Submitted:
31 October 2023
Posted:
01 November 2023
You are already at the latest version
Abstract
. Gut microbiome reflects healthy and predicts possible disease in hosts. A holistic view of this community is needed, focused on identification species, and on the dissection of how species interact with their host and with each other, regardless of whether their presence is beneficial, inconsequential, or detrimental. Distribution of gut-associated eukaryotes within and across NHPs is likely driven by host behavior and ecology. To ascertain the existence of free-living amoebae (FLA) in the gut of wild and captive non-human primates (NHPs), 01 sample stools were collected and submitted to culture-dependent and microscopy examination and DNA sequencing. FLAs were detected in 45.4% (46/101) of fecal samples analysed and their morphological characteristics matching those of Acanthamoeba spp, Vermamoeba spp, and fan-shaped amoebae of the family Vannellidae. Sequence analysis of the PCR- products revealed that the suspected amoebae are highly homologous (99% identity and 100% query coverage) with Acanthamoeba genotype T4 and Vermamoeba vermiformis amoebae. The results showed a great diversity of FLAs in the NHP microbiome, which may pose a potential risk to the health of NHPs. To the best of our knowledge, this is the first report the presence of FLAs in NHPs naturally infected. However, it is not known whether gut-borne FLA are exploiting a viable ecological niche, or whether they are simply transient residents in the gut.
Keywords:
microbiome
; culture
; morphology
; Acanthamoeba
; Vermamoeba
1. Introduction
The complex network of microorganisms that reside in the intestinal microbiome regulates host health through immune, endocrine, and metabolic pathways (e.g., food decomposition, nutrient intake, and drug metabolism) [1,2]. Emerging evidence shows that the majority of the members of this complex remain unknown [3,4]. New insights into the composition of this community are crucial for determining its potentially significant implications for animal health and biological diversity. Despite recent advances of eukaryotic enrichment protocols of metagenomics approaches, studies focused on microeukaryotic organisms are still a young field [5,6]. Indeed, the databases, analysis of the data, and also assembly-based metagenomic tools have been showed less robust when targeting eukaryotic microbes [7]. Lately, a study reported that captive Non-Human Primates (NHPs) harbor microbial species that are more similar to the human ones compared to wild NHPs, and the extent of microbiome overlap is strongly lifestyle dependent [8]. Several parasites that can be found in NHPs, such as Babesia, Cryptosporidium, Amoeba, Toxoplasma, Trypanosoma, Coccidia, nematodes and cestodes, possibly constitute a risk for primates, including humans [9,10,11].
Recently. the DNA of free-living amoebae (FLA) has been detected by classical approaches and PCR amplification, along with high-throughput sequencing in fecal samples [12]. However, a limited number of studies have attempted to address the presence of free-living amoebae (FLA) in the gut microbiome of mammals and other animals [13,14,15,16,17]. Whether those amoebae are therefore likely to have a profound impact on gut, which if so, has been largely overlooked.
FLAs are aerobic, mitochondriate, underexplored eukaryotic protists, but that are ubiquitous in diverse environmental sources [18,19,20]. Among protists, free-living amoebae are the dominant bacterial consumers [21] that may favor true colonization or a transitory passage in the intestinal microbiome. Recently, a metagenomic analysis revealed the presence of Acanthamoebidae and Vahlkampfiidae families in the microbiome of NHPs and other studies have reported the presence of FLA in the intestine [12]. Emerging evidence has linked several FLAs to human disease. Some species of the genera Acanthamoeba spp., Balamuthia spp., Sappinia spp., and Naegleria spp. are potentially pathogenic for humans and other animals [22,23,24,25,26,27,28,29]. Acanthamoeba spp. are found in an aquatic environment, soil, and air and can be isolated from the rhino pharynx and the stool of healthy humans [13,14,15,19,22,23,24,25]. Acanthamaeba keratitis being not uncommon in immunocompetent individuals. On the other hand, brain or disseminated infection in immunocompromised individuals caused by Acanthamoeba sp. are considered rare [28,29]. Primary amoebic meningoencephalitis (PAM), due to Naegleria fowleri and Granulomatous amoebic encephalitis (GAE) by Balamuthia mamdrillaris, are relatively rare too, but perhaps they are under detected and thus, underestimated [18,29]. Indeed, clinical disease are considered to have a very poor prognosis. The symptoms are idiopathic which can be mistaken for other bacterial and viral diseases, and the clinicians who treat the patients are not familiar with FLA [29]. And not surprisingly, almost all cases diagnosed is made at autopsy. Additionally, reliable diagnostic tests are unavailable and, limited diagnostic expertise [18,28,29].
Despite all the issues stressed above, FLA have gained public attention in recent years due to their capability of harboring pathogenic and nonpathogenic microorganisms, through transient or symbiotic relationships [30,31,32,33]. It is not clear what role these organisms play in terms of host survival, infectivity, and invasiveness. Due to their resistance to chlorine, the amoebic cysts are considered to be vectors of viruses, bacteria, fungi and protozoa [34]. Thus, FLAs have the potential to act as a vector of transmission for several microorganisms, however, Acanthamoeba is known to interact with a plethora of microorganisms, although not always to their own avail [32,33,35]. L. pneumophila can infect, replicate, and kill the Acanthamoeba upon escape [36]. While most studies investigate the interactions of Acanthamoeba with several microorganisms, other free-living amoebae should also be considered. Recently, it has been demonstrated the potential of Vermamoeba vermiformis to not only carry and allow multiplication of Helicobacter pylori but to revive the bacteria to a culturable state [37]. It is important to mention that the presence of FLAs in the intestinal microbiome has been reported yet remain poorly explored. Yet, there is the emerging sense that microbes interact in complex and subtle ways that we have yet to fully appreciate, much less understand. Despite their obvious importance, very little is known about it. It is becoming increasingly more evident that to minimize the risks of acanthamoeba and pathogens interactions to human health the gut microbial community should be considered, and further research should aim to better understand the interactions between FLAs-gut microbiota.
Notoriously, with the recent development of “culture-independent” methods of microbial characterization, high-throughput sequencing in fecal samples have been very useful, but they do not directly answer the most critical questions of host–microbiota interactions. To fully understand the presence of FLAs in the animal’s gut microbiota, the establishment of stable cultures is required to explain the ecology of microbiota relative to the biology and potential pathogenicity as well as FLAs–microbiota interactions. In this study, we used culture-dependent approaches to isolate FLA from the gut microbiome, combined with morphological analysis and different staining techniques and DNA sequencing. The NHP gut microbiome may provide an important evolutionary context for understanding the human gut microbiome. This study highlights the importance of FLA in the gut of mammals, with potential relevance to future studies on mucosal immunology, and potential pathogenicity of FLA. Added to this, human-pathogenic, amoeba-resisting microorganisms can be considered as an important emerging field.
2. Materials and Methods
2.1. Sampling
Fecal samples were collected across three conditions: (i) fecal samples collected after defecation on the floor surface of cages; (ii) rectal washouts of captive non-human primates after attaining deep anesthesia, and (iii) fecal samples directly collected from the intestine of wild animals after Euthanasia.
Samples in this cohort were collected from 101 individuals representing three primate species of captive non-human primates. A total of 96 fecal samples were collected of Macaca mulata (N = 65), Saimiri ustus (N = 01), and Saimiri sciureus (10) and 20 rectal washout samples of Macaca mulatta from the Institute of Biomodel Science and Technology (ICTB, Fiocruz). Feces samples from captive animals were collected as soon as possible after defecation on the floor surface of their cages to minimize contamination, or directly from the rectum under anesthesia. In addition to these, five stools samples of Callithris sp. (N = 5) from the Fiocruz Atlantic Forest Campus of the Oswaldo Cruz Foundation (CFMA) (22°56’18”S 43°24’11”W). These stools were collected in the field laboratory after dissection of the viscera, stored in labeled plastic containers, and transported to the main laboratory. All samples were collected from animals habituated to humans, making it possible to collect from specific individuals.
2.2. Culture and Morphological Analyses
Briefly, 1 g of fecal material was homogenized in 2 mL of phosphate-buffered saline; 50 µL of the homogenate filtered through membrane filters with an 8 µm pore size were added to 1.5% non-nutrient agar (NNA) plates containing 2 mL of Page’s amoeba saline, without killed Escherichia coli. All plates were sealed and incubated at 27 °C. After two days, the plate was washed three-time media Page’s amoeba saline, to remove the organic debris and bacteria layer. After that, media Page’s and heat-killed Escherichia coli was added to plate. Each two days, plate was examined using an inverted microscope. When positive, sub-cultured by cutting out a small piece of agar and placing it onto a fresh plate overlaid with a layer of heat-killed Escherichia coli. Finally, within a week to 2 weeks was obtained a robust culture that can be maintained by periodic transfer of aliquots to new culture. The plates were observed daily for amoebic growth up to 30 days after inoculation using an inverted microscope at 200× and 400× magnification.
The presence of FLAs was confirmed from the cyst and trophozoite morphology and sub-cultured by cutting out a small piece of agar and placing it onto a fresh plate overlaid with a layer of heat-killed Escherichia coli. Positive cultures were used to study the morphology and motion characteristics of each isolate using both bright-field and phase-contrast microscopy and permanent stained smears. The material from the surface of the plate was removed, fixed, and stained with Giemsa [38], and Panoptic® staining and examined at a higher magnification (1000×). The smears were dipped for 5 second in each solution, A (fixative number 1), B (Eosin Panoptic No. 2), and C (Blue Panoptic No. 3) without washes in between them. Excess reagent was drained from the slides between solutions and the buffer solution, pH 7.2 x 1second [39].
2.3. Molecular Identification of Free-Living amoeba DNA Sequencing
Samples with a positive result for FLAs at microscopy were re-assessed by a direct PCR method. DNA extraction from the isolates was performed with the commercial QIAmp DNA Mini Kit (Qiagen), according to the manufacturer’s instructions. PCR assay was performed according to the morphological criteria of the amoebae in positive plates using different sets of primers. The genus Acanthamoeba was confirmed by polymerase chain reaction (PCR), using primer forward JDP1 (5´-GGCCCAGATCGTTTACCGTGAA-3´) and reverse JDP2 (5´-TCTCACAAGCTGCTAG GGAGTCA-3´,) which amplifies a 423- to 551-bp fragment for the 18 S rDNA [40]. Likewise, the PCR were performed for Vermamoeba spp. and Vannella spp using primers: forward NA1 5-GCTCCAATAG CGTATATTAA-3 and reverse NA2 5-AGAAAGAGCTATCAATCTGT-3 [41]. Fragments of approximately 650 and 700-800 bp were identified in gel electrophoresis. Amplicons were purified using the Wizard® SV gel and PCR Clean-Up System kit (Promega, Madison, WI, USA) and sequenced for both strands using the PCR primers. DNA cycle sequencing reactions were performed using the BigDye® Terminator v.3.1 Cycle Sequencing Kit and loaded in the ABI 3730 Sequencing Platform (both - Applied Biosystems, Foster City, CA, USA). Raw bi-directional sequences reads were trimmed, assembled into contigs, and manually edited using SeqMan (DNASTAR software package, DNASTAR Inc., Madison, WI, USA) and exported in FASTA format. The consensus sequences were compared with previously published sequences using the Basic Local Alignment Search Tool (BLASTn) available in the GenBank sequence database.
2.4. Assessment of Viability
Acanthamoeba castellanii (ATCC NEF 30010) and Acanthamoeba polyphaga (ATCC 30461) trophozoite growth were cultured at 37°C in the peptone-yeast extract-glucose medium (PYG). For acidic exposure, we evaluated the impact of acidic culture medium on the viability of amoebae, the trophozoite were incubated in media of over a pH range (pH of 2.0 to 6.0), and pHs were adjusted to a required level (2, 3, 4, 5, and 6) and regulated automatically by HCl. Cultures were incubated for 12, 24, 36, 48, 72, and 96 h at 37 °C, with a starting density of 5×104 amoebae/mL. Trophozoite density was determined by cell counting under a light microscope using a Neubauer chamber hemocytometer.
3. Results
Out of 101 fecal samples obtained from PNHs, 46 (45.4%) were positive for potentially pathogenic FLAs, based on the morphological and locomotion criteria, by inverted microscopy (magnification ×200 and ×400) using standard taxonomic identification sources. A great diversity of free-living amoebae was observed in the fecal samples of Callithrix sp. (N=5), Macaca mulatta (N=26), Saimiri sciureus (N=8). And, in rectal washout samples of Macaca mulatta (N=7). All samples had mix-species infections where two or more morphotypes were associated in the same culture. Unfortunately, several cultures of FLA were lost due to fungal heavily overgrowth, even if when added anti-fungal drugs.
Altogether, different groups of FLA were identified, including Acanthamoeba sp., Vermamoeba sp, Naegleria sp., and fan-shaped amoebae of the family Vannellidae (Figure 1b–e). The trophozoites assumed a monopodial form, and the amoeboid-form organism changed to the transient, flagellate form with flagella at the broad end (Figure 1d). In addition, typical mobility was observed with either spinning or jerky movements (Additional file 1), and a cyst form with thick double walls was observed.
Acanthamoeba sp. were identified in the form of double-walled cysts. The ectocyst (outer wall) was differentiated from the variably stained surrounding background, and the endocyst (inner wall) with a stellated, polygonal, square, round, or oval aspect were visually distinguished from the spherical outline of the ectocyst (Figure 2i–l). Unidentified amoeba with rounded cysts ( Figure 2 m) and mix-species infections where two or more morphotypes were associated in the same culture
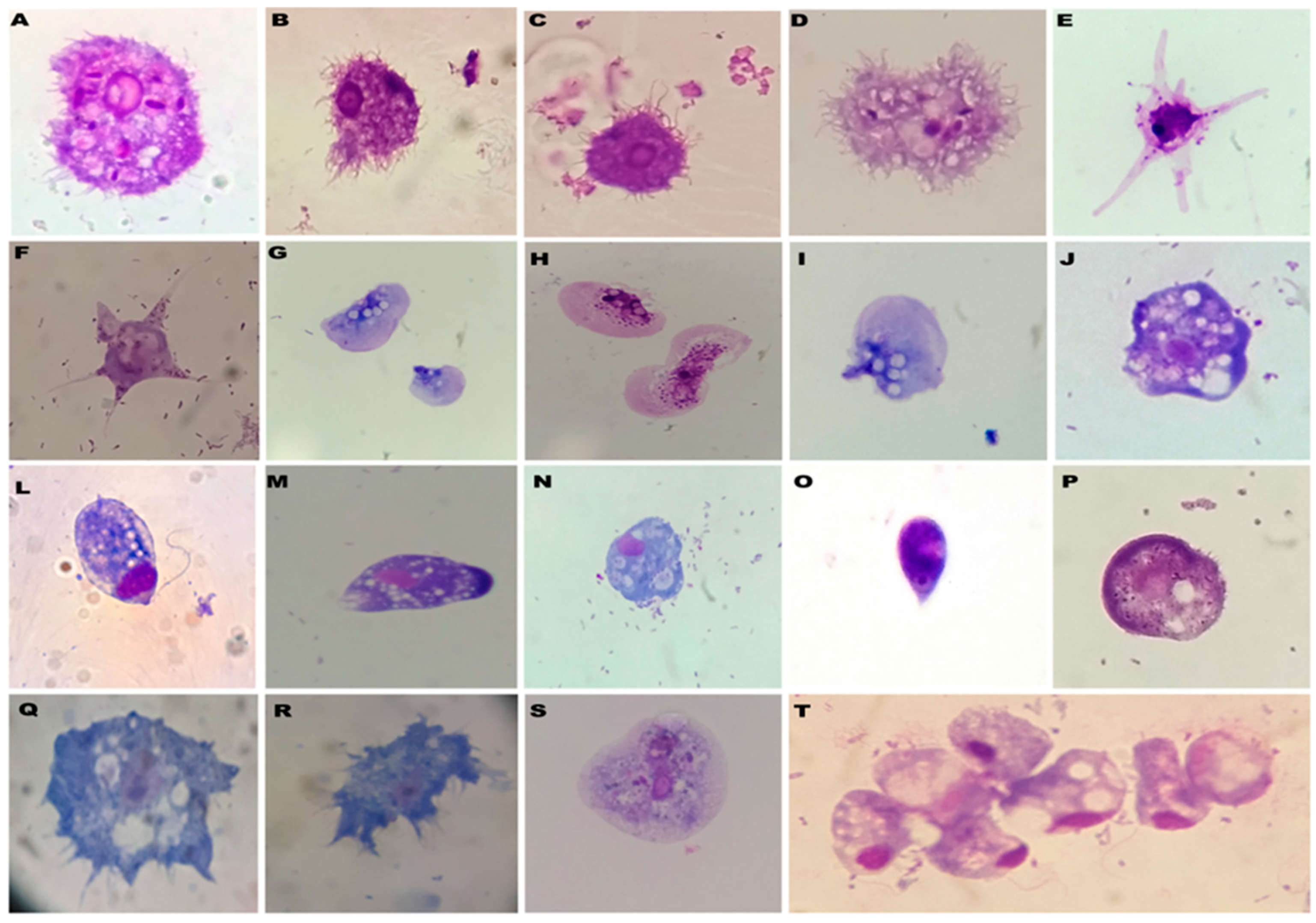
Numerous pine-like pseudopods (acanthopodia) gave the cell a spiny appearance (Figure 1e, Figure 3a–d,p–r), with a single nucleus with well-defined central nucleolus visible in the trophozoites (Figure 3b). Flotation forms of amoebae (Figure 3E–F) and fan-shaped amoebae of the family Vannellidae (Figure 3G–I). Unidentified trophozoites.
All samples positive were mixed with their different genera. Subculturing helped isolation; however, despite our hard trials, only five samples were successfully mono-isolated.
The results of permanent-stained slides showed that the most consistent stain for identifying Acanthamoeba cysts and trophozoites was Panoptic® stain, followed by the Giemsa stain. The Giemsa stain gave poor visibility for acanthopodia. Trophozoite, flagellate, and cyst forms of Naegleria were observed. The Vermamoeba-like trophozoite was detected in 2 of the 46 positive samples, (Figure 1c), slightly oval appearance with a single wall cyst and trophozoite had well-visible cylindrical monopodia in medium. In turn, fan-shaped amoebae of the family Vannellidae were identified by locomotive and floating forms, main characteristics for this amoebae genus (Figure 3 e-i). Besides, trophozoites were semi-circular or fan-shaped, and pronounced areas of the frontal hyaloplasm and the cytoplasm filled with numerous granules (Additional file 2) containing numerous optically empty vacuoles were observed (Figure 1b).
Of the total of 46 samples positive for AVLs from captive and free-living PNHs, cultivated in Agar PAGE, only 36 were submitted to DNA amplification by PCR. Rest of samples (N=10) were not tested due to excessive fungal growth. PCR using genus-specific primers (JDP1 and JDP2) confirmed Acanthamoeba spp. in 77.8% (36/28), amplifying an expected fragment of approximately 500 bp. Sequencing of PCR products and BLAST analysis revealed that 18% (5/28) of sequences belonged to the T4 genotypes when compared to the reference sequences deposited at GenBank. The percentual of identity ranged from 99% to 100% (accession number: MK713911, MT378247, MT378246, MF197422, MF100900, MT378235). The rest of the sequencing of PCR products showed chromatograms suggestive of the presence of mixed infections. In turn, when used the set of primers NA1/NA2, 800 bp PCR products were obtained in 92% (33/36) isolates. However, mixed infections were clearly detected visually in the chromatograms with the sequencing trace has two or more peaks in the same location. Of the 33 PCR products, only one product was successfully sequenced. The sequence analysis revealed that the amoeba had a high homology of 99% to V. vermiformis (access number: KX856374.1, KP792393.1, MN238712.1). Moreover, PCR assay failed to show any positive results for Naegleria-like flagella.
Along the whole pH range studied, remarkably, at an acid pH of 2, A. castellanii and A. polyphaga were observed moving for 24 h but are nonviable within 48 h. While at a pH of 3 through 6, the amoebae remain viable for more than 96 h.
4. Discussion
Eukaryotic diversity in the gut microbiome remains relatively unexplored. Our current understanding of gut microbial communities is shaped by a disproportionate focus on the role of bacteria; however, eukaryotes also shape the community through predation, resource competition, and interaction with the host immune system.
Free-living amoebae are microbial eukaryotes that are widely distributed in the natural environment. In the last decade, those amoebae have attracted considerable research interest, mainly as environmental hosts of several intracellular pathogens [42]. However, we have only scratched the surface of the role of FLAs in the complex and extreme habitat of the mammal’s gut. Arguably, FLAs are an underappreciated group within microbiota, and their interaction with the immune system is an open question, as well as the risk that this could represent to the mammals. However, there have been no studies focused on the issue. In general, the studies have used a limited sample size on isolated members of the eukaryotic community and/or parasite identification under a pathogenic condition [43,44]. Recently, a report based on the metagenomic analysis described the presence of Acanthamoebidae and Vahlkampfiidae families and other protists in the microbiome of NHPs [12]. Conversely in further studies, studies based on stool cultures confirm the presence of FLA in the gut of other vertebrates and invertebrates [13,14,15,16].
In the present study, an unexpected diversity of free-living amoebae in the fecal microbiota of non-human primates was observed. Among the many free-living amoebae existent in nature, Acanthamoeba spp., Balamuthia mandrilaris, Vermamoeba vermiformis, Naegleria fowleri, and Sappinea pedata have been studied much more studied due to their association with disease [19,21,45]. They may cause skin ulcers, keratitis, encephalitis, and amoebic meningoencephalitis in primates and other animals (dogs, ovines, bovines, horses, and kangaroos as well as birds, reptiles, amphibians, fish, and even invertebrates) [19,21,22,46]. Acanthamoeba spp occasionally infects humans and animals as opportunistic pathogens. A case of fulminant amoebic meningoencephalitis and pneumonitis in a simian immunodeficiency virus (SIV)–infected rhesus macaque was reported due to Acanthamoeba spp. [47]. In turn, B. mandrillaris was first discovered in a mandrill baboon (Papio sphinx) that died of encephalitis at the San Diego Zoo Wildlife Park in California in 1986 [48]. In further studies, this amoeba was isolated in Gorilla gorilla gorilla, Pongo pygmaeus), and Old-World primates, including a colobus monkey (Colobus guereza kikuyuensis) and a gibbon (Hylobates concolor leucogenys) [49,50,51], but have been reported in both immunocompetent and immunocompromised individuals of all ages.
Although Vannella spp. is not considered pathogenic in itself, they are capable of harboring pathogenic intracellular organisms such as Microsporidian parasites. Most of the studies focused on the capability of these amoebae to harbor pathogenic and nonpathogenic microorganisms [31,33]. In turn, in the environment, FLAs have played relevant roles in the control of the microbial communities, and some amoebae are considered a “sanctuary” for viruses, bacteria protozoa, and fungi. Some of these microorganisms have no known reservoirs, and many are yet to be discovered [31,32]. Moreover, they have evolved to become resistant to these protists and are emerging pathogens [33].
The gold standard for most main FLAs diagnoses is still culture on E. coli-coated non-nutrient agar plates. However, in the past years, several standard PCR or real-time PCR protocols have become available. In this study, amoebae were isolated and identified at the general level based on morphologic features and at the species level by DNA sequencing. In the presente study, the isolates exhibited morphological characteristics matching those of Acanthamoeba spp, Naegleria spp, Vermamoeba spp, and fan-shaped amoebae of the family Vannellidae. The diversity of FLA described agrees with previous studies [14,15,52,53]. Moura et al., reported the presence of Acanthamoeba, Vahlkampfia, Hartmannella spp (currently named as Vermamoeba spp) and Echinamoeba in human fecal sample by culture-dependent methods. The authors demonstrated in vivo that the isolates of Acanthamoeba spp were capable to produce cerebral lesions [14]. Indeed, an interesting observation was the detection of Vannella spp, which is not one fact unprecedented. Even nonpathogenic Vanella sp can be of clinical relevance, as they can act as vehicles for pathogenic organism. In support of this, Vannella stains and their endocytobiont (resembling microsporidia organism and Pseudomonas aeruginosa) were detected from corneal scrapings [56]. In this study, the primers successfully amplify DNA from samples fan-shaped amoeba of the family Vannellidae, but no PCR-products were successfully sequenced. In turn, our findings of sequence analysis of the PCR products revealed the presence of the Acanthamoeba T4 genotype and V. vermiformis with high similarity with previous studies [57,58]. In general, analyses morphological pointed mixed infections, which is corroborated with chromatogram sequence analyses. Acanthamoeba spp. have been classified into 23 genotypes (T1-T23) based on their 18S rRNA whole gene sequences. However, genotype T4 is the most prevalent in clinical and environmental samples [59]. In the present study, sequences were identical or similar to previously described isolates of the T4 genotype, a trend observed in previous studies. However, since the different genotypes of Acanthamoeba differ in their pathogenic potential, it is relevant to assess whether such differences exist among the subtypes/species within the same genotype, mainly the genotype T4, which is related to the majority of Acanthamoeba infections. Indeed, the relatively low level of knowledge of global amoeba biodiversity indicates a very high probability of finding species in any habitat. Several studies demonstrated the occurrence of potentially pathogenic Acanthamoeba and Vermamoeba in the oral and nasal mucosa of patients with suppressed immune status such as a patient with HIV/AIDS, patients undergoing hemodialysis, and healthy subjects [28,60,61].
In the last years, some studies have reported that the gut microbiome composition is shaped predominantly by environmental factors [62]. It seems reasonable to speculate that wild-caught and outdoor housed animals have the potential to be infected with a variety of bacterial organisms, protozoan and metazoan parasite. Therefore, it is not unlikely that the diversity of FLA in feces may simply reflect the ubiquitous distribution of free-living amoeba in the environment (mainly soil and water collections). According to a study, a ubiquitous colonization and opportunistic infection by free-living eukaryotes such as Cercozoa, Acanthamoeba spp., and other Discosea were observed in pigs [16]. Strikingly, the authors reported that FLAs were recovered in from stomach, ileum, caecum, colon, and rectum and at both incubation temperatures, 25◦C and 37◦ C. Diversity was dominated by amoebae: vahlkampfiids, vannellids, Acanthamoeba spp., Hyperamoeba sp. and Vermamoeba vermiformis.
It is worth noting FLAs feed mainly on bacteria and are resilient to harsh abiotic factors. Macaca mulatta gastric pH, and acid output resemble that of humans [63]. So, how do FLAs survive through the stomach? Notoriously, amoebae possess protection mechanisms against harsh environment. The capacity of the FLAs to adapt in any imaginable set of conditions is remarkable. However, this question is poorly understood and neglected.
In this study, we explore the impact of acid pH on viability of Acanthamoeba spp parasite growth rates were measured in range od from pH 2 to 6. Surprisingly, at an acid pH of 2, A. castellanii and A. polyphaga were observed moving for 24 h, but are nonviable within 48 h. In contrast, in pH ranging from 3 to 6, the amoebae remain viable for more than 96 h. Similar results have been recorded previously with Naegleria fowleri [64]. Under harsh conditions, many amoebas survive by encystment. Acanthamoeba cysts have been shown to be resistant to extreme conditions including freezing, pH 2.0, and ultraviolet irradiation, heavy metals, desiccation, and storage at 4 °C, over long time (20 years) [65]. Likewise, many bacteria that transit gastrointestinal tract (GIT) are neutralophilic but resist acid pH, using other approaches. For example, the ability of H. pylori to survive the low pH of the stomach would seem to suggest that it is an extreme acidophile. In fact, H. pylori is a neutrophile. This bacterium escapes acidity in the stomach by breaking down urea with enzyme-urease producing large amount of ammonia (alkaline) that raising the pH of the immediate environment, in which the pH is nearly neutral [66]. In turn, a study in vitro conditions exhibited no effect on the viability of A. castellanii when cells were inoculated for encystation at different pH with or without the presence of 10% glucose and 50 mM MgCl2. In additional to, acidic pH (3.0) did not serve as a strong stimulus for encystation of A. castellanii, whereas a neutral pH 7.0 served as an optimum medium in which approximately 30-40% encystation. Lastly, the authors reported that light-dark cycles, 5% CO2 and microaerophilic conditions had no effect on encystation of A. castellanii [67]. In the light of these observations, it is worth noting, FLAs feed mainly on the bacteria and are resilient to harsh abiotic factors, what provide clear evidence that some FLAs might remain viable, consequently allowing the colonization and persistence in gut tract.
Although, our findings revealed a diverse occurrence of FLAs infections, environmental contamination related to the sample collects can be excluded, because of part of samples were collected postmortem from Callithrix spp (wild PNH) and the washed gut of captive PNHs. Moreover, we analyzed serial samples on multiple non-consecutive days. The analysis of serial samples collected from several days can help to distinguish between pseudo parasites and real parasites and contamination by soil. In our study, captive NPHs as free primates from the Atlantic Forest region are exposed to soil, water, and air, and hence are at increased risk of getting infected by free-living amoeba. The risk of FLA acquisition is linked to the potential of the host being exposed under local environmental conditions. In many circumstances, NHPs under significant physiological stress or with a compromised immune system are at high risk of severe consequence and death from infectious disease, even those with low virulence. Infectious disease plays a major role in the lives of non-human primates and can have a tremendous negative impact. However, clinically relevant free-living amoebic in animals are rare, but it can show severe and fatal progressions, mainly due to lack of awareness leading to delayed diagnosis and to the shortage of effective treatment.
There is a wide gap in our knowledge regarding FLAs in the mammalian gut, and questions arise as to whether they could represent a long-term colonist or a transient invader in the mammalian intestinal microbiome. However, Acanthamoeba can also be found in the throat, intestine, and nasal region of healthy individuals [14,42,68]. Notoriously, most healthy individuals (humans or other animals) do not develop disease despite regular contact with free-living amoebae. Presumably, the presence of antibodies to FLA in healthy human sera contributes to protecting against infections. Asymptomatic exposures are common in healthy individuals [42,69,70]. In animal models, some studies in vivo and in vitro have shown effective protection against meningoencephalitis and Keratitis after immunization with Acanthamoeba antigen and anti-Acanthamoeba tear IgA provides an immunological barrier, blocking their adherence to epithelial cell [70]. Oral immunization protects against Acanthamoeba keratitis in corneal infections in pigs when administered before the corneal challenge [71] and in hamster animal model [72].
In this study, most of the genera found have already been involved in humans and, or animals infectious. However, our results showed only widespread asymptomatic exposures among the non-human primate. Kollars and Wilhelm reported wild mammals could become infected in nature and produce antibodies against the anti-Naegleria spp. in the manners described for laboratory experiments [73] In general, exposure to antigens via mucosal surfaces in the gut induces the preferential generation of secretory IgA antibodies [74]. Although it is not the objective of this study, we hypothesize that the interaction FLA with mucosal surfaces (gut colonization) may induce a local and systemic humoral immune response that provides solid protection against the development of intestinal and extraintestinal diseases in these animals.
Our findings do not allow for inference on whether the detected FLA are residents or merely transients, nor whether animals acquire natural immune responses. Beyond that, we did not characterize the pathogenicity of the isolates, only detected a genotype considered potentially pathogenic, Acanthamoeba genotype T4. It is necessary to investigate the pathogenicity of the isolated strain. Consequently, future investigations have tremendous potential in understanding FLA in the gut, permitting further scrutiny of natural immune responses of PNHs, chemotherapy, pathogenicity, and dynamics of parasite-host relationships.
5. Conclusions
A great diversity of FLAs was observed in the NHP microbiome, which poses a potential risk to the health of NHPs. Crucially, further studies need to focus on molecular identification of isolates the species level and evaluate their pathogenic potential. However, to the best of our knowledge, this is the first report presenting the occurrence of potentially pathogenic genotype T4 Acanthamoeba, Vermamoeba vermiformis and fan-shaped amoebae of the family Vannellidae.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. The datasets supporting the findings of this article are included within the article and its additional file (video S1: The trophozoites with amoeboid-form changed to the transient, flagellate form typical mobility and video S2: Semi-circular or fan-shaped trophozoites).
Author Contributions
Conceptualization, Helena Lúcia Carneiro Santos; methodology, Igor Rodrigues Cardoso, Clezia Siqueira de Lima, Rhagner Bonono dos Reis, Araujo Pinto, Thalita Pissinatti, Tatiana Kugelmeier, Sócrates Fraga da Costa Neto, and Fabio Alves da Silva; data curation, Igor Rodrigues Cardoso; writing—original draft preparation, Igor Rodrigues Cardoso; writing—review and editing, Helena Lúcia Carneiro Santos.All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Instituto Oswaldo Cruz intramural funding (PAEF II-IOC-22-FIO-18-2-53) and Programa de Pós-Graduação em Medicina Tropical (IOC-FIOCRUZ).
Institutional Review Board Statement
The study was conducted in accordance with protocols that were reviewed and approved by the Committee of Ethics in Animal Expermentation of the Oswaldo Cruz Foundation Rio de Janeiro, Brazil, (protocols number LW5-16 and LW-57/19).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets supporting the findings of this article are included within the article and its additional file. Sequences have been deposited in GenBank database under accession numbers: OR68553, OR685532, OR685533, OR685534, OR685535 and OR685536.
Acknowledgments
The manuscript benefitted from the insightful comments of three anonymous reviewers.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Berrilli F, Di Cave D, Cavallero S, D’Amelio S. Interactions between parasites and microbial communities in the human gut. Front Cell Infect Microbiol. 2012, 2, 141. [Google Scholar] [CrossRef]
- Rastelli M, Cani PD, Knauf C. The Gut Microbiome Influences Host Endocrine Functions. Endocr Rev. 2019, 40, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Thomas AM, Segata N. Multiple levels of the unknown in microbiome research. BMC Biol. 2019, 17, 48. [Google Scholar] [CrossRef]
- Almeida A, Mitchell AL, Boland M, Forster SC, Gloor GB, Tarkowska A, et al. A new genomic blueprint of the human gut microbiota. Nature. 2019, 568, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Hooks KB, O’Malley MA. Contrasting Strategies: Human Eukaryotic Versus Bacterial Microbiome Research. J Eukaryot Microbiol. 2020, 67, 279–295. [Google Scholar] [CrossRef]
- Lind AL, Pollard KS. Accurate and sensitive detection of microbial eukaryotes from whole metagenome shotgun sequencing. Microbiome. 2021, 9, 58. [Google Scholar] [CrossRef]
- Breitwieser FP, Lu J, Salzberg SL. A review of methods and databases for metagenomic classification and assembly. Brief Bioinform. 2019, 20, 1125–1136. [Google Scholar] [CrossRef]
- Manara S, Asnicar F, Beghini F, Bazzani D, Cumbo F, Zolfo M, et al. Microbial genomes from non-human primate gut metagenomes expand the primate-associated bacterial tree of life with over 1000 novel species. Genome Biol. 2019, 20, 299. [Google Scholar] [CrossRef]
- Ashford, R.W. Reid G.D.F., Butynski T.M. The intestinal faunas of man and mountain gorillas in a shared habitat. Ann. Trop. Med. Parasitol. 1990, 84, 337–340. [Google Scholar] [CrossRef]
- Muriuki, S.M.K. Murugu R.K., Munene E., Karere G.M., Chai D.C. Some gastro-intestinal parasites of zoonotic (public health) importance commonly observed in old world non-human primates in Kenya. Acta Trop. 1998, 71, 73–82. [Google Scholar] [CrossRef]
- Gómez, J.M. Nunn C.L., Verdú M. Centrality in primate-parasite networks reveals the potential for the transmission of emerging infectious diseases to humans. Proc. Natl. Acad. Sci. USA. 2013, 110, 7738–7741. [Google Scholar] [CrossRef] [PubMed]
- Gogarten JF, Calvignac-Spencer S, Nunn CL, Ulrich M, Saiepour N, Nielsen HV, et al. Metabarcoding of eukaryotic parasite communities describes diverse parasite assemblages spanning the primate phylogeny. Mol Ecol Resour. 2020, 20, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Mehta AP, Guirges SY. Acute amoebic dysentery due to free-living amoebae treated with methronidazole. J Trop Med Hyg. 1979, 82, 134–136. [Google Scholar]
- de Moura H, Salazar HC, Fernandes O, Lisboa DC, de Carvalho FG. [Free-living amoeba in the human intestine. Evidences of parasitism]. Rev Inst Med Trop Sao Paulo. 1985, 27, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, RS. Free-living amoebae recovered from human stool samples in Strongyloides agar culture. J Clin Microbiol. 2014, 52, 699–700. [Google Scholar] [CrossRef] [PubMed]
- Chavatte N, Lambrecht E, Van Damme I, Sabbe K, Houf K. Free-living protozoa in the gastrointestinal tract and feces of pigs: Exploration of an unknown world and towards a protocol for the recovery of free-living protozoa. Vet Parasitol. 2016, 225, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Mulec, J., Dietersdorfer, E., Ustunturk-Onan, M., and Walochnik, J. Acanthamoeba and other free-living amoebae in bat guano, an extreme habitat. Parasitol Res. 2016, 115, 1375–1383. [CrossRef]
- Angelici MC, Walochnik J, Calderaro A, Saxinger L, Dacks JB. Free-living amoebae and other neglected protistan pathogens: Health emergency signals? Eur J Protistol. 2021, 77, 125760. [Google Scholar] [CrossRef] [PubMed]
- Visvesvara GS, Moura H, Schuster FL. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol Med Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef]
- Carlesso AM, Simonetti AB, Artuso GL, Rott MB. [Isolation and identification of potentially pathogenic free-living amoebae in samples from environments in a public hospital in the city of Porto Alegre, Rio Grande do Sul]. Rev Soc Bras Med Trop. 2007, 40, 316–320. [Google Scholar] [CrossRef]
- Schuster FL, Visvesvara GS. Free-living amoebae as opportunistic and non-opportunistic pathogens of humans and animals. Int J Parasitol. 2004, 34, 1001–1027. [Google Scholar] [CrossRef] [PubMed]
- Pimentel LA, Dantas AF, Uzal F, Riet-Correa F. Meningoencephalitis caused by Naegleria fowleri in cattle of northeast Brazil. Res Vet Sci. 2012, 93, 811–812. [Google Scholar] [CrossRef] [PubMed]
- Henker LC, Lorenzett MP, Dos Santos DL, Virginio VG, Driemeier D, Rott MB, et al. Naegleria fowleri-associated meningoencephalitis in a cow in Southern Brazil-first molecular detection of N. fowleri in Brazil. Parasitol Res. 2021, 120, 2873–2879. [Google Scholar] [CrossRef] [PubMed]
- Madrigal Sesma MJ, Santillana Lopez I. Isolation of free-living amoebas from samples of respiratory origin]. Rev Sanid Hig Publica (Madr). 1989, 63, 63–72. [Google Scholar]
- Rivera F, Medina F, Ramirez P, Alcocer J, Vilaclara G, Robles E. Pathogenic and free-living protozoa cultured from the nasopharyngeal and oral regions of dental patients. Environ Res. 1984, 33, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Santos LC, Oliveira MS, Lobo RD, Higashino HR, Costa SF, van der Heijden IM, et al. Acanthamoeba spp. in urine of critically ill patients. Emerg Infect Dis. 2009, 15, 1144–1146. [Google Scholar] [CrossRef]
- Parija SC, Dinoop K, Venugopal H. Management of granulomatous amebic encephalitis: Laboratory diagnosis and treatment. Trop Parasitol. 2015, 5, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Arab-Mazar Z, Niyyati M, Lasjerdi Z, Spotin A, Alavi Darzam I, Gachkar L. Isolation, identification, and phylogenetic analysis of potentially pathogenic free-living amoebae isolated from nasal and oral mucosa of HIV/AIDS patients in Iran. Parasitol Res. 2019, 118, 3061–3066. [Google Scholar] [CrossRef] [PubMed]
- Kot K, Lanocha-Arendarczyk N, Kosik-Bogacka D. Immunopathogenicity of Acanthamoeba spp. in the Brain and Lungs. Int J Mol Sci. 2021, 22, 3. [Google Scholar] [CrossRef]
- Van der Henst C, Vanhove AS, Drebes Dorr NC, Stutzmann S, Stoudmann C, Clerc S, et al. Molecular insights into Vibrio cholerae’s intra-amoebal host-pathogen interactions. Nat Commun. 2018, 9, 3460. [Google Scholar] [CrossRef]
- Goni P, Fernandez MT, Rubio E. Identifying endosymbiont bacteria associated with free-living amoebae. Environ Microbiol. 2014, 16, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Barker J, Brown MR. Trojan horses of the microbial world: protozoa and the survival of bacterial pathogens in the environment. Microbiology (Reading). 1994, 140 Pt 6, 1253–1259. [Google Scholar] [CrossRef]
- Balczun C, Scheid PL. Free-Living Amoebae as Hosts for and Vectors of Intracellular Microorganisms with Public Health Significance. Viruses. 2017, 9, 65. [Google Scholar] [CrossRef]
- Coulon C, Collignon A, McDonnell G, Thomas V. Resistance of Acanthamoeba cysts to disinfection treatments used in health care settings. J Clin Microbiol. 2010, 48, 2689–2697. [Google Scholar] [CrossRef]
- Dey R, Rieger AM, Stephens C et al. Interactions of Pseudomonas aeruginosa with Acanthamoeba polyphaga Observed by Imaging Flow Cytometry. Cytometry A 2019, 95, 555–564. [Google Scholar] [CrossRef]
- Rowbotham TJ. Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae. J Clin Pathol. 1980, 33, 1179–1183. [Google Scholar] [CrossRef]
- Dey R, Rieger A, Banting, G, Ashbolt N J, Role of amoebae for survival and recovery of ‘non-culturable’ Helicobacter pylori cells in aquatic environments. FEMS Microbiology Ecology 2020, 96, fiaa182.
- De Carli, G.A. Diagnóstico Laboratorial das Parasitoses Humanas - Métodos e Técnicas, 2 edn; Atheneu: São Paulo, Brazil, 2007. [Google Scholar]
- Liu, W. A simplified cytologic staining technic. Am J Clin Pathol. 1970, 54, 767–768. [Google Scholar] [CrossRef] [PubMed]
- Gatti S, Rama P, Matuska S, Berrilli F, Cavallero A, Carletti S, et al. Isolation and genotyping of Acanthamoeba strains from corneal infections in Italy. J Med Microbiol. 2010, 59, 1324–1330. [Google Scholar] [CrossRef]
- Nazar M, Haghighi A, Taghipour N, Ortega-Rivas A, Tahvildar-Biderouni F, Nazemalhosseini Mojarad E, et al. Molecular identification of Hartmannella vermiformis and Vannella persistens from man-made recreational water environments, Tehran, Iran. Parasitol Res. 2012, 111, 835–839. [Google Scholar] [CrossRef]
- Siddiqui R, Khan NA. Biology and pathogenesis of Acanthamoeba. Parasit Vectors. 2012, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Zanzani SA, Gazzonis AL, Epis S, Manfredi MT. Study of the gastrointestinal parasitic fauna of captive non-human primates (Macaca fascicularis). Parasitol Res. 2016, 115, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Drakulovski P, Bertout S, Locatelli S, Butel C, Pion S, Mpoudi-Ngole E, Delaporte E, Peeters M, Mallié M. Assessment of gastrointestinal parasites in wild chimpanzees (Pan troglodytes troglodytes) in southeast Cameroon. Parasitol Res. 2014, 113, 2541–2550. [Google Scholar] [CrossRef] [PubMed]
- Zhang H, Cheng X. Various brain-eating amoebae: the protozoa, the pathogenesis, and the disease. Front Med. 2021, 15, 842–866. [Google Scholar] [CrossRef] [PubMed]
- Schuster FL, De Jonckheere JF, Moura H, Sriram R, Garner MM, Visvesvara GS. Isolation of a thermotolerant Paravahlkampfia sp. from lizard intestine: biology and molecular identification. J Eukaryot Microbiol. 2003, 50, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Westmoreland SV, Rosen J, MacKey J, Romsey C, Xia DL, Visvesvera GS, et al. Necrotizing meningoencephalitis and pneumonitis in a simian immunodeficiency virus-infected rhesus macaque due to Acanthamoeba. Vet pathol. 2004, 41, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Visvesvara GS, Martinez AJ, Schuster FL, Leitch GJ, Wallace SV, Sawyer TK, et al. Leptomyxid ameba, a new agent of amebic meningoencephalitis in humans and animals. J Clin Microbiol. 1990, 28, 2750–2756. [Google Scholar] [CrossRef] [PubMed]
- Anderson MP, Oosterhuis JE, Kennedy S, et al. Pneumonia and meningoencephalitis due to amoeba in a lowland gorilla. J Zoo Anim Med 1976, 17, 87–91. [Google Scholar]
- Canfield PJ, Vogelnest L, Cunningham ML, et al. Amoebic meningoencephalitis caused by Balamuthia mandrillaris in an orangutan. Aust Vet J 1997, 75, 97–100. [Google Scholar] [CrossRef]
- Rideout BA, Gardiner CH, Stalis IH, et al. Fatal infections with Balamuthia mandrillaris (a free-living amoeba) in gorillas and other Old World primates. Vet Pathol 1997, 34, 15–22. [Google Scholar] [CrossRef]
- Vellosa SAG, Mangini ACS, Nunes LR, Schlodtmann AG. Frequência de amebas de vida livre em fezes de indivíduos de uma creche da cidade de Säo Paulo / Frequency free-living amebas in the feces of persons from a nursery in São Paulo city. Rev Inst Adolfo Lutz. 1984, 44, 61–65. [Google Scholar] [CrossRef]
- Zaman, V. Acanthamoeba in human faeces from Karachi. Ann Trop Med Parasitol. 1999, 93, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Michel R, Schmid EN, Boker T, Hager DG, Muller KD, Hoffmann R, et al. Vannella sp. harboring Microsporidia-like organisms isolated from the contact lens and inflamed eye of a female keratitis patient. Parasitol Res. 2000, 86, 514–520.
- Montalbano Di Filippo M, Novelletto A, Di Cave D, Berrilli F. Identification and phylogenetic position of Naegleria spp. from geothermal springs in Italy. Exp Parasitol. 2017, 183, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Esboei BR, Fakhar M, Saberi R, Barati M, Moslemi M, Hassannia H, Dadimoghadam Y, Jalallou N. Genotyping and phylogenic study of Acanthamoeba isolates from human keratitis and swimming pool water samples in Iran. Parasite Epidemiol Control. 2020, 11, e00164. [Google Scholar] [CrossRef] [PubMed]
- Fuerst PA, Booton GC. Species, Sequence Types and Alleles: Dissecting Genetic Variation in Acanthamoeba. Pathogens 2020, 9, 534. [Google Scholar] [CrossRef] [PubMed]
- Memari F, Niyyati M, Lorenzo-Morales J, Jonaydi Z. Isolation and molecular characterization of Acanthamoeba strains isolated from the oral cavity of immunosuppressed individuals in Tehran, Iran. Acta parasitol. 2016, 61, 451–455. [Google Scholar]
- Niyyati M, Arab-Mazar Z, Lasjerdi Z, Lorenzo-Morales J, Espotin A, Yadegarynia D, et al. Molecular characterization of Acanthamoeba strains isolated from the oral cavity of hemodialysis patients in Iran. Parasitol Res. 2017, 116, 2965–2969. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D. Weissbrod, O., Barkan, E. et al. Environment dominates over host genetics in shaping human gut microbiota. Nature. 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Martin ME, Bhatnagar S, George MD, Paster BJ, Canfield DR, Eisen JA. The Impact of Helicobacter pylori Infection on the Gastric Microbiota of the Rhesus Macaque. PLoS ONE. 2013, 8, e76375. [Google Scholar]
- Lam C, He L, Marciano-Cabral F. The Effect of Different Environmental Conditions on the Viability of Naegleria fowleri Amoebae. J Eukaryot Microbiol. 2019, 66, 752–756. [Google Scholar] [CrossRef]
- Sriram R, Shoff M, Booton G, Fuerst P, Visvesvara GS. Survival of Acanthamoeba cysts after desiccation for more than 20 years. J Clin Microbiol. 2008, 46, 4045–4048. [Google Scholar] [CrossRef] [PubMed]
- Ansari S, Yamaoka Y. Survival of Helicobacter pylori in gastric acidic territory. Helicobacter. 2017, 22, 10–1111. [Google Scholar] [CrossRef]
- Aqeel Y, Siddiqui R, Iftikhar H, Khan NA. The effect of different environmental conditions on the encystation of Acanthamoeba castellanii belonging to the T4 genotype. Exp Parasitol. 2013, 135, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Michel, R. Rohl, R., Schneider, H., 1982. Isolation of free-living amoebae from nasal mucosa of healthy individuals. Zentralbl. Bakteriol. Mikrobiol. Hyg. 1982, 176, 155–159. [Google Scholar]
- Lares-Jiménez LF, Borquez-Román MA, Alfaro-Sifuentes R, Meza-Montenegro MM, Casillas-Hernández R, Lares-Villa F. Detection of serum antibodies in children and adolescents against Balamuthia mandrillaris, Naegleria fowleri and Acanthamoeba T4. Exp Parasitol. 2018, 189, 28–33. [Google Scholar] [CrossRef]
- Alizadeh H, He YG, McCulley JP, et al. Successful immunization against Acanthamoeba keratitis in a pig model. Cornea 1995, 14, 180–186. [Google Scholar]
- Leher H, Zaragoza F, Taherzadeh S, Alizadeh H, Niederkorn JY. Monoclonal IgA antibodies protect against Acanthamoeba keratitis. Exp Eye Res. 1999, 69, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Garate M, Alizadeh H, Neelam S, Niederkorn JY, Panjwani N. Oral immunization with Acanthamoeba castellanii mannose-binding protein ameliorates amoebic keratitis. Infect Immun. 2006, 74, 7032–7034. [Google Scholar] [CrossRef]
- Kollars, T.M., Jr. Wilhelm, W.E. The Occurrence of Antibodies to Naegleria Species in Wild Mammals. The Journal of Parasitology. 1996, 82, 73–77. [Google Scholar] [CrossRef]
- Pietrzak B, Tomela K, Olejnik-Schmidt A, Mackiewicz A, Schmidt M. Secretory IgA in Intestinal Mucosal Secretions as an Adaptive Barrier against Microbial Cells. Int J Mol Sci. 2020, 21, 9254. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Microphotograph of unstained free-living amoeba trophozoites observed in cultures usisng a ono-nutrient agar under light microscopy and phase contrst microscopy. A: Mix-species infections where morphotypes of different amoebae islated on non-nutrient agar plate, magnification × 400; B: Fan-shaped amoebae of the family Vannellidae, magnification × 1000, enlargement; C: An elongated cylindricalt trophozoite of Vermamoeba-like amoeba, magnification × 400; D: Naegleria flagellate stage temporary pear-shaped with a pair of flagella (black arrows), magnification × 1000, enlargement; E: Acanthamaoeba spp. show pine-like pseudopods (acanthopodia), magnification × 1000, enlargement; F and G: undentified trophozoite forms, magnification × 400.
Figure 1.
Microphotograph of unstained free-living amoeba trophozoites observed in cultures usisng a ono-nutrient agar under light microscopy and phase contrst microscopy. A: Mix-species infections where morphotypes of different amoebae islated on non-nutrient agar plate, magnification × 400; B: Fan-shaped amoebae of the family Vannellidae, magnification × 1000, enlargement; C: An elongated cylindricalt trophozoite of Vermamoeba-like amoeba, magnification × 400; D: Naegleria flagellate stage temporary pear-shaped with a pair of flagella (black arrows), magnification × 1000, enlargement; E: Acanthamaoeba spp. show pine-like pseudopods (acanthopodia), magnification × 1000, enlargement; F and G: undentified trophozoite forms, magnification × 400.
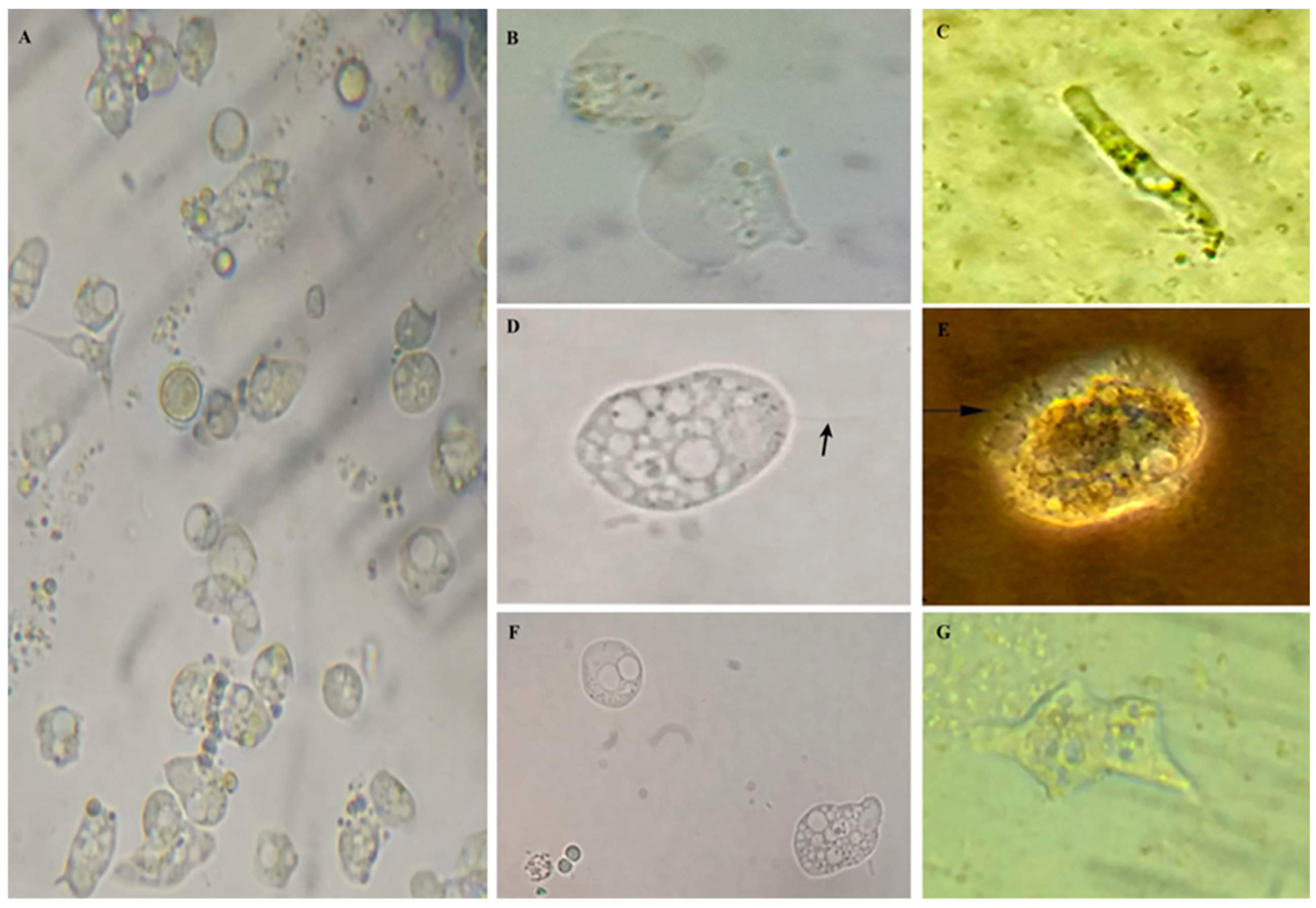
Figure 2.
Microphotograph of free-living amoeba cysts forms found in fecal samples from Non-Human Primates under light and phase contrst microscopy. A-F: rounded and polygonal cysts stained by Panoptic® stain; G and H: unstained cystic forms (round, traingle,and square) evidenced by Differential Interference Contrast (DIC), suggestive of the genus Acanthamoeba; C- Ectocyst (EC) and endocyst (ED). 400x magnification; I-L = Polygonal and stellate cysts, typical of the genus Acanthamoeba shows Ectocyst (EC) and endocyst (ED), 400x magnification. and M= unidentified amoeba with rounded cysts stained by Panoptic® stain.
Figure 2.
Microphotograph of free-living amoeba cysts forms found in fecal samples from Non-Human Primates under light and phase contrst microscopy. A-F: rounded and polygonal cysts stained by Panoptic® stain; G and H: unstained cystic forms (round, traingle,and square) evidenced by Differential Interference Contrast (DIC), suggestive of the genus Acanthamoeba; C- Ectocyst (EC) and endocyst (ED). 400x magnification; I-L = Polygonal and stellate cysts, typical of the genus Acanthamoeba shows Ectocyst (EC) and endocyst (ED), 400x magnification. and M= unidentified amoeba with rounded cysts stained by Panoptic® stain.
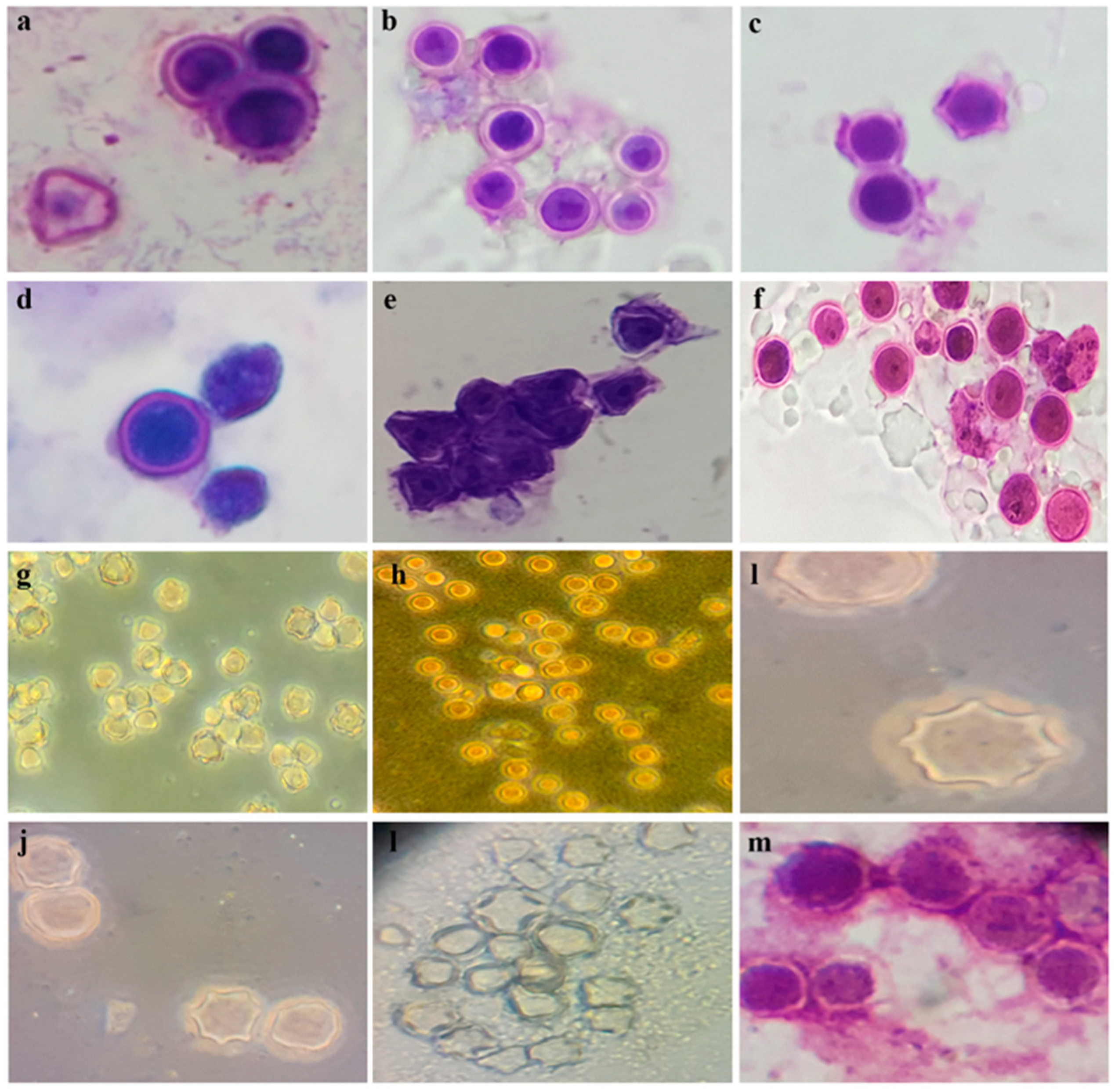
Figure 3.
Microphotograph of free-living amoeba trophozoites forms found in fecal samples from Non-Human Primates, stained by Panoptic stain, x 100 magnification. (A- D and P-R): Acanthamoeba-like trophozoites exibiting fine short acanthopodia; (E-F): Flotation form of the family Vannellidae; (G-I): Fan-shaped amoebae of the family Vannellidae; (L and O): The flagellate trophozoites assumed a monopodial form; (J, P, S -T): Unidentified trophozoites forms, showing nucleus and several vacuoles).
Figure 3.
Microphotograph of free-living amoeba trophozoites forms found in fecal samples from Non-Human Primates, stained by Panoptic stain, x 100 magnification. (A- D and P-R): Acanthamoeba-like trophozoites exibiting fine short acanthopodia; (E-F): Flotation form of the family Vannellidae; (G-I): Fan-shaped amoebae of the family Vannellidae; (L and O): The flagellate trophozoites assumed a monopodial form; (J, P, S -T): Unidentified trophozoites forms, showing nucleus and several vacuoles).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



