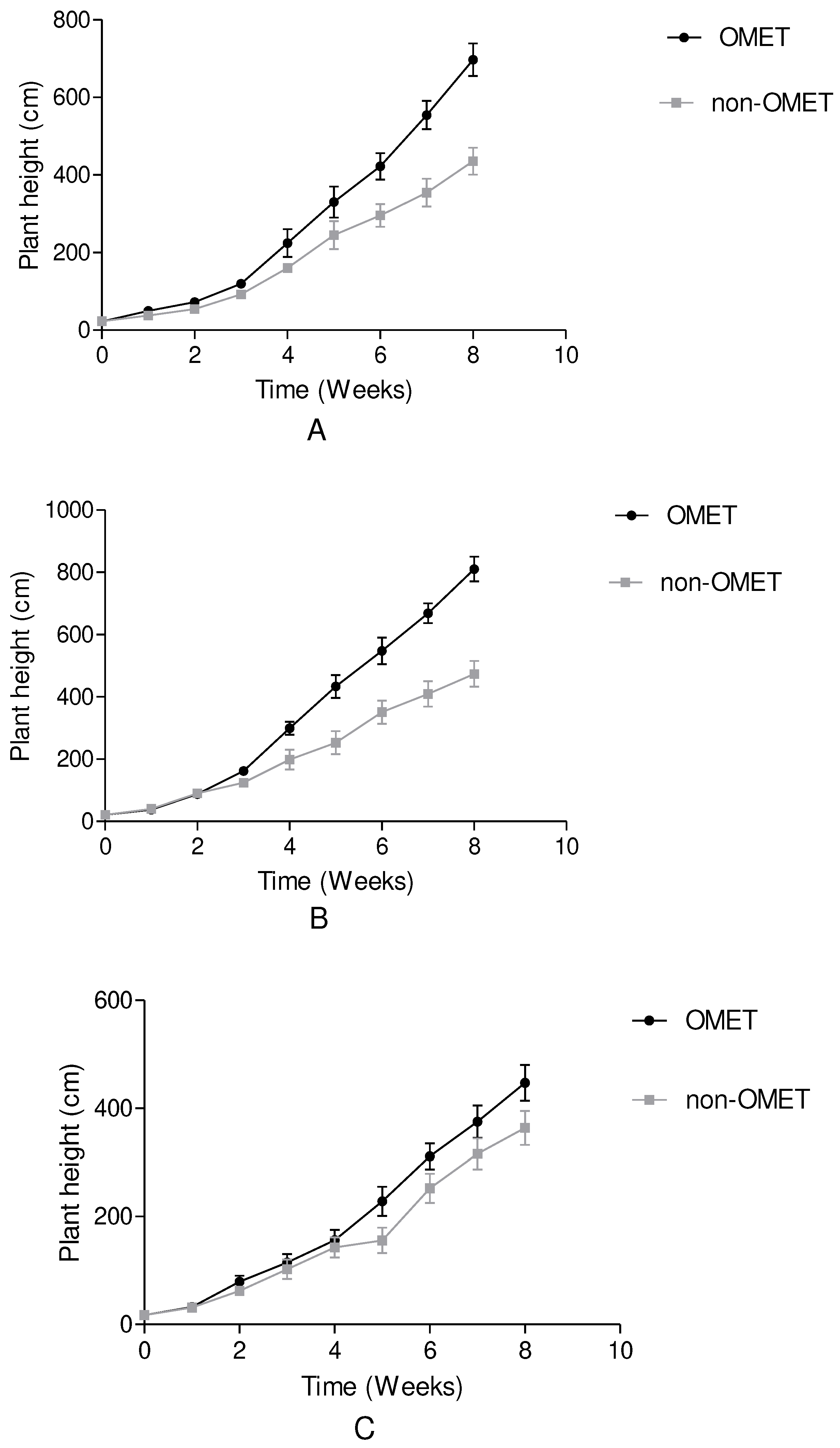
Figure 3.
Cumulative amount of water utilised weekly/plant over a period of 8 weeks under OMET and non-OMET system.
Figure 3.
Cumulative amount of water utilised weekly/plant over a period of 8 weeks under OMET and non-OMET system.
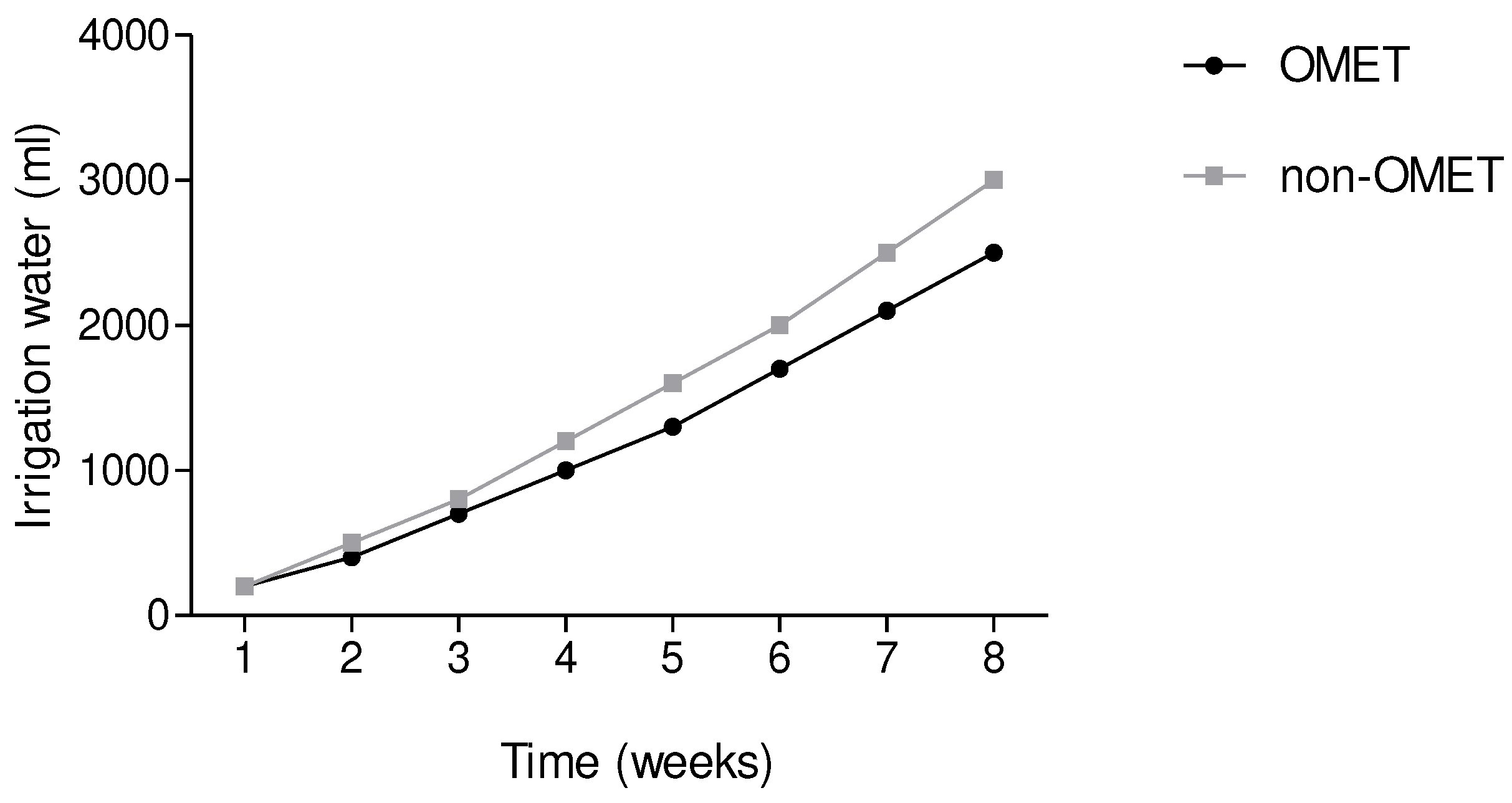
Figure 4.
The effects of OMET system on fresh biomass of three Amaranth species. Results are expressed as the mean values ± standard error (n=12). Bars in the same column marked with different letters indicate significant difference at p≤0.05. Aca. = Amaranthus caudatus; ACr. =Amaranthus cruentus, ATr. = Amaranthus tricolor.
Figure 4.
The effects of OMET system on fresh biomass of three Amaranth species. Results are expressed as the mean values ± standard error (n=12). Bars in the same column marked with different letters indicate significant difference at p≤0.05. Aca. = Amaranthus caudatus; ACr. =Amaranthus cruentus, ATr. = Amaranthus tricolor.
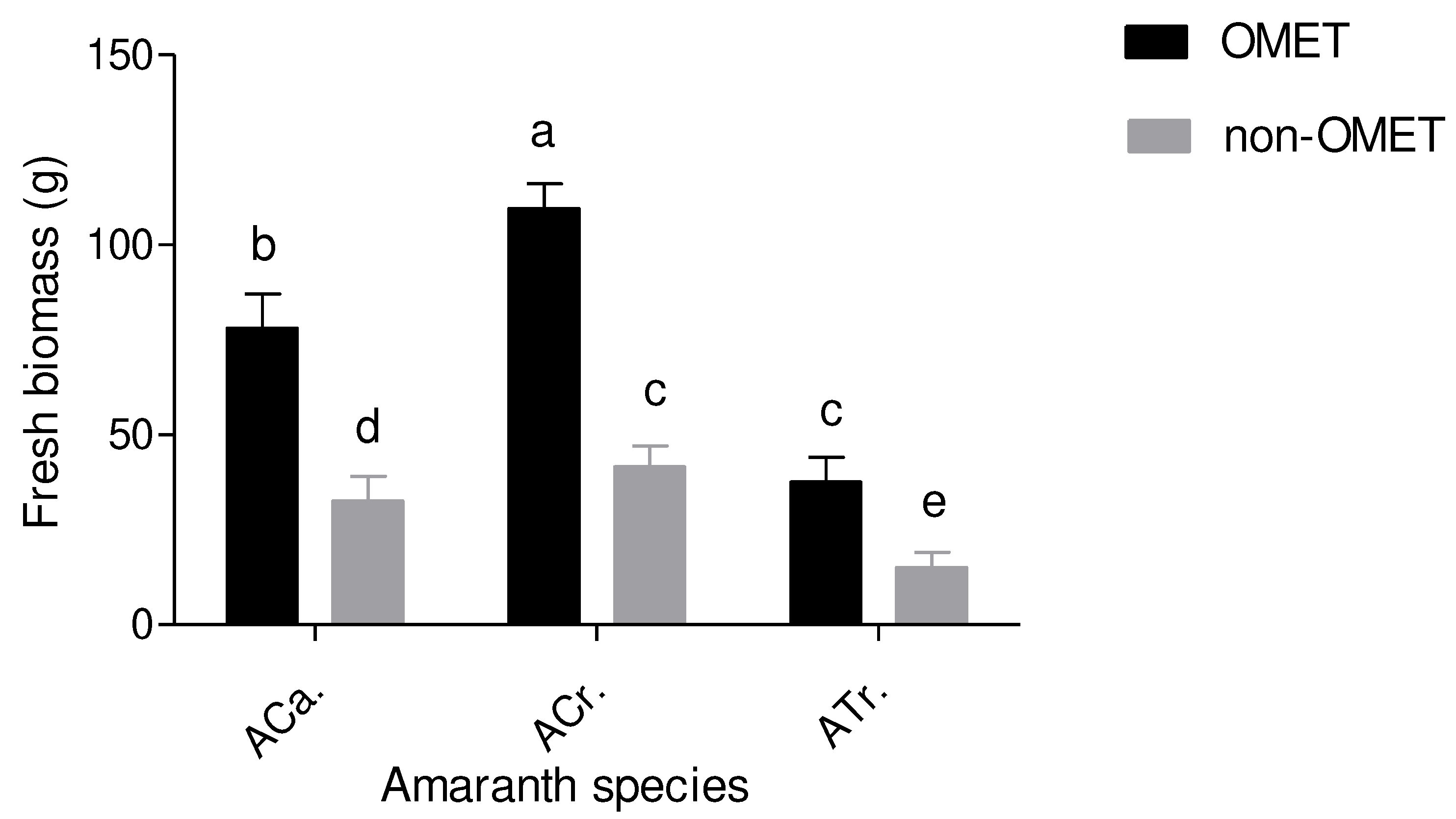
Figure 5.
HPLC-MS-QT of untargeted metabolites profiling of Amaranthus caudatus, Amaranthus cruentus and Amaranthus tricolor, (A) unsupervised PCA and (B) supervised PCA (1= OMET ad 2= non-OMET).
Figure 5.
HPLC-MS-QT of untargeted metabolites profiling of Amaranthus caudatus, Amaranthus cruentus and Amaranthus tricolor, (A) unsupervised PCA and (B) supervised PCA (1= OMET ad 2= non-OMET).
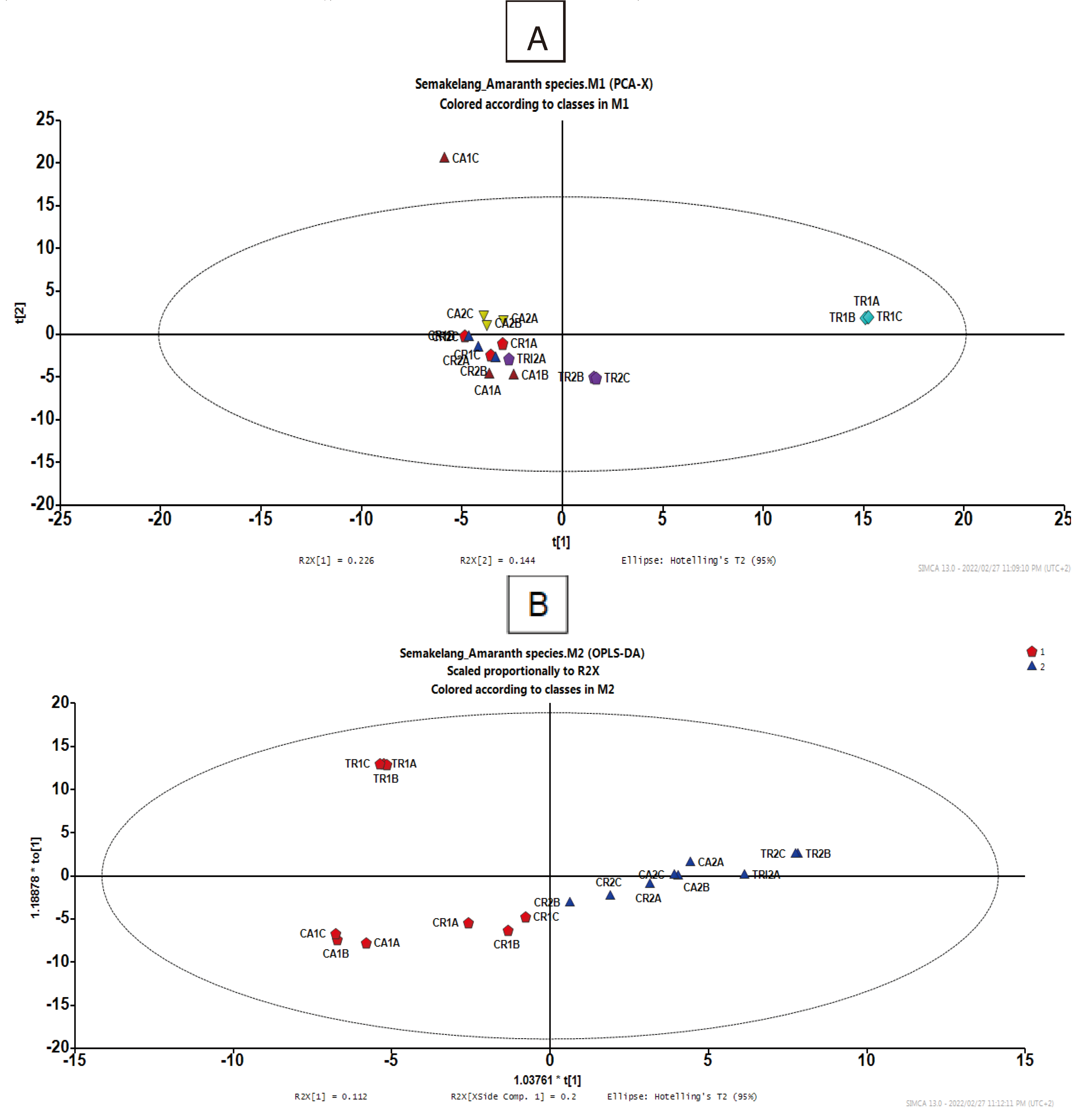
Figure 6.
HPLC-MS-QT of untargeted metabolites profiling of Amaranthus caudatus, (B) Amaranthus cruentus and (C) Amaranthus tricolor; Pareto scaling.
Figure 6.
HPLC-MS-QT of untargeted metabolites profiling of Amaranthus caudatus, (B) Amaranthus cruentus and (C) Amaranthus tricolor; Pareto scaling.
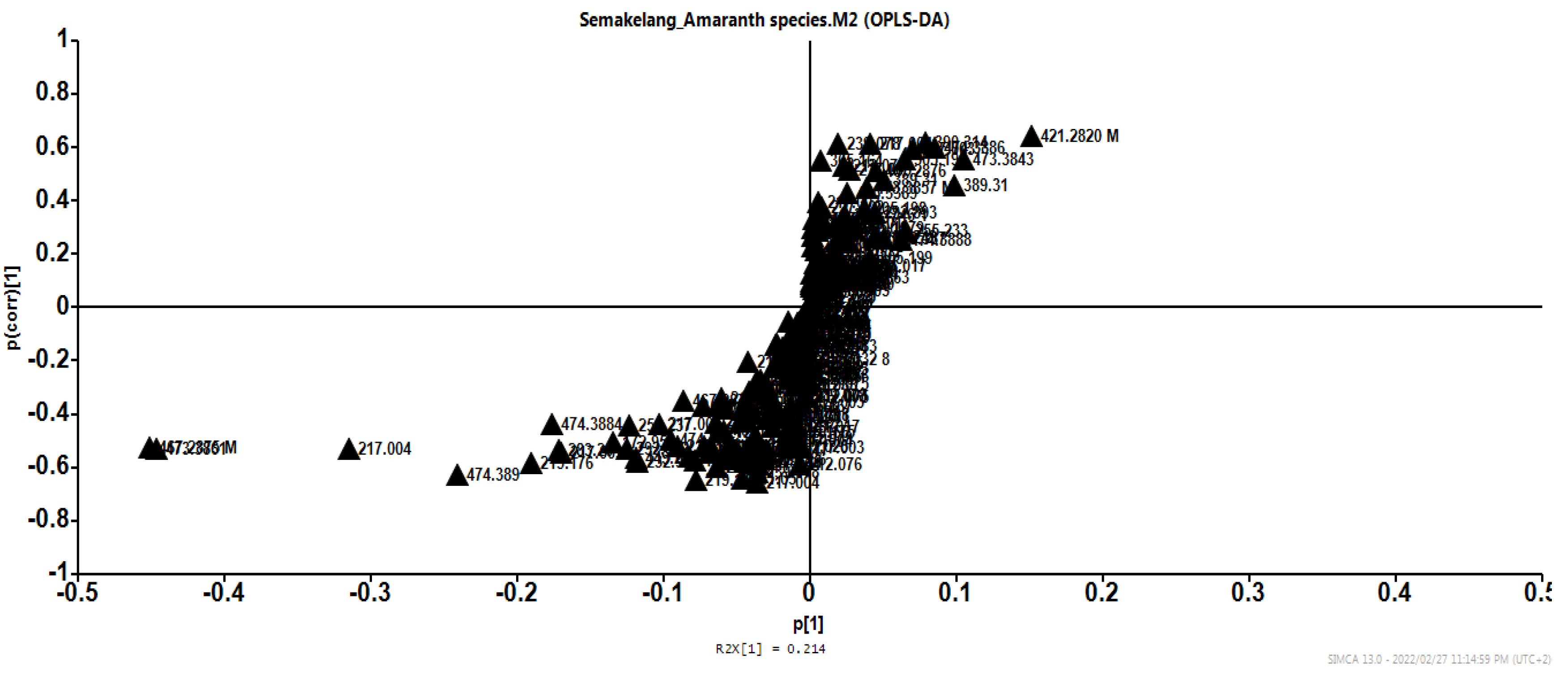

Table 1.
Time of flowering in three Amaranth species grown under OMET and non-OMET system.
| Treatment | Amaranth species | Time of flowering (Weeks) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| OMET system | Amaranthus caudatus | X | |||||||
| Amaranthus cruentus | X | ||||||||
| Amaranthus tricolor | X | ||||||||
| non-OMET system | Amaranthus caudatus | X | |||||||
| Amaranthus cruentus | X | ||||||||
| Amaranthus tricolor | X | ||||||||
Table 2.
The interactive effects of OMET system and Amaranth species on total phenolics, flavonoids and tannins.
Table 2.
The interactive effects of OMET system and Amaranth species on total phenolics, flavonoids and tannins.
| Amaranth species & treatment | Total phenolic (mg GAE/100g DW) | Total flavonoids (mg CA/100g DW) |
Total tannins (mg GAE/100g DW) | |
| A. caudatus | OMET | 210.24±0.033d | 391.26±0.046d | 4.09±0.042d |
| non-OMET | 259.68±0.046b | 453.34±0.036b | 9.60±0.022a | |
| A. cruentus | OMET | 209.65±0.035d | 382.60±0.047e | 3.98±0.043d |
| non-OMET | 262.45±0.028a | 473.28±0.021a | 8.78±0.031b | |
| A. tricolor | OMET | 207.70±0.0.036e | 364.24±0.038f | 3.61±0.038d |
| non-OMET | 257.24±0.040c | 447.59±0.042c | 6.89±0.040c | |
DW= Dry weight, Different letters in the same column indicate significance difference at p≤0.05.
Table 3.
The interactive effects of OMET system and Amaranth species on chlorophyll concentration (mg/kg).
Table 3.
The interactive effects of OMET system and Amaranth species on chlorophyll concentration (mg/kg).
| Chl a (mg/kg) | Chl b (mg/kg) | Chl a+b (mg/kg) | ||
| A. caudatus | OMET | 0.306±0.005a | 47.779±0.004a | 48.085±0.005a |
| non-OMET | 0.301±0.0047b | 47.142±0.003a | 47.443±0.007a | |
| A. cruentus | OMET | 0.301±0.0048b | 47.142±0.008a | 47.443±0.008a |
| non-OMET | 0.296±0.0034c | 46.306±0.003b | 46.603±0.004a | |
| A. tricolor | OMET | 0.289±0.044d | 45.253±0.048c | 45.543±0.045a |
| non-OMET | 0.197±0.036e | 30.749±0.041d | 30.946±0.039b | |
Chl a=Chlorophyll a; Chl b=Chlorophyll b; Chl a+b=Chlorophyll a+b; different letters in the same column indicate significance difference at p≤0.05.
Table 4.
Tentative identification of HPLC-Q-TOF-MS untargeted metabolites of Amaranth species.
| Retention time (Min) |
Exact Mass (g/mol) |
Mass generated ESI (-) TOF MS (g/mol) | Fragmentation | Chemical Formula | Tentative structural assignment |
|---|---|---|---|---|---|
| 15.658 | 474.4557 | 474.08423 | 474.08758:246 475 | C21H30O12 | 6-Feruloylglucose 2,3,4-trihydroxy-3-methylbutylglycoside |
| 8,093 | 219.23 | 218.10211 | 218.10655:4843 219 | C9H17NO5 | Pantothenic acid |
| 21.15 | 468.46 | 467,15897 | 467.13629:1292 468 | C22H28O11 | 6-O-(4-Hydroxybenzoyl)-ajugol |
| 17.63 | 420.41 | 420.3132 | 268.03810:269.04341 | C21H24O9 | Apigenin 7-O-glucoside |
Table 5.
The effect of OMET system on total protein % found in three Amaranth species.
| Amaranth species & treatment | Protein (%) | |
|
A. caudatus |
OMET | 24.1±0.039b |
| non-OMET | 21.4±0.044d | |
|
A. cruentus |
OMET | 28.6±0.46a |
| non-OMET | 24.1±0.040b | |
|
A. tricolor |
OMET | 22.7±0.036c |
| non-OMET | 20.3±0.042d | |
| P-value | 0.04 | |
Different letters shows significance difference at p≤0.05.
Table 6.
Effect of OMET system on the macro (A) and micro (B) elements in three Amaranth species.
| 3A | |||||||||||
| Amaranth species & treatment | Macro elements (mg/kg DW) | ||||||||||
| Ca | Mg | K | P | ||||||||
| A. caudatus | OMET | 104±0.043b | 77.3±0.032b | 251±0.041c | 29.7±0.045b | ||||||
| non-OMET | 104±0.043b | 62.6±0.030d | 244±0.038d | 21.8±0.037c | |||||||
| A. cruentus | OMET | 130±0.038a | 63.3±0.029d | 254±0.046b | 34.7±0.043a | ||||||
| non-OMET | 88.3±0.033d | 56.6±0.025e | 217±0.033f | 22.2±0.039c | |||||||
| A. tricolor | OMET | 104±0.039b | 82.5±0.038a | 276±0.05a | 30.3±0.041b | ||||||
| non-OMET | 90.5±0.041c | 66.2±0.032c | 226±0.035e | 21.5±0.036c | |||||||
| 3B | |||||||||||
| Microelements (mg/kg DW) | |||||||||||
| Cu | Mn | Fe | Se | Zn | |||||||
| A. caudatus | OMET | 0.89±0.023c | 1.73±0.046b | 2.62±0.05b | 8.03±0.049a | 1.11±0.024c | |||||
| non-OMET | 0.84±0.021d | 1.46±0.043d | 2.5±0.050c | 7.01±0.048d | 1.09±0.036c | ||||||
| A. cruentus | OMET | 1.04±0.034a | 1.78±0.048b | 2.52±0.04c | 8.13±0.050a | 1.66±0.048a | |||||
| non-OMET | 0.89±0.023c | 1.17±0.041e | 2.51±0.04c | 7.69±0.05b | 1.28±0.045b | ||||||
| A. tricolor | OMET | 0.93±0.025b | 2.16±0.05a | 3.41±0.39a | 7.28±0.05c | 1.27±0.047b | |||||
| non-OMET | 0.78±0.022e | 1.64±0.04c | 2.47±0.05c | 6.36±0.048e | 0.95±0.039d | ||||||
DW=dry weight, values are expressed as means± standard error; different letters in the same column indicate significance difference at p≤0.05.
Table 7.
Total essential and nonessential amino acids (mg/kg) found in three Amaranth species grown both under OMET and non-OMET system.
Table 7.
Total essential and nonessential amino acids (mg/kg) found in three Amaranth species grown both under OMET and non-OMET system.
| Essential amino acids | |||||||
| Threonine | Valine | Isoleucine | Lysine | Leucine | Phenylalanine | ||
| A. caudatus | OMET | 0.92±0.024b | 1.09±0.043c | 0.97±0.031b | 2.25±0.05a | 1.66±0.044b | 2.36±0.05a |
| non-OMET | 0.66±0.025d | 0.84±0.027d | 0.76±0.026c | 1.3±0.034b | 1.25±0.038c | 1.14±0.035c | |
| A. cruentus | OMET | 1.15±0.034a | 1.45±0.041a | 1.29±0.027a | 2.37±0.05a | 2.21±0.042a | 2.54±0.051a |
| non-OMET | 0.93±0.036b | 1.22±0.042b | 1.12±0.039a | 2.19±0.050a | 1.92±0.051b | 1.95±0.043b | |
| A. tricolor | OMET | 0.88±0.021c | 1.08±0.034c | 0.98±0.028b | 1.62±0.036b | 1.66±0.041b | 1.47±0.038c |
| non-OMET | 0.43±0.021e | 0.52±0.026d | 0.46±0.022d | 0.8±0.012c | 0.74±0.024d | 0.71±0.023d | |
| Non-essential amino acids | |||||||
| Arginine | Serine | Glycine | Aspartate | Glutamate | |||
| A. caudatus | OMET | 1.12±0.047b | 0.97±0.034b | 1.37±0.05b | 1.2±0.041c | 1.52±0.05c | |
| non-OMET | 0.86±0.031c | 0.7±0.030c | 0.95±0.041c | 0.96±0.042c | 1.29±0.049c | ||
| A. cruentus | OMET | 1.37±0.048a | 0.98±0.036b | 1.33±0.044b | 1.33±0.044b | 1.75±0.05b | |
| non-OMET | 1.54±0.05a | 1.16±0.045a | 1.46±0.048a | 2.02±0.05a | 2.55±0.05a | ||
| A. tricolor | OMET | 1.12±0.049b | 0.91±0.042b | 1.21±0.047b | 1.53±0.049b | 1.95±0.05b | |
| non-OMET | 0.53±0.028d | 0.46±0.022c | 0.59±0.029d | 0.6±0.030d | 0.73±0.032d | ||
Values are expressed as means ± standard error; different letters in the same column indicate significance difference at p≤0.05.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



