
Submitted:
26 October 2023
Posted:
27 October 2023
You are already at the latest version
Abstract
: Background: Cartilage harvest and transplantation is a common surgery using costal, auricular, and septal cartilage for craniofacial reconstruction. However, absorption and warping of the cartilage grafts can occur due to inflammatory factors associated with wound healing. Transcriptional factor nuclear factor κB (NFκB) is activated by cytokines, such as interleukin-1 (IL-1), and plays a central role in the transactivation of this inflammatory cytokine gene. Inhibition of NFκB may have anti-inflammatory effects. The aim of this study was to explore the potential of an NFκB decoy oligodeoxynucleotide (Decoy) as a chondroprotective agent. Materials and Methods: Evaluation of safe and efficacious concentrations of Decoy were assessed using rabbit nasal septal chondrocytes (rNSCh) and assays for cytotoxicity, proteoglycan (PG) synthesis, and PG turnover. The efficacious concentration of Decoy determined from the rNSCh was then applied to human nasal septal cartilage (hNSC) in vitro and analyzed for PG turnover, the level of inflammatory markers, and catabolic enzymes in explant-conditioned culture medium. Results: Over the range of Decoy conditions and concentrations, no inhibition of PG synthesis or cytotoxicity was observed. Decoy at 10 μM effectively inhibited PG degradation in the hNSC explant, prolonging PG half-life 63% and decreasing matrix metalloprotease 3 (MMP3) by 70.7% (P = 0.027). Conclusions: Decoy may be a novel chondroprotective therapeutic agent in cartilage transplantation due to its ability to inhibit cartilage degradation due to inflammation cytokines.
Keywords:
nuclear factor-κb
; decoy oligodeoxynucleotide
; cartilage
; resorption
1. Background
Cartilage grafting is a common surgical technique in plastic and reconstructive surgery. Tissue sources include cartilage from the rib, nasal septum, and ear for nasal reconstruction, rhinoplasty, microtia repair, and eyelid reconstruction. However, grafts of cartilage have been associated with post-graft resorption and warping [1,2]. Clinically, NSC is an excellent donor source material [3], but resorption rates in the range from 12 to 50 percent have been reported [4,5]. Factors which tend to affect graft absorption and warping include: high tensile forces [4], compressive forces [6], insufficient nutrition and cell source within the recipient area [3], and the inflammatory process associated with wound healing [7].
It is well known that wound healing involves three phases: inflammation, proliferation and remodeling [8]. The inflammatory phase is triggered by many sources of tissue injury, including surgical procedures. Leukocytes are activated with the release of various cytokines, such as interleukin-1 (IL-1), interleukin-6 (IL-6) and tumor necrosis factor- α (TNF) during this initial phase [9]. This is followed by fibroblast proliferation, angiogenesis, and keratinocyte migration. The release of cytokines can cause net catabolic remodeling of the cartilage graft causing an imbalance in graft matrix degradation relative to matrix synthesis [7]. In recognizing this, Haisch et al. [10] showed that encapsulation of the graft with a polyelectrolyte complex membrane, offers a protective barrier for the cartilage implant against an inflammatory reaction. This suggests that ways of locally inhibiting cartilage catabolism may be beneficial.
The transcription factor nuclear factor κB (NFκB) plays a central role in inflammation and the immune response. NFκB is activated by cytokines such as IL-1and TNF [11]. The activated NFκB transactivates genes which are associated with the inflammation and immune responses such as IL-1, IL-6, and TNF [12]. Theoretically, inhibition of NFκB may provide an anti-inflammatory effect. NFκB decoy oligodeoxynucleotide (Decoy) is a synthetic double stranded deoxyribonucleic acid (dsDNA) containing the NF-κB binding site (GGGATTTCCC) which has a high affinity to activated NFκB. In this regard, activated NFκB binds to Decoy instead of the DNA enhancer elements resulting in inhibition of NFκB (Figure 1) [13,14,15,16]. The effectiveness of Decoy has been shown in a variety of previous studies including pulmonary metastasis by murine osteosarcoma [17], acute renal failure in rats [18], liver grafts in rats [19], rheumatoid arthritis in human synovial cells [20], sun damage of skin in mice [21], ischemic injury in a rat epigastric flap model [22], inflammatory bowel disease in rats [23], and intervertebral disc deterioration in rabbits [16]. The effect of Decoy on nasal septal cartilage (NSC) has not been explored yet.
The level of the load-bearing extracellular matrix (ECM) component, proteoglycan (PG), reflects a cytokine-regulated dynamic balance between PG synthesis by indwelling chondrocytes and PG degradation in articular cartilage [24]. In articular cartilage explants, serum stimulates PG synthesis [25] and inhibits PG degradation [26]. Within serum, insulin-like growth factors normally mediate the stimulation of PG synthesis and inhibition of PG degradation [27], whereas the catabolic factor from synovium [28], found to be IL-1 [29], tips the PG balance to a catabolic state, inhibiting PG synthesis and stimulating PG degradation [30]. It has also been shown that IL-1β regulates the expression of catabolic factors in mouse chondrocytes [31]. IL-1 induces formation of nitric oxide (NO) and matrix-depleting enzymes including matrix metalloproteinase-3 (MMP-3). MMP-3 causes PG depletion and cartilage softening [32], while prolonged IL-1 treatment leads to collagen network damage and loss of cartilage integrity [33]. Anti-cytokine protein agents can prevent experimental arthritis following traumatic injury [34]. Thus, the aim of this study was to assess the potential of Decoy as a chondroprotective agent, inhibiting the degradation of PG in hNSC tissue, associated with NO and MMP-3 formation.
2. Materials and Methods
2.1. Study Design
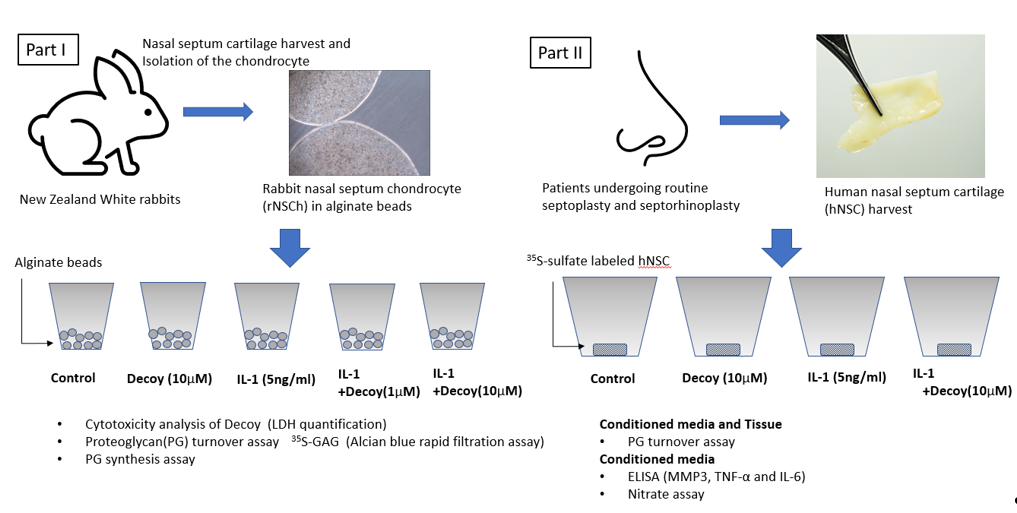
The study design has two parts. In first part, the evaluation of safety and determination of an efficacious concentration of Decoy was performed using rabbit nasal septal chondrocyte (rNSCh). In second part, this determined concentration of Decoy was used for human nasal septal cartilage (hNSC). PG turnover assay and the determination of protein level of inflammation makers were completed. Decoy,5’-CCTTGAAGGGATTTCCCTCC-3’ and 3’-GGAACTTCCCTAAAGGGAGG-5’, was provided by AnGes MG (Osaka, Japan). Decoy is effective in binding the NFκB transcription factor [35,36,37].
2.1.1. Safety and determination of an efficacious concentration of Decoy
2.1.1.1. rNSCh Cell Isolation and Culture in Alginate Beads
After euthanasia, cartilage from the nasal septum was aseptically harvested from 16 New Zealand White rabbits weighing 3.5–4.5kg (Institutional Animal Care and Use Committee approval #S08258). rNSCh were isolated by sequential enzyme digestion for 1 hour at 37 °C with 0.2% pronase (53702, Millipore, Billerica, MA) in Dulbecco’s modified Eagle medium and Ham’s F12 medium (10-090-CV,DMEM/F-12; Corning Inc. Corning, NY) [38]. Afterwards, the tissues were digested overnight with 0.025% collagenase-P (11213865001,Roche Diagnostics, Indianapolis, IN) in DMEM/F12 medium with 5% Fetal bovine Serum (FBS; FB-03, Omega Scientific, Tarzana, CA) [38]. After overnight digestion, the cells were washed with DMEM/F-12 medium, filtered through 70 μm nylon mesh (22-363-548, Fisher Scientific, Pittsburgh, PA) and cell numbers were counted. The cells were then seeded in a T175 flask (CLS431080, Corning Inc.) at 10000 cells/cm2 and cultured for expansion with culture medium (DMEM/F-12 medium containing 10% FBS, 25 μg/mL ascorbic acid [50-81-7, Sigma-Aldrich, St. Louis, MO), 360 μg/mL L-glutamine (25-005-CI, Corning Inc), Antibiotic-Antimycotic Solution (100U/ml penicillin G, 100 μg/mL streptomycin sulfate and 0.25 μg/mL amphotericin B) (30-004-CI, Corning Inc.) and 50 μg/mL gentamicin (15750060, Thermo Fisher Scientific, Waltham, MA)]. The cells were released from monolayer with 0.25% trypsin ethylenediaminetetraacetic acid (EDTA; 17892, Thermo Fisher Scientific) at confluence. The isolated cells were resuspended in a 1.2% low-viscosity sterile alginate (Keltone LV; Kelco, Chicago, IL) solution at a concentration of 2 million cells/ml. Polymerization and formation of the alginate beads was accomplished by injecting the cell-alginate mixture into a 102 mM CaCl2 solution in a dropwise fashion through a 22 gauge needle [38]. After 10 minutes, the newly formed beads (approximately 20,000 cells/beads) were washed three times with a sterile 0.9% NaCl solution followed by two washings with DMEM/F-12. The cells in the beads (9 beads per well) were cultured in 24-well culture plate (CLS3527, Corning Inc.) with complete medium [DMEM/F-12 medium containing 20% FBS, 25 μg/mL ascorbic acid, 360 μg/mL L-glutamine, Antibiotic-Antimycotic Solution and 50 μg/mL gentamicin]. The cultures were maintained at 37°C in a humidified atmosphere of 5% CO2. The medium was changed 3 times per week.
2.1.1.2. Cytotoxicity Analysis of Decoy
As an indicator of cell death, by plasma membrane damage, the release of lactate dehydrogenase (LDH) within the culture medium was measured using a Cytotoxicity Detection Kit (11644793001, Merck, Darmstadt, Germany). Following the culture of rNSCh encapsuled alginate beads for 7 days, Decoy (0 and 10 μM) was transfected to the chondrocytes within the beads. The culture medium was collected after 44h of incubation and assayed for LDH activity according to the manufacturer’s instructions. The in vitro release of LDH from the cells provides an accurate measure of dead cells [39]. Culture medium was collected from the chondrocytes within the beads, 0.9% normal saline served as the control, and administered 0.1% Tween 20 (9005-64-5, Merck) served as the positive control.
2.1.1.3. PG Synthesis Assay
On day 7 of cell culture in alginate beads, the cells were incubated in complete medium in the presence of Decoy and recombinant human IL-1β (IL-1; SIL1B, Thermo Fisher Scientific) (Control, Decoy 10 μM, IL-1 5 ng/ml, IL-1 5 ng/ml +Decoy 1 μM and IL-1 5 ng/ml +Decoy 10 μM). During the last 4 h of culture time, the medium was changed, in each case, to complete medium with the treatment containing [35S]-sulfate (PerkinElmer, Waltham, MA) at a concentration of 20 µCi/ml. After removing the medium, the beads were dissolved with 55 mM sodium citrate, 30 mM EDTA and 150 mM sodium chloride and the two compartments [cell-associate matrix (CM) and further removed matrix (FRM)] were separated by centrifugation at 100 g for 10 min. Each fraction was digested with papain (76216, Merck) and incorporation of [35S]-sulfate was determined using a rapid filtration assay after precipitation of the glycosaminoglycans (GAG) with alcian blue [40]. Rates of [35S]-incorporation were calculated and expressed as CPM [35S]-incorporated/9 beads.
2.1.1.4. PG Turnover Assay
On day 7 of cell culture in alginate beads, the cells were incubated in complete medium containing [35S]-sulfate at a concentration of 20 µCi/ml. Following 16h incubation, the beads were washed 3 times with complete media containing a total of 1.5 mM sulfate in order to removed unincorporated [35S]-sulfate. The medium was changed to complete medium with Decoy (0, 10, 0,1and 10 μM). Following 4h of transfection of Decoy (0, 10, 0, 1 and 10 μM), IL-1 (0, 0, 5, 5 and 5ng/ml) was added. The medium containing Decoy and IL-1 was changed and collected every 24h. On day 5 from the start of treatment, the beads were dissolved and separated to CM and FRM and then digested with papain [35S]-sulfate in the collected media and digested CM and FRM were determined using an alcian rapid filtration assay.
Half-life of PG was calculated using Single Exponential Decay method [41].
2.1.2. Effect of Decoy on hNSC tissue
2.1.2.1. hNSC Harvest and Tissue Culture
hNSC specimens were collected from four patients (30-year-old female, 48-year-old male, 49-year-old male, 58-year-old female) undergoing routine septoplasty and septorhinoplasty at our institution (institutional review board approval, project #130631X). Specimens were transported in sterile saline to the lab. Perichondrium was meticulously removed from cartilage. Within 24 hours of explant, each cartilage specimen was weighed and minced into 10–50mg fragments for tissue culture. The diced cartilage was cultured with complete medium throughout the study. The cultures were maintained at 37 °C in a humidified atmosphere of 5% CO2.
2.1.2.2. Efficiency of Transfection of Decoy
To study transfection efficiency of Decoy in the tissue culture system, fluorescein isothiocyanate (FITC)-labeled Decoy (gifted by AnGes MG) was used. A fragment of whole hNSC was divided in half; one half was cultured for 4h in complete media containing 1 μM of FITC-labeled Decoy. Following washing and incubation for 1h with complete media, the tissue was examined by a confocal spectral laser scanning microscope (Leica Microsystems GmbH, Wetzlar, Germany). To further confirm the cytotoxicity of decoy, the Live-Dead assay of the second half of tissue was performed using LIVE/DEAD Viability/Cytotoxicity Kit for mammalian cells (L3224, Thermo Fisher Scientific) according to manufacturer guidelines.
2.1.2.3. PG Turnover Assay
Following the method described above, after 24h of culture, cartilage was incubated in complete medium containing [35S]-sulfate at a concentration of 20 µCi/ml. Following 16h incubation, the tissue was washed. The medium was changed to complete medium with Decoy and IL-1 (0:0, 10:0, 0:5 and 10 μM:5 ng/ml). The medium containing Decoy and IL-1 was changed and collected every 24h. On day 5 from the start of treatment, the tissue was collected and digested with papain. [35S]-sulfate in the collected media and digested tissue was determined using an alcian blue rapid filtration assay. Half-life of PG was calculated based on fitting [35S] loss to an exponential decay.
2.1.2.4. Enzyme-Linked Immunosorbent Assay (ELISA) Matrix Metalloprotease 3 (MMP-3), TNF and IL-6
After 2 days of treatment with IL-1 and NFκB-Decoy in the tissue culture, the media was assayed by ELISA using human MMP-3 DuoSet (DY513, R&D systems Inc., Minneapolis, MN)), Human TNF High Sensitivity ELISA kit (BMS223HS, Thermo Fisher Scientific) and IL-6 ELISA Ready-SET-GO kit (88-7066-76, Fisher Scientific) according to manufacturer guidelines. All samples were run in duplicate and the resulting quantities were averaged.
2.1.2.5. Measurement of Nitric Oxide (NO)
After 2 days treatment with IL-1 and Decoy in the tissue culture, total Nitrate and Nitrite concentration in the media was also assayed by Nitrate/Nitrite Colorimetric Assay Kit(780001, Cayman Chemical Company, Ann Arbor, MI)according to the manufacturer’s instructions. All samples were run in duplicate and the resulting quantities were averaged.
2.1.2.6. Statistical Analysis
Any assay which showed data to be out of range by the average ± 2 standard deviation (SD) was eliminated. Levene test was used to assess the equality of variances. An unpaired t-test was used to analyze the difference between the two groups. An analysis of variance (ANOVA) was performed with Fisher’s LSD as a post hoc test. If equal variances were rejected, the Games-Howell method was used as a post hoc test. In multi-way ANOVA, Mauchly’s sphericity test was performed and if sphericity was rejected, the degrees of freedom were corrected with ε calculated by Greenhouse-Geisser method (SPSS ver.26, IBM, Chicago, IL, USA). All results were expressed as the mean ± standard error of mean. Statistical significance was taken at P ± 0.05.
3. Results
3.1.1. Decoy inhibiting the release of LDH.
Positive controls were set at 100%. Decoy did not affect cultured rNSCh in terms of cell viability or PG synthesis. Decoy did not show a cytotoxic effect based on the LDH assay (Decoy of 16.5 ± 3.4% vs. control of 19.8 ± 2.0%, P = 0.44, Figure 2).
3.1.2. Decoy not affecting PG synthesis.
For PG synthesis assay, compared with the control, IL-1 administration had a >30% reduction (P <0.0001,), whereas the 10 μM Decoy group did not inhibit PG synthesis ( P = 0.263). Compared with IL-1, Decoy did not inhibit the attenuation of PG synthesis by IL-1 (IL-1+ 1 μM Decoy, P = 0.430, IL-1+ 10 μM Decoy, P = 0.132) (Figure 3).
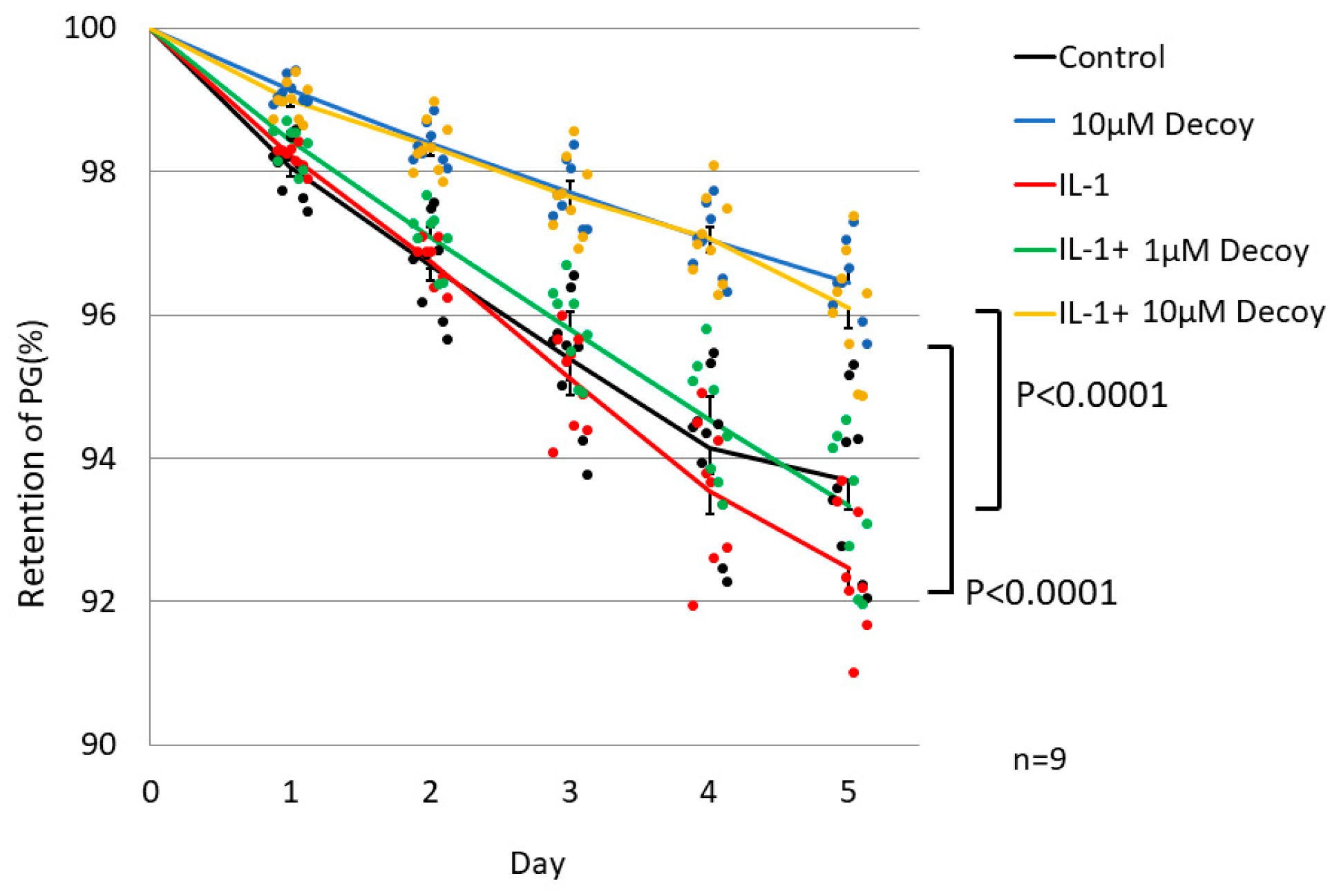
3.1.3. 10 uM Decoy inhibiting PG degradation in rNSCh.
Compared with controls, the 10 μM decoy group showed significant inhibition of PG degradation in the PG turnover assay (10 μM decoy group on day 5: 96.45 ± 0.53% and control on day 5: 93.69 ± 1.1%, P <0.0001). Although PG retention in IL-1 group was lower than the control group, there was no significant difference (IL-1 on day 5: 92.45 ± 0.85%, P = 0.821). In the presence of IL -1, Decoy at 1 μM concentration did not inhibit PG degradation (IL -1+ 1 μM Decoy on day 5: 93.33 ± 0.94%, P = 0.394), while the 10µM concentration of Decoy inhibited degradation of PG significantly (IL -1+ 10 μM Decoy on day 5: 96.1 ± 0.8%, P <0.0001) (Figure 4).
Figure 2.
Cytotoxicity (LDH) assay in rNSCh. Cytotoxicity of Decoy was not demonstrated in the LDH assay (P = 0.44). The plotted values are indicated as mean ± standard error of the mean. (n = 3, biological replicates ). N.S, not significant.
Figure 2.
Cytotoxicity (LDH) assay in rNSCh. Cytotoxicity of Decoy was not demonstrated in the LDH assay (P = 0.44). The plotted values are indicated as mean ± standard error of the mean. (n = 3, biological replicates ). N.S, not significant.
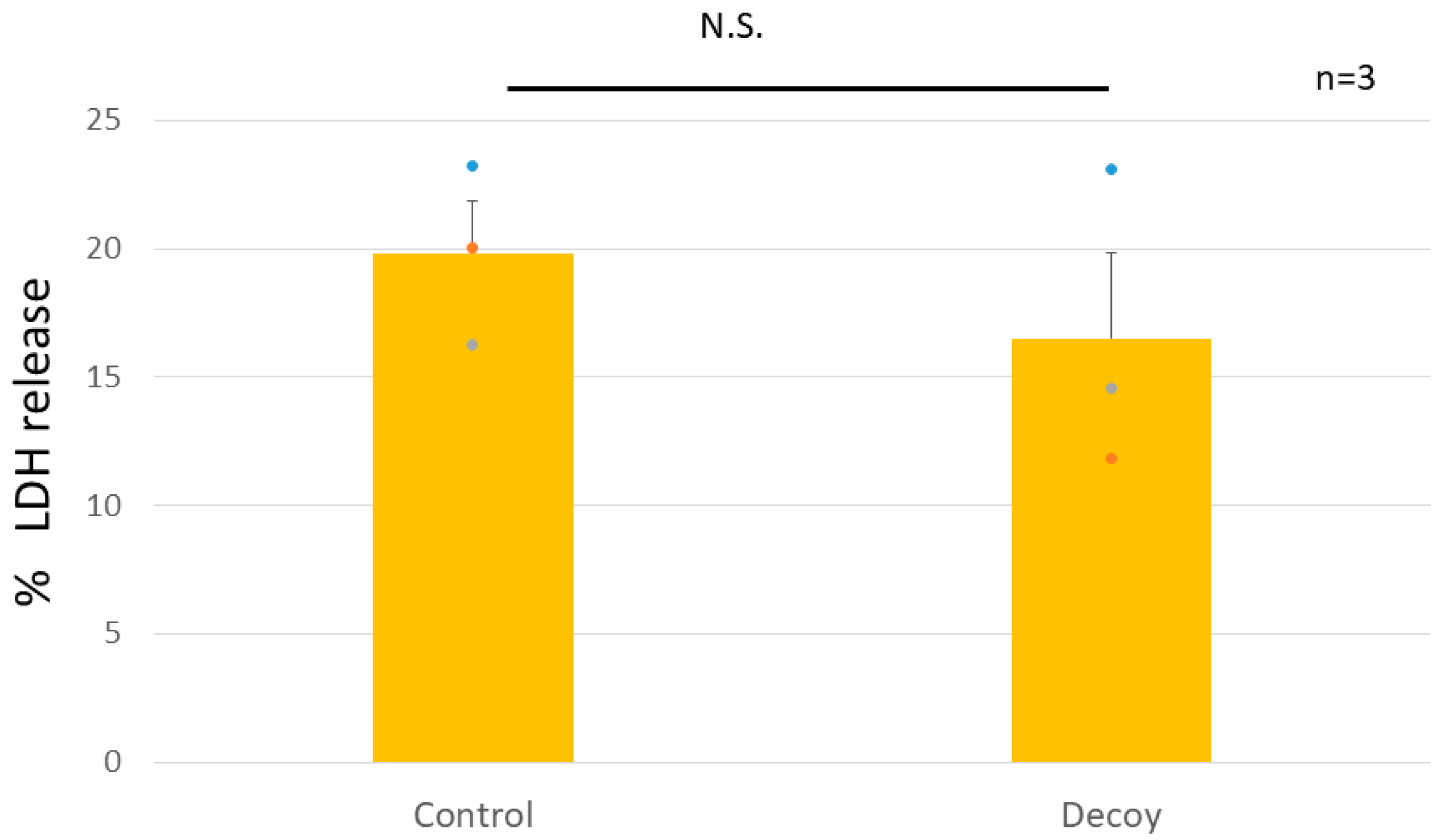
Figure 3.
PG synthesis assay in rNSCh. IL-1 treatment significantly decreased PG synthesis (P <0.0001). The 10 μM Decoy group did not inhibit PG synthesis. Decoy did not inhibit the attenuation of PG synthesis by IL-1. Data are expressed as the mean ± standard error of the mean (n = 9, 3 batches, in triplicate). One-way ANOVA with Games-Howell as a post-hoc test was used.
Figure 3.
PG synthesis assay in rNSCh. IL-1 treatment significantly decreased PG synthesis (P <0.0001). The 10 μM Decoy group did not inhibit PG synthesis. Decoy did not inhibit the attenuation of PG synthesis by IL-1. Data are expressed as the mean ± standard error of the mean (n = 9, 3 batches, in triplicate). One-way ANOVA with Games-Howell as a post-hoc test was used.
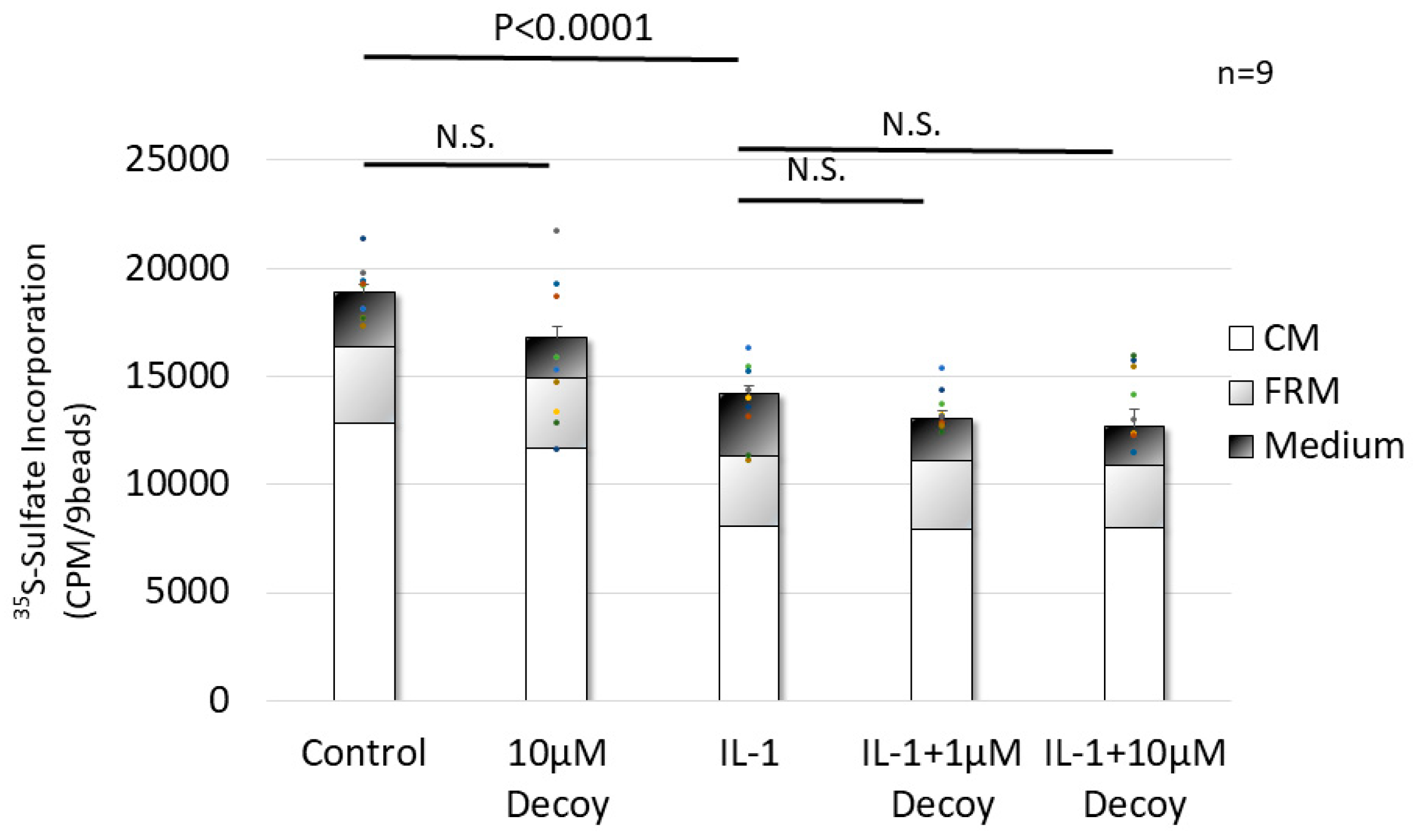
Figure 4.
PG turnover assay in rNSCh. Compared with controls, the 10 μM decoy group showed significant inhibition of PG degradation (P <0.0001). Although PG retention in IL-1 group was lower than the control group, there was no significant difference. In the presence of IL -1, Decoy at 1 μM did not inhibit PG degradation, while 10µM Decoy inhibited degradation of PG significantly (P <0.0001). The line in the graph shows the average for each group. Data are expressed as the mean ± standard error of the mean (n = 9, 3 batches, in triplicate). Two-way repeated ANOVA with Games-Howell as a post-hoc test was used.
Figure 4.
PG turnover assay in rNSCh. Compared with controls, the 10 μM decoy group showed significant inhibition of PG degradation (P <0.0001). Although PG retention in IL-1 group was lower than the control group, there was no significant difference. In the presence of IL -1, Decoy at 1 μM did not inhibit PG degradation, while 10µM Decoy inhibited degradation of PG significantly (P <0.0001). The line in the graph shows the average for each group. Data are expressed as the mean ± standard error of the mean (n = 9, 3 batches, in triplicate). Two-way repeated ANOVA with Games-Howell as a post-hoc test was used.
3.2.1. Decoy transfected into viable cells.
The transfection of FITC-labeled Decoy to the chondrocytes was identifiable 4 hours after initial administration (Figure 5a). The Live-Dead assay demonstrated that most of the cells in the tissue were live cells (Figure 5b, c). From these images, it was inferred that Decoy was incorporated into viable cells.
3.2.2. 10uM Decoy inhibiting PG degradation in hNSC tissue culture.
PG turnover was affected both by individual patient differences (P <0.001), treatment condition (P <0.001) and duration of culture (P <0.001), with an interactive effect (P = 0.001, Figure 6). Decoy significantly inhibited degradation of PG with or without IL-1. On day 5 of culture, the retention of PG for the control group (73.6 ± 7.3%) was increased by Decoy (to 83.6 ± 2.5%, P <0.01), and that for the IL-1 (73.9 ± 7.1%) was also increased for IL-1+Decoy (82.5 ± 2.5%, P <0.01). The half-life of PG was prolonged 63% by addition of Decoy under both conditions (control: 11.5 days, Decoy: 19.3 days, IL-1 11.5 days, and IL-1+ Decoy: 18.7 days). There was no detectable difference between the control group and the IL-1 group (P = 1.00).
3.2.3. Decoy inhibiting MMP3 production (ELISA and Nitrate Assay)
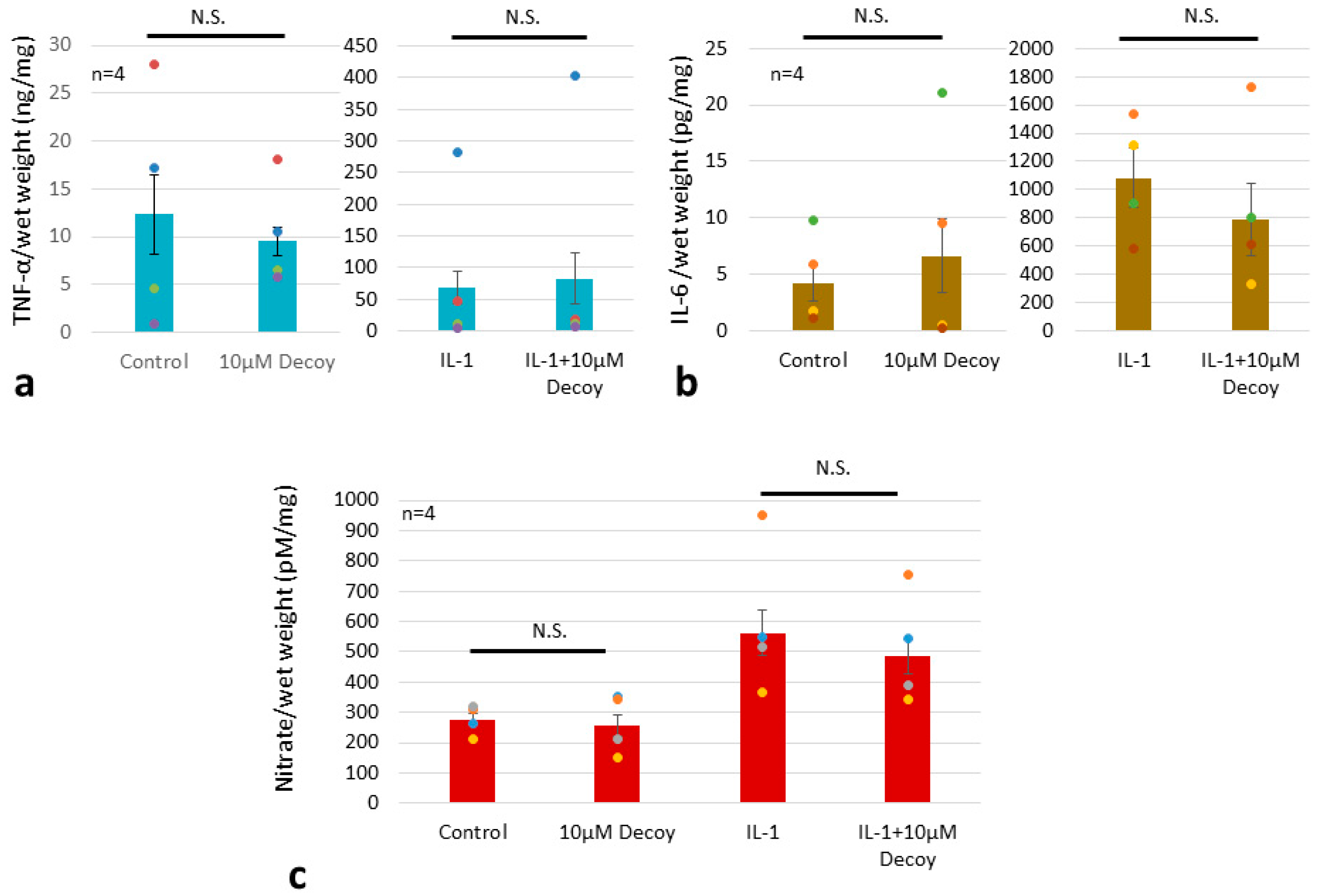
MMP3 was decreased by more than 70% with administration of Decoy compared to controls (P <0.05). IL-1 treatment increased MMP3 to 301.3% of controls (P <0.001). In the presence of IL-1, MMP3 was also significantly reduced by 51.1% with Decoy administration (P <0.001). (Figure 7). Decoy did not inhibit the other inflammation makers, TNF,IL-6, or NO (Figure 8a, b, c).
4. Discussion
NFκB was originally discovered as a lymphoid specific protein that binds to the decameric oligodeoxynucleotide GGGACTTCC present in the intronic enhancer element of the immunoglobulin κ light chain (Iκ) gene [14]. NFκB is normally present in the cytoplasm in an inactivated state by binding inhibitor proteins including IκB. [15,42]. IκB is phosphorylated and undergoes degradation through Multi-subunit IκB kinase (IKK) activation by many kinds of stimulation, including cytokines such as TNF and IL-1, chemokines, and stress [11]. Free NFκB rapidly enters the nucleus and transactivates target genes such as TNF, IL-1, and IL-6 [15,22]. The possibility of a therapeutic agent which uses dsDNA as a “decoy” with a high affinity to targeted transcription factors was first described by Sullenger et al and Bielinska et al [43,44]. Although antisense ODN has the ability to block target genes, decoy ODN is more effective in blocking constitutively expressed factors, as well as multiple transcription factors that bind to the same cis element [13]. The effect of NFκB decoy ODN on reducing the extent of myocardial infarction following reperfusion in a rat model via inhibition of IL-6 and intercellular cell adhesion molecule (ICAM) was first demonstrated in vivo by Morishita et al [35]. Consequently, it was realized that NFκB decoy ODN was not only effective against ischemia and reperfusion injury, but it also had anti-tumor and anti-inflammatory effects [17,18,19,20,21,22,23,45]. In anticipation of this anti-inflammatory effect, a phase 1b clinical trial of Decoy for chronic discogenic lumbar back pain was initiated in the United States in February 2018 [46]. A chimeric decoy ODN, which inhibits both NFκB and the allergic mediator signal transducer and activator of transcription 6 (STAT6), was also developed and used in a mouse model for asthma. It has been reported that chimeric decoy ODN has greater anti-inflammatory effects than decoy ODN with NFκB alone in the mouse asthma model [47].
Methods for quantifying PG include dye-binding methods such as dimethyl methylene blue (DMB), Western blotting, and chromatography have limitations [40,48]. In addition, these methods detect the content of PG, which reflect the results of catabolic and anabolic activities. Our study used the radioactive pre-labeled method followed by the chase (pulse-chase method) which is a gold standard to identify the rate of PG degradation to separate from the anabolic activity [49]. This method is superior to the gene expression analysis, PG content detection and Western blot methods in order to identify the net catabolic status of tissues [24,25,26,27,28,29,30] although the specific enzymes involved cannot be revealed without the newly cleaved epitope analysis.
Major enzymes that degrade ECM of cartilage, such as collagenase, originate from the MMP family as well as disintegrin and metalloproteinase with the thrombospondin motifs (ADAMTS) family. Twenty-eight kinds of MMP have been found so far; MMP-1, -3 and -13 play a central role [50]. MMP-3 can degrade broad spectrum ECM components including collagen, PG, laminins, and fibronectin [51]. The MMP varieties do not exist normally. Their protein synthesis can be initiated following gene transactivation by stimulation of cytkines such as IL-1 and TNF. This newly synthesized MMP remains inactivated unless a serine protease (SP) such as plasmin or trypsin is present. This study revealed significant inhibition of PG degradation of hNSC by Decoy. Despite the significant difference between the control and IL-1 stimulation groups in the protein level of MMP3, there was no significant difference in PG turnover assay in this study. Several factors explain this phenomenon. Firstly, there are influences from the other matrix degradation enzymes which may impact the results (e.g. from other MMP and ADAMTS families). Secondly, pre-upregulation of IL-1 is possible. In one of the specimens, IL-1 may have been upregulated due to previous tissue trauma (data not shown). There may be an association to what is observed with articular cartilage, which occasionally responds poorly to IL-1 stimulation [52]. Cawston et al [53] reported that there was no significant difference in collagen degradation in bovine nasal cartilage and human articular cartilage with or without IL-1. Thirdly, there may be limitations of our model using in vitro conditions. The concentration (10 ng/ml) of SP needs to be higher for in vitro conditions in order to activate MMP [54]. For this reason, the difference noted between the control and IL-1 group in MMP3 protein level did not affect PG turnover. Decoy also inhibited PG degradation in the PG turn over assay of rNSCh even in the absence of IL-1 stimulation. These results suggest that NFκB is involved in PG metabolism (balance between synthesis and degradation) in rNSCh under normal conditions, and that Decoy inhibited NFκB. Decoy did not suppress PG synthesis, and the PG reduction rate was slower by Decoy, suggesting that Decoy does not completely stop metabolism.
Although gene expression of the inflammatory markers (TNF, NO, and IL-6) and enzymes (the MMP and ADAMTS families) have been reported to be attenuated in human intervertebral disc (IVD) cell by Decoy [55], we found that the protein levels of these inflammatory markers were not attenuated by Decoy. Tissue specimens from two patients showed that Decoy inhibited these inflammatory markers (Figure 8), but the rest of the specimens did not. These results may vary by cell type, individual, and presence of extracellular matrix.
In future work, it would be necessary to study the behavior of other inflammation markers and enzymes with the use of Decoy, how differently Decoy interacts between tissues (IVD and NSC, etc.), and whether any variations in the cartilage specimens would make an impact on outcome. Glucocorticoids (GCC) have anti-inflammatory effects by inhibiting NFκB activation [56], but GCC is not suitable for cartilage transplantation because of its side effect of immunosuppression. In addition, GCC inhibits ECM synthesis in human chondrocytes as well [57]. Compared with decoy (10 μM), dexamethasone (1 μM) significantly inhibited PG synthesis in human IVD annuls fibrosus cells in our other experiments (data not shown). In the present study, Decoy showed no attenuation of PG synthesis and was highly effective for PG degradation, suggesting that Decoy may be useful in protecting transplanted cartilage for example, it could be added to implants through pre-incubation prior to implantation or used as an external protective material such as a gel.
5. Conclusion
The integrity and structural form of cartilage grafts which are used for reconstructive purposes can be compromised by degrees of graft resorption, warping from localized biomechanical forces, insufficient nutrition from recipient cells, and the inflammatory process. To curb the deleterious effect of the inflammatory process, this study was conducted to explore the potential of Decoy as a chondroprotective agent for cartilage grafts used for reconstruction. The study results demonstrate significant inhibition of PG decomposition by suppressing MMP3 production in an hNSC explant culture system (cartilage transplantation model). Consequently, it appears that Decoy can maintain the ECM of cartilage and may be considered a novel chondroprotective agent in the cellular environment of inflammation which occurs after cartilage transplantation. Methods of application of Decoy into the wound bed are reserved for further studies in this work.
Author Contributions
H.N. was responsible for the study design, data collection, analysis, and evaluation, and the drafting of this manuscript. D.S. contributed to data acquisition and analysis. D.W. contributed to the study idea, harvesting of the tissue, and manuscript editing. K.M. contributed to the evaluation of the study design, interpretation of the data, and supervised this project.
Acknowledgments
NFκB decoy oligodeoxynucleotide was provided by AnGes Inc (Osaka, Japan). In remembrance, Mary Ellen Lenz, M.S. (1945 – 2022) who contributed to our work.
References
- Fu, Y.; He, A.; Xie, Y.; Zhu, Y.; Li, C.; Zhang, T. The Effect of Fixation Materials on the Long-Term Stability of Cartilage Framework for Microtia Reconstruction. Facial Plast. Surg. Aesthetic Med. 2023, 25, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, X.; Deng, Y. Complications Associated with Autologous Costal Cartilage Used in Rhinoplasty: An Updated Meta-Analysis. Aesthetic Plast. Surg. 2022, 47, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Cervelli, V. Nasal Tip Remodeling Using Autologous Cartilage Grafts: Systematic Review. J. Craniofacial Surg. 2022, 33, 2035–2040. [Google Scholar] [CrossRef] [PubMed]
- Brent, B. The Versatile Cartilage Autograft: Current Trends in Clinical Transplantation. Clin. Plast. Surg. 1979, 6, 163–180. [Google Scholar] [CrossRef] [PubMed]
- De Gabory, L.; Fricain, J.C.; Stoll, D. [Resorption of cartilage grafts in rhinoplasty: fundamental basis]. Revue de laryngologie - otologie - rhinologie 2010, 131, 83–88. [Google Scholar] [PubMed]
- Bujía, J. Determination of the viability of crushed cartilage grafts: clinical implications for wound healing in nasal surgery. Ann. Plast. Surg. 1994, 32, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Hom, D. The wound healing response to grafted tissues. Otolaryngol. Clin. N. Am. 1994, 27, 13–24. [Google Scholar] [CrossRef]
- Janis, J.E.; Harrison, B. Wound Healing: Part I. Basic Science. Plast. Reconstr. Surg. 2014, 133, 199e–207e. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hsu, S.; Chang, M.; Huang, Y. Reducing scar formation by regulation of IL-1 and MMP-9 expression by using sustained release of prednisolone-loaded PDLL microspheres in a murine wound model. J. Biomed. Mater. Res. Part A 2013, 101, 1165–1172. [Google Scholar] [CrossRef]
- Haisch, A.; Gröger, A.; Radke, C.; Ebmeyer, J.; Sudhoff, H.; Grasnick, G.; Jahnke, V.; Burmester, G.; Sittinger, M. Macroencapsulation of human cartilage implants: pilot study with polyelectrolyte complex membrane encapsulation. Biomaterials 2000, 21, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A. S., Gedin, P., Hugo, A., et al. Activation of NF-κB in Synovium versus Cartilage from Patients with Advanced Knee Osteoarthritis: A Potential Contributor to Inflammatory Aspects of Disease Progression. Journal of immunology (Baltimore, Md : 1950) 2018;201:1918-1927. [CrossRef]
- Yamasaki, K., Asai, T., Shimizu, M., et al. Inhibition of NFκB activation using cis-element ‘decoy’of NFκB binding site reduces neointimal formation in porcine balloon-injured coronary artery model. Gene therapy 2003;10:356-364. [CrossRef]
- Morishita, R.; Tomita, N.; Kaneda, Y.; Ogihara, T. Molecular therapy to inhibit NFκB activation by transcription factor decoy oligonucleotides. Curr. Opin. Pharmacol. 2004, 4, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Baltimore, D. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell 1986, 46, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T. D. The Rel/NF-kappaB signal transduction pathway: introduction. Oncogene 1999;18:6842. [CrossRef]
- Kato, K., Akeda, K., Miyazaki, S., et al. NF-kB decoy oligodeoxynucleotide preserves disc height in a rabbit anular-puncture model and reduces pain induction in a rat xenograft-radiculopathy model. European cells & materials 2021;42:90-109. [CrossRef]
- Nishimura, A., Akeda, K., Matsubara, T., et al. Transfection of NF-κB decoy oligodeoxynucleotide suppresses pulmonary metastasis by murine osteosarcoma. Cancer gene therapy 2011;18:250-259. [CrossRef]
- Cao, C. C., Ding, X. Q., Ou, Z. L., et al. In vivo transfection of NF-κB decoy oligodeoxynucleotides attenuate renal ischemia/reperfusion injury in rats. Kidney international 2004;65:834-845. [CrossRef]
- Xu, M.-Q., Shuai, X.-R., Yan, M.-L., Zhang, M.-M., Yan, L.-N. Nuclear factor-kappaB decoy oligodeoxynucleotides attenuates ischemia/reperfusion injury in rat liver graft. World Journal of Gastroenterology 2005;11:6960. [CrossRef]
- Tomita, T.; Takano, H.; Tomita, N.; Morishita, R.; Kaneko, M.; Shi, K.; Takahi, K.; Nakase, T.; Kaneda, Y.; Yoshikawa, H.; et al. Transcription factor decoy for NFκB inhibits cytokine and adhesion molecule expressions in synovial cells derived from rheumatoid arthritis. Rheumatology 2000, 39, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Abeyama, K., Eng, W., Jester, J. V., et al. A role for NF-κB–dependent gene transactivation in sunburn. Journal of Clinical Investigation 2000;105:1751-1759.
- Uemura, T.; Tsujii, M.; Akeda, K.; Iino, T.; Satonaka, H.; Hasegawa, M.; Sudo, A. Transfection of nuclear factor-kappaB decoy oligodeoxynucleotide protects against ischemia/reperfusion injury in a rat epigastric flap model. J. Gene Med. 2012, 14, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Tahara, K., Samura, S., Tsuji, K., et al. Oral nuclear factor-κB decoy oligonucleotides delivery system with chitosan modified poly (D, L-lactide-co-glycolide) nanospheres for inflammatory bowel disease. Biomaterials 2011;32:870-878. [CrossRef]
- Morales, T.I.; Hascall, V.C. Factors involved in the regulation of proteoglycan metabolism in articular cartilage. Arthritis Rheum. 1989, 32, 1197–1201. [Google Scholar] [CrossRef] [PubMed]
- Hascall, V.C.; Handley, C.J.; McQuillan, D.J.; Hascall, G.K.; Robinson, H.; Lowther, D.A. The effect of serum on biosynthesis of proteoglycans by bovine articular cartilage in culture. Arch. Biochem. Biophys. 1983, 224, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.A.; Handley, C.J.; Hascall, V.C.; Campbell, R.A.; Lowther, D.A. Turnover of proteoglycans in cultures of bovine articular cartilage. Arch. Biochem. Biophys. 1984, 234, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Luyten, F.P.; Hascall, V.C.; Nissley, S.; Morales, T.I.; Reddi, A. Insulin-like growth factors maintain steady-state metabolism of proteoglycans in bovine articular cartilage explants. Arch. Biochem. Biophys. 1988, 267, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Fell, H.B.; Jubb, R.W. The Effect of Synovial Tissue on the Breakdown of Articular Cartilage in Organ Culture. Arthritis Rheum. 1977, 20, 1359–1371. [Google Scholar] [CrossRef] [PubMed]
- Saklatvala, J.; Pilsworth, L.M.C.; Sarsfield, S.J.; Gavrilovic, J.; Heath, J.K. Pig catabolin is a form of interleukin 1. Cartilage and bone resorb, fibroblasts make prostaglandin and collagenase, and thymocyte proliferation is augmented in response to one protein. Biochem. J. 1984, 224, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Morales, T.I.; Hascall, V.C. Effects of Interleukin-1 and Lipopolysaccharides on Protein and Carbohydrate Metabolism in Bovine Articular Cartilage Organ Cultures. Connect. Tissue Res. 1989, 19, 255–275. [Google Scholar] [CrossRef] [PubMed]
- Jenei-Lanzl, Z., Meurer, A., Zaucke, F. Interleukin-1β signaling in osteoarthritis - chondrocytes in focus. Cellular signalling 2019;53:212-223. [CrossRef]
- Bonassar, L.J.; Frank, E.H.; Murray, J.C.; Paguio, C.G.; Moore, V.L.; Lark, M.W.; Sandy, J.D.; Wu, J.; Eyre, D.R.; Grodzinsky, A.J. Changes in cartilage composition and physical properties due to stromelysin degradation. Arthritis Rheum. 1995, 38, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Temple, M.M.; Xue, Y.; Chen, M.Q.; Sah, R.L. Interleukin-1alpha induction of tensile weakening associated with collagen degradation in bovine articular cartilage. Arthritis Rheum. 2006, 54, 3267–3276. [Google Scholar] [CrossRef] [PubMed]
- Furman, B.D.; Mangiapani, D.S.; Zeitler, E.; Bailey, K.N.; Horne, P.H.; Huebner, J.L.; Kraus, V.B.; Guilak, F.; A Olson, S. Targeting pro-inflammatory cytokines following joint injury: acute intra-articular inhibition of interleukin-1 following knee injury prevents post-traumatic arthritis. Arthritis Res. Ther. 2014, 16, R134. [Google Scholar] [CrossRef] [PubMed]
- Morishita, R., Sugimoto, T., Aoki, M., et al. In vivo transfection of cis element “decoy” against nuclear factor-κB binding site prevents myocardial infarction. Nature medicine 1997;3:894-899. [CrossRef]
- Sawa, Y.; Morishita, R.; Suzuki, K.; Kagisaki, K.; Kaneda, Y.; Maeda, K.; Kadoba, K.; Matsuda, H. A novel strategy for myocardial protection using in vivo transfection of cis element ’decoy’ against NFkappaB binding site: evidence for a role of NFkappaB in ischemia-reperfusion injury. Circulation 1997;96:II-280-284; discussion II-285.
- Yoshimura, S., Morishita, R., Hayashi, K., et al. Inhibition of intimal hyperplasia after balloon injury in rat carotid artery model using cis-element ‘decoy’of nuclear factor-kB binding site as a novel molecular strategy. Gene therapy 2001;8:1635-1642. [CrossRef]
- Chiba, K.; Andersson, G.B.J.; Masuda, K.; Thonar, E.J.-M.A. Metabolism of the Extracellular Matrix Formed by Intervertebral Disc Cells Cultured in Alginate. Spine 1997, 22, 2885–2893. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, D.; Yin, L.; Zhang, C.; Qu, H.; Xu, J. Neuroglobin protects against cerebral ischemia/reperfusion injury in rats by suppressing mitochondrial dysfunction and endoplasmic reticulum stress-mediated neuronal apoptosis through synaptotagmin-1. Environ. Toxicol. 2023, 38, 1891–1904. [Google Scholar] [CrossRef] [PubMed]
- Masuda, K.; Shirota, H.; Thonar, E. Quantification of 35S-Labeled Proteoglycans Complexed to Alcian Blue by Rapid Filtration in Multiwell Plates. Anal. Biochem. 1994, 217, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Mok, S.S.; Masuda, K.; Häuselmann, H.J.; Aydelotte, M.B.; Thonar, E.J. Aggrecan synthesized by mature bovine chondrocytes suspended in alginate. Identification of two distinct metabolic matrix pools. J. Biol. Chem. 1994, 269, 33021–33027. [Google Scholar] [CrossRef] [PubMed]
- Liu, T., Zhang, L., Joo, D., Sun, S. C. NF-κB signaling in inflammation. Signal transduction and targeted therapy 2017;2:17023. [CrossRef]
- Sullenger, B.A.; Gallardo, H.F.; Ungers, G.E.; Gilboa, E. Overexpression of TAR sequences renders cells resistant to human immunodeficiency virus replication. Cell 1990, 63, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Bielinska, A.; Shivdasani, R.A.; Zhang, L.; Nabel, G.J. Regulation of Gene Expression with Double-Stranded Phosphorothioate Oligonucleotides. Science 1990, 250, 997–1000. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, H., Ishida, Y., Hosomichi, J., et al. Ultrasound microbubble-mediated transfection of NF-κB decoy oligodeoxynucleotide into gingival tissues inhibits periodontitis in rats in vivo. PloS one 2017;12:e0186264. [CrossRef]
- NIH. AMG0103 in Subjects With Chronic Discogenic Lumbar Back Pain - Full Text View - ClinicalTrials.gov. Available at: https://clinicaltrials.gov/ct2/show/NCT03263611. Accessed 28, Jan. 2023.
- Miyake, T., Miyake, T., Sakaguchi, M., Nankai, H., Nakazawa, T., Morishita, R. Prevention of Asthma Exacerbation in a Mouse Model by Simultaneous Inhibition of NF-κB and STAT6 Activation Using a Chimeric Decoy Strategy. Molecular therapy Nucleic acids 2018;10:159-169. [CrossRef]
- Allaith, S.; Tew, S.; Hughes, C.; Clegg, P.; Canty-Laird, E.; Comerford, E. Characterisation of key proteoglycans in the cranial cruciate ligaments (CCLs) from two dog breeds with different predispositions to CCL disease and rupture. Veter- J. 2021, 272, 105657. [Google Scholar] [CrossRef] [PubMed]
- Hascall, V., Luyten, F., Plaas, A., Sandy, J. Steady-state metabo-lism of proteoglycans in bovine articular cartilage explants. In A. Maroudas, K. E. Kuettner eds., Methods in Cartilage Research London: Academic Press; 1990:108-112.
- Peng, Z.; Sun, H.; Bunpetch, V.; Koh, Y.; Wen, Y.; Wu, D.; Ouyang, H. The regulation of cartilage extracellular matrix homeostasis in joint cartilage degeneration and regeneration. Biomaterials 2021, 268, 120555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, C.; He, B.; Zhang, X.; Hao, H.; Hou, Y.; Li, A.; Wang, Y.; Wang, Y. Macrophage Migration Inhibitory Factor Promotes Expression of Matrix Metalloproteinases 1 and 3 in Spinal Cord Astrocytes following Gecko Tail Amputation. J. Integr. Neurosci. 2023, 22, 29. [Google Scholar] [CrossRef] [PubMed]
- Tyler, J., Bolis, S., Dingle, J., et al. Mediators of matrix metabolism. In K. Kuettner, R. Schleyerbach, J. Peyron eds., Articular Cartilage and Osteoarthritis. New York: Raven Press; 1992:251-264.
- Cawston, T., Curry, V., Summers, C., et al. The role of oncostatin M in animal and human connective tissue collagen turnover and its localization within the rheumatoid joint. Arthritis & Rheumatism 1998;41:1760-1771. [CrossRef]
- Okada, Y. Regulation of Matrix Metalloprotease (in Japanese). Jikken igaku 1992;10:256-262.
- Pichika, R., Akeda, K., Gemba, T., Miyamoto, K., An, H. S., Masuda, K. Transcription Factor Decoy For Nfkb Inhibits The Appearance Of Active Mmps And Adamts4 In The Medium Of Human Intervertebral Disc Cells Cultured In Alginate. Paper presented at: 2005 Annual Meeting of the Orthopaedic Research Society May 20-23; Chicago, Ill.
- Nelson, H. S., Leung, D. Y., Bloom, J. W. Update on glucocorticoid action and resistance. Journal of allergy and Clinical Immunology 2003;111:3-22. [CrossRef]
- Song, Y.W.; Zhang, T.; Wang, W.B. Gluococorticoid could influence extracellular matrix synthesis through Sox9 via p38 MAPK pathway. Rheumatol. Int. 2012, 32, 3669–3673. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanism of Decoy: a) NFκB is normally present in the cytoplasm in an inactivated state by binding inhibitor proteins including IκB. NFκB is normally present as an inactive, Igκ-bound complex in cytoplasm. Multi-subunit IκB kinase (IKK) is activated by stimuli such as IL-1 and TNF, and IκB is inducibly degraded by activated IKK. Free NFκB rapidly enters the nucleus and transactivates target genes such as IL-1, TNF, iNOs, MMPs, COX-2, and IL-6. (b) Under the presence of Decoy, transactivation will not begin because the activated NFκB is bound to Decoy. Abbreviations: iNOS (inducible nitric oxide synthase), MMPs (matrix metalloproteinases), COX-2 (cyclooxygenase-2).
Figure 1.
Mechanism of Decoy: a) NFκB is normally present in the cytoplasm in an inactivated state by binding inhibitor proteins including IκB. NFκB is normally present as an inactive, Igκ-bound complex in cytoplasm. Multi-subunit IκB kinase (IKK) is activated by stimuli such as IL-1 and TNF, and IκB is inducibly degraded by activated IKK. Free NFκB rapidly enters the nucleus and transactivates target genes such as IL-1, TNF, iNOs, MMPs, COX-2, and IL-6. (b) Under the presence of Decoy, transactivation will not begin because the activated NFκB is bound to Decoy. Abbreviations: iNOS (inducible nitric oxide synthase), MMPs (matrix metalloproteinases), COX-2 (cyclooxygenase-2).
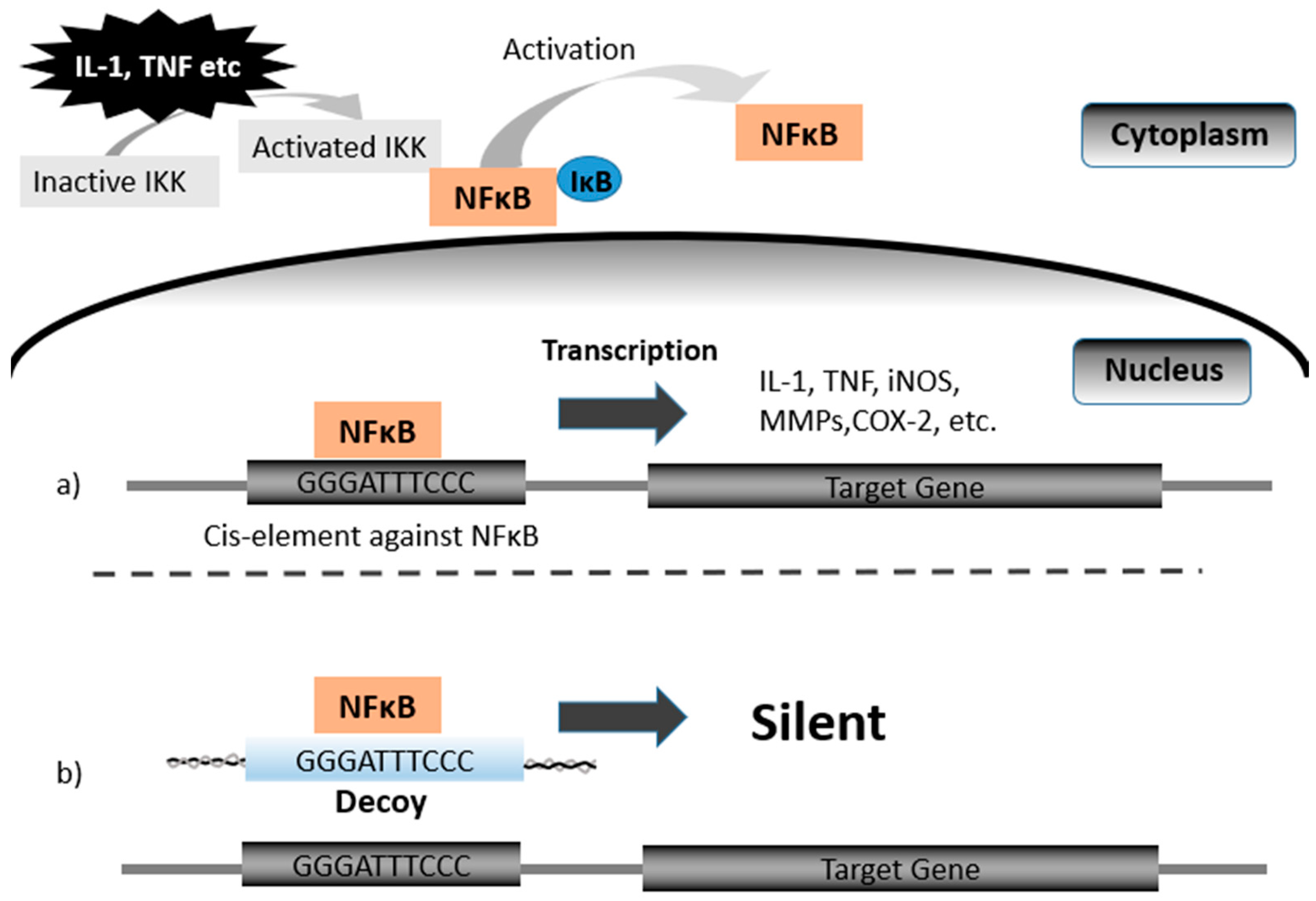
Figure 5.
Efficiency of transfection of NFκB-Decoy ODN. Live-Dead assay was separately performed using human nasal septal cartilage tissues. (a) Green indicates FITC-labeled Decoy 6h hours after the transfection of FITC decoy. (b and c) The separate tissues were subjected to Live/Dead assay. Green indicates viable cells. Red indicates dead cells. Most of cells were viable in the tissues. From these images, it was inferred that Decoy was incorporated into viable cells.
Figure 5.
Efficiency of transfection of NFκB-Decoy ODN. Live-Dead assay was separately performed using human nasal septal cartilage tissues. (a) Green indicates FITC-labeled Decoy 6h hours after the transfection of FITC decoy. (b and c) The separate tissues were subjected to Live/Dead assay. Green indicates viable cells. Red indicates dead cells. Most of cells were viable in the tissues. From these images, it was inferred that Decoy was incorporated into viable cells.
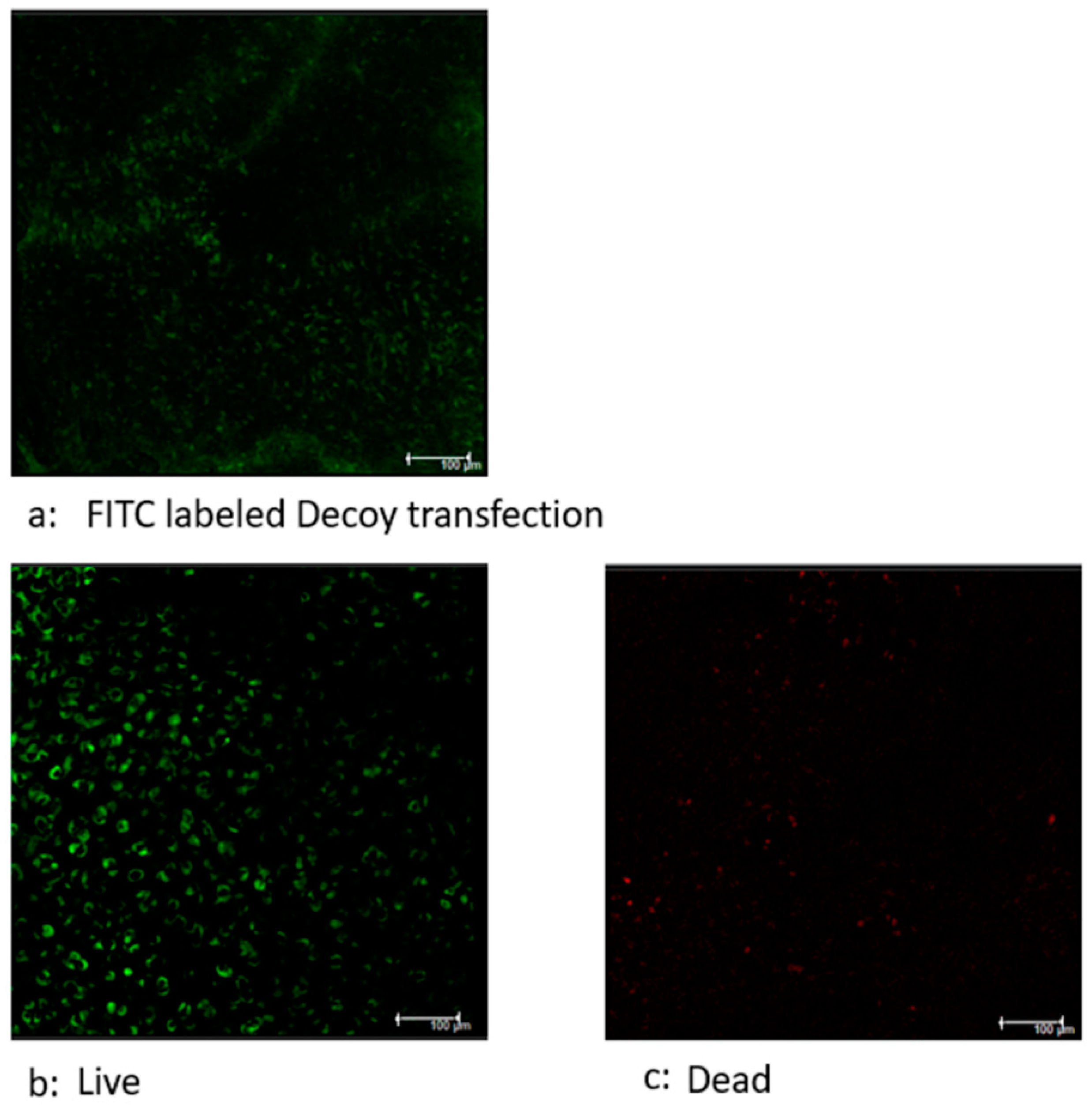
Figure 6.
PG turnover assay in tissue culture. Decoy significantly inhibited degradation of PG with or without IL-1 (P <0.01, P <0.01). There was no significant difference between control group and IL-1 group. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Three-way repeated ANOVA with Games-Howell as a post-hoc test was used.
Figure 6.
PG turnover assay in tissue culture. Decoy significantly inhibited degradation of PG with or without IL-1 (P <0.01, P <0.01). There was no significant difference between control group and IL-1 group. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Three-way repeated ANOVA with Games-Howell as a post-hoc test was used.
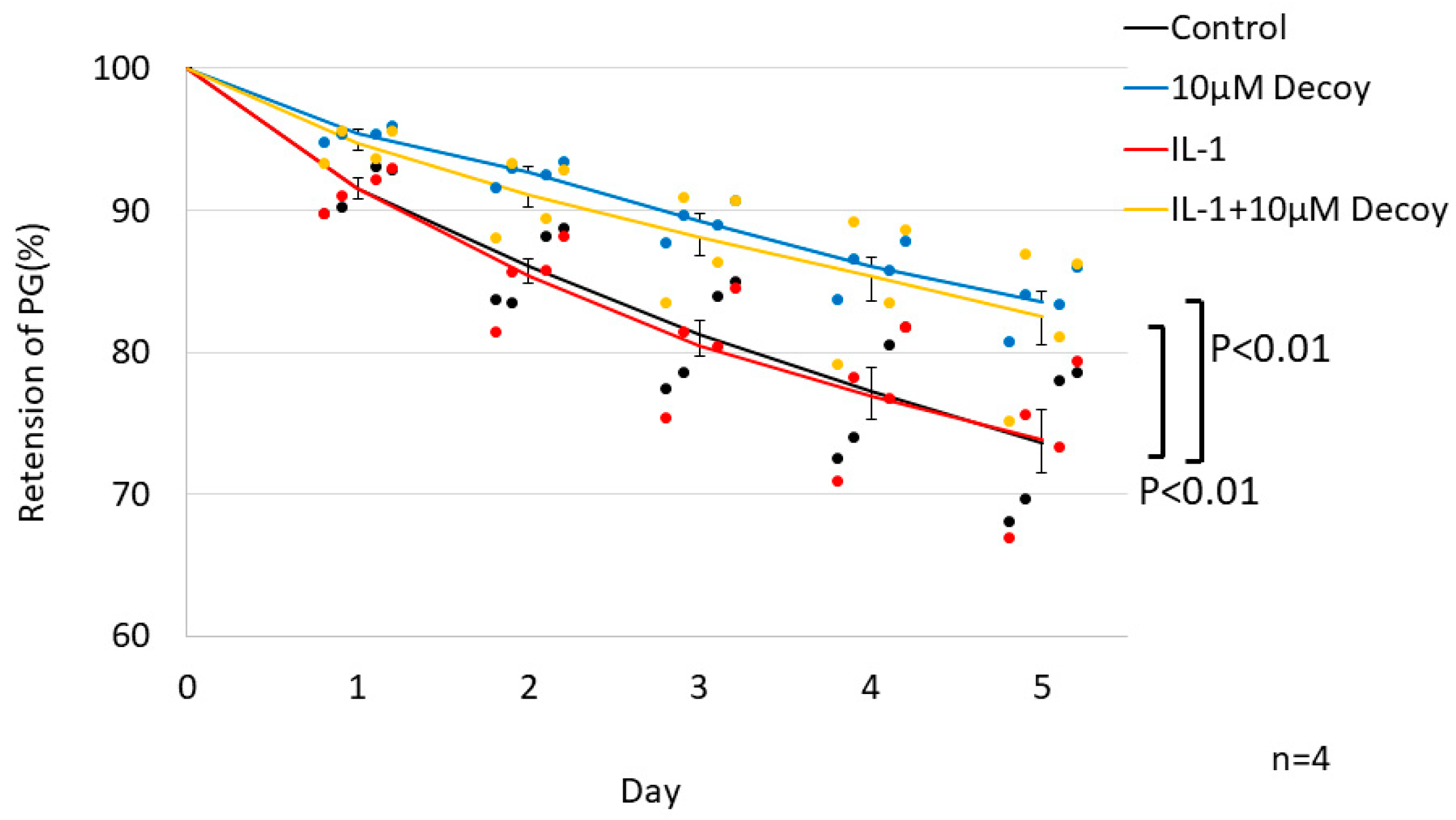
Figure 7.
ELISA MMP3. MMP3 was decreased by more than 70% with administration of Decoy compared to controls (P <0.05). IL-1 treatment increased MMP3 to 301.3% of controls (P <0.001). In the presence of IL-1, MMP3 was also significantly reduced by 51.1% with Decoy administration (P <0.001). Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Games-Howell as a post-hoc test was used.
Figure 7.
ELISA MMP3. MMP3 was decreased by more than 70% with administration of Decoy compared to controls (P <0.05). IL-1 treatment increased MMP3 to 301.3% of controls (P <0.001). In the presence of IL-1, MMP3 was also significantly reduced by 51.1% with Decoy administration (P <0.001). Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Games-Howell as a post-hoc test was used.
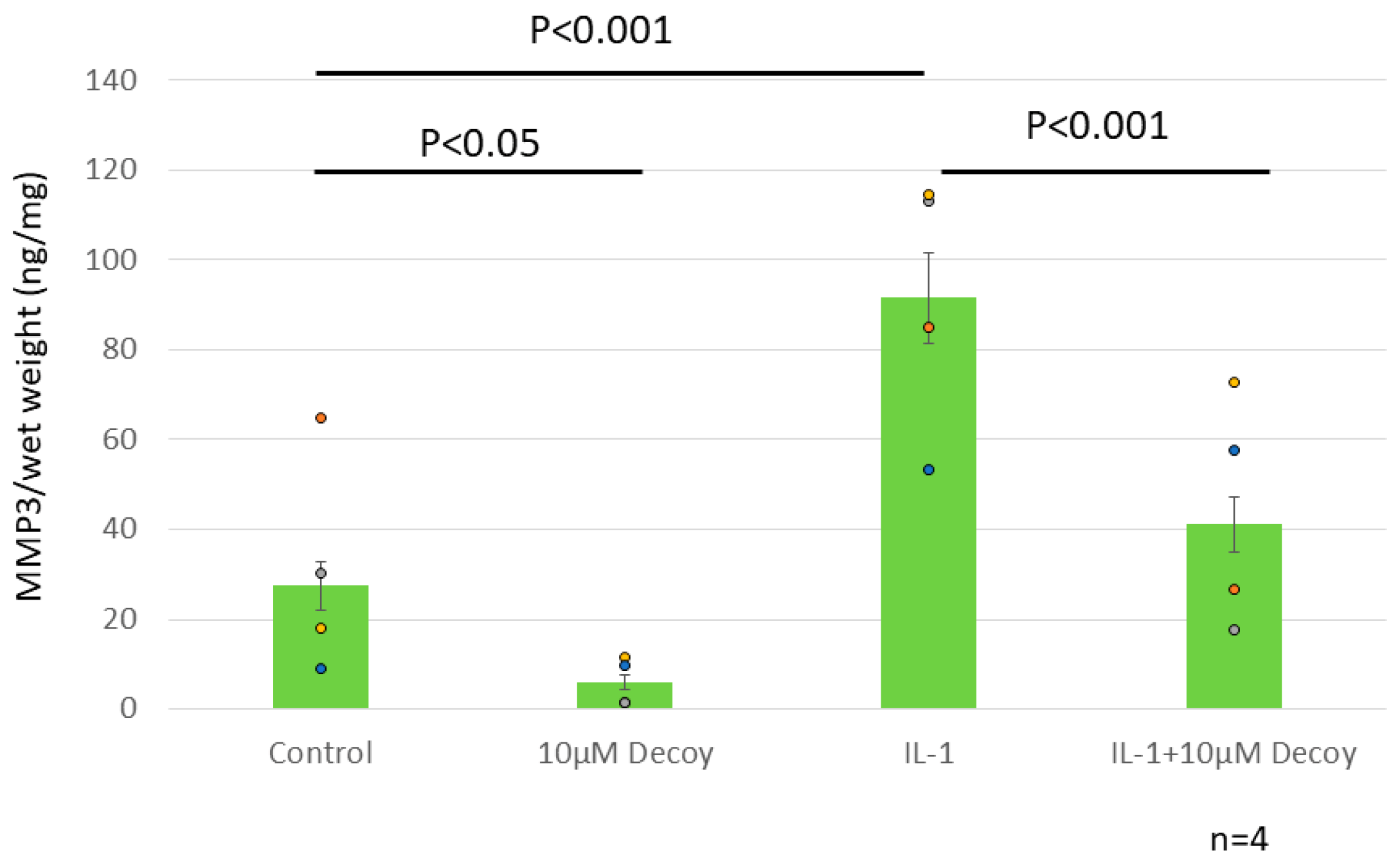
Figure 8.
ELISA TNF, IL-6 and Nitrate assay. (a) ELISA TNF. Decoy did not inhibit TNF. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Games-Howell as a post-hoc test was used. ELISA IL-6. (b) Decoy did not inhibit IL-6. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Games-Howell as a post-hoc test was used. (c) Nitrate assay. Decoy did not inhibit Nitrate. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Fisher’s LSD as a post-hoc test was used.
Figure 8.
ELISA TNF, IL-6 and Nitrate assay. (a) ELISA TNF. Decoy did not inhibit TNF. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Games-Howell as a post-hoc test was used. ELISA IL-6. (b) Decoy did not inhibit IL-6. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Games-Howell as a post-hoc test was used. (c) Nitrate assay. Decoy did not inhibit Nitrate. Data are expressed as the mean ± standard error of the mean (n=4 patients, in triplicate). Two-way ANOVA with Fisher’s LSD as a post-hoc test was used.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



