
Submitted:
02 November 2023
Posted:
03 November 2023
You are already at the latest version
Abstract
Alzheimer’s disease manifests itself as a complex pathological condition with neuroinflammation, oxidative stress and cholinergic dysfunction being a few of the many pathological changes. Due to the complexity of the disease current therapeutic strategies aim at a multitargeted approach often relying on a combination of sub-stances with versatile and complementary effects. In the present study, unique combination of α-lipoic acid, citicoline, extracts of leaves from olive tree and green tea, Vitamin D3, selenium and an immune supporting complex was tested in a scopolamine-induced dementia in rats. Using behavioral and biochemical methods we assessed the effects of this combination on learning and memory, and elucidated the mechanisms under-lying its effects. Our results showed that as compared to the components, experimental combination was most efficient in improving short- and long-term memory assessed by the step-through method as well as spatial memory, assessed by T-maze and Barnes maze underlying decrease in AChE activity and LPO, in-creases in SOD activity in cortex; activities of catalase and GPx, levels of BDNF and pCREB in the hippocam-pus. No significant histopathological changes or blood parameter changes were detected, making the experi-mental combination an effective and safe candidate in a multitargeted treatment of AD.
Keywords:
Alzheimer’s disease
; α-lipoic acid
; citicoline
; extract of leaves green tea
; extract of leaves olive tree
; vitamin D3
; selenium
; scopolamine
1. Introduction
The Alzheimer’s Disease (AD), the most common form of dementia (contributing to 60-70% of cases), is often referred to as the “epidemic of the 21st century”. According to the World Health Organization, at present, more than 55 million people suffer from dementia, the number increasing with 10 million new cases each year. AD and other forms of dementia are revealed as leading causes for death and disability, being classified among the top 10 causes of death worldwide and ranking 3rd in both the Americas and Europe in 2019 and described as one of the most important sources of incapacity and dependency among older people globally (turning the neurodegenerative condition into a major healthcare challenge).
Although deposition of extracellular senile plaques and the presence of intracellular neurofibrillary tangles are considered the major characteristics of AD, it manifests as a complex pathological condition with neuroinflammation, oxidative stress, mitochondrial dysfunction and changes in the cholinergic and monoaminergic neurotransmitter systems being of significant importance [1,2,3,4,5,6,7,8,9]. Apoptosis, altered brain connectivity and abberant neuronal activity as well as brain imbalance of metal ions such as iron, copper, zink and calcium and impaired glucose metabolism are also shown to play an important role [10,11,12,13]. Thus, modern AD therapeutic strategies are increasingly aiming at applying a multi-targeted approach to account for the large complexity of the neurodegenerative condition, whose treatment and prevention so far remain ineffective. Due to their many-sided mechanisms of action and versatile beneficial effects for a range of diseases, biologically active substances of natural origin are coming steadily in the focus of attention of researchers and clinicians as possible treatment and preventive drugs. Many research groups, including ours, report on the favorable effects of natural substances and their synthetically modified analogues in experimental models of dementia and AD based on their antioxidative, anticholinesterase, anti-inflammatory, immunomodulatory and other properties [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30].
Thus, relying on the combined effects and synergistic action of their ingredients components pharmacological combinations based on naturally occurring substances can demonstrate improved effectiveness and turn promising tools as treatment and preventive AD drugs [31,32].
Lipoic acid (LA), also known as α-lipoic acid and thioctic acid, is a low molecular dithiol, naturally occurring in cells and functioning as an essential cofactor for mitochondrial enzymes -ketoacid dehydrogenases) [33]. It is found in both vegetable and animal-based food, most abundantly in spinach, broccolis, and tomatoes as well as bovine kidney, heart, and liver [34]. It affects multiple pathways and shows an abundance of effects that can improve cognitive performance, ensure and sustain neuroprotection and be considered in neurodegenerative disorders, especially AD. Lipoic acid is a powerful antioxidant as, additional to reducing oxidative stress by scavenging reactive oxygen species (ROS), hypochlorous acid and NO and preventing oxidation of lipids and proteins, it regenerates other oxidants like Vit C and E and increases the intracellular levels of glutathione and ubiquinone [35,36,37]. As generally known GSH is an essential key antioxidant with fundamental role in neutralizing ROS and regulating the intrinsic redox homeostasis.
Furthermore, LA and its reduced form dihydrolipoic acid (DHLA) chelate redox-active metals such as Co, Fe and Zn, whose accumulation mediates free radical generation and oxidative damage related to neurodegeneration [38,39]. There is evidence that the iron-chelating ability decreases the phosphorylation of tau and the aggregation of Aß monomers into harmful fibrous forms, thus delaying cell death [40]. Additionally, LA is effective against Aβ-induced cytotoxicity most probably due to its free radical scavenging capacity as Aβ in senile plaques is demonstrated to stimulate production of free radicals [41].
LA possesses also anti-inflammatory properties once, by being a strong antioxidant, as elevated oxidative stress is related to chronic inflammation and second, by inhibiting the NF-κB signaling pathway which involves various genes expression implicated in cell apoptosis and inflammatory responses [42]. LA increases acetylcholine (ACh) production by activation of choline acetyltransferase and by increasing glucose uptake, thus supplying more acetyl-CoA for the production of ACh [43]. LA demonstrates antiapoptotic properties by inhibiting both caspase-dependent and caspase independent cell death pathways [44]. And last but not least treatment with LA has been shown to increase the levels of neurotransmitters like dopamine, serotonin, and norepinephrine in various brain regions. [45].
In view of the numerous favourable properties of LA a wealth of evidence shows its ameliorating effects in cell and animal models of AD as well as when used by patients [46,47,48,49,50,51]. Several studies examine the outcome also from the use of combinations of LA with other natural substances like curcumin, piperine, epigallocatechin gallate, N-acetylcysteine, B vitamins, vitamin C and omega-3 fatty reporting positive effects on cognition and AD pathology both in an AD mouse model and AD patients [31,32,52]. Thus, LA poses itself as a serious candidate for taking a central part in a possible multitargeted AD therapy.
Citicoline (cytidine 5’-diphosphocholine), normally present in human cells, is an essential intermediate in the biosynthesis of cell membrane structural phospholipids, particularly, phosphatidylcholine and a source of choline, used in the biosynthesis of acetylcholine [53]. Citicoline appears not less significant than LA as a possible AD treatment and preventive drug as it demonstrates an equally impressive array of physiological functions related to neuroprotective and procognitive effects. Apart from being involved in the biosynthesis of phosphatidylcholine, a primary component of neuronal membranes and ACh with a major role in memory and learning, it advances the synthesis of several other membrane phospholipids, including phosphatidylethanolamine and phosphatidylserine, contributing to neuronal repair and regeneration and acts as a selective agonist at nicotinic Ach receptors (choline at α7-nAChR ), which have an important role in maintaining normal cognition. In cerebral ischemia and models of neurodegeneration citicoline has been shown to inhibit apoptosis [53,54], to reduce hippocampal neuronal degeneration, to ameliorate learning and memory [54], to normalize norepinephrine and dopamine release [55], to inhibit phospholipase A2 (PLA2) thus preventing ROS and proinflammatory mediators generation from fatty acid, especially arachidonic acid [56]. Oxidative metabolism of arachidonic acid is considered a significant source of ROS [57]. Arachidonic acid is the direct precursor of bioactive lipid metabolites of eicosanoids such as prostaglandins, leukotrienes and epoxyeicosatrienoic acid, considered proinflammatory molecules, as they trigger oxidative stress and stimulate the immune response [58]. Moreover, citicoline stimulates glutathione synthesis and the activity of glutathione reductase, attenuates lipid peroxidation, facilitates mitochondrial function by increasing the activity of cytochrome oxidase [53] and restoring the activity of the mitochondrial ATPase [59,60] and levels of cardiolipin, a phospholipid indispensable for mitochondrial electron transport [53]. Actually, citicoline has been long recognized for its procognitive properties and is one of the most commonly prescribed drugs for cognitive impairments in several European countries [61]. Several recent reviews on human studies and clinical trials on the use of citicoline strongly support a role for citicoline as coadjuvant treatment of cognitive impairment in dementia, chronic degenerative diseases (including AD), stroke and cerebral ischemia and traumatic head injury [52,62,63,64]. Citicoline seems to be effective in improving cognitive abilities in normal healthy people as well [65].
Selenium (Se) is an important trace element involved in a number of important physiological functions [66,67,68] It has been demonstrated that a decrease in (the plasma concentrations of) selenium is related to cognitive decline (and that the cognitive decline is positively correlated with the Se decrease) [69] and several reports show reduced levels of Se in the plasma of AD [70,71,72]. Hence Se levels appear important for proper cognition and Se supplementation appears to hold potential for AD prevention and therapy.The relevance of Se to AD is not without a ground and relies on its strong antioxidant properties not in the last place stemming from the element being indispensable in enzymes such as glutathion peroxidases (GPx1-4, GPx6), thioredoxine reductases (TrxR1-3), selenoprotein P (SelP) and other selenoproteins, involved in reducing oxidative damage to cells and contributing to their oxidative stress defence [73]. Additionally, many studies reveal that Se can alleviate AD-associated pathology by interfering with both the formation of amyloid-β (Aβ) peptide plaques and neurofibrillary tangles, by affecting AD-disordered neurotransmitter metabolism and influencing AD-related signal transduction [74,75,76,77,78].
Olive leaves extract (OLE) has robust anti-oxidant and anti-inflammatory properties due to a remarkable content of bioactive phenols such as phenolic acids, phenolic alcohols, secoiridoid derivatives, flavanols, flavones, flavonols and others, amongst which oleuropein and hydroxytyrosil, in which OLE is particularly abundant and with proven favourable effects regarding AD [79]. Indeed, plentiful research shows that OLE and the bioactive phenols in it are capable of preventing and reducing deteriorating changes related to AD by decreasing the total Aβ brain levels due to increased clearance and reduced production of Aβ, by diminishing neurofibrillary tangles formation, by inhibiting choline esterases, by decreasing oxidative stress and reducing neuroinflammation by inhibiting the NF-κB pathway and upregulating SIRT1, by enhancing autophagy and restoring loss of proteostasis, reflected in lower toxic protein aggregation [80,81,82].
Green tea with a multitude of bioactive compounds such as catechins, caffeine and theanine and particularly epigallocatechin gallate (EGCG), the main cathechin in green tea, has also been targeted as a promising (suitable) option for AD modulation. Catechins, and especially EGCG, have been proven to act as powerful antioxidant systems functioning in several different (additive) ways. First, they directly neutralize ROS, reactive nitrogen species and other free radicals with the antioxidant capacity of EGCG being even greater than that of vitamin E and C, thanks to a number of free OH groups available to sequester radicals. Second, they show indirect antioxidant effects by increasing the activity of SOD and catalase and by binding to antioxidant regulatory elements, inducing the expression of stress response genes relieving oxidative stress. Additional to its potent antioxidant effects EGCG reveals an overabundance of other synergistic actions that shape its neuroprotective role, including reducing tau hyperphosphorylation and its aggregation; preventing the formation of Aβ and its subsequent accumulation by facilitating the nonamyloidogenic processing of the amyloid precursor protein (APP); chelating copper and iron preventing iron-mediated promotion of amyloid and tau pathology; modulating multiple intracellular signalling pathways related to neuroprotection and cell survival and associated with AD such as the protein kinase C (PKC), mitogenactivated protein kinases (MAPK) and phosphoinositide 3-kinase (PI3K/Akt) pathways [83].
Vitamin D is largely recognized for its effect on calcium and bone metabolism, but it has neuroprotective, antioxidant and anti-inflammatory properties as well. Some of the mechanisms by which calcitriol supports the normal brain function and exerts neuroprotective effects include upregulation of the synthesis of neurotrophins such as nerve growth factor (NGF), neurotrophin 3 (NT3 ) and glial cell line-derived neurotrophic factor (GDNF); modulation of neuronal Ca2+ homeostasis by downregulation of L-type voltage-sensitive Ca2+ channels and induction of the synthesis of Ca2+-binding proteins, such as parvalbumin and calbindin-D28K, protecting neurons from calcium excitotoxicity and neuronal death [84,85]; reduction of glutamate neurotoxicity [86,87]. Furthermore, calcitriol has been reported to inhibit the synthesis of inducible nitric oxide synthase (iNOS), thus preventing excessive NO production and generation of toxic hydroxyl and nitrogen dioxide radicals; to increase GSH levels and to upregulate γ-glutamyl transpeptidase activity with a crucial role in maintaining GSH homeostasis [88]; to decrease the secretion of many inflammatory cytokines including IL-2, IL-6, IFN-γ and others and these are just a few of the calcitriols many antioxidant and anti-inflammatory properties. In relation to AD, numerous research using cellular and rodent models has shown that Vitamin D relieves AD pathology by reducing Aβ accumulation through decreasing Aβ formation and increasing its degradation and by inhibiting tau phosphorylation accompanied by prolific cellular changes reducing neuronal loss, oxidative stress and neuroinflammation [89,90]. In human vitamin D levels have been found to positively correlate with memory and cognition and Vit D deficiency has been demonstrated and postulated to be a risk factor for dementia and AD [91,92,93,94,95,96]. Several recent meta-analysis reports provided additional comprehensive evidence for low Vitamin D levels presenting an increased risk for developing both dementia and AD and secured a place for the vitamin in AD preventive and adjuvant therapy especially in vitamin D deficient patients [97,98,99].
In view of the described in detail numerous versatile beneficial effects of the above substances (confirmed for LA in studies of ours as well (Tancheva et al, 2022; Staykov et al, 2022)) underlied by complementary mechanisms of action, the aim of the present study was to study the effect of an experimental combination consisting of α-lipoic acid, citicoline, extract of leaves of green tea, extract of leaves of olive tree, Vitamin D3, сelenium, and an immune supporting complex in an experimental model of Alzheimers-type dementia in rats as a possible part of AD supporting therapy.
2. Materials and Methods
2.1. Experimental combination
The original experimental combination was developed by our team and was patented (patent document № 4391 U1 from 27/10/2022). It contains seven natural bioactive components, namely: α-lipoic acid (Productos Quimicos Gonmisol S.A., Spain), citicoline (as CognizinR, a trade mark by KYOWA HAKKO BIO CO., LTD, JAPAN), extract of leaves from green tea Camellia sinensis, L (PLANTEX, France), extract of leaves from olive tree Olea Europea, L (PLANTEX, France), vitamin D3 (ProTec Nutra Ltd, UK), selenium (as L-selenomethionin - Lallemand, Canada/Europe) and an immune supporting complex, Bulgarian patent (Bul Bio-National Center for Infectious and Parasitic Disesases). The overall technology for preparing the combination is ensured by NATSTIM (www.natstim.eu).
2.2. Laboratory animals
The experiments were conducted on male adult Wistar rats (160-180 g), bred in Experimental Breeding Facility, Slivnitsa, Sofia. The animals were further transported and habituated to the conditions of the local vivarium at Institute of Neurobiology, Bulgarian Academy of Sciences, in plastic cages (n=5) and under standard laboratory conditions- 12-h light/dark cycle, unrestricted access to water and food, optimal temperature, humidity and ventilation of the rooms.
All experimental procedures were conducted in conformity with the institutional guidelines (Bioethics Committee at the Institute of Neurobiology, Bulgarian Academy of Sciences, Bulgaria) in compliance with the national and international laws and policies (the new Directive 2010/63/EU 22.09.2010) of the European Parliament and the Council of the European Union replacing the older Council Directive (86/609/EEC) on the protection of animals used for scientific purposes; the National Institute of Health (NIH) Guide for the Care and Use of Laboratory animals, NIH Publication No. 85-23, 1985 and Bulgarian laws (Ordinance No. 20 of 01.11.2012 on the minimum requirements for the protection and welfare of experimental animals and the facilities for their use, breeding and/or supply, issued by the Ministry of Food and Agriculture, Official Gazette No. 87 of 09.11.2012 and in force from 01.01.2013.
2.3. Experimental groups
Rats were divided into the following experimental groups with 10 animals each:
i) Control group (control) - receiving tap water daily orally (0.5 ml/100 g) for 51 days and at the end of the treatment, together with saline (0.5 ml/100 g, i.p.) for 11 days;
ii) Scopolamine group (Sco) - receiving tap water daily orally (0.5 ml/100 g) for 51 days and at the end of the treatment, together with 2 mg/kg scopolamine (0.5 ml/100 g, i.p. ) for 11 days;
iii) Lipoic acid group (Sco+LA) -receiving LA (daily orally (100 mg/kg; 0.4 ml/100 g) for 51 days and at the end of the treatment together with injected scopolamine 2.0 mg/kg for 11 days;
iv) Citicoline group (Sco+citicoline) -receiving citicoline daily orally (100 mg/kg 0.5 ml/100 g) (51 days) and at the end of the treatment together with injected scopolamine 2.0 mg/kg for 11 days;
v) Lipoic acid and citicoline group (Sco+LA+citicoline) - receiving for 51 days the combination ALA+Citicoline daily orally (respectively 100 mg/kg, 0.4 ml/100 g LA and 100 mg/kg; 0.5 ml/100 g citicoline) and at the end of the treatment together with injected scopolamine 2.0 mg/kg for 11 days;
vi) Experimental combination group (Sco+E)- receiving for 51 days the combination of α-lipoic acid, citicoline, extract of leaves of green tea, extract of leaves of olive tree, Vitamin D3, selenium, and an immune supporting complex (in the results and figures indicated as experimental combination, E) orally (0.5 ml/100 g) and at the end of the treatment together with injected scopolamine 2.0 mg/kg for 11 days.
2.4. Experimental design
All behavioural, biochemical and histological changes induced by the experimental combination, lipoic acid, citicoline and the combination of lipoic acid and citicoline were evaluated using a model of Alzheimer’s type dementia in rats. Experimental dementia of Alzheimer’s type was chemically induced by repeated administrations of scopolamine hydrobromide (11 days, 2.0 mg/kg intraperitoneally ) in laboratory male Wistar rats based on previous studies of ours [23].
The experimental combination as well as LA, citicoline and LA and citicoline together were applied daily orally in the corresponding doses for 40 days before and 11 days simultaneously with Sco (i.e. 51 days).
In the course of the experiment, several behavioral tests such as step-through inhibitory avoidance, T-maze, Barnes maze, Novel object recognition were conducted (Figure 1).
On the day after behavioral tests were performed (12th day from the start of scopolamine treatment) rats were euthanized under CO2 inhalation.
C reactive protein and parameters of toxicity (ASAT, ALAT, creatinine, albumin, globuline, total protein) were determined in blood serum/plasma of the rats.
The internal organs of animals - liver, kidneys, small intestines and stomach - were taken for histological examination and for evaluation of possible toxic effects of the substances.
Brains were quickly removed and two brain structures related to memory- hippocampus and prefrontal cortex were carefully isolated on ice and used for biochemical studies: acetylcholinesterase (AChE) activity, oxidative stress and antioxidant defence system (LPO; levels ot tGSH, SOD, catalase, GPx activity, brain-derived neurotrophic factor (BDNF) and phosphorylated cAMP response element-binding protein (pCREB).
Advanced exploratory analysis of the principle components (rotation Equamax) and additional hierarchical cluster analysis were used for calculation of the index of recovery in each group.
2.5. Behavioral experimental methods
Several behavioural tests were used to evaluate changes in the memory and learning abilities of the experimental animals after application of scopolamine and to assess the restoring capacity ot the experimental combination and its components.
2.5.1. Step-through passive avoidance
The test is one of the most commonly used to assess changes in learning and memory in rodents and especially suitable to study both the animals’ short and long-term memory as recall can be monitored (tested) easily in a one-trial session, at different times after acquisition [100].
The apparatus consists of two chambers, a dark and an illuminated one, separated by a guillotine door. The floor of the dark chamber is lined with a still grid for delivering electric shocks whenever the animal has stepped with all four paws in the chamber. The experiment was conducted in two phases (sessions), a training and a test session as described previosly [23]. In the training session, preceding the start of scopolamine injections, rats were placed in the illuminated chamber and, after a habituation period of 1 min, by opening the guilotine door, allowed to enter the dark chamber where they received an unescapable electric foot shock (0.7 mA, 3 s, once). The time the animals needed to enter the dark chamber was recorded as initial latency (IL). In the test session, performed at the 1st and 24th hour and at the 12th day after scopolamine injections, rats were placed again in the illuminated chamber and again allowed to enter the dark compartment but without receiving a foot shock. The time the animals needed to enter the dark chamber was recorded as step-through latency (STL) (cut-off time 180 sec) and used to evaluate memory and learning impairements in the different experimental groups.
2.5.2. T-maze
The test is particularly appropriate for studying specific aspects of spatial working memory and is less stressful compared to similar tests like the Moris water maze [101], working memory defined as short-time encoding of spatial information to adjust subsequent behavior. Dopaminergic signaling in the prefrontal cortex (PFC), the hippocampus and the striatum has been identified to be involved in the modulation of working memory [102]. The T-maze consists of a starting arm (50x10) and two other arms (40x20) arranged in a T-shape. It is based on the spontaneous or food-stimulated alternation of the T-shaped arms of the maze as animals enter them.
In the second variant, used by us, the innate tendency of the animals to spontaneously alternate shoulders was stimulated by sequentially placing food in the two arms of the maze. The study was conducted in two phases (stages) - "training" - before the start of application of any of the tested substances (pretreatment) and a "test phase" on the 10th, 20th, 30th, 40th day of the pretreatment and on the 12th day after the application of scopolamine following a described protocol [23] using a modified protocol of [103]. Briefly, after a few days when the animals were getting used to the experimenter’s touches (handling), in the next 5 days ("training") they were habituated to the T-maze and prepared for the subsequent test phase. On the first day, the animals were allowed to freely explore the maze for about 15 minutes, with small pieces of food scattered across the maze at short distances to prompt the rodent to associate the maze with food and to motivate it to move forward after being placed at the base of the starting arm. On the second day, the food was only located at the end of the two T-shaped arms to motivate the animal to choose and enter one of those arms. In the most important part of the training, for three consecutive days, each day, the following experiment was conducted (consisting of 10 trials for each animal): the animal was placed at the base of the maze, in a compartment with a sliding (guillotine) door, and after 10 seconds released to spontaneously enter one of the T-shaped arms of the maze. In the first "forced trial", the animal could only enter one arm (the other was closed with a guillotine door) and this was the arm where the food was located. In the following nine trials, both arms were accessible to the animal, but food was to be found only in the arm opposite to the one in which the food was in the previous trial. Entries into a T-arm were recorded as successful if the arm was with food and as unsuccessful if in the opposite arm. If the animal made an unsuccessful entry, it was allowed to continue exploring the maze until finding the food in the opposite arm and eating it. Calculated and compared between groups was the number of successful entries (correct choices) divided by all trials (10) and multiplied by 100 (T-maze performance in %). The ’test phase’ on the corresponding day of the experiment comprised 10 trials and was conducted as described above.
2.5.3. Barnes maze (B-maze)
The test is frequently (ordinarily) used to evaluate changes in spatial learning and memory in rodents [104]. It employs an open, circular platform with a diameter of 122 cm, located 80 cm above the ground, holding 20 holes situated regularly at the platform periphery with a dark box for the animal’s escape under one the holes. A repulsive (aversive) stimulus of strong light (3000 lx) and loud noise (76 dB) was applied to encourage the animal to escape from the open space of the platform, (where placed during the experiment), and hide in the escape box. The evaluation of spatial memory was carried out (achieved) by comparing the times it took the animals to find and enter the hole with the (underneath) escape box (total latency) (and the number of "error" holes before reaching it). The experiment was run, conducted, in two sessions, a “training” and a “test” session, following, with slight modifications, a previously described protocol [28]. The "training" was carried out, performed immediately before the start of the scopolamine injections or, for a group of animals, on the fourth day of scopolamine injections, and the "test session" - on the 12th day after the administration of scopolamine. The training took place over 4 days and during the first day the animals were placed in the centre of the platform and allowed to explore it freely for 5 minutes. During the next 3 days, each day, 4 trials were conducted for each animal, 15 minutes apart, with the animal being placed in the center of the maze in a transparent starting chamber, from which it was released after 30 seconds to move freely in the maze. The time to find and enter the hole with the escape box underneath (and the "error" holes before finding it) was recorded. On the first of the three days, training was conducted without the aversive stimulus, and on the other two days with the strong light and loud noise being on. The test session (on the 12th day after scopolamine administration) was a repetition of the last day of training: 4 trial sessions of 15 min each for each animal, recording the time to find the escape box and the number of unsuccessful trials until it was reached. The maze and escape box, whose location was important not to be changed, were regularly cleaned with 70% ethanol to prevent olfactory cues.
2.5.4. The Novel Object Recognition (NOR)
This test is used to assess changes in non-spatial memory and is a measure of visual (object) recognition memory [105]. It is centered on the recognition of familiar and novel objects using the innate curiosity of rodents and their predisposition to explore the new and unfamiliar and on the fact that if the rat encounters a familiar and a new object it will spend less time at the familiar one.
The experimental set up is a dark plexiglass chamber (60 x 60 x 40 cm) in which the rat can move without any constraints. The study was conducted on two consecutive days. On the first day, the animal was gently placed in the chamber and, for 5 min allowed to freely explore it without any objects in it. On the same day a second five-min phase followed when the animal was again introduced in the chamber but with two identical objects (A+A) in it. On the following day was the actual testing, when the animal was placed in the same environment with two objects, one of which, however, was the old (familiar) one and the other was similar, but different (A+B). The time spent exploring the old and new object was recorded over a period of 5 min. A discrimination index as a measure of recognition memory (RI) was calculated using the formula:
RI= (N/ (N+F)) x100,
where N is the time exploring the new object B and F-the time exploring the familiar object A.
Recognition index values range from 0 to 100%, with RI >50% indicating that animals remember and discriminated objects, and RI <50 are indicative of recognition memory impairments.
2.6. Biochemical methods
2.6.1. Tissue preparation
On the day after the behavioural tests were conducted, the animals were decapitated, brains were quickly removed on ice and cortex and hippocampus separated. Both structures priorly frozen at -25° were cut into smaller tissue fragments and weighted. Fragments were mixed with PBS (0,01 mol/L, pH 7,4) and homogenized (tissue weight (g): PBS (ml), volume 1:9) with a Potter-Elvehjem glass homogenizer with a Teflon pestle. Homogenized tissue was centrifuged for 5 min at 5000 g and the supernatant was used for determining acetylcholinesterase (AChE) activity, the levels of lipid peroxidation (LPO) and the content of total glutathione (tGSH), as well as to measure BDNF and pCREB. Portion of the same diluted homogenate was centrifuged for 20 min at 12,000g and the resulting postmitochondrial supernatant was used for the determination of the antioxidant enzyme activities: superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx). All procedures were kept under temperature control, between 0° and +4°C.
2.6.2. AChE activity
Assessed in the brain’s structures based on the Ellman’s method [106]. The Ellman’s method is a spectrophotometry-based assay in which the increase in the yellow colour of thiocholine produced by the catalytic action of acetylcholinesterase when 5,5’- dithio-bis(2-nitrobenzoic acid (DTNB) is added, is followed. 100 µl of the supernatant were incubated with the Ellman reagent: 2.9 mL of 0.1M phosphate buffer (pH 8), 100 µl of 0.1M DTNB and 20 µl of 0.075M freshly prepared acetylthiocholine iodide. 500 µl of the reaction mixture were injected into the Semi-auto Chemistry Analyzer and kinetics of the reaction was monitored for 3 min at 405 nm.
All biomarkers for oxidative stress were measured spectrophotometrically using kits, purchased from Sigma-Aldrich Co. LLC, USA.
2.6.3. Lipid peroxidation was determined by Lipid Peroxidation (MDA) Assay Kit MAK085 and was expressed as nmoles malondialdehyde (MDA) mg/protein, using a molar extinction coefficient 1.56 x 106M-1 cm-1.
2.6.4. Total glutathione concentration was measured using Glutathione Assay Kit CS0260. The reaction gave a yellow color compound with an absorption peak at 412 nm. The amount of glutathione was calculated from the reference standard and expressed as ng/mg protein.
2.6.5. Superoxide dismutase activity was measured by SOD Assay Kit-WST 19160. The water-soluble tetrazolium (WST-1) was reduced by the superoxide radicals, generated in the xanthine-xanthine oxidase system, to formazan. The inhibition of the WST-1 reduction was considered a measure of SOD activity as one unit was defined as the amount of enzyme needed to inhibit the WST reduction by 50%.
2.6.6. Catalase аctivity was determined by using Catalase Assay Kit CAT100. The absorption decrease at 240 nm corresponded to the decomposition of H2O2 and was a measure of the catalase activity. Enzyme activity was expressed as ΔA240/min/mg protein.
2.6.7. Glutathione peroxidase activity was measured using Glutathione Peroxidase Cellular Activity Assay CGP1. The enzyme activity was expressed as units/mg protein using a molar extinction coefficient 6.22 mM-1cm-1 for NADPH.
2.6.8. Protein concentrations were measured according to Lowry et al. [107] by using a standard curve for bovine serum albumin.
2.6.9. BDNF was measured using a commercially available sandwich ELISA kit (MyBioSource.Inc., USA, Catalog No MBS457896). Briefly, 100 µl of BDNF standards (0.156–10 ng/ml) and samples were added to the BDNF pre-coated 96-well plate. The plate was covered and incubated for 90 minutes at 37° and further the liquid was removed from each well. Next, 100 µl Detection Solution A (Catalog No MBS457896) were added to each well and the plate incubated for 45 minutes at 37°, followed by a wash for 1-2 minutes 3 times with a wash buffer. In the next step 100 µl Detection Solution B (Catalog No MBS457896) were added to each well and the plate incubated for 45 minutes at 37°. Further the plate was washed for 5 times with a wash-buffer, followed by incubation with TMB Substrate Solution for 15-25 minutes. The addition of 3,3′,5,5′-tetramethylbenzidine (TMB) gave a start to the color reaction, stopped 10 min later with a Stop solution, followed by the immediate recording of the absorbance at 450 nm (MR-96A microplate reader). Samples and standards were run and BDNF concentrations calculated using a standard curve.
2.6.10. pCREB was measured using a commercially available sandwich ELISA kit (MyBioSource.Inc., USA, Catalog No MBS1600625). Briefly, 50 µl of pCREB standards (0-12 ng/ml) and samples were added to the pCREB pre-coated 96-well plate. The plate was covered and incubated for 60 minutes at 37° and further the liquid was removed from each well. Next, 50 µl Detection Solution A and 50 µl Detection solution B were added to each well and the plate incubated for 10 minutes at 37°. In the next step 50 µl Stop solution were added to each well followed by the immediate recording of the absorbance at 450 nm (MR-96A microplate reader). Samples and standards were run and pCREB concentrations calculated using a standard curve.
2.6.11. ALAT and ASAT activities: Blood was collected in serum separator tubes, centrifuged at 3000 rpm for 10 minutes to obtain serum, which was stored at -20° in aliquots. The activity of the two enzymes was determined using Giesse Diagnostics kit, Italy. Liquid reagents and serums were left at room temperature (15-25°) before use. 1000 µl of Reagent A and Reagent B (Catalog No 4194, volume 4:1) were pipetted into test tubes. Next 100 µl serum were added and the mixture gently shaken and left at 37° for 5 minutes. 500 µl of the reaction mixture were injected into the Semi-auto Chemistry Analyzer and kinetics was monitored for 3 minutes at 340 nm, using water as a reference.
2.6.12. Creatinine: Blood was collected in serum separator tubes, centrifuged at 3000 rpm for 10 minutes to obtain serum, which was stored at -20° in aliquots. The activity of creatinine was determined using Giesse Diagnostics kit, Italy. Liquid reagents and serums were left at room temperature (15-25°) before use. 1000 µl of Reagent A and Reagent B (volume 1:1) were pipetted into test tubes. Next 100 µl serum were added and the mixture gently shaken and left at 37° for 30 seconds. 500 µl of the reaction mixture were injected into the Semi-auto Chemistry Analyzer and kinetics was monitored for 3 minutes at 340 nm, using water as a reference.
2.6.13. Total protein was determined by using Reagent ready for use (Giesse Diagnostics kit, Italy). Liquid reagents and serums were left at room temperature (15-25°) before use. 1000 µl of Reagent were mixed with 12.5 µl serum, gently shaken and left at 37° for 10 min. 500 µl of the reaction mixture were injected into the Semi-auto Chemistry Analyzer and read at 546 nm against a reagent blank.
2.6.14. Albumin was determined by using Reagent ready for use (BioSino Bio-Technology and Science Inc., Switzerland). Liquid reagents and serums were left at room temperature (15-25°) before use. 1500 µl of Reagent were pipetted into test tubes. Next 5 µl serum were added, the mixture gently shaken and left at 37° for 5 min. 500 µl of the reaction mixture were injected into the Semi-auto Chemistry Analyzer and read at 630 nm against a reagent blank. Globulins were determined as Globulins = Total protein – Albumin
2.6.15. C-reactive protein (CRP): We used a commercially available sandwich ELISA kit (MyBioSource.Inc., USA, Catalog No MBS457896a). Blood was collected in serum separator tubes, centrifuged at 3000 rpm for 10 minutes to obtain serum, which was further used at 500-fold dilution. Briefly, 100 µl CRP standards (0.78–50 ng/ml) and samples were applied to the CRP pre-coated 96-well plate. The plate was covered and incubated for 60 minutes at 37° and further the liquid was removed from each well. Next, 100 µl Detection Solution A were added to each well and the plate incubated for 60 minutes at 37°, followed by a wash for 1-2 minutes 3 times with a wash buffer. In the next step 100 µl Detection Solution B were added to each well and the plate incubated for 30 minutes at 37°. Further the plate was washed for 5 times with a wash-buffer, followed by incubation with Substrate Solution for 10-20 minutes. The reaction was stopped with a Stop solution, followed by the immediate recording of the absorbance at 450 nm (MR-96A microplate reader).
2.7. Histological studies
After dissection and macroscopic examination of the thoracic and abdominal cavity of the rats, stomach, liver, both kidneys and small intestine were separated, and the materials were fixed in 10% neutral buffered formalin for 24 to 48 hours. After samples were taken for histological analysis, they were placed in biopsy cassettes for further processing (DIAPATH model: DONATELLO FAST). For the purpose of the study, the materials were stained by routine histological staining with hematoxylin-eosin.
2.8. Statistical analysis
Results are presented as means ± SEM. Statistical analysis of the parameters was performed using GraphPad Prism 5.0 and one-way analysis of variance (ANOVA) followed by the Dunnett’s post hoc test. In all statistical analyses, an acceptable level of confidence probability P < 0.05, divided into three ascending classes: P < 0.05, P < 0.01 (high significance) and P < 0.001 (very high statistical significance) was assumed.
3. Results
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3.1. Behavioural assessment of the effects of the experimental combination on learning and memory
In Figure 2A–C it is shown that scopolamine treatment significantly decreases the step-through latency (STL) at 1h and 24 h without having an effect on the 12th day after the beginning of scopolamine treatment. At 1h, all four groups (Sco+E; Sco+citicoline; Sco+LA; Sco+LA+citicoline) demonstrated no significant differences with the control group with, however, the group treated with the experimental combination showing STL values closest to those of the control group. At 24h, the groups treated with citicoline, the combination of LA and citicoline and the experimental combination showed a significant difference with the scopolamine-treated group increasing the STL values, but it was the group treated with the experimental combination that showed the highest difference with the scopolamine-treated group the experimental combination practically completely restoring the decreased by scopolamine STL values. At the 12th day the effects of all 5 groups were very much comparable with no significant differences between neither of the groups, although STL values for the group treated with the experimental combination (and with LA) were highest.
In Figure 2D it is shown that scopolamine significantly decreases the performance of the rats in the T-maze. Although all four groups (Sco+E; Sco+citicoline; Sco+LA; Sco+LA+citicoline) demonstrated no significant differences with the control group, it was the group treated with the experimental combination that showed a significant difference with the scopolamine treated group and it was the experimental combination that practically completely restored the impaired by scopolamine T-maze performance.
Figure 3 presents the effect on the Barnes maze performance of the experimental combination and its components when the Barnes maze training was carried out before scopolamine treatment (A) and when the training was carried out under scopolamine treatment (starting on the 4th day of scopolamine treatment). In the first case scopolamine did not change the total latency, neither did citicoline, LA or the two antioxidants together. However, the experimental combination significantly reduced the total latency compared to control showing its favourable effect on spatial memory. In the second case, scopolamine significantly increased the total latency that was restored to control levels by all four groups, but most efficiently by the experimental combination and LA+citicoline which groups showed a significant decrease in total latency values compared to the scopolamine group.
Figure 4 demonstrates that scopolamine treatment resulted in significant lowering of the RI values in the NOR test and significant impairments of recognition memory as an RI index lower than 50% indicates recognition memory decline. In all four groups this decline was effectively counteracted as those groups showed no differences in their RI values compared to the control group. The best effect was shown by the combination LA and citicoline which most efficiently increased RI values showing (out of all other groups) the biggest difference with the scopolamine group.
3.2. Biochemical assessment of the effects of the experimental combination
3.2.1. Effect of the experimental combination and its components on brain AChE activity
The experimental combination and its components inhibited the scopolamine-increased acetylcholinesterase activity in rats with Alzheimer’s type dementia, both in cortex and in hippocampus (Figure 5). More pronounced were the effects in hippocampus, where both the AChE-increasing effect of scopolamine and, in turn, the AChE-decreasing effects of the experimental combination and its components, reAChEd statistical significance. The decrease of the scopolamine–induced increase in AChE activity was comparable for the experimental combination and its components.
3.2.2. Antioxidant potential of the experimental combination and its components
The level of MDA, thus LPO, was significantly increased by scopolamine in both cortex and hippocampus (Figure 6A,B). In cortex, all four groups (Sco+E; Sco+citicoline; Sco+LA; Sco+LA+citicoline) showed no significant differences with the control group and a significant decrease compared to the scopolamine group. The effects of the experimental combination, citicoline and lipoic acid alone and the combination of citicoline+lipoic acid on scopolamine-increased MDA levels, were almost identical. In hippocampus, in the groups Sco+E and Sco+citicoline the MDA levels were restored to control levels and both groups showed no significant difference with the control group and a significant decrease compared to the scopolamine group. The strongest beneficial effect was held by the experimental combination showing the biggest difference with the scopolamine group.
Lower content (although not reaching statistical significance) of the tGSH in the cortex of the rats from the scopolamine group in comparison to the control group was observed (Fig 5, C). In the other four groups (Sco+E; Sco+citicoline; Sco+LA; Sco+LA+citicoline) the level of the tGSH was comparable with that of the control group. The Sco+citicoline group is standing out showing a significant increase in tGSH levels compared with the scopolamine-treated group. In hippocampus, the level of tGSH was comparable in the different groups with only the group treated with the combination of LA and citicoline showing significantly elevated tGSH levels compared to control animals (Fig 6, D).
3.2.3. Effect of the experimental combination and its components on SOD, catalase and GPx activity
SOD activity was not statistically significantly different between the scopolamine and control group in both cortex and hippocampus. In both cortex and hippocampus, however, SOD activity was higher in the group treated with the Experimental combination reaching statistical significance in hippocampus when compared to both the control and scopolamine group (Figure 7A,B).
Both in cortex and hippocampus no significant differences in the CAT activity of the scopolamine-treated and control group were measured. From the tested substances and their combinations, however, in the cortex, the treatment with citicoline and especially with LA led to a significant decrease in the CAT activity compared to the control group (also compared to the scopolamine group in the case of LA). In hippocampus, the treatment with the experimental combination; the combination of LA and citicoline and, especially, with citicoline, lead to a significant increase in the CAT activity compared to the control group (also compared to to the scopolamine group in the case of citicoline) (Figure 6C,D).
In the cortex, GPx activities remained unchaged in all groups. In hippocampus, however, a statistically significant increase (elevation) in the GPx activity was observed in the groups treated with the experimental combination, the combination of citicoline and LA and especially with citicoline. In the latter case the increase was significant not only compared to the control group, but compared to the scopolamine-treated group as well (Figure 7E,F).
3.3. Effect of the experimental combination and its components on the brain BDNF/CREB signaling pathway
Scopolamine significantly decreased BDNF levels both in cortex and hippocampus (Figure 8A,B). In cortex, the experimental combination, citicoline and the combination of citicoline and LA restored the decreased by scopolamine BDNF levels showing no difference with the control group and a significant difference with the scopolamine-treated group. LA failed to restore the decreased by scopolamine BDNF levels and the BDNF level in this group was comparable to the BDNF level in the scopolamine group. In hippocampus the experimental combination, LA and the combination of LA and citicoline returned BDNF levels to control values, with the best effect shown by the experimental combination and the combination of LA and citicoline demonstrating the biggest and most significant increase in BDNF levels compared to the scopolamine group.
Scopolamine significantly decreased pCREB levels both in cortex and hippocampus (Figure 8C,D). In cortex none of the tested substances managed to restore pCREB values with all four groups showing a significant decrease in pCREB levels compared to the control group. In hippocampus the experimental combination and the combination of lipoic acid and citicoline increased the decreased by scopolamine pCREB values and the groups treated with the experimental combination as well as with LA+citicoline showed no differences with the control group of animals. The groups treated with LA and citicoline alone showed pCREB levels similar to those of the scopolamine group and a significant decrease compared to the control group.
3.4. Histological evaluation of pathological changes in stomach, small intestine, liver and kidney of male Wistar rats treated with the еxperimental combination and its components
The data for the combination are compared with those of the individual components in Table 1.
Table 1. Evaluation of pathological changes in stomach, small intestine, liver and kidney of male Wistar rats treated with the experimental combination.
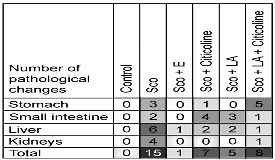
The table presents the number of histopathological changes reflecting inflammatory processes, distributed by organs and studied groups (15 histological preparations per group were studied). Animals treated with scopolamine showed the highest number of lesions, while in the controls practically no inflammatory processes were observed.
Closest to healthy animals is the group treated with the experimental combination (only one inflammatory change in the liver). Rats treated daily with citicoline, lipoic acid, and the combination of citicoline and lipoic acid occupied an intermediate position, with 7, 5, and 8 pathological changes, respectively.
The evaluation of the pathological changes in the stomach, small intestine, liver and kidney of male Wistar rats treated long-term with the experimental combination recorded on the 12th day after the daily administration of scopolamine shows almost identical histological findings compared to the control group treated only with saline: the stomach lacks inflammatory changes, mucinous availability is reduced; in the small intestine, no significant differences were found with the controls - rate of changes (=0); no significant changes in the liver, except for a single initially forming intraparenchymal lymphoid aggregate - rate of changes (=1); in kidneys, no significant differences with the controls were found - rate of changes (=0).
Animals treated with the combination had the highest level of recovery from scopolamine, followed by those treated with lipoic acid alone, citicoline alone, and the combination of lipoic acid and scopolamine.
3.5. Biochemical evaluation of toxicity of the experimental combination and its components (ALAT, ASAT, creatinine, albumin, globulin, total protein assays)
In Figure 9 the effects of scopolamine and the experimental combination and its components on ASAT and ALAT activity and creatinine, albumin, globulin and total protein levels are presented. Any differences between neither of the groups were observed for ASAT and ALAT activity and for creatinine, globulin and total protein levels. The only effect was observed for albumin where scopolamine significantly decreased the the control levels with all the substances counteracting this change and all four groups showing no differences with the control group.
3.6. Evaluation of the anti-inflammatory effect of the combination and its components (C-reactive protein assay)
Scopolamine showed no significant inflammatory effect with CRP levels in blood being only slightly increased in the scopolamine group of animals compared to the control group (Figure 10). It is interesting, however, that both the experimental combination and its components significantly reduced the level of CRP compared to the scopolamine animals and even to the control animals (for lipoic acid and the combination of lipoic acid and citicoline) indicative for a certain anti-inflammatory effect of all the substances.
3.7. Mathematical processing of the experimental data
3.7.1. Evaluation of grouping of the experimental data into separate clusters
Figure 11 presents the results of a principal component analysis pooling histopathology and behavioral memory assessment data at 1 hour, 24 hours, and 12 days after scopolamine administration. Statistical analyses which integrated behavioral and histological data showed clearly distinguished groups organized in separate clusters.
Each dot corresponds to an individual animal – ten in a group; where fewer markers are visible means overlapping or exactly the same values. The formation of distinct groups - clusters testifies to the presence of a sensitive pharmacological effect. Clustering was observed corresponding to the type of experimental treatment of the individual groups. Scopolamine-treated rats (black circles) are grouped in the extreme left position, while healthy animals (white circles) are in the extreme right position.
Citicoline-treated animals also formed a distinct cluster. The remaining experimental groups strongly overlapped. It is noted that the individual values in the demented animals are the most scattered, while those in the controls are the most densely clustered.
The rats treated with alpha lipoic acid and its combination with citicoline occupied an intermediate position between the two extreme groups. Closest to the healthy animals are those treated with the experimental combination. The remaining groups are highly overlapping. The results of this analysis are consistent with the data from the histopathological examination.
3.7.2. Relationship between pathological changes and rate of recovery-combined behavioural and histology data
Figure 12 demonstrates that the experimental combination-treated dement group is very close to healthy controls (restoration coefficient 91 %) followed by those treated with lipoic acid, citicoline and the combination of citicoline and lipoic acid.
4. Discussion
In the present study, using behavioral and biochemical methods, we examined the effect on learning and memory and the neuroprotective effect of an experimental combination consisting of α-lipoic acid, citicoline, extract of leaves of green tea, extract of leaves of olive tree, Vitamin D3, Selenium, and an immune supporting complex in a model of scopolamine-induced Alzheimer-type dementia in rats. We further evaluated the mechanism of the effect of the experimental combination as well as its toxicity and safety.
Scopolamine is a muscarinic cholinergic receptor (mACh Rs) antagonist, which causes memory and cognitive deficits (impairments) elicited by dysfunction of the brain cholinergic system, unfavourable changes in the monoaminergic neurotransmitter systems, increased oxidative stress and impaired antioxidant defense, mitochondrial damage, apoptosis, neuroinflammation and reduced BDNF-CREB, pathological changes found in AD as well [4,5,6,7,8,9,108,109]. Furthermore, scopolamine contributes directly to AD pathology by stimulating Aβ generation and increasing Aβ levels as well as by increasing the amount of the phosphorylated tau protein [110,111,112,113]. The former effect is possibly carried out in part by hampering muscarinic transmission as treatment with selective muscarinic agonists has been shown to decrease cerebrospinal fluid levels of Aβ in AD patients [114,115]. Thus, we considered scopolamine a suitable tool to assess the value of the experimental combination both for memory and cognition and as a possible assisting therapy in AD.
Several behavioural methods such as step-through inhibitory avoidance, T-maze, Barnes maze and NOR (the novel object recognition task) were used to evaluate the changes in learning and memory induced by scopolamine and the capacity of the experimental combination and its components to counteract them.
Step-through inhibitory avoidance is one of the most widely used behavoiural methods to assess memory and cognition. The step-through inhibitory avoidance task is acquired through associative learning with an emotional and spatial memory component to it and is dependent on the integrated activity of several brain areas such as the entorhinal and posterior parietal cortex and hippocampus with dorsal hippocampus being of utmost importance in the acquisition, consolidation and retrieval of the rat’s passive avoidance response [116]. The activation of the cholinergic system through binding of Ach released by cholinergic fibers to muscarinic cholinergic receptors in the hiposcampus plays a key role in inhibitory avoidance processes as administration of mAChRs antagonists, such as atropine, has been shown to worsen and of mAChRs agonists, such as oxotremorine or the muscarinic toxin MT2, to facilitate the acquisition of inhibitory avoidance tasks [117,118,119].
As a muscarinic cholinergic antagonist scopolamine administered intraperitoneally, intracerebrally or intrahippocampally has been plentifully shown to impair the performance of inhibitory avoidance tasks [120]. Our results showing scopolamine-induced decline in recall by significantly reducing the step-through latency at 1h and 24 hours is in agreement with data in the literature showing decrease in this parameter at 1 and 24 hours both in mice and rats [18,121,122,123]. Interestingly, but not surprisingly, scopolamine did not impair the inhibitory avoidance task on the 12th day after daily scopolamine injections. This effect, which we observed in previous experiments as well, is corroborated by findings that different molecular mechanisms may underlie short-term and long-term memory formation [23,124]. In the present experiments the experimental combination restored the STL values of scopolamine-treated rats to that of control rats at both 1h and 24 h showing the best effect compared to citicoline and LA acid alone and the combination of LA and citicoline, especially at 24 hours.
Various mazes are used to assess spatial learning and memory in rats. In our studies we chose the T-maze that relies on positive reinforcement and the Barnes maze, that relies on repulsive agitation (incitement) such as the combination of bright light and noise. Barnes maze is similar to the traditionally used Moris water maze, but avoids a strong aversive stimulus such as swimming as high levels of stress are known to influence the animals’’ performance [125]. Spatial learning and memory are dependent on the hippocampus, on proper functioning of the cholinergic system and very much on the activation of the hippocampal M1 muscarinic cholinergic receptors. Indeed, tasks requiring spatial information processing and spatial memory formation are impaired in patients with hippocampal brain damage and animal studies have shown that hippocampal lesions produce deficits in performance of the water maze [126,127] and the radial arm maze [128] tasks. Furthermore (next), lesions of the cholinergic neurons in the medial septum providing cholinergic input to the hippocampus have been shown to impair both acquisition and maintenance of spatial memory in the T-maze (e.g. Johnson et al, 2002), the water maze [129,130,131,132] tasks. Finally, drugs acting selectively on M1 cholinergic receptors have been demonstrated to influence spatial-memory related tasks [134]. For example, the selective M1 muscarinic receptor antagonist pirenzepine significantly reduces the spontaneous alteration performance in an Y maze and McN-A-343, a selective M1 muscarinic receptor agonist, significantly improved the pirenzepine-induced impairment. As an M1 muscarinic receptor antagonist, though not selective and thus with even broader effects on cognition, scopolamine has been shown to disrupt spatial learning and memory in the radial arm maze [135,136] in the Moris water maze [137] as well as the T-maze [28,138,139] and the Barnes maze [28,140]. In agreement with the literature, in our studies, scopolamine impaired both the T-maze and the Barnes maze tasks reflected in lower T-maze performance and higher total latency in the Barnes maze in scopolamine treated rats. In the case of the Barnes maze task the effect of scopolamine was significant only when training was carried out under scopolamine. This is in line with the finding that it is during acquisition of spatial learning, that ACh is released in hippocampus and cortex (whereas during consolidation of spatial memory ACh levels remain low), underlining the importance for spatial memory formation of ACh receptor blockade during acquisition. In all spatial memory tests the experimental combination showed the best effects. In the T-maze test although no differences in performance were observed between scopolamine-injected rats treated with the experimental combination, citicoline, LA or citicoline+LA and control rats, it was scopolamine-injected rats treated with the experimental combination that showed significantly higher performance compared to rats treated only with scopolamine and it was the experimental combination that restored the control values completely. In the Barnes maze, when scopolamine was applied post training, total latency was lowest (thus animals fastest to find the safety box) in the group treated with the experimental combination and the total latency values of this group were significantly lower even than the values of the control group. When training was performed under scopolamine and scopolamine significantly increased total latency it was again the experimental combination (but also citicoline) that most efficiently counteracted the effect of scopolamine, significantly decreasing the scopolamine-raised values and most effectively returning them to control values.
Additional to the effect on spatial memory we evaluated the effect of the experimental combination on recognition memory using NOR. The test is used to assess changes in non-spatial memory and is a measure of visual (object) recognition memory. It is based entirely on the rats’ spontaneous behavior and can be considered a "pure" test for working memory without a reference component, dependent on the rodent’s natural exploratory instinct, in the absence of externally imposed rules or punishments [105]. Recognition memory involves processing of several types of information such as objects and locations, is mediated through several brain regions, especially the hippocampus and the the perirhinal and medial prefrontal cortices and is distinct from spatial memory [141]. Similar to spatial memory, however, scopolamine impairs recognition memory too [142] as shown in our experiments as well. Additional to improving scopolamine-impaired spatial memory the experimental combination is capable of restoring recognition memory as well by increasing the values of the discrimination index returning them to control values.
Scopolamine is well known to induce cholinergic system dysfunction. Additional to being a muscarinic receptor antagonist and muscarinic cholinergic receptors activation has a central role in synaptic communication and plasticity, learning and memory formation and cognitive function [143,144,145] it reduces acetylcholintransferase activity involved in the synthesis of ACh and increases AChE and butyrylcholinesterase activities, relevant to the degradation of ACh [146,147,148]. In agreement with data in the literature our results also show increased AChE activity in cortex and hippocampus of scopolamine-treated rats with the values in hippocampus of scopolamine-treated animals being significantly higher than those of control rats. The experimental combination decreased AChE activity and returned it to control levels. AChE, being the enzyme that breaks down acetylcholine is fundamental for correct cholinergic transmission and its excessive activity has been many times shown to be associated with cognitive and memory impairments in different experimental models [149,150]. However, excessive AChE activity is further detrimental by aggravating AD pathology by interacting with Aβ and promoting amyloid fibril formation and by forming AChE-Aβ complexes which are more harmful than the amyloid fibrils alone [151,152]. Thus, the alleviating effect of the experimental combination on the increase of AChE activity in scopolamine treated rats explains its memory improving effect, but, even more important, prompts its use not only in conditions of memory and learning deficits, but as part of multifactorial AD treatment as well.
The beneficial effect of the experimental combination on memory and cognition in scopolamine-treated rats could be further explained with the observed decreased oxidative stress induced by decreases in lipid peroxidation and increases in CAT, GPx and SOD activities.
Oxidative stress, defined as the imbalance between oxidant molecules and the organism antioxidant defense system and reflected in increased lipid peroxidation, generation of excessive free radicals and altered antioxidant enzyme activities, has been shown to be related to impairments in learning and memory and is considered part of the pathophysiological mechanism involved in the progression of cognitive dysfunction [153,154,155]. Oxidative damage along with accompanying enhanced lipid peroxidation and decreased activity of the major antioxidant enzymes such as SOD, CAT and GPx, has been shown to be a characteristic feature of AD as well [156]. Furthermore, Esposito and colleagues have shown that in human amyloid precursor protein (hAPP) transgenic mice, inactivating one Sod2 allele (Sod2 +/-) and reducing Sod2 activity exacerbates Alzheimer’s disease-like pathology and have suggested that increasing Sod2 activity might be of therapeutic benefit whereas Clausen and colleagues have explicitly demonstrated prevention of cognitive deficits and brain oxidative stress with superoxide dismutase/catalase mimetics in aged mice [157,158].
Our results show that the experimental combination is increasing the activity of SOD in cortex and catalase and GPx activity in hippocampus, which together with its demonstrated AChE-inhibiting ability is setting it as an attractive candidate in the prevention and treatment of cognitive (age) decline and AD.
Although, in comparison with LA and citicoline alone and LA+citicoline, the experimental combination is showing most consistent effects in attenuating oxidative stress (for example LA, compared to both control and even scopolamine, significantly decreases catalase activity in cortex, rather than increasing it; citicoline also decreases catalase activity in cortex), citicoline is also noticeable for its favorable effects, especially in hippocampus. Indeed, in line with our results, data in the literature shows that citicoline is reducing oxidative damage in a variety of conditions (such as cerebral ischemia, lead-induced oxidative injury; arsenic-induced hepatotoxicity and others) including by decreasing lipid peroxidation and increasing the activity of catalase and GPx [53,159,160,161].
While it looks like that the oxidative stress relief carried out by the experimental combination is mostly due to citicoline, its added value compared to the single antioxidant is certainly owed also to Vit D, the green tea and olive tree leaves extract and for sure Se, as GPx enzymes are selen-containing enzymes. Thus, it has been shown that in rats with spinal cord injury sodium selenite treatment greatly decreases the levels of the lipid peroxidation products malondialdehyde and 4-hydroxynonenal and increases the protein and mRNA expression of glutathione peroxidase 4 (Chen et al, 2022). Regarding AD, it has been shown that selenomethionine (one of the main natural sources of Se and found in many food) is protective against Aβ and iron/hydrogen peroxide-mediated hippocampal neuronal death, an effect mediated by increased GPx protein and activity (Xiong et al, 2007). Vitamin D assists in anti-oxidation by up-regulating the expression of glutathione and superoxide dismutase and down-regulating the expression of NO and the inducible nitric oxide synthase (iNOS) [12,162].
Green tea and olive tree leaves extract have also been shown to favourably affect lipid peroxidation and SOD, CAT and GPx, exerting neuroprotective effects and reducing oxidative damage and possibly contributing to the overall effect of the experimental combination [163,164,165,166].
Decyphering further the molecular mechanisms underlying the effect of the experimental combination we examined the changes in BDNF in cortex and hippocampus of scopolamine treated rats and rats treated with the experimental combination and its main components. BDNF is an important neurotrophin involved in neurogenesis, neuronal differentiation and neuronal protection and crucial for learning and memory since it regulates synaptic function, synaptic plasticity and long-term potentiation in hippocampus [108,167]. Accordingly, it has been shown that interference with the BDNF/TrkB signaling pathway either by genetic manipulations or pharmacological interventions causes memory and cognitive impairments and that chronic BDNF deficiency in BDNF knock out mice results in age-dependent learning deficits. Evidence from both patients and animal models has suggested relation of BDNF to AD and its involvement in AD pathology as well. Thus, research has shown alterations in the BDNF levels in AD patients as well as low BDNF mRNA and protein levels in postmortem AD brain samples [108]. Regarding the role of BDNF in the disease amyloid or tau pathologies, although not completely elucidated, protective effect of BDNF against Aβ- induced neurotoxicity in vitro and in vivo in rats has been observed [168]. Scientific reports show that scopolamine decreases BDNF, which is in in agreement with our results, which demonstrate significant reduction in BDNF levels both in cortex and hippocampus of scopolamine treated animals [18,137,169]. The experimental combination most consistently increases BDNF levels, restoring the neurothrophin control values both in cortex and hippocampus. For comparison, although able in hippocampus, in cortex, LA fails to restore BDNF levels. Citicolin alone and in combination with LA is also restoring scopolamine-decreased BDNF values pointing to a significant role of citicoline in the BDNF-increasing effect of the experimental combination. This is not surprising as, recently, alone and in combination with Coenzyme Q and Vit E, citicoline has been reported to exert complex protective effects in neuronal cells exposed to oxidative stress by reducing the gene expression proinflammatory factors such as of IL-6 and TNFα, and by inducing the gene expression of BDNF [170].
The binding of BDNF to its receptor tyrosine kinase TrkB leads to the dimerization and autophosphorylation of the receptor and subsequent activation of intracellular signaling cascades through which BDNF induces phosphorylation and activation of the cAMP response element-binding protein (CREB) [171]. Phosphorylation of the transcription factor CREB, the further recruitment of transcription cofactors such as CREB binding protein (CBP) and the following transcription of genes such as Egr-1 is thought to play a major role in synaptic plasticity and long-term potentiation and thus in memory formation [172]. Similar to our studies in which scopolamine reduces the level of phosphorylated CREB, both in cortex and hippocampus, other authors have also shown scopolamine-induced reduction of pCREB expression [18,137]. Neither the experimental combination nor its components managed to restore control pCREB values in cortex, whereas in hippocampus the experimental combination and the combination of LA and citicoline significantly increased the reduced by scopolamine levels of pCREB so that no difference was observed between the control group and the group treated with E and the combination of LA and citicoline.
Together with decreasing oxidative stress and reducing AChE activity, we believe it is to a great extent upregulation of BDNF (and pCREB levels) in scopolamine-treated animals that underlies the memory improving effect of the experimental combination shown in all used behavioural tests (regarding both spatial memory revealed in the Barnes maze and the T-maze and the fear-motivated associative learning and memory revealed in the step-through passive avoidance test). Indeed, several lines of evidence point to a substantial part being played by BDNF and activation of its receptor TrkB in hippocampus-dependent memory tasks such as the various mazes and the passive avoidance tests. First, the Morris water maze and passive avoidance tests are linked to increases in BDNF mRNA expression in the hippocampus [171,173]. Second, genetic manipulations in rodents causing BDNF deficiencies or defects in its receptor TrkB result in compromised hippocampus-dependent memory tasks performances. Thus, BDNF knockout mice show cognitive deficits in the T-maze [174] and BDNF mutant mice and mice overexpressing truncated trkB perform significantly worse in a Morris water maze task [175]. Functional blocking of BDNF, through anti-BDNF antibodies, for example, is again shown to be reflected in memory losses shown in the water maze and passive avoidance tests [176,177]. Additionally, CREB mutant mice and rats subjected to heat stress related to perturbations in the BDNF/ERK1/2/CREB axis show impaired hippocampus -dependent special memory [178,179].
Additional to examining the effects of the experimental combination on learning and different types of memory and unravelling the mechanisms of these effects through studies on AChE activity, oxidative stress and antioxidant defence systems markers as well as BDNF/pCREB signaling we assessed its toxicity and safety of use. Histological evaluation of the pathological changes in stomach, small intestine, liver and kidney of the experimental animals revealed no significant inflammatory processes in the rats treated with the experimental combination and showed histological findings almost identical with those in the control group treated only with saline. The lack of inflammation is also confirmed by the lack of changes in CRP in the group treated with the experimental combination compared to the control group of animals. Additional examination of blood biochemical parameters such as ALAT, ASAT, creatinine, albumin, globulin and total protein confirmed the lack of side effects on liver and kidney function and further excluded any inflammatory effects of the experimental combination. Actually, it is worth noting that the groups treated with the experimental combination and its components all showed significantly lower CRP values compared to the scopolamine treated group, with the groups treated with LA and LA+citicoline showing even lower CRP values that those in the control group.
Finally, to better assess and visualize the effects of the combination and its components, histology and behavioural memory assessment data were subjected to principal component and cluster analyses. The applied statistical methods showed clear clustering of the data corresponding to the type of experimental treatment in the individual groups. The group of scopolamine treated rats and that of the healthy controls are clearly distinguishable. The citicoline group also forms a distinct cluster. Closest to the healthy controls are the rats treated with the experimental combination, also located in a separate cluster. It is noteworthy that the individual values in the demented animals are the most scattered, while those in the controls are the most densely clustered. Furthermore, based on the extracted regression components (PC1 and PC2) a “recovery factor” (index of recory) was calculated indicative for the degree of recovery from scopolamine effects. This factor was set as 100 % for the control group, 0 % for the scopolamine group and appeared to be 91 % for the experimental combination demonstrating the best neuroprotective and memory effects for the experimental combination in comparison with its separate components, showing much lower indexes.
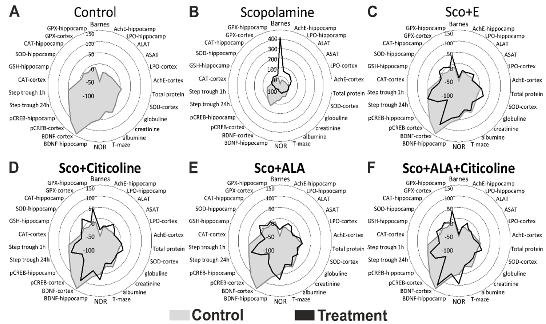
Diagram 1. Radar plots showing the overall treatment profiles, represented as percent change from baseline for each experimental parameter. A profile of healthy controls - depicted in gray, is superimposed on each treatment profile - depicted in black, to illustrate the extent of recovery or impairment (in the case of scopolamine treatment) for each parameter. A. Profile of control treatment; B. Profile of scopolamine treatment; C. Profile of the treatment with scopolamine and the experimental combination (Sco+E); D. Profile of the treatment with scopolamine and citicoline (Sco+Citicoline); E. Profile of the treatment with scopolamine and lipoic acid (Sco+ALA); F. Profile of the treatment with scopolamine and the combination of lipoic acid and citicoline (Sco+ALA+Citicoline).
In summary, our study examined the effect of an inique combination of seven components such as α-lipoic acid, citicoline, extract of leaves of green tea, extract of leaves of olive tree, Vitamin D3, selenium and an immune supporting complex, all of which with well known and proven neuroprotective and memory and cognition preserving and enhancing properties. Compared to its components the experimental combination showed the most complex beneficial effect in a scopolamine model of Alzheimer’s type dementia in rats. It restored efficiently various types of memory such as short-term and long-term memory (as judged by the step-through inhibitory avoidance test) as well as spatial (T-maze, Barnes-maze) and recognition memory (NOR) based on favourable effects as assessed by AChE activity, LPO, tGSH content, SOD, catalase, GPx activity, BDNF and pCREB, which on many occasions exceeded the effects of the separate components (diagram below). The multifaceted beneficial effects of the experimental combination underlined by the versatile and complementary mechanisms of actions of its components combined with lack of toxicity and its anti-inflammatory effects make the experimental combination a suitable candidate for a multitargeted cognition improvement and AD therapy.
5. Patents
The original experimental combination which we investigated in the present study was patented (document № 4391 U1 from 27/10/2022).
Author Contributions
Conceptualization, L.T.; R.K., and P.P.-K.; methodology, L.T. and P.P.-K; software, Y.H.; mathematical analysis, Y.H.; investigation, L.T.; B.M.; D.U.; K.T.; E.T; A.G.; A.A.; M.S.; A.S.; V.G. and M.L.; writing—original draft preparation, PP-K; writing—review and editing, R.K.; visualization, P.P.-K. and Y.H; project administration, R.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Grant D01-217/30.11.2018 under the National Research Program “BioActiveMed” from the Bulgarian Ministry of Education and Science, and also partially supported by Grant KP-06-DV/10-21.12.2019 from the Bulgarian National Science Fund.
Institutional Review Board Statement
Experiments were approved by the Bioethics Committee of the Institute of Neurobiology at the Bulgarian Academy of Sciences according to the "Regulations for Breeding and Experimental Work with Laboratory Animals”. The experimental protocol was in accordance to the requirements of European Communities Council Directive 86/609/EEC.
Acknowledgments
The autors are very grateful to NATSTIM EOOD, and especially to its director Professor Rahamin Shekerdzijski for kind donation of substances used in experiments. The work of Dr. Tsvetomir Garev who performed histopathological evaluations is also very much acknowledged.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be regarded as a potential conflict of interest.
References
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimers Dement. 2018, 6, 575–590. [Google Scholar] [CrossRef]
- Munafò, A.; Burgaletto, C.; Di Benedetto, G.; Di Mauro, M.; Di Mauro, R.; Bernardini, R.; Cantarella, G. Repositioning of Immunomodulators: A Ray of Hope for Alzheimer’s Disease? Front. Neurosci. 2020, 14, 614643. [Google Scholar] [CrossRef]
- Smith, C.D.; Carney, J.M.; Starke-Reed, P.E.; Oliver, C.N.; Stadtman, E.R.; Floyd, R.A.; Markesbery, W.R. Excess brain protein oxidation and enzyme dysfunction in normal aging and in Alzheimer disease. Proc. Natl. Acad. Sci. USA 1991, 88, 10540–10543. [Google Scholar] [CrossRef]
- Baldeiras, I.; Santana, M.T.; Garrucho, R.; Pascoal, A.; Rodrigues, D.; Duro, C.R.; Oliveira. Peripheral oxidative damage in mild cognitive impairment and mild Alzheimer’s disease. J. Alzheimers Dis. 2008, 15, 117–128. [Google Scholar] [CrossRef]
- Padurariu, M.; Ciobica, A.; Hritcu, L.; Stoica, B.; Bild, W.; Stefanescu, C. Changes of some oxidative stress markers in the serum of patients with mild cognitive impairment and Alzheimer’s disease. Neurosci. Lett. 2010, 469, 6–10. [Google Scholar] [CrossRef]
- Kamat, P.K.; Kalani, A.; Rai, S.; Swarnkar, S.; Tota, S.; Nath, C.; Tyagi, N. Mechanism of oxidative stress and synapse dysfunction in the pathogenesis of Alzheimer’s disease: Understanding the therapeutics strategies. Mol. Neurobiol. 2016, 53, 648–661. [Google Scholar] [CrossRef]
- Verri, M.; Pastoris, O.; Dossena, M.; Aquilani, R.; Guerriero, F.; Cuzzoni, G.; Venturini, L.; Ricevuti, G.; Bongiorno, A.I. Mitochondrial alterations, oxidative stress and neuroinflammation in Alzheimer’s disease. Int. J. Immunopathol. Pharmacol. 2012, 25, 345–353. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Nahon-Crystal, E.; ShteinferKuzmine, A.; Gupta, R. VDAC1, mitochondrial dysfunction, and Alzheimer’s disease. Pharmacol. Res. 2018, 131, 87–101. [Google Scholar] [CrossRef]
- Šimić, G.; Babić Leko, M.; Wray, S.; Harrington, C.R.; Delalle, I.; Jovanov-Milošević, N.; Bažadona, D.; Buée, L.; de Silva, R.; Di Giovanni, G.; Wischik, C.M.; Hof, P.R. Monoaminergic neuropathology in Alzheimer’s disease. Prog. Neurobiol. 2017, 151, 101–138. [Google Scholar] [CrossRef]
- Hou, X.Q.; Wu, D.W.; Zhang, C.X.; Yan, R.; Yang, C.; Rong, C.P.; Zhang, L.; Chang, X.; Su, R.Y.; Zhang, S.J.; He, W.Q.; Qu, Z.; Li, S.; Su, Z.R.; Chen, Y.B.; Wang, Q.; Fang, S.H. Bushen-Yizhi formula ameliorates cognition deficits and attenuates oxidative stress-related neuronal apoptosis in scopolamine-induced senescence in mice. Int. J. Mol. Med. 2014, 34, 429–439. [Google Scholar] [CrossRef]
- D’Amelio, M.; Rossini, P.M. Brain excitability and connectivity of neuronal assemblies in Alzheimer’s disease: from animal models to human findings. Prog Neurobiol. 2012, 99, 42–60. [Google Scholar] [CrossRef]
- Wang, L.; Yin, Y.L.; Liu, X.Z.; Shen, P.; Zheng, Y.G.; Lan, X.R.; Lu, C.B.; Wang, J.Z. Current understanding of metal ions in the pathogenesis of Alzheimer’s disease. Transl. Neurodegener. 2020, 9, 10. [Google Scholar] [CrossRef]
- An, Y.; Varma, V.R.; Varma, S.; Casanova, R.; Dammer, E.; Pletnikova, O.; Chia, C.W.; Egan, J.M.; Ferrucci, L.; Troncoso, J.; Levey, A.I.; Lah, J.; Seyfried, N.T.; Legido-Quigley, C.; O’Brien, R.; Thambisetty, M. Evidence for brain glucose dysregulation in Alzheimer’s disease. Alzheimers Dement. 2018, 14, 318–329. [Google Scholar] [CrossRef]
- Yadang, F.S.A.; Nguezeye, Y.; Kom, C.W.; Betote, P.H.D.; Mamat, A.; Tchokouaha, L.R.Y.; Taiwe, G.S.; Agbor, G.A.; Bum, E.N. Scopolamine-induced memory impairment in mice: Neuroprotective effects of Carissa edulis (Forssk.) Valh (Apocynaceae) aqueous extract. Int. J. Alzheimers Dis. 2020, 6372059. [Google Scholar] [CrossRef]
- Haider, S.; Batool, Z.; Ahmad, S.; Siddiqui, R.A.; Haleem, D.J. Walnut supplementation reverses the scopolamine-induced memory impairment by restoration of cholinergic function via mitigating oxidative stress in rats: a potential therapeutic intervention for age related neurodegenerative disorders. Metab. Brain Dis. 2018, 33, 39–51. [Google Scholar] [CrossRef]
- Song, S.H.; Choi, S.M.; Kim, J.E.; Sung, J.E.; Lee, H.A.; Choi, Y.H.; Bae, C.J.; Choi, Y.W.; Hwang, D.Y. α-Isocubebenol alleviates scopolamine-induced cognitive impairment by repressing acetylcholinesterase activity. Neurosci Lett. 2017, 638, 121–128. [Google Scholar] [CrossRef]
- Puangmalai, N.; Thangnipon, W.; Soi-Ampornkul, R.; Suwanna, N.; Tuchinda, P.; Nobsathian, S. Neuroprotection of N-benzylcinnamide on scopolamine-induced cholinergic dysfunction in human SH-SY5Y neuroblastoma cells. Neural Regen Res. 2017, 12, 1492–1498. [Google Scholar]
- Eun, C.S.; Lim, J.S.; Lee, J.; Lee, S.P.; Yang, S.A. The protective effect of fermented Curcuma longa L. on memory dysfunction in oxidative stress-induced C6 gliomal cells, proinflammatory-activated BV2 microglial cells, and scopolamine-induced amnesia model in mice. BMC Complement. Altern. Med. 2017, 17, 367. [Google Scholar] [CrossRef]
- Chen, W.; Cheng, X.; Chen, J.; Yi, X.; Nie, D.; Sun, X.; et al. Lycium barbarum Polysaccharides Prevent Memory and Neurogenesis Impairments in Scopolamine-Treated Rats. PLoS ONE 2014, 9, e88076. [Google Scholar] [CrossRef]
- Giridharan, V.V.; Thandavarayan, R.A.; Sato, S.; Ko, K.M.; Konishi, T. Prevention of scopolamine-induced memory deficits by schisandrin B, an antioxidant lignan from Schisandra chinensis in mice. Free Radic. Res. 2011, 45, 950–958. [Google Scholar] [CrossRef]
- Chan, E.W.L.; Yeo, E.T.Y.; Wong, K.W.L.; See, M.L.; Wong, K.Y.; Gan, S.Y. Piper sarmentosum Roxb. Root Extracts Confer Neuroprotection by Attenuating Beta Amyloid-Induced Pro-Inflammatory Cytokines Released from Microglial Cells. Curr. Alzheimer Res. 2019, 16, 251–260. [Google Scholar] [CrossRef]
- Jahanshahi, M.; Nickmahzar, E.G.; Babakordi, F. The effect of Ginkgo biloba extract on scopolamine-induced apoptosis in the hippocampus of rats. Anat. Sci. Int. 2013, 88, 217–222. [Google Scholar] [CrossRef]
- Lazarova, M.; Tancheva, L.; Alexandrova, A.; Tsvetanova, E.; Georgieva, A.; Stefanova, M.; Tsekova, D.; Vezenkov, L.; Kalfin, R.; Uzunova, D.; Petkova-Kirova, P. Effects of New Galantamine Derivatives in a Scopolamine Model of Dementia in Mice. J. Alzheimers Dis. 2021, 84, 671–690. [Google Scholar] [CrossRef]
- Aleksandrova, S.; Alexova, R.; Dragomanova, S.; Kalfin, R.; Nicoletti, F.; Fagone, P.; Petralia, M.C.; Mangano, K.; Tancheva, L. Preventive and Therapeutic Effects of Punica granatum L. Polyphenols in Neurological Conditions. Int. J. Mol. Sci. 2023, 24, 1856. [Google Scholar] [CrossRef]
- Dragomanova, S.; Lazarova, M.; Munkuev, A.; Suslov, E.; Volcho, K.; Salakhutdinov, N.; Bibi, A.; Reynisson, J.; Tzvetanova, E.; Alexandrova, A.; Georgieva, A.; Uzunova, D.; Stefanova, M.; Kalfin, R.; Tancheva, L. New Myrtenal-Adamantane Conjugates Alleviate Alzheimer’s-Type Dementia in Rat Model. Molecules. 2022, 27, 5456. [Google Scholar] [CrossRef]
- Dragomanova, S.; Pavlov, S.; Marinova, D.; Hodzev, Y.; Petralia, M.C.; Fagone, P.; Nicoletti, F.; Lazarova, M.; Tzvetanova, E.; Alexandrova, A.; Kalfin, R.; Tancheva, L. Neuroprotective Effects of Myrtenal in an Experimental Model of Dementia Induced in Rats. Antioxidants (Basel). 2022, 11, 374. [Google Scholar] [CrossRef]
- Staykov, H.; Lazarova, M.; Hassanova, Y.; Stefanova, M.; Tancheva, L.; Nikolov, R. Neuromodulatory Mechanisms of a Memory Loss-Preventive Effect of Alpha-Lipoic Acid in an Experimental Rat Model of Dementia. J. Mol. Neurosci. 2022, 72, 1018–1025. [Google Scholar] [CrossRef]
- Tancheva, L.; Lazarova, M.; Velkova, L.; Dolashki, A.; Uzunova, D.; Minchev, B.; Petkova-Kirova, P.; Hassanova, Y.; Gavrilova, P.; Tasheva, K.; Taseva, T.; Hodzhev, Y.; Atanasov, A.G.; Stefanova, M.; Alexandrova, A.; Tzvetanova, E.; Atanasov, V.; Kalfin, R.; Dolashka, P. Beneficial Effects of Snail Helix aspersa Extract in an Experimental Model of Alzheimer’s Type Dementia. J. Alzheimers Dis. 2022. [Google Scholar] [CrossRef]
- Tancheva, L.P.; Popatanasov, A.B.; Dragomanova, S.D.; Tzvetanova, E.R.; Aleksandrova, S.M.; Alova, L.G.; Stefanova, M.; Kalfin, R.E. New mechanisms in preventive effect of ellagic acid on cognition in mice with Alzheimer’s disease type dementia. Bul. Chem. Commun. 2018, 50, 20–24. [Google Scholar]
- Aleksandrova, S.M.; Tancheva, L.; Dragomanova, S.; Alova, L.; Stefanova, M.; Georgieva, A.; Simeonova, L.; Pavlova, E.; Kalfin, R. Preventive effect of ellagic acid on cognitive disorders in two mouse models of oxidative stress (influenza infection and scopolamine-induced dementia). Amino Acids 2015, 47, 1651–1651. [Google Scholar]
- Shinto, L.; Quinn, J.; Montine, T.; Dodge, H.H.; Woodward, W.; Baldauf-Wagner, S.; Waichunas, D.; Bumgarner, L.; Bourdette, D.; Silbert, L.; Kaye, J. A randomized placebo-controlled pilot trial of omega-3 fatty acids and alpha lipoic acid in Alzheimer’s disease. J. Alzheimers Dis. 2014, 38, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Sharman, M.J.; Gyengesi, E.; Liang, H.; Chatterjee, P.; Karl, T.; Li, Q.X.; Wenk, M.R.; Halliwell, B.; Martins, R.N.; Münch, G. Assessment of diets containing curcumin, epigallocatechin-3-gallate, docosahexaenoic acid and α-lipoic acid on amyloid load and inflammation in a male transgenic mouse model of Alzheimer’s disease: Are combinations more effective? Neurobiol. Dis. 2019, 124, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Molz, P.; Schröder, N. Potential Therapeutic Effects of Lipoic Acid on Memory Deficits Related to Aging and Neurodegeneration. Front. Pharmacol. 2017, 8, 849. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Tritschler, H.J.; Wessel, K. Neuroprotection by the metabolic antioxidant alpha-lipoic acid. Free Radic. Biol. Med. 1997, 22, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Ahmadvand, H.; Jamor, P. Effects of alpha lipoic acid on level of NO and MPO activity in diabetic rats. Ann. Res. Antioxid. 2017, 2, 2. [Google Scholar]
- Seifar, F.; Khalili, M.; Khaledyan, H.; et al. α-Lipoic acid, functional fatty acid, as a novel therapeutic alternative for central nervous system diseases: a review. Nutr. Neurosci. 2019, 22, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar] [CrossRef]
- Ou, P.; Tritschler, H.J.; Wolff, S.P. Thioctic (lipoic) acid: a therapeutic metal-chelating antioxidant? Biochem. Pharmacol. 1995, 50, 123–126. [Google Scholar] [CrossRef]
- Kaur, D.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Chigurupati, S.; Alhowail, A.; Abdeen, A.; Ibrahim, S.F.; Vargas-De-La-Cruz, C.; Sachdeva, M.; Bhatia, S.; Al-Harrasi, A.; Bungau, S. Decrypting the potential role of α-lipoic acid in Alzheimer’s disease. Life Sci. 2021, 284, 119899. [Google Scholar] [CrossRef]
- Huang, X.; Atwood, C.S.; Hartshorn, M.A.; Multhaup, G.; Goldstein, L.E.; Scarpa, R.C.; Cuajungco, M.P.; Gray, D.N.; Lim, J.; Moir, R.D.; Tanzi, R.E.; Bush, A.I. The A beta peptide of Alzheimer’s disease directly produces hydrogen peroxide through metal ion reduction. Biochemistry 1999, 38, 7609–7616. [Google Scholar] [CrossRef]
- Li, G.; Fu, J.; Zhao, Y.; Ji, K.; Luan, T.; Zang, B. Alpha-lipoic acid exerts anti-inflammatory effects on lipopolysaccharidestimulated rat mesangial cells via inhibition of nuclear factor kappa B (NF-κB) signaling pathway. Inflammation 2015, 38, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Holmquist, L.; Stuchbury, G.; Berbaum, K.; Muscat, S.; Young, S.; Hager, K.; Engel, J.; Münch, G. Lipoic acid as a novel treatment for Alzheimer’s disease and related dementias. Pharmacol. Ther. 2007, 113, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Santos, P.S.; Feitosa, C.M.; Saldanha, G.B.; TomÃ, A.D.; Feng, D.; de Freitas, R.M. Lipoic acid inhibits caspasedependent and -independent cell death pathways and is neuroprotective against hippocampal damage after pilocarpine-induced seizures. Pharmacol. Biochem. Behav. 2011, 97, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Arivazhagan, P.; Panneerselvam, C. Neurochemical changes related to ageing in the rat brain and the effect of DL-alpha-lipoic acid. Exp. Gerontol. 2002, 37, 1489–1494. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xing, G.Q.; Barker, J.L.; Chang, Y.; Maric, D.; Ma, W.; Li, B.S.; Rubinow, D.R. Alpha-lipoic acid protects rat cortical neurons against cell death induced by amyloid and hydrogen peroxide through the Akt signalling pathway. Neurosci. Lett. 2001, 312, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.F.; Bussiere, J.R.; Hammond, R.S.; Montine, T.J.; Henson, E.; Jones, R.E.; Stackman, R.W., Jr. Chronic dietary alpha-lipoic acid reduces deficits in hippocampal memory of aged Tg2576 mice. Neurobiol. Aging. 2007, 28, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wang, W.; Xu, L.; et al. Activation of Nrf2/ARE pathway alleviates the cognitive deficits in PS1V97L-Tg mouse model of Alzheimer’s disease through modulation of oxidative stress. J. Neurosci. Res., 2019, 97, 492–505. [Google Scholar] [CrossRef]
- Fonte, J.; Miklossy, J.; Atwood, C.; Martins, R. The severity of cortical Alzheimer’s type changes is positively correlated with increased amyloid-beta levels: resolubilization of amyloid-beta with transition metal ion chelators. J. Alzheimers Dis. 2001, 3, 209–219. [Google Scholar] [CrossRef]
- Hager, K.; Kenklies, M.; McAfoose, J.; Engel, J.; Münch, G. Alpha-lipoic acid as a new treatment option for Alzheimer’s disease--a 48th months follow-up analysis. J. Neural. Transm. 2007, Suppl. 72, 189–193. [Google Scholar]
- Fava, A.; Pirritano, D.; Plastino, M.; Cristiano, D.; Puccio, G.; Colica, C.; Ermio, C.; De Bartolo, M.; Mauro, G.; Bosco, D. The Effect of Lipoic Acid Therapy on Cognitive Functioning in Patients with Alzheimer’s Disease. J. Neurodegener. Dis. 2013, 2013, 454253. [Google Scholar] [CrossRef]
- Parachikova, A.; Green, K.N.; Hendrix, C.; LaFerla, F.M. Formulation of a medical food cocktail for Alzheimer’s disease: beneficial effects on cognition and neuropathology in a mouse model of the disease. PLoS One 2010, 5, e14015. [Google Scholar] [CrossRef] [PubMed]
- Secades, J.J.; Gareri, P. Citicoline: pharmacological and clinical review, 2022 update. Rev. Neurol. 2022, 75(s05), S1–S89. [Google Scholar]
- Alvarez, X.A.; Sampedro, C.; Lozano, R.; Cacabelos, R. Citicoline protects hippocampal neurons against apoptosis induced by brain beta-amyloid deposits plus cerebral hypoperfusion in rats. Methods Find Exp. Clin. Pharmacol. 1999, 21, 535–540. [Google Scholar] [CrossRef]
- Schauss AG, Nakazaki E.; 67 - Citicoline (CDP-Choline), Editor(s): Joseph E. Pizzorno, Michael T. Murray, Textbook of Natural Medicine (Fifth Edition), Churchill Livingstone, 2020, Pages 515-525.
- Adibhatla, R.M.; Hatcher, J.F. Citicoline decreases phospholipase A2 stimulation and hydroxyl radical generation in transient cerebral ischemia. J. Neurosci. Res. 2003, 73, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Katsuki, H.; Okuda, S. Arachidonic acid as a neurotoxic and neurotrophic substance. Prog. Neurobiol. 1995, 46, 607–636. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Sun, J.; Zhang, W.; Guo, Z.; Ma, Q. Arachidonic acid metabolism in health and disease. Med. Comm. 2023, 4, e363. [Google Scholar] [CrossRef]
- Jasielski, P.; Piędel, F.; Piwek, M.; Rocka, A.; Petit, V.; Rejdak, K. Application of citicoline in neurological disorders: a systematic review. Nutrients 2020, 12, 3113. [Google Scholar] [CrossRef]
- Secades, J.J. Citicoline: pharmacological and clinical review. Rev. Neurol. 2016, 63, S1–S73. [Google Scholar]
- Gareri, P.; Castagna, A.; Cotroneo, A.M.; Putignano, S.; De Sarro, G.; Bruni, A.C. The role of citicoline in cognitive impairment: pharmacological characteristics, possible advantages, and doubts for an old drug with new perspectives. Clin. Interv. Aging 2015, 10, 1421–1429. [Google Scholar] [CrossRef]
- Bonvicini, M.; Travaglini, S.; Lelli, D.; Antonelli Incalzi, R.; Pedone, C. Is Citicoline Effective in Preventing and Slowing Down Dementia: A Systematic Review and a Meta-Analysis. Nutrients 2023, 15, 386. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, P.E.; Dorado, R.; Zea-Sevilla, M.A. Role of Citicoline in Patients with Mild Cognitive Impairment. Neurosci. Insights 2023, 18, 26331055231152496. [Google Scholar] [CrossRef]
- Piamonte, B.L.C.; Espiritu, A.I.; Anlacan, V.M.M. Effects of Citicoline as an Adjunct Treatment for Alzheimer’s Disease: A Systematic Review. J. Alzheimers Dis. 2020, 76, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I. Research Paper: Citicoline Improves Human Vigilance and Visual Working Memory: The Role of Neuronal Activation and Oxidative Stress. Basic Clin. Neurosci. 2020, 11, 423–432. [Google Scholar]
- Rayman, M.P. The importance of selenium to human health. The Lancet 2000, 356, 233–241. [Google Scholar] [CrossRef]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: recent insights into functions and regulation. Cell Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From selenium to selenoproteins: synthesis, identity, and their role in human health. Antioxid. Redox. Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef]
- Akbaraly, N.T.; Hininger-Favier, I.; Carriere, I.; Arnaud, J.; Gourlet, V.; Roussel, A.N.; Berr, C. Plasma selenium over time and cognitive decline in the elderly. Epidemiology 2007, 18, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Smorgon, C.; Mari, E.; Atti, A.R.; Dalla Nora, E.; Zamboni, P.F.; Calzoni, F.; Passaro, A.; Fellin, R. Trace elements and cognitive impairment: an elderly cohort study. Arch. Gerontol. Geriatr. 2004, 9, 393–402. [Google Scholar] [CrossRef]
- Cardoso, B.R.; Ong, T.P.; Jacob-Filho, W.; Jaluul, O.; Freitas, M.I.; Cozzolino, S.M. Nutritional status of selenium in Alzheimer’s disease patients. Br. J. Nutr. 2010, 103, 803–806. [Google Scholar] [CrossRef]
- Vural, H.; Demirin, H.; Kara, Y.; Eren, I.; Delibas, N. Alterations of plasma magnesium, copper, zinc, iron and selenium concentrations and some related erythrocyte antioxidant enzyme activities in patients with Alzheimer’s disease. J. Trace Elem. Med. Biol. 2010, 24, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Loef, M.; Schrauzer, G.N.; Walach, H. Selenium and Alzheimer’s disease: a systematic review. J. Alzheimers Dis. 2011, 26, 81–104. [Google Scholar] [CrossRef]
- Gwon, A.R.; Park, J.-S.; Park, J.-H.; Baik, S.-H.; Jeong, H.-Y.; Hyun, D.-H.; Park, K.W.; Jo, D.-G. Selenium attenuates A beta production and A beta-induced neuronal death. Neurosci. Lett. 2010, 469, 391–395. [Google Scholar] [CrossRef]
- Du, X.; Wang, C.; Liu, Q. Potential Roles of Selenium and Selenoproteins in the Prevention of Alzheimer’s Disease. Curr. Top. Med. Chem. 2016, 16, 835–848. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Shi, Q.; Zhao, Y.; Xie, Y.; Li, X.; Liu, Q.; Iqbal, J.; Zhang, H.; Liu, X.; Shen, L. Se-Methylselenocysteine (SMC) Improves Cognitive Deficits by Attenuating Synaptic and Metabolic Abnormalities in Alzheimer’s Mice Model: A Proteomic Study. ACS Chem. Neurosci. 2021, 12, 1112–1132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Wu, Q.-Y.; Chen, C.; Zheng, R.; Chen, Y.; Liu, Q.; Ni, J.-Z.; Song, G.-L. Selenomethionine Attenuates the Amyloid-Beta Level by Both Inhibiting Amyloid-Beta Production and Modulating Autophagy in Neuron-2a/AbetaPPswe Cells. J. Alzheimers Dis. 2017, 59, 591–602. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Wu, Q.-Y.; Zheng, R.; Chen, C.; Chen, Y.; Liu, Q.; Hoffmann, P.R.; Ni, J.-Z.; Song, G.-L. Selenomethionine Mitigates Cognitive Decline by Targeting Both Tau Hyperphosphorylation and Autophagic Clearance in an Alzheimer’s Disease Mouse Model. J. Neurosci. 2017, 37, 2449–2462. [Google Scholar] [CrossRef]
- Romero-Márquez, J.M.; Forbes-Hernández, T.Y.; Navarro-Hortal, M.D.; Quirantes-Piné, R.; Grosso, G.; Giampieri, F.; Lipari, V.; Sánchez-González, C.; Battino, M.; Quiles, J.L. Molecular Mechanisms of the Protective Effects of Olive Leaf Polyphenols against Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 4353. [Google Scholar] [CrossRef]
- Abdallah, I.M.; Al-Shami, K.M.; Yang, E.; Wang, J.; Guillaume, C.; Kaddoumi, A. Oleuropein-Rich Olive Leaf Extract Attenuates Neuroinflammation in the Alzheimer’s Disease Mouse Model. ACS Chem. Neurosci. 2022, 13, 1002–1013. [Google Scholar] [CrossRef]
- Han, Y.; Tong, Z.; Wang, C.; Li, X.; Liang, G. Oleanolic acid exerts neuroprotective effects in subarachnoid hemorrhage rats through SIRT1-mediated HMGB1 deacetylation. Eur. J. Pharmacol. 2021, 893, 173811. [Google Scholar] [CrossRef]
- Peng, X.P.; Li, X.H.; Li, Y.; Huang, X.T.; Luo, Z.Q. The protective effect of oleanolic acid on NMDA-induced MLE-12 cells apoptosis and lung injury in mice by activating SIRT1 and reducing NF-κB acetylation. Int. Immunopharmacol. 2019, 70, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Valverde-Salazar, V.; Ruiz-Gabarre, D.; García-Escudero, V. Alzheimer’s Disease and Green Tea: Epigallocatechin-3-Gallate as a Modulator of Inflammation and Oxidative Stress. Antioxidants (Basel). 2023, 12, 1460. [Google Scholar] [CrossRef] [PubMed]
- Brewer, L.D.; Thibault, V.; Chen, K.C.; Langub, M.C.; Landfield, P.W.; Porter, N.M. Vitamin D hormone confers neuroprotection in parallel with downregulation of L-type calcium channel expression in hippocampal neurons. J. Neurosci. 2001, 21, 98–108. [Google Scholar] [CrossRef]
- Alexianu, M.E.; Robbins, E.; Carswell, S.; Appel, S.H. 1Alpha, 25 dihydroxyvitamin D3-dependent up-regulation of calcium-binding proteins in motoneuron cells. J. Neurosci Res. 1998, 51, 58–66. [Google Scholar] [CrossRef]
- Garcion, E.; N Wion-Barbot, C.N. Montero-Menei, F. Berger, D. Wion, New clues about vitamin D functions in the nervous system. Trends Endocrinol. Metab. 2002, 13, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Masakazu, I.; Hideyuki, S.; Miki, N.; et al. Protective effects of 1 alpha,25-(OH)(2)D(3) against the neurotoxicity of glutamate and reactive oxygen species in mesencephalic culture. Neuropharmacology 2001, 40, 761–771. [Google Scholar]
- Ndrepepa, G.; Colleran, R.; Kastrati, A. Gamma-glutamyl transferase and the risk of atherosclerosis and coronary heart disease. Clin. Chim. Acta 2018, 476, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, Y.; Meng, X. Vitamin D and neurodegenerative diseases. Heliyon 2023, 9, e12877. [Google Scholar] [CrossRef] [PubMed]
- Lasoń, W.; Jantas, D.; Leśkiewicz, M.; Regulska, M.; Basta-Kaim, A. The Vitamin D Receptor as a Potential Target for the Treatment of Age-Related Neurodegenerative Diseases Such as Alzheimer’s and Parkinson’s Diseases: A Narrative Review. Cells 2023, 12, 660. [Google Scholar] [CrossRef]
- Balion, C.; Griffith, L.E.; Strifler, L.; et al. Vitamin D, cognition, and dementia: a systematic review and metaanalysis. Neurology 2012, 79, 1397–1405. [Google Scholar] [CrossRef]
- Llewellyn, D.J.; Lang, I.A.; Langa, K.M.; et al. Vitamin D and risk of cognitive decline in elderly persons. Arch Intern. Med. 2010, 170, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Soni, M.; Kos, K.; Lang, I.A.; Jones, K.; Melzer, D.; Llewellyn, D.J. Vitamin D and cognitive function. Scand. J. Clin. Lab. Invest. 2012, 72, 79–82. [Google Scholar]
- Lee, D.M.; Tajar, A.; Ulubaev, A.; Pendleton, N.; O’Neill, T.W.; O’Connor, D.B.; Bartfai, G.; Boonen, S.; Bouillon, R.; Casanueva, F.F.; et al. Association between 25-hydroxyvitamin D levels and cognitive performance in middle-aged and older European men. J. Neurol. Neurosurg. Psychiatry 2009, 80, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Littlejohns, T.J.; Henley, W.E.; Lang, I.A.; Annweiler, C.; Beauchet, O.; Chaves, P.H.M.; Fried, L.; Kestenbaum, B.R.; Kuller, L.H.; Langa, K.M.; et al. Vitamin D and the risk of dementia and Alzheimer disease. Neurology 2014, 83, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Navarro, S.G.; Mimenza-Alvarado, A.J.; Jiménez-Castillo, G.A.; Bracho-Vela, L.A.; Yeverino-Castro, S.G.; Ávila-Funes, J.A. Association of Vitamin D with mild cognitive impairment and Alzheimer’s dementia in older Mexican adults. Rev. Invest. Clin. 2019, 71, 381–386. [Google Scholar] [PubMed]
- Pinzon, R.T.; Handayani, T.; Wijaya, V.O.; Buana, R.B. Low vitamin D serum levels as risk factor of Alzheimer’s disease: a systematic review and meta-analysis. Egypt J. Neurol. Psychiatry Neurosurg. 2023, 59, 88. [Google Scholar] [CrossRef]
- Jayedi, A.; Rashidy-Pour, A.; Shab-Bidar, S. Vitamin D status and risk of dementia and Alzheimer’s disease: A meta-analysis of dose-response. Nutr. Neurosci. 2019, 22, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Chai, B.; Gao, F.; Wu, R.; Dong, T.; Gu, C.; Lin, Q.; Zhang, Y. Vitamin D deficiency as a risk factor for dementia and Alzheimer’s disease: an updated meta-analysis. BMC Neurol. 2019, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Jarvik, M.E.; Kopp, R. An improved one-trial passive avoidance learning situation. Psychol. Rep. 1967, 21, 221–224. [Google Scholar] [CrossRef]
- Wenk, G.L. Assessment of spatial memory using the T maze. Curr. Protoc. Neurosci. Chapter 2001, 8. Unit 8.5B. [Google Scholar] [CrossRef]
- Bezu, M.; Malikovic, J.; Kristofova, M.; Engidawork, E.; Hoger, H.; Lubec, G.; Korz, V. Spatial working memory in male rats: Pre-experience and task dependent roles of dopamine D1- and D2- like receptors. Front Behav. Neurosci. 2017, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.M.; Bezu, M.; Korz, V. Evaluating working memory on a T-maze in male rats. Bio. Protoc. 2018, 8, e2930. [Google Scholar] [CrossRef] [PubMed]
- Barnes, C.A. Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J. Comp. Physiol. Psychol. 1979, 93, 74–104. [Google Scholar] [CrossRef] [PubMed]
- Ennaceur, A.; Delacour, J. A new one-trial test for neurobiological studies of memory in rats. 1: Behavioral data. Behav. Brain Res. 1988, 31, 47–59. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, J.R.; Featherstone, R.M. A new and rapid colori-metric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.W.; Rosenbrough, N.J.; Farr, A.L.; Randal, R.J. Protein measurement with folin phenol reagent. J. Biol. Chem. 1951, 193, 256–275. [Google Scholar] [CrossRef]
- Budni, J.; Bellettini-Santos, T.; Mina, F.; Garcez, M.L.; Zugno, A.I. The involvement of BDNF, NGF and GDNF in aging and Alzheimer’s disease. Aging Dis. 2015, 6, 331–341. [Google Scholar] [PubMed]
- Tanila, H. The role of BDNF in Alzheimer’s disease. Neurobiol. Dis. 2017, 97 Pt B, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Liskowsky, W.; Schliebs, R. Muscarinic acetylcholine receptor inhibition in transgenic Alzheimer-like Tg2576 mice by scopolamine favours the amyloidogenic route of processing of amyloid precursor protein. Int. J. Dev. Neurosci. 2006, 24, 149–156. [Google Scholar] [CrossRef]
- Bihaqi, S.W.; Singh, A.P.; Tiwari, M. Supplementation of Convolvulus pluricaulis attenuates scopolamineinduced increased tau and amyloid precursor protein (AbetaPP) expression in rat brain. Indian J. Pharmacol. 2012, 44, 593–598. [Google Scholar] [CrossRef]
- Safar, H.H.; Arab, S.M.; Rizk, S.A. El-Maraghy, Bone marrow-derived endothelial progenitor cells protect against scopolamine-induced Alzheimer-like pathological aberrations. Mol. Neurobiol. 2016, 53, 1403–1418. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, D.K.; CA Ismail, D.A. Ghareeb, Differential metformin dose-dependent effects on cognition in rats: role of Akt. Psychopharmacology 2016, 233, 2513–2524. [Google Scholar] [CrossRef]
- Hock, C.; Maddalena, A.; Heuser, I.; Naber, D.; Oertel, W.; Von Der Kammer, H.; Wienrich, M.; Raschig, A.; Deng, M.; Growdon, J.H.; Nitsch, R.M. Treatment with the selective muscarinic agonist talsaclidine decreases cerebrospinal fluid levels of total amyloid beta-peptide in patients with Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2000, 920, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Nitsch, R.M.; Deng, M.; Tennis, M.; Schoenfeld, D.; and Growdon, J.H. The selective muscarinic M1 agonist AF102B decreases levels of total Abeta in cerebrospinal fluid of patients with Alzheimer’s disease. Ann. Neurol. 2000, 48, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Lorenzini, C.A.; Baldi, E.; Bucherelli, C.; Sacchetti, B.; Tassoni, G. Role of dorsal hippocampus in acquisition, consolidation and retrieval of rat’s passive avoidance response: a tetrodotoxin functional inactivation study. Brain Res 1996, 730, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Boccia, M.M.; Blake, M.G.; Acosta, G.B.; Baratti, C.M. Atropine, an anticholinergic drug, impairs memory retrieval of a high consolidated avoidance response in mice. Neurosci. Lett. 2003, 345, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, I.; da Cunha, C.; Rosat, R.; Jerusalinsky, D.; Ferreira, M.B.; Medina, J.H. Neurotransmitter receptors involved in post-training memory processing by the amygdala, medial septum, and hippocampus of the rat. Behav. Neural. Biology, 1992, 58, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Jeruzalinsky, D.; Cervenansky, C.; Walz, R.; Bianchin, M.; Izquierdo, I. A peptide muscarinic toxin from the Green Mamba venom shows agonist-like action in an inhibitory avoidance learning task. Eur. J. Pharm. 1993, 240, 103–105. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, M.G.; Lana, D.; Pepeu, G. The integrated role of ACh, ERK and mTOR in the mechanisms of hippocampal inhibitory avoidance memory. Neurobiol. Learn. Mem. 2015, 119, 18–33. [Google Scholar] [CrossRef]
- Cruz-Morales, S.E.; Duran-Arevalo, M.; Diaz Del Guante, M.A.; Quirarte, G.; PradoAlcalá, R.A. A threshold for the protective effect of over-reinforced passive avoidance against scopolamine-induced amnesia. Behav. Neural. Biology 1992, 57, 256–259. [Google Scholar] [CrossRef]
- Nomura, Y.; Nishiyama, N.; Saito, H.; Matsuki, N. Role of cholinergic neurotransmission in the amygdala on performances of passive avoidance learning in mice. Biol. Pharm. Bull. 1994, 17, 490–494. [Google Scholar] [CrossRef]
- Giovannini, M.G.; Pazzagli, M.; Malmberg-Aiello, P.; Della, C.L.; Rakovska, A.D.; Cerbai, F.; et al. Inhibition of acetylcholine-induced activation of extracellular regulated protein kinase prevents the encoding of an inhibitory avoidance response in the rat. Neuroscience 2005, 136, 15–32. [Google Scholar] [CrossRef]
- Lana, D.; Cerbai, F.; Di Russo, J.; Boscaro, F.; Giannetti, A.; Petkova-Kirova, P.; Pugliese, A.M.; Giovannini, M.G. Hippocampal long-term memory: effect of the cholinergic system on local protein synthesis. Neurobiol. Learn. Mem. 2013, 106, 246–257. [Google Scholar] [CrossRef]
- Holscher, C. Stress impairs performance in spatial water maze learning tasks. Behav. Brain Res. 1999, 100, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.G.; Garrud, P.; Rawlins, J.N.; O’Keefe, J. Place navigation impaired in rats with hippocampal lesions. Nature, 1982, 297, 681–683. [Google Scholar] [CrossRef]
- Clark, R.E.; Broadbent, N.J.; Squire, L.R. Impaired remote spatial memory after hippocampal lesions despite extensive training beginning early in life. Hippocampus 2005, 15, 340–346. [Google Scholar] [CrossRef]
- Bouffard, J.P.; Jarrard, L.E. Acquisition of a complex place task in rats with selective ibotenate lesions of hippocampal formation: combined lesions of subiculum and entorhinal cortex versus hippocampus. Behav. Neurosci. 1988, 102, 828–834. [Google Scholar] [CrossRef]
- Johnson, D.A.; Zambon, N.J.; Gibbs, R.B. Selective lesion of cholinergic neurons in the medial septum by 192 IgG-saporin impairs learning in a delayed matching to position T-maze paradigm. Brain Res. 2002, 943, 132–141. [Google Scholar] [CrossRef]
- Hagan, J.J.; Salamone, J.D.; Simpson, J.; Iversen, S.D.; Morris, R.G. Place navigation in rats is impaired by lesions of medial septum and diagonal band but not nucleus basalis magnocellularis. Behav. Brain Res. 1988, 27, 9–20. [Google Scholar] [CrossRef]
- Brandner, C.; Schenk, F. Septal lesions impair the acquisition of a cued place navigation task: attentional or memory deficit? Neurobiol. Learn. Mem. 1998, 69, 106–125. [Google Scholar] [CrossRef]
- Janis, L.S.; Glasier, M.M.; Fulop, Z.; Stein, D.G. Intraseptal injections of 192 IgG saporin produce deficits for strategy selection in spatial-memory tasks. Behav. Brain Res. 1998, 90, 23–34. [Google Scholar] [CrossRef]
- Lehmann, O.; Grottick, A.J.; Cassel, J.C.; Higgins, G.A. A double dissociation between serial reaction time and radial maze performance in rats subjected to 192 IgGsaporin lesions of the nucleus basalis and/or the septal region. Eur. J. Neurosci. 2003, 18, 651–666. [Google Scholar] [CrossRef]
- Brandeis, R.; Dachir, S.; Sapir, M.; Levy, A.; Fisher, A. Reversal of age-related cognitive impairments by an M1 cholinergic agonist, AF102B. Pharmacol. Biochem. Behav. 1990, 36, 89–95. [Google Scholar] [CrossRef]
- Hodges, D.B., Jr.; Lindner, M.D.; Hogan, J.B.; Jones, K.M.; Markus, E.J. Scopolamine induced deficits in a battery of rat cognitive tests: comparisons of sensitivity and specificity. Behav. Pharmacol. 2009, 20, 237–251. [Google Scholar] [CrossRef]
- Kay, C.; Harper, D.N.; Hunt, M. Differential effects of MDMA and scopolamine on working versus reference memory in the radial arm maze task. Neurobiol. Learn. Memory, 2010, 93, 151–156. [Google Scholar] [CrossRef]
- Kwon, S.H.; Ma, S.X.; Joo, H.J.; Lee, S.Y.; Jang, C.G. Inhibitory Effects of Eucommia ulmoides Oliv. Bark on Scopolamine-Induced Learning and Memory Deficits in Mice. Biomol. Ther. (Seoul) 2013, 21, 462–469. [Google Scholar] [CrossRef]
- Spowart-Manning, L.; van der Staay, F. The T-maze continuous alternation task for assessing the effects of putative cognition enhancers in the mouse. Behav. Brain Res. 2004, 151, 37–46. [Google Scholar] [CrossRef]
- Yadang, F.S.A.; Nguezeye, Y.; Kom, C.W.; Betote, P.H.D.; Mamat, A.; Tchokouaha, L.R.Y.; Taiwe, G.S.; Agbor, G.A.; Bum, E.N. Scopolamine-induced memory impairment in mice: Neuroprotective effects of Carissa edulis (Forssk.) Valh (Apocynaceae) aqueous extract. Int. J. Alzheimers Dis 2020, 6372059. [Google Scholar] [CrossRef]
- George, A.; Ng, C.P.; O’Callaghan, M.; Jensen, G.S.; Wong, H.J. In vitro and ex-vivo cellular antioxidant protection and cognitive enhancing effects of an extract of Polygonum minus Huds (Lineminus™) demonstrated in a Barnes Maze animal model for memory and learning. BMC Complement Altern. Med. 2014, 14, 161. [Google Scholar] [CrossRef]
- Okada, K.; Nishizawa, K.; Kobayashi, T.; Sakata, S.; Kobayashi, K. Distinct roles of basal forebrain cholinergic neurons in spatial and object recognition memory. Sci. Rep. 2015, 5, 13158. [Google Scholar] [CrossRef]
- Kim, S.K.; Kwon, D.A.; Kim, Y.S.; Lee, H.S.; Kim, H.K.; Kim, W.K. Standardized Extract (HemoHIM) Protects against Scopolamine-Induced Amnesia in a Murine Model. Evid. Based Complement Alternat. Med. 2021, 2021, 8884243. [Google Scholar] [CrossRef]
- Fernández de Sevilla, D.; Núñez, A.; Buño, W. Muscarinic Receptors, from Synaptic Plasticity to its Role in Network Activity. Neuroscience 2021, 456, 60–70. [Google Scholar] [CrossRef]
- Drever, B.D.; Riedel, G.; Platt, B. The cholinergic system and hippocampal plasticity. Behav. Brain Res. 2011, 221, 505–514. [Google Scholar] [CrossRef]
- Power, A.E.; Vazdarjanova, A.; McGaugh, J.L. Muscarinic cholinergic influences in memory consolidation. Neurobiol. Learn Mem. 2003, 80, 178–193. [Google Scholar] [CrossRef]
- Xu, Q.Q.; Xu, Y.J.; Yang, C.; Tang, Y.; Li, L.; Cai, H.B.; Hou, B.N.; Chen, H.F.; Wang, Q.; Shi, X.G.; Zhang, S.J. Sodium tanshinone IIA sulfonate attenuates scopolamine-induced cognitive dysfunctions via improving cholinergic system. Biomed. Res. Int. 2016, 2016, 9852536. [Google Scholar] [CrossRef]
- Sinha, A.; Tamboli, R.S.; Seth, B.; Kanhed, A.M.; Tiwari, S.K.; Agarwal, S.; Nair, S.; Giridhar, R.; Chaturvedi, R.K.; Yadav, M.R. Neuroprotective role of novel triazine derivatives by activating Wnt/ catenin signaling pathway in rodent models of Alzheimer’s disease. Mol. Neurobiol. 2015, 52, 638–652. [Google Scholar] [CrossRef]
- Akinyemi, A.J.; Oboh, G.; Oyeleye, S.I.; Ogunsuyi, O. Anti-amnestic effect of curcumin in combination with donepezil, an anticholinesterase drug: Involvement of cholinergic system. Neurotox. Res. 2017, 31, 560–569. [Google Scholar] [CrossRef]
- Deshmukh, R.; Kaundal, M.; Banslav, V. Caffeic acid attenuates oxidative stress, learning and memory deficit in intra-cerebroventricular streptozotocin induced experimental dementia in rats. Biomed. Pharmacother. 2015, 81, 56–62. [Google Scholar] [CrossRef]
- Ngoupaye, G.T.; Yassi, F.B.; Bahane, D.A.N.; Bum, E.N. Combined corticosterone treatment and chronic restraint stress lead to depression associated with early cognitive deficits in mice. Metab. Brain Dis. 2017, 33, 1–11. [Google Scholar] [CrossRef]
- Reyes, A.E.; Perez, D.R.; Alvarez, A.; Garrido, J.; Gentry, M.K.; Doctor, B.P.; Inestrosa, N.C. A monoclonal antibody against acetylcholinesterase inhibits the formation of amyloid fibrils induced by the enzyme. Biochem. Biophys. Res. Commun. 1997, 232, 652–655. [Google Scholar] [CrossRef]
- Alvarez, A.; Alarcon, R.; Opazo, C.; Campos, E.O.; Munoz, F.J.; Calderon, F.H.; Dajas, F.; Gentry, M.K.; Doctor, B.P.; De ´ Mello, F.G.; Inestrosa, N.C. Stable complexes involving acetylcholinesterase and amyloid-beta peptide change the biochemical properties of the enzyme and increase the neurotoxicity of Alzheimer’s fibrils. J. Neurosci. 1998, 18, 3213–3223. [Google Scholar] [CrossRef]
- Sato, H.; Takahashi, T.; Sumitani, K.; Takatsu, H.; Urano, S. Glucocorticoid generates ROS to induce oxidative injury in the Hippocampus, leading to impairment of cognitive function of rats. J. Clin. Biochem. Nutr. 2010, 47, 224–232. [Google Scholar] [CrossRef]
- Wang, S.; Irving, G.; Jiang, L.; Wang, H.; Li, M.; Wang, X.; Han, W.; Xu, Y.; Yang, Y.; Zeng, T.; Song, F.; Zhao, X.; Xie, K. Oxidative Stress Mediated Hippocampal Neuron Apoptosis Participated in Carbon Disulfide-Induced Rats Cognitive Dysfunction. Neurochem. Res. 2017, 42, 583–594. [Google Scholar] [CrossRef]
- Chauhan, N.R.; Kumar, R.; Gupta, A.; Meena, R.C.; Nanda, S.; Mishra, K.P.; Singh, S.B. Heat stress induced oxidative damage and perturbation in BDNF/ERK1/2/CREB axis in hippocampus impairs spatial memory. Behav. Brain Res. 2021, 396, 112895. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, C. Oxidative stress in Alzheimer’s disease. Neurosci. Bull. 2014, 30, 271–281. [Google Scholar] [CrossRef]
- Clausen, A.; Doctrow, S.; Baudry, M. Prevention of cognitive defcits and brain oxidative stress with superoxide dismutase/catalase mimetics in aged mice. Neurobiol. Aging 2010, 31, 425–433. [Google Scholar] [CrossRef]
- Esposito, L.; Raber, J.; Kekonius, L.; Yan, F.; Yu, G.Q.; Bien-Ly, N.; Puoliväli, J.; Scearce-Levie, K.; Masliah, E.; Mucke, L. Reduction in mitochondrial superoxide dismutase modulates Alzheimer’s disease-like pathology and accelerates the onset of behavioral changes in human amyloid precursor protein transgenic mice. J. Neurosci. 2006, 26, 5167–5179. [Google Scholar] [CrossRef]
- Adibhatla, R.M.; Hatcher, J.F.; Dempsey, R.J. Citicoline: neuroprotective mechanisms in cerebral ischemia. J. Neurochem. 2002, 80, 12–23. [Google Scholar] [CrossRef]
- Aminzadeh, A.; Salarinejad, A. Citicoline protects against lead-induced oxidative injury in neuronal PC12 cells. Biochem. Cell. Biol. 2019, 97, 715–721. [Google Scholar] [CrossRef]
- Nikravesh, M.; Mahdavinia, M.; Neisi, N.; Khorsandi, L.; Khodayar, M.J. Citicoline ameliorates arsenic-induced hepatotoxicity and diabetes in mice by overexpression of VAMP2, PPAR-γ, As3MT, and SIRT3. Pestic. Biochem. Physiol. 2023, 192, 105391. [Google Scholar] [CrossRef] [PubMed]
- Alamro, A.A.; Alsulami, E.A.; Almutlaq, M.; Alghamedi, A.; Alokail, M.; Haq, S.H. Therapeutic Potential of Vitamin D and Curcumin in an In Vitro Model of Alzheimer Disease. J. Cent. Nerv. Sys. Dis. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, O.; Amit, T.; Mandel, S.; Youdim, M.B. Neuroprotective molecular mechanisms of (-)-epigallocatechin-3-gallate: a reflective outcome of its antioxidant, iron chelating and neuritogenic properties. Genes Nutr. 2009, 4, 283–296. [Google Scholar] [CrossRef]
- Somova, L.I.; Shode, F.O.; Ramnanan, P.; Nadar, A. Antihypertensive, antiatherosclerotic and antioxidant activity of triterpenoids isolated from Olea europaea, subspecies africana leaves. J. Ethnopharmacol. 2003, 84, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Dekanski, D.; Selaković, V.; Piperski, V.; Radulović, Z.; Korenić, A.; Radenović, L. Protective effect of olive leaf extract on hippocampal injury induced by transient global cerebral ischemia and reperfusion in Mongolian gerbils. Phytomedicine 2011, 18, 1137–1143. [Google Scholar] [CrossRef] [PubMed]
- Asghari, A.A.; Hosseini, M.; Bafadam, S.; Rakhshandeh, H.; Farazandeh, M.; Mahmoudabady, M. Olea europaea L. (olive) leaf extract ameliorates learning and memory deficits in streptozotocin-induced diabetic rats. Avicenna J. Phytomed. 2022, 12, 163–174. [Google Scholar] [PubMed]
- Yamada, K.; Nabeshima, T. Brain-derived neurotrophic factor/TrkB signaling in memory processes. J. Pharmacol. Sci. 2003, 91, 267–270. [Google Scholar] [CrossRef]
- Arancibia, S.; Silhol, M.; Moulière, F.; Meffre, J.; Höllinger, I.; Maurice, T.; Tapia-Arancibia, L. Protective effect of BDNF against beta-amyloid induced neurotoxicity in vitro and in vivo in rats. Neurobiol. Dis. 2008, 31, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Cho, N.; Lee, K.Y.; Huh, J.; Choi, J.H.; Yang, H.; Jeong, E.J.; Kim, H.P.; Sung, S.H. Cognitive-enhancing effects of Rhus verniciflua bark extract and its active flavonoids with neuroprotective and anti-inflammatory activities. Food Chem. Toxicol. 2013, 58, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Mastropasqua, L.; Agnifili, L.; Ferrante, C.; Sacchi, M.; Figus, M.; Rossi, G.C.M.; Brescia, L.; Aloia, R.; Orlando, G. Citicoline/Coenzyme Q10/Vitamin B3 Fixed Combination Exerts Synergistic Protective Effects on Neuronal Cells Exposed to Oxidative Stress. Nutrients 2022, 14, 2963. [Google Scholar] [CrossRef] [PubMed]
- Tyler, W.J.; Alonso, M.; Bramham, C.R.; Pozzo-Miller, L.D. From the acquisition to consolidation: on the role of brain-derived neurotrophic factor signaling in hippocamal-dependent learning. Learn. Mem. 2002, 9, 224–237. [Google Scholar] [CrossRef]
- Bartolotti, N.; Lazarov, O. CREB signals as PBMC-based biomarkers of cognitive dysfunction: A novel perspective of the brain-immune axis. Brain Behav. Immun. 2019, 78, 9–20. [Google Scholar] [CrossRef]
- Yamada, K.; Mizuno, M.; Nabeshima, T. Role for Brain-derived neurotrophic factor in learning and memory. Life Sci. 2002, 70, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Rossetti, A.C.; Brandwein, C.; Riva, M.A.; Gass, P.; Elsner, P.; Hesse-Macabata, J.; Hipler, U.C.; Smesny, S.; Milleit, B. Brain Derived Neurotrophic Factor Deficiency is Associated with Cognitive Impairment and Elevated Phospholipase A2 Activity in Plasma of Mice. Neuroscience 2022, 480, 167–177. [Google Scholar] [CrossRef]
- Saarelainen, T.; Pussinen, R.; Koponen, E.; Alhonen, L.; Wong, G.; Sirviö, J.; Castrén, E. Transgenic mice overexpressing truncated trkB neurotrophin receptors in neurons have impaired long-term spatial memory but normal hippocampal LTP. Synapse 2000, 38, 102–104. [Google Scholar] [CrossRef]
- Mu, J.-S.; Li, W.-P.; Yao, Z.-B.; Zhou, X.-F. Deprivation of endogenous brain-derived neurotrophic factor results in impairment of spatial learning and memory in adult rats. Brain Res. 1999, 835, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.; Vianna, M.R.M.; Depino, A.M.; e Souza, T.M.; Pereira, P.; Szapiro, G.; Viola, H.; Pitossi, F.; Izquierdo, I. and Medina JH: BDNF-triggered events in the rat hippocampus are required for both short- and long-term memory formation. Hippocampus 2002, 12, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.R.; Kumar, R.; Gupta, A.; Meena, R.C.; Nanda, S.; Mishra, K.P.; Singh, S.B. Heat stress induced oxidative damage and perturbation in BDNF/ERK1/2/CREB axis in hippocampus impairs spatial memory. Behav. Brain Res. 2021, 396, 112895. [Google Scholar] [CrossRef]
- Bourtchuladze, R.; Frenguelli, B.; Blendy, J.; Cioffi, D.; Schutz, G.; Silva, A.J. Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 1994, 79, 59–68. [Google Scholar] [CrossRef]
Figure 1.
Schematic timeline of the experimental paradigm.
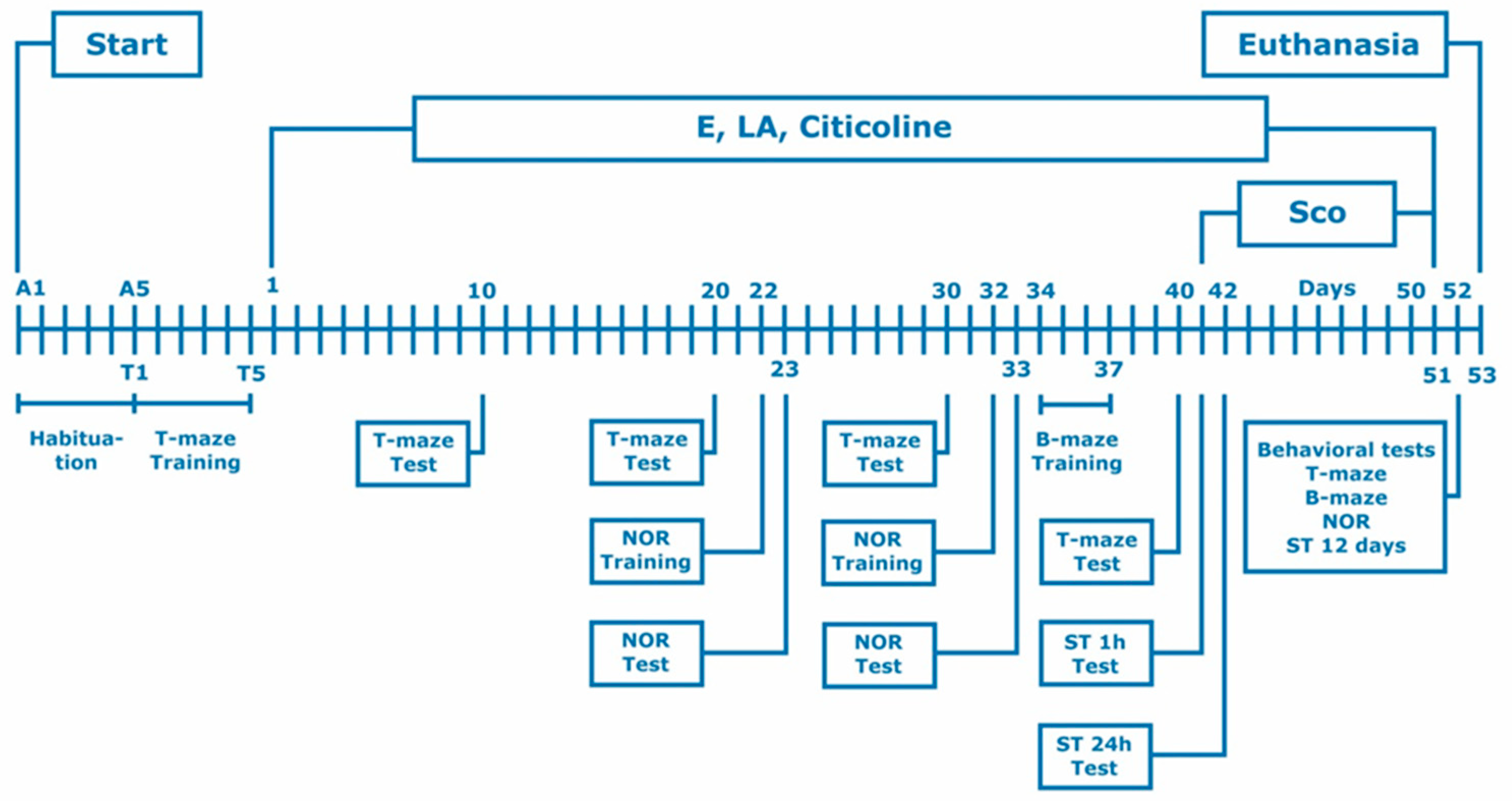
Figure 2.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on learning and memory in a scopolamine model of dementia in rats. Step-through latency is assessed at 1 h, 24 h, and on the 12th day (A–C, respectively) and T-maze performance is evaluated on the 12th day after the beginning of scopolamine treatment (D) for the control, Sco, Sco + E, Sco + citicoline; Sco + LA; Sco+LA+citicoline. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
Figure 2.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on learning and memory in a scopolamine model of dementia in rats. Step-through latency is assessed at 1 h, 24 h, and on the 12th day (A–C, respectively) and T-maze performance is evaluated on the 12th day after the beginning of scopolamine treatment (D) for the control, Sco, Sco + E, Sco + citicoline; Sco + LA; Sco+LA+citicoline. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
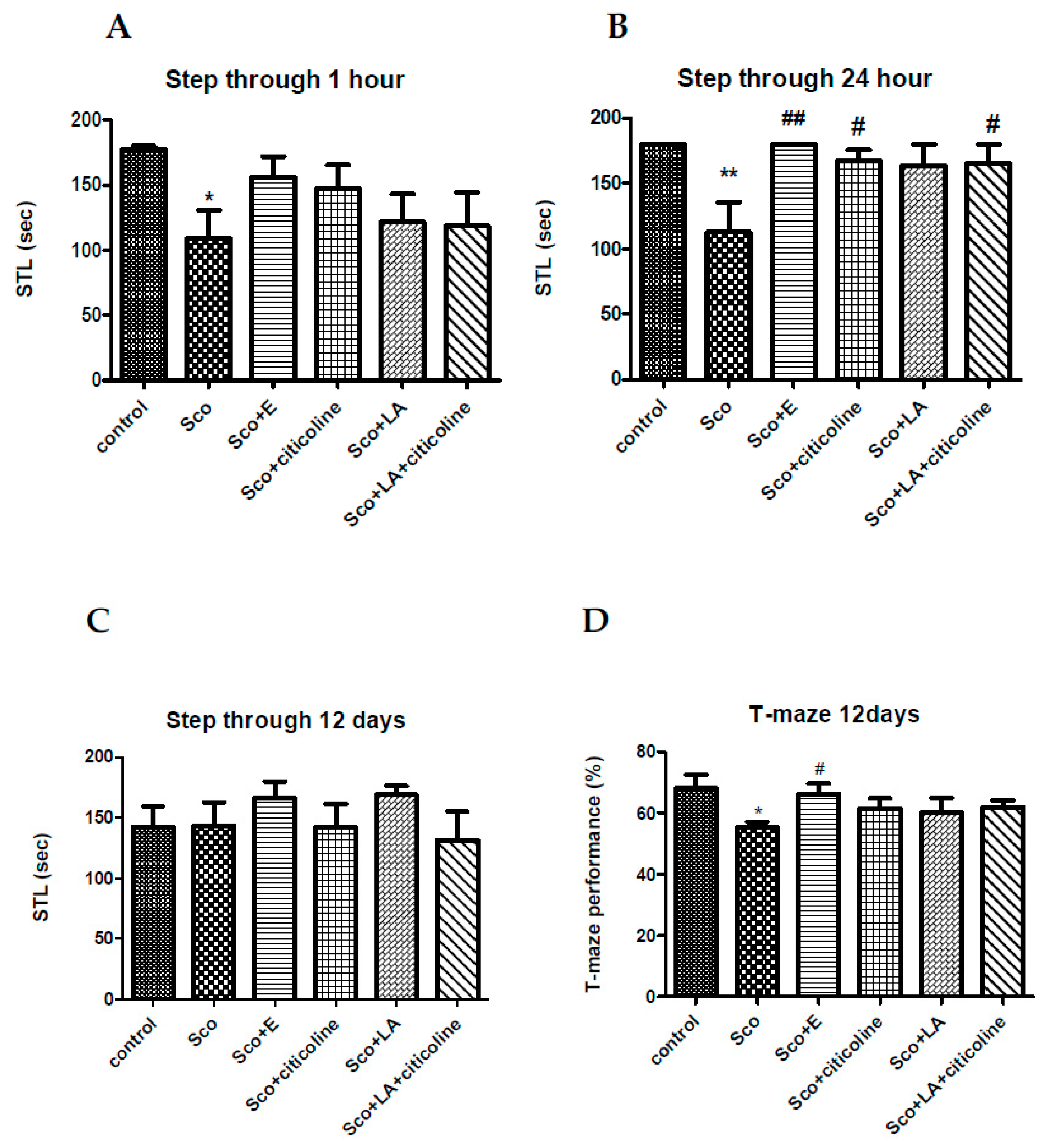
Figure 3.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on learning and memory in a scopolamine model of dementia in rats. Barnes maze total latency is assessed when Barnes maze training was performed before scopolamine treatment (A) and under scopolamine treatment (B) for the control, Sco, Sco + E, Sco + citicoline; Sco + LA; Sco+LA+citicoline. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
Figure 3.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on learning and memory in a scopolamine model of dementia in rats. Barnes maze total latency is assessed when Barnes maze training was performed before scopolamine treatment (A) and under scopolamine treatment (B) for the control, Sco, Sco + E, Sco + citicoline; Sco + LA; Sco+LA+citicoline. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
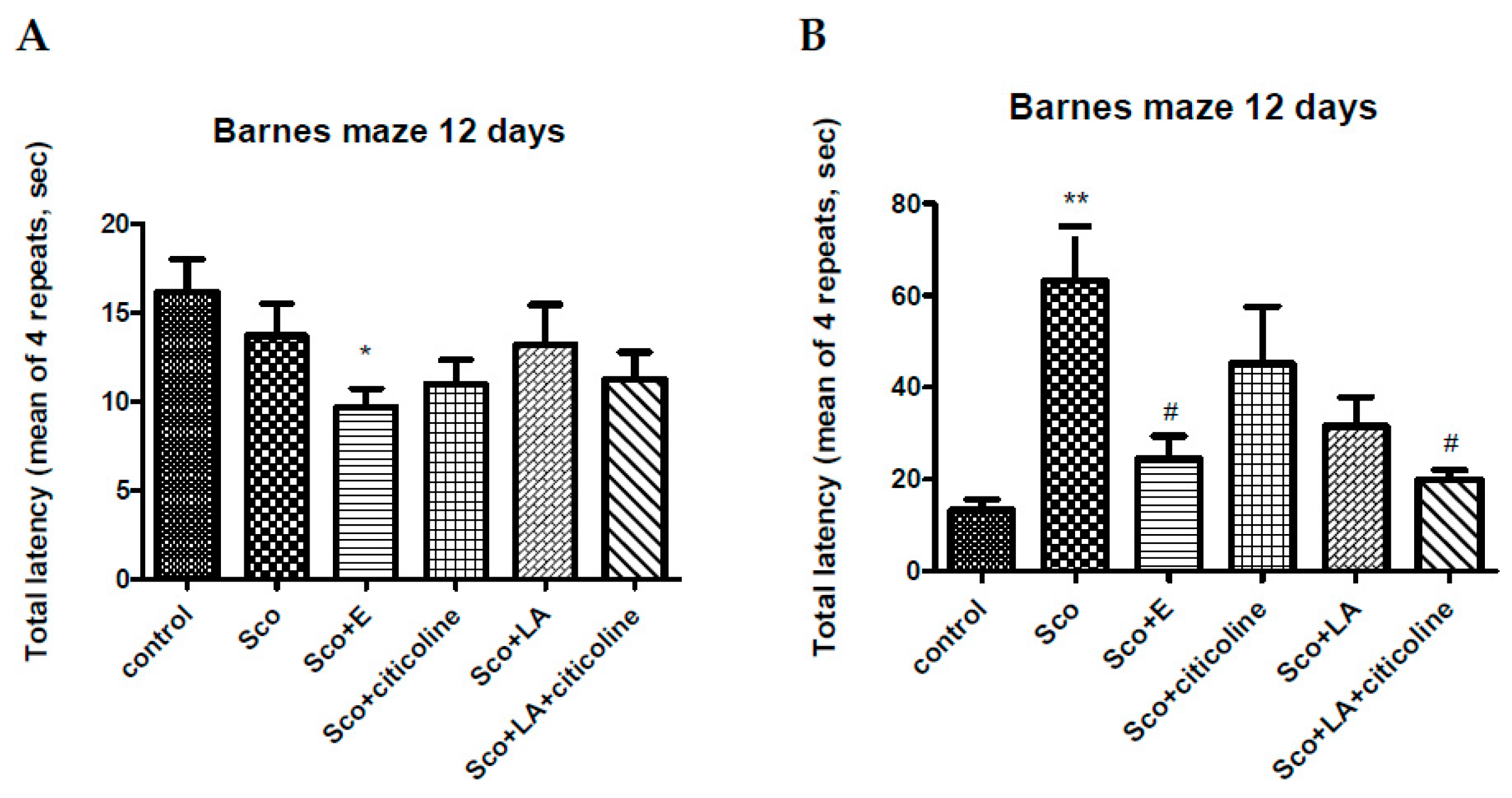
Figure 4.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on learning and memory in a scopolamine model of dementia in rats. RI is evaluated on the 12th day after the beginning of scopolamine treatment (D) for the control, Sco, Sco + E, Sco + citicoline; Sco + LA; Sco+LA+citicoline. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
Figure 4.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on learning and memory in a scopolamine model of dementia in rats. RI is evaluated on the 12th day after the beginning of scopolamine treatment (D) for the control, Sco, Sco + E, Sco + citicoline; Sco + LA; Sco+LA+citicoline. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
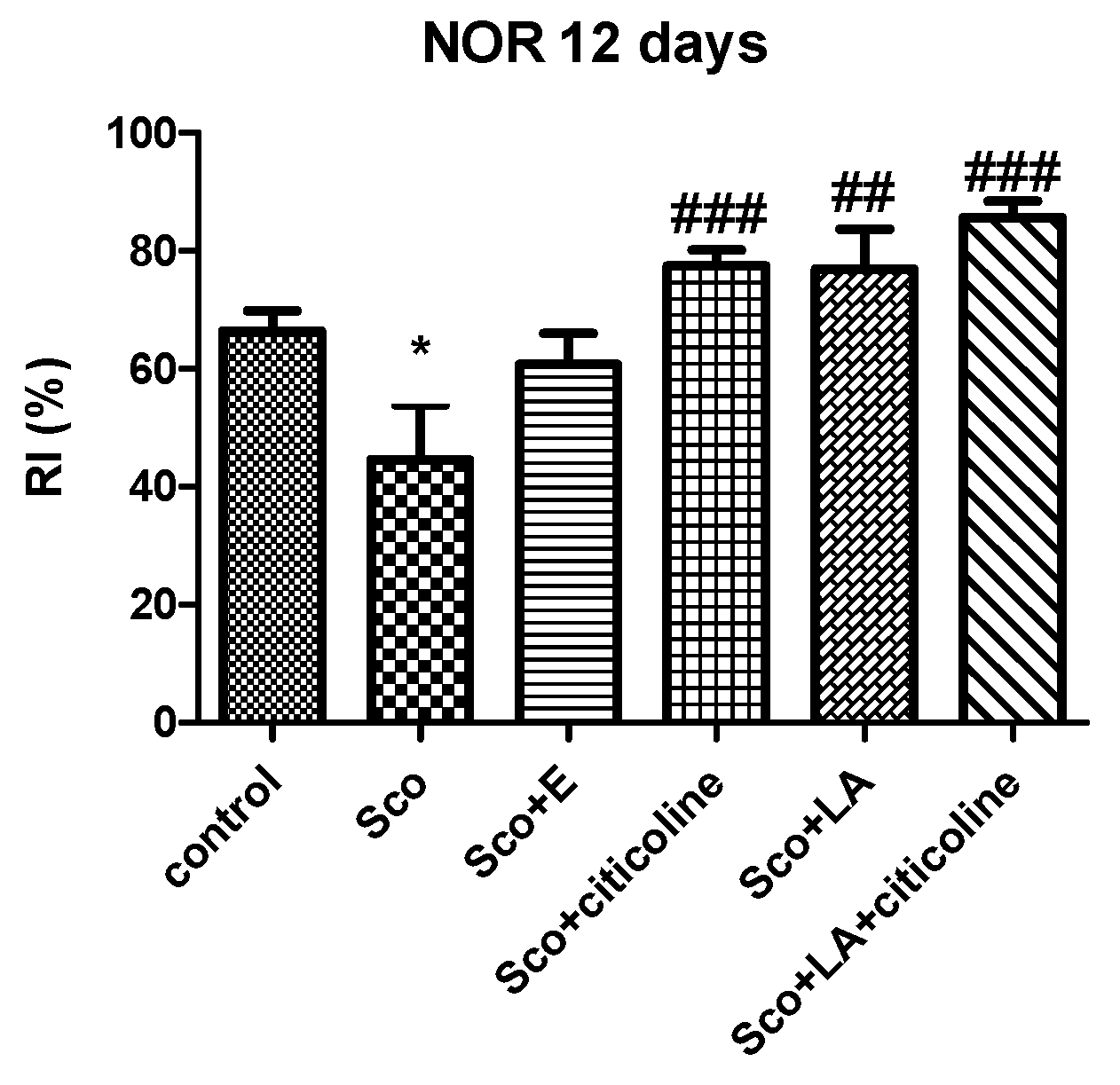
Figure 5.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline), on AChE in cortex (A) and hippocampus (B) of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗”or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
Figure 5.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline), on AChE in cortex (A) and hippocampus (B) of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗”or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
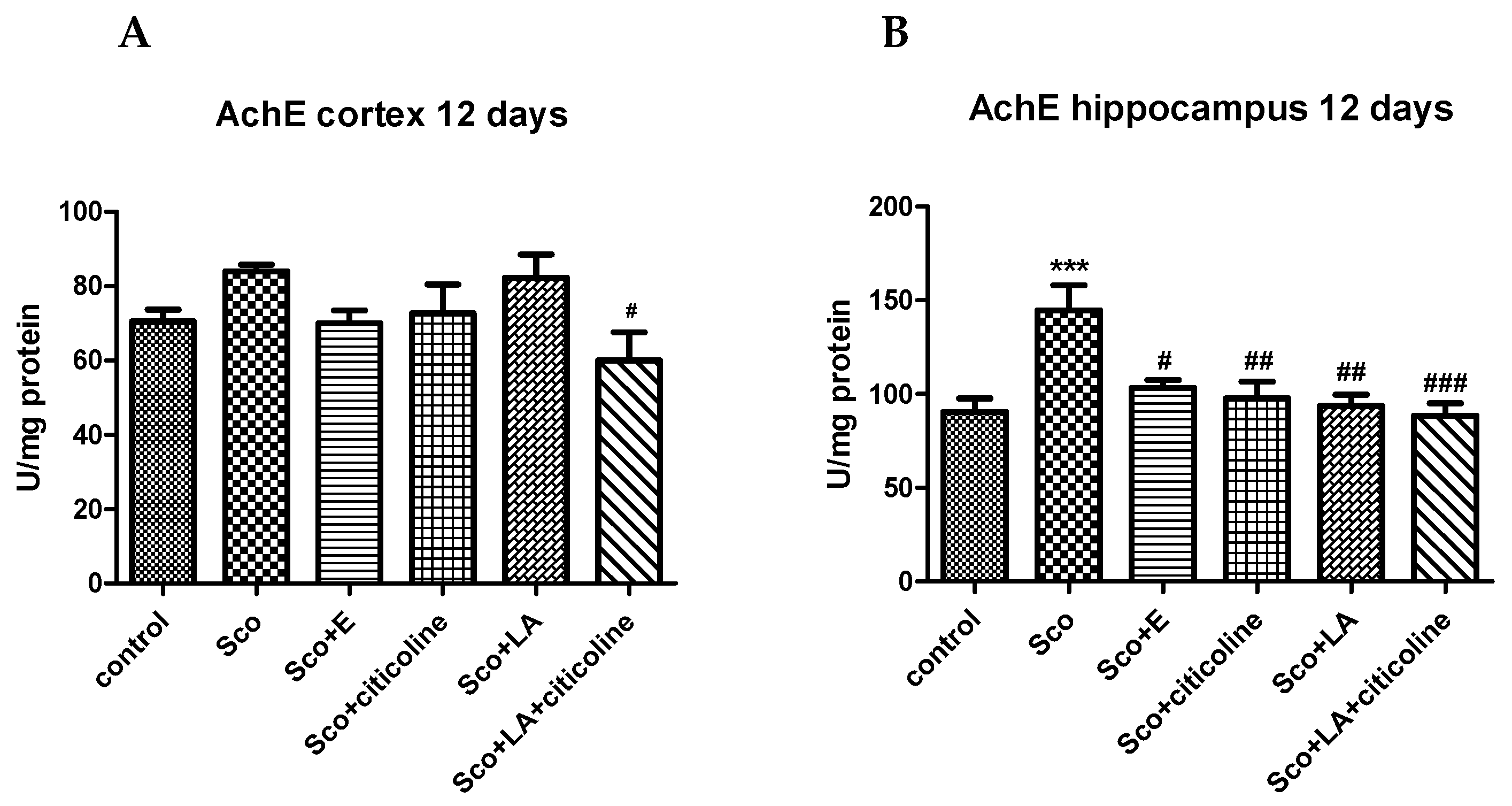
Figure 6.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline), on the MDA levels (as a measure of lipid peroxidation, LPO) and tGSH in cortex (A,C) and hippocampus (B,D) of scopolamine-treated rats.Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
Figure 6.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline), on the MDA levels (as a measure of lipid peroxidation, LPO) and tGSH in cortex (A,C) and hippocampus (B,D) of scopolamine-treated rats.Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
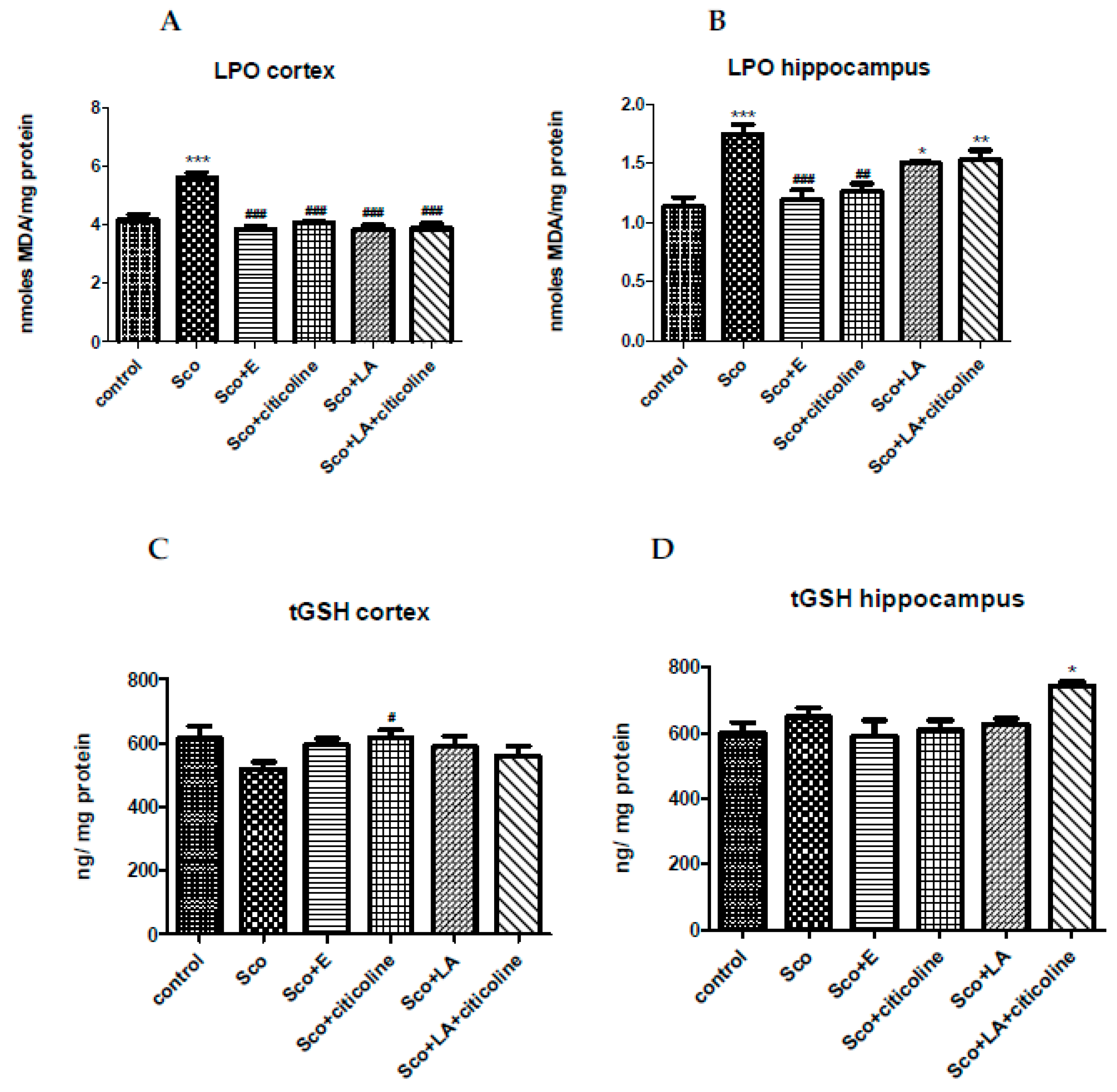
Figure 7.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on SOD, catalase and GPx activity in cortex (A,C,E) and on SOD, catalase and GPx activity in hippocampus (B,D,F) of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
Figure 7.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on SOD, catalase and GPx activity in cortex (A,C,E) and on SOD, catalase and GPx activity in hippocampus (B,D,F) of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
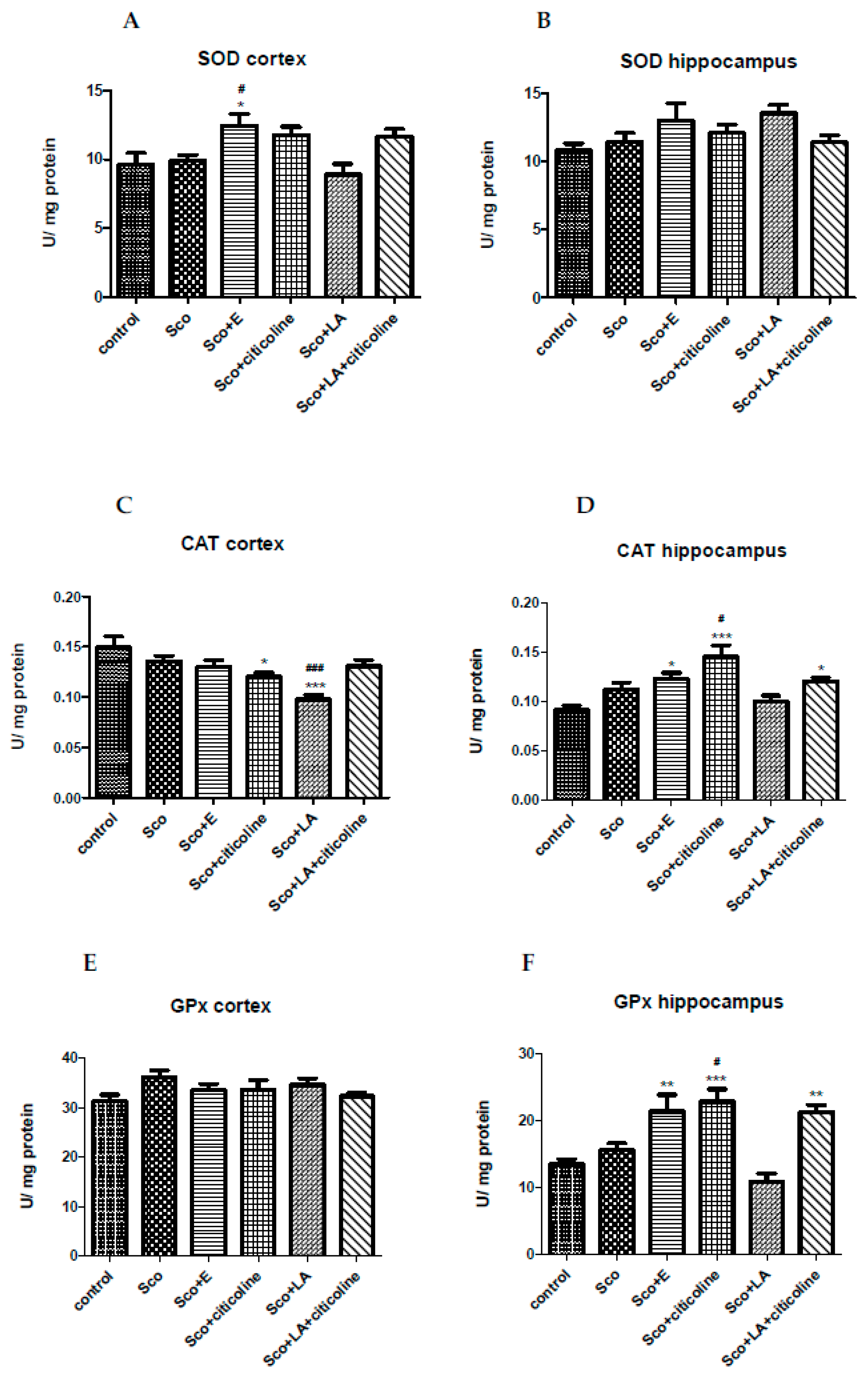
Figure 8.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on BDNF levels in cortex and hippocampus (A,B) and on pCREB levels in cortex and hippocampus (C,D) of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or“#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
Figure 8.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on BDNF levels in cortex and hippocampus (A,B) and on pCREB levels in cortex and hippocampus (C,D) of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or“#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001.
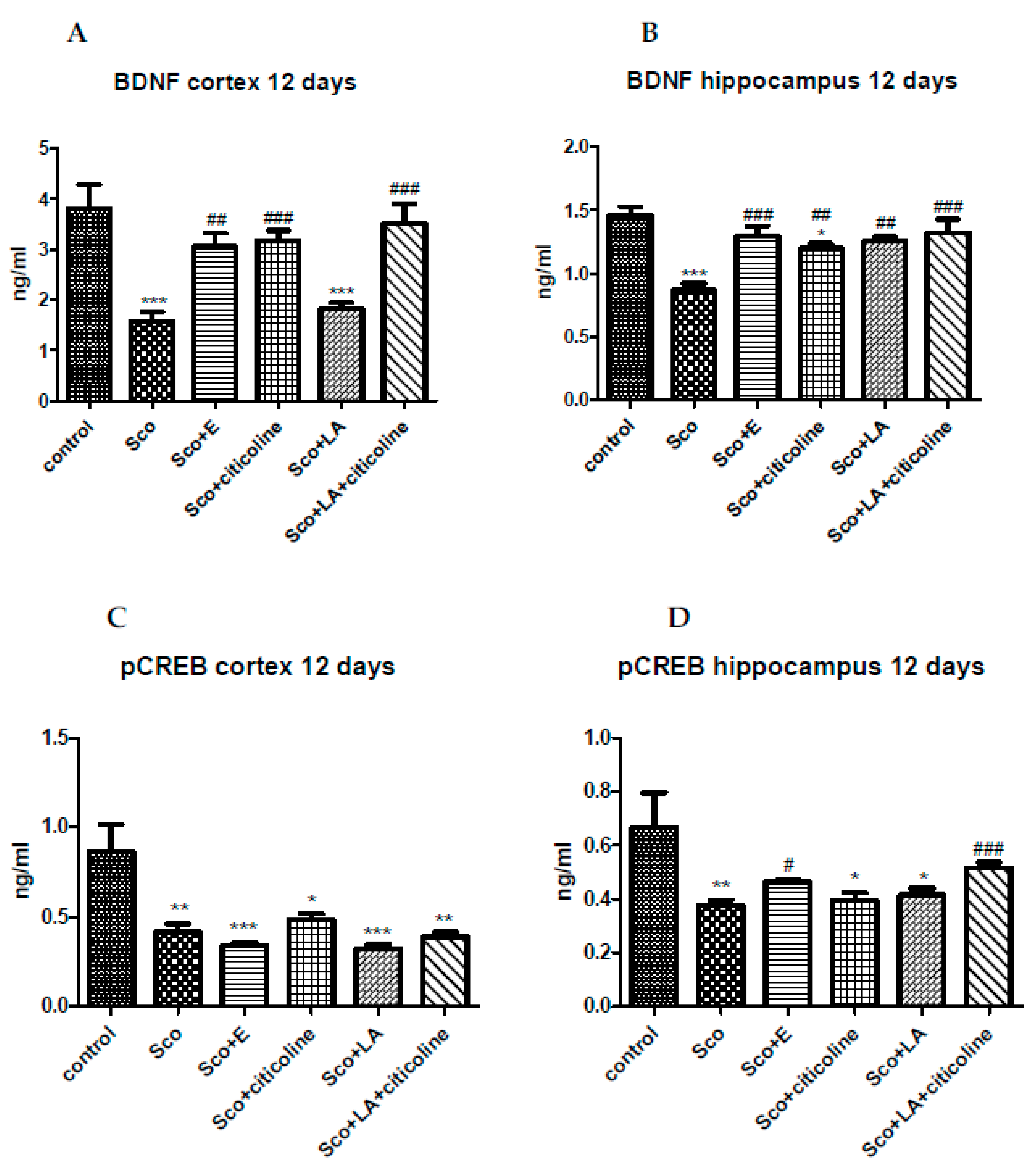
Figure 9.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on ASAT and ALAT activity and creatinine, albumin, globulin and total protein levels of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
Figure 9.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on ASAT and ALAT activity and creatinine, albumin, globulin and total protein levels of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
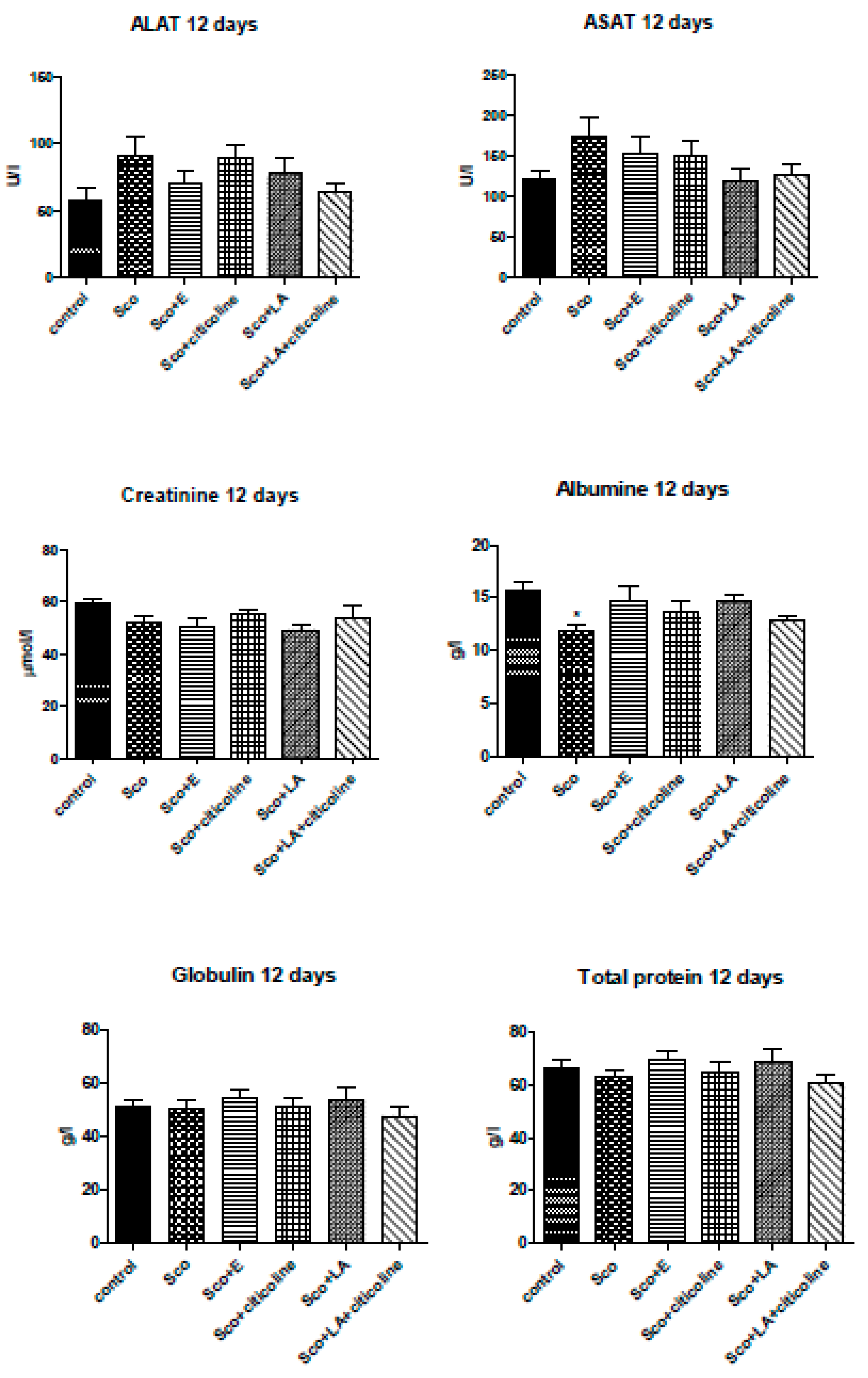
Figure 10.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on CRP levels of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
Figure 10.
Effects of the experimental combination and its components, citicoline, alpha-lipoic acid (LA) and alpha-lipoic acid and citicoline (LA+citicoline) on CRP levels of scopolamine-treated rats. Data are expressed as mean±SEM; significant differences among experimental groups are denoted with “*” whenever groups are compared with the saline (control) group and denoted with “#” whenever groups are compared with the dementia (Sco) group. The number of “∗” or “#” above bars is assigned as follows: # or ∗ at p < 0.05; ## or ∗∗ at p < 0.01; ### or ∗∗∗ at p < 0.001 and #### or ∗∗∗∗ at p < 0.0001.
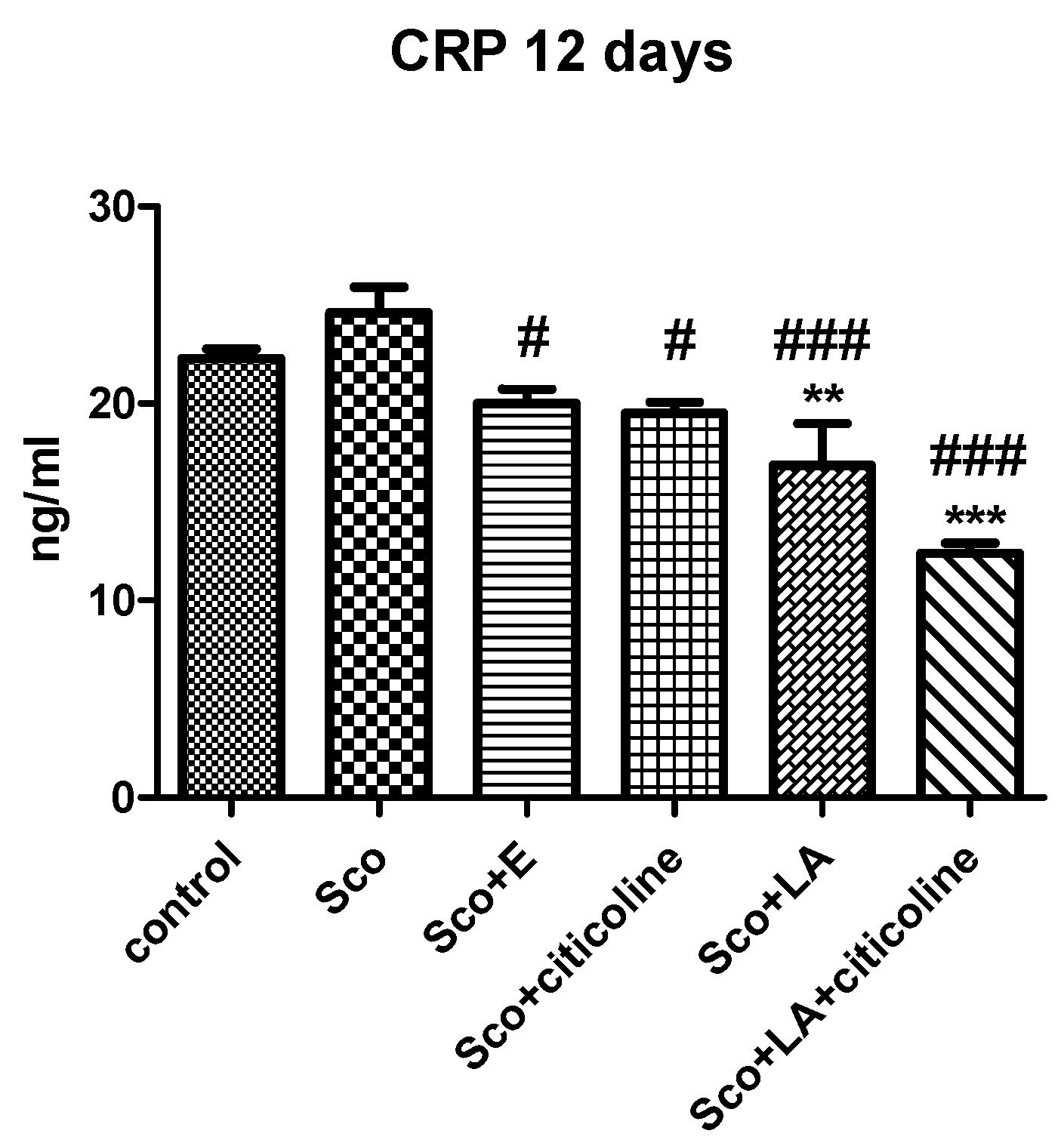
Figure 11.
Analysis of the first two principal components - PC 1 and PC 2, representing a linear combination of the values of the histopathological and behavioral indicators for each experimental group.
Figure 11.
Analysis of the first two principal components - PC 1 and PC 2, representing a linear combination of the values of the histopathological and behavioral indicators for each experimental group.
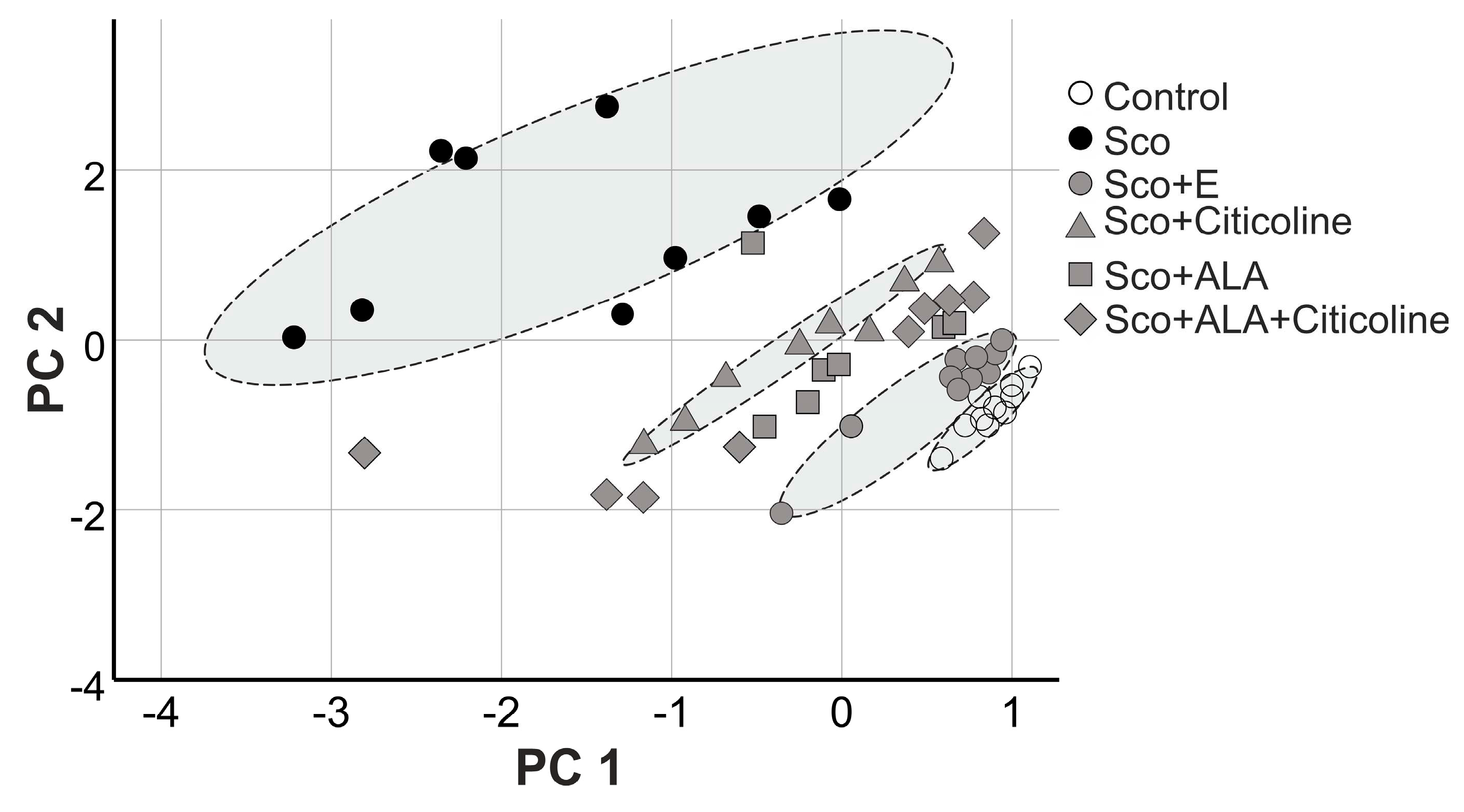
Figure 12.
Correlation between the coefficients of pathological damage and the indicator of recovery in the different experimental groups.
Figure 12.
Correlation between the coefficients of pathological damage and the indicator of recovery in the different experimental groups.
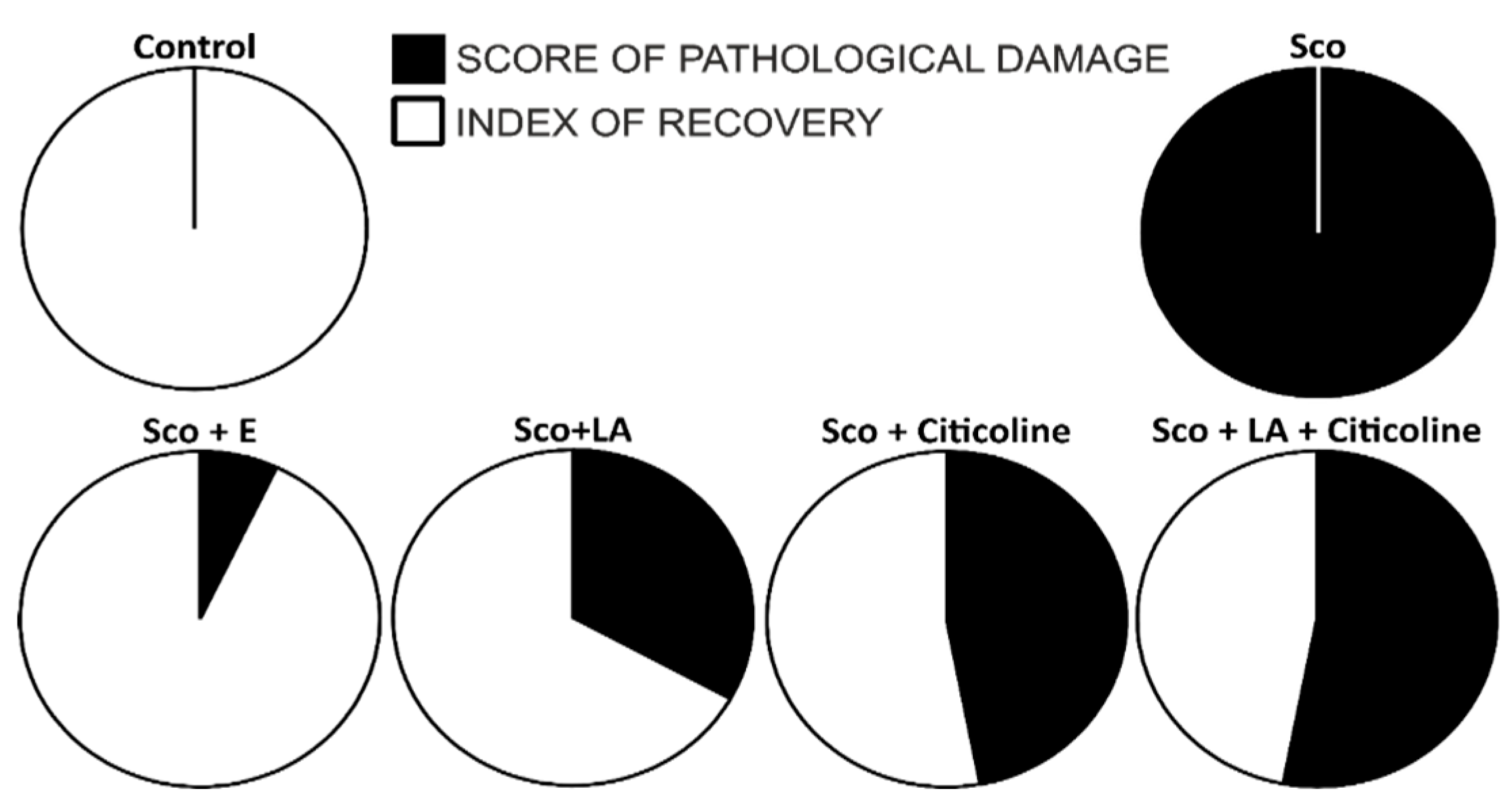
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



