Submitted:
31 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
Soil cadmium (Cd) contamination has become a serious problem in China. In this study, the goals of the field experiments were to understand how the application of nitrogen fertilizer at the full heading stage and filling stage reduced the uptake of Cd in rice by affecting the distribution of Cd in iron plaques on the root surfaces and subcellular in the root and flag leaf. The hydroponic culture experiments aimed to explore the effect of interaction or deficiency of nitrogen and cadmium on cadmium accumulation in rice at the late growth stage. The results showed that under the condition of having enough nitrogen supply during the early growth stage, applying nitrogen fertilizer during the full heading stage and filling stage resulted in a significant increase in the concentration of Fe and Cd on the root plaques at the milk stage and mature stage. Additionally, it increased the concentration and proportion of Cd in the soluble fraction of subcellular in the flag leaves at the milk stage. On the other hand, when there was a deficiency of nitrogen, the concentration of Fe in the root plaques increased significantly, while the concentration of Cd in the root plaques decreased significantly. Moreover, the proportion of Cd in the flag leaf cell walls increased significantly. Regardless of whether there was sufficient or deficient nitrogen supply during the early growth stage, applying nitrogen at the full heading stage reduced the Cd concentration in brown rice by 35.11% and 57.25%, respectively. Under hydroponic culture conditions with both Cd exposure and later-stage nitrogen supply significantly increased the Cd concentration in brown rice. However, in the absence of Cd, nitrogen supply significantly reduced the Cd concentration in brown rice. The accumulation of Cd in brown rice was significantly correlated with the accumulation of Fe and Mn at the late growth stage. Our findings indicate that when there is sufficient nitrogen nutrition during the early growth stage of rice, topdressing nitrogen fertilizer at the full heading stage combined with lime with mineral element fertilizers such as Fe and Mn is an effective approach to reduce the presence of cadmium in rice grains.
Keywords:
Rice cadmium
; Nitrogen fertilizer
; Fe plaque
; Cell wall
; available Cd
1. Introduction
Rice (Oryza sativa L.) is a major grain crop that provides food for more than half of the world’s population [1]. It is worth mentioning that rice is more susceptible to cadmium (Cd) absorption than other crops, resulting in a higher dietary Cd intake for populations with a rice-based diet through the food chain [2]. The issue of Cd-contaminated soils in China is particularly serious in the acidic paddy soil of Southern China. Soil acidification worsens the movement and accessibility of Cd in the soil, resulting in more severe rice Cd pollution compared to other regions in China [3]. Long-term rice consumption with excess Cd harms human health; thus, feasible strategies are urgently required to mitigate Cd risk in rice. Several strategies have been utilized, involving soil remediation, water and nutrient management, and planting uses with low Cd accumulation to minimize Cd accumulation in rice [3,4,5]. Fertilizer is basic required for normal growth in rice cultivation and application of fertilizers has also been tried as a strategy for reducing metal bioavailability to the plants [6,7,8]. Nitrogen management is typically seen as the most affordable, convenient, and effective process for minimizing Cd deposition in crops [9].
Nitrogen fertilizer is the most widely used fertilizer in agricultural production, and nitrogen plays an important role in plant growth and development. As a result, enough nitrogen fertilizer has been applied to the soil around the world to ensure crop yields [10]. However, nitrogen has been proven to be the most effective element involved in regulating the absorption and transport of Cd. After nitrogen application, the soil electrical conductivity increased, and the soil components of Cd were replaced by cations such as Fe2+, Ca2+, Zn2+, and so on, thus increasing the soluble Cd2+ in the soil [9,11]. Jalloh [12] discovered that there is a synergistic effect between NO3- and Cd, while there is an antagonistic effect between NH4+ and Cd. Hassan [13] further confirmed that rice grains treated with NO3- had a 35.7% higher Cd content compared to rice grains treated with NH4+. Wu [14] reported that increasing ammonium nutrition contributes to the inhibition of Cd uptake, xylem transport, and subsequent accumulation in rice, but not affect roots-to-shoots Cd transport. Several studies investigated the effect of urea on cadmium accumulation in rice. Some reports claim that the application of urea significantly decreased the Cd accumulation of rice grain [15]. Deng [16] reported that the use of urea instead of compound fertilizer has clearly resulted in a decrease in the concentration of Cd in brown rice, applying Mn fertilizer as a topdressing has further reduced the accumulation of Cd in the grain. Previous studies have shown that the absorption and accumulation of Cd by rice is closely related to the dose of nitrogen fertilizer. Increasing NH4+ ratios, rather than NO3-, has a greater ability to reduce the gene expression related to Cd transport by roots [14]. Other report also found that excessive NO3- enhanced Cd uptake by up-regulating the gene expression of OsIRT1 and OsNramp1 in rice under Cd stress. N can regulate the absorption and transport of Cd by regulating the non-specific gene expression of Cd and divalent cation transporter in plants. In addition, the application of N fertilizers enhances the antioxidant enzyme systems of rice and increases pectin and hemicellulose content in cell walls, which restricts Cd transport and helps control brown rice concentration [17].
N regulates the isolation and chelating ability of cell walls physiologically, thus regulating the adaptation of plants to Cd [9]. Cell walls isolation is the first contact and retention mechanism of Cd uptake by plant roots. Rice cell wall adsorption and vacuole interception had significant effects on grain Cd content. Due to the partition of the cell walls and vacuole, the retention of Cd in plant roots could limit the transport of Cd through the xylem to the aboveground part. Plant cell walls and vacuole partition could reduce the content of Cd entering organelles and reduce the damage of Cd to organelles and membrane structure, thus alleviating the toxicity of Cd to normal cell metabolism [18]. According to a recent study, under low Cd conditions, NH4+ supply promoted the transfer of Cd from the root cell walls to the cell and increased the pectin and protein binding form of intracellular Cd. However, at higher Cd levels, NH4+ promoted the deposition of Cd in the root cell wall and the accumulation of Cd in the root [19].
The late growth stage of rice is the critical period of Cd accumulation in grains, and the Cd absorbed by roots can be quickly transferred to grains. In addition, Cd accumulated in vegetative organs in the early stage of growth is also transported to grains with photosynthetic matter after grain filling. It was found that taking appropriate measures during the critical period of Cd accumulation in rice grains could effectively reduce Cd accumulation in rice grains [20,21,22]. Keeping a certain nitrogen level in the later growth stage of rice can maintain a high level of photosynthetic capacity of leaves, prolong grain filling time, delay leaf senescence, and inhibit the redistribution of elements in old leaves [23]. However, the effect of nitrogen application during the late growth stage on Cd accumulation in rice grains is still not clear. In this study, urea was used as a nitrogen source to study the effects of nitrogen application at the full heading stage and filling stage on iron and Cd concentrations on root plaque, Cd distribution in subcellular in roots and flag leaves, and Cd accumulation in rice grains. In addition, the effects of nitrogen, and Cd interaction or deficiency on grain Cd accumulation of rice after the full heading, and the relationship between mineral element accumulation in plants and Cd accumulation in grain under hydroponic culture were studied. Furthermore, the underlying mechanisms were clarified by studying the Cd accumulation effect by the application of nitrogen fertilizer during the late growth stage of rice.
2. Materials and methods
2.1. Field experimental site and soils
The paddy field of this research was in the village of Liuyang (28°.18′20″ N, 113.49′26″ E), Hunan Province, China. This area is in a subtropical monsoon humid climate zone, with a mean annual precipitation of 1400 mm and a mean temperature of 16.8–17.2 °C. The main soil type is loam soil. The present study conducted field experiments in the same field in 2018 and 2019. The basic physicochemical properties of the paddy soils were as follows: organic matter (OM) 33.17 g kg-1; pH (H2O) 5.22; cation exchange capacity (CEC) 11.98 cmol kg-1; total N 1.91 g kg-1; total P 0.64 g kg-1; total K 7.26 g kg-1; total Cd 0.72 mg kg-1; CaCl2-extracted Cd 0.34 mg kg-1.
2.2. Fertilizers and rice materials
The main fertilizers used in this experiment were urea (total nitrogen content ≥46%), calcium superphosphate (P2O5 content ≥12%), and potassium chloride (K20 content ≥60%) were purchased from a local agricultural shop in Liuyang, China. The conventional rice cultivar Yuzhenxiang was used for this experiment, which is the most commonly used rice variety in Hunan Province, China.
2.3. Experimental design
2.3.1. Experiment 1
The following treatments were applied to the field in 2018: application of nitrogen fertilizer 144 kg ha-1 and 36 kg ha-1 before transplanting and during panicle differentiation, respectively, as a Control treatment (CK), application of nitrogen fertilizer 108 kg ha-1, 36 kg ha-1 and 36 kg ha-1 before transplanting, during panicle differentiation and at the full heading stage, respectively (T1), application of nitrogen fertilizer 108 kg ha-1, 36 kg ha-1 and 36 kg ha-1 before transplanting, during panicle differentiation and at the filling stage (T2, 9 days after the full heading stage), respectively. According to local fertilizer practices, 750 kg ha-1 calcium superphosphate and 120 kg ha-1 potassium chloride were applied to each plot as base fertilizer, and 120 kg ha-1 potassium chloride was applied at the full heading stage. Base fertilizers were applied to the soil plough horizon (0-20 cm) 2 days before rice transplanting. Each treatment had three replications. Each plot area was 42 m2 with 7 m long and 6 m wide, including 50 cm paths between plots, each of the treatment plots was irrigated separately to avoid cross-contamination among different treatments. 25 days after germination, healthy and uniform seedlings were transplanted into the plots at a planting density of 20 cm by 20 cm on July 18, 2018. The plots were flooded (2-3 cm above the soil surface) from the full heading stage and continued until 7 days before maturity. All crop management was conducted in the field, with pests and diseases immediately prevented and controlled.
The seedlings were transplanted to the same field on July 20, 2019. No application of nitrogen fertilizer during the whole growth period as Control treatment (CK); application of nitrogen fertilizer 36 kg ha-1 at the full heading stage (F1); application of nitrogen fertilizer 36 kg ha-1 at filling stage (T2). Each treatment had three replications. Other measures are in line with 2018.
2.3.2. Experiment 2
The 25 days seedlings were transplanted to a Cd-contaminated field to the full heading stage, healthy and growing rice plants at the full heading stage were selected and returned to Hunan Agricultural University’s rice research institute. The soil of rice root was carefully cleaned, then cultured in deionized water for 12 hours, and then divided into 4 groups. The plants were transplanted into individual tall pots (28.1 cm high, 22 cm wide) with a 5 L hydroponic solution. The initial composition of chemicals used in-stock solutions in preparation for making nutrients solutions was: 2.9 mM N L-1(Urea), 0.32 mM P L-1 (NaH2PO4·2H2O), 1 mM K L-1 (K2SO4), 1mM Ca L-1 (CaCl2), 1.7 mM Mg L-1 (MgSO4·7H2O), 9.1×10-3 mM Mn L-1 (MnCl2·4H2O), 5.2×10-4 mM Mo L-1 Na2MoO4·2H2O2, 1.8×10-2 mM B L-1 (H3BO3), 1.5×10-4 mM Zn L-1 (ZnSO4·7H2O), 1.6×10-4 mM Cu L-1 (CuSO4·5H2O), 3.6×10-2 mM Fe L-1 (FeCl3·6H2O, Citric acid (monohydrate)). Two groups of rice plants were allowed to grow in a nutrient solution with nitrogen (+N), with the remaining grown in a -N solution accordingly. Then plants with and without N treatments were separated into two subgroups, and their growth continued to mature in the corresponding nutrient solution with 50×10-2 mM Cd L-1 (CdCl2·5H2O) and without Cd, allotted to each group individually. There were four treatments: (treatment A, +N +Cd), (treatment B, –N +Cd), (treatment C, +N -Cd), and (treatment D, –N -Cd), all of which were replicated in six pots for a total of 24 experimental units. There was also one plant placed into each container. At maturity, two plants from each treatment were used as one sample, resulting in a total of 12 plant samples for lab analysis. The nutrient solution was completely renewed once every 5 days. Adding deionized water every 2 days to 5 L to supplement the loss of rice transpiration, the solution pH adjustment to the value of 5.5 was made with the addition of 0.1 M HCl or NaOH to the media.
2.4. Rice and soil sampling
In experiment 1, the rice plants collected were collected at the milky stage and mature stage, and plant samples were rinsed with deionized water and separated into roots, shoots, panicles (milky stage), and brown rice. The fresh roots and flag leaves were selected for further analysis at the milky stage in 2019. Then, they were collected and placed in a − 80 °C freezer after being transported back to the laboratory. Soil samples were collected using the "five-points" sampling method at 0, 5, 7, 10, and 15 days after application of nitrogen fertilizer as well as at maturity in 2019. All soil samples were air-dried, ground, and passed through a 2.00 mm and a 0.15 mm nylon sieve, respectively. The rice yield of each plot was measured after threshing and sun-drying the seed.
Plant samples for the hydroponic experiment were collected at the mature stage in experiment 2, and rice plants were separated into brown rice (BR), husks (H), rachises (R), flag leaf (FL), internodes I (I-1), sheath I (S-1), node I (N-1, leaf II (L-2), internodes II (I-2), sheath II (S-2), node II (N-2), leaf III (L-3), internodes III (I-3), sheath III (S-3), node III (N-3), and old tissues (OT). There were sixteen parts to the hydroponic experiment.
All samples were dried at 105 °C; for 30 min and then brought to a constant weight at 70 °C, and dry matter was weighed and ground to pass through a 0.15 mm sieve for chemical analysis.
2.5. Chemical analysis of samples
The Fe/Mn plaques on fresh roots were extracted according to the dithionite-citrate-bicarbonate method (DCB) as described in detail by Zhou [24]. The powdered plant tissues were digested in a 4:1 solution of HNO3 and HClO4; a 1:2.5 soil-to-water ratio was used to determine the pH of the soil; soil available Cd was extracted with a 0.01M CaCl2 (pH 7.3) solution [25], and the extracts were analyzed by ICP-MS within two days. Cd contents in all samples were evaluated using an inductively coupled plasma optical emission spectrometry (ICP-MS, Agilent, USA). All samples were examined with certified reference materials (GBW07428-GSS-14 (soil) and GBW07603-GSV-2 (plant) from the Chinese Academy of Geological Sciences, Beijing) and sample blanks.
The Cd subcellular distributions in the roots and flag leaves were discovered using a differential centrifugation technique [26]. Using a chilled extraction buffer [1 mM DTT (dithiothreitol), 250 mM sucrose, and 50 mM Tris-HCl, pH 7.5] to homogenize samples of frozen flag leaf samples is a quick process. The homogenate was centrifuged at 3000 rpm for 15 minutes, and the precipitate—which mostly consisted of cell wall and cell wall debris was filtered through an 80-μm nylon fabric. At 4 °C, the filtrate was centrifuged for 30 minutes at 12,000 rpm. The resulting supernatant and residue were referred to as the soluble fraction and the cell organelle fraction, respectively.
2.6. Data and statistical analyses
The translocation factor (TF) was used to evaluate each plant tissue transport capacity was calculated using the following equations:
TFa-b= Cd concentrations of b/Cd concentrations of a.
All results were analyzed by one-way ANOVA with Duncan’s New Multiple Range Test to detect any significant difference between treatments (p<0.05). The correlation analysis was conducted using a Pearson’s correlation test with p<0.05 (two-tailed) significance level. All statistical analyses were performed using the SPSS 24.0 and all figures were produced using Origin 2021 software.
3. Results
3.1. Rice yields
The treatments decreased the yield from 6.82% to 11.93% compared to CK in 2018. The T2 treatment produced the lowest grain, and a significant difference was observed with CK (Figure 1). Compared with the control, the rice yields were boosted after topdressing of urea at the full heading and the filling stage, increasing 26.26% and 19.20%, respectively, as shown in Figure 1 in 2019.
3.2. Cd concentrations in rice different parts
The concentrations of Cd in different rice plant parts at the milky stage were all significantly affected by the application of nitrogen fertilizer at the full heading stage or filling stage in 2018 (Figure 2A). Compared with the CK, the T1 and T2 treatments decreased the Cd concentrations in roots and shoots by 39.65-51.95%, and 11.02-35.99%, respectively. T1 treatment decreased the Cd concentrations in panicles by 66.68%, and T2 treatment increased the Cd concentrations in panicles by 25.06%. At the mature stage, the T1 treatment decreased the concentrations in roots, shoots, and brown rice by 16.98%, 11.52%, and 35.11% compared with CK. The T2 treatment increased the Cd concentrations in roots and brown rice by 129.69% and 14.17%, respectively, and significantly decreased the Cd concentrations in shoots by 25.52% (Figure 2C). Compared with CK, the T1 and T2 treatments significantly increased the TFR-S by 35.84-225.67%, decreasing the TFS-B by 55.96-77.44% (Figure 2E).
Compared with CK, the F1 and F2 treatments markedly decreased the Cd concentration in roots, shoots, and panicles at the milky stage by 34.69-51.27%, 21.93-77.54%, and 82.94-83.31%, respectively (Figure 2B), significantly decreasing the Cd concentration in roots, shoot at the mature stage by 40.10-56.69%, 37.21-49.16%, respectively. There was a sharp decrease in brown rice Cd concentration of 57.25% in the F1 treatment, whereas no remarkable change was found in the Cd concentration of brown rice in the F2 treatment (Figure 2D). The risation of TFRS in the F1 and F2 treatments (14.45%, 2.52%), and the reduction of TFSB in the F1 treatment (15.26%) were small, however, the F2 treatment significantly increased TFSB by 59.59% (Figure 2F).
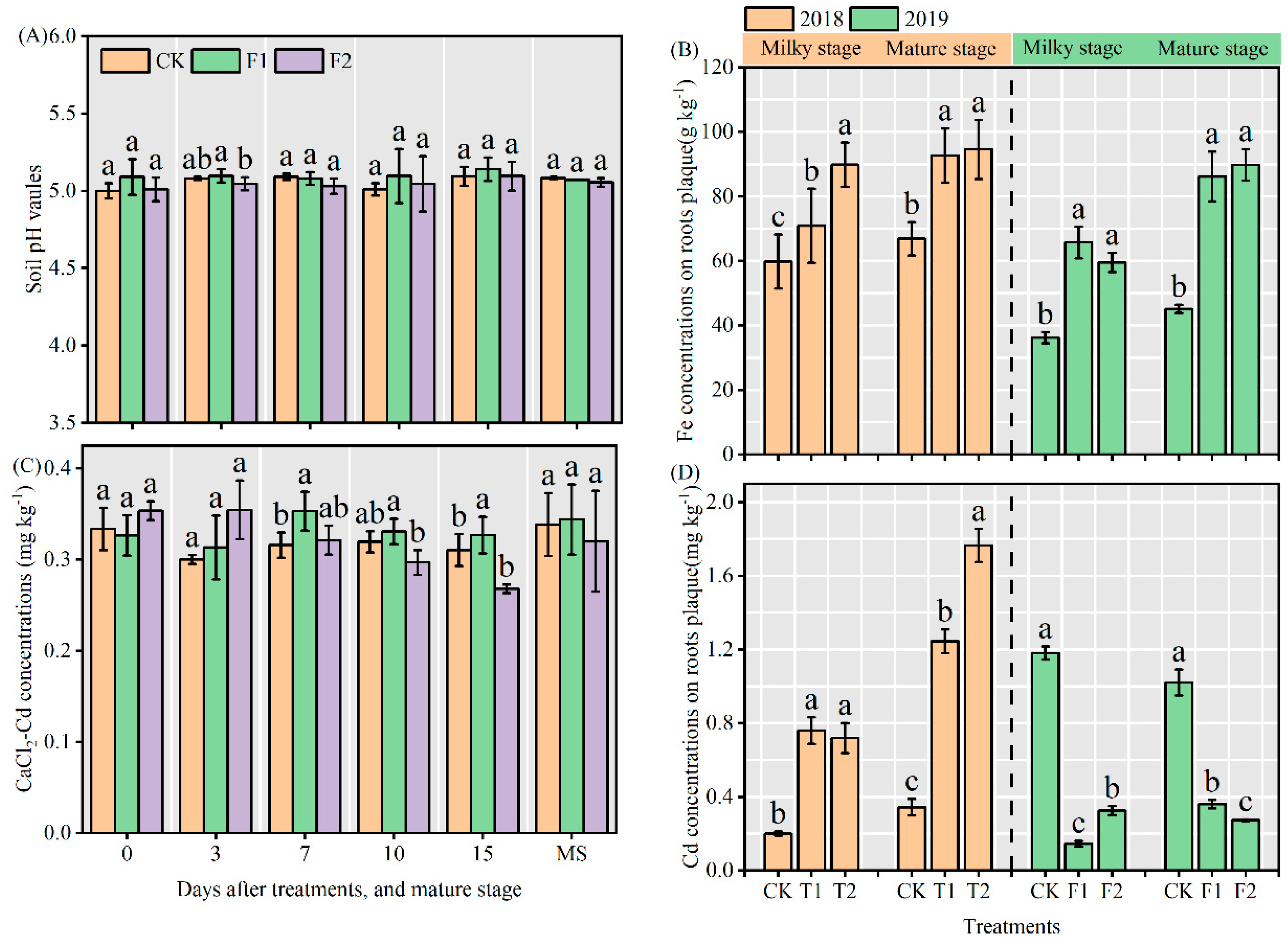
3.3. Soil pH values and CaCl2-Cd concentrations in soil
Compared with CK, F1, and F2 treatments had no significant effect on soil pH, but F2 treatment decreased significantly compared with F1 treatment 3 days after nitrogen fertilizer application. In general, F1 and F2 treatments slightly affected soil pH, reaching the maximum at 15 days after fertilization (Figure 3A). As shown in Figure 3C, compared with CK, F1 significantly increased the concentration of CaCl2 extracted Cd in the soil at 7 and 15 days after nitrogen application. From 7 days after treatments to the mature stage, the concentration of CaCl2-extracted Cd in soil was F1 > CK > F2. The concentration of CaCl2-extractable Cd in soil treated with F2 decreased gradually 3-15 days after nitrogen application.
3.4. Concentrations of Fe and Cd on roots plaque
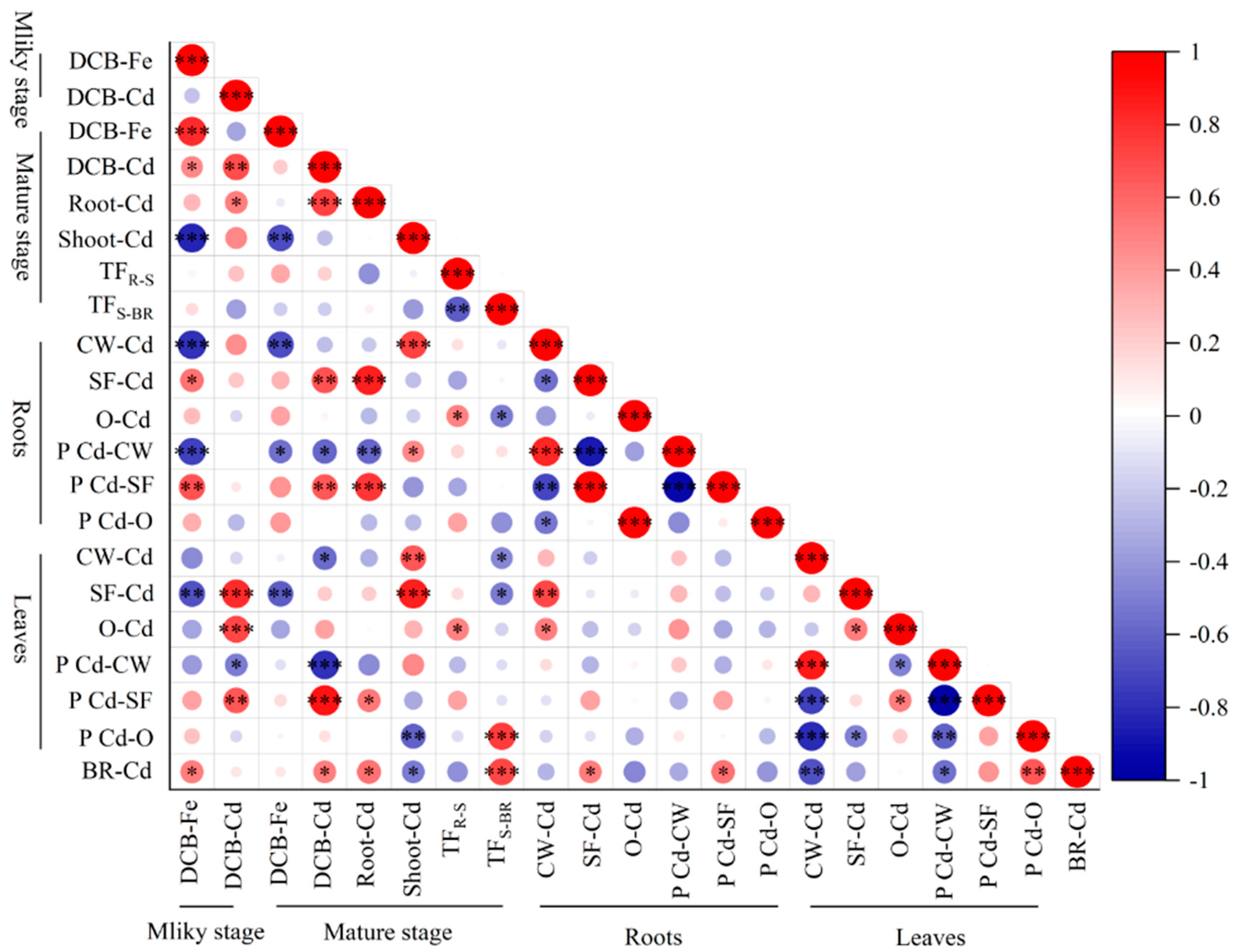
Generally, the Fe concentrations in roots plaque at the milky stage and mature stage were significantly increased by all nitrogen treatments compared with CK (Figure 3B), and the Fe concentrations in roots plaque at the mature stage were higher than that at the milky stage, which indicated that Fe in roots plaque accumulated with the progress of growth stage, meanwhile, applied of nitrogen fertilizer at the late growth stage of rice could promote the accumulation of Fe in the roots plaque. The Cd concentrations in roots plaque in 2018 were significantly increased by all of the nitrogen treatments, whereas the Cd concentrations in roots plaque showed the opposite trend in 2019 (Figure 3D). There was a significantly positively correlation between the Fe concentrations in root plaque at the milky stage, and the Cd concentrations in root plaque at mature stage and brown rice Cd concentrations (p<0.05, Figure 5).
Figure 3.
Soil pH vaules (a), CaCl2-extractable Cd concentration (c), Fe concentration on roots plaque (b), Fe concentration on roots plaque (d). Small letters indicate that there are significant differences among different treatments (p < 0.05).
Figure 3.
Soil pH vaules (a), CaCl2-extractable Cd concentration (c), Fe concentration on roots plaque (b), Fe concentration on roots plaque (d). Small letters indicate that there are significant differences among different treatments (p < 0.05).
3.5. Cd concentration of subcellular in roots and leaves
As evinced by Figure 4, compared with CK, the Cd concentrations and proportion of Cd in the cell walls in roots under T2 treatment were significantly decreased by 61.29% and 50%, respectively (Figure 4A); whereas the Cd concentrations and Cd distribution in soluble fraction were significantly increased by 351.83%, 44.13%. Under the T1 and T2 treatments, the Cd concentrations and Cd distribution in organelle were significantly increased by 215.84-362.23%, and 5.81-7.92%, respectively (Figure 4C). For the control, the Cd concentrations and Cd distribution of the flag leaf of the CK were ranked as follows: cell walls > soluble fraction > organelle, however, under the T1 and T2 conditions as follows: soluble fraction > cell walls > organelle, the T1 and T2 treatments respectively had signification 11.63% and 8.25% decreases in the Cd concentrations in the cell wall and significantly increased the Cd concentrations in the soluble fraction by 411.36-707.99%. Similar results were found on the effect of T1 and T2 treatments on the Cd distribution in the flag leaf.
Compared with CK, the Cd concentrations in the cell walls of roots were significantly decreased by 58.86% and 30.72% under the F1 and F2 treatments (Figure 4B), and the F1 treatment significantly increased the Cd concentrations in the organelle by 219.81%. F1 treatment significantly reduced the proportion of the cell walls distribution of Cd and significantly boosted the Cd distribution in the organelle. For the Cd concentrations in the flag leaf, F1 and F2 treatments boosted the Cd concentrations in the cell walls, and the F2 treatment significantly differed from CK. There was a significant negative correlation between the Cd concentration (p < 0.01), and the Cd distribution proportion (p < 0.05) of the cell walls and the Cd concentrations in brown rice (Figure 5). Moreover, the Cd concentrations in the soluble fraction and organelle were significantly decreased by 74.22-78.10%, and 52.43-62.45%, respectively. Similar results were found on the Cd distribution in the flag leaf (Figure 4D).
Figure 4.
Cd concentration (a, b) and proportion (c, d) of subcellular components in rice roots and flag leaves. Small letters indicate that there are significant differences among different treatments (p < 0.05).
Figure 4.
Cd concentration (a, b) and proportion (c, d) of subcellular components in rice roots and flag leaves. Small letters indicate that there are significant differences among different treatments (p < 0.05).
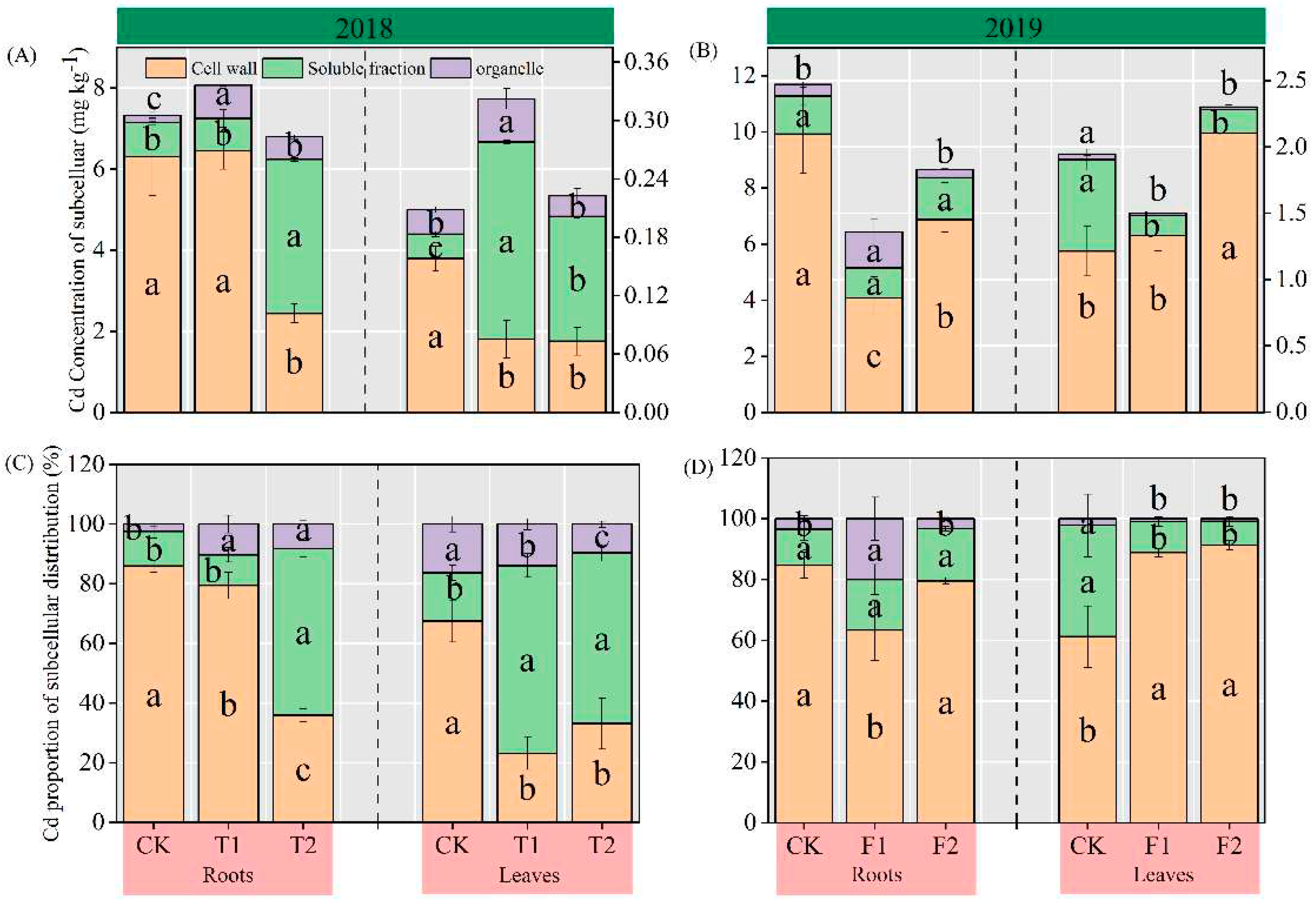
Figure 5.
Spearman’s correlation analysis of the relative abundances of brown rice Cd concentration, Fe and Cd concentration in the root plaque, Cd concentration and proportion of subcellular components in rice roots and flag leaves. Blue and red represent negative and positive correlations, respectively, with darker colors rep-resenting higher correlations. *, p < 0.05, **, p < 0.05, ***, p < 0.01.
Figure 5.
Spearman’s correlation analysis of the relative abundances of brown rice Cd concentration, Fe and Cd concentration in the root plaque, Cd concentration and proportion of subcellular components in rice roots and flag leaves. Blue and red represent negative and positive correlations, respectively, with darker colors rep-resenting higher correlations. *, p < 0.05, **, p < 0.05, ***, p < 0.01.
3.6. Concentrations of Cd in the aerial parts and brown rice in the Hydroponic culture
With the Cd condition provided, treatment A greatly increased the quantities of OT, I-3, N-1, I-1, S-1, FL, H, and BR Cd contents compared to treatment B, and it significantly decreased the amounts of Cd concentrations in N-2, L-2, S-2 (Figure 6A). There was a significant positive correlation between the Cd concentrations of I-3, N-1, I-1, S-1, H and brown rice Cd concentrations, and a significant negative correlation between the Cd concentrations of N-2, L-2, S-2 and brown rice Cd concentrations (Figure 6F). As depicted in Figure 6 B-F, compared with treatment B, it was observed that treatment A led to a significant decrease in the concentrations of Cu, Fe, Mn and Zn in N-3 and L-2, Fe, Mn, and Zn in I-2, and Cu, Fe, and Zn in S-2. Furthermore, all of these exhibited a negative correlation with the Cd content in brown rice.
The Cd contents of S-3, N-2, L-2, FL, I-1, and S-1 considerably increased in the absence of the Cd condition compared to the C and D treatments, whereas the content of S-1, R and H Cd greatly decreased (Figure 6A). There was a significant positive correlation between the Cd concentrations of I-1, N-1, R, H and brown rice Cd concentrations, and a significant negative correlation between the Cd concentrations of OT, I-3, N-2, L-2 and brown rice Cd concentrations (Fig. 6G). Compared to treatment D, treatment C decreased the content of Cu, Fe, Mn, and Zn in N-2, I-2, L-2, S-2, and H, as well as Fe, Mn, and Zn in FL and S-3, and Cu, Fe, and Zn in R. However, it increased the concentration of Cu, Fe, Mn, and Zn in I-3, L-3, and N-1 (Figure 6B-E). There was a positive correlation and significant positive correlation between the levels of Cu, Fe, Mn, and Zn in N-2, I-2, L-2, S-2, and H, as well as Mn in S-3, Cu, Fe, and Mn in R, with the content of Cd in brown rice. Conversely, there was a negative correlation and significant negative correlation between the levels of Cu, Fe, Mn, Zn in L-3 and N-1, Cu, Fe, Mn in I-3, and Zn in R, with the content of Cd in brown rice (Figure 6G).
4. Discussion
4.1. The effect of late nitrogen application on Cd of brown rice was not caused by soil
Cd accumulation in brown rice is a complex process in agroecosystems and is affected by various factors, including climate, soil characteristics, and nutrient transport. Of these factors, soil characteristics, particularly pH, have a significant impact on Cd solubility, mobility, and speciation in soil, as well as play a crucial role in determining Cd accumulation in rice plants [27]. Previous research has demonstrated that the presence of NO3- in rice plants stimulates the secretion of organic acids into the rhizosphere. This leads to an increase in soil CEC and the concentration of H+. Furthermore, it stimulates the augmentation of water-soluble Cd concentrations in the soil and the accumulation of Cd in rice crops [13]. On the other hand, when plants absorb NH4+, it results in the release of H+ into the soil, leading to soil acidification. This acidification, in turn, enhances the bioavailability of Cd in the soil [28]. After urea application, the hydrolysis of urea produces large amounts of ammonium, causing a short-term increase in pH. However, after NH4+ nitrification occurs, the soil pH decreases [29]. All the evidence supports the conclusion that irrespective of whether it is NO3-, NH4+, or urea, all of these substances pose a potential threat of decreasing soil acidity and enhancing the bioavailability of Cd. In the present study, the effects of nitrogen application at different stages of growth were examined. It was found that applying nitrogen at the full heading stage (F1) did not significantly affect soil pH (Figure 3A). However, it did result in significantly higher levels of CaCl2-Cd on the 7th and 15th day after treatment. There was no significant difference in soil pH or CaCl2-Cd in the treatment where nitrogen was applied at the filling stage (F2). Furthermore, we observed that applying nitrogen at the full heading stage led to a significant decrease in brown rice Cd concentrations, with reductions of 35.11% in 2018 and 57.25% in 2019 compared to the CK (Figure 2C-D). On the other hand, applying nitrogen at the filling stage resulted in a 14.17% increase in brown rice concentrations in 2018 compared to the control, with no obvious change in 2019. According to this study, it can be concluded that brown rice Cd concentrations varied opposite when nitrogen was applied during the full heading and filling stages. Interestingly, these changes were observed to be not directly caused by the effects of nitrogen fertilizer on soil pH and CaCl2-Cd.
The vegetative organs of rice possess the capability to impede the transportation of Cd. This ability allows them to accumulate a substantial quantity of Cd in the cell wall or store it in the vacuole. Deposition of Cd in the cell wall is an important mechanism that restricts the accumulation and movement of Cd in plants [30]. The study by Deng [26] suggests that pectin, polysaccharide components, hemicellulose 1, and functional groups, which are all constituents of the plant cell wall, are responsible for the majority of the Cd deposition in the cell wall of rice flag leaves and inhibit its transfer into the rice grain, they exhibit a significant positive correlation with the concentration of Cd in brown rice. 70-90% of Cd is bound to the cell walls due to the presence of functional groups like carboxyl (COO-), hydroxy (-OH), and thiol (-SH) [18]. The cell walls can bind with heavy metal cations, such as Cd, and prevent their transport into the cells [31]. Similar results were observed in our experiment, in that a significant increase in the proportion of Cd in the cell wall of the flag leaf, while the proportion of Cd in the organelles declined noticeably after applying N at the full heading stage and filling stage (Figure 4D). There was a significant positive correlation between the Cd proportion in the cell wall of the flag leaf and brown rice Cd concentrations (Figure 5). Interestingly, under the condition of nitrogen supply in the early stage of rice growth, regardless of whether the nitrogen fertilizer was topdressing at the full heading stage or filling stage, the content and proportion of cadmium in the cell wall of flag leaves did not decrease. Surprisingly, the content and proportion of Cd in the soluble fraction increased significantly (Figure 4A, C). Recent research has shown that the presence of nitrate ions increases the number of functional groups, including -OH, C=O, and -COOH, in the root cell walls of rice, as well as the amount of pectin and hemicellulose. On the other hand, the presence of ammonium ions decreases the amount of pectin, hemicellulose, and functional groups in the cell walls [32,33]. The assimilates required for grain filling in rice come from post-flowering photosynthesis and non-structural carbohydrates (NSC) stored in the leaf sheath and other organs before flowering. The latter contributes to about 1/6 to 1/3 of grain yield and its quantity depends on the growth conditions and nitrogen supply level. In addition, the pre-flowering stored substances are also important for initiating grain filling, and the transport rate and quantity of these substances play a crucial role in the early stage of grain filling [34,35]. We suspect that without N supply during the early stage of rice development, the application of nitrogen fertilizer at the full heading stage and filling stages, to meet the NSC needed for grain filling, a significant amount of nitrogen was absorbed and transported to compensate for the insufficient amount of photosynthetic products caused by the lack of nitrogen supply during the early stage. This, in turn, promotes the accumulation of Cd in the cell walls of the flag leaf. However, under conditions where there is N supply during the early stage, the rice plants are not sensitive to N when nitrogen fertilizer is applied at the full heading stage and filling stages, this can be observed from the rice grain yield (Figure 1).
Rice roots absorb Cd and quickly transport it to the grain at the filling stage, in addition, Cd stored in shoots before heading made a great contribution to the accumulation of Cd in grains [22,24]. In some recent studies, the focus has been on the processes of Cd transport in rice, including Cd uptake by the root, xylem loading, root-to-shoot translocation, phloem transfer at the stem, and transportation via the phloem to the grain [14,17]. In the current study, under the circumstance of nitrogen supply during the initial growth stage, there was a significant increase in TFR-S and a significant decrease in TFS-B at the full heading stage and filling stage (Figure 2E). Furthermore, the range of variation in the F1 treatment was greater than that in the F2 treatment. On the other hand, when there was no nitrogen supply during the early growth stage, there was a slight increase in TFR-S and a significant increase in TFS-BR in the F2 treatment (Figure 2F). Low nitrogen can promote the transport of carbon and nitrogen assimilates in stem sheaths, during the heading stage, the root system has stronger vitality compared to the grain-filling stage, allowing it to absorb more nutrients from the soil. This indicates that its leaves have stronger physiological activity (photosynthetic capacity). The photosynthetic products produced by functional leaves can meet most of the grain filling needs, reducing relatively the proportion of assimilates transported from old leaves and stems to rice grains. As a result, the entry of Cd into rice grains is reduced through nutrient transport pathways. That was the possible reason for the Cd concentration in brown rice of T1 and F1 were lower than others.
4.2. Fe and Mn accumulation in rice significantly affected Cd accumulation in brown rice.
We conducted a solution culture experiment to investigate how the supply of N affects the accumulation of Cd in rice grains during the late growth stage. The purpose was to eliminate any influence from soil factors. Our findings indicate that the presence of Cd, and N supply resulted in a 57.69% increase in the concentration of Cd in brown rice, compared to N deficiency (Figure 6A). This outcome aligns with previous research, which has demonstrated that N promotes the accumulation of cadmium in brown rice. However, when Cd is not present, the supply of N significantly reduces the Cd content in brown rice. Further analysis found that the addition of Cd or did not, N supply decreased the contents of Cu, Fe, Mn, and Zn in N-2, I-2 L-2, and S-2 (except for Fe in N-2 and Mn in S-2 under A treatment, Figure 6 B–E). Cd has the ability to enter plant cells using the same uptake systems as essential mineral elements like Fe2+, Mn2+, and Zn2+ due to their similar physiochemical properties [2,17]. Cd has the ability to compete with Fe for transporters like OsIRT1/2, which are responsible for transporting Fe2+ and are sensitive to Cd. Cd can also compete with Mn for transporters like OsNRAMP5, which are involved in the uptake of both Cd and Mn. This competition among Fe, Mn, and Cd for the same uptake pathway in plants is a contributing factor to the reduction of Cd uptake in plants [36]. Our results show that the condition of the addition of Cd, most of these elements in N-2, I-2 L-2, and S-2, especially Fe and Mn were significantly negatively correlated with Cd content in brown rice (Figure 6F), while they were positively correlated with brown rice Cd content without the condition of Cd (Figure 6G). In other words, the application of N can regulate the distribution of Fe, Mn and other elements in rice plants, and then accumulate Cd in brown rice.
Another reason for Fe can reduce Cd uptake in plants is Fe absorbed into root tissue could compete with Cd for adsorption sites, while Cd without adsorption sites was excluded from the cell wall of the rice root surface [36]. Iron plaque that forms on the root surfaces of rice could adsorb Cd without adsorption sites on the root surface, which can affect the chemical behavior and bioavailability of heavy metals in the soil through adsorption and coprecipitation and plays an important role in the absorption of heavy metals by roots and internal transport within plants [4,6,16]. Zhang [37] reported that low Fe or excess Fe facilitated the uptake of Cd in rice roots, as low Fe up-regulated the expression of Cd-transport related genes and excess Fe enhanced Cd enrichment on the root by ion plaque, and soil fertilization with Fe at the mature stage strongly reduced the grain Cd concentration. Recent research reported that the application of basal alkaline fertilizers [16], microbial organic fertilizer [6], and Fe fertilizer [38] could increase the adsorption and fixation of Cd by iron plaque, and more Cd could not be absorbed directly into the rice. In the present study, we found that applying nitrogen during the late growth stage significantly increased the Fe content on the root plaque of rice at the milky stage and mature stage, and significantly increased the Cd content on root plaque when N was supplied during the early growth stage and decreased significantly under the condition of N deficiency in the early growth stage. The analysis of Figure 5 revealed that there is no strong correlation between the Fe content and Cd content on the root plaque, this finding aligns with previous studies, suggesting that iron plaque does not affect the adsorption of Cd [39]. However, it was negatively correlated with shoots Cd content at the mature stage, the content and proportion of Cd in the root cell walls, and the content of Cd in the soluble fraction of the flag leaf at the milky stage. Thus, the application of N increased the Fe concentration on the roots plaque and reduced Cd entering the cell walls of roots, also limiting the distribution of Cd from roots to shoots.
5. Conclusion
Our findings show that: (1) The application of nitrogen fertilizer at the full heading stage increased the distribution of Cd in the cell walls and soluble fraction of flag leaves at the milk stage, increased Fe plaque on the root surfaces, decreased the transport of Cd from shoots to brown rice, and significantly decreased the Cd concentration of brown rice. (2) The effect of N supply on Cd accumulation of brown rice during the late growth stage depends on whether there is available Cd in the growth medium. It promotes Cd accumulation in brown rice when there is the presence of Cd, but it is the opposite when there is Cd deficiency. The accumulation of Cd in brown rice was significantly correlated with that of Fe and Mn in rice. Accordingly, on the premise of ensuring the normal growth of rice in the early stage, topdressing nitrogen fertilizer at the full heading stage combined with lime and micro-fertilizer containing mineral elements such as Fe and Mn can be used as a feasible way to reduce rice cadmium.
Author Contributions
Y.Z.: Data curation, Formal analysis, Methodology, Visualization, Writing—original draft. H.X.: Methodology, Writing—review and editing. Q.C.: Investigation, Project administration. Q.C.: Investigation, Project administration. H.A.: Funding acquisition, Supervision, Writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Joint Funds of the National Natural Science Foundation of China (32172107), and National Rice Industry Technology System Cultivation and soil Fertilizer Post expert Project of China (CARS-01).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
Special thanks to anonymous reviewers for their valuable comments. In addition, the authors gratefully acknowledge every teacher, classmate, and friend who helped the authors with their experiment and writing.
Conflicts of Interest
The authors declare no competing interests.
References
- Wang, P.; Chen, H.; Kopittke, P.M.; Zhao, F.-J. Cadmium contamination in agricultural soils of China and the impact on food safety. Environ. Pollut. 2019, 249, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Uraguchi, S.; Fujiwara, T. Cadmium transport and tolerance in rice: perspectives for reducing grain cadmium accumulation. Rice. 2012, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, E.; Agapiou, A.; Anastopoulos, I.; Omirou, M.; Ioannides, I.M. The effects of different soil nutrient management schemes in nitrogen cycling. J. Environ. Manag. 2019, 243, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, H.; Huang, D.; Xu, C.; Zhu, H.; Zhu, Q. Water managements limit heavy metal accumulation in rice: Dual effects of iron-plaque formation and microbial communities. Sci. Total Environ. 2019, 687, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Mao, P.; Zhuang, P.; Li, F.; Mcbride, M.B.; Ren, W.D.; Li, Y.Y.; Li, Y.W.; Mo, H.; Fu, H.Y.; Li, Z.A. Phosphate addition diminishes the efficacy of wollastonite in decreasing Cd uptake by rice (Oryza sativa L.) in paddy soil. Sci. Total Environ. 2019, 687, 441–450. [Google Scholar] [CrossRef]
- Kong, F.; Lu, S. Effects of microbial organic fertilizer (MOF) application on cadmium uptake of rice in acidic paddy soil: Regulation of the iron oxides driven by the soil microorganisms. Environ. Pollut. 2022, 307, 119447. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, C.; Wang, C.; Huang, Y.; Liu, Z. Increasing phosphate inhibits cadmium uptake in plants and promotes synthesis of amino acids in grains of rice. Environ. Pollut. 2019, 257, 113496. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Zhu, T.; Gao, Y.; Wang, Y.; Ning, C.; Björn, L.O.; Chen, D.; Li, S. Effects of Ca addition on the uptake, translocation, and distribution of Cd in Arabidopsis thaliana. Ecotoxicol. Environ Saf. 2017, 139, 228–237. [Google Scholar] [CrossRef]
- Yang, Y.; Xiong, J.; Tao, L.; Cao, Z.; Tang, W.; Zhang, J.; Yu, X.; Fu, G.; Zhang, X.; Lu, Y. Regulatory Mechanisms of Nitrogen (N) on Cadmium (Cd) Uptake and Accumulation in Plants: A Review. Sci. Total Environ. 2019, 708, 135186. [Google Scholar] [CrossRef]
- Chen, Z.; Li, X.; Liu, T.; Fu, H.; Yuan, X.; Cheng, Q.; Liao, Q.; Zhang, Y.; Li, W.; Sun, Y.; Yang, Z.; Ma, J.; Li, X. Strategies for fertilizer management to achieve higher yields and environmental and fertilizer benefits of rice production in China. Sci. Total Environ. 2023, 904, 166325. [Google Scholar] [CrossRef]
- Akahane, I.; Makino, T.; Maejima, Y. Effects of Nitrogen Fertilizer, pH, and Electrical Conductivity on the Solubility of Cadmium in Soil Solution. Pedologist. 2010, 53, 101–107. [Google Scholar] [CrossRef]
- Jalloh, M.A.; Chen, J.; Zhen, F.; Zhang, G. Effect of different N fertilizer forms on antioxidant capacity and grain yield of rice growing under Cd stress. J. Hazard. Mater. 2009, 162, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.J.; Wang, F.; Ali, S.; Zhang, G. Toxic Effect of Cadmium on Rice as Affected by Nitrogen Fertilizer Form. Plant Soil. 2005, 277, 359–365. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, W.; Xu, S.; Shi, S.; Wen, D.; Huang, Y.; Peng, L.; Deng, T.; Du, R.; Li, F.; Wang, X.; Wang, F. Increasing ammonium nutrition as a strategy for inhibition of cadmium uptake and xylem transport in rice (Oryza sativa L.) exposed to cadmium stress. Environ. Exp. Bot. 2018, 155, 734–741. [Google Scholar] [CrossRef]
- Xu, C.; Wu, Z.; Zhu, Q.; Zhu, H.; Zhang, Y.; Huang, D. Effect of coated urea on cadmium accumulation in Oryza sativa L. grown in contaminated soil. Environ. Monit. Assess. 2015, 187, 716. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Chen, Y.; Yang, Y.; Lu, L.; Yuan, X.; Zeng, H.; Zeng, Q. Cadmium accumulation in rice (Oryza sativa L.) alleviated by basal alkaline fertilizers followed by topdressing of manganese fertilizer. Environ. Pollut. 2020, 262, 114289. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiong, J.; Chen, R.; Fu, G.; Chen, T.; Tao, L. Excessive nitrate enhances cadmium (Cd) uptake by up-regulating the expression of OsIRT1 in rice (Oryza sativa). Environ. Exp. Bot. 2016, 122, 141–149. [Google Scholar] [CrossRef]
- Liu, J.; Qu, P.; Zhang, W.; Dong, Y.; Li, L.; Wang, M. Variations among rice cultivars in subcellular distribution of Cd: The relationship between translocation and grain accumulation. Environ. Exp. Bot. 2014, 107, 5–31. [Google Scholar] [CrossRef]
- Chai, M.; Li, R.; Shen, X.; Tam, N.F.Y.; Zan, Q.; Li, R. Does ammonium nitrogen affect accumulation, subcellular distribution and chemical forms of cadmium in Kandelia obovata? Ecotoxicol. Environ Saf. 2018, 162, 430–437. [Google Scholar] [CrossRef]
- Tian, T.; Zhou, H.; Gu, J.; Jia, R.; Li, H.; Wang, Q.; Zeng, M.; Liao, B. Cadmium accumulation and bioavailability in paddy soil under different water regimes for different growth stages of rice (Oryza sativa L.). Plant Soil. [CrossRef]
- Rodda, M.S.; Li, G.; Reid, R.J. The timing of grain Cd accumulation in rice plants: the relative importance of remobilisation within the plant and root Cd uptake post-flowering. Plant Soil. 2011, 347, 105–114. [Google Scholar] [CrossRef]
- Fujimaki, S.; Suzui, N.; Ishioka, N.S.; Kawachi, N.; Ito, S.; Chino, M.; Nakamura, S.I. Tracing Cadmium from Culture to Spikelet: Noninvasive Imaging and Quantitative Characterization of Absorption, Transport, and Accumulation of Cadmium in an Intact Rice Plant. Plant Physiol. 2010, 152, 1796–1806. [Google Scholar] [CrossRef]
- GARNETT, T.P. Distribution and Remobilization of Iron and Copper in Wheat. Ann. Bot. 2005, 95, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhu, W.; Yang, W.; Gu, J.; Gao, Z.; Chen, L.; Du, W.; Zhang, P.; Peng, P.; Liao, B. Cadmium uptake, accumulation, and remobilization in iron plaque and rice tissues at different growth stages. Ecotoxicol. Environ Saf. 2018, 152, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, J.; Huang, Q.; Tang, S.; Wang, J.; Hu, P.; Shao, G. Can liming reduce cadmium (Cd) accumulation in rice (Oryza sativa) in slightly acidic soils? A contradictory dynamic equilibrium between Cd uptake capacity of roots and Cd immobilisation in soils. Chemosphere. 2018, 193, 547–556. [Google Scholar] [CrossRef]
- Deng, X.; Chen, B.; Chen, Y.; Jiang, L.; Hu, Y.; Yang, Y.; Rong, X.; Peng, L.; Zeng, Q. Flag leaf cell wall functional groups and components play a crucial role in the accumulation and translocation of Cd in rice grain via foliage application of humic acid. Ecotoxicol. Environ Saf. 2022, 239, 113658. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Deng, X.; Ma, Q.; Zhao, Y.; Wang, A.; Zhang, X.; Zeng, Q. Cadmium accumulation in brown rice (Oryza sativa L.) depends on environmental factors and nutrient transport: A three-year field study. Sci. Total Environ. 2023, 903, 166942. [Google Scholar] [CrossRef]
- Hinsinger, P.; Plassard, C.; Tang, C.; Jaillard, B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: A review. Plant Soil. 2003, 248, 43–59. [Google Scholar] [CrossRef]
- Hong-Duck, R.; Min-Seob, K.; Chung, E.; Baek, U.; Kim, S.; Kim, D.; Kim, Y.; Lee, J. Assessment and identification of nitrogen pollution sources in the Cheongmi River with intensive livestock farming areas, Korea. Environ. Sci. Pollut. Res. 2018, 25, 13499–13510. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, Z. Mechanisms of cadmium phytoremediation and detoxification in plants. Crop J. 2021, 9, 521–529. [Google Scholar] [CrossRef]
- Li, A.; Deng, H.; Jiang, Y.; Ye, C.; Yu, B.; Zhou, X.; Ma, A. Superefficient removal of heavy metals from wastewater by Mg-loaded biochars: adsorption characteristics and removal mechanisms. Langmuir. 2020, 36, 9160–9174. [Google Scholar] [CrossRef]
- Wang, W.; Zhao, X.; Chen, F.; Dong, X.; Lan, P.; Ma, J.; Shen, R. Altered cell wall properties are responsible for ammonium-reduced aluminium accumulation in rice roots. Plant Cell Environ. 2015, 38, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Nkoh, J.N.; Abdulaha-Al Baquy, M.; Dong, G.; Li, J.; Xu, R. Plants alter surface charge and functional groups of their roots to adapt to acidic soil conditions. Environ. Pollut. 2020, 267, 115590. [Google Scholar] [CrossRef] [PubMed]
- Okamura, M.; Arai-Sanoh, Y.; Yoshida, H.; Mukouyama, T.; Adachi, S.; Yabe, S.; Nakagawa, H.; Tsutsumi, K.; Taniguchi, Y.; Kobayashi, N.; Kondo, M. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield. Field Crop Res. 2018, 219, 139–147. [Google Scholar] [CrossRef]
- Li, G.; Pan, J.; Cui, K.; Yuan, M.; Hu, Q.; Wang, W.; Mohapatra, P.; Nie, L.; Huang, J.; Peng, S. Limitation of Unloading in the Developing Grains Is a Possible Cause Responsible for Low Stem Non-structural Carbohydrate Translocation and Poor Grain Yield Formation in Rice through Verification of Recombinant Inbred Lines. FRONT PLANT SCI. 2017, 8, 1369. [Google Scholar] [CrossRef] [PubMed]
- Hiromi, N.; Ippei, O.; Yasuhiro, I.; Satoshi, M.; Naoko, K.N. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci. Plant Nutr. 2006, 52, 464–469. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, D.; Xu, C.; Zhu, H.; Feng, R.; Zhu, Q. Fe fortification limits rice Cd accumulation by promoting root cell wall chelation and reducing the mobility of Cd in xylem. Ecotoxicol. Environ Saf. 2022, 240, 113700. [Google Scholar] [CrossRef]
- Huang, G.; Ding, C.; Hu, Z.; Cui, C.; Zhang, T.; Wang, X. Topdressing iron fertilizer coupled with pre-immobilization in acidic paddy fields reduced cadmium uptake by rice (Oryza sativa L.). Sci. Total Environ. 2018, 636, 1040–1047. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Zhang, F. Role of iron plaque in Cd uptake by and translocation within rice (Oryza sativa L.) seedlings grown in solution culture. Environ. Exp. Bot. 2007, 59, 314–320. [Google Scholar] [CrossRef]
Figure 1.
Rice yield. Small letters indicate that there are significant differences among different treatments (p < 0.05).
Figure 1.
Rice yield. Small letters indicate that there are significant differences among different treatments (p < 0.05).
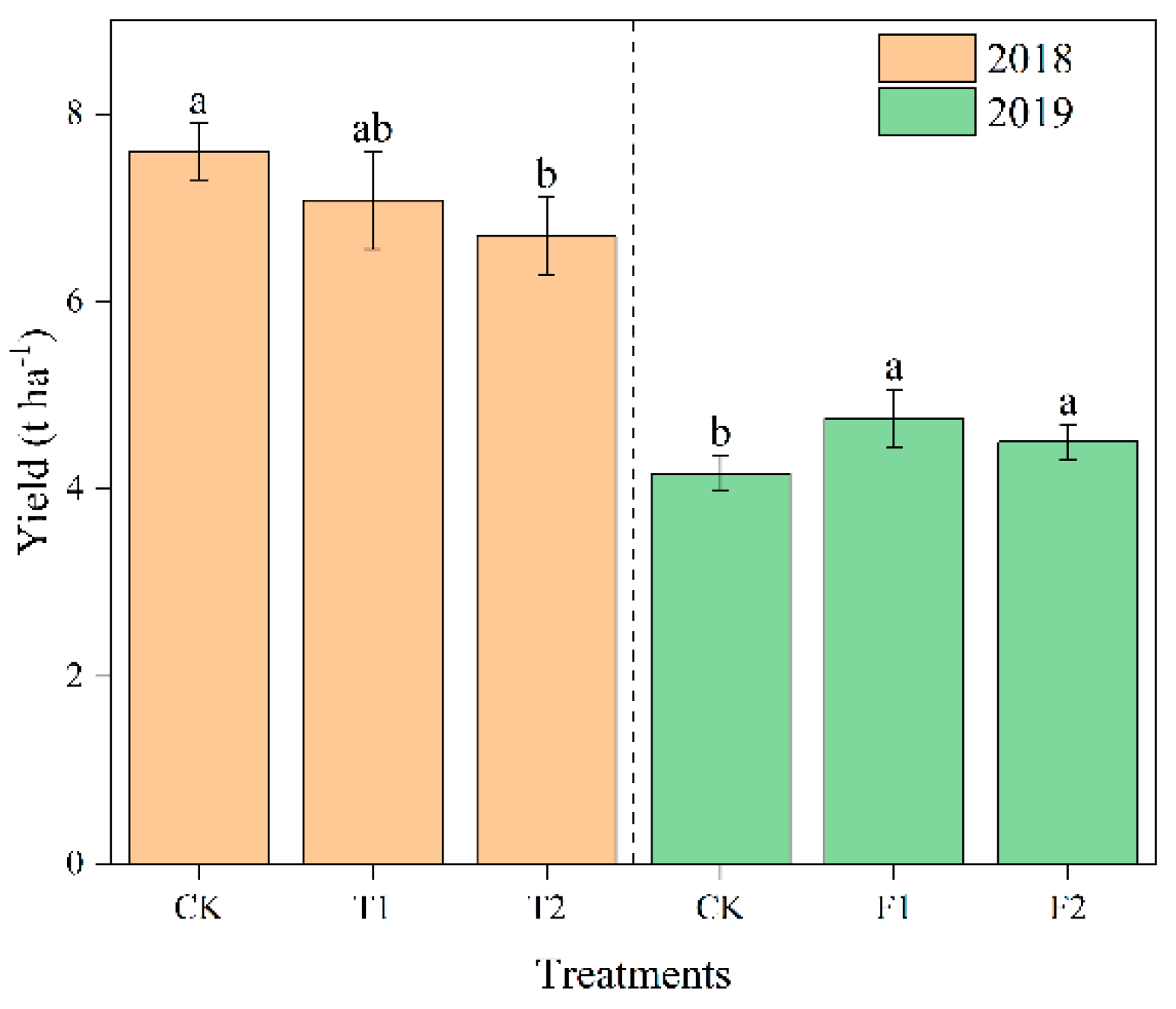
Figure 2.
Cadmium concentration in different parts of rice at the milky stage (a, b) and mature stage (c, d) and Cd transport coefficient at the mature stage (e, f). Small letters indicate that there are significant differences among different treatments (p < 0.05).
Figure 2.
Cadmium concentration in different parts of rice at the milky stage (a, b) and mature stage (c, d) and Cd transport coefficient at the mature stage (e, f). Small letters indicate that there are significant differences among different treatments (p < 0.05).
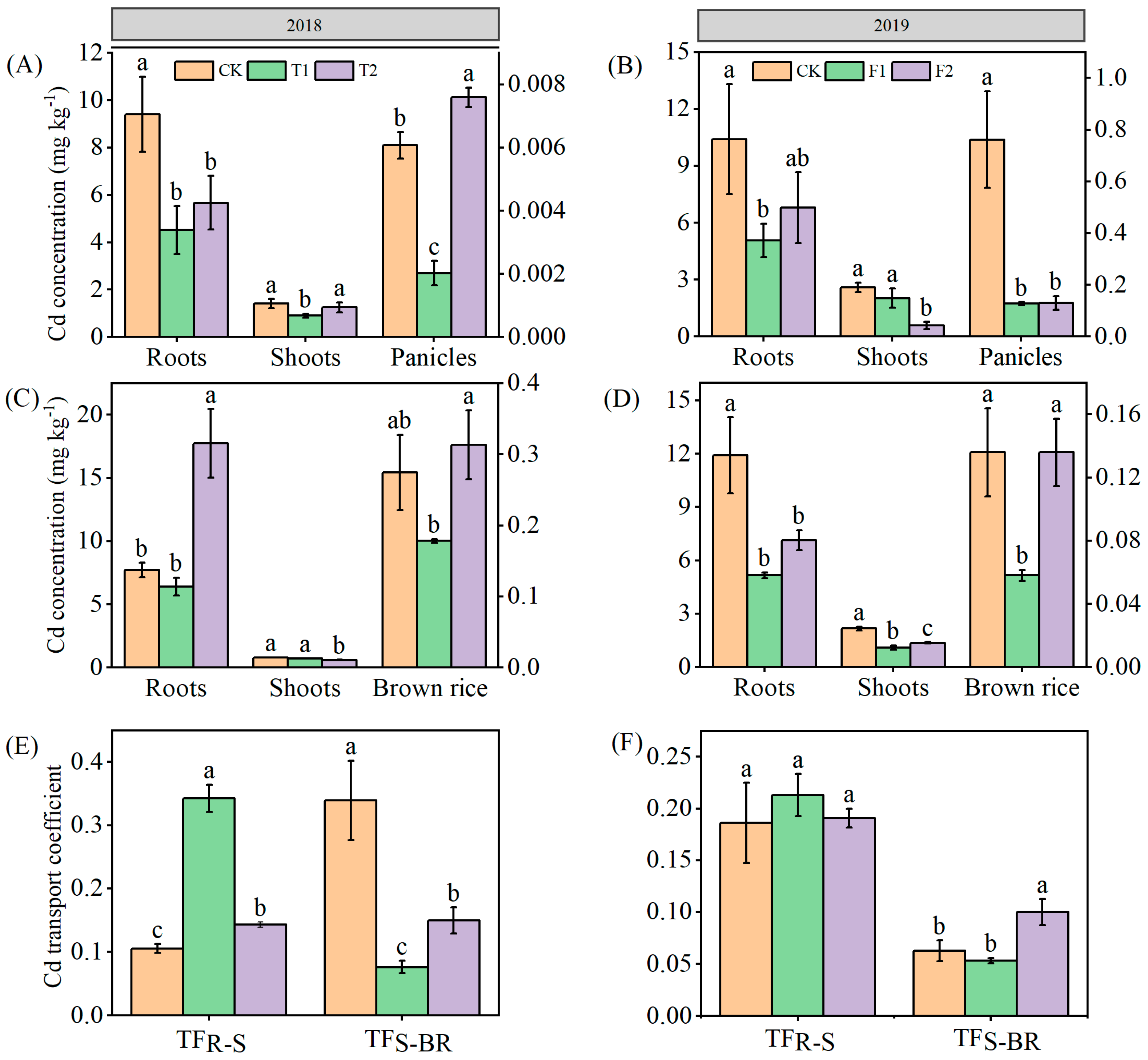
Figure 6.
a, b, c, d, and e represent the content of Cd, Cu, Fe, Mn, and Zn in rice (mg kg-1), respectively. f, and g represent the correlation coefficient between the element content and Cd content of brown rice under the A and B, C and D treatment, respectively. *, p < 0.05, **, p < 0.05, ***, p < 0.01.
Figure 6.
a, b, c, d, and e represent the content of Cd, Cu, Fe, Mn, and Zn in rice (mg kg-1), respectively. f, and g represent the correlation coefficient between the element content and Cd content of brown rice under the A and B, C and D treatment, respectively. *, p < 0.05, **, p < 0.05, ***, p < 0.01.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



