Submitted:
30 October 2023
Posted:
31 October 2023
You are already at the latest version
Abstract
China faces a persistent deficiency in feed protein resources. Enhancing the utilization efficiency of indigenous feed protein resources emerges as a viable strategy to alleviate the current deficit in protein feed supply. Corn gluten meal (CGM), characterized by high proportion of crude protein and glutamine, is predominantly employed in animal feed. Nonetheless, the water-insolubility of CGM protein hampers its protein bioavailability when utilized as feed material. This study aimed to augment protein bioavailability, liberate glutamine peptides from CGM, and produce gluta-mine-enriched CGM fermented feed. We executed a co-fermentation protocol using Bacillus subtilis A5, Lactobacillus 02002, and acid protease to generate the CGM fermented feed. Subsequent in vivo experiment with broilers were conducted to assess the efficacy of the fermented product. Findings revealed that the soluble protein, glutamine, small peptides, and lactic acid contents in the fer-mented feed increased by 69.1%, 700%, 47.6%, and 125.9%, respectively. Incorporating CGM fermented feed into the diet markedly enhanced the growth performance and intestinal health of broilers, positively modulated the cecal microbiota structure, and augmented the population of beneficial bacteria, specifically Lactobacillus. These results furnish both experimental and theoretical foundations for deploying CGM fermented feed as an alternative protein feed resource.
Keywords:
by-product
; glutamine peptide
; intestinal health
; gut microbiota
1. Introduction
In China, the scarcity of feed protein resources significantly impedes the swift advancement of the feed industry and livestock breeding. Soybean meal (SBM) serves as the primary resource of high-quality protein in animal diets, yet 80% of the total soybean consumption (precursor of SBM) is dependent on imports. Alternative protein resources, such as cottonseed meal, canola meal, and rapeseed meal, are seldom incorporated into animal diets due to their inferior amino acid profile, higher fiber content relative to SBM, and the presence of macromolecule proteins [1,2,3]. Consequently, enhancing the utilization efficiency of indigenous feed protein resources emerges as a viable strategy to fulfill the existing supply requirements and meet future demands for protein feed resources.
Termed “the second brain”, the small intestine is crucial for nutrient absorption, metabolism, and the recognition of dietary signals. It also establishes a physical and biological defense against detrimental entities, including pathogens and environmental toxins. Disruptions in intestinal architecture and microbiota equilibrium can compromise the intestinal barrier's functionality, resulting in diminished immunity and stunted growth performance in animals. Glutamine (Gln), a nutrient pivotal for gut health, is integral in promoting intestinal epithelial proliferation, preserving intestinal morphological integrity, and safeguarding the intestinal barrier function [4]. Nonetheless, the direct application of Gln in the feed industry is constrained by its instability and poor water solubility. In contrast, glutamine peptides, known for their enhanced stability and solubility, exhibit superior intestinal modulation capabilities compared to the Gln monomer [5,6] and demonstrate potent immunoregulatory and health-boosting properties as feed additives [7,8,9]. Hence, glutamine peptides represent an efficacious substitute for Gln in protecting intestinal health.
Corn gluten meal (CGM), a significant by-product of the corn starch industry, is primarily utilized in animal feed as a protein source. Over the past five years, China's annual CGM yield has reached 3 million tons. Comprising approximately 60-70% crude protein, which includes 65% zein and 30% glutelin, the Gln content accounts for about one-fourth and one-third of the total amino acids in zein and glutelin, respectively [10,11]. Despite its high crude protein content, the bioavailability of CGM protein is restricted due to the water insolubility of zein and glutelin [12]. Moreover, glutamine peptides remain inactive within the CGM protein molecule's sequence, further limiting the biological functions of Gln in CGM. Solid-state fermentation is an efficient and cost-effective method for altering the physical or physicochemical properties of feedstock, enhancing the bioavailability of agricultural by-products, and generating high-value-added products [13].
Both Bacillus subtilis and lactic acid bacteria are extensively employed as fermentation strains. Bacillus subtilis, known for its remarkable extracellular protease production, can break down large proteins into smaller proteins and peptides [14]. Lactobacillus, part of the lactic acid bacteria family, can reduce the fermentation system's pH and curb the growth of certain pathogenic bacteria through its organic acid production [15]. The incorporation of exogenous protease during fermentation enriches the protease system, boosting microorganisms' efficiency in utilizing macromolecular proteins in feed and thereby enhancing protein bioavailability [16]. To augment the bioavailability of CGM protein and liberate its glutamine peptides, this study implemented a co-fermentation protocol using Bacillus subtilis, Lactobacillus, and acid protease. The bioavailability of the fermented feed was then assessed through an in vivo broiler feeding experiment. Notably, existing literature offers no insights into the functionality of glutamine peptides derived from CGM.
2. Materials and Methods
2.1. Materials
CGM (crude protein, 65%) and corn steep liquor were sourced from Longjiang Fufeng Biotechnologies Co., Ltd. (Qiqihar, China). Wheat bran, SBM, and corn germ meal were obtained from a local market. Acid protease (50,000 U/g; pH 2.0-4.0) was procured from Doing-Higher Bio-Tech Co., Ltd. (Nanning, China). Bis-1,1-trifluoroacetoxy-iodobenzene (BTI) was purchased from Sigma-Aldrich Co. (St, Louis, MO, USA). MRS (de Man, Rogosa, and Sharpe) agar and LB (Luria-Bertani) agar were purchased from Hope Bio-Technology Co., Ltd. (Qingdao, China).
Bacillus subtilis A5 and Lactobacillus 02002 were selected based on their CGM degradation and acid production capabilities. Figure A1 shows the CGM degradation and acid production capabilities of Bacillus subtilis A5 and Lactobacillus 02002. All strains were preserved at the laboratory of the Department of Food and Biological Engineering at the University of Qiqihar.
2.2. Glutamine-Enriched CGM Fermented Feed Preparation
Initially, a blend was created using 40% CGM (extruded and milled), 10% SBM, 10% wheat bran, and 40% corn germ meal. Subsequently, a combined bacterial solution was prepared using Bacillus subtilis A5 (2×108 CFU/mL) and Lactobacillus 02002 (2×107 CFU/mL) in a 1:1 volume ratio. Sterile water, the mixed bacterial solution, acid protease liquid (1000 U/mL), and corn steep liquor were then added to the blend to produce a final fermentation mixture with 45% moisture content (v/w), a 3% bacterial inoculation concentration (v/w), 50 U/g acid protease amount and 10% corn steep liquor (v/w). Finally, the fermentation mixture was packaged in multi-layer, polythene bags equipped with a one-way valve and incubated at 30 °C for 72 h to obtain the glutamine-enriched CGM fermented feed (GCFF).
2.3. Nutritional Value Evaluation of the Glutamine-Enriched CGM Fermented Feed
The feed samples were dried at 65 °C for 48 h and ground using an extruder. The dry mater (method 930.15), crude protein (method 976.05), and crude fat (method 920.39) contents were subsequently analyzed, as described by the Association of Official Analytical Chemists (AOAC 1990) [17]. The crude fiber, acid detergent fiber and neutral detergent fiber contents were determined according to the standard of GB/T 6434-2022, NY/T 1459-2022 and GB/T 20806-2022, respectively. The pH value was assessed based on standard GB/T 10468-1989. Soluble protein content was evaluated using the Folin-phenol method, while small peptides were quantified via the trichloroacetic acid method outlined by Wang et al. [18]. Lactic acid content was measured with an ion chromatograph (Dionex ICS-1100, Thermol Fisher, USA). Probiotic populations were assessed using the plate counting method. Briefly, feed sample was serially diluted (1:10) to 10-7, and these dilutions were plated in triplicate onto LB agar for Bacillus subtilis, and MRS agar for Lactobacillus. CFU on the agars was counted and expressed as CFU per gram of wet feed.
2.4. Glutamine Content Evaluation
The Gln content in soluble protein was determined using the method described in our previous report [11]. The glutamine peptide content was characterized by assessing the no-N terminal Gln content, as N terminal Gln in an aqueous solution easily converts to pyroglutamic acid, losing the beneficial effects of Gln. Briefly, 50 g of dried and ground feed was added to 500 mL of solvent, then allowed to stand at room temperature for 2 h before being centrifuged at 10000 r/min for 30 minutes. The supernatant was filtered using filter paper and then freeze-dried to obtain the feed extract powder. Subsequently, 500 μL of feed extract (100 mg/mL), 2 mL BTI acetonitrile-aqueous solution (10 mg/mL), and 500 μL of pyridine (50 μmol/mL) were combined in an ampoule to create a reaction mixture. The Gln protect reaction of mixture was carried out at 50 °C for 2 h, a control mixture was prepared without the BTI protection reaction. The ampoules were then vacuum-sealed and subjected to an acid-hydrolyzed reaction (110 °C for 24 h) following the addition of 5 mL HCl (6 mol/L). After cooling, the glutamic acid content was measured using a glutamate biosensor analyzer (M-100, Sieman Technology Co., Ltd, Shenzhen, China), with the pH adjusted to neutral. The glutamine content was inferred from the difference in glutamic acid levels between the two samples.
2.5. Animal, Design, and Diets
The animal experiments were approved by the Animal Ethics Committee of the College of Food and Bioengineering, Qiqihar University. A total of 160 one-day-old broilers were procured from a local commercial hatchery and randomly allocated to four treatments. Each group consisted of five replicates with eight broilers each. The four experimental treatments were as follows: (1) corn-soybean meal basal diet (CON), (2) diet with 15% SBM replaced by GCFF (GCFF15), (3) diet with 30% SBM replaced by GCFF (GCFF30), (4) diet with 60% SBM replaced by GCFF (GCFF60). The ingredient and chemical composition of the basal diets are detailed in Table 1. The diets were formulated in accordance with the National Research Council (1994) to satisfy the nutritional requirements of the animals. All broilers had ad libitum access to feed and water throughout the 42-day experimental period.
2.6. Sample Collection
On day 21 and 42, broilers were humanly fasted for 12 h prior to sampling. Body weights were recorded to calculate average daily gain (ADG), and feed consumption was monitored to determine the average daily feed intake (ADFI), then the feed conversion rate (FCR) was obtained. At the study's conclusion, broilers were euthanized to collect blood samples, small intestinal tissues, and cecal contents for further analysis.
2.7. Intestinal Morphology Analysis
Jejunum and ileum tissues were preserved in 4% paraformaldehyde for H&E staining. Villus height (VH) and crypt depth (CD) were quantified using the Image-Pro Plus software 6.0 (Media Cybernetics, Inc., Washington, USA). The villus height to crypt depth ratio (VCR) was calculated. Additionally, the small intestine's length was measured.
2.8. Determination of Serum and Jejunum Tissues Immunological Parameters
Levels of IgG, IgA, and IgM in serum and jejunum tissues were quantified using chicken-specific ELISA kits (Jianglai Biotechnology Co., Ltd., Shanghai, China), adhering to the manufacturer’s protocols.
2.9. 16S rRNA High Throughput Sequencing
Total genomic DNA was extracted from cecal samples utilizing a DNA isolation kit (Jianglai Biotechnology Co., Ltd., Shanghai, China). Alterations in intestinal microbiota composition were examined through 16S rRNA sequencing analysis. Data were processed using the Majorbio Cloud Platform (Majorbio, Shanghai, China).
2.10. Statistical Analysis
Chemical compositions and animal experimental data were evaluated using Graph Pad Prism 8 (Graph Pad Software Inc., California, USA). Differences were deemed statistically significant at p < 0.05. All experiments were conducted in triplicate, results are represented as the mean ± standard error of the mean (SEM).
3. Results
3.1. Chemical Compositions of the GCFF
Table 2 presents the chemical composition of the GCFF. Post-fermentation, the contents of soluble protein and small peptides were increased by 69.1% and 47.6%, respectively, while the levels of crude fiber, crude fat, and acid detergent fiber significantly diminished (p < 0.05). The pH value decreased from 5.2 to 4.3, accompanied by a notable increase in lactic acid content by 125.9% (p < 0.05). Populations of Lactobacillus and Bacillus subtilis reached 2.6×106 and 1.1×105, respectively. The rise in soluble protein and small peptides resulted from the combined degradation activity of microbial protease and acid protease on macromolecule proteins. These low molecular weight proteins and peptides are more readily absorbed and digested by animals. The reduction in crude fiber, crude fat, and acid detergent fiber can be attributed to the degradation action of cellulase and lipase, produced by Bacillus subtilis A5. The decline in pH value was primarily due to the lactic acid generated by Lactobacillus 02002, which may also impart a distinctive sour aroma to the feed.
3.2. Glutamine Content of the GCFF
The BTI protection method was employed to ascertain the Gln content in the soluble protein. As depicted in Table 2, the Gln content within the soluble protein of the GCFF significantly rose from 1.6 mg/g to 12.8 mg/g (p < 0.05), evidencing the successful liberation of glutamine peptides from CGM protein.
3.3. Effects of Dietary GCFF Replacing SBM Supplementation on the Growth Performance of Broilers
Table 3 delineates the impact of GCFF substitution for SBM on broilers' growth performance. Broilers on diets with GCFF replacing 15% and 30% SBM registered higher ADG relative to the basal group throughout the entire period. Similarly, FCR values for broilers on the 15% and 30% GCFF diets were lower than those on the CON diet. These outcomes suggest that substituting GCFF for SBM may bolster growth performance, thereby potentially enhancing economic benefits.
3.4. Effects of Dietary GCFF Replacing SBM Supplementation on the Intestinal Morphology of Broilers
The integrity of the intestinal physical barrier is pivotal for animal health. Essential markers of intestinal structural integrity, such as intestinal length, VH, CD, and VCR, directly influence the absorptive efficiency of the intestinal mucosa. Figure 1 illustrates the effect of GCFF substitution for SBM on the morphological structure of broilers' small intestines. Diets with GCFF replacing SBM extended the relative length of the small intestinal (Figure 1a, b). In the jejunum, broilers on the 15% and 30% GCFF diets exhibited significantly higher VH and lower CD compared to those on the CON diet, with the 30% GCFF group displaying the highest VCR among all cohorts (Figure 1c-e). Regarding the ileum, the GCFF30 cohort demonstrated the highest VH and VCR and the lowest CD (Figure 1f-h). These findings suggest that appropriate GCFF substitution for SBM can promote intestinal structural maturation and enhance the physiological health of the intestine.
3.5. Effects of Dietary GCFF Replacing SBM Supplementation on the Serum and Jejunum Tissues Immunoglobulins of Broilers
Serving as the initial defense barrier against pathogenic intrusion, immunoglobulins are integral to the body's immune competence. Table 4 reveals the effects of GCFF substitution for SBM on serum and jejunum tissues immunoglobulins. Dietary substitution with GCFF led to a rise in serum IgG levels, though no significant alterations were observed in IgM and IgA concentrations. In the jejunal tissues, levels of IgG, IgM, and IgA significantly escalated following GCFF substitution for SBM (p < 0.05). These results denote an enhancement in the broilers' humoral immunity attributable to the GCFF substitution for SBM.
3.6. Effects of Dietary GCFF Replacing SBM Supplementation on the Intestinal Microbiota of Broilers
The gut microflora consists of billions of microorganisms, forming a mutually dependence and restriction dynamic balance within the intestinal micro-ecosystem. Healthy intestinal microflora benefits the host by modulating nutrients absorption and maintaining intestinal microecological stability. The cecal bacterial community in broilers was analyzed using 16S rRNA high-throughput sequencing, with results presented in Figure 2. The Sobs and Shannon index serve as indicators of alpha-diversity, describing microbial diversity within a specific region or ecosystem. A Venn diagram was constructed to identify specific and shared operational taxonomic units (OTUs) in broilers cecal samples under various diets. Principal coordinate analysis (PCoA) was employed to discern similarities or differences between groups. The study yielded a total of 723969 effective sequences post-quality control filtration, clustering into 614 OTUs at 97% sequencing identity. Both the sobs index (Figure 2a) and Shannon index (Figure 2b) were higher in the GCFF-replacing SBM groups compared to the CON group, suggesting that dietary GCFF supplementation in place of SBM could enhance the complexity of broilers' intestinal microflora. The GCFF-replacing SBM groups harbored more OTUs than the CON group (Figure 2c), indicating that GCFF supplementation in lieu of SBM could enrich the diversity of intestinal microbiota in broilers. Regarding the PCoA plot, the GCFF replacing SBM groups clustered closely together, separated distinctly and substantially from the CON group (Figure 2d), demonstrating that dietary GCFF replacing SBM supplementation altered the microbial composition.
Intestinal microbiota abundance at the phylum level is depicted in Figure 2e. Firmicutes, Bacteroidota, and Actinobacteria comprised over 98% of the four phyla in each treatment group. The relative abundance of Bacteroidota was greater in the CON group compared to the GCFF-replacing SBM groups, while Firmicutes were significantly less abundant in the CON group. At the genus level (Figure 2f), GCFF markedly affected broilers' intestinal microflora. The relative abundance of the Lactobacillus, Streptococcus, and Parabacteroides were significantly higher in the GCFF-replacing 15% and 30% SBM groups than in the CON group.
4. Discussion
Previous studies have affirmed that supplementation with Gln or glutamine peptides can enhance intestinal morphology, immune responses, intestinal microbiota, and, subsequently, the growth performance of animals. Table A1 shows partial corn protein sequences information in UniProt. Recognized as a superior source of glutamine-enriched protein. CGM undergoes extrusion, milling, and subsequent co-fermentation with Bacillus subtilis A5, Lactobacillus 02002, and acid protease to produce glutamine-enriched fermented feed. Our findings indicate that the co-fermentation process markedly elevates the concentration of glutamine peptides in the substrate. Firstly, during CGM pre-treatment, the extrusion process disrupts the protein bodies within CGM and unfolds the protein structures, enhancing protease accessibility for protein hydrolysis. These outcomes align with those of Zheng et al., who observed that Alcalase more readily hydrolyzed extruded CGM compared to its non-extruded counterpart [19]. Secondly, Bacillus subtilis A5, chosen for its potent CGM protein degradation capability, facilitates the conversion of parent proteins into active peptides, including glutamine peptides. It has also been reported that Bacillus pumilus secretes alkaline protease, demonstrating a robust capacity to hydrolyze zein [20]. Thirdly, acid protease pre-digests CGM protein and disrupts interactions between proteins and other constituents, thereby enhancing the efficacy of microbial protease [21]. Wang et al. reported that rapeseed meal fermented with the assistance of acid protease contained a higher peptide concentration than that fermented solely by microorganisms [3]. Finally, the acidic environment created by organic acids from Lactobacillus 02002 optimizes acid protease catalytic activity, leading to an increase in CGM protein degradation efficiency [22]. The augmented presence of glutamine peptides and other small peptides due to fermentation is likely to benefit both gut health and production performance in broilers.
The results of the current study reveal that dietary supplementation of GCFF, replacing 15% and 30% of SBM, enhanced the ADG and ADFI values in broilers. These findings align with Zhou's previous work, which noted improved daily weight gain in early-weaned calves receiving a diet supplemented with 1.01 g/kg alanyl-glutamine [23]. Similarity, Abdulkarimi et al. found that 0.5% Gln supplementation boosted both ADG and ADFI in broilers [24], while Wang et al. reported that a diet containing 10% fermented CGM increased the ADG in three yellow broilers [1]. Moreover, this study observed a reduction in FCR values when GCFF replaced 15% and 30% of SBM in the diet, suggesting that this substitution could reduce costs and enhance economic benefits. The augmented growth performance in broilers due to the inclusion of fermented diets may be attributable to the heightened nutritional bioavailability within the fermented feed, including a greater proportion of soluble protein and small peptides. These components are readily absorbed, digested, and ultimately deposited into muscle tissue. Additionally, the lactic acid imparted an acidic flavor to the feed, improving its palatability, which in turn likely increased broilers' feed intake and accelerated the rate of nutrient deposition, thereby promoting body weight gain. To elucidate the mechanisms underlying the growth-promoting effects of GCFF, this study further probed the intestinal responses of broilers to GCFF diets.
A robust intestinal structure is pivotal for the growth promotion in animals. An elongated intestinal length, elevated VH, and reduced CD suggest enhanced nutrient digestion and absorption within the small intestine. This research determined that diets with GCFF substituting for SBM augmented intestinal length and VH, diminished CD, and consequently, raised the VCR ratio (Figure 1). These outcomes are consistent with our findings, Gln supplementation improved relative weight and length of the intestine, as well as VH and VCR of the Nile tilapia juveniles [25]. Jing et al. reported that a CGM hydrolysate rich in glutamine peptides mitigated colonic shortening and decreased the permeability of the colonic mucosa in mice [11]. The beneficial effects of GCFF replacing SBM diets on gut morphology are presumably linked to the metabolic activities of Gln present in the fermented feed. Gln can supply energy and metabolic precursors for the proliferation and differentiation of intestinal mucosal cells, facilitate mucosal protein synthesis, regulate intestinal tight junction permeability, and thereby enhance intestinal structure and function comprehensively [26]. Indeed, glutamine peptides have been shown to more effectively stimulate enterocyte proliferation than the Gln monomer.
In harmony with the positive impact of GCFF on intestinal morphology, enhanced immunological function was noted in both the jejunum tissues and serum of broilers supplemented with GCFF replacing SBM. The bolstering of the immune system plays a crucial role in maintaining intestinal mucosal homeostasis and the host's physiological functions [27]. Gln has been documented to safeguard intestinal health and augment growth performance by modulating the immune response [28,29]. Corroborating our findings, Zhou et al. discovered that intravenously administered glutamine peptides elevated serum IgA, IgG, and intestinal mucosal sIgA, culminating in an advantageous impact on weight gain and intestinal structural integrity in early-weaned calves [23]. Similarly, Xu et al. found that dietary supplementation with glutamine peptides heightened the serum concentrations of IgA, IgG, and IgM, thereby enhancing intestinal barrier function more effectively [9]. The underlying mechanism bolstering immunity may be attributed to the Gln or glutamine peptides present in the fermented feed. Gln, recognized as a potent immune stimulant, serves as an essential energy source for innate immune cells, including lymphocytes and macrophages. The proliferation and activation of these immune cells lead to the secretion of various antibodies, thereby regulating the immune response and fostering a robust immune system.
Given the gut microbiota's pivotal role in sustaining intestinal health, orchestrating immune responses, and promoting growth, it is imperative to elucidate the effects of GCFF on broilers' microbiota composition. Currently, a greater diversity and richness in gut bacteria are considered advantageous, helping animals navigate various environmental challenges. Gln and glutamine peptides have been reported to preserve intestinal structure and function, in addition to enhancing growth performance, by modulating gut microbiota [30]. In this study, microbial diversity, richness, and community abundance experienced a significant upsurge when GCFF replaced 15% and 30% of SBM (Figure 2). This observation is consistent with findings that dietary glycyl-glutamine supplementation augmented the alpha-diversity of gut microbiota, thereby improving growth performance indices [31]. Comparable outcomes were documented with alanyl-glutamine peptide and Gln monomer supplementation [9]. Jing et al. also determined that CGM hydrolysate rich in glutamine peptides could regulate the intestinal microbiota's abundance and diversity, mitigate weight loss, and lower colonic mucosa permeability in mice [11]. Gln orchestrates the survival and proliferation of intestinal bacteria through the modulation of amino acid bacterial metabolism [32]. Known as a precursor in arginine biosynthesis, Gln has been shown to influence the metabolism of arginine family amino acids in the small intestine, potentially mediated in part by small-intestinal bacteria [33]. It is hypothesized that glutamine peptides might alter the bacterial utilization and metabolism of arginine-family amino acids within the intestine, thereby affecting bacterial activity and population dynamics, and prompting a shift in the community of commensal bacteria towards those more beneficial for host metabolism. Besides bioactive peptides, probiotics such as Lactobacillus and Bacillus subtilis might also contribute to the observed changes in broilers' intestinal microbial populations. These probiotics can thwart the colonization of harmful pathogens and promote the proliferation of beneficial anaerobic bacteria through the secretion of short-chain fatty acids and bacteriocins, ultimately enhancing mucosal morphology and the overall health of the animals [34,35]. In summary, these findings suggest that GCFF supplementation can positively alter intestinal microflora structure, although further investigation is warranted to ascertain the precise mechanism.
Gut commensal microbes are integral to maintaining the physiologic and metabolic homeostasis of the host, particularly in preserving intestinal health, owing to their efficiency in nutrient absorption and utilization [36]. In this study, at the phylum level, the Firmicutes to Bacteroidetes ratio escalated in tandem with an increasing proportion of GCFF replacing SBM (Figure 2). A higher Firmicutes to Bacteroidets ratio correlates with augmented energy harvest and enhanced growth performance. Prior research indicated that a diet deficient in protein and energy led to a reduction in Firmicutes and a surge in Bacteroidets [37]. Given the results of the current study, it can be inferred that following the introduction of fermented feed, modifications in intestinal microflora enhanced the efficiency of nutrient absorption and utilization, including proteins and peptides. At the genus level, populations of Lactobacillus, Streptococcus, and Parabacteroides expanded in correlation with the proportion of GCFF supplanting SBM. Lactobacillus possesses a robust ability to metabolize carbohydrates into organic acids, thereby inhibiting pathogen growth, modulating gut immune function, and promoting host health [37]. Bacterial species, including Lactobacillus, Streptococcus. and Parabacteroides, exhibit proficiency in protein degradation [38,39]. Streptococcus, a bacterium involved in amino acid fermentation, utilizes Gln as its preferred nitrogen source [40] and exhibits highly activity in dipeptidyl dipeptidase [41]. Our data suggest that an optimal ratio of GCFF replacing SBM in diets could significantly bolster the population of proteolytic bacteria, enhancing nutrient uptake efficiency, and ultimately improving the intestinal structure integrity and growth performance of broilers.
5. Conclusions
In conclusion, the findings of this study unequivocally show that co-fermentation can enhance the protein bioavailability of CGM, establishing CGM as a prime raw material for glutamine peptides preparation. GCFF emerges as a promising alternative protein source in feed. However, the study did not elucidate the specific mechanisms promoting broiler growth, and subsequent research will aim to uncover the health-promoting mechanisms of glutamine peptides. This research presents an innovative approach for creating a functional feed additive with growth-promoting properties and proposes a viable solution to mitigate the scarcity of feed protein resources.
Author Contributions
L.F.: experiments, data analysis and writing—original draft; X.L.: writing—reviewing and editing; Y.D.: supervision; X.Z.: experimental design and funding acquisition. All authors have read and agreed to the published version of the manuscript..
Funding
This research was funded by the National Key Research and Development Plan of China (No. 2021YFD2100900), Double First-Class Project for Collaborative Innovation Achievements in Disciplines Construction in Heilongjiang Province (No. LJGXCG2022-122 and No. LJGXCG2023-055), Talent Foundation of the Central Government Supporting Local Universities (No. 2020GSP08), Heilongjiang Province Key Research and Development Projects (No. 2020ZX06B01) and Theory and Technology of Corn Deep Processing in Heilongjiang Province (No. SPKF202015).
Data Availability Statement
All relevant data are reported in the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ADFI | Average daily feed intake |
| ADG | Average daily gain |
| BTI | Bis-1,1-trifluoroacetoxy-iodobenzene |
| CD | Crypt depth |
| CGM | Corn gluten meal |
| FCR | Feed conversion rate |
| GCFF | Glutamine-enriched corn gluten meal fermented feed |
| Gln | Glutamine |
| OUT | Operational taxonomic units |
| PCoA | Principal coordinate component analysis |
| SBM | Soybean meal |
| UFF | Corn gluten meal unfermented feed |
| VCR | Villus height to crypt depth |
Appendix A
Figure A1.
CGM degradation and acid production capabilities of Bacillus subtilis A5 and Lactobacillus 02002. The hydrolysis circle of Bacillus subtilis A5 on the front (a) and back(b) of corn gluten meal plates; the hydrolysis circle of Lactobacillus 02002 on the front (c) and back (d) of MRS-CaCO3 plates.
Figure A1.
CGM degradation and acid production capabilities of Bacillus subtilis A5 and Lactobacillus 02002. The hydrolysis circle of Bacillus subtilis A5 on the front (a) and back(b) of corn gluten meal plates; the hydrolysis circle of Lactobacillus 02002 on the front (c) and back (d) of MRS-CaCO3 plates.
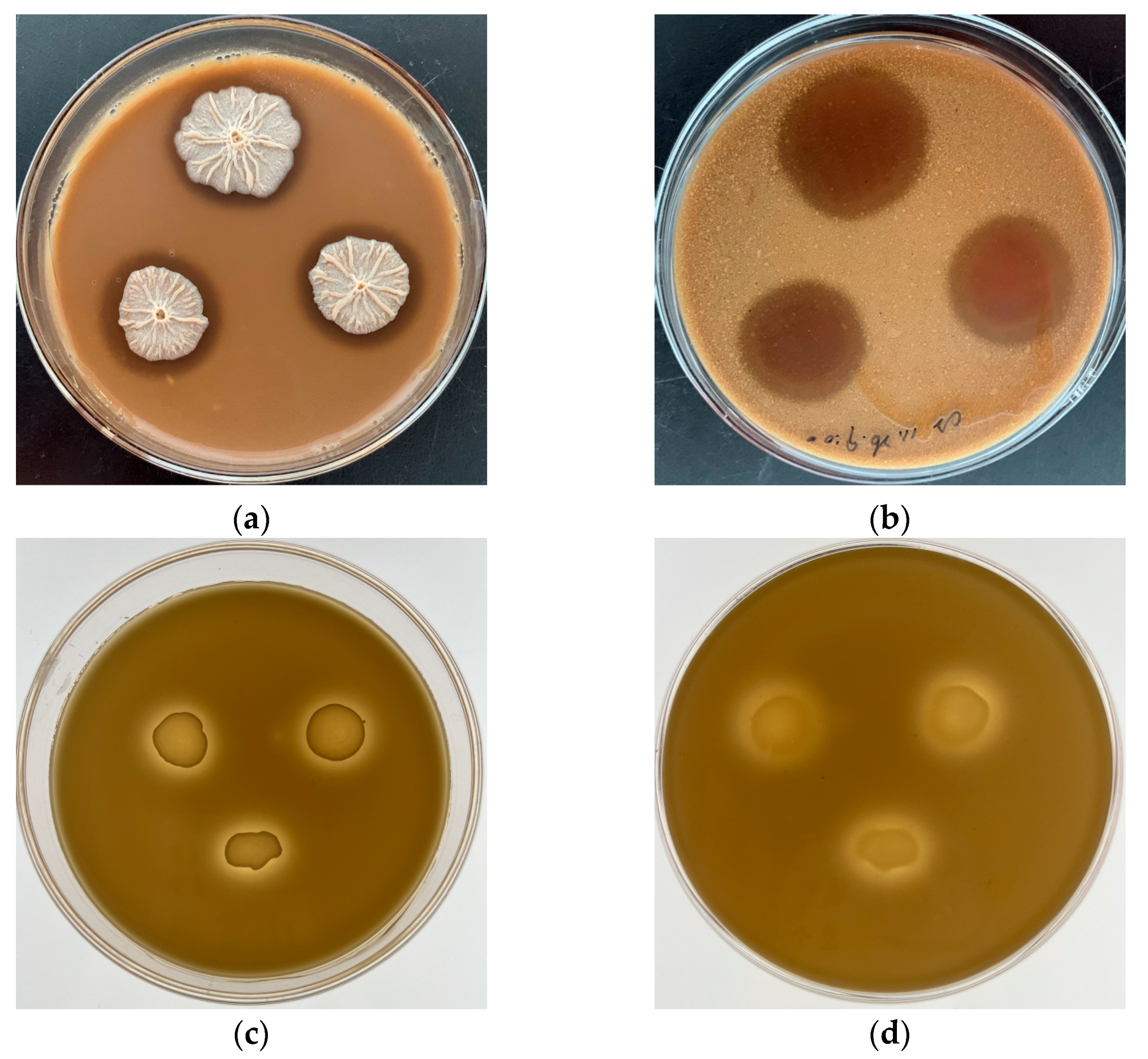

Table A1.
Partial corn protein sequence information in Uniprot.
| No. | Sequence | Position in protein | Protein name | Access No. |
|---|---|---|---|---|
| 1 | QPPRPQP | 88-94 | Glutein-2 | P04706 |
| 2 | LRQQCCQQL | 151-159 | Glutein-2 | P04706 |
| 3 | LQQQPQSGQVA | 180-190 | Glutein-2 | P04706 |
| 4 | AAQIAQQL | 194-201 | Glutein-2 | P04706 |
| 5 | LQQP | 207-210 | Glutein-2 | P04706 |
| 6 | LRQQCCQQQM | 83-92 | Zein-beta | P06673 |
| 7 | AQQLQMMMQL | 101-110 | Zein-beta | P06673 |
| 8 | ALMQQQQQLL | 124-133 | Zein-beta | P06673 |
| 9 | LQQA | 58-61 | Zein-alpha A20 | P04703 |
| 10 | LFQQS | 73-77 | Zein-alpha A20 | P04703 |
| 11 | AQQLQQL | 92-98 | Zein-alpha A20 | P04703 |
| 12 | QQQQFL | 116-121 | Zein-alpha A20 | P04703 |
| 13 | LQQQL | 135-139 | Zein-alpha A20 | P04703 |
| 14 | QQQQLL | 151-156 | Zein-alpha A20 | P04703 |
| 15 | LQQQIL | 170-175 | Zein-alpha A20 | P04703 |
| 16 | LQLQQL | 210-215 | Zein-alpha A20 | P04703 |
Note: Q, glutamine; P, proline; R, arginine; C, cysteine; L, lysine; S, serine; G, glycine; V, valine; A, alanine; I, isoleucine; M, methionine; F, phenylalanine.
References
- Wang, Y.; Liu, X.L.; Jin, L.; Wen, Q.N.; Zhang, Y.G.; Narasimha, K.; Yan, E.Y.; Wang, C.P.; Zheng, Y.B. Effects of Fermented Corn Gluten Meal on Growth Performance, Serum Parameters, Intestinal Morphology, and Immunity Performance of Three-Yellow Broilers. Canadian Journal of Animal Science 2019, 99, 408–417. [Google Scholar] [CrossRef]
- Olukomaiya, O.; Fernando, C.; Mereddy, R.; Li, X.H.; Sultanbawa, Y. Solid-State Fermented Plant Protein Sources in the Diets of Broiler Chickens: A Review. Animal Nutrition 2019, 5, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, H.; Liu, X. A Novel Fermented Rapeseed Meal, Inoculated with Selected Protease-Assisting Screened B. Subtilis Yy-4 and L. Plantarum 6026, Showed High Availability and Strong Antioxidant and Immunomodulation Potential Capacity. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, J.; Soladoye, O.P.; Aluko, R.E.; Fu, Y.; Zhang, Y. Preparation, Receptors, Bioactivity and Bioavailability of Γ-Glutamyl Peptides: A Comprehensive Review. Trends in Food Science & Technology 2021, 113, 301–314. [Google Scholar]
- Satoh, J.; Tsujikawa, T.; Fujiyama, Y.; Bamba, T. Nutritional Benefits of Enteral Alanyl-Glutamine Supplementation on Rat Small Intestinal Damage Induced by Cyclophosphamide. Journal of Gastroenterology and Hepatology 2003, 18, 719–725. [Google Scholar] [CrossRef]
- Tan, B.; Liu, H.L.; He, G.Z.; Xiao, H.; Xiao, D.F.; Liu, Y.H.; Wu, J.P.; Fang, J.; Yin, Y.L. Alanyl-Glutamine but Not Glycyl-Glutamine Improved the Proliferation of Enterocytes as Glutamine Substitution in Vitro. Amino Acids 2017, 49, 2023–2031. [Google Scholar] [CrossRef]
- Zou, T.D.; Deng, C.X.; Wang, Z.R.; Ye, Y.L.; You, J.M. Dietary Alanyl-Glutamine Improves Growth Performance of Weaned Piglets through Maintaining Intestinal Morphology and Digestion-Absorption Function. Animal 2019, 13, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; He, G.; Mai, K.S.; Liu, C.D.; Zhou, H.H.; Wen, H. Dietary Ala-Gln Ameliorated Growth Suppression and Intestinal Injury Induced by Soya Saponin in Zebrafish. Aquaculture 2020, 529. [Google Scholar] [CrossRef]
- Xu, Q.B.; Hu, M.Y.; Li, M.; Hou, J.X.; Zhang, X.H.; Gao, Y.; Chachar, B.; Li, X. Dietary Bioactive Peptide Alanyl-Glutamine Attenuates Dextran Sodium Sulfate-Induced Colitis by Modulating Gut Microbiota. Oxidative Medicine and Cellular Longevity 2021, 2021, 1–17. [Google Scholar] [CrossRef]
- Zheng, X.Q.; Wang, J.T.; Liu, X.L.; Sun, Y.; Zheng, Y.J.; Wang, X.J.; Liu, Y. Effect of Hydrolysis Time on the Physicochemical and Functional Properties of Corn Glutelin by Protamex Hydrolysis. Food Chem 2015, 172, 407–415. [Google Scholar] [CrossRef]
- Jing, Y.; Liu, X.L.; Wang, J.Y.; Ma, Y.Q.; Zheng, X.Q. Production of Corn Protein Hydrolysate with Glutamine-Rich Peptides and Its Antagonistic Function in Ulcerative Colitis in Vivo. Foods 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Zheng, X.Q.; Kopparapu, N.K.; Cong, W.S.; Deng, Y.P.; Sun, X.J.; Liu, X.L. Purification and Evaluation of a Novel Antioxidant Peptide from Corn Protein Hydrolysate. Process Biochemistry 2014, 49, 1562–1569. [Google Scholar] [CrossRef]
- Su, W.F.; Jiang, Z.P.; Wang, C.; Xu, B.C.; Lu, Z.Q.; Wang, F.Q.; Zong, X.; Jin, M.L.; Wang, Y.Z. Dynamics of Defatted Rice Bran in Physicochemical Characteristics, Microbiota and Metabolic Functions During Two-Stage Co-Fermentation. International Journal of Food Microbiology 2022, 362, 109489. [Google Scholar] [CrossRef]
- Razzaq, A.; Shamsi, S.; Ali, A.; Ali, Q.; Sajjad, M.; Malik, A.; Ashraf, M. Microbial Proteases Applications. Frontiers in Bioengineering and Biotechnology 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, A.; Settanni, L. Lactobacilli in Sourdough Fermentation. Food Research International 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Li, W.; Cheng, P.; Zhang, J.B.; Zhao, L.M.; Ma, Y.B.; Ding, K. Synergism of Microorganisms and Enzymes in Solid-State Fermentation of Animal Feed. A Review. Journal of Animal and Feed Sciences 2021, 30, 3–10. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Wang, Y.C.; Xu, K.K.; Lu, F.; Wang, Y.N.; Ouyang, N.N.; Ma, H.L. Increasing Peptide Yield of Soybean Meal Solid-State Fermentation of Ultrasound-Treated Bacillus Amyloliquefaciens. Innovative Food Science & Emerging Technologies 2021, 72. [Google Scholar]
- Zheng, X.Q.; Li, L.T.; Liu, X.L.; Wang, X.J.; Lin, J.; Li, D. Production of Hydrolysate with Antioxidative Activity by Enzymatic Hydrolysis of Extruded Corn Gluten. Appl Microbiol Biotechnol 2006, 73, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Miyaji, T.; Otta, Y.; Nakagawa, T.; Watanabe, T.; Niimura, Y.; Tomizuka, N. Purification and Molecular Characterization of Subtilisin-Like Alkaline Protease Bpp-a from Bacillus Pumilus Strain Ms-1. Letters in Applied Microbiology 2006, 42, 242–247. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, X.; Xu, H.J.; Sun, Y.K.; Zhang, Y.G.; Wang, Y. Improvement of the Nutritional, Antioxidant and Bioavailability Properties of Corn Gluten-Wheat Bran Mixture Fermented with Lactic Acid Bacteria and Acid Protease. Lwt-Food Science and Technology 2021, 144, 111–116. [Google Scholar] [CrossRef]
- Kocabiyik, S.; Ozel, H. An Extracellular—Pepstatin Insensitive Acid Protease Produced by Thermoplasma Volcanium. Bioresource Technology 2007, 98, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Zhang, P.S.; Deng, G.C.; Liu, X.M.; Lu, D.X. Improvements of Immune Status, Intestinal Integrity and Gain Performance in the Early-Weaned Calves Parenterally Supplemented with L-Alanyl-L-Glutamine Dipeptide. Veterinary Immunology and Immunopathology 2012, 145, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Abdulkarimi, R.; Shahir, M.H.; Daneshyar, M. Effects of Dietary Glutamine and Arginine Supplementation on Performance, Intestinal Morphology and Ascites Mortality in Broiler Chickens Reared under Cold Environment. Asian-Australas J Anim Sci 2019, 32, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.L.P.F.; Xavier, W.d.S.; Guimarães, M.G.; Rodrigues, E.J.D.; Furuya, W.M.; Yamamoto, F.Y.; Pezzato, L.E.; Gatlin, D.M.; Barros, M.M. Dietary Glutamine Improves Growth and Intestinal Morphology of Juvenile Gift Tilapia (Oreochromis Niloticus) but Has Limited Effects on Innate Immunity and Antioxidant Capacity. Aquaculture 2023, 563. [Google Scholar] [CrossRef]
- Roth, E. Nonnutritive Effects of Glutamine. Journal of Nutrition 2008, 138, 2025s–2031s. [Google Scholar] [CrossRef] [PubMed]
- Mowat, A.M.; Agace, W.W. Regional Specialization within the Intestinal Immune System. Nature Reviews Immunology 2014, 14, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Hao, L.; Chandra, P.; Jones, D.P.; Willams, I.R.; Gewirtz, A.T.; Ziegler, T.R. Dietary Glutamine and Oral Antibiotics Each Improve Indexes of Gut Barrier Function in Rat Short Bowel Syndrome. American Journal of Physiology-Gastrointestinal and Liver Physiology 2009, 296, G348–G355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, L.; Zhou, Y.; Diao, C.; Han, L.; Yinjie, N.; Liu, S.; Chen, H. Glutamine Ameliorates Mucosal Damage Caused by Immune Responses to Duck Plague Virus. Dose-Response 2017, 15. [Google Scholar] [CrossRef]
- Liu, J.; Zong, C.G.; Yu, X.; Ding, Y.; Chang, B.; Wang, R.Y.; Sang, L.X. Alanyl-Glutamine (Ala-Gln) Ameliorates Dextran Sulfate Sodium (DSS)-Induced Acute Colitis by Regulating the Gut Microbiota, Pi3k-Akt/NF-κB/STAT3 Signaling, and Associated Pulmonary Injury. Acs Infectious Diseases 2023, 979–992. [Google Scholar] [CrossRef]
- Yan, Y.Q.; Xu, B.Y.; Yin, B.Q.; Xu, X.F.; Niu, Y.R.; Tang, Y.M.; Wang, X.K.; Xie, C.L.; Yang, T.; Zhou, S.Y. , et al. Modulation of Gut Microbial Community and Metabolism by Dietary Glycyl-Glutamine Supplementation May Favor Weaning Transition in Piglets. Frontiers in Microbiology 2020, 10, 3125. [Google Scholar] [CrossRef]
- Forchhammer, K. Glutamine Signalling in Bacteria. Frontiers in Bioscience-Landmark 2007, 12, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.L.; Li, X.L.; Xi, P.B.; Zhang, J.; Wu, G.Y.; Zhu, W.Y. L-Glutamine Regulates Amino Acid Utilization by Intestinal Bacteria. Amino Acids 2013, 45, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.; Kim, J.W.; Nyachoti, C.M. Nutrient and Energy Digestibility, and Microbial Metabolites in Weaned Pigs Fed Diets Containing Lactobacillus –Fermented Wheat. Animal Feed Science and Technology 2018, 241, 27–37. [Google Scholar] [CrossRef]
- Canon, F.; Nidelet, T.; Guedon, E.; Thierry, A.; Gagnaire, V. Understanding the Mechanisms of Positive Microbial Interactions That Benefit Lactic Acid Bacteria Co-Cultures. Frontiers in Microbiology 2020, 11, 2088. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Lee, T.T. Effects of Laetiporus Sulphureus-Fermented Wheat Bran on Growth Performance, Intestinal Microbiota and Digesta Characteristics in Broiler Chickens. Animals 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.; Singh, T.P.; Reddi, S.; Malik, R.K.; Kapila, S. Intervention of Probiotic L-Reuteri Fermented Milk as an Adjuvant to Combat Protein Energy Malnourishment Induced Gut Disturbances in Albino Mice. Journal of Functional Foods 2017, 36, 467–479. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Cummings, J.H.; Allison, C. Protein-Degradation by Human Intestinal Bacteria. Journal of General Microbiology 1986, 132, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Korpela, K. Diet, Microbiota, and Metabolic Health: Trade-Off between Saccharolytic and Proteolytic Fermentation. Annual Review of Food Science and Technology, Vol 9 2018, 9, 65–84. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Awati, A.A.; Williams, B.A.; Miller, B.G.; Jones, P.; Stokes, C.R.; Akkermans, A.D.L.; Smidt, H.; De Vos, W.M. Post-Natal Development of the Porcine Microbiota Composition and Activities. Environmental Microbiology 2006, 8, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.L.; Wu, G.Y.; Zhu, W.Y. Amino Acid Metabolism in Intestinal Bacteria: Links between Gut Ecology and Host Health. Frontiers in Bioscience-Landmark 2011, 16, 1768–1786. [Google Scholar] [CrossRef]
Figure 1.
Intestinal morphology of broilers fed experimental diets with or without GCFF replacing SBM. (a) relative length of small intestinal; (b) jejunum length at days 42; (c) jejunum villus height; (d) jejunum crypt depth; (e) jejunum villus height to crypt depth; (f) ileum villus height; (g) ileum crypt depth; (h) ileum villus height to crypt depth. CON, corn and soybean meal basal diet; GCFF, glutamine-enriched corn gluten meal fermented feed; GCFF15, diet substituted 15% soybean meal with GCFF; GCFF30, diet substituted 30% soybean meal with GCFF; GCFF60, diet substituted 60% soybean meal with GCFF. a-c, different superscript letters indicate significant difference between groups (p < 0.05).
Figure 1.
Intestinal morphology of broilers fed experimental diets with or without GCFF replacing SBM. (a) relative length of small intestinal; (b) jejunum length at days 42; (c) jejunum villus height; (d) jejunum crypt depth; (e) jejunum villus height to crypt depth; (f) ileum villus height; (g) ileum crypt depth; (h) ileum villus height to crypt depth. CON, corn and soybean meal basal diet; GCFF, glutamine-enriched corn gluten meal fermented feed; GCFF15, diet substituted 15% soybean meal with GCFF; GCFF30, diet substituted 30% soybean meal with GCFF; GCFF60, diet substituted 60% soybean meal with GCFF. a-c, different superscript letters indicate significant difference between groups (p < 0.05).

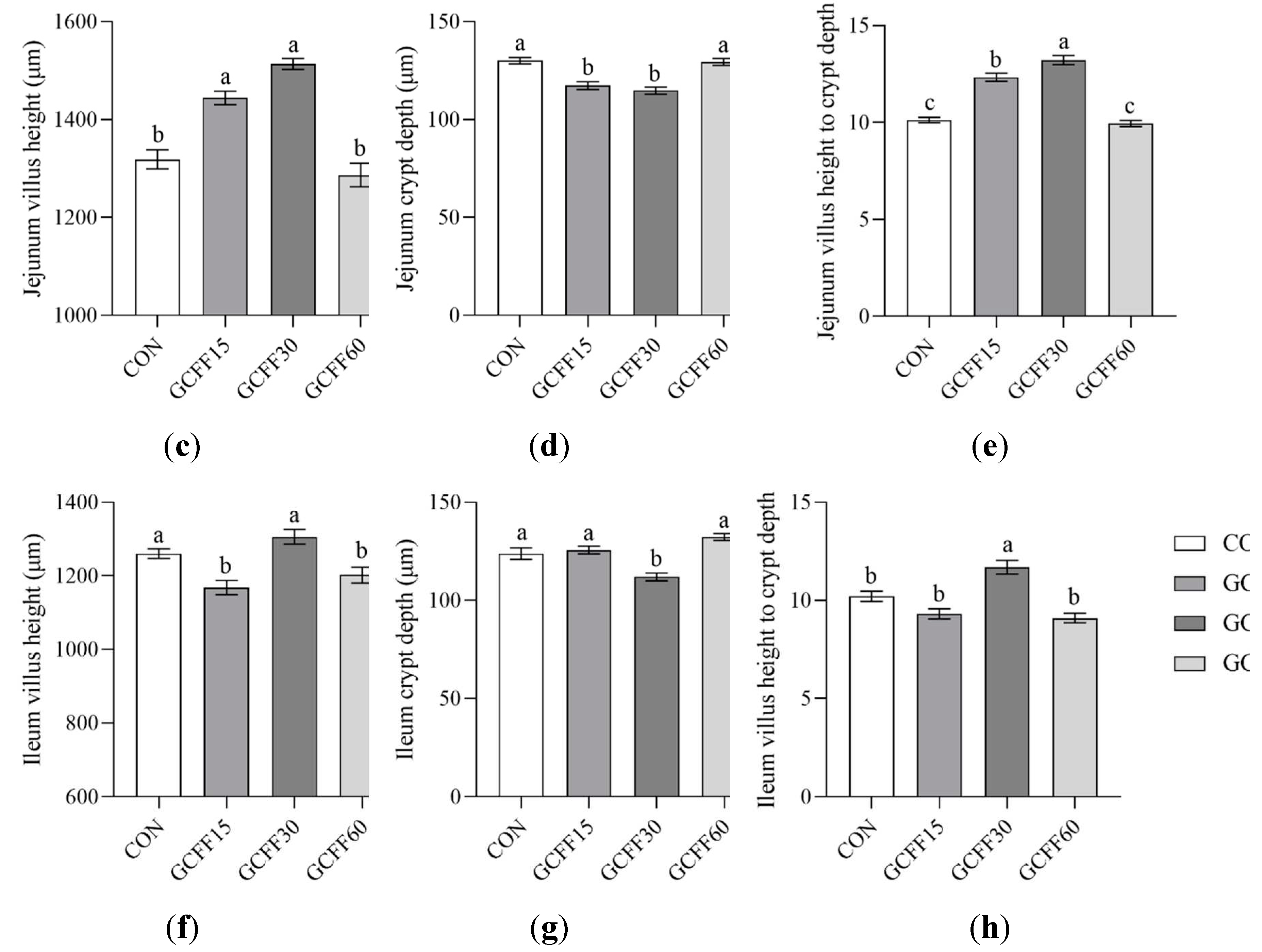
Figure 2.
Intestinal microbiota of broilers fed experimental diets with or without GCFF replacing SBM. (a) rarefaction curves; (b) Shannon curves; (c) Venn diagrams of OTU; (d) principal coordinate analysis (PCoA); (e) microbiota analysis at the phylum level; (f) microbiota analysis at the genus level. CON, corn and soybean meal basal diet. GCFF, glutamine-enriched corn gluten meal fermented feed; GCFF15, diet substituted 15% soybean meal with GCFF; GCFF30, diet substituted 30% soybean meal with GCFF; GCFF60, diet substituted 60% soybean meal with GCFF.
Figure 2.
Intestinal microbiota of broilers fed experimental diets with or without GCFF replacing SBM. (a) rarefaction curves; (b) Shannon curves; (c) Venn diagrams of OTU; (d) principal coordinate analysis (PCoA); (e) microbiota analysis at the phylum level; (f) microbiota analysis at the genus level. CON, corn and soybean meal basal diet. GCFF, glutamine-enriched corn gluten meal fermented feed; GCFF15, diet substituted 15% soybean meal with GCFF; GCFF30, diet substituted 30% soybean meal with GCFF; GCFF60, diet substituted 60% soybean meal with GCFF.
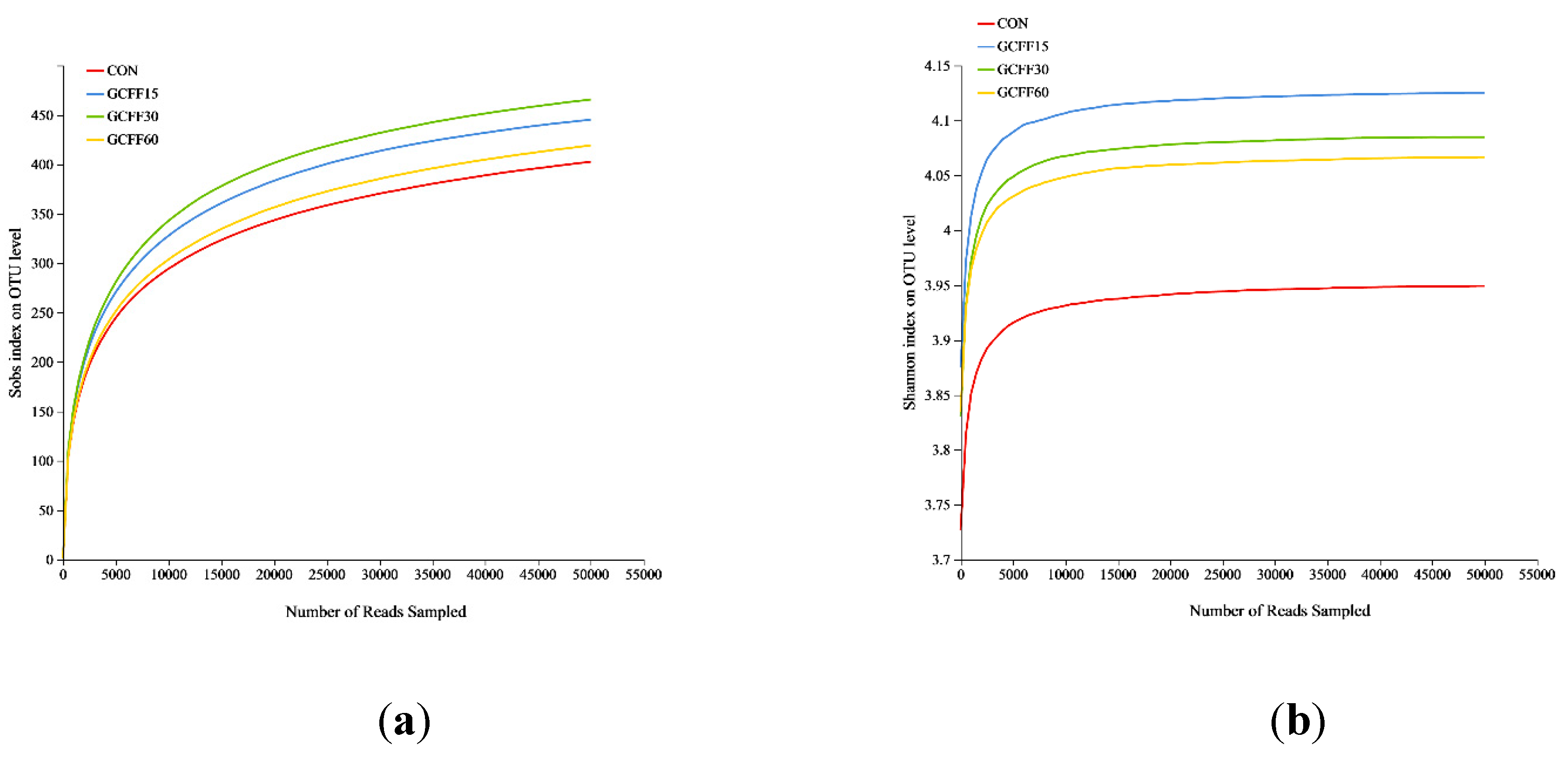
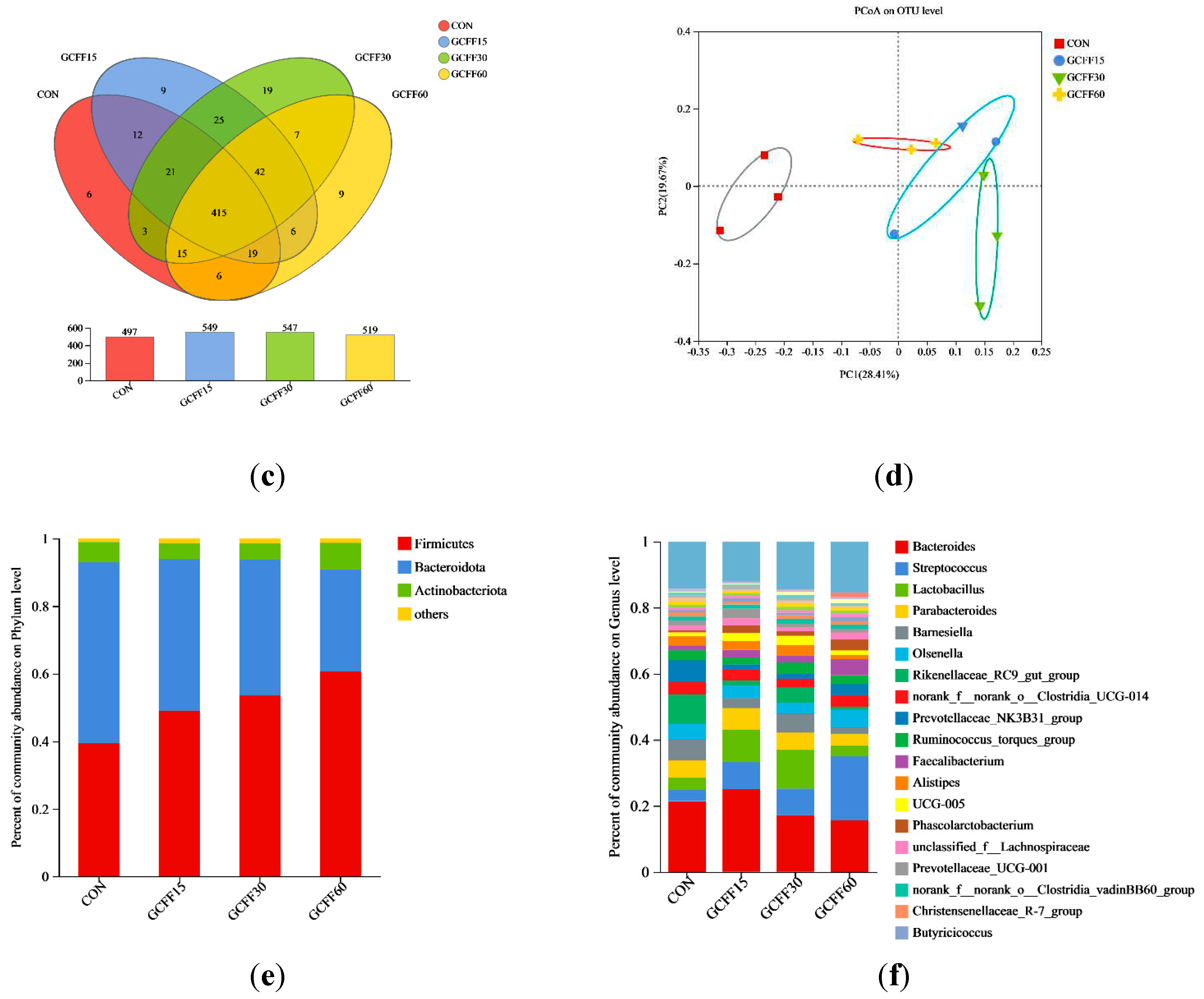
Table 1.
Composition of experimental diets.
| Item | 1-21 d | 22-42 d | ||||||
|---|---|---|---|---|---|---|---|---|
| CON | GCFF15 | GCFF30 | GCFF60 | CON | GCFF15 | GCFF30 | GCFF60 | |
| Ingredients (%, as-fed basis) | ||||||||
| Corn | 57.47 | 59.77 | 57.47 | 57.97 | 62.00 | 63.40 | 62.00 | 62.60 |
| Soybean meal | 35.80 | 29.63 | 25.06 | 14.32 | 31.07 | 26.41 | 21.75 | 12.43 |
| CGM fermented feed | 0.00 | 5.37 | 10.74 | 21.48 | 0.00 | 4.66 | 9.32 | 18.64 |
| Soybean oil | 3.00 | 1.50 | 3.00 | 2.50 | 3.60 | 2.20 | 3.60 | 3.00 |
| Limestone | 1.27 | 1.27 | 1.27 | 1.27 | 1.21 | 1.21 | 1.21 | 1.21 |
| Dicalcium phosphate | 1.45 | 1.45 | 1.45 | 1.45 | 1.20 | 1.20 | 1.20 | 1.20 |
| Choline chloride | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 | 0.20 |
| NaCl | 0.35 | 0.35 | 0.35 | 0.35 | 0.35 | 0.35 | 0.35 | 0.35 |
| Premix 1 | 0.22 | 0.22 | 0.22 | 0.22 | 0.22 | 0.22 | 0.22 | 0.22 |
| DL-Methionine (98%) | 0.20 | 0.20 | 0.20 | 0.20 | 0.12 | 0.12 | 0.12 | 0.12 |
| L-Lysine-HCl (99%) | 0.02 | 0.02 | 0.02 | 0.02 | 0.01 | 0.01 | 0.01 | 0.01 |
| Antioxidant | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 | 0.02 |
Note: 1 The premix provided the following per kilogram of diet: Vitamin A, 9500U; Vitamin D3, 0.0625 mg; Vitamin K3, 2.65 mg; Vitamin B1, 0.025 mg; Vitamin B2, 6 mg; Vitamin E, 30 U; biotin, 0.0325 mg; folic acid, 1.25 mg; pantothenic acid, 12 mg; niacin, 50 mg; Cu, 8 mg; Zn, 75mg; Fe, 80 mg; Mn, 100 mg; Se, 0.15 mg; I, 0.35 mg. CON, corn and soybean meal basal diet; GCFF, glutamine-enriched corn gluten meal fermented feed; GCFF15, diet substituted 15% soybean meal with GCFF; GCFF30, diet substituted 30% soybean meal with GCFF; GCFF60, diet substituted 60% soybean meal with GCFF.
Table 2.
The chemical analysis of the UFF and GCFF.
| Item | UFF | GCFF |
|---|---|---|
| Dry matter (%) | 90.52 ± 0.19 | 90.13 ± 0.28 |
| Crude protein (%) | 40.46 ± 0.28 | 41.33 ± 0.46 |
| Crude fiber (%) | 12.98 ± 0.09 a | 9.10 ± 0.04 b |
| Crude fat (%) | 2.50 ± 0.11 a | 2.17 ± 0.04 b |
| Acid detergent fiber (%) | 28.36 ± 0.18 a | 23.53 ± 0.25 b |
| Neutral detergent fiber (%) | 8.43 ± 0.12 | 8.22 ± 0.09 |
| pH | 5.2 a | 4.6 b |
| Glutamine content (mg/g soluble protein) | 1.60 ± 0.16 b | 12.80 ± 0.14 a |
| Small peptide content (mg/g) | 76.10 ± 0.96 b | 112.30 ± 1.05 a |
| Soluble protein content (mg/g) | 82.40 ± 0.82 b | 140.70 ± 1.98 a |
| Lactic acid content (mg/g) | 13.18 ± 0.03 b | 29.78 ± 0.19 a |
| Lactobacillus (CFU/g) | Undetected | 2.6 × 106 |
| Bacillus subtilis (CFU/g) | Undetected | 1.1 × 105 |
Note: UFF, corn gluten meal unfermented feed; GCFF, glutamine-enriched corn gluten meal fermented feed. a, b, different lowercase superscripts indicate significant differences (p < 0.05) in the same row.
Table 3.
Growth performance of broilers fed experimental diets with or without GCFF replacing SBM.
| Item | Time | CON | GCFF15 | GCFF30 | GCFF60 | SEM | p-value |
|---|---|---|---|---|---|---|---|
| ADG (g) | 1-21 d | 34.0 c | 37.4 a | 35.8 b | 33.4 c | 0.919 | <0.001 |
| 22-42 d | 82.9 c | 92.0 a | 88.3 b | 76.0 d | 3.491 | <0.001 | |
| 1-42 d | 60.7 c | 66.3 a | 62.2 b | 54.6 d | 2.413 | <0.001 | |
| ADFI (g) | 1-21 d | 53.3 a | 54.5 a | 54.9 a | 52.3 b | 0.599 | 0.063 |
| 22-42 d | 172.7 a | 173.2 a | 173.5 a | 160.2 b | 3.237 | <0.001 | |
| 1-42 d | 113.0 a | 113.8 a | 114.2 a | 106.2 b | 1.877 | <0.001 | |
| FCR | 1-21 d | 1.56 | 1.52 | 1.52 | 1.60 | 0.019 | 0.156 |
| 22-42 d | 1.98 b | 1.82 c | 1.98 b | 2.08 a | 0.054 | <0.001 | |
| 1-42 d | 1.86 b | 1.74 c | 1.84 b | 1.94 a | 0.042 | <0.001 |
Note: CON, corn and soybean meal basal diet; GCFF, glutamine-enriched corn gluten meal fermented feed; GCFF15, diet substituted 15% soybean meal with GCFF; GCFF30, diet substituted 30% soybean meal with GCFF; GCFF60, diet substituted 60% soybean meal with GCFF. ADG, average daily gain; ADFI, average daily feed intake; FCR, feed conversion rate. a-d, different lowercase superscripts indicate significant differences (p < 0.05) in the same row.
Table 4.
Immunological parameters of broilers fed experimental diets with or without GCFF replacing SBM.
Table 4.
Immunological parameters of broilers fed experimental diets with or without GCFF replacing SBM.
| Item | CON | GCFF15 | GCFF30 | GCFF60 | SEM | p-value |
|---|---|---|---|---|---|---|
| Serum | ||||||
| IgM (μg/mL) | 624.9 | 632.2 | 624.9 | 605.1 | 5.039 | 0.414 |
| IgG (μg/mL) | 1793.9 b | 2135.9 a | 2039.5 a | 2056.6 a | 63.997 | 0.006 |
| IgA (μg/mL) | 215.0 | 232.9 | 207.4 | 205.8 | 5.376 | 0.096 |
| Jejunum tissues | ||||||
| IgM (μg/mL) | 548.6 b | 603.8 a | 605.9 a | 566.2 b | 12.269 | < 0.001 |
| IgG (μg/mL) | 1609.9 b | 1793.5 a | 1850.6 a | 1798.3 a | 45.617 | < 0.001 |
| IgA (μg/mL) | 210.4 b | 238.0 a | 226.9 a | 220.9 ab | 4.994 | 0.005 |
Note: CON, corn and soybean meal basal diet; GCFF, glutamine-enriched corn gluten meal fermented feed; GCFF15, diet substituted 15% soybean meal with GCFF; GCFF30, diet substituted 30% soybean meal with GCFF; GCFF60, diet substituted 60% soybean meal with GCFF. IgM, immunoglobulin M; IgG, immunoglobulin G; IgA, immunoglobulin A. a, b, values in the same row with different superscript letters were significantly different (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



