
Submitted:
26 October 2023
Posted:
27 October 2023
You are already at the latest version
Abstract
The study aimed to isolate and characterize bacteriocin-producing Enterococcus species from Ethiopian traditional fermented foods and beverages. The bacteriocin bioassay was tested against food-borne pathogens at various temperatures, pH, solvents, additives, and enzymes. A solvent extraction method was established to concentrate bacteriocins from Enterococcus species. The study found that crude extracts with proteinase K, pepsin, and trypsin deactivated antimicrobial activity, confirming their proteinaceous nature. Organic solvents were more effective at extracting bacteriocins from CFS, with chloroform alone and in combination providing rapid and efficient recovery. The optimal bacteriocin production and strong inhibition activity were achieved at 37°C with a pH range of 5.5 to 6.5 incubated for 16-18 hours. The six Enterococcus faecium species were the best bacteriocin-producing isolates. Optimizing bacteriocin production and changing environmental growing circumstances can enhance the application of bacteriocins as food preservation agents. Traditional fermented products are potential sources of bacteriocin-producing LAB.
Keywords:
antimicrobial activity
; bacteriocin producing lactic acid bacteria
; bacteriocins
; Ethiopian
; solvent extraction and traditional fermented products
1. INTRODCUTION
Foodborne diseases (FBDs) have been a comprehensive concern; their prevention and control require multidisciplinary approaches that involve search for human beneficial natural products like probiotics in order to combat pathogens and their associated health risks (Sivapalasingam et al., 2004). Despite the use of modern food preservation methods, the rate of food-related illness continues to rise and is a major cause of death, particularly in nations with inadequate systems in place to monitor food safety (Amit et al., 2017). Due to the consumption of contaminated foods including contaminated canned food, meat, poultry, and fermented dairy products, more than 75% of the world's population currently suffers from food-related diseases (Akhtar et al., 2013). As a result, food safety is a current global concern that has an immediate link to human health and is gravely threatened by pathogenic microbes during transportation, storage, and production time (Parada & Aguilera, 2007).
Furthermore, due to consumers' preferences towards minimally processed foods with no additives or chemical preservatives, the world is shifting its attention toward the application of natural preservatives, including bacteriocin and bacteriocin producing bacteria, as promising natural food preservatives (Molognoni et al., 2019; Santos et al., 2018; J. Yang et al., 2014). Studies on bacteriocins producing lactic acid bacteria (LAB) were widely investigated due to their potential application as bio-preservatives in the food industry as many strains have been given “generally recognized as safe” (GRAS) status (O’Sullivan et al., 2002). The preservative effect of LAB is due to the production of antimicrobial substances, including organic acids, hydrogen peroxide, diacetyl, bacteriocins, and bacteriocin-like antimicrobial substances (Freitas et al., 2015; Sanlibaba et al., 2015). Among these antimicrobial components, bacteriocins have received more attention in recent years because of their potential application in the food industry as natural preservatives targeting multi-drug-resistant pathogens (J. Yang et al., 2014).
The most dominant lactic acid bacteria that produce bacteriocins and bacteriocins like substances, including Lactococcus lactis, Pediococcus pentosaceus and Enterococcus faecium, were isolated and characterized from various traditional fermented foods and beverages. Some species of bacteriocin-producing Enterococcus isolated from various traditional fermented foods and beverages showed potential antagonistic activities against L. Monocytogenes, B. cereus and S. aureus, including multidrug resistant strains such as methicillin-resistant S. aureus. Enterococcus faecium isolated from traditional fermented dairy products are reported to have potential antagonistic activity against both gram-positive and gram-negative bacterial pathogens (do Nascimento et al., 2010; Fugaban et al., 2021; Vimont et al., 2017).
Although many Ethiopian traditional fermented foods, condiments, and beverages potentially contain probiotic Enterococcus species capable of producing various antimicrobial substances of industrial use, there are no research reports undertaken focusing on isolation and characterization of bacteriocin producing Enterococcus species. Hence, it is necessary to isolate, screen and characterize bacteriocins producing Enterococcus species to determine optimal growth conditions for bacteriocin production and application the product as bio-preservative (Santos et al., 2018; Weaver & Lawton, 2007). Bacteriocins-producing capability of Enterococcus species made them important not only for food preservation but also for treating certain multi drug resistant pathogens. Therefore, this study was conducted to isolate, screen and characterize bacitracin producing Enterococcus faecium from Ethiopian traditional fermented foods and beverages; and to characterize their antimicrobial substances Produced by E. faecium isolates.
2. MATERIAL AND METHODS
2.1. Bacterial growth conditions
The bacteria used in this research are among the LAB isolated in previous study (Table 1). They were found in traditionally fermented foods and drinks from Ethiopia, and they may be probiotic LAB. In MRS agar, all probiotic isolates were grown. Additionally, the Ethiopian Public Health Institute (EPHI), in Addis Abeba, Ethiopia, provided some food-borne pathogens (Table 1).
2.2. Preliminary screening of bacteriocins producers
2.2.1. Agar Well Diffusion method
Bacteriocin activity was confirmed through an agar well diffusion test. Bacteriocin producer cells were grown in MRS broth for 18 hours at 37°C. After centrifugation, cell-free supernatant was added to indicator bacteria on seeded plates. The zone of inhibition around the well was considered a positive sample (Goh & Philip, 2015).
2.2.2. Antagonistic activity of crude bacteriocins
Enterococcus species were inoculated in MRS media and incubated at 37°C for 16-18 hours. The cell-free supernatant was collected, and the pH was adjusted to 6.5-7.0 with 1N NaOH to neutralize acids and remove inhibition activity. Catalase enzyme was used to rule out hydrogen peroxide inhibitory activity. Crude bacteriocins were tested against food-borne pathogens on Muller Hinton Agar plates. A 24 hour-old broth culture was swabbed and the cultural supernatant was placed in wells and incubated at 37°C for 24 hours. The zone of inhibition was measured and recorded in millimeters after complete incubation (Erb et al., 2006; Ogunbanwo et al., 2003).
2.3. Evaluating bacteriocins extraction methods
2.3.1. Convectional bacteriocins extraction
To extract bacteriocins, 200 ml of LAB culture was adjusted to pH 6.5 with a 1 M NaOH solution, stirred for 30 minutes, and heated at 70 °C. LAB cells were harvested, washed twice, and re-suspended in 50 mL of 100 mM NaCl at pH 2.0. The cell suspension was centrifuged, and the cell-free supernatant was adjusted to pH 6.5 with a 1 M NaOH solution. The bacteriocin extract was filtrated through a 0.2-m pore-size cellulose acetate, and their bioactivity against selected human pathogens was tested (Wray et al., 2018).
2.3.2. Ammonium sulphate precipitation.
The cell-free supernatant was precipitated with 60-80% ammonium sulfate and stirred overnight. Proteins were precipitated by adding ammonium sulfate to 40% saturation and stirring for 24 hours. The precipitate was collected and re-suspended in Tris-buffer, pH 7. Additional precipitations were performed for 40-60% and 60-80% ammonium sulfate saturation. The 0-40% fraction was dissolved in Tris buffer, dried, and re-suspended in Tris buffer, pH 7, and assayed for bacteriocin activity (Burianek & Yousef, 2000;Zommiti, Bouffartigues, et al., 2018).
2.4. Bacteriocin extraction using solvents
2.4.1. Solvent extraction
The MRS broth was inoculated with 1% of an overnight bacteriocin-producing Enterococcus species, and incubated at 37 C for 16–18 hours. The culture was centrifuged, and the cell-free supernatant was stirred with organic solvents (1:1, v/v), like Methanol, Petroleum ether, chloroform, and isopropanol, and with their combinations of two or three (2:1, V/V). The culture supernatant fluid was acidified with acetic acid and extracted with chloroform and methanol (Ivy et al., 2022; Vimont et al., 2017).
2.4.2. Ethyl acetate extraction
The solvent extraction method was employed to extract bacteriocins from selected strains. Bacteria were inoculated into MRS broth and incubated at 37°C for 24 hours. . In 500ml separating funnel, equal volume of bacterial cell and ethyl acetate were added and the culture broth was shaken vigorously for 10 minutes. The upper organic layer containing bacteriocins recovered, while the rest part was separately separated to the new container. The solvent was removed under vacuum using a rotary evaporator. The dry extract was dissolved in methanol, adjusted to pH 7, and treated with 5 mg/ml catalase to remove hydrogen peroxide antagonistic activity. The extract was filtered through a 0.20μm membrane filter, and the filtered supernatant was stored in glass vials (Barale et al., 2022; Muhammad et al., 2017).
2.4.3. Butanol Extraction
To extract bacteriocins using butanol, 50 ml of n-butanol and 5g NaCl were mixed with cell-free supernatant of bacteriocin producing LAB. The mixture was centrifuged for 10 minutes, evaporated at 45 ºC, and re-suspended in saline (50ml, 9g/L). The partially purified crude bacteriocins were used to determine their characteristics and antagonistic activities against foodborne pathogens (Elyass et al., 2015).
2.4.4. Petroleum ether extraction
The study used petroleum ether for bacteriocins extraction by inoculating LAB into MRS broth and incubating at 37°C for 24 hours. The culture broth was transferred to a separating funnel, previous inoculated with petroleum ether. The separating funnel was shaken for 10 minutes, and the content settled into two distinct layers. The upper organic layer contained bacteriocins, and the rest of the broth culture was separated. The solvent was removed under vacuum using a rotary evaporator. The dried extract was dissolved in methanol, adjusted to pH 7.00 with 1M NaOH, and treated with 5 mg/ml catalase to remove antagonistic activity from hydrogen peroxide. The extract was filtered through a 0.20μm pore size membrane and evaluated for bacteriocins activity (Yang et al., 2012).
2.4.5. Chloroform extraction.
A MRS broth was inoculated with 1% of an overnight culture of bacteriocin-producing LAB and incubated for 18 hours at 37 °C. Cell-free supernatants were recovered after centrifugation at 8,000 rpm for 20 min. at 4 °C. Chloroform was added to the cell-free supernatants, stirred vigorously, and centrifuged at 8,000 rpm for 20 minutes. The sediment and solids at the interface were recovered by carefully pouring off the top aqueous layer and holding back floating interfacial precipitates. Tris-buffer was used to re-suspend the contents of the bottles, and the mixtures were combined in a test tube. The sediments were separated from the remaining chloroform and medium. The pellet was transferred to a small aluminum pan for drying in a chemical hood overnight. A second extraction with chloroform yielded minimal amounts of material at the interface and did not add any antimicrobial activity to the sample. A portion of the powder was suspended in a 1 ml Tris buffer and held overnight in the refrigerator to rehydrate and dissolve particles (Ahmed et al., 2016; Sasidharan et al., 2011; Xie et al., 2021).
2.5. Physic-chemical characterization of bacteriocins
The crude bacteriocins extracted from all LAB isolates with strong inhibition zone was characterized with respect to stability to temperature, pH, and enzymes and others parameters.
2.5.1. Effect of Temperature
In order to test the effect of temperature (heat resistance), 10 ml of crude bacteriocin preparation was exposed for 15 minutes to various heat treatments: 40℃, 60℃, 80℃ and 100℃, respectively. Residual bacteriocins activity was detected against selected pathogenic bacteria at each of these temperatures by using agar-well diffusion assay (Ogunbanwo et al., 2004).
2.5.1.1. Effect of pH
The sensitivity of crude bacteriocins to different pH values was tested by adjusting the bacteriocins' pH to 6.5, 8.5, and 12. The cell-free supernatant was adjusted with hydrochloric acid and sodium hydroxide, and incubated for 4 hours at room temperature. Residual activity against selected pathogenic bacteria was determined using agar-well diffusion assay. Treatment control, organism control and media control was kept (Kelleher et al., 2003).
2.5.1.2. Effect of enzymes
Effect of enzymes was tested on the antagonistic activity of crude bacteriocins preparation by treatment with amylase, lipase, and protease each at a final concentration of 1mg per ml. It was then incubated at room temperature for 2 hours and residual activity of bacteriocins was assayed along with bacteriocins and salts (Yamada et al., 1983).
2.5.1.3. Effect of solvents
Sensitivity of bacteriocins to different solvents concentration was investigated by the addition of 50% volume of methanol, ethanol, acetone and ethyl acetate (Ammor et al., 2005). Samples with and without of solvents was incubated for 12 h at 37°C and the residual activity was determined by the well diffusion assay.
2.5.1.4. Effect of surfactants
Surfactant sensitivity of bacteriocins was investigated by the addition of 1% (final volume) SDS, CTAB, Triton-X 100, Tween 80 and Tween 20 (Ammor et al., 2005) to cell free supernatant. Samples with and without of surfactants were incubated for 12 h at 37°C and residual activity was determined by well diffusion method.
2.6. Optimization for enhanced bacteriocins production
2.6.1. Media selection for bacteriocins optimization
To identify the potential media for bacteriocins production and optimization commercially available media such as Nutrient Broth, Tryptone Soy Broth, Brine Heart Infusion Broth and MRS Broth and other were prepared in laboratory to optimize carbon to nitrogen ratio. About 100 ml of broth was prepared in 250 ml conical flask and 10% of inoculum was added, and then kept without agitation for 16 h at 35°C. The fermented broth was centrifuged and analyzed for antimicrobial potential of bacteriocins.
2.6.2. Effect of carbon sources on bacteriocins production
In order to evaluate the effect of carbon sources on the growth of bacteriocins producing lactic acid bacteria and bacteriocins production, 2% dextrose, glucose, maltose, fructose, sucrose, starch and lactose were supplemented in MRS broth and were incubated at 37°C for 48 hours. The cell biomass and bacteriocins activities were determined(Ellis, 2001).
2.6.3. Effect of nitrogen sources bacteriocins production
To evaluate the effect of different nitrogen on the growth of bacteriocins producing lactic acid bacteria and bacteriocins production, MRS broth was supplemented with 2% nitrogenous compounds (meat extract, yeast extract, tryptone and bacteriological peptone) and were incubated at 37°C for 48 hours. The cell biomass and bacteriocins activities were determined (Nelson et al., 2001).
2.6.4. Effect of Incubation Period on Bacteriocins Production
The effect of incubation period on the production of bacteriocins was carried out. Media was incubated for different periods like 16h, 18 h, 24h, 32h and 37h, respectively. Finally, to determine the efficiency of bacteriocins, after each activity the bacteriocins antimicrobial activity was carried out.
2.7. Data analysis
All experiments were carried out in triplicate. The data is displayed as mean standard deviation. A one-way ANOVA was employed to evaluate statistical differences, followed by Tukey's multiple comparison test at P< 0.05 was used to determine whether a difference was significant.
3. Results
3.1. Evaluating bacteriocins producing potential
In this study, 60 Enterococcus species were identified based on morphology, biochemistry, and physiological features, and out of the total isolates, six of them were screened for bacteriocin-producing ability and antagonistic activity against foodborne pathogens. All of the isolates showed strong inhibition zones ranging from 13mm to 35mm. As the result, all six Enterococcus faecium showed strong inhibitory action against Escherichia coli, Staphylococcus aurous, Salmonella Typhimurium, Pseudomonas aeuroginosa, and Candida albicans. Enterococcus faecium JULAB-Br39 showed the highest inhibition zone with the diameter of inhibition zone 20.50, 21.00 and 20.17 mm against, Escherichia coli, Pseudomonas aeuroginosa and Salmonella Typhimurium, respectively. The inhibition zone showed significant variation at P<0.05. Generally, Enterococcus faecium JULAB-E23, JULAB-Br39, and JULAB-S36, were the potential bacteriocin-producing candidate showed the highest inhibition zone against all selected food borne pathogens (Table 2). For the final confirmation, proteinaceous nature of the bacteriocins confirmed by evaluating their sensitivity to photolytic enzymes. Hence, bacteriocins of six Enterococcus faecium exhibited complete inactivation of antimicrobial activity after the treatment of bacteriocins with proteinase K confirming its proteinaceous nature.
3.2. Antimicrobial activity of bacteriocins crude extract
3.2.1. Determination of antimicrobial substance
Based on the basic properties of the protein to determine whether an antimicrobial substance is a natural protein. Antimicrobial compounds produced by Enterococcus faeium inhibited by proteolytic enzymes such as pepsin, trypsin, and proteinase K (Table 3). The mean values of the inhibition zone resulted due to suspected bacteriocin producing Enterococcus faecium showed a significance variation at p<0.05(Table 3).
3.2.2. Preparation of bacteriocin crude extract
The diameter of the inhibition zone of the crude extract of bacteriocin obtained by ammonium sulfate precipitation was less than 18 mm (Table 4). The antibacterial ability of the crude extract gradually increased in the increase in ammonium sulfate concentration. The diameter of the inhibition zone reached the maximum when 80% ammonium sulfate and pH 7-phosphate buffer solution was reconstituted.
In this study, different solvents were used to extract bacteriocins, the diameter of the inhibition zone of the crude extract extracted with all solvents listed in Table 5. Mostly, the zone of inhibitions of crude bacteriocins extracted with chloroform, isopropanol and ethyl acetate and above 18 mm, indicating that all of these three solvents had good extraction effects on bacteriocin. The crude bacteriocin extract obtained by using theses solvents reconstituted in phosphate buffer solution to pH 7.0 had the best antimicrobial activity, which the diameter of the inhibition zone was above 28 mm. It was found that the antibacterial activity of the crude extract prepared by extraction of chloroform, isopropanol and ethyl acetate was significantly higher than that of the ammonium sulfate fractionation after the comparison of ammonium sulfate precipitation and organic solvent extraction.
3.3. Characterization of the partially purified supernatants
3.3.1. Effects of Temperature
Temperature plays a crucial role in bacteriocin activity. There was a general trend showing a decrease in bacteriocin activity with an increase in temperature. The bacteriocin extracted from Enterococcus faecium with JULAB23, JUALBBr39, and JULABS36, were quite active even passing through high temperature and pressure during sterilization, indicating it is a heat-stable protein. Generally, bacteriocin produced by many bacteriocins producers were active and outstanding range of temperatures from 45-50°C. On the other hand, no activity was detected in autoclaved supernatant even after 10 minutes only (Figure 1 and Figure 2).
3.3.2. Effect of pH on the inhibitory activity
The effects of changing the pH of the supernatants to various values have exhibited dramatic alterations, as indicated in Table 6. After the pH adjustment, the supernatant obtained from bacteriocin-producing Enterococcus faecium had the highest inhibitory activity at pH 5.0, decreasing slightly at pH 7.0 and pH 3.0, and showing the most remarkable decrease at pH 9.0, which results in a 40% reduction from the pH 5.00 values.
3.3.3. Effect of enzymes
Table 7 shows the effects of subjecting the supernatants obtained from each of the isolates to the enzymes proteinase-k, pepsin, trypsin and α-amylase, and then using them to inhibit growth of some selected indicator bacteria (E. coli, S. aureus and L. monocytogenes). It can be seen that no inhibition was produced by any of the cell-free supernatants in presence of the protein-digesting enzymes (proteinase-k and pepsin) indicating complete digestion of the inhibitory substance in the supernatants, and asserting its proteinaceus nature; while no reduction in the inhibitory activity was observed in the presence of the carbohydrate-degrading αamylase. No inhibitory activity was shown in un-inoculated medium containing no enzyme (negative control).
3.3.4. Effects of surfactants and additives of the antimicrobial activity of bacteriocins
The antimicrobial activity of bacteriocin producing Enterococcus faecium were evaluated by treating with different organic compounds, the antagonistic activity of bacteriocin like inhibitor substance remain active. However, the antibacterial effect was reduced to some extent when methanol was added to supernatants possibly because the surface structure of various antimicrobial agents caused intolerance to methanol. In addition, CFS of bacteriocin producing Enterococcus faecium were treated with additives, such as sodium citrate, potassium and tween, to verify the effect of food additives and other chemicals on the antibacterial components of bacteriocin producing lactic acid bacteria LAB (Table 6). Hence, CFS of the bacteriocins producers showed stable antimicrobial activity against the indicator bacteria treated when treated with sodium citrate and potassium. However, after Tween-80 was added, the antimicrobial activity remarkably differed and relatively reduced its potential antimicrobial activity.
3.4. Bacteriocins production
The bacteriocin production started during the late log phase and early stationary phase, however, the bacteriocin concentration level decreased in period of stationary phase. This is may be due to the partial degradation or adsorption of on produced cells. Regarding to the physico-chemical property of bacteriocins, which tolerate a wide range of temperatures and pH, but it was inactivated at pH 2 and 10. The activity of bacteriocin was not lost after treatments with surfactants and most of the organic solvents.
To determine the maximum bacteriocins production, the antimicrobial activity of bacteriocins producing Enterococcus faecium was evaluated using four different media, and then modified MRS broth results the maximum antimicrobial activity followed by Brain Heart infusion broth and Tryptosa soya broth, while nutrient broth produce the least antimicrobial activity. To evaluate the media potency, the antimicrobial activity of bacteriocin producing Enterococcus faecium was tested against S.aurues (Figure 3).
3.5. Effects of carbon sources on antimicrobial activity of Enterococcus faecium
In this study, the effect of the carbon source, on the antimicrobial activity of bacteriocin producing Enterococcus faecium was determined using MRS media supplemented with 2% of different carbon sources (glucose, lactose, fructose, Maltose, and sucrose) as proposed by (Todorov, 2009). The strongest antimicrobial activity of bacteriocin producing Enterococcus faecium was observed in the MRS media supplemented with glucose, sucrose and maltose after their inhibitory activity tested against L.monocytogens (Table 6).
3.6. Effects of Nitrogen sources on antimicrobial activity of Enterococcus faecium
Similarly, the effects of nitrogen sources on the antimicrobial activity of Enterococcus faecium was investigated, hence the MRS broth supplemented with yeast and meat extract produced potential antagonistic effects against Listeria monocytogenes followed by tyrptone, while MRS broth supplemented with peptone produced the least inhibition results (Figure 4).
4. Discussion
In this study, six Enterococcus faecium were identified and characterized from cabbage, shamita, borde, and ergo, for their bacteriocin producincing capacity. Bacteriocin-producing Enterococcus faecium demonstrated antimicrobial activity against Escherichia coli, Staphylococcus aureus, Listeria monocytogenes, Pseudomonas auroginosa, Salmonella Typhimurium, and Candida albicans. Therefore, all six Enterococcus faecium showed the strong and potential antagonistic activity against all selected food borne pathogens. Similar to this result, Balla et al. (2000) demonstrated that Enterococcus faecium, convened for bacteriocin-producing capability, showed broad antagonistic activity against Listeria monocytogenes. Besides this, Gaaloul et al. (2015) reported that antimicrobial substances produced by Enterococcus faecium GGN7 demonstrated potential antagonistic effects against Listeria monocytogenes. In more recent studies, bacteriocin-producing Enterococcus faecium isolated from various sources has shown potential antimicrobial effects against several foodborne pathogens (Cocolin et al., 2007; Ghrairi et al., 2008; Ohmomo et al., 2000; Rehaiem et al., 2010). In addition, Franzetti et al (2004); Gaaloul et al. (2015); Hosseini et al. (2009); Quintela-Baluja et al. (2022) and Silva et al. (2018) large numbers of bacteriocin producing Enterococcus faecium were isolated from raw milk and fermented food origin and various food stuff and demonstrated potential bacteriocin producing ability. These findings confirmed that Enterococcus faecium isolated from Ethiopian traditional fermented foods and beverages was found to be the best potential food preservative because of its capacity to produce various antimicrobial substances, including bacteriocins.
The CFS of bacteriocin-producing Enterococcus faecium demonstrated broad antimicrobial activity against both gram-positive pathogens (Listeria monocytogenes and Staphylococcus aureus) and gram-negative pathogens (Salmonella Typhimurium and Escherichia coli), with inhibition zones ranging from 13.00 to 35.31 mm. Abanoz & Kunduhoglu (2018) reported that Enterococcus faecium exhibited inhibition zones against Listeria monocytogenes, Staphylococcus aureus, Salmonella Typhimurium, and Escherichia coli ranging from 20 to 28 mm. This implies that Enterococcus faecium's antimicrobial activity may vary depending on the source, the method of isolation, and the best selection criteria used to identify the bacteriocin-producing Enterococcus faecium.
Similarly, Perumal & Venkatesan, (2017) demonstrated that Enterococcus faecium exhibited broad antimicrobial activity against both gram-positive bacteria (L. monocytogenes and S. aureus) and gram-negative bacteria (Salmonella sp., S. thypi, and E. coli), with inhibition zones ranging from 13 to 18 mm. As a result, those Enterococcus faecium isolates were able to synthesize bacteriocin-like antimicrobial peptides with potential antagonism against gram-positive and gram-negative bacteria (Drider et al., 2010).
Enterococcus faecium produces antimicrobial substances that are inactivated by pepsin, trypsin, and proteinase K, but perform well with -amylase enzyme. These peptides are considered as bacteriocin-like substances, with a proteinase nature. Correspondingly, Ghrairi et al. (2008) reported that the antimicrobial activity of bacteriocin producing Enterococcus faecium MMT21, remain active after treated by pepsin, trypsin and proteinase K. Likewise, Cocolin et al. (2007), Ghrairi et al. (2008) and Zommiti et al. (2018) reported that, bacteriocin like antimicrobial substances produced by Enterococcus faecium MMT22, Enterococcus faecium M241 and M249, Enterococcus faecium GHB21 was totally deactivated with the inoculation these enzymes and therefore antimicrobial substances produced by Enterococcus faecium isolated in this study is considered as bacteriocin like antimicrobial substances.
Bacteriocin-producing Enterococcus faecium showed stable antibacterial activity against S. aureus at 80°C, but not at 100°C, indicating its effectiveness at high temperatures (80-100°C).This suggests that Enterococcus faecium may be a potential candidate for food preservation at high temperatures. Similar to these findings, bacteriocin like substance produced by Enterococcus faecium was reported to be more stable and effectively inhibit the growth of food borne pathogens even at the high temperature 80-100 °C for 10-20 minutes (Du et al., 2017).
It was also discovered that the bacteriocin produced by E. faecium M241 and GHB21 remained entirely stable following a 30-minute after heat treatment at 100 °C, although activity reduced at higher temperatures (Abanoz & Kunduhoglu, 2018; Cocolin et al., 2007), which is quite similar to these findings. On other hand, Ben Braïek et al. (2017) and Phumisantiphong et al. (2017), observed that the antibacterial activity of bacteriocin-producing E. faecium EF478 and E. faecium Q1 shown potential stability at 60 °C for 30 minutes and 60 °C for 1 hour, respectively. Therefore, based on the thermal stability and proteinase nature of the antimicrobial substance produced by Enterococcus faecium, these isolates be used for industrial applications as a novel and promising food preservatives, since many food processing procedures involve a heating step (Abanoz & Kunduhoglu, 2018).
The stability of bacteriocin producing Enterococcus faecium was evaluated against S. aureus ATCC @25923 from pH 2-12 values. All six Enterococcus faecium produced potential antimicrobial activity after treated with different pH values. The maximum bacteriocin activity was observed at pH 5 and 7. In line to this study, Kumar et al. (2010) and Sarkar et al. (2020), reported that antimicrobial activity of bacteriocin producing E. faecium LR/6 remain effective in between pH, 2 -7, but ,about 20 % of the inhibition activity reduced, while the pH increased to pH 8.00. Similarly, to this study, several bacteriocin producing Enterococcus faecium showed potential antagonistic activity at wide pH range, from 2.00 to 8.00 (Kang & Lee, 2005; Moreira et al., 2020). As a result, the effect of pH on the antimicrobial activity of bacteriocin-producing Enterococcus faecium revealed that the antimicrobial peptides produced by these isolates were found to be active over a wide pH range of 2-8.500, allowing their use as food preservatives in acidic and alkaline environments(Isleroglu et al., 2012).
In this study, the effects of different surfactants and food additives on bacteriocin efficiency was evaluated , the antibacterial activity of bacteriocin producing Enterococcus faecium remain active having active antagonistic activity against S. aureus after treated with different food additive and surfactants. Similarly, several researchers reported that, the CFS of bacteriocin produced by Enterococcus faecium treated with organic solvents, detergents demonstrated potential antimicrobial activity (Rajeswari et al., 2010; Rehaiem et al., 2010; Santos et al., 2018).
Furthermore, the resistance of bacteriocin-producing Enterococcus faecium to organic solvents, surfactants, and detergents facilitates the widespread use of antimicrobial substances, by allowing the isolates to retain structural and functional properties during various purification processes. As a result, bacteriocin stability under conditions such as organic solvents, surfactants, and detergents, acid resistance, and heat stability is highly beneficial and enables their application in food processing and preservation of various foods and drinks(V. Kumar et al., 2016; Rushdy & Gomaa, 2013).
The study evaluated various media for bacteriocin production, finding that Enterococcus faecium cultivated with Modified MRS broth and BHI broth achieved the best antimicrobial activity. MRS medium contains carbon and nitrogen sources, while BHI is a general-purpose nutrient medium suitable for various microorganisms. Growth kinetics showed that BHI broth with an initial pH of 7.4 was ideal for LAB growth, resulting in potential antimicrobial activity. This finding is more consistent with the research reported by, as the researcher reported that, the highest bacteriocin antimicrobial was obtained when the Enterococcus faecium were incubated in MRS broth at 32°C for 16-24h, while BHI medium was suitable for bacteriocin production pH 7.4(Gutiérrez-Cortés et al., 2018; Todorov, 2009; Weine & Kim, 2019; Zhang et al., 2018). Therefore, modified MRS broth and BHI broth supplemented with different carbon and nitrogen sources are the most suitable media for bacteriocin production.
In this study, ammonium sulfate precipitation and solvent extraction methods developed to recovery bacteriocin production efficiency from E.faecium. Accordingly, various techniques, ammonium sulfate precipitation, chloroform, ethyl acetate, isopropanol, petroleum, methanol and ethanol based solvent extraction were used to recovery bacteriocin producing potential of E.faecium. Hence, the bacteriocin precipitated by 40 and 60 % w/v ammonium sulphate showed less effective antagonistic activities as compared to antimicrobial activity precipitated by 80% ammonium sulphate. Similarly, Barale et al. (2022) and Xu et al. (2009) reported that the antimicrobial activity of bacteriocin is greater when it precipitates at a higher concentration of ammonium sulfate than when it precipitates at a lower percent or concentration, because the higher concentration recovers the bacteriocin (up to 80% ammonium sulfate efficiency).
The recovery of bacteriocin from E. faecium using various solvents was found to be most promising for extracting bacteriocin-like compounds with high antibacterial activity. Chloroform and its mixture showed the greatest recovery, followed by isopropanol and ethyl acetate. These solvents are believed to be capable of extracting antimicrobial peptides from MRS broth of putative bacteriocin-producing E. faecium. In line to this result, Barale et al (2022) found that chloroform and ethyl acetate were the most effective solvents for extracting bacteriocin and lipo-peptides. Furthermore, the authors demonstrated that, methanol was the best solvent for extracting bacteriocin with good antimicrobial activity against B. cereus NCIM 2703, S. aureus NCIM 2654, and B. cereus ATCC 10,876 with zones of inhibition of 24 mm, 22 mm, and 14 mm, respectively (Barale et al., 2022). As a result, it is obvious that combining two or more approaches might be used to efficiently purify and recover bacteriocin from bacteriocin-producing lactic acid bacteria isolated from diverse traditional fermented foods and beverages.
Ethiopian traditional fermented beverages and dairy products are rich sources of potent bacteriocin-producing strains. Six Enterococcus faecium isolates, JULABBr39, JULABBr40, JULABBS36, JULABS43, JULABE23, and JULABE36, were isolated from Ethiopian fermented foods. These isolates produce heat-stable, pH resistant bacteriocin-like substances, and exhibit potential antagonism against Gram-positive and Gram-negative bacteria. These isolates have a wide activity spectrum against various bacteria, including Escherichia coli, Staphylococcus aureus, Listeria monocytogenes, Pseudomonas auroginoasa, Salmonella Typhimurium, and Candida albicans
Based on these fascinating properties, the bacteriocin-producing Enterococcus faecium strains isolated in this study could be a promising candidate to be employed as a potential food preservative, particularly due to their stability, ability to withstand some extreme conditions that may be encountered while preparing some processed foods, and ability to inhibit some pathogenic and/or spoilage bacteria. However, for further it needs some detail study and research focusing on isolation characterization of bacteriocin and other active peptides using completely genomic sequencing to identify the types, as well as the position of cluster genes and encode gene responsible for bacteriocin produced by Enterococcus faecium, as well as determination of antibactiotic resistance gene identification, in order to qualify Enterococcus faecium and their bacteriocins as a safe bio preservative.
Authors Contributions
Dessalegn Amenu and Ketema Bacha proposed the research concept and provided the necessary tools for the experiments and experimental instructions. Dessalegn Amenu designed and conducted the experiments and analyzed the data. The authors read and approved the manuscript.
Funding
The research work is supported by Jimma University, College of Natural sciences (Staff research grant, at JU, Natural science).
Availability of data and materials
The study's original contributions are included in the article/ supplementary file, any questions should be directed to the corresponding author.
Ethics approval and consent to participate
This research study does not include any of the authors' experiments involving human or animal subjects.
Consent for publication
This article contains no human participants.
Acknowledgment
The author is also extremely appreciative of the financial support provided by Jimma University's College of Natural Sciences for the best completion of this project.
Competing interests
The authors declare that, the authors have no any competing interests
Abbreviation
LAB= lactic acid bacteria
GRAS= Generally recognized as safe
CFS= Cell free supernatant
FBD= Foodborne diseases
EPHI= Ethiopian Public Health Institute
JU= Jimma university
References
- Abanoz, H. S., & Kunduhoglu, B. (2018). Antimicrobial Activity of a Bacteriocin Produced byEnterococcus faecalis KT11 against Some Pathogens andAntibiotic-Resistant Bacteria. Korean Journal for Food Science of Animal Resources, 38(5) 1064. [CrossRef]
- Ahmed, D., Fatima, K., Saeed, R., & Masih, R. (2016). Isolation and identification of bioactive compounds from chloroform fraction of methanolic extract of Carissa opaca roots. Natural Product Research, 30(17), 2012–2016. [CrossRef]
- Akhtar, M. S., Panwar, J., & Yun, Y. S. (2013). Biogenic synthesis of metallic nanoparticles by plant extracts. ACS Sustainable Chemistry and Engineering, 1(6). [CrossRef]
- Amit, S. K., Uddin, M. M., Rahman, R., Islam, S. M. R., & Khan, M. S. (2017). A review on mechanisms and commercial aspects of food preservation and processing. Agriculture and Food Security, 6(1), 1–22. [CrossRef]
- Ammor, S., Dufour, E., Zagorec, M., Chaillou, S., & Chevallier, I. (2005). Characterization and selection of Lactobacillus sakei strains isolated from traditional dry sausage for their potential use as starter cultures. Food Microbiology, 22(6). [CrossRef]
- Balla, E., Dicks, L. M. T., Du Toit, M., Van Der Merwe, M. J., & Holzapfel, W. H. (2000). Characterization and cloning of the genes encoding enterocin 1071A and enterocin 1071B, two antimicrobial peptides produced by Enterococcus faecalis BFE 1071. Applied and Environmental Microbiology, 66(4), 1298–1304. [CrossRef]
- Barale, S. S., Ghane, S. G., & Sonawane, K. D. (2022a). Purification and characterization of antibacterial surfactin isoforms produced by Bacillus velezensis SK. AMB Express, 12(1), 1–20. [CrossRef]
- Barale, S. S., Ghane, S. G., & Sonawane, K. D. (2022b). Purification and characterization of antibacterial surfactin isoforms produced by Bacillus velezensis SK. AMB Express, 12(1), 1–20. [CrossRef]
- Ben Braïek, O., Ghomrassi, H., Cremonesi, P., Morandi, S., Fleury, Y., Le Chevalier, P., Hani, K., Bel Hadj, O., & Ghrairi, T. (2017). Isolation and characterisation of an enterocin P-producing Enterococcus lactis strain from a fresh shrimp (Penaeus vannamei). Antonie van Leeuwenhoek, International Journal of General and Molecular Microbiology, 110(6), 771–786. [CrossRef] [PubMed]
- Burianek, L. L., & Yousef, A. E. (2000). Solvent extraction of bacteriocins from liquid cultures. Letters in Applied Microbiology, 31(3), 193–197. [CrossRef] [PubMed]
- Cocolin, L., Foschino, R., Comi, G., & Grazia Fortina, M. (2007). Description of the bacteriocins produced by two strains of Enterococcus faecium isolated from Italian goat milk. Food Microbiology, 24(7–8), 752–758. [CrossRef]
- do Nascimento, M. da S., Moreno, I., & Kuaye, A. Y. (2010). Antimicrobial activity of Enterococcus faecium FAIR-E 198 against gram-positive pathogens. Brazilian Journal of Microbiology, 41(1), 74–81. [CrossRef]
- Drider, D., Belguesmia, Y., Choiset, Y., Prévost, H., Dalgalarrondo, M., & Chobert, J. M. (2010). Partial purification and characterization of the mode of action of enterocin S37: A bacteriocin produced by enterococcus faecalis S37 isolated from poultry feces. Journal of Environmental and Public Health, 2010. [CrossRef]
- Du, L., Liu, F., Zhao, P., Zhao, T., & Doyle, M. P. (2017). Characterization of Enterococcus durans 152 bacteriocins and their inhibition of Listeria monocytogenes in ham. Food Microbiology, 68, 97–103. [CrossRef]
- Ellis, M. V. (2001). Harmful supervision, a cause for alarm: Comment on Gray et al. (2001) and Nelson and Friedlander (2001). In Journal of Counseling Psychology (Vol. 48, Issue 4). [CrossRef]
- Elyass, M. E., Mahdi, A. A., & Attitalla, I. H. (2015). Characterization and Evaluation of Antimicrobial Activity of Bacteriocins from Lactobacillus Curvatus and Pediococcus Pentosaceus. Infectious & Non Infectious Diseases, 1(1), 1–7. [CrossRef]
- Erb, D. K., Steidel, C. C., Shapley, A. E., Pettini, M., Reddy, N. A., & Adelberger, K. L. (2006). Hα Observations of a Large Sample of Galaxies at z ∼ 2: Implications for Star Formation in High-Redshift Galaxies. The Astrophysical Journal, 647(1). [CrossRef]
- Franzetti, L., Pompei, M., Scarpellini, M., & Galli, A. (2004). Phenotypic and genotypic characterization of Enterococcus spp. of different origins. Current Microbiology, 49(4), 255–260. [CrossRef]
- Freitas, T. K. F. S., Oliveira, V. M., de Souza, M. T. F., Geraldino, H. C. L., Almeida, V. C., Fávaro, S. L., & Garcia, J. C. (2015). Optimization of coagulation-flocculation process for treatment of industrial textile wastewater using okra (A. esculentus) mucilage as natural coagulant. Industrial Crops and Products, 76, 538–544. [CrossRef]
- Fugaban, J. I. I., Bucheli, J. E. V., Holzapfel, W. H., & Todorov, S. D. (2021). Characterization of partially purified bacteriocins produced by enterococcus faecium strains isolated from soybean paste active against listeria spp. and vancomycin-resistant enterococci. Microorganisms, 9(5), 1085. https://www.mdpi.com/2076-2607/9/5/1085/htm.
- Gaaloul, N., ben Braiek, O., Hani, K., Volski, A., Chikindas, M. L., & Ghrairi, T. (2015). Isolation and characterization of large spectrum and multiple bacteriocin-producing Enterococcus faecium strain from raw bovine milk. Journal of Applied Microbiology, 118(2), 343–355. [CrossRef] [PubMed]
- Ghrairi, T., Frere, J., Berjeaud, J. M., & Manai, M. (2008). Purification and characterisation of bacteriocins produced by Enterococcus faecium from Tunisian rigouta cheese. Food Control, 19(2), 162–169. [CrossRef]
- Goh, H. F., & Philip, K. (2015). Purification and Characterization of Bacteriocin Produced by Weissella confusa A3 of Dairy Origin. PLOS ONE, 10(10), e0140434. [CrossRef]
- Gutiérrez-Cortés, C., Suarez, H., Buitrago, G., Nero, L. A., & Todorov, S. D. (2018). Characterization of bacteriocins produced by strains of Pediococcus pentosaceus isolated from Minas cheese. Annals of Microbiology, 68(6), 383–398. https://annalsmicrobiology.biomedcentral.com/articles/10.1007/s13213-018-1345-z.
- Hosseini, S. V., Arlindo, S., Böhme, K., Fernández-No, C., Calo-Mata, P., & Barros-Velázquez, J. (2009). Molecular and probiotic characterization of bacteriocin-producing Enterococcus faecium strains isolated from nonfermented animal foods. Journal of Applied Microbiology, 107(4), 1392–1403. [CrossRef]
- Isleroglu, H., Yildirim, Z., Tokatli, M., Oncul, N., & Yildirim, M. (2012). Partial characterisation of enterocin KP produced by Enterococcus faecalis KP, a cheese isolate. International Journal of Dairy Technology, 65(1), 90–97. [CrossRef]
- Ivy, J., Fugaban, I., Enrique Vazquez Bucheli, J., Park, J.Y., Suh, H.D., Jung, E. S., de Melo Franco, B.D.M., Ivanova, I.V., Holzapfel, W.H., Todorov, S.D., Paulo, S., Correspondence, B., & Dimitrov Todorov, S. (2022). Antimicrobial properties of Pediococcus acidilactici and Pediococcus pentosaceus isolated from silage. Journal of Applied Microbiology, 132(1), 311–330. [CrossRef]
- Kang, J. H., & Lee, M. S. (2005). Characterization of a bacteriocin produced by Enterococcus faecium GM-1 isolated from an infant. Journal of Applied Microbiology, 98(5), 1169–1176. [CrossRef]
- Kelleher, D. J., Karaoglu, D., Mandon, E. C., & Gilmore, R. (2003). Oligosaccharyltransferase isoforms that contain different catalytic STT3 subunits have distinct enzymatic properties. Molecular Cell, 12(1). [CrossRef]
- Kumar, M., Tiwari, S. K., & Srivastava, S. (2010). Purification and characterization of enterocin LR/6, a bacteriocin from enterococcus faecium LR/6. Applied Biochemistry and Biotechnology, 160(1), 40–49. [CrossRef]
- Kumar, V., Sheoran, P., Gupta, A., Yadav, J. P., & Tiwari, S. K. (2016). Antibacterial property of bacteriocin produced by Lactobacillus plantarum LD4 isolated from a fermented food. Annals of Microbiology, 66(4), 1431–1440.
- Molognoni, L., Daguer, H., Motta, G. E., Merlo, T. C., & Lindner, J. D. D. (2019). Interactions of preservatives in meat processing: Formation of carcinogenic compounds, analytical methods, and inhibitory agents. In Food Research International (Vol. 125). [CrossRef]
- Moreira, S. M., Picinin, C. T., do Amarante, C. C., Pilatti, L. A., & Pedroso, B. (2020). Simulation of the new proposed method by CAPES for the Qualis 2017-2020 classification of the Brazilian archives of biology and technology. Brazilian Archives of Biology and Technology, 63. [CrossRef]
- Muhammad Shahid Riaz Rajoka a, Hafiza Mahreen Mehwish c, Muhammad Siddiq b, Zhao Haobin a, Jing Zhu a, Li Yan a, Dongyan Shao a, Xiaoguang Xu a, J. S., Riaz Rajoka, M. S., Mehwish, H. M., Siddiq, M., Haobin, Z., Zhu, J., Yan, L., Shao, D., Xu, X., & Shi, J. (2017). Identification, characterization, and probiotic potential of Lactobacillus rhamnosus isolated from human milk. 84, 271–280. [CrossRef]
- O’Sullivan, L., Ross, R. P., & Hill, C. (2002). Potential of bacteriocin-producing lactic acid bacteria for improvements in food safety and quality. In Biochimie (Vol. 84, Issues 5–6). [CrossRef]
- Ogunbanwo, S. T., Sanni, A. I., & Onilude, A. A. (2003). Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. African Journal of Biotechnology, 2(8). [CrossRef]
- Ogunbanwo, S. T., Sanni, A. I., & Onilude, A. A. (2004). Effect of bacteriocinogenic Lactobacillus spp. on the shelf life of fufu, a traditional fermented cassava product. World Journal of Microbiology and Biotechnology, 20(1), 57–63. [CrossRef]
- Ohmomo, S., Murata, S., Katayama, N., Nitisinprasart, S., Kobayashi, M., Nakajima, T., Yajima, M., & Nakanishi, K. (2000). Purification and some characteristics of enterocin ON-157, a bacteriocin produced by Enterococcus faecium NIAI 157. Journal of Applied Microbiology, 88(1), 81–89. [CrossRef]
- Parada, J., & Aguilera, J. M. (2007). Food microstructure affects the bioavailability of several nutrients. Journal of Food Science, 72(2). [CrossRef]
- Perumal, V., & Venkatesan, A. (2017). Antimicrobial, cytotoxic effect and purification of bacteriocin from vancomycin susceptible Enterococcus faecalis and its safety evaluation for probiotization. LWT, 78, 303–310. [CrossRef]
- Phumisantiphong, U., Siripanichgon, K., Reamtong, O., & Diraphat, P. (2017). A novel bacteriocin from Enterococcus faecalis 478 exhibits a potent activity against vancomycin-resistant enterococci. PLoS ONE, 12(10). [CrossRef]
- Quintela-Baluja, M., Jobling, K., Graham, D. W., Tabraiz, S., Shamurad, B., Alnakip, M., Böhme, K., Barros-Velázquez, J., Carrera, M., & Calo-Mata, P. (2022). Rapid Proteomic Characterization of Bacteriocin-Producing Enterococcus faecium Strains from Foodstuffs. [CrossRef]
- Rajeswari, T., Venugopal, A., Viswanathan, C., Kishmu, L., Venil, C. K., & Kumar, S. (2010). Antibacterial activity of honey against Staphylococcus aureus from infected wounds. Pharmacologyonline.Silae.It, 1, 537–541. https://pharmacologyonline.silae.it/files/newsletter/2010/vol1/59.Kumar.pdf.
- Rehaiem, A., Martínez, B., Manai, M., & Rodríguez, A. (2010). Production of enterocin A by Enterococcus faecium MMRA isolated from ‘Rayeb’, a traditional Tunisian dairy beverage. Journal of Applied Microbiology, 108(5), 1685–1693. [CrossRef]
- Rushdy, A. A., & Gomaa, E. Z. (2013). Antimicrobial compounds produced by probiotic Lactobacillus brevis isolated from dairy products. Annals of Microbiology, 63(1). [CrossRef]
- Sanlibaba, P., Güçer, Y., & Şanlıbaba, P. (2015). ANTIMICROBIAL ACTIVITY OF LACTIC ACID BACTERIA. Agriculture & Food ISSN, 3. www.scientific-publications.net.
- Santos, R., Oliva-Teles, A., Saavedra, M., Enes, P., & Serra, C. (2018). Bacillus spp. as source of Natural Antimicrobial Compounds to control aquaculture bacterial fish pathogens. Frontiers in Marine Science, 5. [CrossRef]
- Sarkar, S. L., Hossain, M. I., Monika, S. A., Sanyal, S. K., Roy, P. C., Hossain, M. A., & Jahid, I. K. (2020). Probiotic potential of pediococcus acidilactici and enterococcus faecium isolated from indigenous yogurt and raw goat milk. Microbiology and Biotechnology Letters, 48(3). [CrossRef]
- Sasidharan, S., Chen, Y., Saravanan, D., Sundram, K. M., & Yoga Latha, L. (2011). Extraction, Isolation and Characterization of Bioactive Compounds from Plants’ Extracts. African Journal of Traditional, Complementary, and Alternative Medicines, 8(1), 1. [CrossRef]
- Silva, C. C. G. C. C. G., Silva, S. P. M. S. C. G., & Ribeiro, S. C. (2018). Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. 9(APR), 594. www.frontiersin.org.
- Sivapalasingam, S., Friedman, C. R., Cohen, L., & Tauxe, R. V. (2004). Fresh produce: A growing cause of outbreaks of foodborne illness in the United States, 1973 through 1997. In Journal of Food Protection (Vol. 67, Issue 10). [CrossRef]
- Todorov, S. D. (2009). Bacteriocinas de Lactobacillus plantarum - produção, organização genética e modo de ação. Brazilian Journal of Microbiology, 40(2), 209–221. [CrossRef]
- Vimont, A., Fernandez, B., Hammami, R., Ababsa, A., Daba, H., & Fliss, I. (2017). Bacteriocin-Producing Enterococcus faecium LCW 44: A High Potential Probiotic Candidate from Raw Camel Milk. Frontiers in Microbiology, 8(MAY). [CrossRef]
- Weaver, D. B., & Lawton, L. J. (2007). Twenty years on: The state of contemporary ecotourism research. Tourism Management, 28(5). [CrossRef]
- Weine, E. R., & Kim, N. S. (2019). Systematic distortions in clinicians’ memories for client cases: Increasing causal coherence. Journal of Experimental Psychology: Learning Memory and Cognition, 45(2). [CrossRef]
- Wingfield, P. T. (2001). Wingfield, P. T. (2001). Protein Precipitation Using Ammonium Sulfate. Current Protocols in Protein Science / Editorial Board, John E. Coligan... [et Al.], APPENDIX 3, AppendixPrecipitation Using Ammonium Sulfate. [CrossRef]
- Wray, N. R., Ripke, S., Mattheisen, M., Trzaskowski, M., Byrne, E. M., Abdellaoui, A., Adams, M. J., Agerbo, E., Air, T. M., Andlauer, T. M. F., Bacanu, S. A., Bækvad-Hansen, M., Beekman, A. F. T., Bigdeli, T. B., Binder, E. B., Blackwood, D. R. H., Bryois, J., Buttenschøn, H. N., Bybjerg-Grauholm, J., … Sullivan, P. F. (2018). Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nature Genetics, 50(5). [CrossRef]
- Xie, Y., Peng, Q., Ji, Y., Xie, A., Yang, L., Mu, S., Li, Z., He, T., Xiao, Y., Zhao, J., & Zhang, Q. (2021). Isolation and Identification of Antibacterial Bioactive Compounds From Bacillus megaterium L2. Frontiers in Microbiology, 12, 662. [CrossRef]
- Xu, H., Jeong, H. S., Lee, H. Y., & Ahn, J. (2009). Assessment of cell surface properties and adhesion potential of selected probiotic strains. Letters in Applied Microbiology, 49(4). [CrossRef]
- Yamada, R., Sato, M., Kawabata, M., Nakatsuka, H., Nakamura, K., & Takashima, S. (1983). Hepatic artery embolization in 120 patients with unresectable hepatoma. Radiology, 148(2). [CrossRef]
- Yang, E., Fan, L., Jiang, Y., Doucette, C., & Fillmore, S. (2012). Antimicrobial activity of bacteriocin-producing lactic acid bacteria isolated from cheeses and yogurts. AMB Express, 2(1), 1–12. [CrossRef]
- Yang, J., Ji, Y., Park, H., Lee, J., Park, S., Yeo, S., Shin, H., & Holzapfel, W. H. (2014). International Journal of Food Microbiology Selection of functional lactic acid bacteria as starter cultures for the fermentation of Korean leek ( Allium tuberosum Rottler ex Sprengel.). International Journal of Food Microbiology, 191, 164–171. [CrossRef] [PubMed]
- Zhang, J., Yang, Y., Yang, H., Bu, Y., Yi, H., Zhang, L., Han, X., & Ai, L. (2018). Purification and partial characterization of bacteriocin Lac-B23, a novel bacteriocin production by lactobacillus plantarumJ23, isolated from Chinese traditional fermented milk. Frontiers in Microbiology, 9(OCT). [CrossRef]
- Zommiti, M., Bouffartigues, E., Maillot, O., Barreau, M., Szunerits, S., Sebei, K., Feuilloley, M., Connil, N., & Ferchichi, M. (2018). In vitroassessment of the probiotic properties and bacteriocinogenic potential of pediococcus pentosaceusMZF16 isolated from artisanal tunisian meat "dried ossban. Frontiers in Microbiology, 9(NOV), 2607. [CrossRef]
- Zommiti, M., Cambronel, M., Maillot, O., Barreau, M., Sebei, K., Feuilloley, M., Ferchichi, M., & Connil, N. (2018). Evaluation of probiotic properties and safety of Enterococcus faecium isolated from artisanal Tunisian Meat ‘Dried Ossban’. Frontiers in Microbiology, 9(AUG). [CrossRef]
Figure 1.
Effects of Temperatures on the antimicrobial activity of bacteriocins producing Enterococcus faecium.
Figure 1.
Effects of Temperatures on the antimicrobial activity of bacteriocins producing Enterococcus faecium.
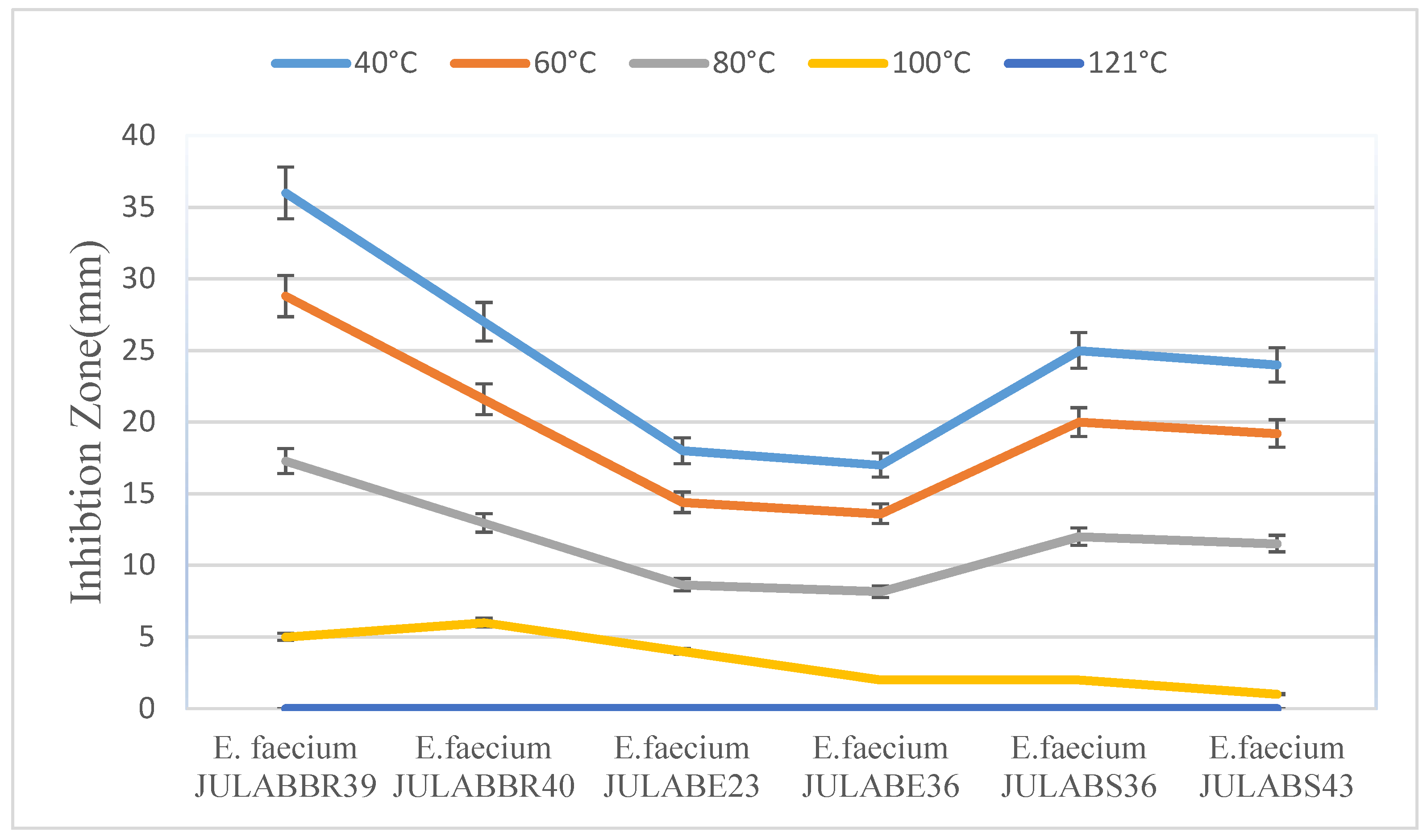
Figure 2.
Effects of pH on the antimicrobial activity of bacteriocins producing Enterococcus faecium.
Figure 2.
Effects of pH on the antimicrobial activity of bacteriocins producing Enterococcus faecium.
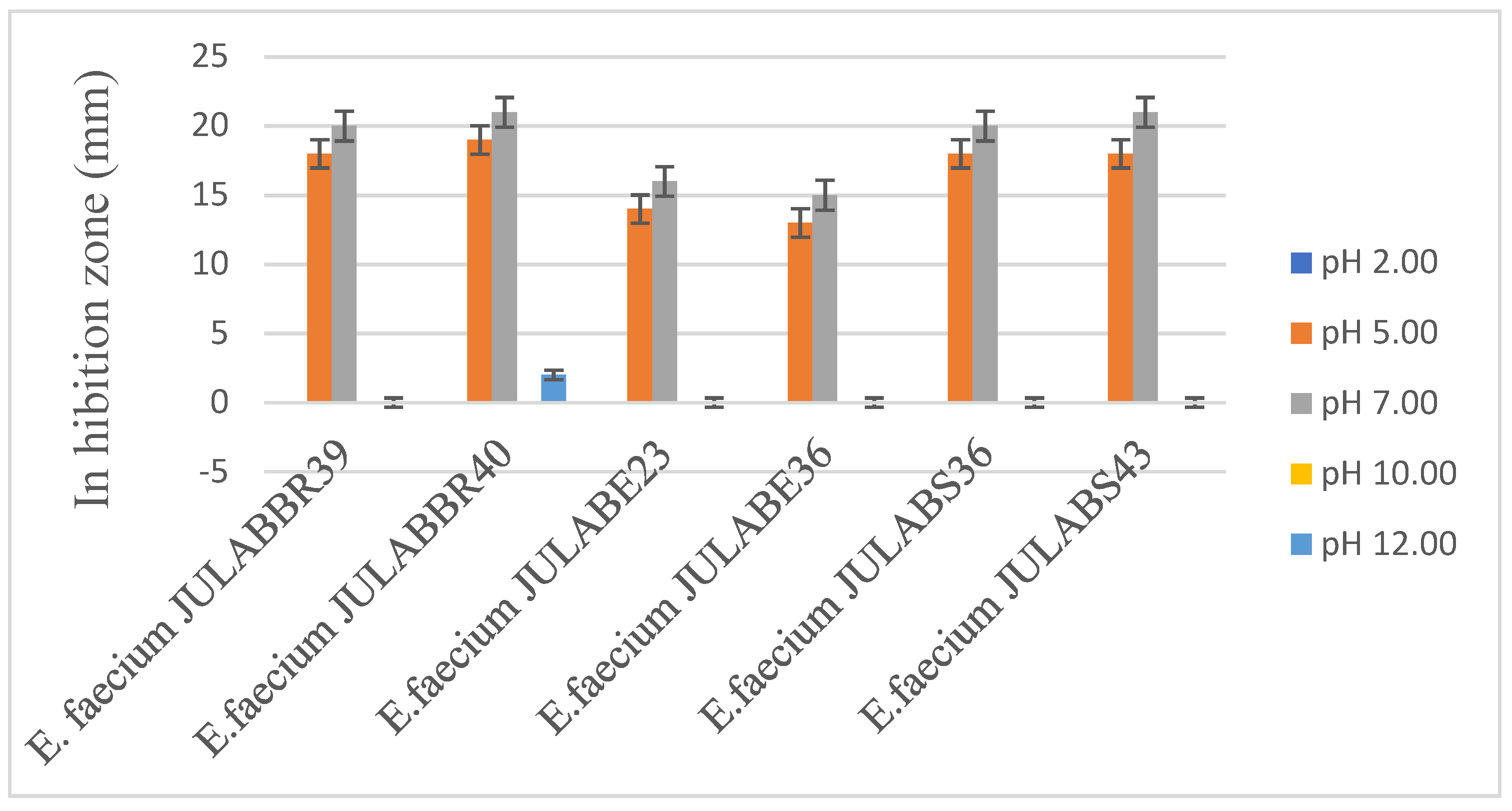
Figure 3.
Effects of different media composition for the optimum antimicrobial activity of bacteriocin producing Enterococcus faecium isolated from Ethiopian traditional fermented foods and beverages.
Figure 3.
Effects of different media composition for the optimum antimicrobial activity of bacteriocin producing Enterococcus faecium isolated from Ethiopian traditional fermented foods and beverages.
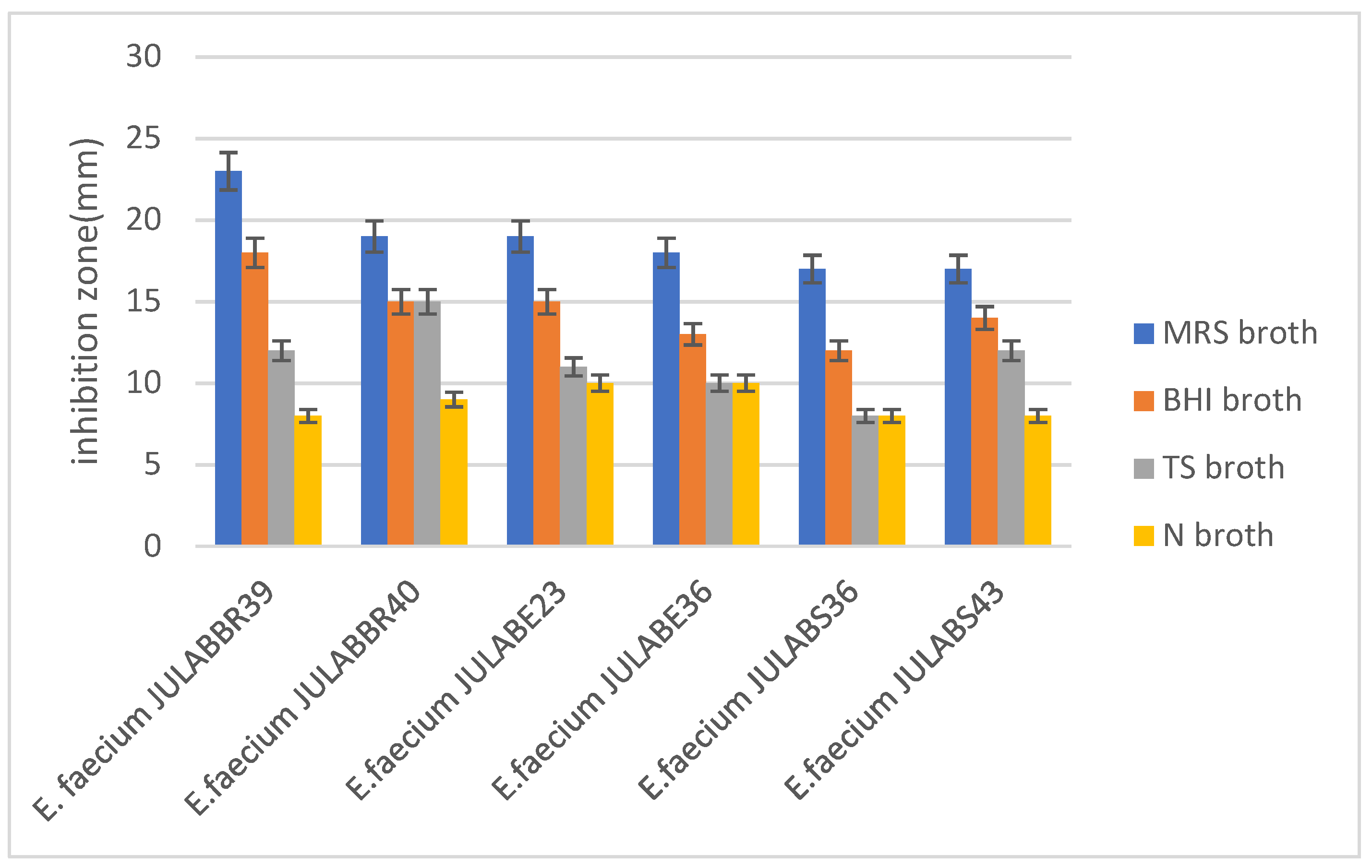
Figure 4.
Effects of nitrogen sources on antimicrobial activity of bacteriocin producing Enterococcus faecium after MRS broth supplemented with different carbon sources, against Listeria monocytogenes.
Figure 4.
Effects of nitrogen sources on antimicrobial activity of bacteriocin producing Enterococcus faecium after MRS broth supplemented with different carbon sources, against Listeria monocytogenes.
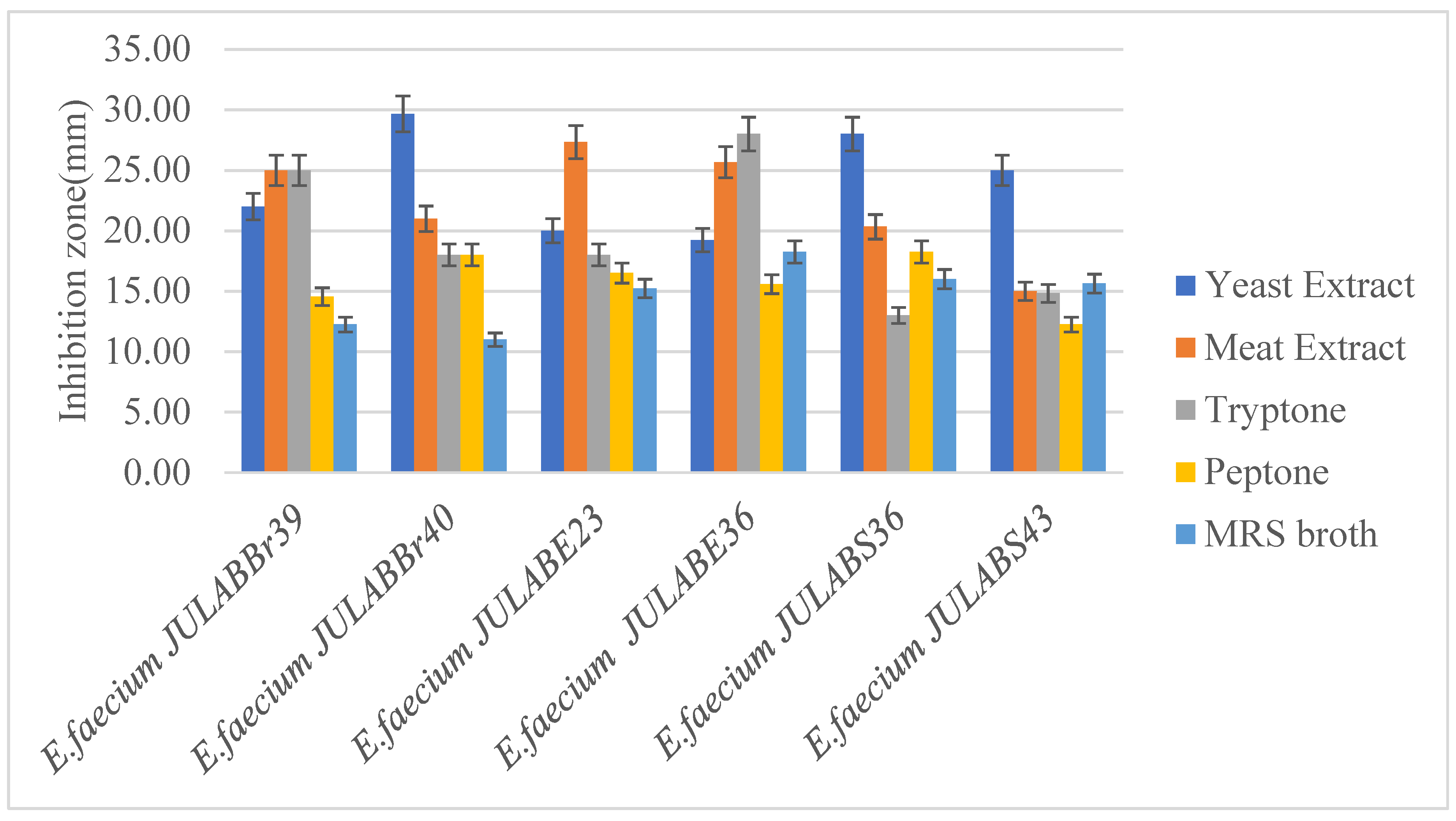

Table 1.
Sources of Enterococcus species and test pathogenic bacteria used in the study.
| Sources of isolates | Designation/code | Identity |
| EPHI* | ATCC@25922 | Escherichia. Coli |
| EPHI | ATCC@25923 | Staphylococcus aureus |
| EPHI | ATCC@7644 | Listeria monocytogens |
| EPHI | ATCC@27853 | Pseudomonas auroginoasa |
| EPHI | ATCC@13311 | Salmonella Typhimurium |
| EPHI | ATCC@ 14053 | Candida albicans |
| Borde | JULAB-Br40 | Enterococcus faecium |
| Borde | JULAB-Br39 | Enterococcus faecium |
| Ergo | JULAB-E23 | Enterococcus faecium |
| Ergo | JULAB-E36 | Enterococcus faecium |
| Cabbage-Shamita | JULAB-S43 | Enterococcus faecium |
| Cabbage-Shamita | JULAB-S36 | Enterococcus faecium |
*EPHI, Ethiopian Public Health Institute.
Table 2.
Antimicrobial activity of putative bacteriocins producing Enterococcus species against food borne pathogens.
Table 2.
Antimicrobial activity of putative bacteriocins producing Enterococcus species against food borne pathogens.
| S/N | LAB isolates |
E.coli ATCC®25922 |
S.aurues ATCC®25923 |
L.monocytogenes ATCC®7644 |
P.auroginosa ATCC®27853 |
S. Tphimurium ATCC®13311 |
C.albicans ATCC®14053 |
| 1) | JULABBE23 | 13.50±0.71d | 16.00±1.41bc | 35.31±0.44a | 20.50±0.71bc | 18.88±0.18abc | 16.13±0.18d |
| 2) | JULABBr39 | 20.50±0.71a | 17.50±0.71ab | 20.26±0.37de | 21.25±0.35a | 20.17±0.71a | 15.50±0.71ef |
| 3) | JULABBR40 | 17.75±0.35b | 17.50±0.71ab | 28.32±0.45bc | 16.13±0.18c | 15.13±0.18c | 24.38±0.53cd |
| 4) | JULABE36 | 18.50±0.71b | 18.00±0.00a | 28.25±0.35bc | 13.25±0.35de | 20.17±0.71a | 21.50±0.71c |
| 5) | JULABS36 | 20.25±0.35a | 17.50±0.71ab | 25.25±0.35c | 16.13±0.18c | 19.13±0.18ab | 19.26±0.37de |
| 6) | JULABS43 | 17.93±0.11b | 15.50±0.7ca | 20.13±0.18de | 14.13±0.18de | 18.13±0.18abc | 30.26±0.37a |
| Tetracycline | 18.75±1.06b | 17.50±0.71ab | 18.38±0.53d | 14.13±0.18de | 20.13±0.18a | 25.26±0.37cd |
Mean values denoted by the same letter within a column are not significantly different at p<0.05.
Table 3.
Antimicrobial activity of crude bacteriocins produced by Enterococcus faecium (after Organic acids and Hydrogen peroxides treatments).
Table 3.
Antimicrobial activity of crude bacteriocins produced by Enterococcus faecium (after Organic acids and Hydrogen peroxides treatments).
| Diameter of inhibition zone (mm) | ||||
| S/N | LAB isolates | E.coli ATCC@25922 | S.aurues ATCC@25923 | L.monocytogenes ATCC@7644 |
| 1) | JULABE36 | 18.5±0.00ab | 18.55±0.00b | 17.00±0.00c |
| 2) | JULABE23 | 20.5±0.18a | 18.4±0.18b | 18.22±0.18c |
| 3) | JULABS43 | 14.75±0.35c | 17.05±0.71bc | 24.00±0.71bc |
| 4) | JULABS36 | 14.5±0.18c | 17.5±0.18bc | 25.25±0.37bc |
| 5) | JULABBR40 | 18.93±0.18ab | 18.00±0.18b | 27.00±0.53b |
| 6) | JULABBR39 | 19.00±0.71a | 20.22±0.18a | 36.00±0.18a |
Note: Mean values denoted by the same letter within a column are not significantly different at p< 0.05.
Table 4.
Antimicrobial activity of bacteriocin extracted by ammonium sulfate.
|
E.coli ATCC@25922 |
S.aurues ATCC@25923 |
L.monocytogenes ATCC@7644 | |||||||||||
| S/N | LAB isolates | 40% | 50% | 60% | 80% | 40% | 50% | 60% | 80% | 40% | 50% | 60% | 80% |
| 1.00 | JULABBR39 | – | – | * | **** | – | – | * | **** | – | – | * | **** |
| 2.00 | JULABBR40 | – | – | * | **** | – | – | * | **** | – | – | * | **** |
| 3.00 | JULABE23 | – | – | * | **** | – | – | * | **** | – | – | * | **** |
| 4.00 | JULABE36 | – | – | * | **** | – | – | * | **** | – | – | * | **** |
| 5.00 | JULABS36 | – | – | * | **** | – | – | * | **** | – | – | * | **** |
| 6.00 | JULABS43 | – | – | * | **** | – | – | * | **** | – | – | * | **** |
Note: - no inhibition zone, *=<10 mm of diameter of inhibition zone, -**=10-20 diameter of inhibition zone, ***=20-30 diameter of inhibition zone, **** =>30 mm.
Table 5.
Antimicrobial activity of bacteriocin extracted with different organic solvents.
| S/N | LAB isolates | Chloroform | Iso-propaly | Ethyl Acetate | Petroleum ether | Methanol | Butanol | Ethanol |
| 1) | JULABBR39 | **** | **** | *** | ** | ** | * | * |
| 2) | JULABBR40 | **** | **** | *** | ** | ** | * | * |
| 3) | JULABE23 | **** | *** | *** | ** | * | * | * |
| 4) | JULABE36 | **** | *** | *** | ** | * | * | – |
| 5) | JULABS36 | **** | *** | *** | ** | * | * | – |
| 6) | JULABS43 | **** | *** | *** | ** | * | * | * |
Note: - no inhibition zone, *=<10 mm of diameter of inhibition zone, -**=10-20 diameter of inhibition zone, ***=20-30 diameter of inhibition zone, **** =>30 mm.
Table 6.
Antimicrobial activity of bacteriocin producing Enterococcus faecium after MRS broth supplemented with different carbon sources, against Listeria monocytogens.
Table 6.
Antimicrobial activity of bacteriocin producing Enterococcus faecium after MRS broth supplemented with different carbon sources, against Listeria monocytogens.
| Enterococcus faecium strains | Glucose | Sucrose | Maltose | Fructose | Lactose | MRS broth |
| JULABBR39 | 31.33±0.41a | 29.67±1.70a | 25.00±0.82ab | 18.00±0.82c | 16.00±0.82b | 15.67±1.70abc |
| JULABBR40 | 30.67±0.47ab | 28.00±0.82a | 27.33±0.47a | 28.00±0.00a | 17.33±0.94a | 17.00±0.82a |
| JULABE23 | 29.00±0.81ab | 25.00±0.83ab | 25.67±0.47ab | 25.00±0.85b | 15.00±0.82b | 13.67±2.36bc |
| JULABE36 | 25.00±0.82b | 22.00±0.85abc | 21.00±0.82c | 18.00±0.00c | 13.00±1.41bc | 10.67±0.94d |
| JULABS36 | 21.00±0.82bc | 18.90±0.97b | 15.00±0.74d | 13.00±0.82de | 15.33±1.89b | 16.33±1.25ab |
| JULABS43 | 17.50±0.40c | 18.33±0.48b | 20.33±0.52c | 14.83±0.24d | 12.33±0.47bd | 15.00±0.82abc |
Note: Mean values denoted by the same letter within a column are not significantly different at p< 0.05.
Table 7.
Effects of enzymes on the potency of antimicrobial activity of bacteriocins produced by Enterococcus faecium isolated from Ethiopian traditional fermented foods and beverages.
Table 7.
Effects of enzymes on the potency of antimicrobial activity of bacteriocins produced by Enterococcus faecium isolated from Ethiopian traditional fermented foods and beverages.
| S/N | LAB isolates | Pepsin (5mg/1ml) | Trypsin (5mg/1ml) |
Proteinase K (5mg/1ml) |
α-amylase | Enzyme-free cell supernatants |
| 1 | JULABBR39 | - | - | - | + | + |
| 2 | JULABBR40 | - | - | - | + | + |
| 3 | JULABE23 | - | - | - | + | + |
| 4 | JULABE36 | - | - | - | + | + |
| 5 | JULABS36 | - | - | - | + | + |
| 6 | JULABS43 | - | - | - | + | + |
Note: -= no inhibition, += positive inhibition.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



