
Submitted:
29 January 2024
Posted:
30 January 2024
You are already at the latest version
Abstract
Spodopotera frugiperda is a worldwide invasive pest that has caused major economic damage. According to classical biological control theory, natural enemies that can control invasive pests come from the area of origin as the pests that have gone through coadaptation processes. However, the new association’s approach suggests that local natural enemies are equally capable to control invasive pests. Due to the lack of data on the association of S. frugiperda and local natural enemies, research was conducted through a rapid survey to study the population density of S. frugiperda in Yogyakarta, Indonesia and to study the diversity of parasitoid associated with S. frugiperda. The results showed the difference in S. frugiperda population and parasitism rate, with 15 parasitoid species associated with S. frugiperda. Four egg parasitoids, eight larval parasitoids, and three pupal parasitoids were found to be associated with S. frugiperda for three years after it was first discovered in Indonesia. A new association was found between S. frugiperda and three parasitoid species, consisting of two egg parasitoids (Platygasteridaesp.01 and Platygasteridaesp.02) and a larval parasitoid (Eupelmidaesp.01). A relatively new association was found between S. frugiperda and two larval parasitoids (Coccygidium sp. and Stenobracon sp.), and an existing association was found between seven species including an egg parasitoid Telenomus remus, three larval parasitoids (Cotesia sp., Microplitis sp., Campoletis sp.), and three pupal parasitoids (Brachymeria lasus, B. femorata, and Charops sp.). Telenomus remus is the most dominant parasitoid, with a higher abundance and parasitism rate. This study also reported the first findings of the association of S. frugiperda with the larval parasitoid Megaselia scalaris in Indonesia. The result suggests another method for biological control using the new association approach since local natural enemies can foster quick adaptation to invasive pests.
Keywords:
Brachymeria
; Eupelmus
; host-parasite interaction
; local adaptation
; Platygasteridae
; Telenomus remus
; Stenobracon
1. Introduction
The appearance of invasive pests is a problem that requires attention because it may threaten agriculture and the variety of local species [1] and cause biotic homogenization [2]. Spodoptera frugiperda J.E. Smith (Lepidoptera: Noctuidae) is an invasive pest from America. It has become a new pest in Indonesia since early 2019 [3]. This pest has a wide range of distribution. It has now spread to 32 provinces in Indonesia, including Sumatra [4], Java [5], Kalimantan [6], and Sulawesi [7]. Spodoptera frugiperda infestations should be severely considered since a population of 0.2 to 0.8 S. frugiperda larvae/plant can decrease maize productivity by 20 to 50% [3]. Reports of damage due to S. frugiperda have been reported in several countries, such as Ethiopia and Kenya (32–47%) [8], Zimbabwe (32–48%) [9], Ghana (22–67%) [10], and Indonesia (60%) [11]. Spodoptera frugiperda has reportedly replaced the position of Asian corn borer Ostrinia furnacalis Guenée (Lepidoptera: Crambidae) as the primary pest of maize in China [12]. Rizali et al. [2] mentioned that the presence of S. frugiperda significantly decreases the intensity of attack of other lepidopteran pests and indirectly causes negative effects on the diversity of their natural enemies (particularly predators) in different maize fields in Indonesia.
A strategy to control S. frugiperda can be done using natural enemies such as parasitoids [13]. According to the classical biological control method, invasive pest control can be carried out by importing parasitoids from the original habitat of the pest, which have been proven to be potential biological agents [14]. For example, the introduction of Rodolia cardinalis Mulsant (Coleoptera: Coccinellidae) to control the cotton aphid, Icerya purchasi Maskell (Hemiptera: Margarodidae) in California [15], and the introduction of Urophora affinis Frauenfeld and U. quadrifasciata Meigen (Diptera: Tephritidae) to control spotted knapweed Centaurea maculosa Lam. and C. diffusa Lamarck in western North America [16]. In Indonesia, the introduction of biological control agents was also carried out several times, such as the introduction of Curinus coeruleus Mulsant (Coleoptera: Coccinellidae) from Hawaii to control Heteropsylla cubana Crawford (Hemiptera: Psyllidae) on mimosoid tree Leucaena leucocephala [17], the introduction of Anagyrus lopezi De Santis (Hymenoptera: Encyrtidae) from Thailand to control cassava mealybugs Phenacoccus manihoti Matile-Ferrero (Hemiptera: Pseudococcidae) [18], and the introduction of Cecidochares connexa Macquart (Diptera: Tephritidae) from Columbia to control the invasive species-siam weed Chromolaena odorata L. [19]. The latest example provides important information that 28 years after introduction, C. connexa has been reported to be associated with several local parasitoid species [20,21,22,23]. Another example is the association between leaf miner Liriomyza spp. and local parasitoids in Indonesia. Liriomyza spp. is an invasive pest found in Indonesia from America in the 1990s [24]. Liriomyza spp. was reported to be associated with 11 parasitoid species in 2000 [25], and in 2022, Liriomyza spp. was associated with 18 local parasitoid species [26]. These findings support Hokkanen and Pimentel’s old hypothesis [27] on the possibility of new associations that can be formed between herbivores and their local natural enemies.
Several studies have been reported since S. frugiperda was first reported in Indonesia. These studies were mostly focused on the presence/absence, diversity, infestation level, and ecology of S. frugiperda [5,28,29,30,31]. Research on the performance of local parasitoid species in Indonesia in parasitizing S. frugiperda on a lab scale has even been reported [32,33]. Surveys on the infestation level of S. frugiperda, the association between S. frugiperda with local parasitoids, and its associated parasitism rate have also been carried out but are limited to a specific period [2,5,34,35,36,37,38,39,40,41,42,43]. Preliminary research in Yogyakarta, Indonesia, showed a low attack rate from local parasitoids toward S. frugiperda [44]. Thus, there is scattered information regarding the possibility of an association between S. frugiperda and local parasitoids. Therefore, this research aimed to study the population density of S. frugiperda and the diversity of parasitoids associated with S. frugiperda for three years after it was first discovered in Indonesia. This is important to study as an effort to prepare local biological control agents potentially to be used in controlling S. frugiperda.
2. Materials and Methods
2.1. Sampling Location Determination
Sampling locations were determined using a purposive sampling method in four central regencies in Yogyakarta, including Sleman, Bantul, Kulon Progo, and Gunung Kidul. Parasitoid sampling activities were carried out in 17 districts of Sleman, 17 districts of Bantul, 12 districts of Kulon Progo, and 18 districts of Gunung Kidul as replication. A total of 2–3 villages were selected from each district as sampling points. From each village, a maize field was chosen as a sampling point using GPS Essentials (mictale.com), resulting in 133 sampling points (Figure 1). Sampling of parasitoids was carried out on maize fields during the vegetative phase (2–3 weeks old) because the highest S. frugiperda infestation occurs at this stage [28].
2.2. Sampling
The survey was carried out from January 2020 to May 2022. Sampling was carried out once on each field. Parasitoids were collected by collecting eggs and larvae of S. frugiperda, found on maize plants in every field. Sampling was carried out purposively by taking eggs and larvae found. The samples obtained were brought from the field to the laboratory using an insect-rearing plastic container (21 × 21 cm). The parasitoids from the eggs are placed in an Eppendorf containing 90% ethanol. Meanwhile, the hatched larvae are transferred to and kept individually in plastic cups (400 ml) containing baby corn as a food source until moths or larval/pupal parasitoids emerge. Rearing was maintained under laboratory conditions (26 ± 1 °C, 60–80% r.h.). Parasitoids that emerged were counted, recorded, and grouped based on similar morphological characteristics, then preserved in a 1.5 ml microtube filled with 70% ethanol for further identification.
2.3. Parasitoid Identification
The emerging parasitoids were identified at the Plant Protection Laboratory, Department of Agrotechnology, Universitas Muhammadiyah Yogyakarta. Identification of parasitoids was carried out by observing and matching the morphological characteristics of the parasitoids with some relevant literature [38,45,46,47]. The identified parasitoids were photographed at certain magnifications using a Leica S6E Stereo Microscope (Leica Microsystems, Germany) at the Biological Control Laboratory, Department of Plant Protection, Faculty of Agriculture, IPB University. The parasitoid photo is then processed using TrueChromeII, TCapture 5.1 software (Fuzhou Tucsen Photonics Co., Ltd., China) by adjusting the magnification size to the desired unit (mm) to get the parasitoid body size. All identified parasitoids were confirmed at the Ecology and Systematics Laboratory, Faculty of Applied Science and Technology, Ahmad Dahlan University.
Associations between S. frugiperda and parasitoids are defined as new associations when a parasitoid has never been reported in the Western Hemisphere and another region, relatively new associations when a parasitoid was present in other regions but not in the Western Hemisphere, and existing associations when a parasitoid was not originally from but currently present in Western Hemisphere.
2.4. Data Analysis
A General Linear Model (GLM) was used to analyze the total number of S. frugiperda and the total parasitism rate in each regency. Both data were subjected to stepwise simplification before GLM analysis to determine the appropriate model based on the AIC number. A GLM with Gaussian family and log link function was used to analyze the total number of S. frugiperda, while the total parasitism rate was analyzed using a GLM with Gamma family and identity link function. The mean difference of those data between each district was further tested using Tukey’s HSD at a 95% significance level. Areas with zero abundance were not included in the analysis. The GLM analysis was performed using R Statistic version 4.2.2 [48].
The parasitism rate of the egg was observed under the Nikon SMZ18 Stereo Microscope (Nikon Instruments Inc, USA). Eggs were photographed using Optilab Advance, and the number of eggs was calculated using Image Raster software (PT. Miconos, Indonesia). The parasitism rate was calculated by dividing the number of parasitized hosts by the total number of hosts. Furthermore, parasitoid distribution was mapped based on sampling points (regional administrative data) using ArcGis 10 (ESRI, Environmental Systems Research Institute, California, USA).
3. Results
According to this survey, S. frugiperda was discovered to be present in four of Yogyakarta’s central regencies, with significantly different population number (GLM: F3,42 = 9.5495, P < 0.0001). The highest population of S. frugiperda is found in Bantul. The population in Gunung Kidul is almost half of Bantul’s population. Meanwhile, the lowest populations were found in Kulonprogo and Sleman. Additionally, the parasitism rate varies significantly amongst districts (GLM: F3,42 = 3.6889, P < 0.01). However, the highest parasitism rate occurred in Kulonprogo, and the lowest occurred in Gunung Kidul. Meanwhile, the parasitism rate in Bantul and Sleman was similar (Tabel 1).
Fifteen species of parasitoids were associated with S. frugiperda. Four species were egg parasitoids, eight were larval parasitoids, and three were pupal parasitoids. The egg parasitoids found were Platygasteridae.sp01 (Figure 2a), Platygasteridae.sp02 (Figure 2b), Trichogramma sp. (Figure 2c), and Telenomus remus (Figure 2d). The larval parasitoids found were Diptera, such as Archytas marmoratus (Figure 2e) and Megaselia sp. (Figures 2f), Hymenoptera, such as Cotesia sp. (Figure 2g), Campoletis sp. (Figure 2h), Coccygidium sp. (Figure 2i), Eupelmidae.sp01 (Figure 2j), Microplitis sp. (Figure 2k), Stenobracon sp. (Figure 2l). Meanwhile, the pupal parasitoids found were Brachymeria femorata (Figure 2m), Brachymeria lasus (Figure 2n), and Charops sp. However, not all parasitoid species were found in every location. Eight species of parasitoids were found in Bantul, seven in Sleman, six in Gunung Kidul, and only three in Kulonprogo.

Table 1.
Number of S. frugiperda (egg and larvae) and total parasitism rate in different districts of Yogyakarta, Indonesia.
Table 1.
Number of S. frugiperda (egg and larvae) and total parasitism rate in different districts of Yogyakarta, Indonesia.
| District | Number of S. frugiperda (Mean ± S.E.) |
Parasitism rate (%) (Mean ± S.E.) |
N |
| Bantul | 1638.60 ± 364.99 b | 22.95 ± 5.18 ab | 17 |
| Gunung Kidul | 649.27 ± 281.13 a | 13.54 ± 6.69 a | 10 |
| Kulonprogo | 108.28 ± 41.57 ab | 54.68 ± 16.46 b | 7 |
| Sleman | 140.11 ± 27.55 ab | 23.46 ± 7.45 ab | 12 |
* Means with different letters in a column are significantly different by Tukey HSD Test (α = 0.05). S.E. Standard Error, N. Replication.
A new association was found between S. frugiperda and three parasitoid species, consisting of two egg parasitoids (Platygasteridaesp.01 and Platygasteridaesp.02) and a larval parasitoid (Eupelmidaesp.01) because these parasitoids have never been reported in another region including the Western Hemisphere. A relatively new association was found between S. frugiperda and two larval parasitoids (Coccygidium sp. and Stenobracon sp.) since these parasitoids exist elsewhere but not in the Western Hemisphere, and an existing association was found between seven species, including an egg parasitoid Telenomus remus, three larval parasitoids (Cotesia sp., Microplitis sp., Campoletis sp.), and three pupal parasitoids (Brachymeria lasus, B. femorata, and Charops sp.) because these parasitoids originated outside of the Western Hemisphere but are now found there.
Telenomus remus had the highest parasitism rate (14.74–71.97%). Relatively high parasitism rates were discovered in Microplitis sp. and Platygasteridae.sp02. Other parasitoids, such as Cotesia sp. and Stenobracon sp., had maximum parasitism rates of 16.67% and 11.54%, respectively. Meanwhile, Trichogramma sp., Coccygidium sp., Eupelmidae.sp01, B. lasus, and B. formata had a maximum parasitism rate of less than 10%. Meanwhile, the other parasitoids, including Platygasteridaesp.01, Campoletis sp., Megaselia scalaris, Archytas marmoratus, and Charops sp., had a parasitism rate of less than 5% (Table 2).
In contrast to the parasitism rate, the richest and most abundant species of parasitoids were found in Bantul. This amount is far higher than in other places. In total, 8753 parasitoids were obtained in Bantul, 2478 parasitoids in Sleman, 924 in Gunung Kidul, and 479 in Kulon Progo (Table 3).
Based on the population of S. frugiperda data and mapping analysis of the parasitoid species distribution found in the field, it can be concluded that S. frugiperda has spread almost throughout the Yogyakarta region (Figure 3). Telenomus remus was the most dominant parasitoid because of its abundance. However, T. remus was only distributed in a few areas (Figure 4), with the highest abundance found in Bantul.
4. Discussion
Our research shows the different numbers of S. frugiperda found across all regencies. Bantul has the highest abundance of S. frugiperda compared to other locations because of the large cornfield in this area [49]. Corn is also planted in the Kulonprogo and Gunung Kidul, but the corn in these areas is a fodder crop, while corn in Bantul is sweet corn. Spodoptera frugiperda prefers sweet corn to fodder corn [50]. Meanwhile, sweet corn plants in Sleman are sprayed with pesticides more often, resulting in lower S. frugiperda populations in this region.
The abundance and parasitism rates show the opposite, where the highest parasitism rate occurs in Kulonprogo, while the highest abundance is found in Bantul. This happens because of variations in the numbers of pests and parasitized pests overall. Bantul has the highest abundance of parasitoids because of the characteristics of the sampling site, where Bantul Regency serves as Yogyakarta’s primary maize-producing hub [49], making hosts (S. frugiperda) more accessible than other districts. According to Kishinevsky et al. [51], an individual parasitoid would be more prevalent in a site if its host population is more numerous, as are its hosts.
The results of this research indicate that biological control of invasive pests does not always have to be controlled using Classical Biological Control methods, which emphasizes that invasive pests are controlled by the natural enemy from the country of origin because local natural enemies cannot control the invasive pest [14], a new association between invasive pests and local natural enemies will not result in suppression/regulation of the pest because adaptation might take too long. In fact, adaptation can happen relatively quickly, as we found in this study. Elton [52] said that when a parasite species is introduced into an ecosystem with a host or hosts it has never been associated with, the parasite population often rises quickly, often to outbreak levels, and its host population is suppressed.
The result shows an increase in the number of parasitoids associated with S. frugiperda. These findings indicate a similar pattern in other areas of Indonesia. For example, one parasitoid was associated with S. frugiperda at the beginning of survey activities. According to Jindal et al. [53], no parasitoids were seen in India in 2019 because S. frugiperda had recently infected the crop in the late season. Maharani et al. [5] also reported no parasitoids directly associated with S. frugiperda in Bandung and Garut, West Java, Indonesia, 2019. However, A. marmoratus and Hymenoptera larvae were found from Mythymna separata obtained from the same field where S. frugiperda was collected. Furthermore, Pu’u and Mutiara [54] reported no parasitoids associated with S. frugiperda in Ende, East Nusa Tenggara, in 2020. Suroto et al. [39] reported one parasitoid (Apanteles sp.) associated with S. frugiperda in Banyumas, Central Java, in 2021. Then, Minarni et al. [36] reported the association of S. frugiperda with one egg parasitoid (T. remus) and three larval parasitoids from the Braconidae, Ichneumonidae, and Chalcididae families in the same location in 2022. Other studies, such as Tawakkal in 2020 [55], reported the association of S. frugiperda with six parasitoids in Bogor, West Java. This study reports 15 parasitoids associated with S. frugiperda three years after their discovery in Indonesia. Numerous factors, including the degree of pest infestation [56], geography such as landscape structure [57], and regional variations in agricultural production practices [58], all impact the diversity of parasitoids.
Some parasitoids are similar to those found in the Western Hemisphere, while others differ. Chelonus insularis (Hymenoptera: Ichneumonidae) is identified as the primary parasitoid of S. frugiperda in most investigations conducted in North, Central, and South America. However, Eiphosoma laphygmae (Hymenoptera: Icheumonidae) is more recommended as a prospective candidate for introduction because of its specificity and significance as a parasitoid of the pest across most of its natural habitat [13]. These two parasitoids were not discovered during our research. They were not associated with S. frugiperda in other investigations, including those conducted in Cameroon [59] and India [60].
The most prevalent parasitoid identified in this investigation is T. remus. This study reported similar results from other investigations [61,62], where T. remus was the dominant parasitoid for S. frugiperda because this parasitoid has a high abundance and parasitism rate (14.74-71.97%%). Kumela et al. [8] also reported that T. remus was a parasitoid of S. frugiperda eggs, with the highest parasitism rate (69.3%) in Kenya and Southern China (30-50%) [62]. Sari et al. [32] reported the potential of T. remus as a biological agent of S. frugiperda in Indonesia, with a parasitism rate of 69.40%. This value is comparable to other egg parasitoids such as T. chilotraeae [63]. The parasitism rate of T. remus may be higher in a situation with many potential hosts. Junaedi et al. [64] said that the availability of hosts for parasitoid survival could increase the parasitism rate. During sampling in the field, the population of S. frugiperda eggs was abundant. The high parasitism rate is also due to T. remus’ ability to find and recognize its host [65]. Goulart et al. [66] said that the ability of T. remus to search and recognize its hosts is better than other S. frugiperda egg parasitoids such as Trichogramma pretiosum.
Telenomus remus is a native egg parasitoid from Malaysia and Papua New Guinea [67]. Telenomus remus is a common egg parasitoid used to control pests in the Noctuidae group, especially the Spodoptera genus [68]. Telenomus remus has been introduced to many countries, including America [68], despite having little success in controlling S. frugiperda [69]. In contrast to studies in Indonesia and other countries like Africa (Benin, Cameroon, Côte d’Ivoire, Ghana, Kenya, Niger, Nigeria, Uganda, South Africa, Tanzania, and Zambia) and Asia (China, India, and Nepal) [70] which claimed that T. remus is a potential biological control agent for controlling S. frugiperda. This could occur because S. frugiperda is not native to these regions. When a parasitoid can attack a new host species, the host typically suffers greatly [71]. In contrast to America, the existence of other parasitoids cannot successfully control S. frugiperda because they have evolved to their original natural enemies, making it resistant to effective control by other parasitoids [72].
Other egg parasitoids found were Platygasteridaesp.01 and Platygasteridaesp.02. Morphologically, these parasitoid species differ from T. remus, even though they both come from the Platygasteridae family. These parasitoids are characterized by eight flagellum segments and a wider second metasoma segment, different from T. remus, which has dilation in the third metasoma [73]. Unfortunately, due to minimal sample conditions, identification could not be carried out to the genus level. Platygasteridae was found, with a lower abundance than T. remus. This might happen because the two are not the primary parasitoids of S. frugiperda eggs. Platygasteridaesp.01 has a similar morphological character to Platygaster oryzae, the main parasitoid of Asian rice gall midge Orseolia oryzae [74]. However, the species Platygasteridaesp. 01 could not be confirmed, even though this parasitoid emerged from the rearing of S. frugiperda egg clusters collected from maize fields adjacent to rice fields. Meanwhile, Platygasteridaesp.02 has a different body color from P. oryzae. Platygaster oryzae has a metallic black body color [75,76], while the parasitoids found were a bright yellow. Platygasteridaesp.02 has a fairly high parasitism rate (42%) compared to Platygasteridaesp.01. However, the parasitism incident of Platygasteridaesp.02 was only found in one egg cluster. These two egg parasitoids may not be potential candidates for S. frugiperda. Nevertheless, the discovery of two different egg parasitoid species from T. Remus and Trichogramma indicates the existence of two new associations between S. frugiperda and local egg parasitoids. Another association has been found between S. frugiperda and larval parasitoids from Hymenoptera order, including Microplitis sp., Cotesia sp., Campoletis sp., Coccygidium sp., Eupelmidaesp.01, and Stenobracon sp. Microplitis sp., Cotesia sp., and Campoletis sp. are present in the Western Hemisphere. However, Cotesia like C. ruficrus imported from the Australia [77], M. manilae from the Thailand [78], and C. chloridae from the India [79] to the US. Microplitis sp. is a larval parasitoid with the highest parasitism rate (39.7–61.29%) compared to other S. frugiperda larval parasitoids. The genus Microplitis, reported as a larval parasitoid of S. frugiperda, includes M. manilae [78]. Microplitis is a genera widely distributed throughout all biogeographic zones, with macrolepidopterans as their primary hosts [80]. Moreover, Cotesia also has a significant parasitism rate (5.02–16.67%). Supeno et al. [38] and Suroto et al. [39] also reported the incidence of parasitism of Cotesia on S. frugiperda larvae with a 17–22% parasitism rate. Association of S. frugiperda larvae with Microplitis and Cotesia was also reported from Bogor, West Java, with 12.3 and 0.39 % parasitism rates, respectively [55]. Campoletis had a 0.2–12.69% parasitism rate. Campoletis has also been reported in India, with a 2-4% parasitism rate [60]. The results of this study also indicate that Microplitis, Cotesia, and Campoletis are parasitoids of S. frugiperda larvae that potentially develop as biological control agents. Surprisingly, Coccygidium sp., Eupelmidaesp.01, and Stenobracon sp. have never been found in the western hemisphere [81]. Coccygidium had a 0.2–17.54% parasitism rate. Coccygidium was also reported in India, with a 0.001% parasitism rate [60], and in Ghana, with a 3.9–19.3% parasitism rate [45]. Stenobracon sp. was also reported from India as the predominant larval parasitoid of S. frugiperda [53]. No incidence of parasitism has ever been reported for Eupelmid families in S. frugiperda. Thus, this is a new association discovered between S. frugiperda and larval parasitoid Eupelmidaesp.01.
Apart from the Hymenoptera order, there were also S. frugiperda larvae parasitoids from the Diptera order, such as Archytas marmoratus and Megaselia scalaris. Archytas marmoratus is a potential parasitoid for S. frugiperda in the American field [82]. However, the parasitism rate found in this study was very low. The parasitism level of M. scalaris was also very low. Megalia scalaris was first reported in Asian regions, including India [83] and China [84]. Megalia scalaris was also found in the Mexican region [85]. This finding is also the first report of an association between S. frugiperda and M. scalaris in Indonesia. Megaselia scalaris is an insect found in various regions, usually in decaying organic matter [86]. Besides being reported as a larval parasitoid of S. frugiperda, M scalaris was also reported to be associated with peach fruit fly, Bactrocera zonata (Saunders), Mediterranean fruit fly, Ceratitis capitata (Wiedemann) in Egypt [87], and fruit-piercing moths Thyas coronota (Fabricius) (Lepidoptera: Erebidea) in India [88]. However, according to a recent study, M. scalaris is not recommended as a potential biological control agent for S. frugiperda. Megaselia scalaris prefers to consume deceased larvae instead of acting as an endoparasitoid with parasitism rates of 2.2 and 0.7% in third- and fifth-instar larvae of S. frugiperda, respectively [89].
Besides the association between S. frugiperda and several egg and larval parasitoids, there was another association with three pupal parasitoids, such as Brachymeria lasus and B. femorata from the Chalcididae family, and Charops sp. from Ichneumonid family. Several chalcidid families reported to be associated with S. frugiperda include B. flavipes (=robusta) (Fabricius), B. ovata (Say), Conura femorata (Fabricius), C. hirtifemora (Ashmead), C. igneoides (Kirby), C. immaculata (Cresson), and C. meteori (Burks) [81]. These two S. frugiperda pupal parasitoids were only found in Bantul. Brachymeria lasus and B. femorata usually attack hidden insects, such as the banana leafroller caterpillar Erinota thrax (Lepidoptera: Hesperiidae) [90], fire caterpillars, and other Noctuidae families. The proximity of banana plants to the sampling site in Bantul Regency suggests that there may be other hosts for these parasitoids. However, these two parasitoids have also been found parasitizing the pupae of S. frugiperda in Egypt [91]. Lastly, Charops sp. was identified through its pupal characteristics. Charops sp. was also reported as a pupal parasitoid of S. frugiperda in Cameroon [92], Ghana, and Benin [59].
Our findings support Hokkanen and Pimentel’s theory on using the New Association biological control approach for controlling S. frugiperda with local parasitoids such as T. remus and Microplitis sp. Telenomus remus has been used as a biological agent for S. exigua [93], and Microplitis manilae has also been around for a long time and is associated with other Spodoptera species, such as S. litura in Indonesia [94]. This evidence led us to conclude that these parasitoids existed earlier than S. frugiperda in Indonesia. Hokkanen and Pimental [27] also said that the original host of the most effective new association biocontrol agents is closely connected to the new host of the agent introduced. The original and subsequent hosts have typically belonged to the same genus. Longer taxonomic “jumps’’ to a new host from distinct families of hosts are also feasible, as exemplified by a pupal parasitoid B. lasus. Therefore, T. remus and Microplitis sp. are thus possible biological control agent options for use in control initiatives within the new association of biological control concepts equipped to regulate S. frugiperda.
5. Conclusions
In conclusion, this study reveals the difference in the number of S. frugiperda and the parasitism rate across regions. Fifteen parasitoids are associated with S. frugiperda, including three new associations, two relatively new associations, and seven existing associations. Our study suggests the possibilities of biological control with a new association approach using local parasitoids for controlling S. frugiperda. Moving forward, further investigations into the scalability in wider agricultural settings will be pivotal for developing sustainable and locally adapted pest control measures.
Author Contributions
Conceptualization, I.N., I.L.I.P., and D.B.; methodology, I.N., I.L.I.P., and D.B.; software, I.N., and F.S.; validation, I.N., I.L.I.P., and D.B.; formal analysis, I.N., I.L.I.P., F.S.; investigation, I.N., and I.L.I.P.; resources, I.N., I.L.I.P, and F.S.; data curation, I.N., and F.S.; writing—original draft preparation, I.N., I.L.I.P, and F.S.; writing—review and editing, I.N., I.L.I.P., F.S. and D.B.; visualization, I.N., and F.S.; supervision, D.B.; project administration, I.N. and I.L.I.P.; funding acquisition, I.N. and I.L.I.P. All authors have read and agreed to the published version of the manuscript.
Funding
The Plant Protection Laboratory, UMY, and Zoology Laboratory UAD funded this research and publication. The publication process was also partially supported by the World Class Professor program from the Directorate General of Higher Education (Ditjen Dikti) of the Ministry of Education and Culture, Republic of Indonesia, in 2023.
Data Availability Statement
The data that support the findings of this study are available on request from the author (I.N.).
Acknowledgments
The authors thank M. Agung Faturohman, Annisa Harmaningtyas Novinda Putri, Siti Munawarah, Ahmad Nguzairon, Nuriya Laelatus Sifa’iyah, Anggarsih Triyono, and Izaz Hadaya Amajid for their help in collecting sample. The authors also thank Risa Rahma Dewi, Lidia Sari, and Meri Eliza for their assistance in imaging the parasitoid samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hill, M.P.; Clusella-Trullas, S.; Terblanche, J.S.; Richardson, D.M. Drivers, impacts, mechanisms and adaptation in insect invasions. Biological Invasion 2016, 18, 883–891. [Google Scholar] [CrossRef]
- Rizali, A.; Oktaviyani, O.; Putri, S.; Doananda, M.; Linggani, A. Invasion of fall armyworm Spodoptera frugiperda, a new invasive pest, alters native herbivore attack intensity and natural enemy diversity. Biodiversitas Journal of Biological Diversity 2021, 22, 3482–3488. [Google Scholar] [CrossRef]
- Nonci, N.; Kalqutny, S.H.; Muis, A.; Azrai, M.; Aqil, M. Pengenalan Fall Armyworm (Spodoptera frugiperda JE Smith) Hama Baru pada Tanaman Jagung di Indonesia (in Indonesian); Balai Penelitian Tanaman Serealia: 2019.
- Girsang, S.S.; Nurzannah, S.E.; Girsang, M.A.; Effendi, R. The distribution and impact of fall army worm (Spodoptera frugiperda) on maize production in North Sumatera. In Proceedings of the International Conference on Sustainable Cereals and Crops Production Systems in the Tropics Makassar, Indonesia, 2020; p. 012099. [CrossRef]
- Maharani, Y.; Dewi, V.K.; Puspasari, L.T.; Rizkie, L.; Hidayat, Y.; Dono, D. Cases of fall army worm Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) attack on maize in Bandung, Garut and Sumedang district, West Java. CROPSAVER-Journal of Plant Protection 2019, 2, 38–46. [Google Scholar] [CrossRef]
- Subiono, T. Preferences of Spodoptera frugiperda (Lepidoptera: Noctuidae) in several feed sources (in Indonesian). Jurnal Agroekoteknologi Tropika Lembab 2020, 2, 130–134. [Google Scholar] [CrossRef]
- Noer, H. Population and attack rate Spodoptera frugiperda on corn plants in Tulo Village, Sigi Regency (in Indonesian). Jurnal Agrotech 2020, 10, 66–68. [Google Scholar] [CrossRef]
- Kumela, T.; Simiyu, J.; Sisay, B.; Likhayo, P.; Mendesil, E.; Gohole, L.; Tefera, T. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. International Journal of Pest Management 2018, 65, 1–9. [Google Scholar] [CrossRef]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda JE Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Protection 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J. Fall armyworm: impacts and implications for Africa. Outlooks on Pest Management 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Lubis, A.A.N.; Anwar, R.; Soekarno, B.P.; Istiaji, B.; Dewi, S.; Herawati, D. Coray wood corn (Spodoptera frugiperda) caterpillars in corn crops in Petir Village, Daramaga Sub-District, Bogor District, and its control potential using Metarizhium rileyi (in Indonesian). Jurnal Pusat Inovasi Masyarakat 2020, 2, 931–939. [Google Scholar]
- Song, Y.; Yang, X.; Li, H.; Wu, K. The invasive Spodoptera frugiperda (JE Smith) has displaced Ostrinia furnacalis (Guenée) as the dominant maize pest in the border area of southwestern China. Pest Management Science 2023, 79, 3354–3363. [Google Scholar] [CrossRef] [PubMed]
- Kenis, M. Prospects for classical biological control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in invaded areas using parasitoids from the Americas. Journal of Economic Entomology 2023, 116, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Hajek, A.E. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: 2012. [CrossRef]
- Caltagirone, L.E.; Doutt, R.L. The history of the vedalia beetle importation to California and its impact on the development of biological control. Annual Review of Entomology 1989, 34, 1–16. [Google Scholar] [CrossRef]
- Winston, R.; Schwarzländer, M.; Hinz, H.L.; Day, M.D.; Cock, M.J.; Julien, M. Biological control of weeds: A world catalogue of agents and their target weeds; USDA Forest Service, Forest Health Technology Enterprise Team: 2014.
- USAID. Environmental Assessment for Implementation of Biological Control for The Leucaena Psyllid in Asia and Africa: Case Studies from India, Indonesia, Laos, Malaysia, Nepal, the Philippines and Thailand; Center for the Environment: Washington DC, 1995. [Google Scholar]
- Wyckhuys, K.A.G.; Rauf, A.; Ketelaar, J. Parasitoids introduced into Indonesia: part of a region-wide campaign to tackle emerging cassava pests and diseases. Biocontrol News and Information 2014, 35, 35N–37N. [Google Scholar]
- Tjitrosemito, S. The establishment of Procecidochares connexa in West Java, Indonesia: a biological control agent of Chromolaena odorata. Biotropia - The Southeast Asian Journal of Tropical Biology 1999, 12, 19–24. [Google Scholar] [CrossRef]
- Buchori, D.; Rizali, A.; Lukvitasari, L.; Triwidodo, H. Insect communities associated with siam weed: evaluation after three decades of Cecidochares connexa release as biocontrol agent. Diversity 2020, 12, 344. [Google Scholar] [CrossRef]
- Lukvitasari, L.; Triwidodo, H.; Rizali, A.; Buchori, D. Effects of locations on gall fly Cecidochares connexa (Macquart) attacks on invasive exotic plant Chromolaena odorata (L.) King & Robinson and its interaction with local insect communities (in Indonesian). Jurnal Entomologi Indonesia 2021, 18, 127–139. [Google Scholar] [CrossRef]
- McFadyen, R.E.C.; Desmier de Chenon, R.; Sipayung, A. Biology and host specificity of the Chromolaena stem gall fly, Cecidochares connexa (Macquart)(Diptera: Tephritidae). Australian Journal of Entomology 2003, 42, 294–297. [Google Scholar] [CrossRef]
- Safi’i, I. Parasitisasi pada Cecidochares connexa di beberapa tempat di Jawa Barat. Undergraduate Thesis, IPB University, Bogor, 2006. [Google Scholar]
- Rauf, A. Liriomyza: hama pendatang baru di Indonesia (in Indonesia). Buletin HPT 1995, 8, 46–48. [Google Scholar]
- Rauf, A.; Shepard, B.M.; Johnson, M.W. Leafminers in vegetables, ornamental plants and weeds in Indonesia: surveys of host crops, species composition and parasitoids. International Journal of Pest Management 2000, 46, 257–266. [Google Scholar] [CrossRef]
- Tapahillah, T. Survei lalat pengorok daun Liriomyza spp. (Diptera: Agromyzidae) dan parasitoidnya pada berbagai tumbuhan inang dan ketinggian tempat di Jawa Barat. Undergraduate Thesis, IPB University, Bogor, 2002. [Google Scholar]
- Hokkanen, H.M.; Pimentel, D. New associations in biological control: theory and practice. The Canadian Entomologist 1989, 121, 829–840. [Google Scholar] [CrossRef]
- Trisyono, Y.A.; Suputa, S.; Aryuwandari, V.E.F.; Hartaman, M.; Jumari, J. Occurrence of heavy infestation by the fall armyworm Spodoptera frugiperda, a new alien invasive pest, in corn Lampung Indonesia. Jurnal Perlindungan Tanaman Indonesia 2019, 23, 156–160. [Google Scholar] [CrossRef]
- Megasari, D.; Putra, I.L.I.; Martina, N.D.; Wulanda, A.; Khotimah, K. Biology of Spodoptera frugiperda JE Smith on some types of feed in the laboratory (in Indonesian). Agrovigor: Jurnal Agroekoteknologi 2022, 15, 63–67. [Google Scholar] [CrossRef]
- Putra, I.L.I.; Martina, N.D. Life cycle of Spodoptera frugiperda with feeding kale and leeks in laboratory (in Indonesian). Jurnal Ilmu Pertanian Indonesia 2021, 26, 386–391. [Google Scholar] [CrossRef]
- Nurkomar, I.; Trisnawati, D.W.; Fahmi, F.; Buchori, D. Survival, development, and fecundity of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) on various host plant species and their implication for the pest management. Insects 2023, 14, 629. [Google Scholar] [CrossRef]
- Sari, A.; Buchori, D.; Nurkomar, I. The potential of Telenomus remus Nixon (Hymenoptera: Scelinoidae) as biocontrol agent for the new fall armyworm S. frugiperda (Lepidoptera: Noctuidae) in Indonesia. Planta Tropika 2020, 8, 69–74. [Google Scholar] [CrossRef]
- Putra, I.L.I.; Wati, C.D.N.S. Parasicity level of Telenomus sp. parasitoid against Spodoptera frugiperda JE Smith eggs in the laboratory. Journal of Natural Sciences and Mathematics Research 2020, 6, 73–77. [Google Scholar] [CrossRef]
- Polem, E.S.N. Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae): Serangan dan Parasitoidnya di Nagan Raya, Aceh (in Indonesian). Undergraduate Thesis, IPB University, Bogor, 2021. [Google Scholar]
- Rongkok, H.T.; Pasaru, F. Identification of paracitoids of Spodoptera frugiperda (Lepidoptera: Noctuidae) and its paracity levels in farmer-owned corn plantation in District Sigi and in District Donggala (in indonesian). Jurnal Agrotekbis 2021, 9, 972–978. [Google Scholar]
- Minarni, E.W. Eksplorasi musuh alami hama Spodoptera frugiperda pada pertanaman jagung di lahan kelompok tani raden Kelurahan Pabuwaran Kecamatan Purwokerto Utara Kabupaten Banyumas (in Indonesia). In Proceedings of the Prosiding Seminar Nasional LPPM Unsoed; 2023; pp. 139–147. [Google Scholar]
- Sari, W.; Nelly, N. Natural enemies of Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) on corn plants in West Sumatera. In Proceedings of the 2nd Agrifood System International Conference Padang, Indonesia; 2023; p. 012045. [Google Scholar] [CrossRef]
- Supeno, B.; Tarmizi, T.; Meidiwarman, M.; Haryanto, H. Keragaman parasitoid yang berasosiasi dengan telur hama baru Spodoptera frugiperda di Pulau Lombok (in Indonesian). Prosiding SAINTEK 2021, 3, 418–423. [Google Scholar]
- Suroto, A.; Soesanto, L.; Bahrudin, M. Attack rate and natural enemies of Spodoptera frugiperda JE Smith on corn plants in five districts in Banyumas regency (in Indonesian). Proceedings Series on Physical & Formal Sciences 2021, 2, 44–49. [Google Scholar] [CrossRef]
- Tarigan, S.; Maryana, N.; Mubin, N. Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae): Attacks and their natural enemies on corn plantations in Munte Village, Munte Sub-district, Karo District, North Sumatera. In Proceedings of the International Conference on Modern and Sustainable Agriculture 2022; 2023; p. 012030. [Google Scholar] [CrossRef]
- Tawakkal, M.; Buchori, D.; Maryana, N. New association between Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) and native natural enemies: Bioprospection of native natural enemies as biological control agents. In Proceedings of the IOP Conference Series: Earth and Environmental Science; 2021; p. 012030. [Google Scholar] [CrossRef]
- Wahyuningsih, R.D.; Harjaka, T.; Suputa, S.; Trisyono, Y.A. Parasitization levels of Spodoptera frugiperda eggs (Smith)(Lepidoptera: Noctuidae) in three different corn ecosystems in East Java. Jurnal Perlindungan Tanaman Indonesia 2022, 26, 28–39. [Google Scholar] [CrossRef]
- Waruwu, A.; Tobing, M.C.; Siregar, A.Z. Ekplorasi parasitoid telur Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) pada tanaman jagung di Desa Purwobinangun Kec. Sei Bingai, Kab. Langkat (in Indonesian). Jurnal Online Agroteknologi 2023, 11, 10–18. [Google Scholar] [CrossRef]
- Nurkomar, I.; Putra, I.; Trisnawati, D.; Saman, M.; Pangestu, R.; Triyono, A. The existence and population dynamic of new fall armyworm species Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) in Yogyakarta, Indonesia. In Proceedings of the The 3rd International COnference on Sustainable Agriculture, Yogyakarta, Indonesia, 2021; p. 012023. [CrossRef]
- Agboyi, L.K.; Mensah, S.A.; Clottey, V.A.; Beseh, P.; Glikpo, R.; Rwomushana, I.; Day, R.; Kenis, M. Evidence of leaf consumption rate decrease in fall armyworm, Spodoptera frugiperda, larvae parasitized by Coccygidium luteum. Insects 2019, 10, 410. [Google Scholar] [CrossRef]
- Shylesha, A.; Jalali, S.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K. Studies on new invasive pest Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae) and its natural enemies. Journal of Biological control 2018, 32, 1–7. [Google Scholar] [CrossRef]
- Wei, Y.-W.; Zhou, Y.-B.; Zou, Q.-C.; Sheng, M.-L. A new species of Campoletis Förster (Hymenoptera, Ichneumonidae) with a key to species known from China, Japan and South Korea. ZooKeys 2020, 1004, 99–108. [Google Scholar] [CrossRef]
- RCoreTeam. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, 2015. [Google Scholar]
- DinasPerindustrianPerdagangandanKoperasiKabupatenBantul. Prospektus Investasi Sektor Pangan Kabupaten Bantul (in Indonesian). Available online: https://dpmptsp.bantulkab.go.id/web/potensi_investasi/detail/19-sektor-pangan (accessed on 12 September).
- Sunari, A.A.A.A.S.; Sumiartha, I.K.; Supartha, I.W.; Mahaputra, I.F.; Yudha, I.K.W.; Utama, I.W.E.K.; Wiradana, P.A. Potential yield reduction of sweet and glutinous corn varieties damaged by the invasive pest Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae): Field trial scale. In Proceedings of the 2nd International Conference on Education and Technology (ICETECH 2021); 2022; pp. 14–19. [Google Scholar] [CrossRef]
- Kishinevsky, M.; Keasar, T.; Bar-Massada, A. Parasitoid abundance on plants: Effects of host abundance, plant species, and plant flowering state. Arthropod-Plant Interactions 2017, 11, 155–161. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Elton, CS 1958: The ecology of invasions by animals and plants. London: Methuen. Progress in Physical Geography 2007, 31, 659–666. [Google Scholar] [CrossRef]
- Jindal, J.; Sharma, K.P.; Shera, P.S.; Cheema, H.K. Native Parasitoids of Fall Army Worm Spodoptera frugiperda (J E Smith) in Maize. Indian Journal of Entomology 2022, 84, 865–867. [Google Scholar] [CrossRef]
- Pu’u, Y.M.; Mutiara, C. Attacks of invasive pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera:Noctuidae) on corn plantation in Ende District Flores, Indonesia [in Indonesian]. Jurnal Entomologi Indonesia 2021, 18, 153–153. [Google Scholar] [CrossRef]
- Tawakkal, M.; Buchori, D.; Maryana, N. New association between Spodoptera frugiperda JE Smith (Lepidoptera: Noctuidae) and native natural enemies: Bioprospection of native natural enemies as biological control agents. In Proceedings of the 2nd International Symposium on Transdisciplinarity Approach for Knowledge Co-Creation in Sustainability - Understanding Complexity and Transdisciplinarity for Environmental Sustainability Bogor, Indonesia; 2021; p. 012030. [Google Scholar] [CrossRef]
- Salazar-Mendoza, P.; Peralta-Aragón, I.; Romero-Rivas, L.; Salamanca, J.; Rodriguez-Saona, C. The abundance and diversity of fruit flies and their parasitoids change with elevation in guava orchards in a tropical Andean forest of Peru, independent of seasonality. Plos one 2021, 16, e0250731. [Google Scholar] [CrossRef]
- Syahidah, T.; Prasetyo, L.; Buchori, D. Landscape structure and parasitoid diversity as measures of sustainable landscape. In Proceedings of the IOP Conference Series: Earth and Environmental Science; 2019; p. 012012. [Google Scholar] [CrossRef]
- Schoeninger, K.; Souza, J.L.; Krug, C.; Oliveira, M.L. Diversity of parasitoid wasps in conventional and organic guarana (Paullinia cupana var. sorbilis) cultivation areas in the Brazilian Amazon. Acta Amazonica 2019, 49, 283–293. [Google Scholar] [CrossRef]
- Abang, A.; Nanga, S.; Fotso Kuate, A.; Kouebou, C.; Suh, C.; Masso, C.; Saethre, M.; Fiaboe, K. Natural Enemies of Fall Armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Different Agro-Ecologies. Insects 2021, 12, 509. Insect 2021, 12, 509. [Google Scholar] [CrossRef]
- Sharanabasappa, S.; Kalleshwaraswamy, C.; Poorani, J.; Maruthi, M.; Pavithra, H.; Diraviam, J. Natural enemies of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), a recent invasive pest on maize in South India. The Florida Entomologist 2019, 102, 619–623. [Google Scholar] [CrossRef]
- Kenis, M.; Du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G. Telenomus remus, a candidate parasitoid for the biological control of Spodoptera frugiperda in Africa, is already present on the continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef]
- Liao, Y.-L.; Yang, B.; Xu, M.-F.; Lin, W.; Wang, D.-S.; Chen, K.-W.; Chen, H.-Y. First report of Telenomus remus parasitizing Spodoptera frugiperda and its field parasitism in southern China. Journal of Hymenoptera Research 2019, 73, 95–102. [Google Scholar] [CrossRef]
- Sari, A.; Nurkomar, I.; Buchori, D. Comparison of Spodoptera frugiperda parasitoid performance under laboratory conditions. In Proceedings of the 3rd International Symposium on Transdisciplinary Approach for Knowledge Co-Creation in Sustainability, Bogor, Indonesia, 2023; p. 012031. [CrossRef]
- Junaedi, E.; Yunus, M.; Hasriyanty, H. Parasitoids and it is parasitisme on white rice stem borer (Scirpophaga innotata Walker) in two different altitudes of rice fields (Oryza sativa l.) in District of Sigi (in Indonesian). e-Journal Agrotekbis 2016, 4, 280–287. [Google Scholar]
- Legault, S.; Hébert, C.; Blais, J.; Berthiaume, R.; Bauce, E.; Brodeur, J. Seasonal ecology and thermal constraints of Telenomus spp.(Hymenoptera: Scelionidae), egg parasitoids of the hemlock looper (Lepidoptera: Geometridae). Environmental Entomology 2012, 41, 1290–1301. [Google Scholar] [CrossRef]
- Goulart, M.M.P.; Bueno, A.d.F.; Bueno, R.C.O.d.F.; Diniz, A.F. Host preference of the egg parasitoids Telenomus remus and Trichogramma pretiosum in laboratory. Revista Brasileira de Entomologia 2011, 55, 129–133. [Google Scholar] [CrossRef]
- Hernandez, D.; Ferrer, F.; Linares, B. Introducción de Telenomus remus Nixon (Hym.: Scelionidae) para controlar Spodoptera frugiperda (Lep.: Noctuidae) en Yaritagua-Venezuela. Agronomía Tropical 1989, 39, 45–61. [Google Scholar]
- Cave, R.D. Biology, ecology and use in pest management of Telenomus remus. Biocontrol News and Information 2000, 21, 21N–26N. [Google Scholar]
- Colmenarez, Y.C.; Babendreier, D.; Ferrer Wurst, F.R.; Vásquez-Freytez, C.L.; de Freitas Bueno, A. The use of Telenomus remus (Nixon, 1937)(Hymenoptera: Scelionidae) in the management of Spodoptera spp.: potential, challenges and major benefits. CABI Agriculture and Bioscience 2022, 3, 1–13. [Google Scholar] [CrossRef]
- Copeland, R. Fall armyworm natural enemy of the day: Telenomus remus (Hymenoptera: Scelionidae). Available online: http://www.icipe.org/content/fall-armyworm-natural-enemy-day-telenomus-remus (accessed on 12 September).
- Chandler, A.C.; Read, C. Introduction to Parasitology; John Willey and Sons: New York, USA, 1961. [Google Scholar]
- Pimentel, D. Introducing parasites and predators to control native pests. The Canadian Entomologist 1963, 95, 785–792. [Google Scholar] [CrossRef]
- Polaszek, A.; Kimani, S.W. Telenomus species (Hymenopetra: Scelionidae) attacking eggs of pyralid pests (Lepidoptera) in Africa: A review and guide to identification. Bulletin of Entomological Research 1990, 80, 57–71. [Google Scholar] [CrossRef]
- Kalyanasundaram, M.; Kamala, I.M. Parasitoids. In Ecofriendly Pest Management for Food Security; 2016; pp. 109-138. [CrossRef]
- Buhl, P.N. Species of Platygastrinae and Sceliotrachelinae from rainforest canopies in Tanzania, with keys to the Afrotropical species of Amblyaspis, Inostemma, Leptacis, Platygaster and Synopeas (Hymenoptera, Platygastridae). Tijdschrift voor Entomologie 2011, 154, 75–126. [Google Scholar] [CrossRef]
- Buhl, P.N. Key to Platygaster (Hymenoptera: Platygastridae) from Denmark, with descriptions of new species. Steenstrupia 2006, 29, 127–167. [Google Scholar]
- McCutcheon, G.S.; Salley, W.; Turnipseed, S. Biology of Apanteles ruficrus, an imported parasitoid of Pseudoplusia includens, Trichoplusia ni, and Spodoptera frugiperda (Lepidoptera: Noctuidae). Environmental Entomology 1983, 12, 1055–1058. [Google Scholar] [CrossRef]
- Rajapakse, R.H.; Ashley, T.R.; Waddill, V.H. Biology and host acceptance of Microplitis manilae (Hymenoptera: Braconidae) raised on fall armyworm larvae Spodoptera frugiperda (Lepidoptera: Noctuidae). Florida Entomologist 1985, 68, 653–657. [Google Scholar] [CrossRef]
- Alam, M. Attempts at the biological control of major insect pests of maize in Barbados, WI. Proceeding of the Caribbean Food Crops Society 1978, 15, 127–135. [Google Scholar] [CrossRef]
- Whitfield, J.B.; Austin, A.D.; Fernandez-Triana, J.L. Systematics, biology, and evolution of microgastrine parasitoid wasps. Annual Review of Entomology 2018, 63, 389–406. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Meagher, R.L. The Spodoptera frugiperda host strains: What they are and why they matter for understanding and controlling this global agricultural pest. Journal of Economic Entomology 2022, 115, 1729–1743. [Google Scholar] [CrossRef]
- Gross, H.R. Little-known fly-promising biocontrol weapon. Agricultural Research 1985, 33, 12. [Google Scholar]
- Deshmukh, S.S.; Kiran, S.; Naskar, A.; Pradeep, P.; Kalleshwaraswamy, C.; Sharath, K. First record of a parasitoid, Megaselia (M) scalaris (Diptera: Phoridae) of fall armyworm, Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae) from India. Egyptian Journal of Biological Pest Control 2021, 31, 94. [Google Scholar] [CrossRef]
- Tang, Y.; Li, Q.; Xiang, L.; Gu, R.; Wu, Y.; Zhang, Y.; Bai, X.; Niu, X.; Li, T.; Wei, J. First report on Megaselia scalaris Loew (Diptera: Phoridae) infestation of the invasive pest Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in China. Insects 2021, 12, 65. [Google Scholar] [CrossRef]
- Ruíz-Nájera, R.E.; Molina-Ochoa, J.; Carpenter, J.E.; Espinosa-Moreno, J.A.; Ruíz-Nájera, J.A.; Lezama-Gutierrez, R.; Foster, J.E. Survey for hymenopteran and dipteran parasitoids of the fall armyworm (Lepidoptera: Noctuidae) in Chiapas, Mexico. Journal of Agricultural and Urban Entomology 2007, 24, 35–42. [Google Scholar] [CrossRef]
- Disney, R.H.L. Natural history of the scuttle fly, Megaselia scalaris. Annuual Review of Entomology 2008, 53, 39–60. [Google Scholar] [CrossRef]
- Daif, M.K.; Mosallam, A.; Ebrahim, A. The scuttle fly, Megaselia scalaris (Loew, 1866)(Diptera: Phoridae): A new threat on laboratory aass production of fruit flies. Journal of Plant Protection and Pathology 2023, 14, 109–113. [Google Scholar] [CrossRef]
- Rajan, R.; Shamsudeen, R. First record of Megaselia scalaris (Diptera: Phoridae) as a parasitoid of Thyas coronata (Lepidoptera: Erebidae). Indian Journal of Entomology 2023, 1–3. [Google Scholar] [CrossRef]
- Acevedo-Alcalá, A.; Lomeli-Flores, J.R.; Rodríguez-Leyva, E.; Rodríguez-Rodríguez, S.E.; Ortiz-Andrade, E. Does Megaselia scalaris (Diptera: Phoridae) have potential as a biological control agent of fall armyworm? Florida Entomologist 2023, 106, 56–58. [Google Scholar] [CrossRef]
- Wibowo, L. The abundance and diversity of the parasitoid of banana leaf skipper pest (Erionota thrax L.) in South Lampung Regency (in Indonesian). Jurnal Hama dan Penyakit Tumbuhan Tropika 2015, 15, 26–32. [Google Scholar] [CrossRef]
- Fawzi, F.S.; Iman, I.a.-S.; Adil, A.H.M., Abd al-Salam. Survey and distribution density of genus Brachymeria species (Hymenoptera: Chalcididae) in Egypt. Egyptian Journal of Plant Protection Research Institute 2020, 3, 415-432.
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Day, R. Parasitoid complex of fall armyworm, Spodoptera frugiperda, in Ghana and Benin. Insects 2020, 11, 68. [Google Scholar] [CrossRef] [PubMed]
- Buchori, D.; Herawati, E.; Sari, A. The effectiveness of Telenomus remus (Nixon)(Hymenoptera: Scelionidae) for controlling welsh onion pest Spodoptera exigua Hübner (Lepidoptera: Noctuidae) (in indonesian). Jurnal Entomologi Indonesia 2008, 5, 81–95. [Google Scholar] [CrossRef]
- Ratna, E.S. Efficiency of parasitization on larval host, Spodoptera litura (F.), by an endoparasitoid Snellenius (=Microplitis) manilae Ashmead in the laboratory (in Indonesian). Jurnal Hama dan Penyakit Tumbuhan Tropika 2008, 8, 8–16. [Google Scholar] [CrossRef]
Figure 1.
Sampling point of S. frugiperda parasitoid in Yogyakarta.
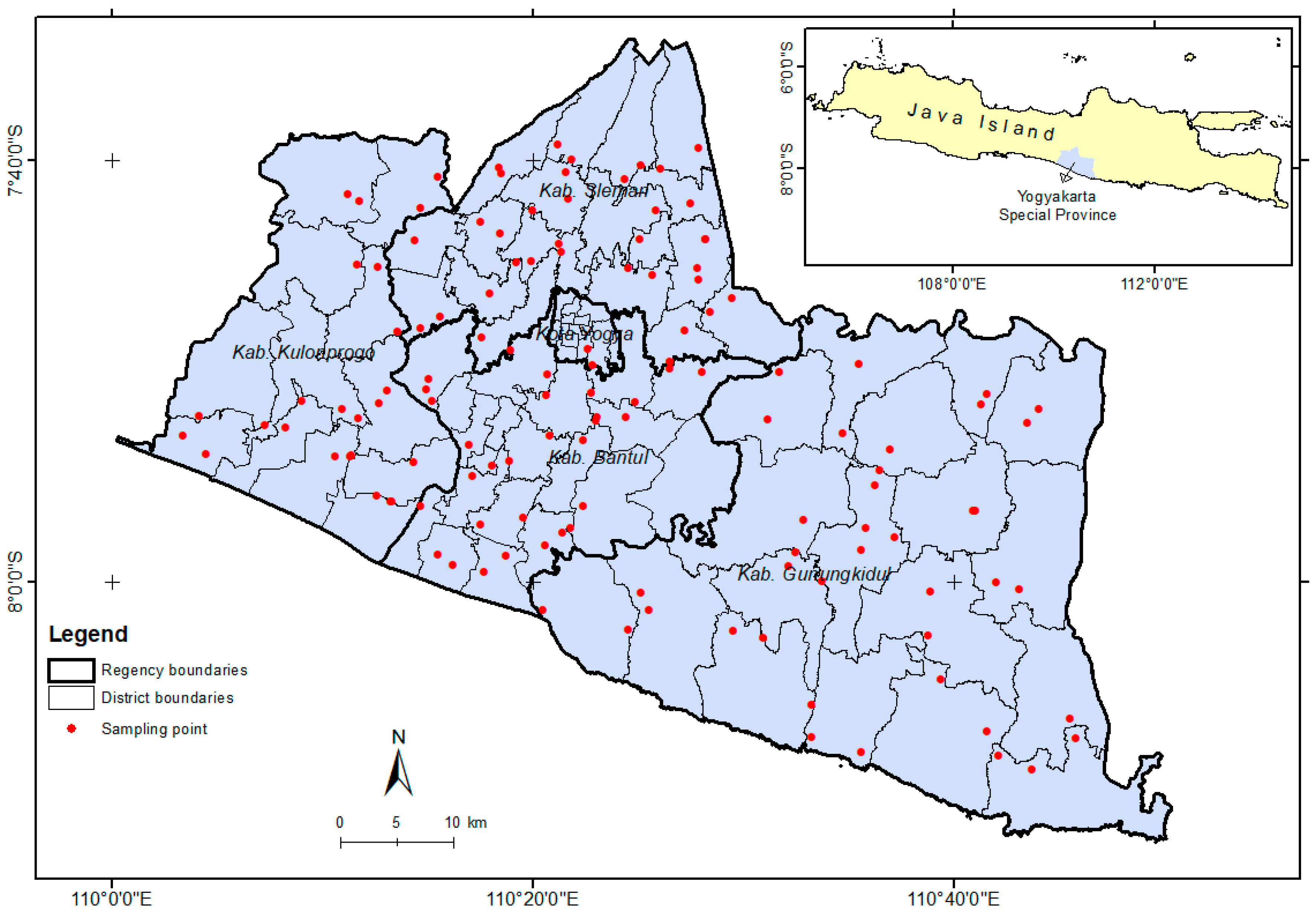
Figure 2.
Parasitoid of Spodoptera frugiperda in Special Region Yogyakarta, Indonesia. a. Platygasteridae.sp01, b. Platygasteridae.sp02, c. Trichogramma sp., d. Telenomus remus. e. Archytas marmoratus, f. Megaselia scalaris, g. Cotesia sp., h. Campoletis sp., i. Coccygidium sp., j. Eupelmidae.sp01, k. Microplitis sp., l. Stenobracon sp. m. Brachymeria femorata, and n. Brachymeria lasus.
Figure 2.
Parasitoid of Spodoptera frugiperda in Special Region Yogyakarta, Indonesia. a. Platygasteridae.sp01, b. Platygasteridae.sp02, c. Trichogramma sp., d. Telenomus remus. e. Archytas marmoratus, f. Megaselia scalaris, g. Cotesia sp., h. Campoletis sp., i. Coccygidium sp., j. Eupelmidae.sp01, k. Microplitis sp., l. Stenobracon sp. m. Brachymeria femorata, and n. Brachymeria lasus.

Figure 3.
Distribution of Spodoptera frugiperda’s parasitoid in Special Region Yogyakarta, Indonesia.
Figure 3.
Distribution of Spodoptera frugiperda’s parasitoid in Special Region Yogyakarta, Indonesia.
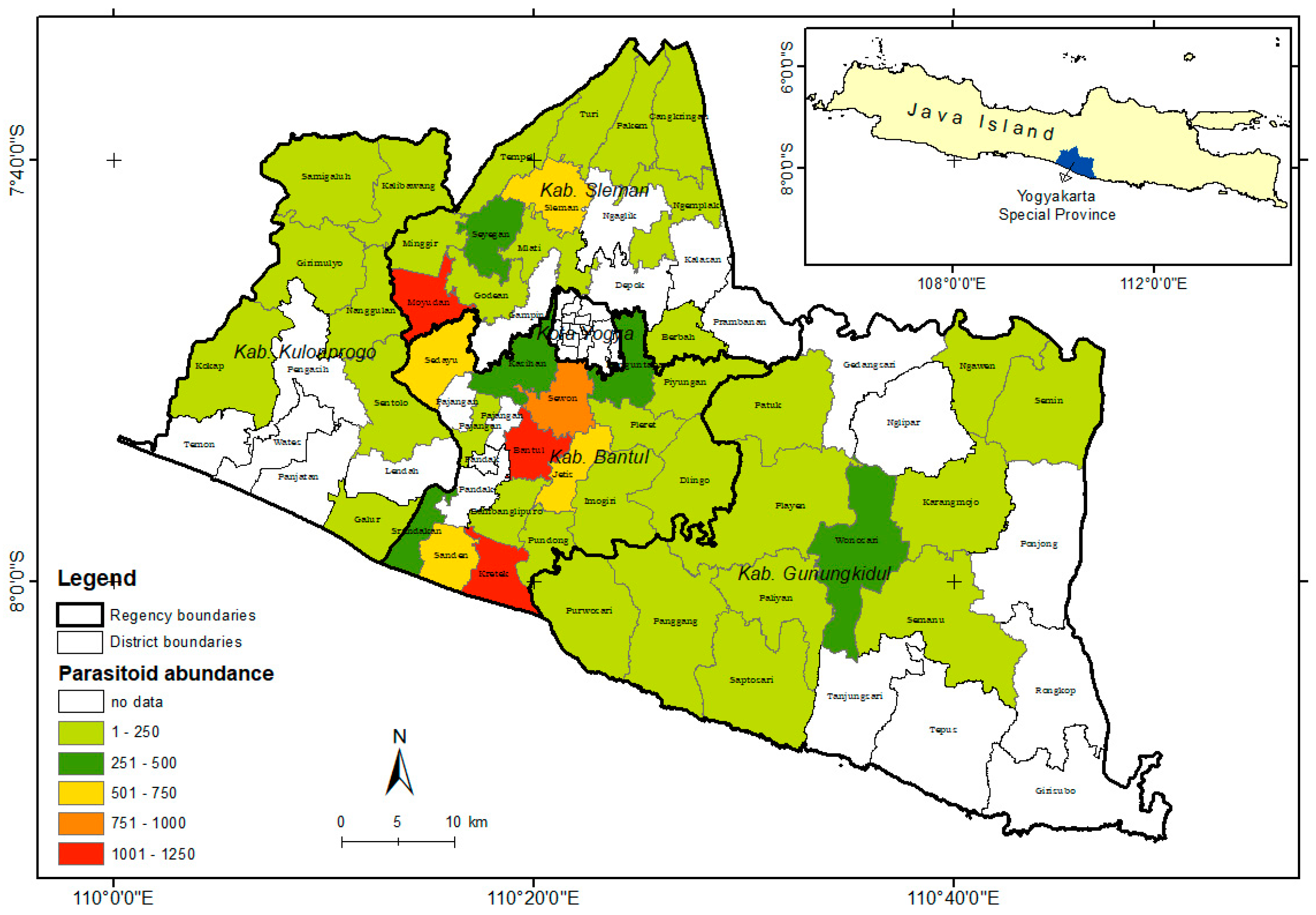
Figure 4.
Distribution of Telenomus remus in Special Region Yogyakarta, Indonesia.
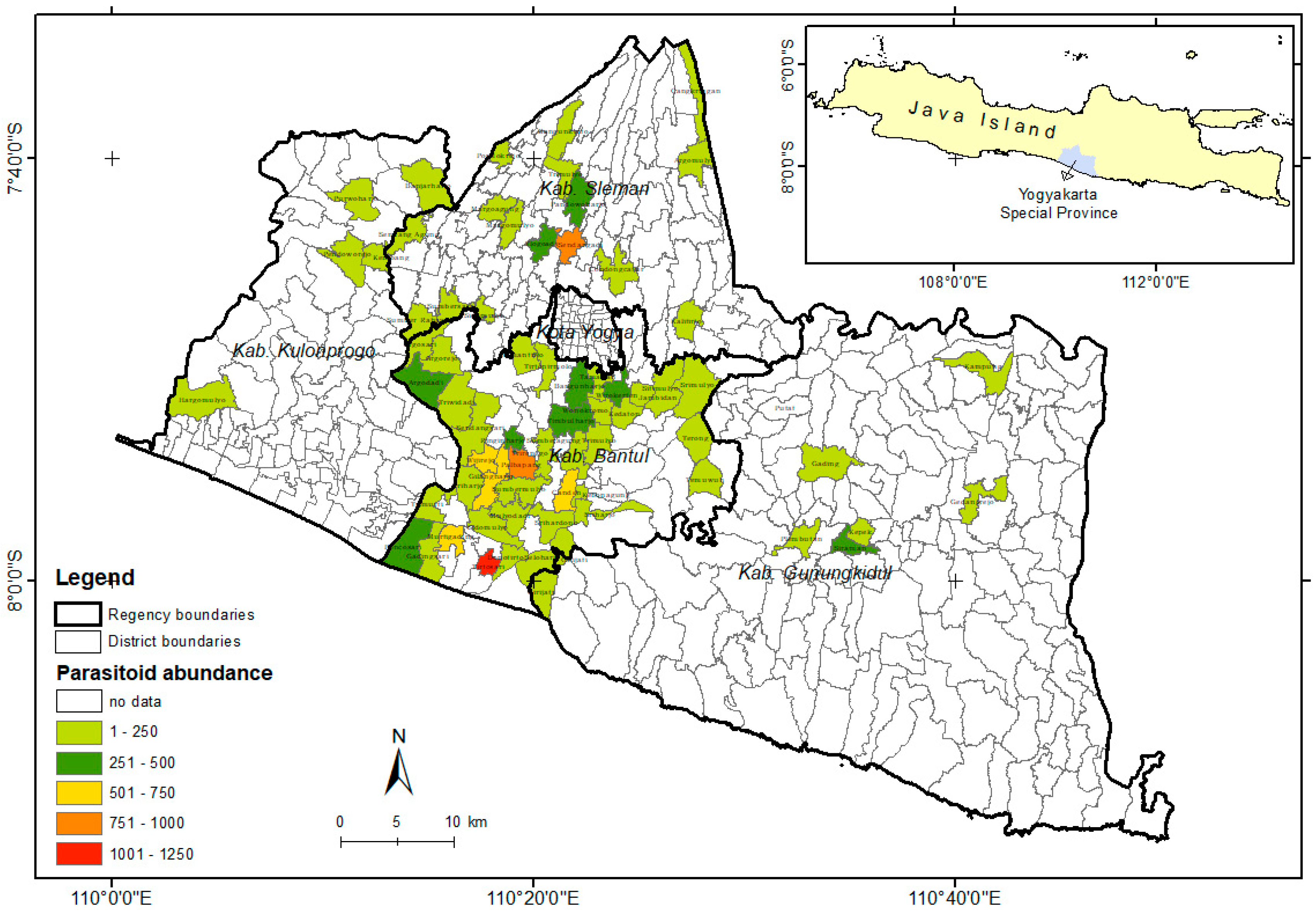
Table 2.
Parasitism rate of Spodoptera frugiperda’s parasitoid in Special Region Yogyakarta, Indonesia.
Table 2.
Parasitism rate of Spodoptera frugiperda’s parasitoid in Special Region Yogyakarta, Indonesia.
| Order | Family | Species | Parasitism rate (Mean, %) | |||
| Bantul | Gunung Kidul |
Kulon progo |
Sleman | |||
| Egg parasitoid | ||||||
| Hymenoptera | Platygasteridae | Platygasteridae.sp01 + | 1.46 | |||
| Platygasteridae | Platygasteridae.sp02 + | 42.00 | ||||
| Platygasteridae | Telenomus remus• | 37.29 | 14.74 | 71.97 | 33.23 | |
| Trichogrammatidae | Trichogramma sp. | 5.78 | ||||
| Larval parasitoid | ||||||
| Hymenoptera | Braconidae | Cotesia sp. • | 11.89 | 16.67 | 5.02 | |
| Ichneumonidae | Campoletis sp. • | 1.61 | ||||
| Braconidae | Coccygidium sp. * | 5.56 | ||||
| Eupelmidae | Eupelmidae.sp01 + | 6.25 | ||||
| Braconidae | Microplitis sp. • | 39.70 | 61.29 | |||
| Braconidae | Stenobracon sp. * | 11.54 | ||||
| Diptera | Tachinidae | Archytas marmoratus | 4.03 | |||
| Phoridae | Megaselia scalaris | 3.70 | ||||
| Pupal parasitoid | ||||||
| Hymenoptera | Chalcididae | Brachymeria femorata• | 6.25 | |||
| Chalcididae | Brachymeria lasusi• | 6.82 | ||||
| Ichneumonidae | Charops sp. • | 1.04 | 4.02 | |||
+: new association, *: relatively new association, •: existing association.
Table 3.
Diversity of Spodoptera frugiperda’s parasitoid in Special Region Yogyakarta, Indonesia.
| Order | Family | Species | Abundance | |||
| Bantul | Gunung Kidul |
Kulon progo |
Sleman | |||
| Egg parasitoid | ||||||
| Hymenoptera | Platygasteridae | Platygasteridae.sp01 | 49 | |||
| Platygasteridae | Platygasteridae.sp02 | 21 | ||||
| Platygasteridae | Telenomus remus | 8536 | 831 | 466 | 2324 | |
| Trichogrammatidae | Trichogramma sp. | 1988 | ||||
| Larval parasitoid | ||||||
| Hymenoptera | Braconidae | Cotesia sp. | 27 | 12 | 110 | |
| Ichneumonidae | Campoletis sp. | 1 | 1 | |||
| Braconidae | Coccygidium sp. | 1 | 1 | |||
| Eupelmidae | Eupelmidae.sp01 | 1 | ||||
| Braconidae | Microplitis sp. | 15 | 19 | |||
| Braconidae | Stenobracon sp. | 3 | ||||
| Diptera | Tachinidae | Archytas marmoratus | 2 | |||
| Phoridae | Megaselia scalaris | 5 | ||||
| Pupal parasitoid | ||||||
| Hymenoptera | Chalcididae | Brachymeria femorata | 5 | |||
| Chalcididae | Brachymeria lasusi | 3 | ||||
| Ichneumonidae | Charops sp. | 1 | 2 | |||
| Species richness | 8 | 6 | 3 | 7 | ||
| Total abundance | 8753 | 924 | 479 | 2478 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



