
Submitted:
02 November 2023
Posted:
02 November 2023
You are already at the latest version
Abstract
The methods used to introduce CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats)/Cas-mediated genome editing into fruit species, as well as the impacts of the application of this technology to activate and knock out of target genes in different fruit trees species including tree development, yield, fruit quality, and tolerance to biotic and abiotic stresses have been firstly described in this review. The application of this gene editing technology could allow the development of new generations of fruit crops with improved traits by targeting different genetic segments or even could facilitate the introduction of traits in elite cultivars without changing other traits. However, at this moment, the scarcity of efficient regeneration and transformation protocols in some species, the fact that many of those procedures are the genotype-dependent or the convenience of segregating the transgenic parts of the CRISPR system represent the main handicaps limiting the potential of genetic editing techniques for fruit trees. Finally, latest news on the legislation and regulations about the use of plants modified through CRISPR/Cas systems has been also discussed.
Keywords:
Plants
; Molecular Biology
; Genomic
; Transcriptomic
; Epigenetic
1. Introduction
Fruit trees are crops of great economic importance worldwide as they are vital components of our food production systems. Abiotic stresses such as salinity or drought, monoculture of disease-susceptible cultivars, excessive use of pesticides and the appearance of new pathogens cause significant economic losses in the production of various fruit species and are important threats to the environment and to the sustainable food production [1]. Fruit trees also play an integral role in the food and nutrition industries due to their invaluable primary and secondary metabolites [2]. Fruits are rich sources of dietary fiber, pectin, antioxidant components, phytoestrogens, cyanogenic glucosides, and vitamins that have been recognized for their role in promoting optimal human health and bolstering the body’s defense against illnesses [3].
On the other hand, perennial fruit trees suffer different biological and environmental challenges throughout their life. Specifically, fruit trees are infected by a wide range of pathogenic agents, including fungi, bacteria, viruses, which can lead to significant economic losses if not properly addressed or managed [1]. Furthermore, in the current climate change scenario that we face, it is increasingly common for fruit trees the lack of enough chilling accumulation during winter due to the increasing temperatures and this crops are also affected by extreme drought in some areas.
Therefore, there is great interest in obtaining improved fruit cultivas with high nutritional quality and resistant to different stresses. Additionally, it becomes imperative to grasp the roles of stress-tolerance-related genes and their regulatory mechanisms for the purpose of developing more resilient cultivars. Breeding of fruit crops through conventional techniques has been effectual in both quality and yield characteristics although, this is a slow breeding method with random consequences due to extrinsic and intrinsic factors such as long juvenile period, self-incompatibility, heterozygosity, long times for selection of the seedlings, and lack of correlations between seedling and mature plants [2]. Traditional breeding methods have been enriched by the inclusion of transgenesis, a valuable tool for plant breeding that enables the introduction or modification of specific and important traits in a single step [4], also allowing the functional genomic studies.
Despite their advantages, transgenesis has its own limitations including the random integration of transgenes into the genome and the fact that many fruit trees species are recalcitrant or transformation-time-consuming species. Therefore, it is paramount to enhance transgenic research and dedicate additional efforts to enhance the efficiency of regeneration and transformation procedures [5]. Besides, in higher plants, achieving the insertion of DNA sequences at a precise genomic location through homologous recombination, known as Gene Targeting (GT), has remained challenging due to the notably low efficiency of homologous recombination (HR) [6]. One approach to enhance HR-dependent gene targeting involves inducing double-strand breaks (DSBs) in the genomic DNA at the desired target site [7].
Among the new technologies developed in the last years, various site-specific nucleases (SSNs) have emerged, enabling the precise creation of DSBs at specific locations within the genome. SSNs have significant economic, time-saving, and streamlined advantages relative to conventional breeding methods which may take up to approximately a decade in order to develop a variety [8]. This methodology can use to study genes involved in traits such as drought tolerance, disease resistance, and higher quality and yield [9]. SSNs can be used for different purposes to modify the structure and function of host genome in agricultural crops, such as targeted mutation, modification, insertion, replacing, stacking, and translational modulation of desired genes [8].
Classification of SSN-based genome editing system is according to the following categories: meganucleases, zinc-finger nucleases (ZFNs), transcription-like effector nucleases (TALENs) and Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) that are associated with the RNA-guided Cas double-stranded DNA-binding protein (CRISPR/Cas system) [5,10]. The main differences among them are their relative specificity and efficiency [11]. ZFNs and TALENs are engineered nucleases and their mode of action is based on protein-DNA interaction. However, CRISPR/Cas system depends on RNA-DNA coupling [10]. Even though the use of the synthetic nucleases ZFNs and TALES has allowed the targeting of many genomic sites, the application of these techniques for edition of plant genomes has been limited [12].
Since its discovery, the most used genome editing tool used in plant research and breeding is CRISPR associated with a designed RNA-guided Cas9 endonuclease [13]. Specifically, CRISPR/Cas9 from Streptococcus pyogenes (SpyCas9) has been successfully used for genome editing in many plant species [14]. However, CRISPR/Cas9 has some inconveniences, such us limitation of target specificity, activity, efficiency and targeting scope [15]. These limitations have been overcome by engineering the basic Crispr/Cas9 system and the discovery of other Cas enzymes from various species, extending the range of genome editing tools [16].
The emergence of CRISPR/Cas technology initiated a new perspective on Genetically Modified Organisms (GMO) regulations. The generation of GMOs through transgenesis involves an insertion of a foreign DNA into the genome of the plant, which is not allowed in many countries around the world. However, the targeted modification of a gene using CRISPR/Cas technology producing mutations is in many cases similar to the application of mutagenic agents that are legally acceptable in some countries.
In this review, we have analyzed the methods used to introduce CRISPR/Cas systems into fruit species, as well as the impacts of CRISPR/Cas on altering plant architecture, improving fruit quality and yield. and tolerance to biotic and abiotic stresses. Furthermore, this review includes the latest news on the legislation and regulations about the use of plants modified through CRISPR/Cas systems.
2. Mechanism of CRISPR/Cas System
In nature, CRISPR/Cas systems provide prokaryotes an RNA-guided adaptive immunity against bacteriophages and plasmids [17]. These systems are encoded by the CRISPR array and the accompanying CRISPR-associated (Cas) genes. The CRISPR array contains two types of sequences, the palindromic repeats and the ‘spacer’ sequences that are derived from viral or plasmid genome. On the other hand, Cas genes codify for different proteins involved in the process [18].
The adaptative immune response consists in three stages: adaptation, expression and maturation, and interference [19]. In the adaptation stage, Cas proteins recognize foreign genetic elements (protospacers) and insert them in between the repeats of the CRISPR array, forming new spacers. The expression and maturation stage consist in the transcription of the CRISPR array into a pre-crRNA that is further processed forming smaller mature crRNAs, one for each spacer. Then, the crRNA forms a complex with Cas proteins, and in some cases with the tracrRNA (transactivating crRNA) [20], which leads to the interference stage. The complex recognizes by base-pairing foreign nucleic acids that are complementary to crRNA sequences. In addition, a specific motif called PAM (protospacer adjacent motif) is necessary for the stable binding to the target DNA and it’s crucial for the discrimination between self and non-self sequences [18]. The recognition of the foreign genetic elements by the Cas-crRNA complex triggers the activation of nuclease activity of Cas proteins which degrade the foreign genome, avoiding the infection [19].
There is a wide diversity of CRISPR/Cas systems that differ in the Cas protein sequences, gene compositions and architecture of the genomic loci. According to Makarova et. al [21] CRISPR/Cas systems can be classified in 2 different classes, 6 types and 33 subtypes, even though this classification is constantly evolving as new systems are being discovered. Class 1 and 2 differ in the number of Cas proteins involved in crRNA processing and interference, being class 2 systems simpler, as they only require one multidomain crRNA-binding protein.
Among all these different systems, CRISPR/Cas9 is the most used for genome engineering technologies because of its properties. This class 2, type II system is capable of cleaving dsDNA with just one Cas protein (Cas9) being necessary for recognition and cleavage of the DNA [22]. Cas9 endonuclease assembles with the crRNA that is interacting with the tracrRNA and binds the dsDNA complementary to crRNA. Furthermore, crRNA and tracrRNA can be engineered as a single RNA chimera (sgRNA) that contains the complementary sequence to the target DNA and drives the complex to the sequence-specific DNA cleavage [23].
Cas9 cleavages the target DNA producing a double strand break (DSB) which can trigger two endogenous DNA repair mechanisms: homologous directed repair (HDR) or nonhomologous end joining (NHEJ) (Figure 1). Both mechanisms are interesting for genome engineering applications [24]. HDR occurs if there is a homologous template, being useful for changing or replacing sequences [25]. In case there is not any homologous template NHEJ is triggered. This mechanism generates small insertions and deletions (indels) in order to ligate the broken ends as fast as possible, leading to the knockout of the gene [25] (Figure 1).
3. Genetic Transformation Technology in Fruit Trees
Transformation of several fruit trees has carried out for many traits and has been improved for a successful genetic transformation so far. Transgenic technology is increasingly used in fruit species to overcome the disadvantages of conventional traditional breeding methods and for gene function research [26]. However, there are several limitations in transformation of fruit trees. Most of fruit trees are recalcitrant for regeneration and/or transformation, the processes are genotype-dependent, long time-consuming compared to other species, and accurate selection with antibiotics or herbicide is necessary to avoid chimeric plants [5]. Furthermore, the lack of available and efficient explants for regeneration and transformation procedures (e.g. seedlings, leaves from micropropagated plants or immature seeds) makes difficult the establishment of effective protocols [27]. Moreover, the public concern and the legislative boundaries on GMO production and commercialization hamper the huge biotechnological potential of the fruit genetic transformation techniques.
Fruit quality improvement, biotic and abiotic tolerance/resistance have been achieved in fruit scion cultivars by direct transformation, but the use of genetically modified rootstocks to confer new characteristics to the non-transformed scion by trans-grafting shows a potential improvement of fruit trees species, in particular those recalcitrant to transformation, and could mitigate public concerns about transgene dispersions or transgenic fruit consumption [27,28]. Additionally, fruit genome editing is a new breeding technology that offers the possibility of producing improved commercial fruit cultivars and could help to address some of the regulatory constraints about the cultivation of first-generation transgenic crops [29].
Agrobacterium tumefaciens (=Rhizobium radiobacter)-mediated transformation is the prevalent method used for genetic engineering of tree fruit crops during more than three decades [30]. Successful transformation has been reported for functional genomics studies and genetic improvement of several fruit crops genotypes by using Agrobacterium infection [27,31].
Although grapevine (Vitis vinifera) is considered as a recalcitrant specie for transformation [30], over the past few years, numerous works have reported the successful transformation of various grape rootstocks and cultivars using Agrobacterium-mediated and or biolistic bombardment techniques [32]. These transformations have involved a range of target genes, such as genes involved in resistance and tolerance against diseases, pests, abiotic stresses, as well as enhancing fruit quality [32,33].
In the same way as with the grapevine, genetic transformation using Agrobacterium tumefaciens is the most used method to obtain transgenic apple (Malus domestica) plants [34]. Genetically modification of apples has been feasible since 1989 [35] and in the following years, most studies were focused on increasing transformation efficiency. Agrobacterium-mediated transformation has become a conventional tool for functional genome studies in apple by overexpression or RNAi-based gene silencing [34]. However, only a select number of research teams have managed to effectively use transformation methods for breeding porpoises. It is remarkable the release of a non-browning transgenic apple, known as Artic® apple, which has been developed using a sense-posttranscriptional silencing of a chimeric polyphenoloxidase gene [36]. The Arctic® apple concept is the result of one breeding program by the biotech company Okanagan Specialty Fruits and currently there are three commercial varieties of Arctic® apple [29]. Notable advancements, particularly in recent years, have broadened the range of tools available to breeders and researchers involved in breeding apple efforts like the establishment of protocols using rapid crop cycles breeding, methods for obtaining marker-free genetically modified plants or the production of cisgenic apple plants (plants that contain genes present in the species or in cross-compatible relatives, but not foreign genes). Regarding applications of targeted gene silencing, in addition to traditional RNAi-based silencing via stable transformation with hairpin gene constructs, powerful technologies have emerged such as optimized protocols for virus-induced gene silencing (VIGS) and artificial microRNAs (amiRNAs). Furthermore, the establishment of methods for successful targeted genomic editing in apple trees has also been achieved [34].
Agrobacterium-mediated transformation of citrus was initially reported by Moore et al. [37] using internodal stem segments as explants followed by regeneration of shoots. Extensive research has resulted in the development of improved Agrobacterium protocols for the genetic modification of citrus plants [38,39]. Due to the difficulties of conventional citrus breeding (a complex reproductive biology, juvenility, a high heterozygosity level), genetic transformation has been considered as a possible alternative strategy for citrus improvement [40]. Modified plants from different citrus species have been generated with resistant to diseases such as huanglongbing and citrus canker caused by bacteria [41,42,43], tristeza disease caused by Citrus tristeza virus [44], and also plants tolerant to different environmental stresses [43].
Although in most of woody fruit species and specially in Prunus species transformation and regeneration are frequently limited to a few genotypes [45], among Prunus, European plum (Prunus domestica L.) is the species most frequently transformed [46]. However, Japanese plum (Prunus salicina L.) transformation has been reported with low efficiency [47]. In the first works, several marker genes were introduced into the plum genome [48,49]. Also protocols for alternative selection marker genes introduction and elimination of marker genes to avoid environmental risks have been reported [50,51]. Regarding the introduced traits in European plum, resistance against pests, diseases and/or abiotic stresses, shorter juvenile period, dwarfing and continuous flowering have been the main objectives [46].
Sharka disease caused by Plum pox virus (PPV) is the most important disease of stone fruit and the establishment of new cultivars resistant to sharka is one of the most focused topics in European plum breeding programs [52]. Among different transgenic strategies used to achieve PPV resistance, successful results have been obtained through applications of RNA silencing techniques [53]. The first PPV-resistant transgenic Prunus was the plum C5 or ‘Honeysweet’, that was obtained through Agrobacterium-mediated transformation of plum hypocotyls slices using a binary plasmid carrying the PPV-CP full-length gene [54]. The resistance of ‘Honeysweet’ was due to the post-transcriptional gene silencing (PTGS) of the coat protein (CP) virus gene [55]. ‘HoneySweet’ and is freely available for fruit production in United States and for use as a source of PPV resistance for developing new PPV-resistant plum cultivars worldwide pending regulatory approval [56]. Different ihpRNA PPV-CP constructs have been designed to obtain new PPV-CP silenced transgenic plum lines [57] or other viral sequences were chosen to induce sharka resistance in European plum [58] demonstrating be effective against a wide range of PPV strains [59,60,61]. Recently, the use of PPV-resistant transgenic plum rootstocks has been proposed as a strategy for conferring virus resistance to recalcitrant to transformation cultivars or species, which could mitigate public concerns about transgenes dispersion and eating transgenic food [28].
Engineered plum lines were produced through interference RNA-mediated silencing of the A. tumefaciens oncogenes ipt and iaaM to study the possibility of generate plum transgenic rootstocks resistant to crown gall disease. Several lines were infected Agrobacterium strains at the greenhouse showing a significant reduction in the development of the disease [62].
The use of transgenic Prunus rootstocks resistant to salinity and/or drought could improve productivity in arid and semi-arid regions affected by environmental stresses. Transgenic European plum lines tolerant to salt stress were obtained by overexpressing cytosolic superoxide dismutase (SOD) from spinach and / or cytosolic ascorbate peroxidase (APX) from pea [63,64]. Modulation of the enzymatic antioxidants and enhancement of non-enzymatic antioxidants like glutathione and ascorbate are responsible of the stress tolerance [64]. Additionally, one transgenic line with high APX activity showed tolerance to severe water stress [65].
European plum has also been transformed with the Flowering Locus T1 (FT1) gene from Populus trichocarpa and transgenic plants that expressed high levels of FT1 flowered and produced fruits in the greenhouse within 1 to 10 months [66]. FT plums showed the ability to continuously produce flowers and fruit regardless of day length or chilling time and survive winter temperatures. For these reasons, FT plums are used in crosses at the USDA ARS facility (Kearneysville, West Virginia, USA) in what has been called “FasTrack” breeding [67]. The “FasTrack" system has allowed minimizing the generation cycle of plums plants from 3-7 years to one year-around, it can be used under greenhouse conditions and the system allows the fast incorporation of important traits into plums.
Apricot (Prunus armeniaca L.) is a very recalcitrant specie with important limitations in regeneration and transformation from explants of juvenile or mature origin. There are several works reporting the production of transgenic apricot plants expressing marker genes the marker genes gfp or uidA and nptII [68,69,70]. Also, the generation of marker-free transgenic apricot plants was achieved by using the regeneration-promoting gene ipt and site-specific recombination site-specific recombination [71] and the chemical-inducible Cre-LoxP system [72].
Although the main goal of transgenic research has been the generation of plants resistant to diseases [73], until now there are very few studies indicating the production of transgenic apricot lines with modified target genes for breeding objectives. In this sense, Laimer da Câmara Machado et al. [74] produced some transgenic apricot lines with the CP of PPV that showed resistant to virus infection. More recently, Alburquerque et al. [62] reported the generation of engineered apricot plants with the A. tumefaciens oncogenes ipt and iaaM silenced, although all transgenic lines were gall susceptible to crown gall disease.
Regarding peach (Prunus persica L.), after the first publication reporting regeneration of transformed plants [75], the lack efficient genetic transformation protocols prevents the application of many biotechnological tools in peach breeding programs like RNA interference, cisgenesis/intragenesis, or genome editing [76].
In other fruit crops like papaya (Carica papaya L.) important challenges has been achieved. In 1992, Papaya ringspot virus (PRSV) was detected in Hawaii. Since the disease caused by PRSV was not completely controlled with conventional methods, local researchers generated transgenic papaya lines that contained the coat protein gene of PRSV, utilizing microprojectile-mediated transformation of immature zygotic embryos of the ‘Sunset’ cultivar. Thus, the transgenic papaya ‘SunUp’ cultivar which is completely resistant to PRSV was established [77].
To the best of our knowledge, most gene-editing studies in fruit trees have been performed via Agrobacterium-mediated transformation to stable knockout of genes associated to major agronomic traits, primary and secondary metabolites production, disease resistances, and for improving breeding purposes using the popular CRISPR/Cas9 system to achieve editing [78,79]. Table 1 summarizes the results of different gene-editing studies performed with fruit trees indicating the CRISPR/Cas delivery technique used.
The polyethylene glycol (PEG)-mediated delivery method has also been employed in fruit trees genome editing system (Table 1), since it is especially useful for these species where the production of transgenic plants is very slow [127]. Nevertheless, this method has not been widely used in genome editing of fruit trees because of low efficiency and the limitations in protoplast preparation, transformation, and regeneration techniques [128]. Malnoy et al. [120] transformed grapevine protoplasts targeted in MLO7 to increase resistance to powdery mildew. PEG-mediated transforming protocols in grapevine have been improved for a better editing and protoplast regeneration [123,124]. Also, Malnoy et al. [120] used the same methodology targeted in DIPM-1, DIPM-2, and DIPM-4 in apple to increase resistance to fire blight disease. PEG-mediated transformation has carried out in other fruit trees like orange (Citrus sinensis (L.) Osbeck) [125,126], banana (Musa spp.) [121] and chestnut (Castanea sativa) [122].
Another transient transformation technique to deliver DNA directly into plant cells is particle bombardment (gene gun-mediated transformation). This technique has been established for several fruit trees transformation protocols. However, particle bombardment is less efficient than Agrobacterium-mediated methods and it is limited due to worse explant regeneration and the destruction of genomic sequences during bombardment [31,128]. To the best of our known, this method is yet to be reported for genome editing in fruit trees.
4. CRISPR/Cas-Mediated Gene Knock-in and Knock-out in Fruit Trees
CRISPR/Cas has been applied to activate and knock out of target genes in different fruit trees species including tree development, yield, fruit quality, and tolerance to biotic and abiotic stresses trying to answer to different challenges (Figure 2).
4.1. Tree Growth and Development
The development and growth of the plant is a crucial factor that will determine the size, density, and ultimately the productivity of a plant. To enhance planting density and subsequently increase productivity while improving nutrient and water use efficiency, the cultivation of dwarf fruit trees has become a prominent strategy [129,130]. Nevertheless, adjusting plant height in these dwarf crops poses challenges that have led to investigations centered on phytohormones and genetic manipulation.
Phytohormones play pivotal roles in plant growth and architecture [131]. Among these, gibberellins (GA) are recognized for their ability to stimulate plant elongation [2]. Disruption of genes involved in GA biosynthesis can result in dwarf plant structures [132]. MaGA20ox2 gene is involved in gibberellic acid biosynthesis and plant height in Gros Michel banana cultivar, CRISPR/Cas9 technology has been successfully used to modify the MaGA20ox2 gene and generate semi-dwarf mutants [92]. Given the high conservation of phytohormone pathways across various plant species, researchers have identified several key regulatory genes within these pathways.
In the cytokinin context, Feng et al. [133] observed changes in gene expression associated with the cytokinin metabolic pathway and trans-zeatin concentration in apple rootstocks, distinguishing between vigorous and dwarf variants. They identified decreased expression of the IPT5b gene, characterized by high methylation levels in the promoter region, leading to impaired trans-zeatin synthesis and potentially causing dwarfism.
In some instances, abnormal levels of abscisic acid (ABA) can contribute to dwarfism in fruit trees by influencing plant growth and development. Pang et al. [119] supported this concept by generating six homozygous mutant pear lines (Pyrus betulifolia) with dwarf phenotypes through Agrobacterium-mediated transformation. These mutants exhibited elevated endogenous ABA levels and increased expression of ABA pathway genes, directly linking ABA to dwarfism. Furthermore, as mentioned in Sattar et al. [134] has been demonstrated the potential for genetic improvement in dwarf pear (Pyrus bretschneideri) through CRISPR/Cas9 technology, modifying the PbPAT14 gene function.
Jia et al. [135] conducted research on the MdNAC1 gene in apple, known to be associated with plant dwarfism. Transgenic plants with overexpressed MdNAC1 displayed dwarf characteristics, including shorter stems and roots, reduced leaf area, and decreased levels of brassinosteroids (BR) and ABA. This implies that MdNAC1 regulates dwarfism by affecting BR and ABA production. Subsequently, another follow-up study [136] revealed increased abundance of the MdKNOX15 gene, part of the apple KNOX transcription factor family, in stems of varieties displaying early flowering dwarfism.
Estrigolactone (SL) is a recently identified plant hormone that plays a pivotal role in branching inhibition in plants. Two key genes involved in SL biosynthesis, CCD7 and CCD8, have been investigated in the grapevine. CRISPR/Cas9 technology was employed, specifically for editing the VvCCD7 and VvCCD8 genes [137]. As a result of these genetic edits, it was observed that the ccd8 mutant exhibited a higher number of branches compared to wild-type plants, highlighting the significance of the VvCCD8 gene in grapevine branching regulation [128].
Additionally, mutations in the CNGC2 gene in Arabidopsis have shown pleiotropic effects, including the accumulation of salicylate compounds and the development of dwarf plants [138]. CNGC2 is also involved in processes such as fertilization and leaf development [139,140], with mutations leading to anomalies in anther length and deficient fertilization. The effectiveness of MdCNGC2 as a target gene for genetic improvement in apple cultivars allows the application of CRISPR/Cas9 technology to induce mutations, consistently resulting in the accumulation of salicylate compounds in apple calluses, showcasing the versatility of this tool in genetic manipulation [84].
These genes offer promising avenues for genetic editing aimed at creating fruit crops with dwarf phenotypes or modified branch architecture, with significant potential to boost productivity and efficiency in fruit tree agriculture.
4.2. Early Flowering
The extended juvenile phase observed in many fruit trees leads to a prolonged non-flowering period, which can span from 3 to 15 years, depending on the specific fruit tree [141]. Elevated levels of terminal flowering protein (TFL) are typically associated with this youthful stage. TFL acts as a negative regulator of flowering by inhibiting the expression of several flowering-stimulating proteins, including FLOWERING LOCUS T (FT), LEAFY (LFY), and APETALA1 (AP1) [142].
To address this challenge, [81] utilized CRISPR/Cas9 technology to edit the Apple TFL1 gene. They employed two different sgRNAs to target the TFL1 gene, and they also used the same construct to edit the pear TFL1. It was observed that early flowering in transgenic pear lines (9%) and transgenic apple lines (93%) targeted the PcTFL1.1 and MdTFL1.1 genes, respectively.
Moreover, CRISPR/Cas9 technology was applied to simultaneously target the AcCEN4 and AcCEN genes in kiwifruit (Actinidia deliciosa). This transformation changed a perennial climbing plant, which produces axillary inflorescences after years of youth, into a compact plant with rapid terminal flowering, all achieved through mutations in the CEN genes [113]. Moreover, in the edited CEN lines, early flowering was dependent on the genetic dosage. In other words, plants mutated with all four alleles of CEN and CEN4 exhibited early flowering compared to plants with fewer allelic mutations [2]. The advantage of modifying a perennial plant like kiwifruit to flower year-round is that it allows for a quick reproductive cycle, producing fruits throughout the year instead of seasonal crops [143].
In trifoliate orange, the overexpression of Citrus unshiu FLOWERING LOCUS T (CiFT) led to early flowering just twelve weeks after being transferred to the greenhouse [144]. Additionally, the overexpression of FT in C. excelsa resulted in early flowering and an accelerated citrus breeding program [145].
4.3. Fruit Growth and Development
CRISPR/Cas holds significant potential for fine-tuning valuable quantitative traits in crop improvement, such as fruit size [148]. Nevertheless, it is crucial to emphasize that the manipulation of specific genes can lead to a wide range of effects. For instance, in the case of the SlKLUH gene, its copy number has been observed to positively correlate with fruit weight. Nonetheless, the deletion or reduction of SlKLUH often results in smaller fruits and can lead to other growth defects, such as smaller inflorescences and sterile flowers [149].
Besides, conserved motifs in the KLUH promoter, related to the promotion of plant organ growth, have also been identified in crops such as pepper and sweet cherry. This suggests that the same approach can be used to engineer changes in fruit and seed size using the KLUH gene in these crops [150]. These findings underscore the genetic editing potential for enhancing specific traits in agricultural crops, while emphasizing the importance of understanding the associated side effects and genetic intricacies in this process.
4.4. Shelf-Life and Fruit Ripening
One crucial aspect of postharvest fruit quality revolves around its shelf-life, in which ethylene plays a pivotal role in both ripening and fruit softening. The fruit shelf-life can be extended by controlling ethylene biosynthesis and signal transduction, as highlighted in recent studies conducted on apricots and plums through the application of ethylene-related chemicals, either by inhibiting or increasing ethylene production [151].
Fruits with a prolonged shelf-life can be achieved by modifying methylation patterns or by silencing key genes involved in ethylene biosynthesis, ripening processes, or their signaling pathways in fruit crops.
To illustrate, the CRISPR/Cas9 gene editing technique was employed to eliminate MaACO1 (aminocyclopropane-1-carboxylate oxidase 1), the gene responsible for encoding the enzyme that converts ACC into ethylene. This genetic modification resulted in an extended fruit shelf-life period of up to 40 days when compared to wild-type bananas [152]. Moreover, the edited MA-ACO1 lines yielded fruits with an increased vitamin C content [153].
4.5. Fruit color, Flavor, and Bioactive Compounds
Genomic editing of key genes offers the potential to design fruit crops with elevated levels of functional metabolites and pigments, which can have a significant impact on enhancing the quality and nutritional characteristics of fruits. Red fruits, rich in various bioactive components and nutrients such as antioxidants, minerals, vitamins, and dietary fiber, have been the subject of research [154]. In this context, CRISPR/Cas9 technology has demonstrated its ability to modulate specific traits in fruits. For instance, editing of MdMKK9 through this technology resulted in a positive increase in anthocyanin expression levels, leading to the characteristic red color in apples [86].
Furthermore, plants can produce various secondary metabolites that play a significant role in growth control, component regeneration, and nutrient enhancement. Carotenoids, for instance, participate in processes like photosynthesis and have antioxidant functions, contributing to attractive colors in fruits and other plant organs [155]. Phytoene desaturase (PDS) has been identified as the rate-limiting enzyme in the carotenoid synthesis pathway [128]. Eliminating PDS using the CRISPR/Cas9 system resulted in mutants with an albino phenotype, underscoring the fundamental importance of this gene in carotenoid synthesis.
Enrichment of bananas with β-carotene was achieved by modifying the lycopene epsilon-cyclase (LCYε) gene using CRISPR/Cas9 [134]. Sequence analysis revealed insertion-deletion mutations in the LCYε gene, significantly increasing β-carotene accumulation without adversely affecting agro-morphological characteristics. In the case of pomegranates (Punica granatum L.), a fruit rich in phenolic metabolites, two UDP-glycosyltransferase genes, PgUGT84A23 and PgUGT84A24, were deleted using CRISPR/Cas9, resulting in a reduction of phenolic anthocyanin content [156].
Additionally, the regulation of tartaric acid (TA) biosynthesis in grapes involves the L-idonate dehydrogenase (IdnDH) gene, which was genetically eliminated using CRISPR/Cas9 [105]. In apples, the MdAAT1 gene has been identified as responsible for ester synthesis [157]. Furthermore, a similar regulatory role of NAC transcription factors in flavor ester formation in apples highlights the importance of NAC1 homologs in different fruit species [158].
4.6. Improving Stress Tolerance in Fruit Trees
Abiotic and biotic stresses, such as drought, extreme temperatures, pests, and diseases, pose substantial threats to global food production. These challenges can result in reduced crop yields, lower crop quality, and economic losses for farmers. To mitigate these issues, the advent of gene editing systems has opened up promising avenues for agriculture. Gene editing techniques like CRISPR/Cas9 enable precise modifications to an organism’s DNA, allowing researchers and breeders to target specific genes associated with stress resistance or desired traits [159].
Nowadays, CRISPR/Cas technology has been harnessed to confer tolerance to various environmental stresses, as demonstrated in recent studies [160]. Within this context, it has been observed that the Dehydration-Responsive Element-Binding (DREB) transcription factor (TF) plays a pivotal role in the regulation of several stress-inducible genes. It has been substantiated that DREB2-type proteins, a subtype of DREB proteins, play a significant role in enhancing drought, salinity, and heat tolerance in a variety of plants, including fruit-bearing trees [161]. For instance, the overexpression of MsDREB6.2 has notably ameliorated drought resistance in transgenic apple lines [162], while the overexpression of AtDREB1b, a cold-inducible TF, has bolstered cold resistance in transgenic grapevines [163].
CRISPR/Cas9 has evolved into an effective tool for introducing robust transcriptional regulatory elements into the promoter region of genes governing the expression of stress-responsive genes. Consequently, this augmentation enhances their expression, thereby fortifying plant stress resilience.
The modulation of stomatal activity in response to abscisic acid (ABA) through the AtMYB60 gene in Arabidopsis thaliana has been shown to exert an influence on drought resistance [164]. In an endeavor to enhance drought resistance in grapevines, CRISPR-Cas9 has been employed to mutate the ortholog of AtMYB60, known as the VvMYB60 gene [82].
Moreover, in related research, the overexpression of the apple spermidine synthase gene has bestowed multiple resistances against environmental stressors in pear trees [165].
The involvement of the Calcium-Dependent Protein Kinase (CDPK) in responses to environmental stressors has been extensively documented by Wang and their team [166]. Overexpression of the apple CDPK gene, referred to as MdCIPK6L, has substantially heightened resistance to saline, osmotic, drought, and cold stresses without compromising root growth [167]. Furthermore, a reduction in MdNPR1 expression has been discerned in apple plants in response to drought conditions [168].
The overexpression of drought-responsive genes, achieved through genetic editing via the CRISPR/Cas9 system, holds promising prospects for the cultivation of high-yield, high-quality fruit trees in arid regions, thereby contributing to the global food supply for the world population [169].
Editing genes that negatively regulate plant immunity is a strategy for obtaining disease-resistant crops. A notable example of this is the elimination of the NPR3 gene, a suppressor of defense responses, which has enhanced the resistance of Theobroma cacao leaves to the Phytophthora tropicalis pathogen [95]. Furthermore, the overexpression of AtNPR1 or its counterparts in other plants such as A. thaliana [170], grapevine [171], and apple [172] has demonstrated increased disease tolerance.
In the specific case of the disease caused by the fungus Botrytis cinerea, the VvWRKY52 gene has been identified as being linked to the biological stress response. Utilizing CRISPR/Cas9 technology, this gene was successfully removed in grapevines (Vitis vinifera), resulting in a significant improvement in tolerance to Botrytis cinerea [128].
On another note, citrus canker disease caused by Xanthomonas spp. poses a serious threat to citrus crops. The CsLOB1 gene has been pinpointed as a key player in citrus canker resistance in Duncan grapes. Through CRISPR/Cas9, this gene was modified in Duncan grapefruits, showing varying levels of resistance to citrus canker [98].
Genes from the MLO (MILDEW RESISTANCE LOCUS) family have been associated with susceptibility to powdery mildew in several plant species [173]. Inactivating these genes has led to increased tolerance to powdery mildew in various species. For instance, mutation of the VvMLO3 [111] and VvMLO7 alleles [82] using CRISPR/Cas9 has conferred greater powdery mildew resistance in grapevines. Furthermore, Dalla Costa et al. [82] designed vectors carrying the CRISPR/Cas9 system to knock-out the genes MdDIPM1 and MdDIPM4, which have a role in the susceptibility to fire blight in apple and evaluated some strategies to eliminate exogenous DNA via site-specific removal mechanisms. The elimination of the EIF4E gene has improved papaya immunity against the Papaya ring spot mosaic virus [174], and the removal of the MdCNGC2 gene in apples has increased resistance to Botryosphaeria dothidea [84]. This underscores the effectiveness of genome editing, utilizing the CRISPR/Cas9 system, in enhancing the resistance of various plants to diseases.
5. Off-Target Issue
One of the main advantages of CRISPR/Cas systems is their specificity, however some off-target editing events can occur leading to undesired modifications. Cas nuclease activity can be triggered even if there is an imperfect complementarity between sgRNA and the off-target genomic site [175], mainly when the mismatches are located far from the PAM sequence [176].
In order to decrease off-target modifications some strategies can be followed. First, it is important to design highly specific gRNAs. For this it is possible to use truncated sgRNAs (tru-gRNAs) that are formed of sequences shorter than 20 nucleotides, which reduces the likelihood of complementarity with mismatches while maintaining on-target efficiency [177]. In A. thaliana tru-gRNAs have been used resulting in no off-target modifications [178].
Cas nucleases also play an important role in specificity. Different natural variants of Cas nucleases, that come from different microorganisms can identify diverse PAM sequences. For example, SaCas9 (from Staphylococcus aureus) has been used to obtain higher on-target efficiency and less off-targets compared to SpCas9 (from Streptococcus pyogenes) in A. thaliana. The higher specificity of SaCas9 is due to the requirement of a longer PAM sequence. However, this also results in a lower number of potential target sites [179]. It is also possible to improve Cas nucleases specificity by protein engineering. Slaymaker et al. [176] modified amino acids that are crucial for the interaction between the sgRNA-Cas and the target sequence, obtaining a more specific Cas called eSpCas9 (enhanced specificity) with great results in rice [180].
Another approach is to use paired nCas9 (nickase Cas9). nCas9 is mutated in one of its nuclease domains, therefore it makes a single-stranded break in the DNA. This way, two nCas that are leaded by two paired sgRNAs recognizing the target sequence are necessary to create a double-stranded break, doubling the recognition site [181].
Although different strategies have been described to minimize the problem of off-targets, so far they have not been used in fruit trees. Therefore, it would be interesting to explore its application in this type of crops.
6. Regulatory Limits of Genome Editing
Despite the numerous advantages that genome editing technologies offer to obtain improved fruit trees compared to conventional breeding techniques [134], the legislative limitations that regulate agricultural production frequently hampered the important potential of these biotechnological tools [82]. The regulatory regimes applied to GMO in each country are different, being more permissive in North- and South-America, Australia, and certain parts of Asia. Other countries such as Japan, New Zealand, Norway, Switzerland and the European Union (EU) set up more restrictive regimes and the number of approvals for GMO cultivation and commercial use have been strictly limited [182]. In particular, the current EU legislation of GMOs and derived products for food is based on Directive 2001/18/EC and Regulations 1829/2003 and 1830/2003, which indicate that the authorization regime of a GMO event requires an environmental and human health risk assessment.
The New Genomic Techniques (NGTs), that include targeted mutagenesis and cisgenesis or intragenesis, provide new opportunities to alter the genetic material in a different way from established genomic techniques, with higher precision and speed in introducing the chosen genetic modifications only from a crossable species. Certain targeted mutated crops are indistinguishable from the original plant cultivar, natural mutations or from genetic modifications introduced by conventional breeding techniques.
Nowadays an increasing number of countries have adapted the regulatory status of genome edited plants, by releasing some of the editing technologies from the conventional GMO regulation [183]. This is the case of important exporters of agricultural products countries such as United States, Canada, Brazil, Argentina and Australia [182,184].
On the other hand, in July 2018 the European Court of Justice (ECJ) ruled that plants obtained by NTGs are to be considered as GMOs and as such must comply with the regulations contained in the Directive 2001/18/EC concerning the approval of GMOs [82,183]. However, the organisms obtained through conventional chemical or radiation induced random mutagenesis methods are excluded from the scope of the Directive [185]. After the ECJ ruling, a policy debate within the EU was initiated. In 2019 the Council of the European Union requested the European Commission to conduct a study on the impact of the ECJ ruling considering the technical status of novel genomic techniques, ethics and the views of the EU countries and stakeholders [185]. In 2021 this study concluded that the EU legislation is not fit to regulate plants obtained with some NGTs and that the current legislation needs to be adapted to scientific and technical progress in this area [182,183]. Based on this study, the European Commission (EC) suggested a revision and promotes citizen, stakeholder and Member State consultations in 2022.
Recently, as a result of this consultation, the EC published a proposal (5 July 2023) for a ‘Regulation of the European Parliament and of the Council on plants obtained by certain new genomic techniques and their food and feed, and amending Regulation (EU) 2017/625’. This proposal of Regulation only affects plants obtained by targeted mutagenesis and cisgenesis (including intragenesis), that do not carry genetic material from non-crossable species, and establishes two categories of plants obtained by NGTs: plants comparable to naturally occurring or conventional plants (category 1 NTG), and plants with modifications that are more complex (category 2 NTG). The category 1 NGT plants and their progeny obtained by conventional breeding techniques have comparable risk to conventionally bred plants, and therefore it is proposed to completely derogate the Union legislation on GMOs and GMO-related requirements in sectoral legislation. The recent Regulation must to be adopted by the EU Member States in the Council and the European Parliament, following the ordinary legislative procedure to become a law.
Genetically edited plants that can benefit from this possible law must not have any element of the CRISPR systems, therefore, the production of transgene-free genome-edited plants is paramount, particularly in the case of fruit crops, due to their long growth cycle that make difficult the elimination of CRISPR elements by successive crossing. Although there are several methods to generate this type of plants like delivering of CRISPR components in the form of mRNA or ribonucleoprotein complexes, the use of mRNA or ribonucleoprotein biolistic delivery, PEG-mediated transformation followed by protoplast regeneration [186], they have been successfully reported only in herbaceous crops [187,188,189,190].
7. Conclusions
Successful genome editing studies in fruit trees show that CRISPR/Cas can induce changes in target genes. In the future, the application of gene editing could allow the development of new generations of fruit crops with improved traits by targeting different genetic segments or even could facilitate the introduction of traits in elite cultivars without changing other traits. Although the use of genome edition techniques promises a quick, easy and inexpensive way to develop novel crop varieties with improved traits than before, in the case of fruit crops the scarcity of efficient regeneration and transformation protocols in some species, the fact that many of those procedures are the genotype-dependent or the convenience of segregating the transgenic parts of the CRISPR system are some of the problems that limit the potential of genetic editing techniques for fruit trees. More research is needed to establish efficient regeneration and transformation protocols for fruit trees species. Transgrafting technique using CRISPR-transgenic rootstocks with resistance to root diseases, drought or salt stress for transmission of desired traits to scion may be an alternative strategy to obtain non-transgenic fruit with improved characteristics. The methods for producing transgene-free genome-edited plants or those that avoid the need to remove CRISPR DNA from edited plants still require lengthy tissue culture, and they are not applicable to many species recalcitrant for genetic transformation and regeneration, including most fruit trees. Therefore, the optimization and adaptation of these method is a challenge in these crops. Finally, the new perspective of the society together with the possible changes in the legislative framework pave the way to raise public and regulatory concern about the use of NTG plants including fruit crops.
Author Contributions
Conceptualization, M.M.V., S.R.D. and N.A.; writing—review and editing, M.M.V., S.R.D., G.O.H., C.P.C., S.M.E.M., G.B., J.A.S., P.M.G. and N.A. All authors have read and agreed to the published version of the manuscript.
Funding
This study is part of the AGROALNEXT Program that has been financed by MCIN with NextGenerationEU funds (PRTR-C17.I1) and by the Séneca Foundation with funds from the Autonomous Community of the Region of Murcia (CARM).
Informed Consent Statement
No applicable.
Data Availability Statement
No applicable.
Acknowledgments
Authors thank Mansureh Tamizgran and Khalil Rahimi Devin for their grammatical corrections and valuable suggestions on the manuscript. C.P.C. acknowledges the “Ministerio Español de Ciencia, Innovación y Universidad” for the support though a “Formación de Profesorado Universitario” grant (FPU19/03767). G.O.H. acknowledges Fundación Séneca (Región de Murcia, Spain) for the support through the “21788/FPI/22” fellowship. M.M.V. acknowledges the “Consejo Superior de Investigaciones Científicas” for the support through the JAEINT_22_00399 fellowship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Khan, A.; Korban, S.S. Breeding and Genetics of Disease Resistance in Temperate Fruit Trees: Challenges and New Opportunities. Theor. Appl. Genet. 2022, 135, 3961–3985. [Google Scholar] [CrossRef]
- Savadi, S.; Mangalassery, S.; Sandesh, M.S. Advances in Genomics and Genome Editing for Breeding next Generation of Fruit and Nut Crops. Genomics 2021, 113. [Google Scholar] [CrossRef] [PubMed]
- Vinholes, J.; D, G.P.; Vizzotto, M. Stone Fruits as a Source of Bioactive Compounds. In Natural Bioactive Compounds from Fruits and Vegetables as Health Promoters Part II; Silva, L.R., Silva, B., Eds.; Bentham Science: Covilha-Portugal, 2016; pp. 110–142. [Google Scholar]
- Scorza, R.; Callahan, A.; Dardick, C.; Ravelonandro, M.; Polak, J.; Malinowski, T.; Zagrai, I.; Cambra, M.; Kamenova, I. Genetic Engineering of Plum Pox Virus Resistance:‘HoneySweet’Plum—from Concept to Product. Plant Cell, Tissue Organ Cult. 2013, 115, 1–12. [Google Scholar] [CrossRef]
- Alburquerque, N.; Baldacci-Cresp, F.; Baucher, M.; Casacuberta, J M Collonnier, C.; El Jaziri, M.; Nogué, F.; Burgos, L. New Transformation Technologies for Trees. In Biosafety of Forest Transgenic Trees: Improving the Scientific Basis for Safe Tree Development and Implementation of EU Policy Directives; Vettori, C., Gallardo, F., Kazana, V., Häggman, H., Migliacci, F., Pilate, G., Fladung, M., Eds.; Springer: Dordrecht, 2016; pp. 31–66.
- Hanin, M.; Paszkowski, J. Plant Genome Modification by Homologous Recombination. Curr. Opin. Plant Biol. 2003, 6, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Wehrkamp-Richter, S.; Degroote, F.; Laffaire, J.B.; Paul, W.; Perez, P.; Picard, G. Characterisation of a New Reporter System Allowing High Throughput in Planta Screening for Recombination Events before and after Controlled DNA Double Strand Break Induction. Plant Physiol. Biochem. 2009, 47. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas Genome Editing and Precision Plant Breeding in Agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Puchta, H.; Fauser, F. Synthetic Nucleases for Genome Engineering in Plants: Prospects for a Bright Future. Plant J. 2014, 78, 727–741. [Google Scholar] [CrossRef]
- Abdallah, N.A.; Prakash, C.S.; McHughen, A.G. Genome Editing for Crop Improvement: Challenges and Opportunities. GM Crops Food 2015, 6, 183–205. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-Based Methods for Genome Engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef]
- Puchta, H. Applying CRISPR/Cas for Genome Engineering in Plants: The Best Is yet to Come. Curr. Opin. Plant Biol. 2017, 36, 1–8. [Google Scholar] [CrossRef]
- Wada, N.; Osakabe, K.; Osakabe, Y. Expanding the Plant Genome Editing Toolbox with Recently Developed CRISPR–Cas Systems. Plant Physiol. 2022, 188, 1825–1837. [Google Scholar] [CrossRef]
- Wada, N.; Ueta, R.; Osakabe, Y.; Osakabe, K. Precision Genome Editing in Plants: State-of-the-Art in CRISPR/Cas9-Based Genome Engineering. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Denes, C.E.; Cole, A.J.; Aksoy, Y.A.; Li, G.; Neely, G.G.; Hesselson, D. Approaches to Enhance Precise Crispr/Cas9-mediated Genome Editing. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, Z.; Unver, T.; Zhang, B. CRISPR/Cas: A Powerful Tool for Gene Function Study and Crop Improvement. J. Adv. Res. 2021, 29, 207–221. [Google Scholar] [CrossRef]
- Wang, J.Y.; Pausch, P.; Doudna, J.A. Structural Biology of CRISPR–Cas Immunity and Genome Editing Enzymes. Nat. Rev. Microbiol. 2022, 20, 641–656. [Google Scholar] [CrossRef]
- Jiang, F.; Doudna, J.A. CRISPR-Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46. [Google Scholar] [CrossRef]
- Amitai, G.; Sorek, R. CRISPR-Cas Adaptation: Insights into the Mechanism of Action. Nat. Rev. Microbiol. 2016, 14. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Beisel, C.L. The TracrRNA in CRISPR Biology and Technologies. Annu. Rev. Genet. 2021, 55. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary Classification of CRISPR–Cas Systems: A Burst of Class 2 and Derived Variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Knott, G.J.; Doudna, J.A. CRISPR-Cas Guides the Future of Genetic Engineering. Science (80-. ). 2018, 361. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science (80-. ). 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Hryhorowicz, M.; Lipiński, D.; Zeyland, J.; Słomski, R. CRISPR/Cas9 Immune System as a Tool for Genome Engineering. Arch. Immunol. Ther. Exp. (Warsz). 2017, 65. [Google Scholar] [CrossRef]
- Yin, K.; Gao, C.; Qiu, J.L. Progress and Prospects in Plant Genome Editing. Nat. Plants 2017, 3. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, C.; Xiao, D.; Liang, Y.; Wang, Y. Advances and Perspectives of Transgenic Technology and Biotechnological Application in Forest Trees. Front. Plant Sci. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Song, G.Q.; Prieto, H.; Orbovic, V. Agrobacterium-Mediated Transformation of Tree Fruit Crops: Methods, Progress, and Challenges. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Alburquerque, N.; Pérez-Caselles, C.; Faize, L.; Ilardi, V.; Burgos, L. Effective Transfer of Plum Pox Virus Resistance from Transgenic Plum Rootstocks to Apricot Scions. 2023, 1–11. [CrossRef]
- Lobato-Gómez, M.; Hewitt, S.; Capell, T.; Christou, P.; Dhingra, A.; Girón-Calva, P.S. Transgenic and Genome-Edited Fruits: Background, Constraints, Benefits, and Commercial Opportunities. Hortic. Res. 2021, 8. [Google Scholar] [CrossRef]
- Orbovic, V.; Prieto, H. Editorial: New Developments in Agrobacterium Mediated Transformation of Tree Fruit Crops, Volume II. Front. Plant Sci. 2023, 14, 1–2. [Google Scholar] [CrossRef]
- Wang, X.; Tu, M.; Li, Z.; Wang, Y.; Wang, X. Current Progress and Future Prospects for the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) Genome Editing Technology in Fruit Tree Breeding. CRC. Crit. Rev. Plant Sci. 2018, 37, 233–258. [Google Scholar] [CrossRef]
- Zhang, X. ming; Wu, Y. fei; Li, Z.; Song, C. bing; Wang, X. ping Advancements in Plant Regeneration and Genetic Transformation of Grapevine (Vitis Spp.). J. Integr. Agric. 2021, 20, 1407–1434. [Google Scholar] [CrossRef]
- Campos, G.; Chialva, C.; Miras, S.; Lijavetzky, D. New Technologies and Strategies for Grapevine Breeding Through Genetic Transformation. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Schröpfer, S.; Lempe, J.; Emeriewen, O.F.; Flachowsky, H. Recent Developments and Strategies for the Application of Agrobacterium-Mediated Transformation of Apple Malus × Domestica Borkh. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- James, D.J.; Passey, A.J.; Barbara, D.J.; Bevan, M. Genetic Transformation of Apple (Malus Pumila Mill.) Using a Disarmed Ti-Binary Vector. Plant Cell Rep. 1989, 7, 658–661. [Google Scholar] [CrossRef]
- Stowe, E.; Dhingra, A. Development of the Arctic® Apple. Plant Breed. Rev. 2021, 44, 273–296. [Google Scholar] [CrossRef]
- Moore, G.A.; Jacono, C.C.; Neidigh, J.L.; Lawrence, S.D.; Cline, K. And Regeneration of Transgenic Plants. 1992, 238–242.
- Cervera, M.; Juárez, J.; Navarro, A.; Pina, J.A.; Durán-Vila, N.; Navarro, L.; Peña, L. Genetic Transformation and Regeneration of Mature Tissues of Woody Fruit Plants Bypassing the Juvenile Stage. Transgenic Res. 1998, 7, 51–59. [Google Scholar] [CrossRef]
- Dutt, M.; Grosser, J.W. Evaluation of Parameters Affecting Agrobacterium-Mediated Transformation of Citrus. Plant Cell. Tissue Organ Cult. 2009, 98, 331–340. [Google Scholar] [CrossRef]
- Peña, L.; Navarro L IV Transgenic Citrus. In Biotechnology in Agriculture and Forestry, Vol. 44. ransgenic trees; Bajaj Y P S, Ed.; Springer Berlin Heidelberg: Berlin, Heidelberg, 1999; pp. 39–54.
- Alquézar, B.; Carmona, L.; Bennici, S.; Peña, L. Engineering of Citrus to Obtain Huanglongbing Resistance. Curr. Opin. Biotechnol. 2021, 70, 196–203. [Google Scholar] [CrossRef]
- Soares, J.M.; Tanwir, S.E.; Grosser, J.W.; Dutt, M. Development of Genetically Modified Citrus Plants for the Control of Citrus Canker and Huanglongbing. Trop. Plant Pathol. 2020, 45, 237–250. [Google Scholar] [CrossRef]
- Nehela, Y.; Killiny, N. The Unknown Soldier in Citrus Plants: Polyamines-Based Defensive Mechanisms against Biotic and Abiotic Stresses and Their Relationship with Other Stress-Associated Metabolites. Plant Signal. Behav. 2020, 15. [Google Scholar] [CrossRef]
- Cervera, M.; Esteban, O.; Gil, M.; Gorris, M.T.; Martínez, M.C.; Peña, L.; Cambra, M. Transgenic Expression in Citrus of Single-Chain Antibody Fragments Specific to Citrus Tristeza Virus Confers Virus Resistance. Transgenic Res. 2010, 19, 1001–1015. [Google Scholar] [CrossRef]
- Petri, C.; Burgos, L. Transformation of Fruit Trees. Useful Breeding Tool or Continued Future Prospect? Transgenic Res. 2005, 14. [Google Scholar] [CrossRef]
- Petri, C.; Alburquerque, N.; Faize, M.; Scorza, R.; Dardick, C. Current Achievements and Future Directions in Genetic Engineering of European Plum (Prunus domestica L.). Transgenic Res. 2018, 27, 225–240. [Google Scholar] [CrossRef]
- Urtubia, C.; Devia, J.; Castro, A.; Zamora, P.; Aguirre, C.; Tapia, E.; Barba, P.; Dell’Orto, P.; Moynihan, M.R.; Petri, C.; et al. Agrobacterium-Mediated Genetic Transformation of Prunus salicina. Plant Cell Rep. 2008, 27, 1333–1340. [Google Scholar] [CrossRef]
- Gonzalez-Padilla, I.M.; Webb, K.; Scorza, R. Early Antibiotic Selection and Efficient Rooting and Acclimatization Improve the Production of Transgenic Plum Plants (Prunus domestica L.). Plant Cell Rep. 2003, 22, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Canli, F.A.; Wang, X.; Sibbald, S. Genetic Transformation of Prunus domestica L. Using the Hpt Gene Coding for Hygromycin Resistance as the Selectable Marker. Sci. Hortic. 2009, 119, 339–343. [Google Scholar] [CrossRef]
- Petri, C.; Hily, J.M.; Vann, C.; Dardick, C.; Scorza, R. A High-Throughput Transformation System Allows the Regeneration of Marker-Free Plum Plants (Prunus domestica). Ann. Appl. Biol. 2011, 159, 302–315. [Google Scholar] [CrossRef]
- Wang, H.; Petri, C.; Burgos, L.; Alburquerque, N. Phosphomannose-Isomerase as a Selectable Marker for Transgenic Plum (Prunus Domestica L.). Plant Cell. Tissue Organ Cult. 2013, 113. [Google Scholar] [CrossRef]
- Petri, C.; Ruiz, D.; Faize, M.; Burgos, L.; Alburquerque, N. Prunus Domestica Plum. In Biotechnology of fruit and nut crops; Litz, R.E., Pliego-Alfaro, F., Hormaza, J.I., Eds.; CABI: Florida, 2020; pp. 512–531. [Google Scholar]
- Ilardi, V.; Tavazza, M. Biotechnological Strategies and Tools for Plum Pox Virus Resistance: Trans-, Intra-, Cis-Genesis, and Beyond. Front. Plant Sci. 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Scorza, R.; Ravelonandro, M.; Callahan, A.M.; Cordts, J.M.; Fuchs, M.; Dunez, J.; Gonsalves, D. Transgenic Plums (Prunus domestica L.) Express the Plum Pox Virus Coat Protein Gene. Plant Cell Rep. 1994, 14, 18–22. [Google Scholar] [CrossRef]
- Scorza, R.; Callahan, A.; Levy, L.; Damsteegt, V.; Webb, K.; Ravelonandro, M. Post-Transcriptional Gene Silencing in Plum Pox Virus Resistant Transgenic European Plum Containing the Plum Pox Potyvirus Coat Protein Gene. Transgenic Res. 2001, 10, 201–209. [Google Scholar] [CrossRef]
- Scorza, R.; Ravelonandro, M.; Callahan, A.; Zagrai, L.; Polak, J.; Malinowski, T.; Cambra, M.; Levy, L.; Damsteegt, V.; Krška, B.; et al. Honeysweet (C5), the First Genetically Engineered Plum Pox virus-Resistant Plum (Prunus domestica L.) Cultivar. HortScience 2016, 51, 601–603. [Google Scholar] [CrossRef]
- Hily, J.M.; Ravelonandro, M.; Damsteegt, V.D.; Bassett, C.; Petri, C.; Liu, Z.; Scorza, R. Plum Pox Virus Coat Protein Gene Intron-Hairpin-RNA (IhpRNA) Constructs Provide Resistance to Plum Pox Virus in Nicotiana benthamiana and Prunus domestica. J. Am. Soc. Hortic. Sci. 2007, 132, 850–858. [Google Scholar] [CrossRef]
- Di Nicola-Negri, E. Di; Brunetti, A.; Tavazza, M.; Ilardi, V. Hairpin RNA-Mediated Silencing of Plum Pox Virus P1 and HC-Pro Genes for Efficient and Predictable Resistance to the Virus. Transgenic Res. 2005, 14, 989–994. [Google Scholar] [CrossRef]
- Di Nicola-Negri, E.; Tavazza, M.; Salandri, L.; Ilardi, V. Silencing of Plum Pox Virus 5’UTR/P1 Sequence Confers Resistance to a Wide Range of PPV Strains. Plant Cell Rep. 2010, 29, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Monticelli, S.; Di Nicola-Negri, E.; Gentile, A.; Damiano, C.; Ilardi, V. Production and in Vitro Assessment of Transgenic Plums for Resistance to Plum Pox Virus: A Feasible, Environmental Risk-Free, Cost-Effective Approach. Ann. Appl. Biol. 2012, 161, 293–301. [Google Scholar] [CrossRef]
- García-Almodovar, R.C.; Clemente-Moreno, M.J.; Díaz-Vivancos, P.; Petri, C.; Rubio, M.; Padilla, I.M.G.; Ilardi, V.; Burgos, L. Greenhouse Evaluation Confirms in Vitro Sharka Resistance of Genetically Engineered H-UTR/P1 Plum Plants. Plant Cell. Tissue Organ Cult. 2015, 120, 791–796. [Google Scholar] [CrossRef]
- Alburquerque, N.; Faize, L.; Burgos, L. Silencing of Agrobacterium Tumefaciens Oncogenes Ipt and IaaM Induces Resistance to Crown Gall Disease in Plum but Not in Apricot. Pest Manag. Sci. 2017, 73. [Google Scholar] [CrossRef]
- Faize, M.; Faize, L.; Petri, C.; Barba-Espin, G.; Diaz-Vivancos, P.; Clemente-Moreno, M.J.; Koussa, T.; Rifai, L.A.; Burgos, L.; Hernandez, J.A. Cu/Zn Superoxide Dismutase and Ascorbate Peroxidase Enhance in Vitro Shoot Multiplication in Transgenic Plum. J. Plant Physiol. 2013, 170, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Vivancos, P.; Faize, M.; Barba-Espin, G.; Faize, L.; Petri, C.; Hernández, J.A.; Burgos, L. Ectopic Expression of Cytosolic Superoxide Dismutase and Ascorbate Peroxidase Leads to Salt Stress Tolerance in Transgenic Plums. Plant Biotechnol. J. 2013, 11, 976–985. [Google Scholar] [CrossRef]
- Díaz-Vivancos, P.; Faize, L.; Nicolás, E.; Clemente-Moreno, M.J.; Bru-Martinez, R.; Burgos, L.; Hernández, J.A. Transformation of Plum Plants with a Cytosolic Ascorbate Peroxidase Transgene Leads to Enhanced Water Stress Tolerance. Ann. Bot. 2016, 117, 1121–1131. [Google Scholar] [CrossRef]
- Srinivasan, C.; Dardick, C.; Callahan, A.; Scorza, R. Plum (Prunus domestica) Trees Transformed with Poplar FT1 Result in Altered Architecture, Dormancy Requirement, and Continuous Flowering. PLoS One 2012, 7, 1–11. [Google Scholar] [CrossRef]
- Scorza, R.; Dardick, C.D.; Callahan, A.M.; Srinivasan, C.; Delong, T.; Harper, J.; Raines, D.D.; Castro, S. FasTrack’-a Revolutionary Approach to Long-Generation Cycle Specialty Crop Breeding. In Proceedings of the Xth International Symposium Plum & Prune Genetics; 2012; p. Paper No. 101.
- Petri, C.; Wang, H.; Alburquerque, N.; Faize, M.; Burgos, L. Agrobacterium-Mediated Transformation of Apricot (Prunus armeniaca L.) Leaf Explants. Plant Cell Rep. 2008, 27. [Google Scholar] [CrossRef] [PubMed]
- Petri, C.; López-Noguera, S.; Alburquerque, N.; Egea, J.; Burgos, L. An Antibiotic-Based Selection Strategy to Regenerate Transformed Plants from Apricot Leaves with High Efficiency. Plant Sci. 2008, 175. [Google Scholar] [CrossRef]
- Petri, C.; Wang, H.; Burgos, L.; Sánchez-Navarro, J.; Alburquerque, N. Production of Transgenic Apricot Plants from Hypocotyl Segments of Mature Seeds. Sci. Hortic. 2015, 197. [Google Scholar] [CrossRef]
- López-Noguera, S.; Petri, C.; Burgos, L. Combining a Regeneration-Promoting Gene and Site-Specific Recombination Allows a More Efficient Apricot Transformation and the Elimination of Marker Genes. Plant Cell Rep. 2009, 28, 1781–1790. [Google Scholar] [CrossRef]
- Petri, C.; López-Noguera, S.; Wang, H.; García-Almodóvar, C.; Alburquerque, N.; Burgos, L. A Chemical-Inducible Cre-LoxP System Allows for Elimination of Selection Marker Genes in Transgenic Apricot. Plant Cell. Tissue Organ Cult. 2012, 110, 337–346. [Google Scholar] [CrossRef]
- Alburquerque, N.; Ruiz, D.; Burgos, L.; Petri, C. Prunus armeniaca Apricot. In Biotechnology of Fruit and Nut Crops; Litz, R.E., Pliego-Alfaro, F., Hormaza, J.I., Eds.; CABI: Florida, 2020; pp. 496–511. [Google Scholar]
- Laimer da Câmara Machado, M.; da Câmara Machado, A.; Hanzer, V.; Weiss, H.; Regner, F.; Steinkeliner, H.; Mattanovich, D.; Plail, R.; Knapp, E.; Kalthoff, B.; et al. Regeneration of Transgenic Plants of Prunus armeniaca Containing the Coat Protein Gene of Plum Pox Virus. Plant Cell Rep. 1992, 11, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Smigocki, A.C.; Hammerschlag, F.A. Regeneration of Plants from Peach Embryo Cells Infected with a Shooty Mutant Strain of Agrobacterium. J. Am. Soc. Hortic. Sci. 1991, 116, 1092–1097. [Google Scholar] [CrossRef]
- Ricci, A.; Sabbadini, S.; Prieto, H.; Padilla, I.M.G.; Dardick, C.; Li, Z.; Scorza, R.; Limera, C.; Mezzetti, B.; Perez-Jimenez, M.; et al. Genetic Transformation in Peach (Prunus persica l.): Challenges and Ways Forward. Plants 2020, 9, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Fitch, M.M.M.; Manshardt, R.M.; Gonsalves, D.; Slightom, J.L.; Sanford, J.C. Virus Resistant Papaya Plants Derived from Tissues Bombarded with the Coat Protein Gene of Papaya Ringspot Virus. Bio/Technology 1992, 10, 1466–1472. [Google Scholar] [CrossRef]
- Gouthu, S.; Mandelli, C.; Eubanks, B.A.; Deluc, L.G. Transgene-Free Genome Editing and RNAi Ectopic Application in Fruit Trees: Potential and Limitations. Front. Plant Sci. 2022, 13, 1–13. [Google Scholar] [CrossRef]
- Fernandez i Marti, A.F.; Dodd, R.S. Using CRISPR as a Gene Editing Tool for Validating Adaptive Gene Function in Tree Landscape Genomics. Front. Ecol. Evol. 2018, 6, 1–7. [Google Scholar] [CrossRef]
- Nishitani, C.; Hirai, N.; Komori, S.; Wada, M.; Okada, K.; Osakabe, K.; Yamamoto, T.; Osakabe, Y. Efficient Genome Editing in Apple Using a CRISPR/Cas9 System. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Charrier, A.; Vergne, E.; Dousset, N.; Richer, A.; Petiteau, A.; Chevreau, E. Efficient Targeted Mutagenesis in Apple and First Time Edition of Pear Using the CRISPR-Cas9 System. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Dalla Costa, L.; Piazza, S.; Pompili, V.; Salvagnin, U.; Cestaro, A.; Moffa, L.; Vittani, L.; Moser, C.; Malnoy, M. Strategies to Produce T-DNA Free CRISPRed Fruit Trees via Agrobacterium tumefaciens Stable Gene Transfer. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Malabarba, J.; Chevreau, E.; Dousset, N.; Veillet, F.; Moizan, J.; Vergne, E. New Strategies to Overcome Present CRISPR/CAS9 Limitations in Apple and Pear: Efficient Dechimerization and Base Editing. Int. J. Mol. Sci. 2021, 22. [Google Scholar] [CrossRef]
- Zhou, H.; Bai, S.; Wang, N.; Sun, X.; Zhang, Y.; Zhu, J.; Dong, C. CRISPR/Cas9-Mediated Mutagenesis of MdCNGC2 in Apple Callus and VIGS-Mediated Silencing of MdCNGC2 in Fruits Improve Resistance to Botryosphaeria Dothidea. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, P.; Bozorov, T.A.; Zhang, D. Application of CRISPR/Cas9 Technology in Wild Apple (Malus sieverii) for Paired Sites Gene Editing. Plant Methods 2021, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, X.; Wang, Y.; Xu, J.; Jiang, S.; Zhang, Y. MdMKK9-Mediated the Regulation of Anthocyanin Synthesis in Red-Fleshed Apple in Response to Different Nitrogen Signals. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Kong, K.; Han, J.; Song, S.; Bai, T.; Song, C.; Wang, M.; Yan, Z.; Zhang, H.; Zhang, R.; et al. Field Detection of Multiple RNA Viruses/Viroids in Apple Using a CRISPR/Cas12a-Based Visual Assay. Plant Biotechnol. J. 2021, 19. [Google Scholar] [CrossRef]
- Kaur, N.; Alok, A.; Shivani; Kaur, N.; Pandey, P.; Awasthi, P.; Tiwari, S. CRISPR/Cas9-Mediated Efficient Editing in Phytoene Desaturase (PDS) Demonstrates Precise Manipulation in Banana Cv. Rasthali Genome. Funct. Integr. Genomics 2018, 18. [Google Scholar] [CrossRef]
- Naim, F.; Dugdale, B.; Kleidon, J.; Brinin, A.; Shand, K.; Waterhouse, P.; Dale, J. Gene Editing the Phytoene Desaturase Alleles of Cavendish Banana Using CRISPR/Cas9. Transgenic Res. 2018, 27, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Ntui, V.O.; Tripathi, J.N.; Tripathi, L. Robust CRISPR/Cas9 Mediated Genome Editing Tool for Banana and Plantain (Musa Spp.). Curr. Plant Biol. 2020, 21. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 Editing of Endogenous Banana Streak Virus in the B Genome of Musa Spp. Overcomes a Major Challenge in Banana Breeding. Commun. Biol. 2019, 2. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Wu, S.; Dou, T.; Zhu, H.; Hu, C.; Huo, H.; He, W.; Deng, G.; Sheng, O.; Bi, F.; et al. Using CRISPR/Cas9 Genome Editing System to Create MaGA20ox2 Gene-Modified Semi-Dwarf Banana. Plant Biotechnol. J. 2020, 18. [Google Scholar] [CrossRef]
- Kaur, N.N.; Alok, A.; Shivani; Kumar, P.; Kaur, N.N.; Awasthi, P.; Chaturvedi, S.; Pandey, P.; Pandey, A.K.A.; Pandey, A.K.A.; et al. CRISPR/Cas9 Directed Editing of Lycopene Epsilon-Cyclase Modulates Metabolic Flux for β-Carotene Biosynthesis in Banana Fruit. Metab. Eng. 2020, 59, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Sheng, O.; Deng, G.; He, W.; Dong, T.; Yang, Q.; Dou, T.; Li, C.; Gao, H.; Liu, S.; et al. CRISPR/Cas9-Mediated Genome Editing of MaACO1 (Aminocyclopropane-1-Carboxylate Oxidase 1) Promotes the Shelf Life of Banana Fruit. Plant Biotechnol. J. 2021, 19. [Google Scholar] [CrossRef] [PubMed]
- Fister, A.S.; Landherr, L.; Maximova, S.N.; Guiltinan, M.J. Transient Expression of CRISPR/Cas9 Machinery Targeting TcNPR3 Enhances Defense Response in Theobroma Cacao. Front. Plant Sci. 2018, 9, 268. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Nian, W. Targeted Genome Editing of Sweet Orange Using Cas9/SgRNA. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Jia, H.; Xu, J.; Orbović, V.; Zhang, Y.; Wang, N. Editing Citrus Genome via SaCas9/SgRNA System. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome Editing of the Disease Susceptibility Gene CsLOB1 in Citrus Confers Resistance to Citrus Canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering Canker-Resistant Plants through CRISPR/Cas9-Targeted Editing of the Susceptibility Gene CsLOB1 Promoter in Citrus. Plant Biotechnol. J. 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Orbović, V.; Wang, N. CRISPR-LbCas12a-Mediated Modification of Citrus. Plant Biotechnol. J. 2019, 17. [Google Scholar] [CrossRef]
- Wang, L.; Chen, S.; Peng, A.; Xie, Z.; He, Y.; Zou, X. CRISPR/Cas9-Mediated Editing of CsWRKY22 Reduces Susceptibility to Xanthomonas Citri Subsp. Citri in Wanjincheng Orange (Citrus sinensis (L.) Osbeck). Plant Biotechnol. Rep. 2019, 13, 501–510. [Google Scholar] [CrossRef]
- Jia, H.; Wang, N. Generation of Homozygous Canker-Resistant Citrus in the T0 Generation Using CRISPR-SpCas9p. Plant Biotechnol. J. 2020, 18. [Google Scholar] [CrossRef]
- Dutt, M.; Mou, Z.; Zhang, X.; Tanwir, S.E.; Grosser, J.W. Efficient CRISPR/Cas9 Genome Editing with Citrus Embryogenic Cell Cultures. BMC Biotechnol. 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, Y.; Wang, N. Highly Efficient Generation of Canker-Resistant Sweet Orange Enabled by an Improved CRISPR/Cas9 System. Front. Plant Sci. 2022, 12. [Google Scholar] [CrossRef]
- Ren, C.; Liu, X.; Zhang, Z.; Wang, Y.; Duan, W.; Li, S.; Liang, Z. CRISPR/Cas9-Mediated Efficient Targeted Mutagenesis in Chardonnay (Vitis Vinifera L.). Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, I.; Ban, Y.; Azuma, A.; Onoue, N.; Moriguchi, T.; Yamamoto, T.; Toki, S.; Endo, M. CRISPR/Cas9-Mediated Targeted Mutagenesis in Grape. PLoS One 2017, 12, 1–16. [Google Scholar] [CrossRef]
- Wang, X.; Tu, M.; Wang, D.; Liu, J.; Li, Y.; Li, Z.; Wang, Y.; Wang, X. CRISPR/Cas9-Mediated Efficient Targeted Mutagenesis in Grape in the First Generation. Plant Biotechnol. J. 2018, 16, 844–855. [Google Scholar] [CrossRef]
- Ren, F.; Ren, C.; Zhang, Z.; Duan, W.; Lecourieux, D.; Li, S.; Liang, Z. Efficiency Optimization of CRISPR/CAS9-Mediated Targeted Mutagenesis in Grape. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Li, M.Y.; Jiao, Y.T.; Wang, Y.T.; Zhang, N.; Wang, B.B.; Liu, R.Q.; Yin, X.; Xu, Y.; Liu, G.T. CRISPR/Cas9-Mediated VvPR4b Editing Decreases Downy Mildew Resistance in Grapevine (Vitis vinifera L.). Hortic. Res. 2020, 7. [Google Scholar] [CrossRef]
- Sunitha, S.; Rock, C.D. CRISPR/Cas9-Mediated Targeted Mutagenesis of TAS4 and MYBA7 Loci in Grapevine Rootstock 101-14. Transgenic Res. 2020, 29. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.Y.; Guo, Y.; Cheng, Y.; Hu, Y.; Xiao, S.; Wang, Y.; Wen, Y.Q. CRISPR/Cas9-Mediated Mutagenesis of VvMLO3 Results in Enhanced Resistance to Powdery Mildew in Grapevine (Vitis vinifera). Hortic. Res. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Liu, Y.; Guo, Y.; Duan, W.; Fan, P.; Li, S.; Liang, Z. Optimizing the CRISPR/Cas9 System for Genome Editing in Grape by Using Grape Promoters. Hortic. Res. 2021, 8. [Google Scholar] [CrossRef]
- Varkonyi-Gasic, E.; Wang, T.; Voogd, C.; Jeon, S.; Drummond, R.S.M.; Gleave, A.P.; Allan, A.C. Mutagenesis of Kiwifruit CENTRORADIALIS-like Genes Transforms a Climbing Woody Perennial with Long Juvenility and Axillary Flowering into a Compact Plant with Rapid Terminal Flowering. Plant Biotechnol. J. 2019, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, S.; Li, D.; Zhang, Q.; Li, L.; Zhong, C.; Liu, Y.; Huang, H. Optimized Paired-sgRNA/Cas9 Cloning and Expression Cassette Triggers High-efficiency Multiplex Genome Editing in Kiwifruit. Plant Biotechnol. J. 2018, 16, 1424–1433. [Google Scholar] [CrossRef] [PubMed]
- Herath, D.; Voogd, C.; Mayo-Smith, M.; Yang, B.; Allan, A.C.; Putterill, J.; Varkonyi-Gasic, E. CRISPR-Cas9-Mediated Mutagenesis of Kiwifruit BFT Genes Results in an Evergrowing but Not Early Flowering Phenotype. Plant Biotechnol. J. 2022, 20, 2064–2076. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Herrera, A.; Figueroa-Yáñez, L.; Castaño, E.; Santamaría, J.; Pereira-Santana, A.; Espadas-Alcocer, J.; Sánchez-Teyer, F.; Espadas-Gil, F.; Alcaraz, L.D.; López-Gómez, R.; et al. A Novel Dreb2-Type Gene from Carica Papaya Confers Tolerance under Abiotic Stress. Plant Cell. Tissue Organ Cult. 2016, 125. [Google Scholar] [CrossRef]
- Gumtow, R.; Wu, D.; Uchida, J.; Tian, M. A Phytophthora Palmivora Extracellular Cystatin-like Protease Inhibitor Targets Papain to Contribute to Virulence on Papaya. Mol. Plant-Microbe Interact. 2018, 31, 363–373. [Google Scholar] [CrossRef]
- Pettongkhao, S.; Navet, N.; Schornack, S.; Tian, M.; Churngchow, N. A Secreted Protein of 15 KDa Plays an Important Role in Phytophthora Palmivora Development and Pathogenicity. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Pang, H.G.; Yan, Q.; Zhao, S.; He, F.; Xu, J.F.; Qi, B.X.; Zhang, Y. Knockout of the S-Acyltransferase Gene, Pbpat14, Confers the Dwarf Yellowing Phenotype in First Generation Pear by ABA Accumulation. Int. J. Mol. Sci. 2019, 20, 1–16. [Google Scholar] [CrossRef]
- Malnoy, M.; Viola, R.; Jung, M.H.; Koo, O.J.; Kim, S.; Kim, J.S.; Velasco, R.; Kanchiswamy, C.N. DNA-Free Genetically Edited Grapevine and Apple Protoplast Using CRISPR/Cas9 Ribonucleoproteins. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhu, H.; Liu, J.; Yang, Q.; Shao, X.; Bi, F.; Hu, C.; Huo, H.; Chen, K.; Yi, G. Establishment of a PEG-Mediated Protoplast Transformation System Based on DNA and CRISPR/Cas9 Ribonucleoprotein Complexes for Banana. BMC Plant Biol. 2020, 20. [Google Scholar] [CrossRef] [PubMed]
- Pavese, V.; Moglia, A.; Abbà, S.; Milani, A.M.; Marinoni, D.T.; Corredoira, E.; Martinez, M.T.; Botta, R. First Report on Genome Editing via Ribonucleoprotein (RNP) in Castanea Sativa Mill. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Scintilla, S.; Salvagnin, U.; Giacomelli, L.; Zeilmaker, T.; Malnoy, M.A.; Rouppe van der Voort, J.; Moser, C. Regeneration of Non-Chimeric Plants from DNA-Free Edited Grapevine Protoplasts. Front. Plant Sci. 2022, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.; Bertini, E.; D’Incà, E.; Fasoli, M.; Zenoni, S. DNA-Free Genome Editing in Grapevine Using CRISPR/Cas9 Ribonucleoprotein Complexes Followed by Protoplast Regeneration. Hortic. Res. 2023, 10. [Google Scholar] [CrossRef]
- Mahmoud, L.M.; Kaur, P.; Stanton, D.; Grosser, J.W.; Dutt, M. A Cationic Lipid Mediated CRISPR/Cas9 Technique for the Production of Stable Genome Edited Citrus Plants. Plant Methods 2022, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cheng, Y.; Fang, H.; Roberts, N.; Zhang, L.; Vakulskas, C.A.; Niedz, R.P.; Culver, J.N.; Qi, Y. Highly Efficient Genome Editing in Plant Protoplasts by Ribonucleoprotein Delivery of CRISPR-Cas12a Nucleases. Front. Genome Ed. 2022, 4, 1–11. [Google Scholar] [CrossRef]
- Pak, S.; Li, C. Progress and Challenges in Applying CRISPR/Cas Techniques to the Genome Editing of Trees. For. Res. 2022, 2, 0–0. [Google Scholar] [CrossRef]
- Min, T.; Hwarari, D.; Li, D.; Movahedi, A.; Yang, L. CRISPR-Based Genome Editing and Its Applications in Woody Plants. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Donadio, L.C.; Lederman, I.E.; Roberto, S.R.; Stucchi, E.S. Dwarfing-Canopy and Rootstock Cultivars for Fruit Trees. Rev. Bras. Frutic. 2019, 41. [Google Scholar] [CrossRef]
- Dongariyal, A.; Chandra, A.K.; Dongriyal, A.; Kumar, A.; Sharma, P. Tending Genome Editing via CRISPR/Cas9-Induced Mutagenesis: Opportunity and Challenges for Yield, Quality and Nutritional Improvement of Fruit Crops. Sci. Hortic. 2023, 311. [Google Scholar] [CrossRef]
- Rudikovskii, A. V.; Stolbicova, A. V.; Rudikovskaya, E.G.; Dudareva, L. V. Role of Phytohormones in the Formation of Dwarf and Tall Siberian Crabapple (Malus Bаccata l. Borkh.). Zemdirbyste 2019, 106. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, M.; Tan, B.; Jiang, Y.; Zheng, X.; Ye, X.; Guo, Z.; Xiong, T.; Wang, W.; Li, J.; et al. A Single Nucleotide Mutation in GID1c Disrupts Its Interaction with DELLA1 and Causes a GA-Insensitive Dwarf Phenotype in Peach. Plant Biotechnol. J. 2019, 17, 1723–1735. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, X.; Wu, T.; Xu, X.; Han, Z.; Wang, Y. Methylation Effect on IPT5b Gene Expression Determines Cytokinin Biosynthesis in Apple Rootstock. Biochem. Biophys. Res. Commun. 2017, 482. [Google Scholar] [CrossRef]
- Sattar, M.N.; Iqbal, Z.; Al-Khayri, J.M.; Jain, S.M. Induced Genetic Variations in Fruit Trees Using New Breeding Tools: Food Security and Climate Resilience. Plants 2021, 10, 1347. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Gong, X.; Li, M.; Li, C.; Sun, T.; Ma, F. Overexpression of a Novel Apple NAC Transcription Factor Gene, MdNAC1, Confers the Dwarf Phenotype in Transgenic Apple (Malus domestica). Genes 2018, 9. [Google Scholar] [CrossRef]
- Jia, P.; Xing, L.; Zhang, C.; Chen, H.; Li, Y.; Zhang, D.; Ma, J.; Zhao, C.; Han, M.; Ren, X.; et al. MdKNOX15, a Class I Knotted-like Transcription Factor of Apple, Controls Flowering and Plant Height by Regulating GA Levels through Promoting the MdGA2ox7 Transcription. Environ. Exp. Bot. 2021, 185. [Google Scholar] [CrossRef]
- Ren, C.; Guo, Y.; Kong, J.; Lecourieux, F.; Dai, Z.; Li, S.; Liang, Z. Knockout of VvCCD8 Gene in Grapevine Affects Shoot Branching. BMC Plant Biol. 2020, 20. [Google Scholar] [CrossRef]
- Clough, S.J.; Fengler, K.A.; Yu, I.C.; Lippok, B.; Smith, R.K.; Bent, A.F. The Arabidopsis Dnd1 “Defense, No Death” Gene Encodes a Mutated Cyclic Nucleotide-Gated Ion Channel. Proc. Natl. Acad. Sci. U. S. A. 2000, 97. [Google Scholar] [CrossRef]
- Ma, W.; Smigel, A.; Walker, R.K.; Moeder, W.; Yoshioka, K.; Berkowitz, G.A. Leaf Senescence Signaling: The Ca2+-Conducting Arabidopsis Cyclic Nucleotide Gated Channel2 Acts through Nitric Oxide to Repress Senescence Programming. Plant Physiol. 2010, 154. [Google Scholar] [CrossRef] [PubMed]
- Rehman, R.U. Cyclic Nucleotide-Gated Channels: Essential Signaling Components in Plants for Fertilization and Immunity Responses. In Plant Signaling: Understanding the Molecular Crosstalk; 2014; Vol. 9788132215.
- Corte, L.E.D.; Mahmoud, L.M.; Moraes, T.S.; Mou, Z.; Grosser, J.W.; Dutt, M. Development of Improved Fruit, Vegetable, and Ornamental Crops Using the CRISPR/Cas9 Genome Editing Technique. Plants 2019, 8. [Google Scholar]
- Pillitteri, L.J.; Lovatt, C.J.; Walling, L.L. Isolation and Characterization of a Terminal Flower Homolog and Its Correlation with Juvenility in Citrus. Plant Physiol. 2004, 135. [Google Scholar] [CrossRef] [PubMed]
- Eshed, Y.; Lippman, Z.B. Revolutions in Agriculture Chart a Course for Targeted Breeding of Old and New Crops. Science (80-. ). 2019; 366. [Google Scholar]
- Endo, T.; Shimada, T.; Fujii, H.; Kobayashi, Y.; Araki, T.; Omura, M. Ectopic Expression of an FT Homolog from Citrus Confers an Early Flowering Phenotype on Trifoliate Orange (Poncirus trifoliata L. Raf.). Transgenic Res. 2005, 14. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, K.; Agüero, J.; Vives, M.C.; Aleza, P.; Pina, J.A.; Moreno, P.; Navarro, L.; Guerri, J. Precocious Flowering of Juvenile Citrus Induced by a Viral Vector Based on Citrus Leaf Blotch Virus: A New Tool for Genetics and Breeding. Plant Biotechnol. J. 2016, 14. [Google Scholar] [CrossRef] [PubMed]
- Callahan, A.M.; Srinivasan, C.; Dardick, C.; Scorza, R. Rapid Cycle Breeding: Application of Transgenic Early Flowering for Perennial Trees. In Plant Breeding Reviews; 2016; Vol. 40.
- Prudencio, A.S.; Devin, S.R.; Mahdavi, S.M.E.; Martínez-García, P.J.; Salazar, J.A.; Martínez-Gómez, P. Spontaneous, Artificial, and Genome Editing-Mediated Mutations in Prunus. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Sapkota, M.; van der Knaap, E. Perspectives of CRISPR/Cas-Mediated Cis-Engineering in Horticulture: Unlocking the Neglected Potential for Crop Improvement. Hortic. Res. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D.; et al. Major Impacts of Widespread Structural Variation on Gene Expression and Crop Improvement in Tomato. Cell 2020, 182. [Google Scholar] [CrossRef]
- Li, Q.; Feng, Q.; Snouffer, A.; Zhang, B.; Rodríguez, G.R.; van der Knaap, E. Increasing Fruit Weight by Editing a Cis-Regulatory Element in Tomato KLUH Promoter Using CRISPR/Cas9. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Salazar, J.A.; Ruiz, D.; Zapata, P.; Martínez-García, P.J.; Martínez-Gómez, P. Whole Transcriptome Analyses of Apricots and Japanese Plum Fruits after 1-MCP (Ethylene-Inhibitor) and Ethrel (Ethylene-Precursor) Treatments Reveal New Insights into the Physiology of the Ripening Process. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef]
- Hu, H.; Scheben, A.; Verpaalen, B.; Tirnaz, S.; Bayer, P.E.; Hodel, R.G.J.; Batley, J.; Soltis, D.E.; Soltis, P.S.; Edwards, D. Amborella Gene Presence/Absence Variation Is Associated with Abiotic Stress Responses That May Contribute to Environmental Adaptation. New Phytol. 2022, 233. [Google Scholar] [CrossRef]
- Fizikova, A.; Tikhonova, N.; Ukhatova, Y.; Ivanov, R.; Khlestkina, E. Applications of CRISPR/Cas9 System in Vegetatively Propagated Fruit and Berry Crops. Agronomy 2021, 11. [Google Scholar] [CrossRef]
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive Compounds and Antioxidant Activity in Different Types of Berries. Int. J. Mol. Sci. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Dall’Osto, L.; Holt, N.E.; Kaligotla, S.; Fuciman, M.; Cazzaniga, S.; Carbonera, D.; Frank, H.A.; Alric, J.; Bassi, R. Zeaxanthin Protects Plant Photosynthesis by Modulating Chlorophyll Triplet Yield in Specific Light-Harvesting Antenna Subunits. J. Biol. Chem. 2012, 287. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Wu, S.; Tian, L. Effective Genome Editing and Identification of a Regiospecific Gallic Acid 4-O-Glycosyltransferase in Pomegranate (Punica granatum L.). Hortic. Res. 2019, 6. [Google Scholar] [CrossRef]
- Goulet, C.; Kamiyoshihara, Y.; Lam, N.B.; Richard, T.; Taylor, M.G.; Tieman, D.M.; Klee, H.J. Divergence in the Enzymatic Activities of a Tomato and Solanum Pennellii Alcohol Acyltransferase Impacts Fruit Volatile Ester Composition. Mol. Plant 2015, 8. [Google Scholar] [CrossRef]
- Cao, X.; Wei, C.; Duan, W.; Gao, Y.; Kuang, J.; Liu, M.; Chen, K.; Klee, H.; Zhang, B. Transcriptional and Epigenetic Analysis Reveals That NAC Transcription Factors Regulate Fruit Flavor Ester Biosynthesis. Plant J. 2021, 106. [Google Scholar] [CrossRef]
- Zaidi, S.M.S.; Saud, A. Future of US-China Relations: Conflict, Competition or Cooperation? Asian Soc. Sci. 2020, 16. [Google Scholar] [CrossRef]
- Joshi, R.K.; Bharat, S.S.; Mishra, R. Engineering Drought Tolerance in Plants through CRISPR/Cas Genome Editing. 3 Biotech 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Zhao, X.; Zhao, Q.; Wang, G.; Li, Y.; Hao, Y. MdDREB2A in Apple Is Involved in the Regulation of Multiple Abiotic Stress Responses. Hortic. Plant J. 2021, 7. [Google Scholar] [CrossRef]
- Liao, X.; Guo, X.; Wang, Q.; Wang, Y.; Zhao, D.; Yao, L.; Wang, S.; Liu, G.; Li, T. Overexpression of MsDREB6.2 Results in Cytokinin-Deficient Developmental Phenotypes and Enhances Drought Tolerance in Transgenic Apple Plants. Plant J. 2017, 89, 510–526. [Google Scholar] [CrossRef]
- Jin, W.; Dong, J.; Hu, Y.; Lin, Z.; Xu, E.; Han, Z. Improved Cold-Resistant Performance in Transgenic Grape (Vitis Vinifera L.) Overexpressing Cold-Inducible Transcription Factors AtDREB1b. HortScience 2009, 44. [Google Scholar] [CrossRef]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Leonhardt, N.; Dellaporta, S.L.; Tonelli, C. A Guard-Cell-Specific MYB Transcription Factor Regulates Stomatal Movements and Plant Drought Tolerance. Curr. Biol. 2005, 15. [Google Scholar] [CrossRef]
- Wen, X.P.; Pang, X.M.; Matsuda, N.; Kita, M.; Inoue, H.; Hao, Y.J.; Honda, C.; Moriguchi, T. Over-Expression of the Apple Spermidine Synthase Gene in Pear Confers Multiple Abiotic Stress Tolerance by Altering Polyamine Titers. Transgenic Res. 2008, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, T.; Li, T.; Wang, M.; Yang, G.; He, G. A CBL-Interacting Protein Kinase TaCIPK2 Confers Drought Tolerance in Transgenic Tobacco Plants through Regulating the Stomatal Movement. PLoS One 2016, 11. [Google Scholar] [CrossRef]
- Kaur, N.; Awasthi, P.; Tiwari, S. Fruit Crops Improvement Using CRISPR/Cas9 System; Elsevier Inc., 2020; ISBN 9780128181409.
- Bhat, J.A.; Ali, S.; Salgotra, R.K.; Mir, Z.A.; Dutta, S.; Jadon, V.; Tyagi, A.; Mushtaq, M.; Jain, N.; Singh, P.K.; et al. Genomic Selection in the Era of Next Generation Sequencing for Complex Traits in Plant Breeding. Front. Genet. 2016, 7, 221. [Google Scholar] [CrossRef] [PubMed]
- Devin, S.R.; Prudencio, Á.S.; Mahdavi, S.M.E.; Rubio, M.; Martínez-García, P.J.; Martínez-Gómez, P. Orchard Management and Incorporation of Biochemical and Molecular Strategies for Improving Drought Tolerance in Fruit Tree Crops. Plants 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Li, X.; Dong, X. Generation of Broad-Spectrum Disease Resistance by Everexpression of an Essential Regulatory Gene in Systemic Acquired Resistance. Proc. Natl. Acad. Sci. U. S. A. 1998, 95. [Google Scholar] [CrossRef]
- Le Henanff, G.; Farine, S.; Kieffer-Mazet, F.; Miclot, A.S.; Heitz, T.; Mestre, P.; Bertsch, C.; Chong, J. Vitis Vinifera VvNPR1.1 Is the Functional Ortholog of AtNPR1 and Its Overexpression in Grapevine Triggers Constitutive Activation of PR Genes and Enhanced Resistance to Powdery Mildew. Planta 2011, 234. [Google Scholar] [CrossRef]
- Malnoy, M.; Jin, Q.; Borejsza-Wysocka, E.E.; He, S.Y.; Aldwinckle, H.S. Overexpression of the Apple MpNPR1 Gene Confers Increased Disease Resistance in Malus x domestica. Mol. Plant-Microbe Interact. 2007, 20. [Google Scholar] [CrossRef]
- Kusch, S.; Panstruga, R. Mlo-Based Resistance: An Apparently Universal “Weapon” to Defeat Powdery Mildew Disease. Mol. Plant-Microbe Interact. 2017, 30.
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of Broad Virus Resistance in Non-Transgenic Cucumber Using CRISPR/Cas9 Technology. Mol. Plant Pathol. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Gao, C. Genome Engineering for Crop Improvement and Future Agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef]
- Slaymaker, I.M.; Gao, L.; Zetsche, B.; Scott, D.A.; Yan, W.X.; Zhang, F. Rationally Engineered Cas9 Nucleases with Improved Specificity. Science (80-. ). 2016, 351. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Sander, J.D.; Reyon, D.; Cascio, V.M.; Joung, J.K. Improving CRISPR-Cas Nuclease Specificity Using Truncated Guide RNAs. Nat. Biotechnol. 2014, 32. [Google Scholar] [CrossRef]
- Osakabe, Y.; Watanabe, T.; Sugano, S.S.; Ueta, R.; Ishihara, R.; Shinozaki, K.; Osakabe, K. Optimization of CRISPR/Cas9 Genome Editing to Modify Abiotic Stress Responses in Plants. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Steinert, J.; Schiml, S.; Fauser, F.; Puchta, H. Highly Efficient Heritable Plant Genome Engineering Using Cas9 Orthologues from Streptococcus Thermophilus and Staphylococcus Aureus. Plant J. 2015, 84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhang, H.; Li, T.; Chen, K.; Qiu, J.L.; Gao, C. Perfectly Matched 20-Nucleotide Guide RNA Sequences Enable Robust Genome Editing Using High-Fidelity SpCas9 Nucleases. Genome Biol. 2017, 18. [Google Scholar] [CrossRef]
- Mikami, M.; Toki, S.; Endo, M. Precision Targeted Mutagenesis via Cas9 Paired Nickases in Rice. Plant Cell Physiol. 2016, 57. [Google Scholar] [CrossRef]
- Spök, A.; Sprink, T.; Allan, A.C.; Yamaguchi, T.; Dayé, C. Towards Social Acceptability of Genome-Edited Plants in Industrialised Countries? Emerging Evidence from Europe, United States, Canada, Australia, New Zealand, and Japan. Front. Genome Ed. 2022, 4, 2683–2691. [Google Scholar] [CrossRef] [PubMed]
- Sprink, T.; Wilhelm, R.; Hartung, F. Genome Editing around the Globe: An Update on Policies and Perceptions. Plant Physiol. 2022, 190, 1579–1587. [Google Scholar] [CrossRef]
- Eriksson, D.; Kershen, D.; Nepomuceno, A.; Pogson, B.J.; Prieto, H.; Purnhagen, K.; Smyth, S.; Wesseler, J.; Whelan, A. A Comparison of the EU Regulatory Approach to Directed Mutagenesis with That of Other Jurisdictions, Consequences for International Trade and Potential Steps Forward. New Phytol. 2019, 222, 1673–1684. [Google Scholar] [CrossRef]
- Van Der Meer, P.; Angenon, G.; Bergmans, H.; Buhk, H.J.; Callebaut, S.; Chamon, M.; Eriksson, D.; Gheysen, G.; Harwood, W.; Hundleby, P.; et al. The Status under EU Law of Organisms Developed through Novel Genomic Techniques. Eur. J. Risk Regul. 2023, 14, 93–112. [Google Scholar] [CrossRef]
- Hu, J.; Gao, C. CRISPR-Edited Plants by Grafting. Nat. Biotechnol. 2023, 41, 909–910. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Chen, K.; Li, T.; Zhang, Y.; Wang, Y.; Zhao, Q.; Liu, J.; Zhang, H.; Liu, C.; Ran, Y.; et al. Efficient DNA-Free Genome Editing of Bread Wheat Using CRISPR/Cas9 Ribonucleoprotein Complexes. Nat. Commun. 2017, 8, 6–10. [Google Scholar] [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Mark Cigan, A. Genome Editing in Maize Directed by CRISPR-Cas9 Ribonucleoprotein Complexes. Nat. Commun. 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.W.; Kim, J.; Kwon, S. Il; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-Free Genome Editing in Plants with Preassembled CRISPR-Cas9 Ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Turesson, H.; Nicolia, A.; Fält, A.S.; Samuelsson, M.; Hofvander, P. Efficient Targeted Multiallelic Mutagenesis in Tetraploid Potato (Solanum tuberosum) by Transient CRISPR-Cas9 Expression in Protoplasts. Plant Cell Rep. 2017, 36, 117–128. [Google Scholar] [CrossRef]
Figure 1.
Gene editing mechanism of CRISP/Cas9. A single RNA chimera (sgRNA) drives the complex CRISPR/Cas9 to the target DNA and the protospacer adjacent motif (PAM) enables the stable binding. Cas9 produces a double strand break (DSB) that can trigger two endogenous DNA repair mechanisms: homologous directed repair (HDR) or nonhomologous end joining (NHEJ).
Figure 1.
Gene editing mechanism of CRISP/Cas9. A single RNA chimera (sgRNA) drives the complex CRISPR/Cas9 to the target DNA and the protospacer adjacent motif (PAM) enables the stable binding. Cas9 produces a double strand break (DSB) that can trigger two endogenous DNA repair mechanisms: homologous directed repair (HDR) or nonhomologous end joining (NHEJ).
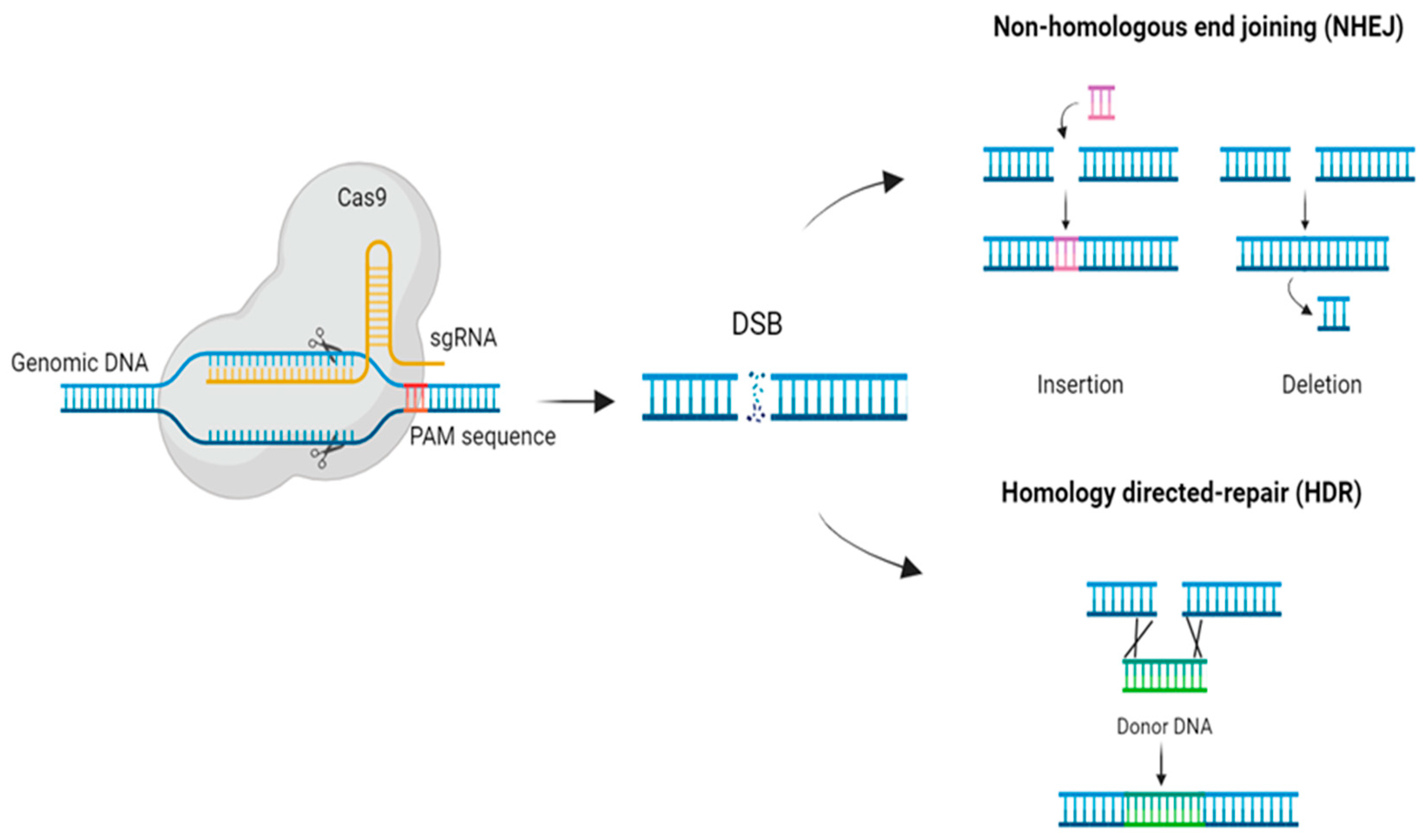
Figure 2.
Overview of the applications and challenges of the CRISPR-Cas9 gene editing technique in fruit trees.
Figure 2.
Overview of the applications and challenges of the CRISPR-Cas9 gene editing technique in fruit trees.

Table 1.
Collection of traits improved by CRISPR/Cas9 and CRISPR/Cas12 in different fruit trees.
| CRISPR-Cas delivery | Fruit trees | Target gene edited | Improved Traits | References |
|---|---|---|---|---|
| A. tumefaciens-mediated | Apple | PDS | Enhanced biosynthesis of carotenoid | [80] |
| PDS and TFL1 | Albino phenotype and early flowering | [81] | ||
| MdDIPM1 and MdDIPM4 | Fire blight resistance | [82] | ||
| ALS | Chlorsulfuron resistance | [83] | ||
| CNGC2 | B. dothidea resistance | [84] | ||
| MsPDS | Albino phenotype | [85] | ||
| MdMKK9 | Increased anthocyanin content | [86] | ||
| Detection viruses and viroids CRISPR | [87] | |||
| Banana | RAS-PDS1 and 2 | Albino phenotype | [88] | |
| PDS | Albino phenotype and dwarfing | [89,90] | ||
| eBSV | Control of virus pathogenesis | [91] | ||
| MaGA20ox2 | Dwarf phenotype | [92]) | ||
| LCYε | Carotene biosynthesis | [93] | ||
| MaACO1 | Fruit ripening | [94] | ||
| Cacao | TcNPR3 | Phytophthora tropicalis resistance | [95] | |
| Citrus | CsPDS | Method optimization | [96] | |
| Cs7g03360 | Phenotypic changes in Carrizo leaf | [97] | ||
| CsLOB1 | Citrus canker resistance | [98] | ||
| CsLOB1 promoter | Citrus canker resistance | [99] | ||
| PDS and CsLOB1 | Albino phenotype and canker resistance | [100] | ||
| CsWRKY22 | Citrus canker resistance | [101] | ||
| CsLOB1 | Citrus canker resistance | [102] | ||
| pC-PDS1 and Pc-PDS2 | Chlorophyll and carotenoid content | [103] | ||
| CsLOB1 | Citrus canker resistance | [104] |
Table 1.
Collection of traits improved by CRISPR/Cas9 and CRISPR/Cas12 in different fruit trees (Cont.).
Table 1.
Collection of traits improved by CRISPR/Cas9 and CRISPR/Cas12 in different fruit trees (Cont.).
| CRISPR-Cas delivery | Fruit trees | Target gene edited | Improved Traits | References |
|---|---|---|---|---|
| Grape | IdnDH | High tartaric acid biosynthesis | [105] | |
| VvPDS | Increased carotenoid biosynthesis | [106] | ||
| VvWRKY52 | Botrytis cinerea resistance | [107] | ||
| VvPDS | Albino phenotype | [108] | ||
| VvMLO7 | Powdery mildew resistance | [82] | ||
| VvPR4b | Downy mildew resistance | [109] | ||
| MYBA5/6/7 and TAS4a/b | Anthocyanin accumulation | [110] | ||
| VvMLO3 and VvMLO4 | Powdery mildew resistance | [111] | ||
| TMT1 and TMT2 | Reduced sugar accumulation | [112] | ||
| Kiwifruit | CEN4 and CEN | Terminal flower and fruit | [113] | |
| AcPDS | Albino phenotype (leaves) | [114] | ||
| AcBFT2 | Reduced dormancy and early bud break | [115] | ||
| Papaya | CpDreb2 | Gene disruption for water stress | [116] | |
| PpalEPIC8 | Phytophthora palmivora resistance | [117] | ||
| Ppal15kDa | Phytophthora palmivora resistance | [118] | ||
| Pear | TFL1 | Early flowering | [81] | |
| PbPAT14 | Dwarf yellowing phenotype | [119] | ||
| PDS and ALS | Albino phenotype and chlorsulfuron resistance | [83] | ||
| PEG-meditated | Apple | DIPM-1, -2 and -4 | Fire blight resistance | [120] |
| Banana | PDS | Method optimization | [121] | |
| Chestnut | PDS | Method optimization | [122] | |
| Grape | MLO7 | Enhanced biotic resistance against powdery mildew | [120] | |
| DMR6 and MLO6 | Downy- and powdery-mildew resistance | [123] | ||
| GFP | Method optimization | [124] | ||
| Orange | CsNPR3 | Induced biotic stress tolerance | [125] | |
| PH5 | Method optimization | [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



