
Submitted:
31 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
Healthcare faces a major problem with the increased emergence of antimicrobial resistance due to the over-prescription of antibiotics. This causes dysbiosis and major disruption of the gut microbiome, leading to the development of intestinal diseases, abnormalities in the regulation of immune responses, malfunctioning of entero-endocrine signaling, and an imbalance in communication with the central nervous system (CNS). Bacteriophages may provide a solution to the treatment of bacterial infections given their specificity. This approach increased rapidly over the last ten years, because of the rapid rise of multi-drug-resistant bacteria worldwide coupled with a decline in the development and production of novel antibacterial agents. Eradication of multidrug-resistant bacteria is often only possible with bacteriophage treatment. Phage proteins are used to stimulate immune responses against specific pathogens, improve antibiotic susceptibility, prevent biofilm formation, and characterize bacterial pathogens. This review discusses the therapeutic potential of bacteriophage-derived proteins.
Keywords:
Bacteriophage-derived proteins
; therapeutic potential
1. Introduction
Per annum, approximately 700,000 deaths related to multidrug-resistant infections are reported worldwide, which may increase to 10 million by 2050 [1]. Treatment with bacteriophages (phages) differs from treatment with antibiotics in that phages are mostly strain-specific [2]. Phage therapy is, however, not an alternative to treatment with antibiotics. Bacteria do have the ability to develop resistance to phage attacks. This is achieved through the modification (or loss) of phage receptors or the secretion of molecules that block adhesion sites [3,4]. Bordetella bronchiseptica uses reverse transcriptase to protect cells against phage infection [5].
The biggest advantage of using phages in treating infections lies in their ability to produce enzymes that interact with bacterial surfaces or destroy biofilms. These enzymes also play an important role in the entry and exit of phages from susceptible hosts [6,7,8]. Endogenous lysins are produced during the final phase of bacterial infection. The proteins holin and spannin, encoded by genes in the lysin operon, assist with the translocation of lysin across the cell membrane to the peptidoglycan layer. Lysins are grouped into two main categories, i.e., endo-β-N-acetylglucosaminidases or N-acetylmuramidases (lysozyme). Endopeptidases degrade protein moieties and amidases (e.g., N-acetylmuramoyl-L-alanine amidase) hydrolyze amide bonds between glycans and peptides [6]. Exolysins, classified as hydrolases or lyases, on phage tail fibers, tail spike proteins (TSPs), and phage baseplates cleave exopolysaccharides (EPS) on the surface of bacterial cells to facilitate adsorption and degrade capsule polysaccharides (CPS) [9]. Some phages produce endosialidases at their tail structures to degrade polysaccharide barriers. Phage polysaccharide lyases cleave 1,4 glycosidic bonds present in hyaluronate, alginate and pectin [9]. Lipases, produced by a select few phages, hydrolyze the carboxyl ester bonds of triacylglycerols, resulting in the release of organic acids and glycerol [10,11]. Depolymerases of phages destroy biofilms to gain access to bacterial cells [12].
This review summarizes the interactions between phages and bacteria, and the resistance mechanisms that bacteria have developed against phage attacks, including immune systems. The role that phages and phage-derived proteins play in the fight against bacterial infections is discussed, and the limitations of phage therapy are highlighted. The improvement of phage infections using genetic engineering is investigated.
2. Classification of Bacteriophages
Phages are classified into four basic morphological groups, i.e., tailed (order Caudovirales), polyhedral (Microviridae), filamentous (Inoviridae), and pleomorphic (Plasmaviridae). Phage genomes are either ssDNA (Microviridae), dsDNA (Myoviridae), dsRNA (Cystoviridae) or ssRNA (Leviviridae), as shown in Figure 1. Genome sizes vary, ranging from 3.3 kbp in Leviviridae to 500 kbp in Bacillus megaterium phage G [13]. In some cases, archaeal viruses and phages share a similar morphology, as observed in Inoviridae [14]. It is likely that archaeal viruses and phages evolved from a common ancestor that infected similar hosts before the divergence of bacteria, archaea, and eukarya [15]. The International Committee on Taxonomy of Viruses (ICTV) classified bacteriophages into 55 orders, 168 families, 103 subfamilies, 1 422 genera, and 6 591 species [16]. The classification of more prominent phages is listed in Table 1.
The specificity of phages differs, for example, Rhizobium etli phage ph09 has a narrow host range and infects only four strains within the species, contrary to Staphylococcus aureus phage ϕ812 that infects 743 strains, including 38 coagulase-negative Staphylococcus spp. [17,18]. Examples of phages infecting hosts from different genera have also been described, e.g. the promiscuous podophage Atoyac that infects species of the genera Aeromonas, Pseudomonas, Yersinia, Hafnia, Escherichia, and Serratia [19].
Figure 1.
Variations in bacteriophage morphology (adapted from Dion et al. [20]). Double-stranded DNA bacteriophages are divided into tailed phages (Myoviridae, Podoviridae, and Siphoviridae) and non-tailed phages (Tectiviridae, Corticoviridae, and pleomorphic Plasmaviridae). Phages with single-stranded genomes are protected by icosahedral capsids (Microviridae), filamentous capsids (Inoviridae), or envelopes (Leviviridae and Cystoviridae).
Figure 1.
Variations in bacteriophage morphology (adapted from Dion et al. [20]). Double-stranded DNA bacteriophages are divided into tailed phages (Myoviridae, Podoviridae, and Siphoviridae) and non-tailed phages (Tectiviridae, Corticoviridae, and pleomorphic Plasmaviridae). Phages with single-stranded genomes are protected by icosahedral capsids (Microviridae), filamentous capsids (Inoviridae), or envelopes (Leviviridae and Cystoviridae).
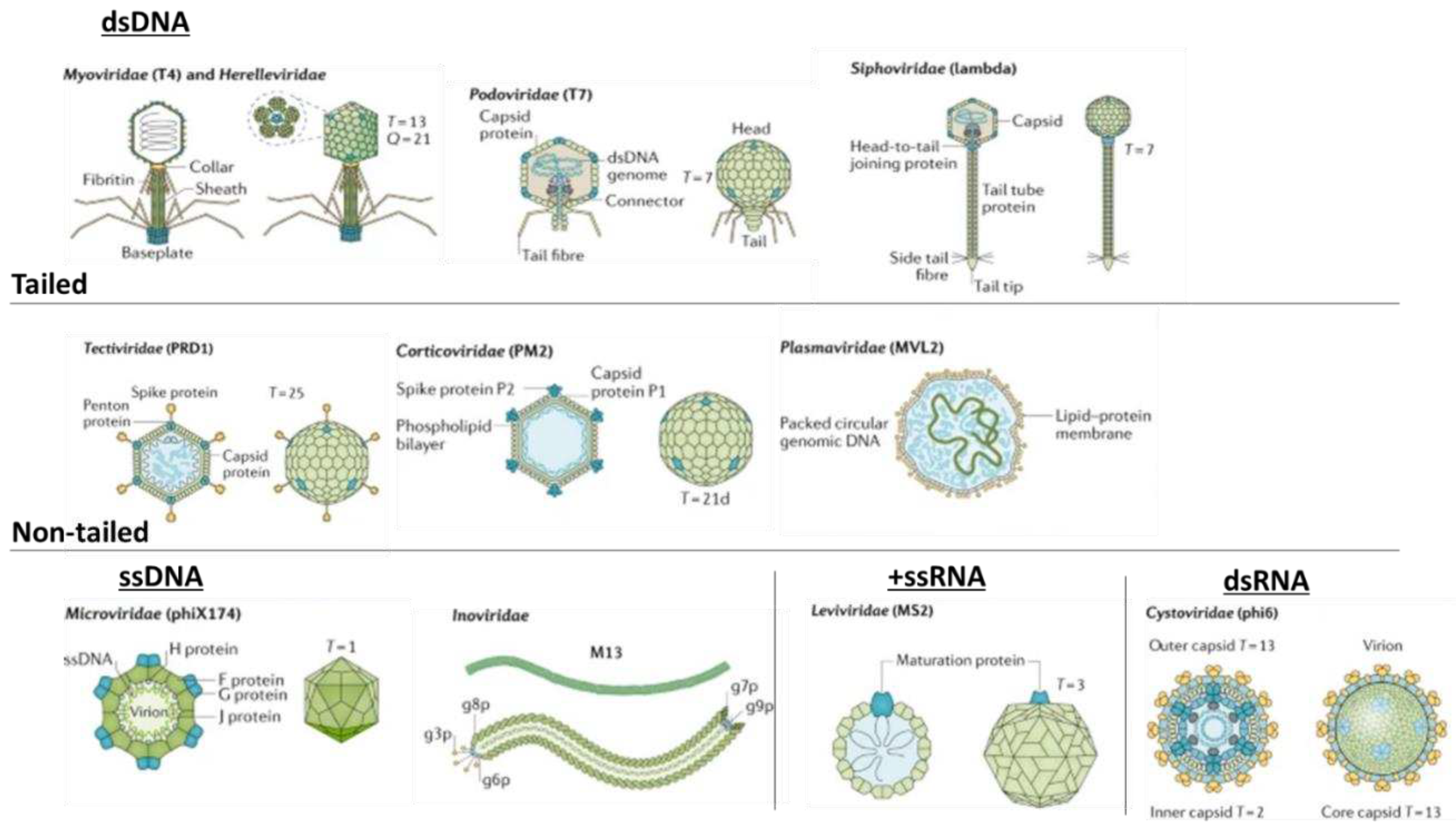

Table 1.
The classification of most prominent phages according to the International Committee on Taxonomy of Viruses (ICTV) based on genome type, morphology, and bacterial host (adapted from Hulo et al. [21]).
Table 1.
The classification of most prominent phages according to the International Committee on Taxonomy of Viruses (ICTV) based on genome type, morphology, and bacterial host (adapted from Hulo et al. [21]).
| Family | Genome | Morphology | Host | Phage |
|---|---|---|---|---|
| Myoviridae | dsDNA | Contractile-tailed, non-enveloped | Enterobacteria, Escherichia coli, and Leptospira spp. | T4, LE3, and phi29 |
| Podoviridae | dsDNA | Short-tailed, non-enveloped | Enterobacteria, E. coli, Bacillus spp., and Bordetella spp. | T7, phi29, and BPP-1 |
| Siphoviridae | dsDNA | Long non-contractile-tailed, non-enveloped | Proteobacteria, Lactococcus spp., Streptomyces spp., and Mycobacterium spp. | Lambda, DS6A, PA-2, phiC31, and D29 |
| Corticoviridae | dsDNA | Non-tailed, non-enveloped | Pseudoalteromonas spp. | PM2 |
| Plasmaviridae | dsDNA | Non-tailed, enveloped, pseudo-spherical | Acholeplasma spp. | MVL2 and AVL2 |
| Tectiviridae | dsDNA | Non-tailed, non-enveloped | Microbacterium spp., Streptomyces spp., and Pseudomonas spp. | PRD1, PR4, and Bam35 |
| Inoviridae | ssDNA | Filamentous, helical capsid | Enterobacteria, Vibrio spp., Spiroplasma spp., Salmonella spp., and Rastonia spp. | M13, fs1, 1-C74, Ike, and RSM1 |
| Microviridae | ssDNA | Non-enveloped, non-tailed, icosahedral capsid | Enterobacteria, Bdellovibrio spp., and Chlamydia spp. | phiX174, Chp1, and PhiMH2K |
| Cystoviridae | dsRNA | Enveloped | Pseudomonas spp. | phi6 |
| Leviviridae | ssRNA | Enveloped | Enterobacteriaceae and E. coli | MS2 |
3. Phage˗Host Adsorption and Cell Entry Strategies
The adsorption of phages to the host involves a series of interactions between binding proteins of the phage and receptors on the surface of the host cell membrane [22]. Examples of phage attachment to bacterial surfaces are shown in Figure 2. Attachment to a host may be due to Brownian motion, dispersion, or flow [23]. Reversible binding allows desorption and the re-infection of another host. Once suitable binding of primary receptors is achieved, irreversible binding of the phage injection machinery is initiated. Enzymatic cleavage of the host cell envelope components reveals specific receptors required by the phage. This leads to conformational changes in phage tail machinery proteins allowing for the injection of the phage genome, and transfection of the host [22]. For some phages, e.g., T5, primary adsorption involves the O-antigen polymannose moiety of lipopolysaccharides (LPS). Irreversible binding occurs at the conical portions of the straight tail which harbor receptor-binding proteins that attach permanently to the ferrichrome outer membrane protein, FhuA [24]. Outer membrane proteins are often hijacked by phages as target sites for receptor-binding proteins (RBPs).
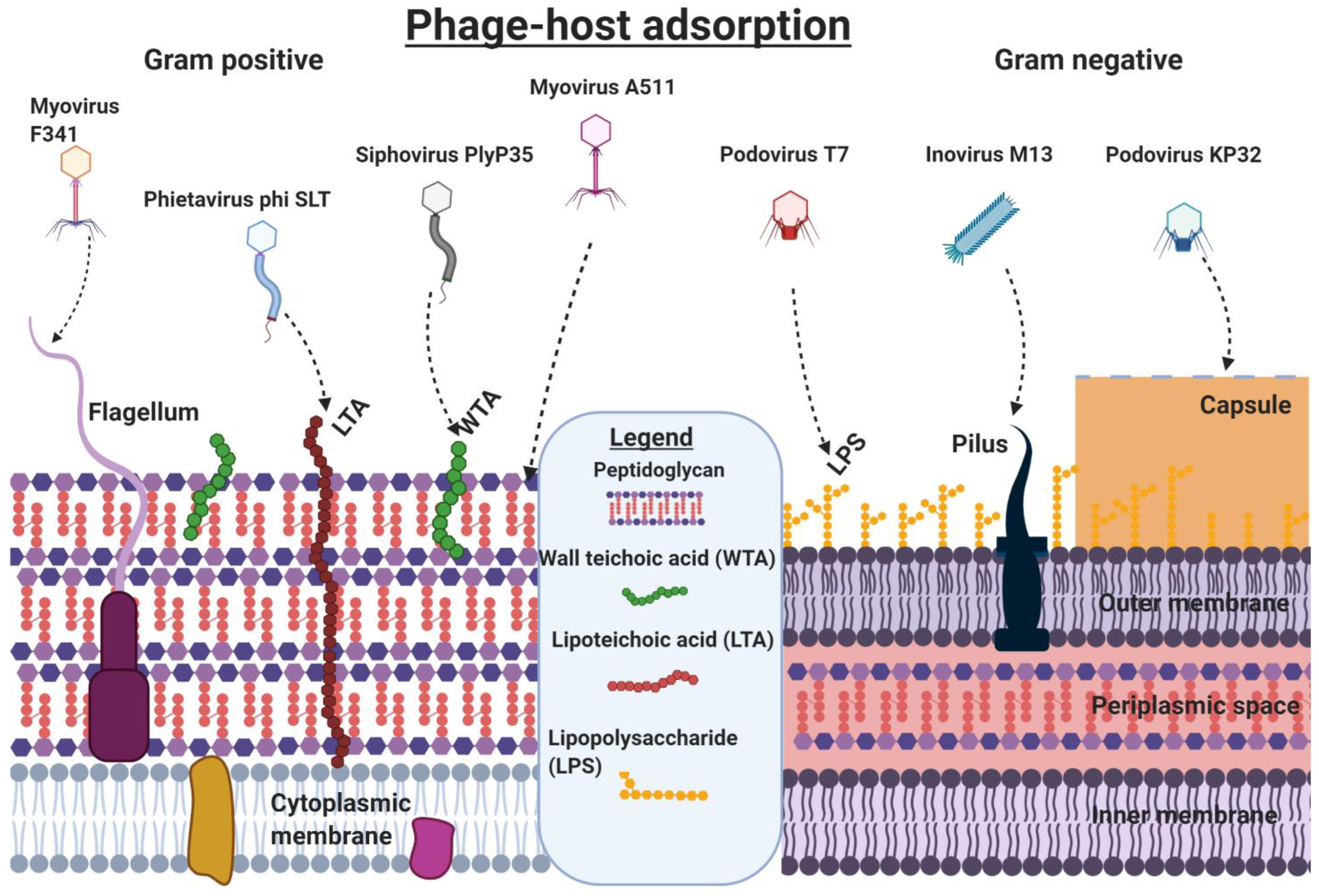
Attachment to Gram-positive bacteria differs from the attachment to Gram-negative bacteria because of variations in the thickness of peptidoglycan layers, and levels of lipoteichoic acids (LTAs), lipopolysaccharides (LPS), and lipoproteins [27]. The different cellular structures facilitating the attachment of phages to bacterial cells are shown in Figure 3. Phage SSU5, isolated from O-antigen deficient Salmonella mutants, could not infect wild-type (WT) Salmonella due to the masking of core polysaccharides by the O-antigen [28]. In contrast, O-antigen-specific phages could not infect O-antigen mutants lacking receptors for core polysaccharides. Specificity is key to phage infections and is particularly apparent for Mu G(+) that targets the terminal Glcα-2Glcα1 or GlcNAcα1-2Glcα1 within LPS [29]. Saccharide moieties are abundant in some Gram-positive bacteria and many phages have taken advantage of these, making them suitable sites for host attachment.
A Siphoviridae LL-H phage targets glucose moieties of LTA and irreversibly binds to a glycerol phosphate group in the LTA of Lactobacillus delbrueckii [30]. Glucose, rhamnose, and galactose in cell wall pneumococcal capsular polysaccharides (PPS) have been used by phages as receptor sites [31]. Phages can also bind directly to peptidoglycan, as observed for Listeria phage A511 which exhibits a broad host range due to the conserved nature of peptidoglycan [32]. In many cases, phages need to gain access to the underlying structure of the bacterial cell wall and degrade a path through the cell envelope to access the necessary receptors responsible for viral entry. This is done by using LPS-specific glycanases and deacetylases. Similarly, phages that infect Gram-positive bacteria produce exolysins that degrade peptidoglycan layers [33,34]. In Gram-positive bacteria, teichoic acid in the cell wall (WTA) provides strength, hydrophobicity, and zwitterionic properties to attract cations such as Ca++ and K+ and may also serve as adhesins to attach to surfaces and other bacteria. A SPP1 phage infecting Bacillus subtilis targets WTA as its primary reversible receptor and then facilitates irreversible adsorption to its host by a secretory system protein YueB [35]. Other mechanisms that play a role in infection are the transmission of signals, and tactics used to inject the virome [36].
Some phages target variable cellular surface structures such as pili, flagella, and polysaccharides. Other phages are plasmid-dependent and will only infect cells carrying, and expressing, genes involved in phage entry [37,38]. Flagellotropic phages with chi-like tail fibers target H-antigens on flagella, as observed for Salmonella serovars [39,40]. Klebsiella pneumoniae is well known for its ability to produce exopolysaccharides. Phages attacking Klebsiella produce depolymerases that degrade the depolysaccharide layer. An example is depoKP36 which degrades capsules and exposes underlying structures required for phage infection [41]. Modification of capsules, or loss of capsule formation, results in the development of phage resistance, as demonstrated by Song et al. [43]. Several phages of Leviviridae, Inoviridae, and Cystoviridae bind to conjugative pili, Type IV pili, and other pili involved in bacterial attachment [44,45,46]. Jalasvuori et al. [47] investigated plasmid-dependant bacteriophages to initiate antibiotic susceptibility in previously resistant strains of Salmonella enterica and E. coli. The plasmid-dependant phage PRD1 drives phage resistance by causing a loss in plasmids containing antibiotic-resistant genes. This allows bacteria to avoid phage infection by silencing the expression of phage receptors.
Figure 3.
Phages use different cell structures as attachment sites, and entering of the host’s cytoplasmic membrane. Gram-positive bacteria are susceptible to phage infiltration through their peptidoglycan (Listeria phage A511), WTA (Listeria phage Plyp35), lipoteichoic acids (LTA, Staphylococcal phage phi SLT), and flagella (Campylobacter phage F341). Gram-negative hosts can be targeted via their lipopolysaccharides (LPS, E. coli Phage T7), or pili (E. coli Phage M13), and capsules (Klebsiella phage KP32). Information was obtained from Dunne et al. [48]. Created using Biorender.
Figure 3.
Phages use different cell structures as attachment sites, and entering of the host’s cytoplasmic membrane. Gram-positive bacteria are susceptible to phage infiltration through their peptidoglycan (Listeria phage A511), WTA (Listeria phage Plyp35), lipoteichoic acids (LTA, Staphylococcal phage phi SLT), and flagella (Campylobacter phage F341). Gram-negative hosts can be targeted via their lipopolysaccharides (LPS, E. coli Phage T7), or pili (E. coli Phage M13), and capsules (Klebsiella phage KP32). Information was obtained from Dunne et al. [48]. Created using Biorender.
Once attachment to the host and degradation of the host’s surrounding cell barriers are achieved, the phage proceeds to enter the host’s cytoplasm through a variety of different measures, depending on the phage’s morphology. Tailed phages use specialized tail structures to deliver their genome into the host with variations occurring across the three subfamilies. Myoviridae that infect Gram-negative bacteria allow for conformational changes to their baseplate and trigger the expulsion of a rigid internal tube, acting as a channel crossing the host cell’s envelope. This is followed by fusion with the cell membrane, driving the phage genome from the capsid into the host’s cytoplasm [49]. This process is similar in Gram-positive bacteria, except that phages use specialized tape measure proteins and the opening of a proximal plug that joins the capsid with the tail and then releases the phage genome. Although the precise role of the tape measure protein is unknown, it is believed to aid movement across the host’s thick cell envelope. Some tail fiber structures produce murein hydrolase that degrades peptidoglycan [50]. Phage T5, a Siphoviridae phage, uses tails to interact with the iron-siderophore receptor FhuA on the host, resulting in conformational changes. This leads to localized degradation of peptidoglycan and pore formation, causing fusion of the host's outer and inner membranes, and providing safety to viral DNA from periplasmic nucleases. It also creates a pore through which the genetic material is released into the host’s cytoplasm [51]. In essence, much is the same for Podoviruses that infect Gram-negative hosts. Once their tail tip reaches the cell membrane, it forms a conduit for genome injection. In Gram-positive hosts, it is believed that Podoviridae burrows a tunnel into the host’s thick peptidoglycan layer with exolysins reaching the cell membrane surface [52].
The filamentous M13 phage enters the host by binding to the host pilus. Then, the pilus retracts toward the host’s inner membrane bringing the phage along with it [53]. The phage’s two N-terminal domains are involved in binding to primary and secondary receptors on the host whilst the C-terminal is responsible for virion uncoating and facilitating the release of DNA into the host’s cytoplasm. Tectivirus PRD1 is an unusual phage in that it directly fuses with the host membrane and, as it is non-enveloped, and has a capsid that surrounds an inner membrane. The phage’s capsid protein, P2, is used for receptor recognition and initiates irreversible binding. A P11 protein is responsible for outer membrane penetration, and a P7 protein digests the peptidoglycan layers after which it is unclear whether cell membrane penetration or fusion occurs. Finally, there is evidence that a tubular structure forms between the capsid and host cell membrane which facilitates DNA injection [54]. Members of the Cystoviridae have a lipid-rich envelope encapsulating their nucleocapsid and are believed to have a similar mechanism of entry to animal viruses. Adsorption to the host is mediated through pilus retraction. The phage envelope and host outer membrane fuse together, followed by peptidoglycan degradation allowing the nucleocapsid to enter the periplasmic space. An invagination of the host cell membrane occurs facilitating entry of the virion into the host cytoplasm in an endocytic-like vesicle. Internal core particles containing the phage genome are released into the host cytoplasm and the segmented dsRNA is polycistronically transcribed into mRNA by viral RNA-dependent RNA polymerase [55,56]. All of this is performed within the core particle and exported out, avoiding host antiviral mechanisms, and providing a template for the translation of viral proteins.
4. Phage-Host Interactions
Phages are obligate parasites and are reliant on bacteria to complete their lifecycle. Bacteria have developed strategies to negate phage infections. Host defense strategies can be categorized into exogenous, physical barriers, and endogenous defensive mechanisms, e.g., clustered regularly interspaced palindromic repeat (CRISPR)-CRISPR-associated proteins (Cas) systems, restriction modifications, and abortive infections. These strategies are often overcome by phages through mutation, or changes by hosts to block certain phages making them susceptible to other phages targeting different or modified receptors. In addition, phages can modulate host virulence, allowing for the survival of their hosts and subsequently the phages when their hosts are under threat from the human immune system [57]. Phages are known to inactivate host-specific RNA polymerases inhibiting host translation and some engage in superinfection exclusion whereby they inhibit further infection of their host by other phages, including the same phages [58,59]. Some modulate receptors are used by other phages, thus blocking entry, and preventing secondary infection of their host. The co-evolution of bacteria to resist phage attacks can impact host growth, virulence, and environmental fitness [60].
5. Resistance to Phage Attack
As bacteria evolve, they generate mechanisms to avoid phage infection but in doing so they encourage many phages to evolve counter mechanisms. Hosts may undergo changes to their cell walls, such that they express new RBPs to combat phage adsorption. In this instance, phages can adapt to these changes. For example, λ phage typically binds to LamB but E. coli can diverge to express a new receptor, OmpF. However, λ phage can evolve to sustain its cell tropism. This is achieved through a mutation to the distal domain of the J protein which in turn is achieved through a combination of mutations to the RBP gene [61]. Some bacteria that produce capsules or exopolysaccharide structures hinder the accessibility to RBPs necessary for phage infection. Interestingly, a strain of capsulated E. coli EV36 was found to avoid T7 phage infection using its K1 capsule and subsequent removal of the capsule led to plaque formation [62]. Some phages can produce a depolymerizing enzyme with the ability to degrade the host capsule. This has been observed in coliphages, K1F and K1-5, that possess endosialidases that hydrolyse K1 capsules [62]. The endosialidase, encoded by gp143, appears at the distal portion of the tail (Myoviridae) and is one of five spike proteins that facilitates accessibility for other RBPs to interact with host receptors [63]. Bacterial capsules differ vastly. Streptococcus pyrogenes synthesizes a capsule composed of hyaluronic acid, whilst Pseudomonas spp. produce exopolysaccharide capsules. Klebsiella spp. produce 77 distinct capsule serotypes which differ in their capsular monomers, stereochemistry, and glycosidic linkages [64]. These bacteria are still susceptible to phages, but they require different depolymerases. Examples are Streptococcus phage H4489A which produces hyaluronidase, and Pseudomonas phage AF which contains an exopolysaccharide hydrolase [65,66]. Alternatively, bacteria can avoid infection by expressing their phage target surface proteins in a stochastic manner, during phase variations or physiological regulations, but phages can counter this by relying on alternative receptors or target host surface structures that are a necessity to the host [67]. An example of the latter was observed in Campylobacter jejuni which avoided phage infection by phase-variable expression of the O-methyl phosphoramidate (MeOPN) moiety in its capsule [68]. Furthermore, Holst Sørensen et al. [69] observed acquired resistance in vivo as phage-resistant C. jejuni were selected either lacking the MeOPN or had gained a 6-O-Me group on the capsule. Lastly, some Vibrio and Escherichia spp. deploy extracellular vesicle decoys displaying RBPs on their surfaces that adsorb phages from the environment, lowering environmental phage titer and lowering host exposure [70].
6. Bacterial Immunity to Phage Infections
Immune systems are not only present in complex multicellular organisms. Prokaryotes also have a primitive immune system conferring adaptive immunity against bacteriophages. They rely on four mechanisms of action, namely a restriction-modification system, a DNA end degradation system, a CRISPR-cas system, and a system stimulating abortive infection [71]. More than 80% of bacterial genomes respond to incoming viral infections with at least one restriction-modification system [72]. Using modification enzymes, bacteria can methylate their own DNA protecting it from restriction endonuclease activity, which cleaves any unmethylated DNA such as phage DNA. To overcome this defense mechanism, phages can encode their own strain-specific modifying enzymes that methylate specific sequences of the phage DNA, thereby preventing degradation by bacterial endonucleases. Many T1 phages mask the recognition sites required by restriction enzymes Sau3AI and DpnI by methylation using Dam methyltransferase during genome packaging [73]. The T7 phage inhibits restriction enzyme activity with an anti-restriction protein, Ocr (overcome classical restriction), an early expressed protein that blocks the active site of type I DNA restriction enzymes by structurally mimicking the phosphate backbone of bent B-form DNA [74].
Alternatively, some E. coli and Salmonella spp. can express nucleases that actively seek out DNA with free ends. The RecBCD enzyme is a helicase-nuclease responsible for dsDNA repair but also protects bacteria by degrading invading linear DNA from phages and extra-chromosomal DNA [75]. This can be countered using gp2, found in T4 phage, which attaches to the free-ended phage DNA thereby blocking accessibility of RecBCD’s active site. Similar activity has been observed in Lambda and Mu phages by Gam protein [76,77]. The CRISPR/Cas-associated genes have been acknowledged as the DNA-encoded and RNA-mediated adaptive immune system of bacteria. This process occurrs in three stages, namely adaption, CRISPR RNA (crRNA) biosynthesis and targeting [78]. During adaption, invading phage nucleic acids are integrated into a CRISPR array, comprised of CRISPR loci (21-48 bp) direct repeats interspaced by the newly acquired phage nucleic acids known as CRISPR spacers. This array is then translated and processed by cas endoribonucleases within the repeated sequences to synthesize small crRNAs. It is important to note that the crRNA and cas proteins form a complex complementary to incoming phages which in turn is identical to the parent phage. This phenomenon induces a sequence-specific cleavage of phage nucleic acids preventing the proliferation of the phage genome and viral progeny [71,78]. Evading adaptive immunity is not only seen in eukaryotes but it has also been reported in bacteriophages. It has been observed that Streptococcus thermophilus phages overcome their host’s resistance in the form of a newly gained spacer. This is facilitated by a single point mutation in the target spacer which inhibits the formation of cas protein-crRNA complex, impacting the specificity of the cas protein and allowing the phage target to continue proliferating within the host [79]. A Vibrio cholera phage, ICP1, counteracts the host antiviral defense islands, phage-inducible chromosomal islands (PICIs) like elements (PLE), by encoding a CRISPR/Cas-like system of its own to mediate DNA cleavage and destruction of PLE [80]. Many more anti-CRISPR genes involved in host defense evasion have been extensively investigated by Pawluk et al. (2014), showing some Pseudomonas phages carry multiple mechanisms to combat different types of CRISPR systems [81,82].
Abortive infection mechanisms instigated by hosts are diverse. In Lactococcus lactis a gene related to abortive infection (abiZ) is known to accelerate the lysis process within the host before matured viral progeny can be produced [83]. Other mechanisms causing abortive infection are E. coli Lit proteins inhibiting translation, and Rex proteins, A and B, recognizing the phage DNA-protein complex thus facilitating membrane depolarization and ultimately cell death [84,85]. These mechanisms are deployed by 'sacrificial’ bacteria to prevent further infection of the entire population.
7. Bacteriophage-Based Therapeutics
7.1. Phage Therapy
Several in vitro studies have suggested that phages may be used in the treatment of CF [86,87,88]. However, studies in humans are mostly limited to case reports. Only a few randomized, placebo-controlled trials have been reported. Whilst a few trials have shown that phages are safe therapeutic agents, they often do not supersede the standard of care (SOC) antibiotics or conventional treatments used in disease management (Table 2). In a clinical trial using phages to treat complicated urinary tract infections (UTIs), the placebo and treatment with antibiotics resulted in a 37% and 28% success rate, respectively. This superseded the 18% success rate reported with phage treatment [89]. In-depth and well-designed clinical trials are required to assess the efficacy of phage therapy and phage-antibiotic therapy.
Phages were successfully used in the treatment of a multi-drug-resistant Acinetobacter baumanii infection [90] and the eradication of P. aeruginosa from aortic grafts [91]. In the latter study, the patient was treated with a lytic phage (OMKO1) bound to the outer membrane protein Mof, and the mexAB- and mexXY-multidrug efflux systems of P. aeruginosa [92]. Targeting these efflux pumps increased the susceptibility of the pathogen to ceftazidime by twofold and to ciprofloxacin by tenfold. In addition to the impact of OMKO1 on antibiotic susceptibility, the phage destroyed the biofilms deposited on the implanted device [92]. A patient suffering from CF who underwent a bilateral orthotopic lung transplant to eradicate a chronic Mycobacterium abscessus pulmonary infection did not respond to treatment with antimycobacterial agents [93]. Treatment with a phage cocktail, also active against Mycobacterium smegmatis, eradicated M. abscessus. The repressor gene from two of the most strain-specific phages in the cocktail had to be deleted to convert the phages from temperate to lytic. For a listing of clinical data published from 2000 to 2021 that involved 2 241 patients who were treated with phage therapy, and the safety and efficacy of phage therapy, the reader is referred to the review by Uyttebroek et al. [94]. In this review the authors summarize the effect of phage therapy in the treatment of pneumology, urology, orthopaedics, dermatology, otorhinolaryngology, ophthalmology, gastroenterology, cardiology, and intensive care medicine. Clinical improvement was seen in 79% of patients and bacterial eradication in 87% of patients who were on phage therapy. Case studies of phage therapy and the outcome of the results are listed in Table 2 and Table 3.
Bacteria that develop extreme resistance to phage treatment are usually eradicated by using phage combinations (cocktails), higher phage titers, or phage cocktails combined with antibiotics. The latter approach is not always successful, as phages may transfer antibiotic resistance genes to bacteria and lead to the development of genetically altered, or extremely resistant, pathogens [95,96,97]. Genes encoding resistance to β-lactams (blaTEM), fluoroquinolones (qnrS), macrolides (ermB), sulphonamides (sulI), and tetracyclines (tetW) have been detected in the viromes of phages present in activated sludge [98], urban wastewater, hospitals [99,100,101,102], freshwater fish [103], and human faeces [104].
Reports of fluoroquinolones and anticoagulants that induce the expression of prophage genes, and the spreading of temperate phages [105] are alarming and may in the future influence phage therapy. Colomer-Lluchn et al. [106] have shown that treatment of wastewater with EDTA or sodium citrate activates the lytic cycle of lysogenic phages, which increases the release of phages from infected cells and the spreading of antibiotic resistance genes located on viromes. Phages isolated from patients infected with antibiotic-resistant bacteria may carry genes encoding resistance to the same antibiotics. This was observed in phages isolated from patients with cystic fibrosis (CF) who received extensive antibiotic treatment [107]. The authors identified 66 genes that may each encode an antibiotic efflux pump. Of these, 15 genes encoded resistance to fluoroquinolone and nine to β-lactamase. Although these findings are of major concern, other studies have shown that the risk of transduction, although possible, is lower than originally anticipated. Enault et al. [108] argued that genes encoding antibiotic resistance are not commonly found in the virome [108]. Furthermore, the methods that are used to detect antibiotic-resistance genes in viromes have been questioned [86].
Table 2.
Post and current phage therapy and phage-associated clinical trials with an impact on human health (updated from Abedon et al. [109]).
Table 2.
Post and current phage therapy and phage-associated clinical trials with an impact on human health (updated from Abedon et al. [109]).
| Infection(s)/phage trial interest | Causative agent(s)/agents of interest | Outcomes/comments | Reference/clinical trial identifier |
|---|---|---|---|
| Suppurative skin infections* | Pseudomonas, Staphylococcus, Klebsiella, Proteus, and E. coli | Thirty-one patients were treated orally and locally for chronically infected skin ulcers with a 74% success rate | [110] |
| Acute postoperative empyema in chronic suppurative lung diseases* | Staphylococcus, Streptococcus, E. coli, and Proteus | Phage-antibiotic combinations were used in the successful treatment of 45 patients | [111] |
| Complications due to bacterial infections in cancer patients* | Staphylococcus and Pseudomonas | 82% (65) successful treatment with phages compared to patients treated with antibiotics 61% (66) | [112] |
| Recurrent subphrenic abscess* | Antibiotic-resistant E. coli | A single patient was successfully treated with phages after 33 days | [113] |
| Urinary tract infections* | Staphylococcus, E. coli, and Proteus | Forty-six UTI patients were treated with phages with 92% making clinical improvements and 84% achieving bacterial clearance | [114] |
| Rhinitis, pharyngitis, dermatitis, and conjunctivitis* | Staphylococcus, Streptococcus, E. coli, Proteus, enterococci, and P. aeruginosa | Patients were treated with phages (360), antibiotics (404), and phage-antibiotic combinations (576). Clinical improvements of 86%, 48%, and 83% across the treatment regimes, respectively | [115] |
| Cerebrospinal meningitis * | K. pneumoniae | Successful treatment with orally administered phages in a newborn. | [116] |
| Bacterial diarrhea | E. coli | Orally administered coliphages showed no improvement in clinical outcome, some dysbiosis with streptococci was observed | [117] |
| Complicated or recurrent UTI patients with transurethral resection of the prostate | Enterococcus, E. coli, streptococci, P. mirabilis, P. aeruginosa, staphylococci | Patients with intravesical administered pyophage cocktail, orally administered antibiotics, and a placebo bladder irrigation. Success rates of 18%, 28% and 37% were observed, respectively | [118] |
| Burn wounds | P. aeruginosa | Phages PP1131 showed no significant difference to standard of care antibiotics - patients treated with PP1131 were found to have phage-resistant P. aeruginosa | [119] |
| Prosthetic joint infections | S. aureus, S. epidermidis, S. lugdunensis, Streptococcus sp., E. faecium, E. faecalis, E. coli, P. aeruginosa, and/or K. pneumoniae | Phage treatment, with intraoperative and intravenous PhageBank™ bacteriophages, in conjunction with standard-of-care antibiotics/Debridement, Antibiotics, and Implant Retention (DAIR) procedures. Completion is predicted in 2024 | [120] |
| Diabetic foot ulcers (DFU) | Staphylococcus sp., wound microbiome | Use of anti-staphylococcal phage gel (Intralytix Inc, Baltimore, Maryland, USA). Effect on bacterial microbiome of DFU wounds and patient outcomes. Trial was abandoned for funding reasons | [121] |
| Probiotic application for overall gut health | Bifidobacterium animalis subsp. lactis BL04 | The use of bacteriophages (PreforPro) increased the survival and efficacy of probiotic bacteria administered vs probiotics only vs placebo | [122] |
| Phages preventing the acquisition of multi-resistant enterobacteria (PHAGE-BMR) | E. coli or K. pneumoniae containing ESBL or carbapenemases | Collection of multidrug-resistant bacteria from patients in intensive care, subsequent search for presence and absence of phages in carriers/non-carriers. Currently active but of unknown status | [123] |
| Phage dynamics and influences during human gut microbiome establishment (METAKIDS) | A broad range of bacteriophage and bacterial hosts. | Characterize phage and bacterial genomes, abundance, and variations during infant gut development. Terminated | [124] |
| Bacterial infection in cystic fibrosis patients | P. aeruginosa | A cocktail of 10 bacteriophages was used to reduce Pseudomonas presence after 6 and 24 h including sensitivity of isolates. Completed with no recorded outcomes | [125] |
| Prebiotic | Escherichia coli and microbiota | Commercial coliphage cocktail effects on the microbiota and systemic inflammation. No disruption to microbiota and no effect on inflammatory markers | [126] |
| Venous leg ulcers | P. aeruginosa, S. aureus, and E. coli | Polyvalent phage preparation of 8 bacteriophages was assessed for their safety and efficacy. No available outcomes but the trial was completed | [127] |
| Lower urinary tract colonization | E. coli | Assess the safety, tolerability, pharmacokinetics, and pharmacodynamics of phage cocktail LBP-EC01 | [128,129] |
| Safety of topical phage solution intended for wound infections | S. aureus | Evaluating the safety and skin reactions to ascending doses of phages compared to the placebo | [130] |
Abbreviation: ESBL - extended spectrum β-lactamase. Those marked with a (*) originated in Poland and the Soviet Union.
Table 3.
Recent individual case studies of personalized phage therapy that impacted patients with multi-drug resistant infections.
Table 3.
Recent individual case studies of personalized phage therapy that impacted patients with multi-drug resistant infections.
| Infection(s) | Bacterial specie(s) | Outcome/comment | Reference |
|---|---|---|---|
| Complicated necrotizing pancreatitis | Acinetobacter baumannii | Clearance of A. baumannii and return to health using intravenously (IV) and percutaneously administered (9) phages screened from a phage bank | [131] |
| Bacteremia | P. aeruginosa | An IV-administered bacteriophage cocktail comprised of two phages cleared the bacteremia, but the patient succumbed to other complications | [132] |
| Lung infection and transplant recipient | P. aeruginosa | An IV and nebulizer-administered bacteriophage cocktail, AB-PA01 and Navy, with the patient recovering from pneumonia | [133] |
| Infection of left ventricular assist device | P. aeruginosa | Six-week IV-administered (3) phage cocktail, the patient was clear and then relapsed but a change in antibiotics led to recovery | [132] |
| Osteomyelitis | A. baumannii and K. pneumoniae | The patient developed post-operative infection with multidrug-resistant isolates. IV bacteriophage-antibiotic combination led to the patient’s full recovery without the need for amputation | [134] |
| UTI | ESBL E. coli | Phage treatment with two phages over 23 days in conjunction with antibiotic treatment led to negative urine cultures and full recovery of the patient | [132] |
| CNS infection of a recovering trauma patient | A. baumannii | IV treatment with an A. baumannii phage for 8 days led to CSF cultures coming back negative for A. baumannii but positive for K. pneumoniae and S. aureus. The patient was declared brain dead and later announced deceased | [132] |
| Lung infection of cystic fibrosis patient | Achromobacter xylosoxidans | Cefiderocol and phage treatment were performed for 5 days followed by continuous phage therapy. The patient recovered and was discharged | [135] |
Abbreviations: IV - intravenous; CNS - central nervous system; UTI,- urinary tract infection; CSF – cerebrospinal fluid.
7.2. Therapeutic Potential of Phage-Derived Proteins
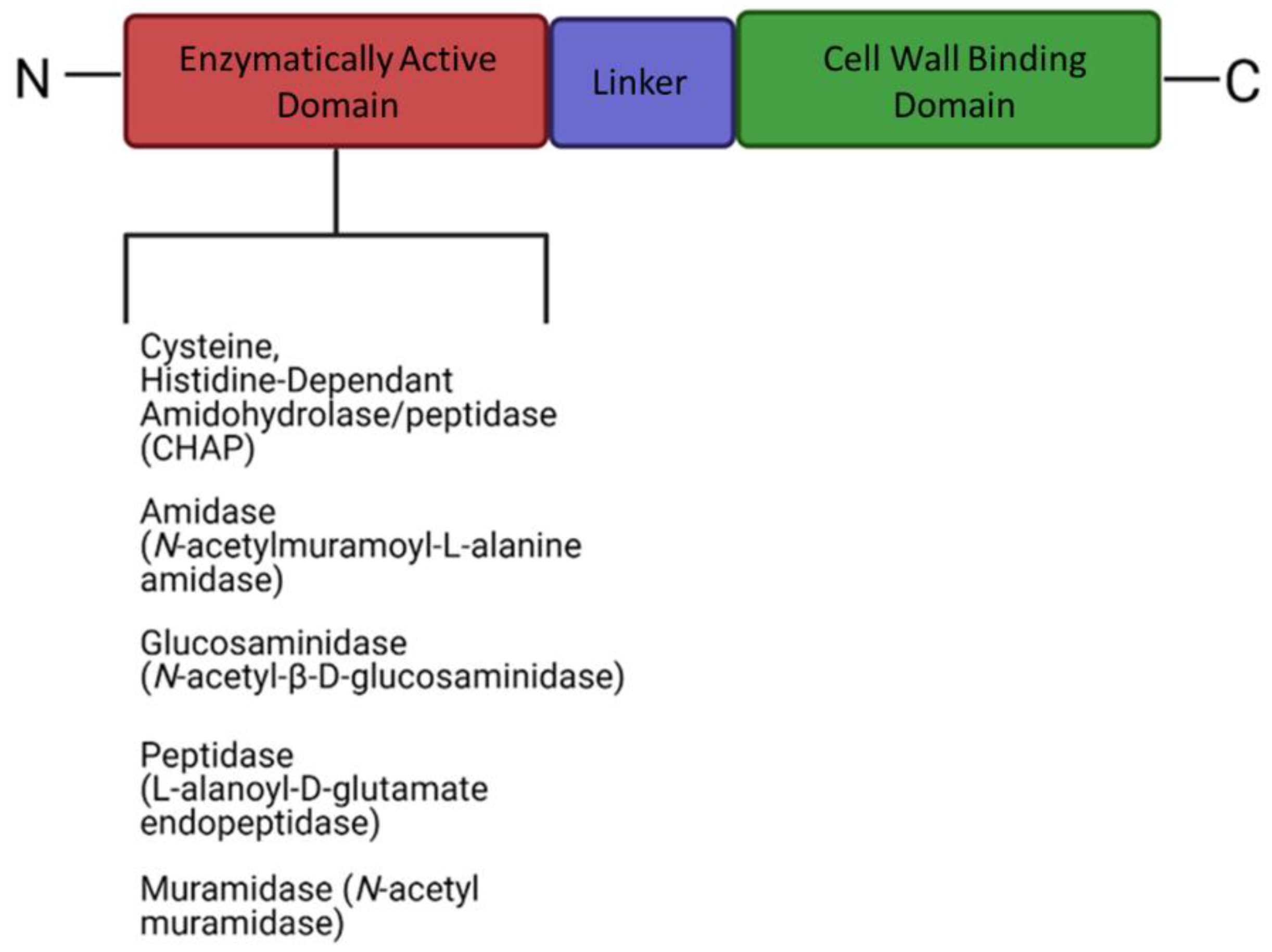
Bacteriophages produce a range of enzymatically active proteins required for their adsorption, entry, and exit from their susceptible hosts. During the late phase of infection, bacteriophages produce endogenous lysins, allowing host lysis and subsequent release of viral progeny. Lysins are part of a lysis cassette and rely on two other genes, namely holin and spannin, to help with the translocation of lysin across the cell membrane to peptidoglycan [6,7,8]. Phage lysins are usually composed of a two-domain structure (Figure 4), although exceptions have been reported. PlyPalA is an important lysin against Paenibacillus larvae, the causative agent of American foulbrood, which is detrimental to honeybees [136]. The activity of endolysins can vary and activity has been observed against sugars constituting the bacterial cell wall i.e., they may be endo-β-N-acetylglucosaminidases or N-acetylmuramidases (lysozyme). Endopeptidases, which degrade protein moieties, amidases such as N-acetylmuramoyl-L-alanine amidase which degrade amide bonds between glycans and peptides have also been reported. Lysins can also fall under a broader class of Cysteine Histidine-dependent Amidohydrolase/Peptidases (CHAPs) with one example of such observed in Streptococcus pyogenes producing a CHAP-like lysin that hydrolyzes the 1,4-β-glycosidic bonds between N-acetyl-d-glucosamine and N-acetylmuramic acid together in the peptidoglycan chain [137]. Contrary to the activity of the N-terminal, the C-terminal domain is usually involved in substrate binding and host specificity. Substrates include carbohydrates found in the cell wall of bacteria and the C-terminal is paramount for efficient cleavage of cell wall substrates [9]. These lysins form pores in the cell wall by hydrolyzing peptidoglycan, disrupting cell wall integrity and in turn hypertonic lysis. Although the impact of lysins on Gram-positive bacteria is promising, little activity is observed against Gram-negative bacteria which could likely be due to the bioavailability of peptidoglycan being blocked by the gram-negative cell envelope. Few endolysins are endogenously active in vivo, such as SPN9CC, PlyF307, and CfP1gp153. Lysins traverse the outer membrane with the help of external agents [138,139,140]. The mode of action of lysins has led to lysin-based medicinal applications, such as lysin-antibiotic combinations that can combat antibiotic resistant bacteria. Djurkovic et al. (2005) found various antibiotic combinations efficacious such as gentamicin and penicillin, with a streptococcal phage lysin, CpI-1 [141].
They also found that a combination of penicillin and CpI-1 was highly active against previously penicillin-resistant strains. There have been recent successful results in a randomized controlled trial using an anti-staphylococcal lysin (exebacase) to treat bloodstream infections involving methicillin-resistant S. aureus [142]. They found exebacase in conjunction with antibiotics proved more efficacious than antibiotics alone, and that treatment reduced hoptizilization time by 4 days and readmission of patients by 48%.
Figure 4.
The generalized structure of endolysins containing an N-terminal enzymatic domain and a cell wall binding domain on the C-terminal (created using Biorender).
Figure 4.
The generalized structure of endolysins containing an N-terminal enzymatic domain and a cell wall binding domain on the C-terminal (created using Biorender).
Contrary to endolysins, exolysins or phage-encoded depolymerases (Table 4) are usually found on phage tail fibers, tail spike proteins (TSPs), or the phage baseplate. Importantly, they cleave polysaccharides located on the bacterial cell envelope and are involved in host adsorption. Exolysins can be classified into two main classes, i.e., hydrolases and lyases, which act on a carbohydrate substrate such as capsule polysaccharides (CPS), extracellular polysaccharide (EPS) matrices, and O-polysaccharides. Based on the substrate hydrolases act upon, they can be further subclassed into groups such as sialidases, rhamnosidases, levanases, xylanases, and dextranases. Many hydrolases rely on a water molecule to specifically cleave the O-glycosidic bonds between polysaccharide monomers [143]. Sialic acid capsules are used by several bacterial species including E. coli K1, Haemophilus influenza, Streptococcus spp., and Campylobacter jejuni. Capsules promote pathogenesis by improving adherence to surfaces, evasion of host immune responses, biofilm formation, and acting as a nutrient source [144]. Phages encode endosialidases within their tail structures to overcome this carbohydrate barrier. Activity has been seen against a neuropathogenic E. coli K1 strain by podovirus K1E which encodes a hydrolytic tail spike protein that specifically binds and cleaves the K1 capsule [145]. An endosialidase, Endo92, from phage phi92 was capable of digesting K1 and K92 capsules of E. coli and is uniquely able to cleave both the α-2,8- and α-2,9-linkages of sialic acid [146]. Levanases are predominately found in bacterial species such as Bacillus and Pseudomonas and can hydrolyze the β-2,6-linked D-fructofuranosyl residues of levan [147,148]. Levan is an important structure in the development of a robust biofilm for Bacillus spp., however it is not a necessity. It plays a role in the stability of floating biofilms, can provide a nutritional reserve, and was found to be the majority polysaccharide present in the EPS matrix [149]. Levanases have been found in several B. subtilis phages (SP10, ϕNIT1 and SPG24) and assist phages by exposing receptors [150]. Endorhamnosidase activity was first observed in Salmonella (ser.) Typhimurium phage P22 which degrades the O-antigen present on the LPS of S-strains [151]. Specific cleavage by the P22 tail spike protein targets α-rhamnosyl 1-3 galactose linkages of the O-antigen, which is also seen in several other Salmonella phages [31]. Often mutations in the LPS lead to insensitivity to certain bacteriophages but also contribute to less virulent strains of bacteria [152]. An earlier study found that Klebsiella phages exhibit galactosidase or glucosidase activities which cause degradation of side chains present in CPS [153].
Phage polysaccharide lyases cleave the 1,4 glycosidic bonds using a β-elimination mechanism. These enzymes appear to act on three types of polysaccharides including hyaluronate, alginate and pectin, although not exclusively. Hyaluronidases first drew attention with several bacterial species producing them, and it was attributed to be a virulence factor for tissue permeability and pathogen invasion. It is thought that this same enzyme is used in streptococcal prophages to penetrate hyaluronic acid capsules, likely facilitating host entry. Alginate lyases can be mannuronate or guluronate lyases that degrade the two 1,4-glycosidic linked monomers, α-L-guluronic acid and β-D-mannuronic acid, within alginate. Alginate provides structural integrity in brown algae but is also synthesized in Pseudomonas and Azotobacter species shown to contribute to biofilm formation. Alginate lyases are encoded in tail components of Pseudomonas and Azotobacter phages assisting penetration of phages across the acetylated poly(M)-rich EPS matrix allowing phages to bind to the cell envelope. Uropathogenic E. coli (UPEC) produce a capsular polysaccharide rich in colonic acid which allow protection against hostile environments and promote pathogenicity [154]. This negatively charged polymer contains glucose, galactose, fucose and glucuronic acid and is upregulated in established biofilms [155]. There is evidence of phages overcoming this carbohydrate barrier, for example Phi92 contains a colanidase tail spike protein which degrades colonic acid allowing secondary tail spikes to degrade and/or bind to the cell envelope [146]. Lipases are rarely seen in phage genomes but are ubiquitous in nature. They have a broad specificity and often multifunctional properties. Phage lipases hydrolyze the carboxyl ester bonds of triacylglycerols releasing organic acids and glycerol. The role of lipases in phages has yet to be elucidated [10,11]. There is some evidence that a lipase or esterase could be used to modify the O-antigen present on the LPS preventing further phage infections [156].
Table 4.
Bacteriophage-encoded depolymerases that contribute to host adsorption.
| Enzyme class | Phage/enzyme | Polymer substrates | Targeted genera | Reference |
|---|---|---|---|---|
| Hydrolases | ||||
| Sialidases | Phi92 | Polysialic acid | E. coli K1 & K92 | [146] |
| K1E | E. coli K5 | [157] | ||
| K1F | E. coli K1 | [158] | ||
| Levanase | SP10 | Levan | Bacillus species | [150] |
| SPG24 | ||||
| Rhamnosidase | Sf6 | O-antigen LPS | Shigella flexneri | [151,159] |
| P22 | Rhamnogalacturonan | Salmonella (ser.) Typhimurium | ||
| Cellulases | S6 | Cellulose | Erwinia amylovora | [160] |
| Peptidases | CHAPK | Pentaglycine cross-bridge peptidoglycan | Staphylococcus aureus | [161,162] |
| phiNIT1 | Poly-γ-glutamate | Bacillus spp. | ||
| Lyases | ||||
| Hyaluronidases | Prophages | Hyaluronan | Streptococcus equi | [163,164] |
| H4489A | Streptococcus pyogenes | |||
| Alginate lyases | PT 6 | Alginic acid | P. aeruginosa | [12,65] |
| AF | P. putida | |||
| Pectin/pectate lyases | ΦIPLA7 | Pectin* | Staphylococcal spp. | [165] |
| Others | ||||
| Colanidase | Phi92 | Colonic acid | E. coli | [63] |
| Lipases/triacylglycerol hydrolases | Phi3ST:2 | Carboxyl ester bonds* | Cellulophaga spp. | [166] |
| Tf | Pseudomonas spp. | |||
*The exact role or substrate degraded by the phage-derived depolymerase is yet to be defined.
Researchers are looking into exploiting phage-derived depolymerases to make bacteria less virulent, assist in antibiotic treatment, act as prophylactics on medical devices, and improve immune responses to bacterial infections. There is strong evidence that phage depolymerases have potential as anti-biofilm agents, for example, phage alginate lyases can reduce biofilm formation of P. aeruginosa [12]. Alginate lyases can also improve antibiotic killing of mucoid P. aeruginosa [167]. Removal of the alginic acid EPS matrix is important for antibiotic efficacy as the EPS can block the bioavailability of gentamicin or tobramycin. Furthermore, the biofilm can directly bind aminoglycosides and cationic antibiotics [168,169]. Importantly, the removal of EPS-related virulence factors increases macrophage uptake of bacteria and exposure to immune complement, both contributing to the elimination of bacterial burden during infection [170,171]. Similar anti-virulent agents have been observed in Klebsiella phages producing capsular depolymerases that degrade CPS, reducing virulence of carbapenem-resistant K. pneumoniae and exposing it to serum complement for effective killing [172]. Phage depolymerase-antibiotic combinations have been investigated, where phage depolymerase Dpo71, degraded A. baumannii CPS and reduced biofilm formation. Furthermore, the removal of CPS improved the antibacterial activity of colistin in a Galleria mellonella infection model [173]. Contrary to the success of Chen et al., a similar study performed using a CPS-degrading depolymerase, depoKP36, for K. pneumoniae noted that combination therapy did not improve antibiotic efficiency. Interestingly, no drug interference was observed with antibiotic-depoKP36 combinations. Removing CPS can improve phagocytosis and complement mediated opsonization, therefore further study should account for these immune responses when evaluating the use of phage depolymerases.
Bacteriophages have been investigated for their potential prophylactic use in lining medical equipment, especially catheters. A recent study by Rice et al. [174] found a promising pectate lyase domain containing tail spike protein in a Proteus phage which reduced biofilm formation of P. mirabilis. The authors concluded that such a tail spike could be used for the treatment of catheter-associated UTIs (CAUTIs) and other studies have shown that catheters coated with bacteriophages can prevent biofilm formation [175]. Yet there are very few studies looking into depolymerases derived from phages on their own, most investigated the use of whole phage cocktails. A study by Shahed-Al-Mahmud et al. [176] evaluated the anti-fouling capabilities of a phage tail spike protein against A. baumannii biofilms on catheter sections, however it did not noting inhibited colonization. The therapeutic effect was further evaluated in a zebrafish model which showed the tail spike protein increased survivability of zebrafish by 80% when challenged with A. baumannii. This warrants further investigation into the use of phage-derived depolymerases as prophylactic coatings on medical devices. Clinical trials into the use of phage depolymerase cocktails in combination with antibiotics are important.
8. Limitations of Phage Therapy
A major limitation of the use of phages in the treatment of infections is the possibility of overstimulation of the immune system. Phage capsids, tails, and tail fibers are proteins and are thus recognized by the immune system. Some studies reported no major changes in the immune response of patients treated with phages [2]. These findings are supported by the observation that phages are phagocytosed within a few minutes after administration, at least according to tests conducted on animals and mammalian tissue cells [177]. In immunocompromised mice, phage T7 was eliminated within 60 min after injection [178]. In most cases, treatment with phage T7 did not increase pro-inflammatory cytokines and reactive oxygen species (ROS) and did not damage tissue [179]. Similar studies were reported with phage T4. The intraperitoneal injection of phage T4 head proteins in mice did not stimulate the production of interleukin (IL)-1a, IL-1b, IL-2, IL- 6, IL-10, IL-12 p40/p70, interferon (IFN)-γ, tumour necrosis factor (TNF)-α, monocyte chemoattractant protein (MCP-1), monokine induced by gamma (MIG), RANTES, granulocyte colony-stimulating factor (GCSF), granulocyte-macrophage colony-stimulating factor (GM-CSF) and reactive oxygen species (ROS) [180]. These findings were supported by the findings of Hwang, et al. (2016). [181]. Phages used to treat Burkholderia cenocepacia pulmonary infection in mice controlled the increase in bacterial cell numbers and did not stimulate the production of macrophage inflammatory protein 2 (MIP-2) and TNF-α [182]. Despite findings that the human immune system is not altered by phage therapy, antibodies against E. coli T4 phage were detected in more than 80% of patients who have not been treated with phage T4 [183]. Findings such as these expose our limited knowledge of the interactions between phage and host.
9. Genomic Engineering of Phages
A research field that has been neglected is the genetic engineering of phages. The introduction of rpsL and gyrA in lysogenic phages increased the sensitivity of pathogens towards streptomycin and nalidixic acid, respectively [184]. This approach may be used in the treatment of methicillin-resistant S. aureus, as shown in the treatment of skin infections [185] and bacterial infections associated with wounds [186], burns [187,188], and diabetic leg and foot ulcers [186,189,190,191]. Phages have also been used in the treatment of wound sepsis caused by multidrug-resistant P. aeruginosa [192]. Positive results obtained with phages in the treatment of bacterial skin infections were questioned by data generated from a clinical trial in which phage treatment was compared with sulfadiazine cream [188]. Concluded from this study, the phages used in the treatment were unstable, which resulted in patients receiving 1,000-fold to 10,000-fold fewer phages than initially prescribed.
Genome engineering of phages may be done in several ways and include recombination between phage DNA and plasmids [193], the use of shuttle plasmids [194], cloning of specific genes [195], recombineering [196,197], CRISPR-Cas selection [197], and a combination of recombineering and CRISPR-mediated counter-selection [198]. For a review on the advances of genetically engineered phages, the reviewer is referred to Pires et al. [199].
Although the oral administration of phages prevented cholera [200], randomized controls were not included. In other studies, inconclusive results were obtained when patients with E. coli diarrhea were treated with phage cocktails [201,202,203,204]. An increase in intestinal levels of Streptococcus gallolyticus and Streptococcus salivarius was recorded [201], which may indicate an imbalance in gut microbiota. The authors ascribed these changes to the fact that only 60% of the 120 patients that were enrolled in the trial showed E. coli in their stool and that the population of phages used in the treatment did not increase in the GIT.
Phages may be used to control biofilm formation, as shown with the ability of some phages to produce depolymerases [205]. Recently, three depolymerases were identified in the genome of a Klebsiella pneumoniae phages [206]. The depolymerases destroyed K. pneumoniae capsule serotypes K7, K20, and K27, and revealed promising results in vivo in a Mus musculus survival study. Majkowska-Skrobek et al. [207] showed that a phage depolymerase sensitized K. pneumoniae against serum-mediated killing and phagocytosis. Similar findings were reported for phages used in the treatment of multidrug-resistant Acinetobacter baumannii [173], E. coli [208,209], P. aeruginosa [210] and Proteus mirabilis [174]. Lu and Collins [205] engineered a phage to express biofilm-degrading enzymes and destroy an E. coli biofilm. The engineered phage reduced bacterial biofilm cell counts by approximately 99.9%. In-depth studies on depolymerases produced by phages may lead to the development of vaccines against capsules of antibiotic-resistant bacterial strains.
10. Diagnostic Potential of Phages and Phage-Derived Proteins
The diagnostic potential of phage-derived proteins has been investigated. Bioinformatic tools such as Kaptive was developed to rapidly identify capsule and lipopolysaccharide (K and O) types of K. pneumoniae and A. baumannii [211]. Recently, Vibrio parahaemolyticus was added to this database [212]. Rapid capsule typing tools have several advantages over traditional capsule typing which includes serological reactivity assays or polymerase chain reaction-based sequencing [213]. Serotyping was first introduced in 1926 and has been used ever since to type capsules. For example, capsule serotyping has identified 77 capsule types (K-types) of K. pneumoniae, while genotyping has found 134 KL (K-locus) types [64,213]. Although important, several limitations of serotyping and genotyping have been reported. Limitations include the inability to distinguish between genotypically similar KL-types and expensive and difficulty to produce antisera.
Alternative methods could include using whole phages or phage-derived proteins for capsule typing. Success has been seen in this regard, where whole phages and phamolge-derived proteins have been used not only for capsule and LPS typing but also for other diagnostic purposes. Li et al. (2021) discovered a novel phage, Klebsiella phage SH-KP152410, that specifically recognized the KL64 capsule type of clinical K. pneumoniae strains [214]. The authors also showed that the depolymerase (K64-ORF41) could reliably type K64 (serotyping) and KL64 K. pneumoniae strains in agreement with genotyping and serotyping [214]. They also demonstrated one of the limitations of genotyping where clinical strains were erroneously typed because an insertion occurred in the wcaJ gene sequence which encodes WcaJ responsible for initiation of capsule biosynthesis. Park and Park [215] identified an O-antigen (located in the LPS) active depolymerase, Dpo10, in the genome of an Escherichia siphophage. The authors tested the specific typing capabilities of Dpo10 and showed that the depolymerase only acted upon E. coli O157:H7 strains. This study further substantiates the viability of applying phage depolymerases to specifically type bacterial CPS and LPS. Moreover, several other studies have proven the feasibility of phages and phage-derived proteins in fundamental studies regarding bacterial defense and virulence mechanisms. Dunstan et al. [216] proved that depolymerase can be used to characterize the monomers in the capsule of K. pneumoniae. This fundamental information can be used to generate capsule-specific vaccines. Other articles have used whole phages in combination with nuclear magnetic resonance and mass spectrometry to identify the constituents and structures of capsules.
Reliable, sensitive, and fast detection of nosocomial pathogens such as E. coli, K. pneumoniae, P. mirabilis, and A. baumannii is urgently required [217]. Therefore, multiple efforts have focused on creating point-of-care biosensor devices that specifically recognize pathogens. Chen et al. [217] created a biosensor that could specifically and reliably (100% recognition) detect A. baumannii strains (n = 77). The authors used a heterologously expressed phage-derived receptor binding protein, called Gp50, that showed higher specificity than the whole phage (100% vs 27.3%). They were also able to quantify A. baumannii in colony-forming units per mL and observed 100% recognition even in complex sample matrices.
11. Conclusions
Intact bacteriophages and phage-derived proteins have various advantageous properties when used as therapeutic agents and diagnostic probes. Therapeutically, whole phage preparations have shown promise but with a more personalized medicine approach. This approach requires a rapid pipeline for the isolation, purification, and characterizing of therapeutically suitable phages. Screening for phage resistance must include studies on phage-resistant mutant populations, the antibiotic susceptibility of bacteria, hypovirulence factors, and susceptibility to immune responses. The concept of using phage-derived proteins such as endolysins and depolymerases, alone and in combination with, conventional antibiotics to treat previously antibiotic-resistant bacteria requires further research. Antibiotic treatment could be improved by the simultaneous use of depolymerases that could decapsulate or degrade biofilms, thereby increasing antibiotic bioavailability. This approach could be the answer to the treatment of chronic infections, especially since the development of new antibiotics is an expensive and time-consuming process. Although conceptually promising, the widespread implementation of phage therapy in routine clinical practices is restricted by the lack of safety and efficacy data collected according to clinical trial standards and regulations. Care should be taken when results are interpreted, as clinical studies differ in design and aim, e.g., treatment with single phages vs phage cocktails, routes of administration (intravenous, oral, local, or combined), and treatment in combination with antibiotics.
References
- Wall, S. Prevention of Antibiotic Resistance – an Epidemiological Scoping Review to Identify Research Categories and Knowledge Gaps. Glob Health Action 2019, 12, 1756191. [Google Scholar] [CrossRef]
- Forde, A.; Hill, C. Phages of Life – the Path to Pharma. Br J Pharmacol 2018, 175, 412–418. [Google Scholar] [CrossRef]
- Seed, K.D. Battling Phages: How Bacteria Defend against Viral Attack. PLoS Pathog 2015, 11, e1004847. [Google Scholar] [CrossRef]
- Riede, I.; Eschbach, M.-L. Evidence That TraT Interacts with OmpA of Escherichia Coli. FEBS Lett 1986, 205, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Deora, R.; Doulatov, S.R.; Gingery, M.; Eiserling, F.A.; Preston, A.; Maskell, D.J.; Simons, R.W.; Cotter, P.A.; Parkhill, J.; et al. Reverse Transcriptase-Mediated Tropism Switching in Bordetella Bacteriophage. Science (1979) 2002, 295, 2091–2094. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Bacteriophage Lysins as Effective Antibacterials. Curr Opin Microbiol 2008, 11, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.-N.; Smith, D.L.; Young, R. Holins: The Protein Clocks of Bacteriophage Infections. Annu Rev Microbiol 2000, 54, 799–825. [Google Scholar] [CrossRef] [PubMed]
- Young, R. Phage Lysis: Three Steps, Three Choices, One Outcome. Journal of Microbiology 2014, 52, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Bacteriophage Lysins as Effective Antibacterials. Curr Opin Microbiol 2008, 11, 393. [Google Scholar] [CrossRef] [PubMed]
- Akoh, C.C.; Lee, G.-C.; Liaw, Y.-C.; Huang, T.-H.; Shaw, J.-F. GDSL Family of Serine Esterases/Lipases. Prog Lipid Res 2004, 43, 534–552. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Gupta, N.; Rathi, P. Bacterial Lipases: An Overview of Production, Purification and Biochemical Properties. Appl Microbiol Biotechnol 2004, 64, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Glonti, T.; Chanishvili, N.; Taylor, P.W. Bacteriophage-Derived Enzyme That Depolymerizes the Alginic Acid Capsule Associated with Cystic Fibrosis Isolates of Pseudomonas Aeruginosa. J Appl Microbiol 2010, 108, 695–702. [Google Scholar] [CrossRef]
- Hatfull, G.F.; Hendrix, R.W. Bacteriophages and Their Genomes. Curr Opin Virol 2011, 1, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Demina, T.A.; Roux, S.; Aiewsakun, P.; Kazlauskas, D.; Simmonds, P.; Prangishvili, D.; Oksanen, H.M.; Krupovic, M. Diversity, Taxonomy, and Evolution of Archaeal Viruses of the Class Caudoviricetes. PLoS Biol 2021, 19, e3001442. [Google Scholar] [CrossRef]
- Brüssow, H.; Hendrix, R.W. Phage Genomics. Cell 2002, 108, 13–16. [Google Scholar] [CrossRef]
- Adriaenssens, E.M.; Sullivan, M.B.; Knezevic, P.; van Zyl, L.J.; Sarkar, B.L.; Dutilh, B.E.; Alfenas-Zerbini, P.; Łobocka, M.; Tong, Y.; Brister, J.R.; et al. Taxonomy of Prokaryotic Viruses: 2018-2019 Update from the ICTV Bacterial and Archaeal Viruses Subcommittee. Archives of Virology 2020 165:5 2020, 165, 1253–1260. [Google Scholar] [CrossRef]
- Santamaria, R.M.; Innes, N.P.T.; Machiulskiene, V.; Evans, D.J.P.; Splieth, C.H. Caries Management Strategies for Primary Molars: 1-Yr Randomized Control Trial Results. J Dent Res 2014, 93, 1062–1069. [Google Scholar] [CrossRef]
- Pantůcek, R.; Rosypalová, A.; Doskar, J.; Kailerová, J.; Růzicková, V.; Borecká, P.; Snopková, S.; Horváth, R.; Götz, F.; Rosypal, S. The Polyvalent Staphylococcal Phage Phi 812: Its Host-Range Mutants and Related Phages. Virology 1998, 246, 241–252. [Google Scholar] [CrossRef]
- Cazares, D.; Cazares, A.; Figueroa, W.; Guarneros, G.; Edwards, R.A.; Vinuesa, P. A Novel Group of Promiscuous Podophages Infecting Diverse Gammaproteobacteria from River Communities Exhibits Dynamic Intergenus Host Adaptation. mSystems 2021, 6. [Google Scholar] [CrossRef]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage Diversity, Genomics and Phylogeny. Nat Rev Microbiol 2020, 18, 125–138. [Google Scholar] [CrossRef]
- Hulo, C.; de Castro, E.; Masson, P.; Bougueleret, L.; Bairoch, A.; Xenarios, I.; Le Mercier, P. ViralZone: A Knowledge Resource to Understand Virus Diversity. Nucleic Acids Res 2011, 39, D576–D582. [Google Scholar] [CrossRef] [PubMed]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host Receptors for Bacteriophage Adsorption. FEMS Microbiol Lett 2016, 363, fnw002. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, S.; Tsurumi, S.; Imai, N. Crossover Behavior for Brownian Motion. J Chem Phys 1986, 84, 539–540. [Google Scholar] [CrossRef]
- Heller, K.J.; Schwarz, H.; Tubingen, U.; Mikrobiologie, L.I. Irreversible Binding to the Receptor of Bacteriophages T5 and BF23 Does Not Occur with the Tip of the Tail. J Bacteriol 1985, 162, 621–625. [Google Scholar] [CrossRef]
- Wolf, S.G.; Shimoni, E.; Elbaum, M.; Houben, L. STEM Tomography in Biology. 2018, 33–60. [CrossRef]
- Wang, C.; Tu, J.; Liu, J.; Molineux, I.J. Structural Dynamics of Bacteriophage P22 Infection Initiation Revealed by Cryo-Electron Tomography. Nature Microbiology 2019 4:6 2019, 4, 1049–1056. [Google Scholar] [CrossRef]
- Dunne, M.; Hupfeld, M.; Klumpp, J.; Loessner, M.J. Molecular Basis of Bacterial Host Interactions by Gram-Positive Targeting Bacteriophages. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Kim, M.; Kim, S.; Park, B.; Ryu, S. Core Lipopolysaccharide-Specific Phage SSU5 as an Auxiliary Component of a Phage Cocktail for Salmonella Biocontrol. Appl Environ Microbiol 2014, 80, 1026. [Google Scholar] [CrossRef] [PubMed]
- Sandulache, R.; Prehm, P.; Kamp, D. Cell Wall Receptor for Bacteriophage Mu G(+). J Bacteriol 1984, 160, 299–303. [Google Scholar] [CrossRef]
- Munsch-Alatossava, P.; Alatossava, T. The Extracellular Phage-Host Interactions Involved in the Bacteriophage LL-H Infection of Lactobacillus Delbrueckii Ssp. Lactis ATCC 15808. Front Microbiol 2013, 4. [Google Scholar] [CrossRef]
- Wolin, M.J.; Douglas, L.J. Cell Wall Polymers and Phage Lysis of Lactobacillus Plantarum. Biochemistry 1971, 10, 1551–1555. [Google Scholar] [CrossRef]
- Wendlinger, G.; Loessner, M.J.; Scherer, S. Bacteriophage Receptors on Listeria Monocytogenes Cells Are the N-Acetylglucosamine and Rhamnose Substituents of Teichoic Acids or the Peptidoglycan Itself. Microbiology (N Y) 1996, 142, 985–992. [Google Scholar] [CrossRef]
- Moak, M.; Molineux, I.J. Peptidoglycan Hydrolytic Activities Associated with Bacteriophage Virions. Mol Microbiol 2004, 51, 1169–1183. [Google Scholar] [CrossRef]
- Barbirz, S.; Müller, J.J.; Uetrecht, C.; Clark, A.J.; Heinemann, U.; Seckler, R. Crystal Structure of Escherichia Coli Phage HK620 Tailspike: Podoviral Tailspike Endoglycosidase Modules Are Evolutionarily Related. Mol Microbiol 2008, 69, 303–316. [Google Scholar] [CrossRef]
- Baptista, C.; Santos, M.A.; São-José, C. Phage SPP1 Reversible Adsorption to Bacillus Subtilis Cell Wall Teichoic Acids Accelerates Virus Recognition of Membrane Receptor YueB. J Bacteriol 2008, 190, 4989–4996. [Google Scholar] [CrossRef]
- Xu, J.; Xiang, Y. Membrane Penetration by Bacterial Viruses. J Virol 2017, 91. [Google Scholar] [CrossRef]
- Olsen, R.H.; Siak, J.-S.; Gray, R.H. Characteristics of PRD1, a Plasmid-Dependent Broad Host Range DNA Bacteriophage. J Virol 1974, 14, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Ojala, V.; Laitalainen, J.; Jalasvuori, M. Fight Evolution with Evolution: Plasmid-dependent Phages with a Wide Host Range Prevent the Spread of Antibiotic Resistance. Evol Appl 2013, 6, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, R.A.; Pickard, D.; Dougan, S.; Goulding, D.; Cormie, C.; Hardy, J.; Li, F.; Grinter, R.; Harcourt, K.; Yu, L.; et al. The Flagellotropic Bacteriophage YSD1 Targets Salmonella Typhi with a Chi-like Protein Tail Fibre. Mol Microbiol 2019, 112, 1831–1846. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.M.; Dunstan, R.A.; Grinter, R.; Belousoff, M.J.; Wang, J.; Pickard, D.; Venugopal, H.; Dougan, G.; Lithgow, T.; Coulibaly, F. The Architecture and Stabilisation of Flagellotropic Tailed Bacteriophages. Nature Communications 2020 11:1 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Majkowska-Skrobek, G.; Łątka, A.; Berisio, R.; Maciejewska, B.; Squeglia, F.; Romano, M.; Lavigne, R.; Struve, C.; Drulis-Kawa, Z. Capsule-Targeting Depolymerase, Derived from Klebsiella KP36 Phage, as a Tool for the Development of Anti-Virulent Strategy. Viruses 2016, 8. [Google Scholar] [CrossRef]
- Song, L.; Yang, X.; Huang, J.; Zhu, X.; Han, G.; Wan, Y.; Xu, Y.; Luan, G.; Jia, X. Phage Selective Pressure Reduces Virulence of Hypervirulent Klebsiella Pneumoniae Through Mutation of the Wzc Gene. Front Microbiol 2021, 12, 2904. [Google Scholar] [CrossRef]
- Song, L.; Yang, X.; Huang, J.; Zhu, X.; Han, G.; Wan, Y.; Xu, Y.; Luan, G.; Jia, X. Phage Selective Pressure Reduces Virulence of Hypervirulent Klebsiella Pneumoniae Through Mutation of the Wzc Gene. Front Microbiol 2021, 12. [Google Scholar] [CrossRef]
- Tzagoloff, H.; Pratt, D. The Initial Steps in Infection with Coliphage M13. Virology 1964, 24, 372–380. [Google Scholar] [CrossRef]
- Roncero, C.; Darzins, A.; Casadaban, M.J. Pseudomonas Aeruginosa Transposable Bacteriophages D3112 and B3 Require Pili and Surface Growth for Adsorption. J Bacteriol 1990, 172, 1899–1904. [Google Scholar] [CrossRef]
- Manchak, J.; Anthony. G.; Frost, L.S. Mutational Analysis of F-pilin Reveals Domains for Pilus Assembly, Phage Infection and DNA Transfer. Mol Microbiol 2002, 43, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Jalasvuori, M.; Friman, V.-P.; Nieminen, A.; Bamford, J.K.H.; Buckling, A. Bacteriophage Selection against a Plasmid-Encoded Sex Apparatus Leads to the Loss of Antibiotic-Resistance Plasmids. Biol Lett 2011, 7, 902–905. [Google Scholar] [CrossRef] [PubMed]
- Dunne, M.; Hupfeld, M.; Klumpp, J.; Loessner, M. Molecular Basis of Bacterial Host Interactions by Gram-Positive Targeting Bacteriophages. Viruses 2018, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Kanamaru, S.; Leiman, P.G.; Kostyuchenko, V.A.; Chipman, P.R.; Mesyanzhinov, V. v.; Arisaka, F.; Rossmann, M.G. Structure of the Cell-Puncturing Device of Bacteriophage T4. Nature 2002 415:6871 2002, 415, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Mahony, J.; Alqarni, M.; Stockdale, S.; Spinelli, S.; Feyereisen, M.; Cambillau, C.; van Sinderen, D. Functional and Structural Dissection of the Tape Measure Protein of Lactococcal Phage TP901-1. Scientific Reports 2016 6:1 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, P.; Jacquot, P.; Plançon, L.; Chami, M.; Engel, A.; Parquet, C.; Herbeuval, C.; Letellier, L. Phage T5 Straight Tail Fiber Is a Multifunctional Protein Acting as a Tape Measure and Carrying Fusogenic and Muralytic Activities. Journal of Biological Chemistry 2008, 283, 13556–13564. [Google Scholar] [CrossRef]
- Fokine, A.; Rossmann, M.G. Molecular Architecture of Tailed Double-Stranded DNA Phages. Bacteriophage 2014, 4, e28281. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, R.; Bradley, R.; Paranchych, W. The Effect of M13 Phage Infection upon the F Pili of E. Coli. Virology 1973, 54, 220–229. [Google Scholar] [CrossRef]
- Grahn, A.M.; Daugelavičius, R.; Bamford, D.H. Sequential Model of Phage PRD1 DNA Delivery: Active Involvement of the Viral Membrane. Mol Microbiol 2002, 46, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Daugelavičius, R.; Cvirkaitė, V.; Gaidelytė, A.; Bakienė, E.; Gabrėnaitė-Verkhovskaya, R.; Bamford, D.H. Penetration of Enveloped Double-Stranded RNA Bacteriophages Φ13 and Φ6 into Pseudomonas Syringae Cells. J Virol 2005, 79, 5017–5026. [Google Scholar] [CrossRef] [PubMed]
- Cvirkaitė-Krupovič, V.; Poranen, M.M.; Bamford, D.H. Phospholipids Act as Secondary Receptor during the Entry of the Enveloped, Double-Stranded RNA Bacteriophage Φ6. Journal of General Virology 2010, 91, 2116–2120. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Bossi, N.; Uzzau, S.; Maloriol, D.; Bossi, L. Variable Assortment of Prophages Provides a Transferable Repertoire of Pathogenic Determinants in Salmonella. Mol Microbiol 2001, 39, 260–272. [Google Scholar] [CrossRef]
- McAllister, W.T.; Barrett, C.L. Roles of the Early Genes of Bacteriophage T7 in Shutoff of Host Macromolecular Synthesis. J Virol 1977, 23, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage Resistance Mechanisms. Nat Rev Microbiol 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Scanlan, P.D.; Buckling, A.; Hall, A.R. Experimental Evolution and Bacterial Resistance: (Co)Evolutionary Costs and Trade-Offs as Opportunities in Phage Therapy Research. Bacteriophage 2015, 5, e1050153. [Google Scholar] [CrossRef]
- Meyer, J.R.; Dobias, D.T.; Weitz, J.S.; Barrick, J.E.; Quick, R.T.; Lenski, R.E. Repeatability and Contingency in the Evolution of a Key Innovation in Phage Lambda. Science 2012, 335, 428–432. [Google Scholar] [CrossRef]
- Scholl, D.; Adhya, S.; Merril, C. Escherichia Coli K1’s Capsule Is a Barrier to Bacteriophage T7. Appl Environ Microbiol 2005, 71, 4872–4874. [Google Scholar] [CrossRef]
- Schwarzer, D.; Buettner, F.F.R.; Browning, C.; Nazarov, S.; Rabsch, W.; Bethe, A.; Oberbeck, A.; Bowman, V.D.; Stummeyer, K.; Mühlenhoff, M.; et al. A Multivalent Adsorption Apparatus Explains the Broad Host Range of Phage Phi92: A Comprehensive Genomic and Structural Analysis. J Virol 2012, 86, 10384. [Google Scholar] [CrossRef] [PubMed]
- Patro, L.P.P.; Rathinavelan, T. Targeting the Sugary Armor of Klebsiella Species. Front Cell Infect Microbiol 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, A.; Ceyssens, P.-J.; Krylov, V.N.; Noben, J.-P.; Volckaert, G.; Lavigne, R. Identification of EPS-Degrading Activity within the Tail Spikes of the Novel Pseudomonas Putida Phage AF. Virology 2012, 434, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Yang, R.; Fan, K.; Dong, H.; Gao, C.; Wang, S.; Yu, L.; Cheng, Z.; Lei, L. External Lysis of Escherichia Coli by a Bacteriophage Endolysin Modified with Hydrophobic Amino Acids. AMB Express 2019, 9, 106. [Google Scholar] [CrossRef] [PubMed]
- Samson, J.E.; Magadán, A.H.; Sabri, M.; Moineau, S. Revenge of the Phages: Defeating Bacterial Defences. Nat Rev Microbiol 2013, 11, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Holst Sørensen, M.C.; van Alphen, L.B.; Fodor, C.; Crowley, S.M.; Christensen, B.B.; Szymanski, C.M.; Brøndsted, L. Phase Variable Expression of Capsular Polysaccharide Modifications Allows Campylobacter Jejuni to Avoid Bacteriophage Infection in Chickens. Front Cell Infect Microbiol 2012, 2, 11. [Google Scholar] [CrossRef]
- Sørensen, M.C.H.; van Alphen, L.B.; Fodor, C.; Crowley, S.M.; Christensen, B.B.; Szymanski, C.M.; Brøndsted, L. Phase Variable Expression of Capsular Polysaccharide Modifications Allows Campylobacter Jejuni to Avoid Bacteriophage Infection in Chickens. Front Cell Infect Microbiol 2012, 2. [Google Scholar] [CrossRef]
- Manning, A.J.; Kuehn, M.J. Contribution of Bacterial Outer Membrane Vesicles to Innate Bacterial Defense. BMC Microbiol 2011, 11, 258. [Google Scholar] [CrossRef]
- Bernheim, A.; Sorek, R. The Pan-Immune System of Bacteria: Antiviral Defence as a Community Resource. Nat Rev Microbiol 2020, 18, 113–119. [Google Scholar] [CrossRef]
- Murray, N.E. Immigration Control of DNA in Bacteria: Self versus Non-Self. Microbiology (N Y) 2002, 148, 3–20. [Google Scholar] [CrossRef]
- Auer, B.; Schweiger, M. Evidence That Escherichia Coli Virus T1 Induces a DNA Methyltransferase. J Virol 1984, 49, 588–590. [Google Scholar] [CrossRef]
- Walkinshaw, M.D.; Taylor, P.; Sturrock, S.S.; Atanasiu, C.; Berge, T.; Henderson, R.M.; Edwardson, J.M.; Dryden, D.T.F. Structure of Ocr from Bacteriophage T7, a Protein That Mimics B-Form DNA. Mol Cell 2002, 9, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Dillingham, M.S.; Kowalczykowski, S.C. RecBCD Enzyme and the Repair of Double-Stranded DNA Breaks. Microbiology and Molecular Biology Reviews 2008, 72, 642–671. [Google Scholar] [CrossRef]
- Murphy, K.C. The λ Gam Protein Inhibits RecBCD Binding to DsDNA Ends. J Mol Biol 2007, 371, 19–24. [Google Scholar] [CrossRef]
- d’Adda di Fagagna, F.; Weller, G.R.; Doherty, A.J.; Jackson, S.P. The Gam Protein of Bacteriophage Mu Is an Orthologue of Eukaryotic Ku. EMBO Rep 2003, 4, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Marraffini, L.A. CRISPR-Cas Systems: Prokaryotes Upgrade to Adaptive Immunity. Mol Cell 2014, 54, 234–244. [Google Scholar] [CrossRef]
- Deveau, H.; Barrangou, R.; Garneau, J.E.; Labonté, J.; Fremaux, C.; Boyaval, P.; Romero, D.A.; Horvath, P.; Moineau, S. Phage Response to CRISPR-Encoded Resistance in Streptococcus Thermophilus. J Bacteriol 2008, 190, 1390–1400. [Google Scholar] [CrossRef]
- Seed, K.D.; Lazinski, D.W.; Calderwood, S.B.; Camilli, A. A Bacteriophage Encodes Its Own CRISPR/Cas Adaptive Response to Evade Host Innate Immunity. Nature 2013, 494, 489–491. [Google Scholar] [CrossRef]
- Bondy-Denomy, J.; Pawluk, A.; Maxwell, K.L.; Davidson, A.R. Bacteriophage Genes That Inactivate the CRISPR/Cas Bacterial Immune System. Nature 2013, 493, 429–432. [Google Scholar] [CrossRef]
- Pawluk, A.; Bondy-Denomy, J.; Cheung, V.H.W.; Maxwell, K.L.; Davidson, A.R. A New Group of Phage Anti-CRISPR Genes Inhibits the Type I-E CRISPR-Cas System of Pseudomonas Aeruginosa. mBio 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Durmaz, E.; Klaenhammer, T.R. Abortive Phage Resistance Mechanism AbiZ Speeds the Lysis Clock To Cause Premature Lysis of Phage-Infected Lactococcus Lactis. J Bacteriol 2007, 189, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Shinedling, S.; Parma, D.; Gold, L. Wild-Type Bacteriophage T4 Is Restricted by the Lambda Rex Genes. J Virol 1987, 61, 3790–3794. [Google Scholar] [CrossRef]
- Bingham, R.; Ekunwe, S.I.N.; Falk, S.; Snyder, L.; Kleanthous, C. The Major Head Protein of Bacteriophage T4 Binds Specifically to Elongation Factor Tu. Journal of Biological Chemistry 2000, 275, 23219–23226. [Google Scholar] [CrossRef] [PubMed]
- Essoh, C.; Blouin, Y.; Loukou, G.; Cablanmian, A.; Lathro, S.; Kutter, E.; Thien, H.V.; Vergnaud, G.; Pourcel, C. The Susceptibility of Pseudomonas Aeruginosa Strains from Cystic Fibrosis Patients to Bacteriophages. PLoS One 2013, 8, e60575. [Google Scholar] [CrossRef]
- Saussereau, E.; Vachier, I.; Chiron, R.; Godbert, B.; Sermet, I.; Dufour, N.; Pirnay, J.-P.; De Vos, D.; Carrié, F.; Molinari, N.; et al. Effectiveness of Bacteriophages in the Sputum of Cystic Fibrosis Patients. Clinical Microbiology and Infection 2014, 20, O983–O990. [Google Scholar] [CrossRef]
- Kutateladze, M.; Adamia, R. Phage Therapy Experience at the Eliava Institute. Med Mal Infect 2008, 38, 426–430. [Google Scholar] [CrossRef]
- Leitner, L.; Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Chkonia, I.; Rigvava, S.; Chkhotua, A.; Changashvili, G.; McCallin, S.; Schneider, M.P.; et al. Intravesical Bacteriophages for Treating Urinary Tract Infections in Patients Undergoing Transurethral Resection of the Prostate: A Randomised, Placebo-Controlled, Double-Blind Clinical Trial. Lancet Infect Dis 2021, 21, 427–436. [Google Scholar] [CrossRef]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter Baumannii Infection. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef]
- Chan, B.K.; Turner, P.E.; Kim, S.; Mojibian, H.R.; Elefteriades, J.A.; Narayan, D. Phage Treatment of an Aortic Graft Infected with Pseudomonas Aeruginosa. Evol Med Public Health 2018, 2018, 60–66. [Google Scholar] [CrossRef]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage Selection Restores Antibiotic Sensitivity in MDR Pseudomonas Aeruginosa. Sci Rep 2016, 6, 26717. [Google Scholar] [CrossRef] [PubMed]
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered Bacteriophages for Treatment of a Patient with a Disseminated Drug-Resistant Mycobacterium Abscessus. Nat Med 2019, 25, 730–733. [Google Scholar] [CrossRef] [PubMed]
- Uyttebroek, S.; Chen, B.; Onsea, J.; Ruythooren, F.; Debaveye, Y.; Devolder, D.; Spriet, I.; Depypere, M.; Wagemans, J.; Lavigne, R.; et al. Safety and Efficacy of Phage Therapy in Difficult-to-Treat Infections: A Systematic Review. Lancet Infect Dis 2022, 22, e208–e220. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, Y.; Boyd, E.F. Mobilization of the Vibrio Pathogenicity Island between Vibrio Cholerae Isolates Mediated by CP-T1 Generalized Transduction. FEMS Microbiol Lett 2002, 214, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Brabban, A.D.; Hite, E.; Callaway, T.R. Evolution of Foodborne Pathogens via Temperate Bacteriophage-Mediated Gene Transfer. Foodborne Pathog Dis 2005, 2, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Maiques, E.; Úbeda, C.; Tormo, M.A.; Ferrer, M.D.; Lasa, I.; Novick, R.P.; Penadés, J.R. Role of Staphylococcal Phage and SaPI Integrase in Intra- and Interspecies SaPI Transfer. J Bacteriol 2007, 189, 5608–5616. [Google Scholar] [CrossRef]
- Parsley, L.C.; Consuegra, E.J.; Kakirde, K.S.; Land, A.M.; Harper, W.F.; Liles, M.R. Identification of Diverse Antimicrobial Resistance Determinants Carried on Bacterial, Plasmid, or Viral Metagenomes from an Activated Sludge Microbial Assemblage. Appl Environ Microbiol 2010, 76, 3753–3757. [Google Scholar] [CrossRef]
- Marti, E.; Variatza, E.; Balcázar, J.L. Bacteriophages as a Reservoir of Extended-Spectrum β -Lactamase and Fluoroquinolone Resistance Genes in the Environment. Clinical Microbiology and Infection 2014, 20, O456–O459. [Google Scholar] [CrossRef]
- Rodriguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sànchez-Melsió, A.; Borrego, C.M.; Barceló, D.; Balcázar, J.L. Occurrence of Antibiotics and Antibiotic Resistance Genes in Hospital and Urban Wastewaters and Their Impact on the Receiving River. Water Res 2015, 69, 234–242. [Google Scholar] [CrossRef]
- Subirats, J.; Sànchez-Melsió, A.; Borrego, C.M.; Balcázar, J.L.; Simonet, P. Metagenomic Analysis Reveals That Bacteriophages Are Reservoirs of Antibiotic Resistance Genes. Int J Antimicrob Agents 2016, 48, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Marti, R.; Zurfluh, K.; Hagens, S.; Pianezzi, J.; Klumpp, J.; Loessner, M.J. Long Tail Fibres of the Novel Broad-Host-Range T-Even Bacteriophage S16 Specifically Recognize Salmonella OmpC. Mol. Microbiol. 2013, 87, 818–834. [Google Scholar] [CrossRef]
- Marti, E.; Variatza, E.; Balcázar, J.L. Bacteriophages as a Reservoir of Extended-Spectrum β -Lactamase and Fluoroquinolone Resistance Genes in the Environment. Clinical Microbiology and Infection 2014, 20, O456–O459. [Google Scholar] [CrossRef] [PubMed]
- Quirós, P.; Colomer-Lluch, M.; Martínez-Castillo, A.; Miró, E.; Argente, M.; Jofre, J.; Navarro, F.; Muniesa, M. Antibiotic Resistance Genes in the Bacteriophage DNA Fraction of Human Fecal Samples. Antimicrob Agents Chemother 2014, 58, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Torres-Barceló, C. The Disparate Effects of Bacteriophages on Antibiotic-Resistant Bacteria. Emerging Microbes & Infection 2018, 7, 1–12. [Google Scholar] [CrossRef]
- Colomer-Lluch, M.; Jofre, J.; Muniesa, M. Quinolone Resistance Genes (QnrA and QnrS) in Bacteriophage Particles from Wastewater Samples and the Effect of Inducing Agents on Packaged Antibiotic Resistance Genes. Journal of Antimicrobial Chemotherapy 2014, 69, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Fancello, L.; Desnues, C.; Raoult, D.; Rolain, J.M. Bacteriophages and Diffusion of Genes Encoding Antimicrobial Resistance in Cystic Fibrosis Sputum Microbiota. Journal of Antimicrobial Chemotherapy 2011, 66, 2448–2454. [Google Scholar] [CrossRef] [PubMed]
- Enault, F.; Briet, A.; Bouteille, L.; Roux, S.; Sullivan, M.B.; Petit, M.-A. Phages Rarely Encode Antibiotic Resistance Genes: A Cautionary Tale for Virome Analyses. ISME J 2017, 11, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage Treatment of Human Infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef]
- Cislo, M.; Dabrowski, M.; Weber-Dabrowska, B.; Woyton, A. Bacteriophage Treatment of Suppurative Skin Infections. Arch Immunol Ther Exp (Warsz) 1987, 35, 175–183. [Google Scholar]
- Ioseliani, G.D.; Meladze, G.D.; Chkhetiia, N.S.; Mebuke, M.G.; Kiknadze, N.I. [Use of Bacteriophage and Antibiotics for Prevention of Acute Postoperative Empyema in Chronic Suppurative Lung Diseases]. Grudn Khir 1980, 63–67. [Google Scholar]
- Kochetkova, V.A.; Mamontov, A.S.; Moskovtseva, R.L.; Erastova, E.I.; Trofimov, E.I.; Popov, M.I.; Dzhubalieva, S.K. [Phagotherapy of Postoperative Suppurative-Inflammatory Complications in Patients with Neoplasms]. Sov Med 1989, 23–26. [Google Scholar]
- Kwarciński, W.; Lazarkiewicz, B.; Weber-Dabrowska, B.; Rudnicki, J.; Kamiński, K.; Sciebura, M. [Bacteriophage Therapy in the Treatment of Recurrent Subphrenic and Subhepatic Abscess with Jejunal Fistula after Stomach Resection]. Pol Tyg Lek 1994, 49, 535–535. [Google Scholar]
- Perepanova, T.S.; Darbeeva, O.S.; Kotliarova, G.A.; Kondrat’eva, E.M.; Maiskaia, L.M.; Malysheva, V.F.; Baiguzina, F.A.; Grishkova, N. v. [The Efficacy of Bacteriophage Preparations in Treating Inflammatory Urologic Diseases]. Urol Nefrol (Mosk) 1995, 14–17. [Google Scholar]
- Sakandelidze, V.M. [The Combined Use of Specific Phages and Antibiotics in Different Infectious Allergoses]. Vrach Delo 1991, 60–63. [Google Scholar]
- Strój, L.; Weber-Dabrowska, B.; Partyka, K.; Mulczyk, M.; Wójcik, M. [Successful Treatment with Bacteriophage in Purulent Cerebrospinal Meningitis in a Newborn]. Neurol Neurochir Pol 1999, 33, 693–698. [Google Scholar] [PubMed]
- Sarker, S.A.; Sultana, S.; Reuteler, G.; Moine, D.; Descombes, P.; Charton, F.; Bourdin, G.; McCallin, S.; Ngom-Bru, C.; Neville, T.; et al. Oral Phage Therapy of Acute Bacterial Diarrhea With Two Coliphage Preparations: A Randomized Trial in Children From Bangladesh. EBioMedicine 2016, 4, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Leitner, L.; Ujmajuridze, A.; Chanishvili, N.; Goderdzishvili, M.; Chkonia, I.; Rigvava, S.; Chkhotua, A.; Changashvili, G.; McCallin, S.; Schneider, M.P.; et al. Intravesical Bacteriophages for Treating Urinary Tract Infections in Patients Undergoing Transurethral Resection of the Prostate: A Randomised, Placebo-Controlled, Double-Blind Clinical Trial. Lancet Infect Dis 2021, 21, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; le Floch, R.; et al. Efficacy and Tolerability of a Cocktail of Bacteriophages to Treat Burn Wounds Infected by Pseudomonas Aeruginosa (PhagoBurn): A Randomised, Controlled, Double-Blind Phase 1/2 Trial. Lancet Infect Dis 2019, 19, 35–45. [Google Scholar] [CrossRef]
- NIH, U.S. National Library of medicine Bacteriophage Therapy in First Time Chronic Prosthetic Joint Infections. NCT05269121. Available online: https://clinicaltrials.gov/study/NCT05269121 (accessed on 28 October 2023).
- Long Island University, C.D.E.K. Assessing the Efficacy of Anti-Staphylococcal Phages in the Management of Infected Foot Ulcers in Diabetes. NCT04289948. Available online: https://www.cdek.liu.edu/trial/NCT04289948/ (accessed on 28 October 2023).
- National Library of medicine BacterioPHAGE for Gastrointestinal Health 2 Study. NCT04511221. Available online: https://clinicaltrials.gov/study/NCT04511221 (accessed on 28 October 2023).
- National Library of medicine Existence in the Human Digestive Flora of Phages Able to Prevent the Acquisition of Multiresistant Enterobacteria (PHAGO-BMR). NCT03231267. Available online: https://clinicaltrials.gov/study/NCT03231267 (accessed on 28 October 2023).
- Sybesma, W.; Rohde, C.; Bardy, P.; Pirnay, J.-P.; Cooper, I.; Caplin, J.; Chanishvili, N.; Coffey, A.; De Vos, D.; Scholz, A.; et al. Silk Route to the Acceptance and Re-Implementation of Bacteriophage Therapy—Part II. Antibiotics 2018, 7, 35. [Google Scholar] [CrossRef]
- NIH, U.S. National Library of medicine Bacteriophage Effects on Pseudomonas Aeruginosa (MUCOPHAGES). NCT01818206. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT01818206 (accessed on 28 October 2023).
- NIH, U.S. National Library of medicine PHAGE Study: Bacteriophages as Novel Prebiotics. NCT03269617. Available online: https://clinicaltrials.gov/study/NCT03269617 (accessed on 28 October 2023).
- NIH, U.S. National Library of medicine A Prospective, Randomized, Double-Blind Controlled Study of WPP-201 for the Safety and Efficacy of Treatment of Venous Leg Ulcers (WPP-201). NCT00663091. Available online: https://clinicaltrials.gov/study/NCT00663091 (accessed on 28 October 2023).
- WCG CenterWatch Safety Tolerability and PK of LBP-EC01 in Patients With Lower Urinary Tract Colonization Caused by E. Coli. Available online: https://www.centerwatch.com/clinical-trials/listings/234324/safety-tolerability-and-pk-of-lbp-ec01-in-patients-with-lower-urinary-tract-colonization-caused-by-e-coli/ (accessed on 28 October 2023).
- Lenneman, B.R.; Fernbach, J.; Loessner, M.J.; Lu, T.K.; Kilcher, S. Enhancing Phage Therapy through Synthetic Biology and Genome Engineering. Curr Opin Biotechnol 2021, 68, 151–159. [Google Scholar] [CrossRef]
- NIH, U.S. National Library of medicine Ascending Dose Study of the Safety of AB-SA01 When Topically Applied to Intact Skin of Healthy Adults. NCT02757755. Available online: https://clinicaltrials.gov/study/NCT02757755 (accessed on 28 October 2023).
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and Use of Personalized Bacteriophage-Based Therapeutic Cocktails To Treat a Patient with a Disseminated Resistant Acinetobacter Baumannii Infection. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef]
- Duplessis, C.A.; Stockelman, M.; Hamilton, T.; Merril, G.; Brownstein, M.; Bishop-Lilly, K.; Schooley, R.; Henry, M.; Horne, B.; Sisson, B.M.; et al. A Case Series of Emergency Investigational New Drug Applications for Bacteriophages Treating Recalcitrant Multi-Drug Resistant Bacterial Infections: Confirmed Safety and a Signal of Efficacy. J. Intensive Crit. Care 2019, 5. [Google Scholar]
- Aslam, S.; Courtwright, A.M.; Koval, C.; Lehman, S.M.; Morales, S.; Furr, C.L.L.; Rosas, F.; Brownstein, M.J.; Fackler, J.R.; Sisson, B.M.; et al. Early Clinical Experience of Bacteriophage Therapy in 3 Lung Transplant Recipients. Am J Transplant 2019, 19, 2631–2639. [Google Scholar] [CrossRef]
- Nir-Paz, R.; Gelman, D.; Khouri, A.; Sisson, B.M.; Fackler, J.; Alkalay-Oren, S.; Khalifa, L.; Rimon, A.; Yerushalmy, O.; Bader, R.; et al. Successful Treatment of Antibiotic-Resistant, Poly-Microbial Bone Infection With Bacteriophages and Antibiotics Combination. Clin Infect Dis 2019, 69, 2015–2018. [Google Scholar] [CrossRef]
- Gainey, A.B.; Burch, A.K.; Brownstein, M.J.; Brown, D.E.; Fackler, J.; Horne, B.; Biswas, B.; Bivens, B.N.; Malagon, F.; Daniels, R. Combining Bacteriophages with Cefiderocol and Meropenem/Vaborbactam to Treat a Pan-Drug Resistant Achromobacter Species Infection in a Pediatric Cystic Fibrosis Patient. Pediatr Pulmonol 2020, 55, 2990–2994. [Google Scholar] [CrossRef]
- LeBlanc, L.; Nezami, S.; Yost, D.; Tsourkas, P.; Amy, P.S. Isolation and Characterization of a Novel Phage Lysin Active against Paenibacillus Larvae, a Honeybee Pathogen. 2015, 5, e1080787. [Google Scholar] [CrossRef]
- Bateman, A.; Rawlings, N.D. The CHAP Domain: A Large Family of Amidases Including GSP Amidase and Peptidoglycan Hydrolases. Trends Biochem Sci 2003, 28, 234–237. [Google Scholar] [CrossRef]
- Oliveira, H.; Boas, D.V.; Mesnage, S.; Kluskens, L.D.; Lavigne, R.; Sillankorva, S.; Secundo, F.; Azeredo, J. Structural and Enzymatic Characterization of ABgp46, a Novel Phage Endolysin with Broad Anti-Gram-Negative Bacterial Activity. Front Microbiol 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Lood, R.; Winer, B.Y.; Pelzek, A.J.; Diez-Martinez, R.; Thandar, M.; Euler, C.W.; Schuch, R.; Fischetti, V.A. Novel Phage Lysin Capable of Killing the Multidrug-Resistant Gram-Negative Bacterium Acinetobacter Baumannii in a Mouse Bacteremia Model. Antimicrob Agents Chemother 2015, 59, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.A.; Shin, H.; Heu, S.; Ryu, S. Exogenous Lytic Activity of SPN9CC Endolysin against Gram-Negative Bacteria. J Microbiol Biotechnol 2014, 24, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Djurkovic, N.; McCormack, D.; Casimir, G. The Behavioral Reactions of Victims to Different Types of Workplace Bullying. International Journal of Organization Theory & Behavior 2005, 8, 439–460. [Google Scholar] [CrossRef]
- Fowler, V.G.; Das, A.F.; Lipka-Diamond, J.; Schuch, R.; Pomerantz, R.; Jáuregui-Peredo, L.; Bressler, A.; Evans, D.; Moran, G.J.; Rupp, M.E.; et al. Exebacase for Patients with Staphylococcus Aureus Bloodstream Infection and Endocarditis. J Clin Invest 2020, 130, 3750–3760. [Google Scholar] [CrossRef]
- Davies, G.; Henrissat, B. Structures and Mechanisms of Glycosyl Hydrolases. Structure 1995, 3, 853–859. [Google Scholar] [CrossRef]
- Severi, E.; Hood, D.W.; Thomas, G.H. Sialic Acid Utilization by Bacterial Pathogens. Microbiology (N Y) 2007, 153, 2817–2822. [Google Scholar] [CrossRef]
- Tomlinson, S.; Taylor, P.W. Neuraminidase Associated with Coliphage E That Specifically Depolymerizes the Escherichia Coli K1 Capsular Polysaccharide. J Virol 1985, 55, 374–378. [Google Scholar] [CrossRef]
- Schwarzer, D.; Browning, C.; Stummeyer, K.; Oberbeck, A.; Mühlenhoff, M.; Gerardy-Schahn, R.; Leiman, P.G. Structure and Biochemical Characterization of Bacteriophage Phi92 Endosialidase. Virology 2015, 477, 133–143. [Google Scholar] [CrossRef]
- Murakami, H.; Kuramoto, T.; Mizutani, K.; Nakano, H.; Kitahata, S. Purification and Some Properties of a New Levanase from Bacillus Sp. No. 71. Biosci Biotechnol Biochem 1992, 56, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Jathore, N.R.; Bule, M.V.; Tilay, A.V.; Annapure, U.S. Microbial Levan from Pseudomonas Fluorescens: Characterization and Medium Optimization for Enhanced Production. Food Sci Biotechnol 2012, 21, 1045–1053. [Google Scholar] [CrossRef]
- Dogsa, I.; Brloznik, M.; Stopar, D.; Mandic-Mulec, I. Exopolymer Diversity and the Role of Levan in Bacillus Subtilis Biofilms. PLoS One 2013, 8, e62044. [Google Scholar] [CrossRef]
- Maaroufi, H.; Levesque, R.C. Glycoside Hydrolase Family 32 Is Present in Bacillus Subtilis Phages. Virol J 2015, 12, 157. [Google Scholar] [CrossRef] [PubMed]
- Iwashita, S.; Kanegasaki, S. Smooth Specific Phage Adsorption: Endorhamnosidase Activity of Tail Parts of P22. Biochem Biophys Res Commun 1973, 55, 403–409. [Google Scholar] [CrossRef]
- León, M.; Bastías, R. Virulence Reduction in Bacteriophage Resistant Bacteria. Front Microbiol 2015, 06. [Google Scholar] [CrossRef]
- Thurow, H.; Niemann, H.; Rudolph, C.; Stirm, S. Host Capsule Depolymerase Activity of Bacteriophage Particles Active on Klebsiella K20 and K24 Strains. Virology 1974, 58, 306–309. [Google Scholar] [CrossRef]
- Hanna, A.; Berg, M.; Stout, V.; Razatos, A. Role of Capsular Colanic Acid in Adhesion of Uropathogenic Escherichia Coli. Appl Environ Microbiol 2003, 69, 4474. [Google Scholar] [CrossRef]
- Danese, P.N.; Pratt, L.A.; Kolter, R. Exopolysaccharide Production Is Required for Development of Escherichia Coli K-12 Biofilm Architecture. J Bacteriol 2000, 182, 3593–3596. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Prokhorov, N.S.; Shashkov, A.S.; Ovchinnikova, O.G.; Zdorovenko, E.L.; Liu, B.; Kostryukova, E.S.; Larin, A.K.; Golomidova, A.K.; Letarov, A. v. Variations in O-Antigen Biosynthesis and O-Acetylation Associated with Altered Phage Sensitivity in Escherichia Coli 4s. J Bacteriol 2015, 197, 905–912. [Google Scholar] [CrossRef]
- Thompson, J.E.; Pourhossein, M.; Waterhouse, A.; Hudson, T.; Goldrick, M.; Derrick, J.P.; Roberts, I.S. The K5 Lyase KflA Combines a Viral Tail Spike Structure with a Bacterial Polysaccharide Lyase Mechanism. Journal of Biological Chemistry 2010, 285, 23963–23969. [Google Scholar] [CrossRef] [PubMed]
- Hallenbeck, P.C.; Vimr, E.R.; Yu, F.; Bassler, B.; Troy, F.A. Purification and Properties of a Bacteriophage-Induced Endo-N-Acetylneuraminidase Specific for Poly-Alpha-2,8-Sialosyl Carbohydrate Units. Journal of Biological Chemistry 1987, 262, 3553–3561. [Google Scholar] [CrossRef]
- Freiberg, A.; Morona, R.; Van Den Bosch, L.; Jung, C.; Behlke, J.; Carlin, N.; Seckler, R.; Baxa, U. The Tailspike Protein of Shigella Phage Sf6. Journal of Biological Chemistry 2003, 278, 1542–1548. [Google Scholar] [CrossRef] [PubMed]
- Knecht, L.E.; Heinrich, N.; Born, Y.; Felder, K.; Pelludat, C.; Loessner, M.J.; Fieseler, L. Bacteriophage S6 Requires Bacterial Cellulose for Erwinia Amylovora Infection. Environ Microbiol 2022. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Itoh, Y. Characterization of Poly-γ-Glutamate Hydrolase Encoded by a Bacteriophage Genome: Possible Role in Phage Infection of Bacillus Subtilis Encapsulated with Poly-γ-Glutamate. Appl Environ Microbiol 2003, 69, 2491–2497. [Google Scholar] [CrossRef]
- Fenton, M.; Keary, R.; McAuliffe, O.; Ross, R.P.; O’Mahony, J.; Coffey, A. Bacteriophage-Derived Peptidase Eliminates and Prevents Staphylococcal Biofilms. Int J Microbiol 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Bharati, A.P.; Singh, N.; Pandey, P.; Joshi, P.; Singh, K.; Mitra, K.; Gayen, J.R.; Sarkar, J.; Akhtar, Md.S. The Prophage-Encoded Hyaluronate Lyase Has Broad Substrate Specificity and Is Regulated by the N-Terminal Domain. Journal of Biological Chemistry 2014, 289, 35225–35236. [Google Scholar] [CrossRef] [PubMed]
- BAKER, J.R.; DONG, S.; PRITCHARD, D.G. The Hyaluronan Lyase of Streptococcus Pyogenes Bacteriophage H4489A. Biochemical Journal 2002, 365, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Briers, Y.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Role of the Pre-Neck Appendage Protein (Dpo7) from Phage VB_SepiS-PhiIPLA7 as an Anti-Biofilm Agent in Staphylococcal Species. Front Microbiol 2015, 6, 1315. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-Encoded Depolymerases: Their Diversity and Biotechnological Applications. Appl Microbiol Biotechnol 2016, 100, 2141–2151. [Google Scholar] [CrossRef]
- ALKAWASH, M.A.; SOOTHILL, J.S.; SCHILLER, N.L. Alginate Lyase Enhances Antibiotic Killing of Mucoid Pseudomonas Aeruginosa in Biofilms. APMIS 2006, 114, 131–138. [Google Scholar] [CrossRef]
- Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.L.; Parsek, M.R. The Pel Polysaccharide Can Serve a Structural and Protective Role in the Biofilm Matrix of Pseudomonas Aeruginosa. PLoS Pathog 2011, 7, e1001264. [Google Scholar] [CrossRef]
- Bansal, S.; Harjai, K.; Chhibber, S. Depolymerase Improves Gentamicin Efficacy during Klebsiella Pneumoniae Induced Murine Infection. BMC Infect. Dis 2014, 14, 456. [Google Scholar] [CrossRef]
- Domenech, M.; Ramos-Sevillano, E.; García, E.; Moscoso, M.; Yuste, J. Biofilm Formation Avoids Complement Immunity and Phagocytosis of Streptococcus Pneumoniae. Infect Immun 2013, 81, 2606–2615. [Google Scholar] [CrossRef]
- Eftekhar, F.; Speert, D.P. Alginase Treatment of Mucoid Pseudomonas Aeruginosa Enhances Phagocytosis by Human Monocyte-Derived Macrophages. Infect Immun 1988, 56, 2788–2793. [Google Scholar] [CrossRef]
- Liu, Y.; Leung, S.S.Y.; Huang, Y.; Guo, Y.; Jiang, N.; Li, P.; Chen, J.; Wang, R.; Bai, C.; Mi, Z.; et al. Identification of Two Depolymerases From Phage IME205 and Their Antivirulent Functions on K47 Capsule of Klebsiella Pneumoniae. Front Microbiol 2020, 11, 218. [Google Scholar] [CrossRef]
- Chen, X.; Liu, M.; Zhang, P.; Xu, M.; Yuan, W.; Bian, L.; Liu, Y.; Xia, J.; Leung, S.S.Y. Phage-Derived Depolymerase as an Antibiotic Adjuvant Against Multidrug-Resistant Acinetobacter Baumannii. Front Microbiol 2022, 13. [Google Scholar] [CrossRef]
- Rice, C.J.; Kelly, S.A.; O’Brien, S.C.; Melaugh, E.M.; Ganacias, J.C.B.; Chai, Z.H.; Gilmore, B.F.; Skvortsov, T. Novel Phage-Derived Depolymerase with Activity against Proteus Mirabilis Biofilms. Microorganisms 2021, 9, 2172. [Google Scholar] [CrossRef]
- Melo, L.D.R.; Veiga, P.; Cerca, N.; Kropinski, A.M.; Almeida, C.; Azeredo, J.; Sillankorva, S. Development of a Phage Cocktail to Control Proteus Mirabilis Catheter-Associated Urinary Tract Infections. Front Microbiol 2016, 0, 1024. [Google Scholar] [CrossRef]
- Shahed-Al-Mahmud, Md.; Roy, R.; Sugiokto, F.G.; Islam, Md.N.; Lin, M.-D.; Lin, L.-C.; Lin, N.-T. Phage ΦAB6-Borne Depolymerase Combats Acinetobacter Baumannii Biofilm Formation and Infection. Antibiotics 2021, 10, 279. [Google Scholar] [CrossRef]
- Kaźmierczak, Z.; Piotrowicz, A.; Owczarek, B.; Hodyra, K.; Miernikiewicz, P.; Lecion, D.; Harhala, M.; Górski, A.; Dąbrowska, K. Molecular Imaging of T4 Phage in Mammalian Tissues and Cells. Bacteriophage 2014, 4, e28364. [Google Scholar] [CrossRef]
- Srivastava, A.S.; Kaido, T.; Carrier, E. Immunological Factors That Affect the in Vivo Fate of T7 Phage in the Mouse. J Virol Methods 2004, 115, 99–104. [Google Scholar] [CrossRef]
- Park, K.; Cha, K.E.; Myung, H. Observation of Inflammatory Responses in Mice Orally Fed with Bacteriophage T7. J Appl Microbiol 2014, 117, 627–633. [Google Scholar] [CrossRef]
- Miernikiewicz, P.; Dąbrowska, K.; Piotrowicz, A.; Owczarek, B.; Wojas-Turek, J.; Kicielińska, J.; Rossowska, J.; Pajtasz-Piasecka, E.; Hodyra, K.; Macegoniuk, K.; et al. T4 Phage and Its Head Surface Proteins Do Not Stimulate Inflammatory Mediator Production. PLoS One 2013, 8, e71036. [Google Scholar] [CrossRef]
- Hwang, J.-Y.; Kim, J.-E.; Song, Y.-J.; Park, J.-H. Safety of Using Escherichia Coli Bacteriophages as a Sanitizing Agent Based on Inflammatory Responses in Rats. Food Sci Biotechnol 2016, 25, 355–360. [Google Scholar] [CrossRef]
- Carmody, L.A.; Gill, J.J.; Summer, E.J.; Sajjan, U.S.; Gonzalez, C.F.; Young, R.F.; LiPuma, J.J. Efficacy of Bacteriophage Therapy in a Model of Burkholderia Cenocepacia Pulmonary Infection. J Infect Dis 2010, 201, 264–271. [Google Scholar] [CrossRef]
- Dąbrowska, K.; Miernikiewicz, P.; Piotrowicz, A.; Hodyra, K.; Owczarek, B.; Lecion, D.; Kaźmierczak, Z.; Letarov, A.; Górski, A. Immunogenicity Studies of Proteins Forming the T4 Phage Head Surface. J Virol 2014, 88, 12551–12557. [Google Scholar] [CrossRef]
- Edgar, R.; Friedman, N.; Molshanski-Mor, S.; Qimron, U. Reversing Bacterial Resistance to Antibiotics by Phage-Mediated Delivery of Dominant Sensitive Genes. Appl Environ Microbiol 2012, 78, 744–751. [Google Scholar] [CrossRef]
- Międzybrodzki, R.; Fortuna, W.; Weber-Dąbrowska, B.; Górski, A. Phage Therapy of Staphylococcal Infections (Including MRSA) May Be Less Expensive than Antibiotic Treatment Word Count: Coresponding; 2007; Vol. 61.
- Morozova, V.V.; Vlassov, V.V.; Tikunova, N.V. Applications of Bacteriophages in the Treatment of Localized Infections in Humans. Front Microbiol 2018, 9. [Google Scholar] [CrossRef]
- Rose, T.; Verbeken, G.; Vos, D. De; Merabishvili, M.; Vaneechoutte, M.; Lavigne, R.; Jennes, S.; Zizi, M.; Pirnay, J.-P. Experimental Phage Therapy of Burn Wound Infection: Difficult First Steps. Int J Burns Trauma 2014, 4, 66–73. [Google Scholar]
- Jault, P.; Leclerc, T.; Jennes, S.; Pirnay, J.P.; Que, Y.-A.; Resch, G.; Rousseau, A.F.; Ravat, F.; Carsin, H.; Le Floch, R.; et al. Efficacy and Tolerability of a Cocktail of Bacteriophages to Treat Burn Wounds Infected by Pseudomonas Aeruginosa (PhagoBurn): A Randomised, Controlled, Double-Blind Phase 1/2 Trial. Lancet Infect Dis 2019, 19, 35–45. [Google Scholar] [CrossRef]
- Rhoads, D.D.; Wolcott, R.D.; Kuskowski, M.A.; Wolcott, B.M.; Ward, L.S.; Sulakvelidze, A. Bacteriophage Therapy of Venous Leg Ulcers in Humans: Results of a Phase I Safety Trial. J Wound Care 2009, 18, 237–243. [Google Scholar] [CrossRef]
- Markoishvili, K.; Tsitlanadze, G.; Katsarava, R.; Glenn, J.; Morris, M.D., Jr.; Sulakvelidze, A. A Novel Sustained-release Matrix Based on Biodegradable Poly(Ester Amide)s and Impregnated with Bacteriophages and an Antibiotic Shows Promise in Management of Infected Venous Stasis Ulcers and Other Poorly Healing Wounds. Int J Dermatol 2002, 41, 453–458. [Google Scholar] [CrossRef]
- Fish, R.; Kutter, E.; Wheat, G.; Blasdel, B.; Kutateladze, M.; Kuhl, S. Compassionate Use of Bacteriophage Therapy for Foot Ulcer Treatment as an Effective Step for Moving Toward Clinical Trials. In; 2018; pp. 159–170. [CrossRef]
- Abdul-Hassan, H.S.; El-Tahan, E.; Massoud, B.; Gomaa, R. Bacteriophage Therapy of Pseudomonas Burn Wound Sepsis. Ann. Medit. Burn Club 1990, 3, 262–264. [Google Scholar]
- Sarkis, G.J.; Jacobs, W.R.; Hatfulll, G.F. L5 Luciferase Reporter Mycobacteriophages: A Sensitive Tool for the Detection and Assay of Live Mycobacteria. Mol Microbiol 1995, 15, 1055–1067. [Google Scholar] [CrossRef]
- Jacobs, W.R.; Tuckman, M.; Bloom, B.R. Introduction of Foreign DNA into Mycobacteria Using a Shuttle Phasmid. Nature 1987, 327, 532–535. [Google Scholar] [CrossRef]
- Chauthaiwale, V.M.; Therwath, A.; Deshpande, V. V Bacteriophage Lambda as a Cloning Vector. Microbiol Rev 1992, 56, 577–591. [Google Scholar] [CrossRef]
- Marinelli, L.J.; Piuri, M.; Swigoňová, Z.; Balachandran, A.; Oldfield, L.M.; van Kessel, J.C.; Hatfull, G.F. BRED: A Simple and Powerful Tool for Constructing Mutant and Recombinant Bacteriophage Genomes. PLoS One 2008, 3, e3957. [Google Scholar] [CrossRef]
- Hatoum-Aslan, A. Phage Genetic Engineering Using CRISPR–Cas Systems. Viruses 2018, 10, 335. [Google Scholar] [CrossRef]
- Wetzel, K.S.; Guerrero-Bustamante, C.A.; Dedrick, R.M.; Ko, C.-C.; Freeman, K.G.; Aull, H.G.; Divens, A.M.; Rock, J.M.; Zack, K.M.; Hatfull, G.F. CRISPY-BRED and CRISPY-BRIP: Efficient Bacteriophage Engineering. Sci Rep 2021, 11, 6796. [Google Scholar] [CrossRef]
- Pires, D.P.; Cleto, S.; Sillankorva, S.; Azeredo, J.; Lu, T.K. Genetically Engineered Phages: A Review of Advances over the Last Decade. Microbiology and Molecular Biology Reviews 2016, 80, 523–543. [Google Scholar] [CrossRef]
- Summers, W.C. Cholera and Plague in India: The Bacteriophage Inquiry of 1927–1936. J Hist Med Allied Sci 1993, 48, 275–301. [Google Scholar] [CrossRef]
- Sarker, S.A.; Sultana, S.; Reuteler, G.; Moine, D.; Descombes, P.; Charton, F.; Bourdin, G.; McCallin, S.; Ngom-Bru, C.; Neville, T.; et al. Oral Phage Therapy of Acute Bacterial Diarrhea With Two Coliphage Preparations: A Randomized Trial in Children From Bangladesh. EBioMedicine 2016, 4, 124–137. [Google Scholar] [CrossRef]
- Sarker, S.A.; McCallin, S.; Barretto, C.; Berger, B.; Pittet, A.-C.; Sultana, S.; Krause, L.; Huq, S.; Bibiloni, R.; Bruttin, A.; et al. Oral T4-like Phage Cocktail Application to Healthy Adult Volunteers from Bangladesh. Virology 2012, 434, 222–232. [Google Scholar] [CrossRef]
- McCallin, S.; Alam Sarker, S.; Barretto, C.; Sultana, S.; Berger, B.; Huq, S.; Krause, L.; Bibiloni, R.; Schmitt, B.; Reuteler, G.; et al. Safety Analysis of a Russian Phage Cocktail: From MetaGenomic Analysis to Oral Application in Healthy Human Subjects. Virology 2013, 443, 187–196. [Google Scholar] [CrossRef]
- Bourdin, G.; Schmitt, B.; Marvin Guy, L.; Germond, J.-E.; Zuber, S.; Michot, L.; Reuteler, G.; Brüssow, H. Amplification and Purification of T4-Like Escherichia Coli Phages for Phage Therapy: From Laboratory to Pilot Scale. Appl Environ Microbiol 2014, 80, 1469–1476. [Google Scholar] [CrossRef]
- Lu, T.K.; Collins, J.J. Dispersing Biofilms with Engineered Enzymatic Bacteriophage. Proceedings of the National Academy of Sciences 2007, 104, 11197–11202. [Google Scholar] [CrossRef]
- Wu, J.W.; Wang, J.T.; Lin, T.L.; Liu, Y.Z.; Wu, L.T.; Pan, Y.J. Identification of Three Capsule Depolymerases in a Bacteriophage Infecting Klebsiella Pneumoniae Capsular Types K7, K20, and K27 and Therapeutic Application. J Biomed Sci 2023, 30. [Google Scholar] [CrossRef]
- Majkowska-Skrobek, G.; Latka, A.; Berisio, R.; Squeglia, F.; Maciejewska, B.; Briers, Y.; Drulis-Kawa, Z. Phage-Borne Depolymerases Decrease Klebsiella Pneumoniae Resistance to Innate Defense Mechanisms. Front Microbiol 2018, 9. [Google Scholar] [CrossRef]
- Lin, H.; Paff, M.L.; Molineux, I.J.; Bull, J.J. Therapeutic Application of Phage Capsule Depolymerases against K1, K5, and K30 Capsulated E. Coli in Mice. Front Microbiol 2017, 8. [Google Scholar] [CrossRef]
- Chen, Y.; Li, X.; Wang, S.; Guan, L.; Li, X.; Hu, D.; Gao, D.; Song, J.; Chen, H.; Qiana, P. A Novel Tail-Associated O91-Specific Polysaccharide Depolymerase from a Podophage Reveals Lytic Efficacy of Shiga Toxin-Producing Escherichia Coli. Appl Environ Microbiol 2020, 86. [Google Scholar] [CrossRef]
- Mi, L.; Liu, Y.; Wang, C.; He, T.; Gao, S.; Xing, S.; Huang, Y.; Fan, H.; Zhang, X.; Yu, W.; et al. Identification of a Lytic Pseudomonas Aeruginosa Phage Depolymerase and Its Anti-Biofilm Effect and Bactericidal Contribution to Serum. Virus Genes 2019, 55, 394–405. [Google Scholar] [CrossRef]
- Wyres, K.L.; Wick, R.R.; Gorrie, C.; Jenney, A.; Follador, R.; Thomson, N.R.; Holt, K.E. Identification of Klebsiella Capsule Synthesis Loci from Whole Genome Data. Microb Genom 2016, 2, e000102. [Google Scholar] [CrossRef]
- van der Graaf-van Bloois, L.; Chen, H.; Wagenaar, J.A.; Zomer, A.L. Development of Kaptive Databases for Vibrio Parahaemolyticus O- and K-Antigen Genotyping. Microb Genom 2023, 9. [Google Scholar] [CrossRef]
- Pan, Y.-J.; Lin, T.-L.; Chen, C.-T.; Chen, Y.-Y.; Hsieh, P.-F.; Hsu, C.-R.; Wu, M.-C.; Wang, J.-T. Genetic Analysis of Capsular Polysaccharide Synthesis Gene Clusters in 79 Capsular Types of Klebsiella Spp. Sci Rep 2015, 5, 15573. [Google Scholar] [CrossRef]
- Li, J.; Sheng, Y.; Ma, R.; Xu, M.; Liu, F.; Qin, R.; Zhu, M.; Zhu, X.; He, P. Identification of a Depolymerase Specific for K64-Serotype Klebsiella Pneumoniae: Potential Applications in Capsular Typing and Treatment. Antibiotics 2021, 10, 144. [Google Scholar] [CrossRef]
- Park, D.-W.; Park, J.-H. Characterization of a Novel Phage Depolymerase Specific to Escherichia Coli O157:H7 and Biofilm Control on Abiotic Surfaces. J. Microbiol. 2021, 59, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Dunstan, R.A.; Bamert, R.S.; Belousoff, M.J.; Short, F.L.; Barlow, C.K.; Pickard, D.J.; Wilksch, J.J.; Schittenhelm, R.B.; Strugnell, R.A.; Dougan, G.; et al. Mechanistic Insights into the Capsule-Targeting Depolymerase from a Klebsiella Pneumoniae Bacteriophage. Microbiol. Spectr. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, H.; Luo, S.; Wang, L.; Lu, S.; Fu, Z. Engineering Phage Tail Fiber Protein as a Wide-Spectrum Probe for Acinetobacter Baumannii Strains with a Recognition Rate of 100%. Anal Chem 2022. [Google Scholar] [CrossRef] [PubMed]
Figure 2.
(A) T4 phages attach to the surface of E. coli [25]. (B) A cryo-electron microscopy image of P22 phage forming a trans-envelope channel across the cell envelope [26]. (C) A generalized representation of a tailed phage with primary receptors on long tail fibers. Irreversible binding of the central distal tail components punctures the host cell membrane, and the viral genome is released [22].
Figure 2.
(A) T4 phages attach to the surface of E. coli [25]. (B) A cryo-electron microscopy image of P22 phage forming a trans-envelope channel across the cell envelope [26]. (C) A generalized representation of a tailed phage with primary receptors on long tail fibers. Irreversible binding of the central distal tail components punctures the host cell membrane, and the viral genome is released [22].
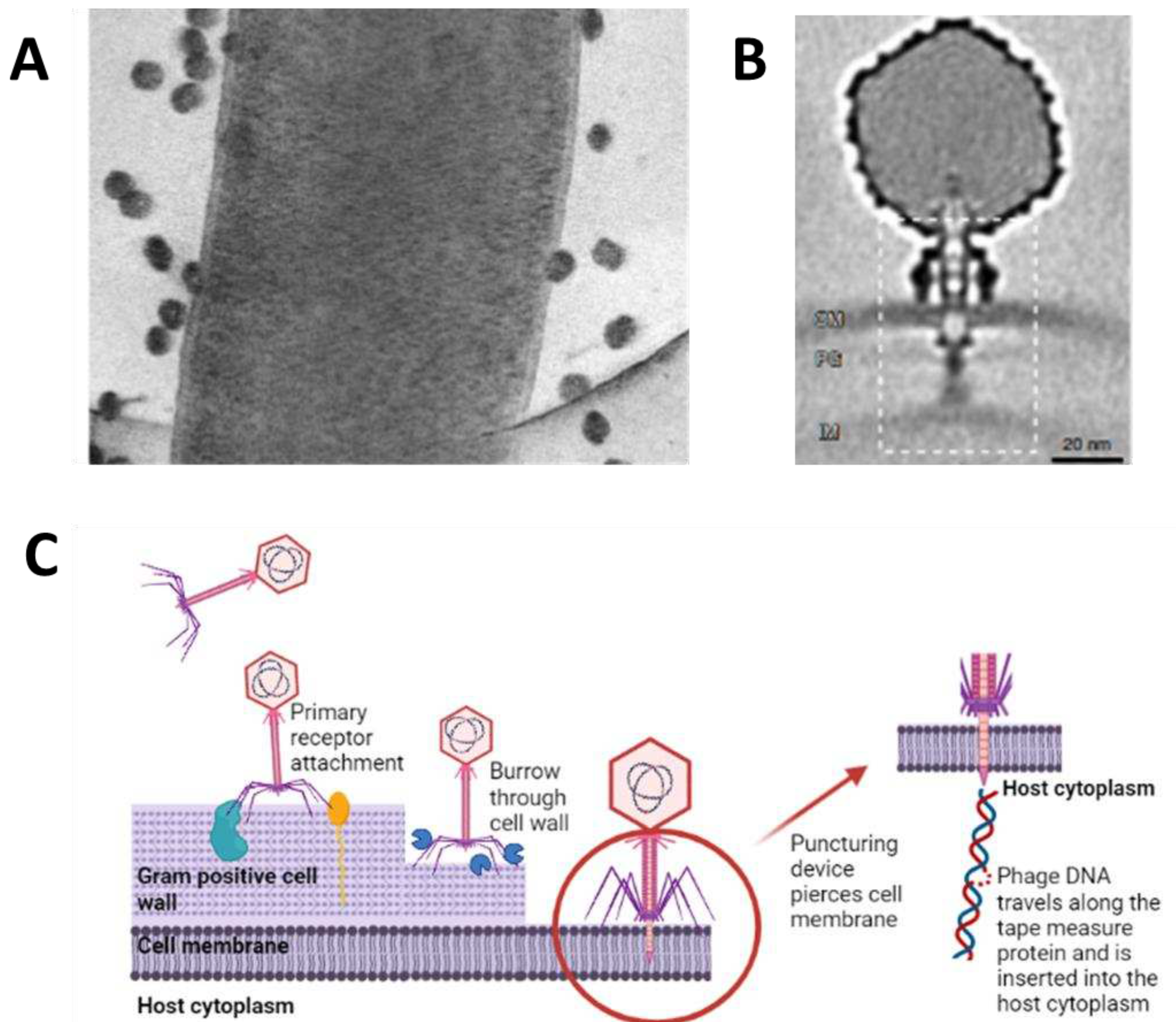
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



