
Submitted:
31 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
Soil sodicity is a major constraint to seedling emergence and crop production, potentially reducing plant growth due to physical and chemical constraints. Examining responses to ion imbalances may assist in the identification of genotypes tolerant to chemical constraints in sodic soils and improve productivity. We evaluated the performance of four wheat (Triticum aestivum. L) genotypes in solutions with five sodium adsorption ratios (SARs) ranging from 0 to 60. For all four genotypes, seedling emergence and shoot dry matter (DM) decreased significantly with increasing SAR. A significant positive correlation was observed between Ca concentration in roots, as well as both root and shoot DM for all genotypes. At SAR values >20, a more tolerant genotype (EGA Gregory), had higher Ca concentrations in root tissues, while a more sensitive genotype (Baxter) exhibited Na-induced Ca deficiency. Thus, selection for genotypes that are able to accumulate Ca in roots in sodic conditions may be a useful trait for selecting genotypes tolerant of soils with high ESP values. However, for soils that restrict plant growth at ESP (SAR) values of 6-10%, it is likely that growth is restricted by physical constraints rather than by a Na-induced Ca deficiency.
Keywords:
biomass
; Ca concentration
; seedling
; emergence
; roots
; youngest mature leaf
1. Introduction
Sodic soils cover 550 million ha of land worldwide, with 340 million ha occur ring in Australia alone [1]. Soil sodicity is a major constraint limiting grain production, occurring in nearly 60% of Australian grain cropping soils, and costing Australian wheat growers an estimated $1.46 billion/year in forfeited grain yields [2]. Indeed, grain yield on sodic soils is often less than 50% of the potential yield in the absence of constraints [3].
In Australia, sodic soils are usually defined as soils that contain a horizon in which the exchangeable sodium percentage (ESP) is 6 or greater, with soils having an ESP greater than 15 considered strongly sodic [4]. In addition, Läuchli and Epstein [5] suggested that the sodium adsorption ratio (SAR) of the saturated soil extract could be used to estimate the sodic hazard to plants. Rengasamy and Olsson [1] also classified sodic soils based upon SAR and considered soils with a soil solution SAR greater than 3 to be sodic.
Sodicity in the surface soil causes swelling and dispersion of clay particles, resulting in degradation of soil structure. This can lead to surface crusting, hard-setting, waterlogging, poor water infiltration and reduced plant available water capacity. These in turn may cause poor seed germination, reduced seedling emergence, reduced root growth, low water use efficiency, reduced plant growth and reduced yield [3,6,7,8].
In addition to the physical constraints caused by dispersion, sodic soils can also have chemical constraints, including Na and Cl toxicity, as well as deficiencies in essential plant nutrients, such as Ca, K, and micronutrients [3]. The deficiency of Ca in particular can be a problem in Australian sodic soils, where Ca concentrations in the soil solution are often very low (<1 mM) [9] and high Na: Ca ratios can potentially cause a nutritional imbalance. Similarly, K is also often found at low levels, and the uptake of K by plants can be markedly suppressed as sodicity increases [10]. Numerous studies have shown that an increase in ESP causes a significant decrease in K availability as well as an increase in Na concentration of the plant tissues, including in cotton (Gossypium hirsutum) [11,12], rapeseed (Brassica napus) [13], sugarcane (Saccharum officinarum) [14] , rice (Oryza sativa) [15], aloe vera (Aloe barbadensis) [16], barley (Hordeum vulgare) [17], and wheat (Triticum aestivum .L) [18,19]. The poor productivity of sodic soils has also been partly attributed to micronutrient deficiencies due to the high pH [20,21]. However, not all studies have found micronutrient deficiencies in sodic soils, with Wright and Rajper [19] finding no differences in the concentrations of Cu, Fe, Mn and Zn in wheat grain and straw, although the total content of these micronutrients was decreased due to lower DM.
Different management practices can be used to improve crop production in sodic soils, particularly amelioration, for example via gypsum application, to reduce soil ESP [21,22,23]. However, although some amelioration strategies have been reported to be effective, they are often too costly and are not economically viable [24,25]. Consequently, there is a demand for crop varieties tolerant to sodicity to maintain productivity in constrained areas.
Plant sensitivity to sodic soils can vary between genotypes within a plant species. For example, differences in K and Na accumulation in solution culture have been observed in the shoots of different wheat genotypes [26]. Studies have also reported that wheat genotypes showed variation in their tolerance to physical constraints of sodic soils [27,28]. However, the selection of tolerant genotypes represents a substantial challenge to plant breeders due to the complex nature of plant tolerance to sodic soils, and the need to identify plants with tolerance to both the physical and chemical challenges presented by sodic conditions [24]. These problems are exacerbated by the practical difficulties of growing plants at high ESP values.
The present study examined four wheat genotypes (Baxter, EGA Gregory, Spitfire and Ventura) that have previously been observed to differ in their performance in the field in a sodic soil with an ESP of 12.5-17% [17]. We grew the plants in nutrient solutions at five different SAR values (0, 10, 20, 30, and 60) to determine if the difference we previously observed in their growth in sodic soils was related to differences in tolerance to chemical imbalances associated with Na toxicity or Ca deficiency. Plant performance was assessed by measuring seedling emergence, shoot and root growth as well as tissue elemental concentrations. We postulated that this information could allow the development of suitable screening techniques to identify tolerant genotypes with improved crop establishment and early plant growth where Na toxicity and/or Na-induced Ca deficiency occur due to high surface soil ESP.
2. Materials and Methods
2.1. Selection of Genotypes and Seed Collection
The four wheat genotypes (Baxter, EGA Gregory, Ventura and Spitfire) used in this study were selected from those found previously to differ in their growth and yield in sodic soil with ESP value of 12.5 (equivalent solution SAR ~9.5) in surface soils and ESP >22% in subsoils (equivalent solution SAR ≥ 20) in the field [17]. In the previous field-based study, Baxter was found to maintain yield on a sodic soil better than Ventura and EGA Gregory, which were comparatively sensitive [17]. Furthermore, these genotypes have also been found to differ in their emergence from a crusted sodic soil in glasshouse experiments [27], where Spitfire and Ventura were better able to emerge through a surface crust compared to EGA Gregory and Baxter.
All seed samples used in this experiment were harvested from increase rows grown under irrigated conditions with non-limiting soil nutrient levels at the Queensland Government research farm at Kingsthorpe, Queensland, Australia (27.52 °S, 151.79 °E) in the absence of soil constraints. Seeds were harvested in November 2016 and were stored in a cold room at 7 °C. One week prior to the experiment, seeds were warmed to 22 °C.
2.2. Solution Preparation and Plant Growth
A solution culture experiment was conducted to investigate the impact of high SAR solutions on growth of wheat. A black 10 L bucket was filled with a basal nutrient solution with µM concentrations of, 1000 Ca, 40 NH4+-N, 102 Mg, 305 K, 20 Na, 10 Fe (Na2FeEDTA), 0.5 Mn, 0.5 Zn, 0.2 Cu, 1320 Cl, 720 NO3--N, 254 SO42–-S, 5 PO4–-P, 1 B, and 0.01 Mo. Solutions were continuously aerated and the unadjusted solution pH was 6.0. From these basal solutions, five different SAR values (0, 10, 20, 30 and 60) were prepared by adding differing quantities of NaCl, NaSO4, CaCl2.2H2O and CaSO4.2H2O (Table 1). A mixture of chloride and sulphate salts was used to avoid chloride toxicity in the solution. Across these five SAR values, ionic strength (I) was held constant at 31 mM. A computer program, PhreeqcI (version 3.1.1.8228, wateq4f database), was used to determine the quantity of NaCl, NaSO4, CaCl2.2H2O and CaSO4.2H2O required to prepare the five SAR solutions at a constant I of 31 mM (Table 1). This value for I was selected because Anzooman, Dang, Christopher, Mumford, Menzies and Kopittke [28] reported that SAR (0-60) solutions that have I values ranging from 25 to 50 mM (corresponding electrical conductivity (EC) values being from 0.15 to 3.0 dS m-1) did not have a marked adverse effect on either seed germination or on seedling emergence of these genotypes. The SAR values selected (0, 10, 20, 30 and 60) are equivalent to approximate soil ESP values of 0, 13, 23, 31 and 47 % when calculated using the Gapon equation described in Sumner and Miller [29]. These ESP values are commonly found in many of the surface and subsoils of the region [25]. Concentrations of selected nutrients (Ca, Cu, Fe, K, Mg, Mn, Na, P, S, and Zn) in the nutrient solution were determined at the start and end of each week using inductively coupled plasma optical emission spectroscopy (ICP-OES, Table S1 and S2 of Supplementary Material).
The experiment was undertaken in a laboratory at 25°C at The University of Queensland, St Lucia (Australia) under high pressure sodium lights. These lights provided photosynthetically active radiation (PAR) of 1500 µmol m-2 s-1 when measured at canopy height, with 12 h of light per day. In each 10 L bucket, there were four holes across which shade cloth was suspended and supported using foam cups. Each cup contained a single genotype (with the four replicates being in the four cups of each container), with 10 seeds per cup. The average of the 10 seeds per cup formed a single replicate of one SAR solution for each genotype. Each solution treatment was replicated four times. Thus, with five SAR values, four genotypes, and four replicates, the experiment consisted of a total of 80 experimental units. The appropriate seeds were placed on the shade cloth (Figure S1 of Supplementary Material), which allowed the seeds to imbibe moisture from below but ensured that they were not covered by the solution. Seeds were then covered using white polypropylene beads to limit evaporation and to limit entry of light into the bucket.
At the end of the experimental period, 14 d after sowing, plants were harvested before being separated into root, stem plus petiole, and youngest mature leaf (YML). A digital camera (Canon PowerShot SX600 HS 16 MP Ultra-Zoom Digital) was used to capture images of the roots from each pot. Root length were measured from these images using ImageJ (version 1.45s, National Institutes of Health).
Roots were washed in deionized water to remove any adhering solution. The various plant parts were dried at 65 °C for 72 h and the DM of the shoots and roots recorded. Shoots and roots were digested in a 1:5 mixture of perchloric acid and nitric acid before measurement of concentrations of Ca, Cu, Fe, K, Mn, Mg, Na, P, S and Zn in the YML and the root tissues using inductively coupled plasma optical emission spectrometry (ICP-OES).
2.3. Statistical Analysis
Linear mixed models were employed to analyse the data using the residual maximum likelihood procedure [30] via the ASReml-R package [31] in the R software environment [32].
For each experiment, the explanatory variables (genotypes, SAR) were treated as fixed effects while replicate block effects were considered as random. Hence, the predictions of the treatment effects obtained from the model for each of the experiments were provided as empirical best linear unbiased estimates (eBLUEs). Significance of treatment and interaction effects were tested via Wald tests using an approximate F-statistic. A 5% significance level was used for both Wald and LSD tests.
There were some seedlings that did not emerge. Hence, root length was only available from the emerged seedlings of each genotype in each treatment. For the analysis of root length, an arcsine transformation was used to meet the assumption of homogeneity of variance. The transformation is calculated by equation (1):
where is emergence (%).
Seedling emergence, root and shoot DM and their relationship with Ca, Na and K were analysed using regression analysis, fitting either polynomial, linear curves or curves of the general form, using equation (2):
where, b is the maximum DM/nutrient concentrations in SAR 0 and Ca sufficient solutions, c is a strength coefficient and increases with the strength of the toxicant and h is a shape coefficient [33]. Regression analyses were conducted using SYSTAT 13 (Cranes Software International Ltd, Bangalore, India).
3. Results
3.1. High SAR Reduced Seedling Emergence
A significant negative correlation was found between seedling emergence and SAR for all four genotypes (P<0.001, Figure 1). At a SAR value of 0, all four genotypes had ≥ 80% emergence (Figure 1). However, emergence rate decreased to ca. 15 % for Spitfire, Ventura, and Baxter at SAR 60. The decrease in emergence for EGA Gregory was less pronounced, with emergence being 50 % at SAR 60 (Figure 1).
3.2. Root Length, Root Mass and Shoot Mass Were Also Reduced
For root DM, a significant interaction was found between the genotype and SAR, indicating that while root DM generally decreased with increasing SAR, the pattern of this decrease differed between the four genotypes (P<0.001, Figure 2). Specifically, for Baxter, Spitfire, and Ventura, the root DM decreased from 0.21-0.25 g at SAR 0 to 0-0.03 g at SAR 60 (Figure 2). However, EGA Gregory was less sensitive to increasing SAR, with root DM remaining comparatively constant (0.20-0.23 g) irrespective of SAR (Figure 2). In a similar manner to root DM, maximum root length of Spitfire, Baxter and Ventura decreased with increasing SAR, but maximum root length of EGA Gregory was less (non-significant, P>0.05) influenced by solution SAR (Figure S2 of Supplementary Material).
The shoot DM of all four genotypes was also observed to decrease with increasing SAR (Figure 3). At SAR 0, shoot DM of the four genotypes ranged from 0.26 to 0.31 g, while at SAR 60, shoot DM ranged from 0.04 to 0.13 g (Figure 3). A significant interaction for shoot DM was found between the genotypes and SAR, indicating that the pattern of the decrease in shoot DM with increasing SAR varied among the genotypes (P=0.03, Figure 3). Specifically, the decrease in shoot DM was lowest for EGA Gregory (decreasing from 0.27 to 0.13 g) and highest for Baxter (0.26 to 0.05 g, Figure 3).
3.3. Elemental concentrations in root tissues
For all four genotypes, the Ca concentration of the root tissues tended to decrease as SAR increased (Figure S3 of Supplementary Material). The magnitude of this decrease was largest for Ventura (decreasing from 0.45 to 0.18%) and smallest for EGA Gregory (decreasing from 0.45 to 0.29%). It was observed that higher Ca concentration in root tissues was correlated with a greater ability to maintain both root DM (R2=0.77; P<0.0001, Figure 4) and shoot DM (R2=0.46, P<0.01, Figure S4 of Supplementary Material). From the regression, a decrease in the Ca concentration of the root tissues to ca. 0.15% was associated with a 50% reduction in root DM. In a similar manner, the K concentration of the root tissues also decreased significantly for all four genotypes with increasing SAR (P<0.01, Figure S3 of Supplementary Material). Again, the magnitude of this decrease was largest for Ventura (decreasing from 3.96 to 1.11%) but it was smallest for Spitfire (decreasing from 2.58 to 2.01%). A significant positive relationship was observed between K concentration in roots and root DM (R2=0.39, Figure 4). In contrast to Ca and K, the concentration of Na in the root tissues increased significantly with SAR, increasing from 0.05-0.06% at SAR 0 to 0.62-1.99% at SAR 10-60 (P<0.0001, Figure S3 of Supplementary Material). However, there was no significant relationship between root DM and root Na concentration (R2 =0.05, Figure 4).
3.4. Elemental concentrations in the YML tissues
For all four genotypes, the tissue Ca concentration of the YML tended to decrease as SAR increased (Figure S5 of Supplementary Material). The magnitude of this decrease was largest for Ventura (decreasing from 0.68 to 0.04%) and smallest for Spitfire (decreasing from 0.58 to 0.33%). Importantly, it was noted that for both Ventura and Baxter, the Ca concentration of the YML decreased to values lower than the critical concentration reported for Ca deficiency (0.25 % [9] at SAR 60, indicating that these plants were likely Ca deficient at high SAR. A significant positive relationship was observed between the Ca concentration in YML and shoot DM for all four genotypes (R2=0.44, P<0.001, Figure 5). Furthermore, for all four genotypes, the K concentration of the YML tended to decrease as SAR increased (Figure S5 of Supplementary Material). This decrease was the largest for Ventura (decreasing from 3.83 to 0.42 %) and smallest for EGA Gregory (decreasing from 4.14 to 3.01 %). Importantly, it was noted that for both Ventura and Baxter, the K concentration of the YML decreased to values lower than the critical concentration for K deficiency (1.6 % [9]) at SAR 60, indicating that these plants were likely K deficient at high SAR. A positive relationship was observed between K concentration in YML and shoot DM (R2= 0.43, P<0,001 Figure 5).
In contrast to Ca and K, the concentration of Na in the YML tissues increased significantly with increasing SAR up to 30 for all four genotypes, with tissue Na concentrations at SAR 60 being similar (or slightly lower) than at SAR 30 (Figure S5 of Supplementary Material). No significant differences (P=0.09) were found between genotypes regarding their YML tissue concentrations of Na. However, a significant relationship was also observed between Na concentration in YML and shoot DM (R2=0.52, Figure 5). We also examined the K: Na ratio in the YML tissues, finding a significant difference between treatments (P<0.0001, Table 2). The tissue K: Na ratio did not vary significantly between the genotypes (P=0.17) at five SARs. A significant positive correlation was found between the K: Na in the YML and shoot DM (R2=0.86, Figure 5).
3.5. Relationship between Root and Shoot DM and Root Length
No significant difference was observed between the root length of all the four genotypes at SAR 0, 10, 20 and 30. However, at SAR 60, the root lengths of Baxter and Ventura decreased sharply. A significant positive correlation was also observed between the root length and root DM of all the four wheat genotypes in all treatments (R2=0.80, Figure S6 of Supplementary Material) and between root and shoot DM (R2=0.56, Figure S7 of Supplementary Material).
3.6. Comparison between Traits
Previous research identified different traits of wheat genotypes that influenced their adaptability in the presence of physical and chemical constraints of sodic soils. Seedling emergence of wheat genotypes decreased in the presence of a surface crust of a thickness and strength relevant to that observed in field conditions (soil ESP 10) [27]. Seedling coleoptile length of wheat genotypes also reduced with an increase of soil ESP and bulk density [34]. Wheat genotypes also showed variation in seedling emergence, seedling emergence force and root angle in sodic conditions [28,34]. In the present study, seedling emergence, and Ca concentration in roots have been identified as potential traits that could be used to identify wheat genotypes tolerant to high SAR conditions. A general comparison between the traits identified in the present study and those in previous studies is presented in Table 3.
4. Discussion
The aim of the present study was to investigate differences in the performance of four wheat genotypes grown in nutrient solutions with five different SAR values. These four wheat genotypes were selected as previous studies had demonstrated that their performance in a sodic soil in the field differed markedly. In the present study, it was found that the performance of these genotypes in sodic soils in the field at an ESP value of 12.5%, or in the presence of soil surface crust at an ESP of 10%, could not be explained by differences in response to chemical constraints (see discussion below and Table 3). Thus, these results suggest that differences in plant performance for this particular sodic soil are likely related to differences in their tolerance to adverse physical conditions. However, these findings will likely be useful for identification of tolerant wheat genotypes in other sodic soils of high ESP (>30%).
4.1. Ca deficiency in the roots contributed to reduced growth at high SAR
Both root and shoot DM generally decreased as SAR increased, although differences were observed between the four genotypes (Figure 2 and 3). Overall, EGA Gregory was found to be the most tolerant of elevated SAR values, with root DM remaining relatively constant across all SAR values examined (Figure 2) and with the reduction in shoot DM being smaller than for the three other genotypes (Figure 3).
This reduction in root and shoot mass observed at elevated SAR was associated with the development of a Na-induced Ca deficiency. Specifically, although increases in SAR resulted in decreasing tissue concentrations of Ca and K in both the roots and shoots as well as increasing tissue concentrations of Na (Figure S3 and S5 of Supplementary Material), the decrease in root DM was most closely associated with decreases in root tissue concentrations of Ca (R2 = 0.77), compared to decreases in root tissue concentrations of K (R2 = 0.39) or increases in Na (R2 = 0.05). Indeed, EGA Gregory, which had the lowest reduction in root DM (Figure 2) also had the lowest reduction in root tissue Ca concentrations (Figure S3 of Supplementary Material).
Although the absolute concentration of Ca in the nutrient solution was adequate to meet nutritional requirements under non-limiting conditions (≥ 160 µM, Table 1) (for example, see Figure 1 of Kopittke, Blamey, Kinraide, Wang, Reichman and Menzies [33]) it is well known that the addition of other salts, including those of Na, K, and Mg can induce Ca deficiency [35,36]. The formation of Na-induced Ca deficiency is associated with a decrease in Ca availability at elevated Na concentrations, due to competition between Na and Ca, and due to a reduction in the Ca2+ availability at the plasma membrane surface [9,37]. In the present study with wheat, a root tissue Ca concentration of 0.15 % was found to correspond to a 50% reduction in root DM. This is similar to previous values reported for reduction in shoot and root DM due to Ca deficiency [38,39,40]. Given that Ca is largely immobile in the phloem [41], it must have adequate availability in the rooting medium to ensure optimal growth.
4.2. Nutritional imbalances in the shoot
Although decreases in shoot DM were also associated with a decrease in shoot tissue Ca concentrations (Figure 5), this relationship was not as strong as that observed for the roots, suggesting that the roots were the primary site of Ca deficiency. Rather in the shoots, the positive correlation between K: Na in YML and shoot DM was stronger (Figure 5) which has been supported by Dang, Christopher and Dalal [17] who found a positive correlation (R2=0.47) between K: Na in YML and grain yield. However, the non-significant difference in K: Na in YML of these genotypes indicated that K: Na is not a significant trait to identify wheat genotypes tolerance in SAR solutions. The K and Na concentration in YML may not be used to identify genotypes tolerant to sodicity as there were no differences in the pattern of K and Na accumulation in the genotypes despite differences in plant growth (Figure 5). Dang, Christopher and Dalal [17] also found no pattern of Na accumulation of tolerant and non-tolerant wheat genotypes in YML and reported that Na in YML was not a causative factor for growth depression in wheat on representative sodic soils in Queensland.
4.3. Is growth in sodic soils related to tolerance to ion imbalances?
Among the four genotypes used in the present experiment EGA Gregory, Ventura, and Baxter have previously been identified as differing in their performance in the field when grown in a sodic soil [17]. In this previous study, it was found that Baxter and Ventura produced significantly higher yields compared to EGA Gregory in crusted sodic soils. These previous results differ clearly from the present study, where EGA Gregory was found to produce the highest root and shoot DM at elevated SAR values (Figure 2 and 3).
These observations are important and provide several insights. Firstly, it is apparent that the superior yield of Baxter and Ventura in the field study of Dang, Christopher and Dalal [17] was not due to a greater tolerance to ion imbalances. Secondly, it is important to note that the ESP of the soil in the field study of Dang, Christopher and Dalal [17] was comparatively modest (12.5%) with a soil solution SAR value of 9.5. Thus, the higher SAR values examined in the present study markedly exceeded those found in the sodic soil of Dang, Christopher and Dalal [17]. The reduction in root and shoot DM at the tested SAR value of 10, which most closely corresponding to that observed in the field (9.5), was comparatively modest. Thus, it is likely that the reduction in yield observed in the field study was likely not due to ion imbalances per se. Finally, it is noteworthy that Anzooman, Christopher, Mumford, Dang, Menzies and Kopittke [27] utilized a glasshouse study to examine whether growth of these four genotypes was influenced by the physical properties of a sodic soil. These authors found among these four genotypes, Ventura and Spitfire had significantly greater seedling emergence compared to Baxter and EGA Gregory in the presence of surface crust in sodic soil (ESP 10%, SAR 7) and that their emergence was associated with greater seedling emergence force and narrower seminal root angle [28,34]. Thus, it is apparent that the differences in growth between these four wheat genotypes when grown in a sodic soil in the presence of surface crust [28] are due to differences in the ability of the genotypes to overcome the physical constraints rather than due to an improved tolerance of ion imbalances. Nevertheless, the findings of the present study illustrate that for sodic soils with high SAR values, differences in the tolerance of genotypes to ion imbalances would potentially allow for the selection of genotypes to improve yield.
In semi-tropical areas of Australia, crops often rely heavily of deep soil moisture late in the season due to increasing water limitation as water from surface layers is exhausted. SAR of sodic soils are often much greater in the deeper layers of the sub-soil, for example below 40 or 60cm, and can commonly exceed the critical SAR of 30 identified in the current study [42]. It is thus quite possible that tolerance to ionic constraints may be more important late in the season during terminal moisture stress when crops need to extract moisture from deeper, more sodic, layers. Conversely, tolerance to physical constraints may be more important for early emergence through crusts, establishment and early growth. Thus, the relative importance of adaptation to chemical versus physical constraints is likely to vary during the crop cycle. Further study is warranted to determine the relative importance of these two classes of adaptation for overall performance in the relevant cropping environments.
5. Conclusions
The results from the present study suggest that at early growth stages, wheat tolerance to the chemical constraints of sodicity might be governed by the ability of genotypes to accumulate Ca in roots. Tolerant genotypes generally had higher Ca concentration in roots, whereas relatively more susceptible genotypes exhibited Na-induced Ca deficiency (at SAR ≥30). The concentration of Ca and K in YML was not below critical levels at lower SAR (<30) for all the genotypes. This indicates that K and Ca concentration in the YML might not affect early plant growth in sodic conditions < 30 SAR. This study identified adaptations to chemical constraints of sodic soil in solution culture and indicates that these do not necessarily correlate with tolerance to physical constraints of sodic soil such as surface crusting. The relative importance of adaptation to chemical constraints versus physical constraints in relevant cropping environments on sodic soils warrants further investigation.is section is not mandatory but can be added to the manuscript if the discussion is unusually long or complex.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization: M.A., J.C., Y.P.D. and P.M.K.; Methodology: M.A. and P.M.K.; Analysis and result interpretation: M.A., P.M.K. and Y.P.D.; Writing—original draft preparation: M.A.; Writing—review and editing: P.M.K., J.C, Y.P.D., and N.W.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received funding from The University of Queensland and Australian Government Research Training Program Scholarship
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Acknowledgments
The authors wish to acknowledge support from The University of Queensland, Department of Agriculture and Fisheries Queensland, the Australian Grains Research and Development Corporation (GRDC) and the grain producers of Australia.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Rengasamy, P.; Olsson, K.A. Sodicity and soil structure. Australian Journal of Soil Research 1991, 29, 935-952. [CrossRef]
- Orton, T.G.; Mallawaarachchi, T.; Pringle, M.J.; Menzies, N.W.; Dalal, R.C.; Kopittke, P.M.; Searle, R.; Hochman, Z.; Dang, Y.P. Quantifying the economic impact of soil constraints on Australian agriculture: A case-study of wheat. Land Degradation & Development 2018, 29, 3866-3875.
- Rengasamy, P. Transient salinity and subsoil constraints to dryland farming in Australian sodic soils: an overview. Animal Production Science 2002, 42, 351-361. [CrossRef]
- Houghton, P.D.; Charman, P.E.V. Glossary of terms used in soil conservation; Soil Conservation Service of NSW: 1986.
- Läuchli, A.; Epstein, E. Plant responses to saline and sodic conditions. Agricultural Salinity Assessment and Management 1990, 71, 113-137. [CrossRef]
- Thimmappa, K.; Singh, Y.P.; Raju, R. Reclamation of sodic soils in India: An economic impact assessment. In Bioremediation of Salt Affected Soils: An Indian Perspective., Arora S., Singh A., Y., S., Eds.; Springer, Cham: 2017. [CrossRef]
- Agassi, M.; Morin, J.; Sheinberg, I. Effect of raindrop impact energy and water salinity on infiltration rates of sodic soils. Soil Science Society of America Journal 1985, 49, 186-190. [CrossRef]
- Rao, P.; Mishra, B.; Gupta, S. Effects of soil salinity and sodicity on grain quality of tolerant, semi-tolerant and sensitive rice genotypes. Rice Science 2013, 20, 284-291. [CrossRef]
- Reuter, D.; Robinson, J.B. Plant Analysis: An Interpretation Manual; CSIRO publishing: 1997.
- Wakeel, A. Potassium–sodium interactions in soil and plant under saline-sodic conditions. Journal of Plant Nutrition and Soil Science 2013, 176, 344-354. [CrossRef]
- Rochester, I. Phosphorus, and potassium nutrition of cotton: interaction with sodium Crop and Pasture Science 2010, 61, 821-834. [CrossRef]
- Dodd, K.; Guppy, C.; Lockwood, P.; Rochester, I. The effect of sodicity on cotton: Plant response to solutions containing high sodium concentrations. Plant and Soil 2010, 330, 239-249. [CrossRef]
- Porcelli, C.A.; Flavio, H.G.B.; Raul, S.L. The K/Na and Ca/Na ratios and rapeseed yield, under soil salinity or sodicity. Plant and Soil 1995, 175, 251-255. [CrossRef]
- Dang, Y.; Mehla, A.; Chhabra, R.; Kumar, S. Sodicity induced yield losses and changes in mineral concentration of sugarcane genotypes. In Proceedings of the International Society of Sugar Cane Technologists XXIII Congress, 1999; pp. 89-97.
- Qadar, A. Potassium and sodium contents of shoot and lamine of rice cultivars and their sodicity tolerance. Journal of Plant Nutrition 1995 18, 2281-2290. [CrossRef]
- Rahi, T.S.; Singh, K.; Singh, B. Screening of sodicity tolerance in aloe vera: An industrial crop for utilization of sodic lands. Industrial Crops and Products 2013, 44, 528-533. [CrossRef]
- Dang, Y.P.; Christopher, J.; Dalal, R.C. Genetic diversity in barley and wheat for tolerance to soil constraints. Agronomy Journal 2016, 6. [CrossRef]
- Chhipa, B.; Lal, P. Na/K ratios as the basis of salt tolerance in wheat. Australian Journal of Agricultural Research 1995, 46, 533-539. [CrossRef]
- Wright, D.; Rajper, I. An assessment of relative effects of adverse physical and chemical properties of sodic soil on the growth and yield of wheat (Triticum aestivum L.). Plant and soil 2000, 223, 277-285. [CrossRef]
- Naidu, R.; Rengasamy, P. Ion interactions and constrains to plant nutrition in Australian sodic soils Soil Research 1993, 31, 801-819. [CrossRef]
- Curtin, D.; Naidu, R. Fertility constraints to plant production. In Sodic Soils: Distribution, Properties, Management and Environmental Consequences, Sumner, M.E., Naidu, R., Eds.; Oxford University Press: New York, 1999.
- Jayawardane, N.; Chan, K. The management of soil physical properties limiting crop production in Australian sodic soils-a review. Soil Research 1994, 32, 13-44. [CrossRef]
- Chartres, C.J. Sodic Soils: an introduction to their formation and distribution in Australia. Australian Journal of Soil Research 1993, 31, 751-760. [CrossRef]
- Richard, R. Current, and emerging environmental challenges in Australian agriculture - The role of plant breeding. Crop and Pasture Science 2002, 53, 881-892.
- Dang, Y.; Dalal, R.C.; Routley, R.; Schwenke, G.D.; Daniel, I. Subsoil constraints to grain production in the cropping soils of the north easter region of Australia: an overview. Australian Journal of Experimental Agriculture 2006, 46, 19-35.
- Gorham, J.; Bridges, J.; Dubcovsky, J.; Dvorak, J.; Hollington, P.; Luo, M.C.; Khan, J. Genetic analysis and physiology of a trait for enhanced K+/Na+ discrimination in wheat. New Phytologist 1997, 137, 109-116. [CrossRef]
- Anzooman, M.; Christopher, J.; Mumford, M.; Dang, Y.P.; Menzies, N.W.; Kopittke, P.M. Selection for rapid germination and emergence may improve wheat seedling establishment in the presence of soil surface crusts. Plant and Soil 2018, 426, 227-239. [CrossRef]
- Anzooman, M.; Dang, Y.P.; Christopher, J.; Mumford, M.H.; Menzies, N.W.; Kopittke, P.M. Greater emergence force and hypocotyl cross sectional area may improve wheat seedling emergence in sodic conditions Plant Science 2018, 277, 188-195. [CrossRef]
- Sumner, M.E.; Miller, W.P. Cation-exchange capacity and exchange coefficients. In Methods of Soil Analysis, Sparks, D.L., Ed.; American Society of Agronomy, Madison, WI: 1996.
- Patterson, H.; Thompson, R. Recovery of inter-block information when block sizes are unequal. Biometrika 1971, 58, 545-554. [CrossRef]
- Butler, D.; Ciullis, B.; Gilmour, A.; Gogel, B. ASReml-R reference manual; The State of Queensland, Department of Primary Industries and Fisheries, Brisbane: 2009.
- R Core Team. R: A Language and Environment for Statistical Computing. 2017.
- Kopittke, P.M.; Blamey, F.P.C.; Kinraide, T.B.; Wang, P.; Reichman, S.M.; Menzies, N.W. Separating multiple, short-term, deleterious effects of saline solutions on the growth of cowpea seedlings. New Phytologist 2011, 189, 1110-1121. [CrossRef]
- Anzooman, M.; Christopher, J.; Dang, Y.P.; Taylor, J.; Menzies, N.W.; Kopittke, P.M. Chemical and physical influence of sodic soils on the coleoptile length and root growth angle of wheat genotypes. Annals of Botany 2019, 124, 1043-1052. [CrossRef]
- Grattan, S.R.; Grieve, C.M. Mineral nutrient acquisition and response by plants grown in saline environments In Handbook of Plant and Crop Stress, Pressarakli, M., Ed.M.Dekker, Ed.; 1993.
- Grattan, S.R.; Grieve, C.M. Salinity–mineral nutrient relations in horticultural crops. Scientia horticulturae 1998, 78, 127-157. [CrossRef]
- Huang, J.; Redmann, R. Responses of growth, morphology, and anatomy to salinity and calcium supply in cultivated and wild barley. Canadian Journal of Botany 1995, 73, 1859-1866. [CrossRef]
- Peverill, K.; Sparrow, L.; Reuter, D. Soil analysis: an interpretation manual; CSIRO publishing: 1999.
- Kurth, E.; Cramer, G.R.; Läuchli, A.; Epstein, E. Effects of NaCl and CaCl2 on cell enlargement and cell production in cotton roots. Plant Physiology 1986, 82, 1102-1106. [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant, Cell, and Environment 2002, 25, 239-250. [CrossRef]
- Cramer, G.R.; Läuchli, A. Ion activities in solution in relation to Na+− Ca2+ interactions at the plasmalemma. Journal of Experimental Botany 1986, 37, 321-330.
- Dang, Y.P.; Dalal, R.C.; Buck, S.R.; Harms, B.; Kelly, R.; Hochman, Z.; Schwenke, G.D.; Biggs, A.; Ferguson, N.; Norrish, S. Diagnosis, extent, impacts, and management of subsoil constraints in the northern grains cropping region of Australia. Soil Research 2010, 48, 105-119. [CrossRef]
Figure 1.
Emergence of four wheat genotypes (a) EGA Gregory, (b) Spitfire, (c) Ventura and (d) Baxter at five different SAR solution treatments (0, 10, 20, 30 and 60), averaged over four replicates. The vertical bars indicate the standard error.
Figure 1.
Emergence of four wheat genotypes (a) EGA Gregory, (b) Spitfire, (c) Ventura and (d) Baxter at five different SAR solution treatments (0, 10, 20, 30 and 60), averaged over four replicates. The vertical bars indicate the standard error.
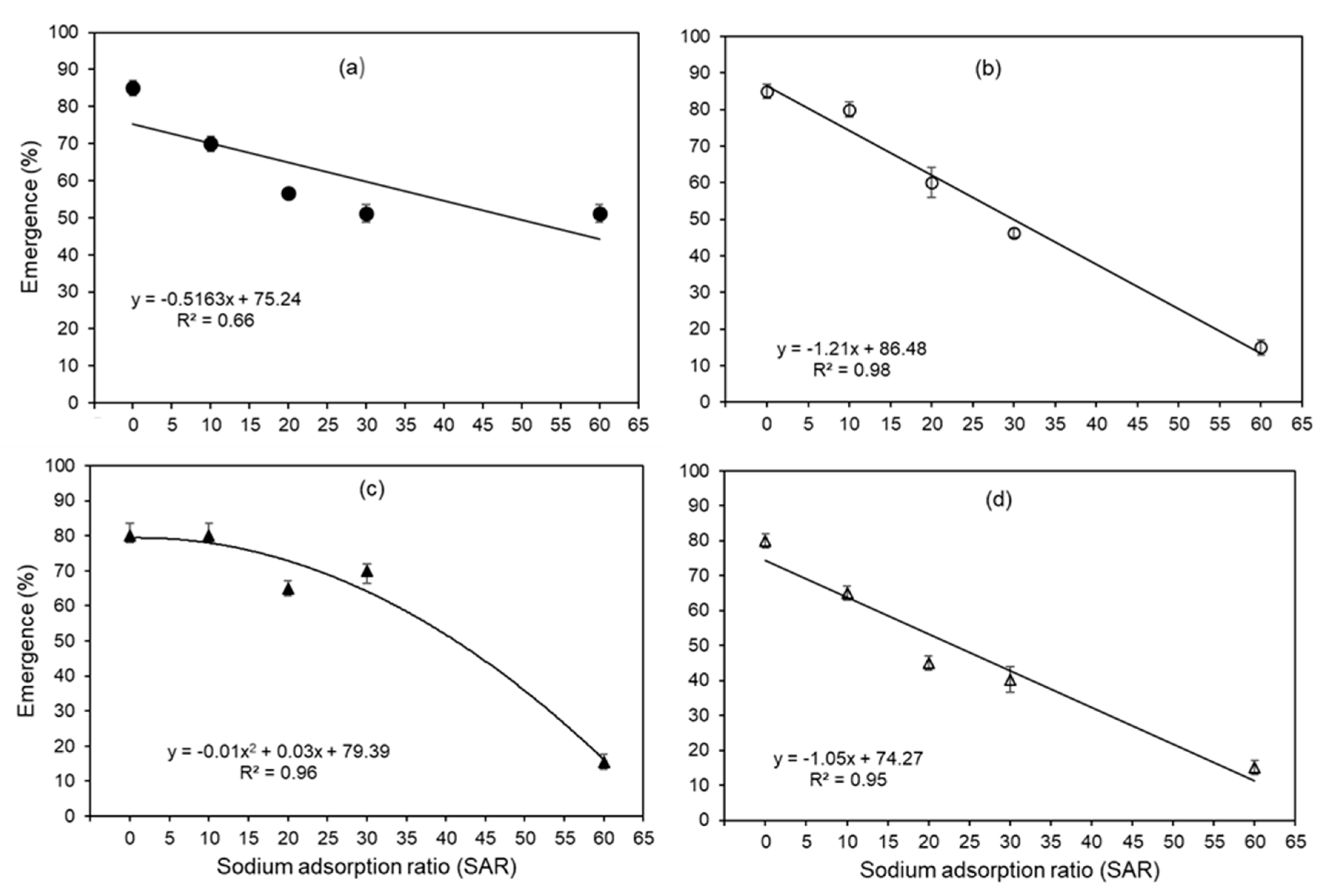
Figure 2.
Root DM of four wheat genotypes (a) EGA Gregory, (b) Spitfire, (c) Ventura, and (d) Baxter in five SAR treatments. The bars indicate the standard error of four replicates.
Figure 2.
Root DM of four wheat genotypes (a) EGA Gregory, (b) Spitfire, (c) Ventura, and (d) Baxter in five SAR treatments. The bars indicate the standard error of four replicates.
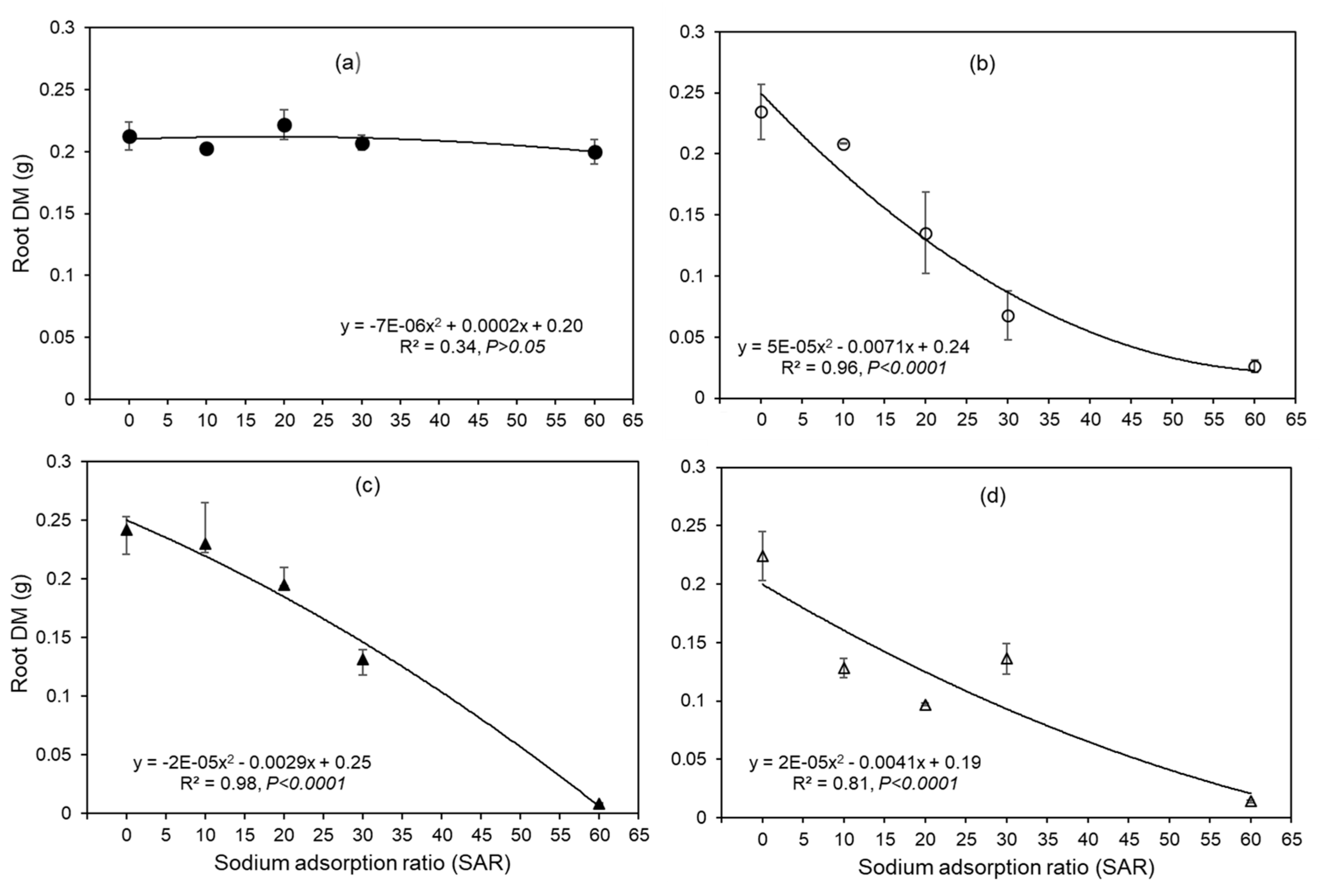
Figure 3.
Shoot DM of four wheat genotypes (a) EGA Gregory, (b) Spitfire, (c) Ventura, and (d) Baxter in five SAR treatments. The bars indicate the standard error of four replicates.
Figure 3.
Shoot DM of four wheat genotypes (a) EGA Gregory, (b) Spitfire, (c) Ventura, and (d) Baxter in five SAR treatments. The bars indicate the standard error of four replicates.
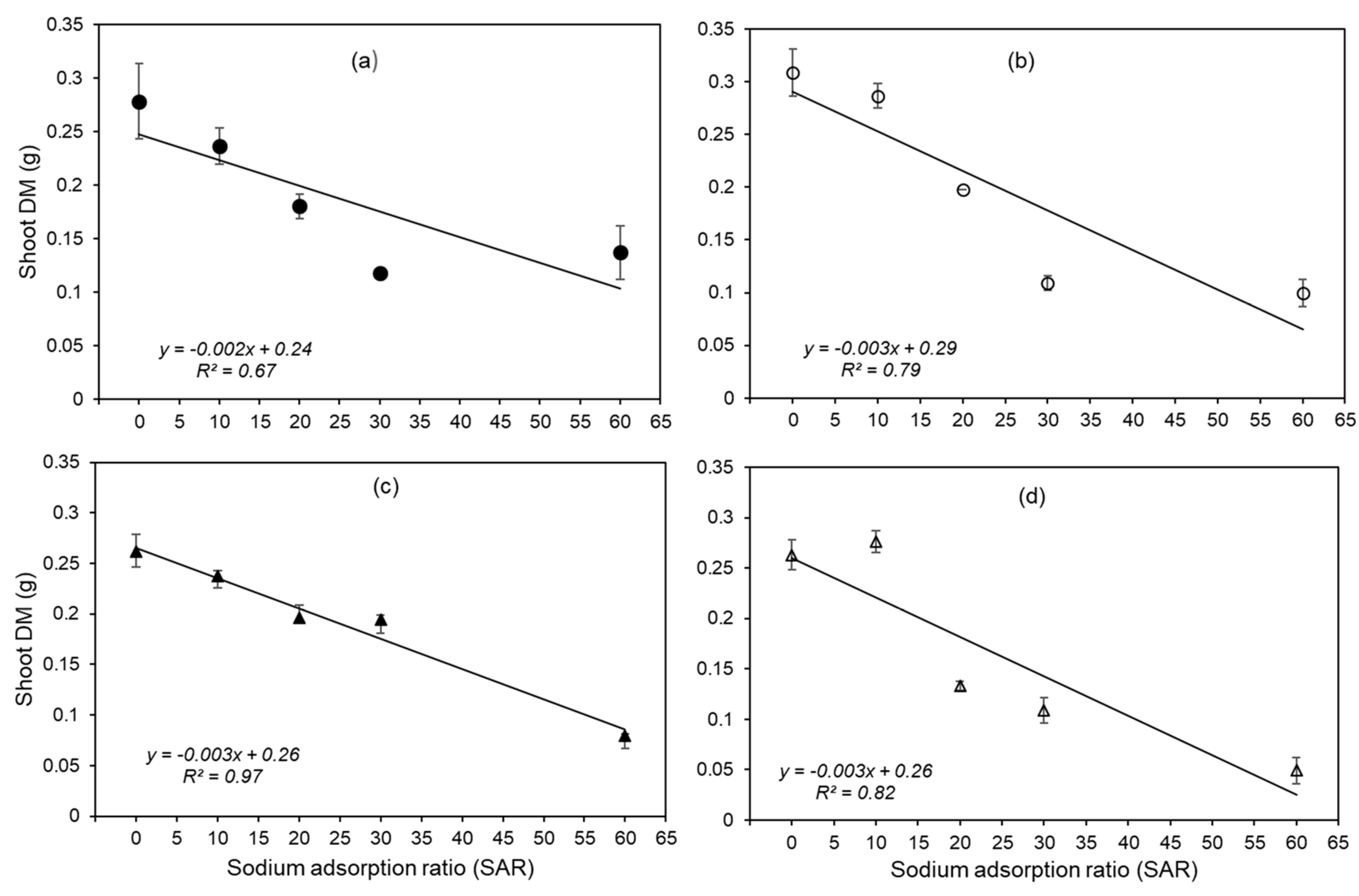
Figure 4.
Relationship between root DM and concentrations of Ca (R2=0.77), K (R2=0.39) and Na (R2=0.05) in root tissues of four wheat genotypes in four replicates grown in five SAR treatments. The bars indicate the standard error. The relationship between root DM and Na concentration in the roots was not significant (P>0.05).
Figure 4.
Relationship between root DM and concentrations of Ca (R2=0.77), K (R2=0.39) and Na (R2=0.05) in root tissues of four wheat genotypes in four replicates grown in five SAR treatments. The bars indicate the standard error. The relationship between root DM and Na concentration in the roots was not significant (P>0.05).
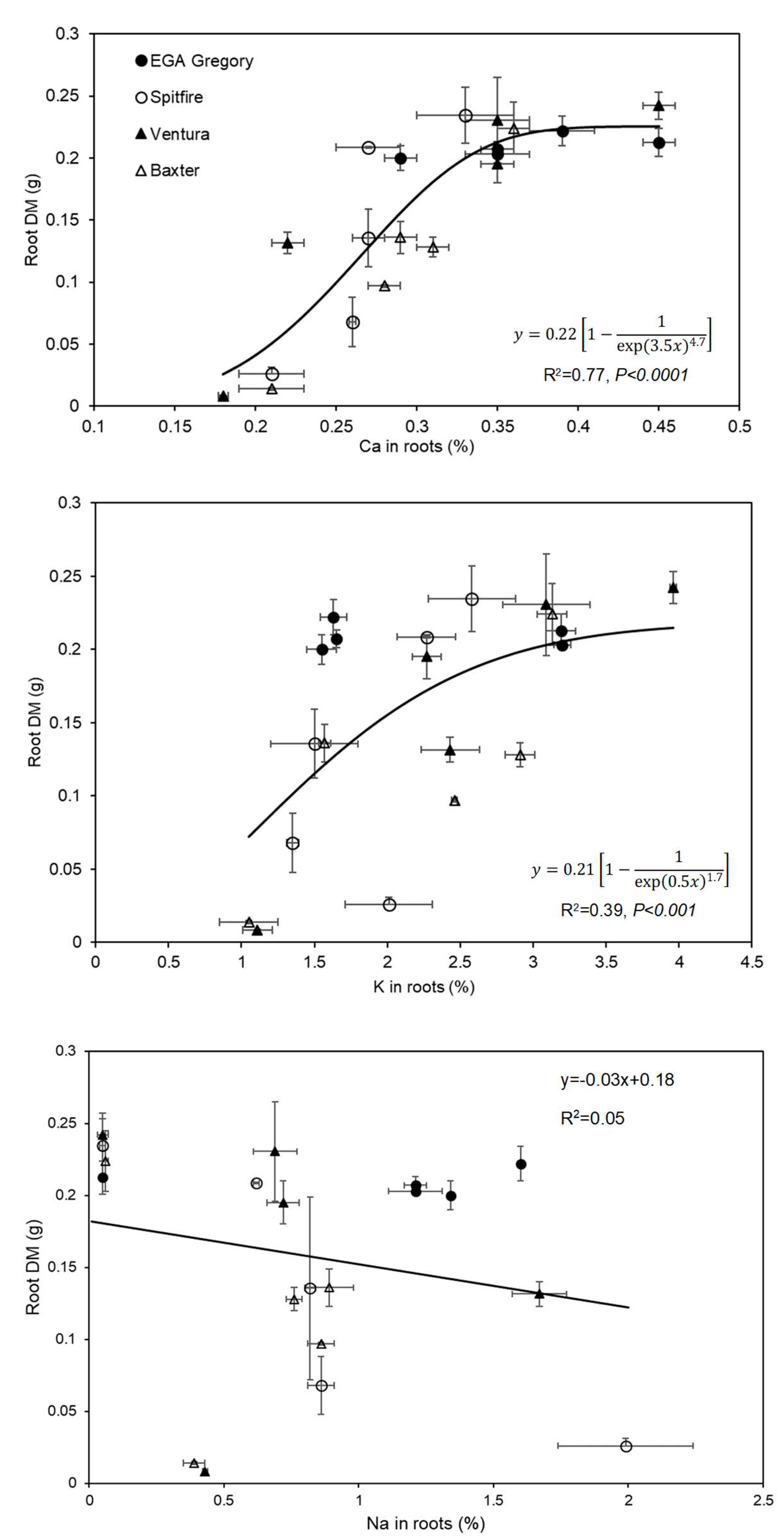
Figure 5.
Relationship between shoot DM and element concentrations Ca (R2=0.44), K (R2=0.43), Na (R2=0.52), and ratio of K: Na (R2=0.86) in YML of four wheat genotypes in four replicates grown in five SAR treatments. The bars inside the graphs present the standard error. The dotted vertical lines present the critical concentration for deficiency for Ca and K and critical concentration for toxicity for Na in YML [9].
Figure 5.
Relationship between shoot DM and element concentrations Ca (R2=0.44), K (R2=0.43), Na (R2=0.52), and ratio of K: Na (R2=0.86) in YML of four wheat genotypes in four replicates grown in five SAR treatments. The bars inside the graphs present the standard error. The dotted vertical lines present the critical concentration for deficiency for Ca and K and critical concentration for toxicity for Na in YML [9].
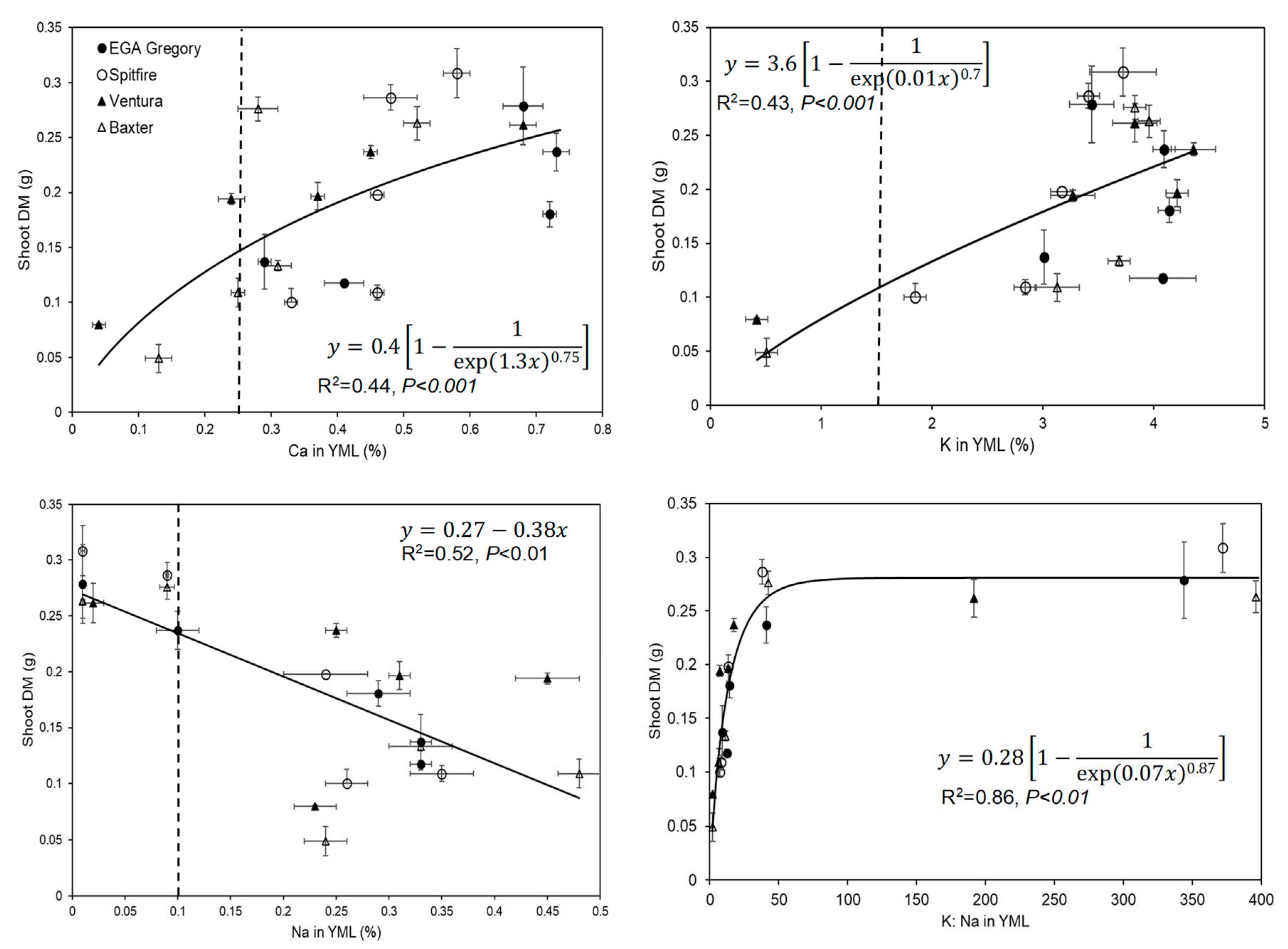

Table 1.
Concentrations of NaCl, Na2SO4, CaCl2.2H2O and CaSO4.2H2O added to nutrient solutions to prepare five different SAR values at constant ionic strength (I) of 31 mM in the basal nutrient solution.
Table 1.
Concentrations of NaCl, Na2SO4, CaCl2.2H2O and CaSO4.2H2O added to nutrient solutions to prepare five different SAR values at constant ionic strength (I) of 31 mM in the basal nutrient solution.
| SAR |
I mM |
NaCl mM |
Na2SO4 mM |
CaCl2.2H2O mM |
CaSO4.2H2O mM |
|---|---|---|---|---|---|
| 0 | 31 | 0.10 | 0.10 | 4.00 | 7.00 |
| 10 | 31 | 5.80 | 5.80 | 1.50 | 1.50 |
| 20 | 31 | 7.00 | 7.00 | 0.55 | 0.55 |
| 30 | 31 | 7.70 | 7.70 | 0.30 | 0.30 |
| 60 | 31 | 8.00 | 8.00 | 0.08 | 0.08 |
Table 2.
K: Na in the youngest mature leaf (YML) of four wheat genotypes in five different SAR treatments averaged over four replicates. The least significant difference (LSD) column allows comparison between genotypes (upper case letters are the same for genotypes which do not differ significantly (P>0.05) and between treatments (lower case).
Table 2.
K: Na in the youngest mature leaf (YML) of four wheat genotypes in five different SAR treatments averaged over four replicates. The least significant difference (LSD) column allows comparison between genotypes (upper case letters are the same for genotypes which do not differ significantly (P>0.05) and between treatments (lower case).
| Genotypes | SAR | K: Na | LSD |
|---|---|---|---|
| EGA Gregory | 0 | 344 | aA |
| 10 | 40.9 | bB | |
| 20 | 14.3 | cC | |
| 30 | 12.4 | cdCD | |
| 60 | 9.12 | cC | |
| Spitfire | 0 | 372 | aA |
| 10 | 37.9 | bcB | |
| 20 | 13.2 | cdCD | |
| 30 | 8.11 | cdCD | |
| 60 | 7.12 | cdCD | |
| Ventura | 0 | 192 | aB |
| 10 | 17.4 | bcBC | |
| 20 | 13.6 | cC | |
| 30 | 7.27 | cC | |
| 60 | 1.83 | fE | |
| Baxter | 0 | 396 | aA |
| 10 | 42.6 | bAE | |
| 20 | 11.2 | cC | |
| 30 | 6.52 | cdCD | |
| 60 | 2.13 | fEF | |
| P between treatments | <0.0001 | ||
| P between genotypes | 0.17 | ||
| Interaction between genotypes and treatments | 0.15 | ||
Table 3.
Comparison of significant traits of four wheat genotypes in this study and significant traits found in [27], Anzooman, Dang, Christopher, Mumford, Menzies and Kopittke [28] and Anzooman, Christopher, Dang, Taylor, Menzies and Kopittke [34]. High, medium and low have been presented as ‘H’, ‘M’ and ‘L’ respectively in the table. Nutrient concentrations are indicated as being either above the minimum critical level (>) or below (<).
Table 3.
Comparison of significant traits of four wheat genotypes in this study and significant traits found in [27], Anzooman, Dang, Christopher, Mumford, Menzies and Kopittke [28] and Anzooman, Christopher, Dang, Taylor, Menzies and Kopittke [34]. High, medium and low have been presented as ‘H’, ‘M’ and ‘L’ respectively in the table. Nutrient concentrations are indicated as being either above the minimum critical level (>) or below (<).
| Genotype | Relative Seedling Emergence in Soila | Rapid Germinationa | Seedling Emergence Forceb | Root Anglec | Ca Concentration in YML (SAR 30 and above)d | K Concentration in YML (SAR 60)d | Ca Concentration in Root (SAR 60)d | K Concentration in Root (SAR 60)d |
|---|---|---|---|---|---|---|---|---|
| EGA Gregory | Sensitive | L | L | L | H (>) | H (>) | H (>) | H (>) |
| Baxter | Sensitive | M | L | L | L (<) | L (<) | L (<) | M (>) |
| Ventura | Tolerant | H | H | H | L (<) | L (<) | L (<) | M (>) |
| Spitfire | Tolerant | H | H | H | H (>) | H (>) | H (>) | H (>) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



