
Submitted:
03 November 2023
Posted:
03 November 2023
You are already at the latest version
Abstract
CD44 is a type I transmembrane glycoprotein, and possesses various isoforms which are largely classified into CD44 standard and CD44 variant (CD44v) isoforms. Some variant-encoded regions play critical roles in tumor progression. However, the function of CD44 variant 4 (CD44v4)-encoded region has not been fully understood. Using peptide immunization, we developed an anti-CD44v4 mAb, C44Mab-108, which is useful for flow cytometry, western blotting, and immunohistochemistry. In this study, we determined the critical epitope of C44Mab-108 by enzyme-linked immunosorbent assay (ELISA). We used the alanine (or glycine)-substituted peptides of the CD44v4-encoded region (amino acids 271-290 of human CD44v3-10), and found that C44Mab-108 did not recognize the alanine-substituted peptides of D280A and W281A. Furthermore, these peptides could not inhibit the recognition of C44Mab-108 in flow cytometry and immunohistochemistry. The results indicate that the critical binding epitope of C44Mab-108 includes Asp280 and Trp281 of CD44v3-10.
Keywords:
CD44
; CD44 variant 4
; monoclonal antibody
; epitope
; enzyme-linked immunosorbent assay
1. Introduction
CD44 has various isoforms, which are generated by the alternative splicing of CD44 pre-mRNA [1]. The mRNA of CD44 standard (CD44s) isoform is produced by constant region exons, including the first five (1 to 5) and the last five (16 to 20) [2]. The mRNAs of CD44 variant (CD44v) isoform are produced by the assembling of variant exons (v1–v10) with the constant region exons of CD44s [3]. CD44s and CD44v receive the post-translational modifications, including N- or O-glycosylation [4]. Both CD44s and CD44v can attach to hyaluronic acid, which is important for cellular adhesion, homing, and motility [5].
CD44v plays important roles in the tumor progression by specific functions of variant exon-encoded regions [6]. The heparin-binding growth factors are recruited to heparan sulphate modified in the v3-encoded region [7,8]. MET, a receptor tyrosine kinase for hepatocyte growth factor, associates with the v6-encoded region [9,10]. These functions are essential for the activation of growth factor signaling and tumor proliferation. However, the roles of CD44 variant 4 (CD44v4)-encoded region have not been investigated. Therefore, specific antibodies against CD44v4 are indispensable for basic research, tumor diagnosis, and therapy.
We have established anti-CD44 monoclonal antibodies (mAbs), which recognize the standard [11,12]- or each variant [13,14,15,16,17,18,19,20]-encoded region. All mAbs can be used for flow cytometry, western blotting, and immunohistochemistry, and are expected to contribute not only to basic research but also to pathogenic diagnosis. We also determined the critical epitopes of C44Mab-5 [21] and C44Mab-46 [22,23]. We previously established an anti-CD44v4 mAb, C44Mab-108 (mouse IgG1, kappa) using the peptide immunization [19]. To clarify further characteristics of C44Mab-108, we performed epitope mapping using enzyme-linked immunosorbent assay (ELISA).
2. Materials and Methods
2.1. Peptides
The CD44v4 peptide (271-AFDHTKQNQDWTQWNPSHSN-290) and 20 alanine (or glycine)-substituted peptides (Table 1) were synthesized by utilizing PEPScreen (Sigma-Aldrich Corp., St. Louis, MO). The number of amino acids (aa) is derived from human CD44v3-10 (accession No.: X66733).
2.2. ELISA
The CD44v4 peptides were immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher Scientific Inc., Waltham, MA) at a concentration of 10 µg/mL for 30 min at 37℃. After washing with phosphate-buffered saline (PBS) containing 0.05% Tween20 (PBST; Nacalai Tesque, Inc., Kyoto, Japan), wells were blocked with 1% bovine serum albumin (BSA)-containing PBST for 30 min at 37℃. The plates were incubated with 10 µg/mL of C44Mab-108, followed by a peroxidase-conjugated anti-mouse immunoglobulins (1:2000 diluted; Agilent Technologies Inc., Santa Clara, CA). Enzymatic reactions were performed using the ELISA POD Substrate TMB Kit (Nacalai Tesque, Inc.). Optical density was measured at 655 nm using an iMark microplate reader (Bio-Rad Laboratories, Inc., Berkeley, CA).
2.3. Flow cytometriy
The N-terminal PA16-tagged CD44v3-10-overexpressed Chinese hamster ovary-K1 (CHO/CD44v3-10) [19] cells were harvested after a brief exposure to 0.25% trypsin in 1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.) and washed with 0.1% BSA in PBS. C44Mab-108 (10 μg/mL) was incubated with the CD44v4 peptides (10 μg/ml) for 30 min at 4°C. The cells were further treated with Alexa Fluor 488-conjugated anti-mouse IgG (1:2000). Fluorescence data were collected using the SA3800 Cell Analyzer (Sony Corp., Tokyo, Japan).
2.4. Immunohistochemical analysis
One formalin-fixed paraffin-embedded (FFPE) tissue of oral squamous cell carcinoma (OSCC) for peptide blocking assay was obtained from Tokyo Medical and Dental University [24]. The tissue slides were autoclaved in citrate buffer (pH 6.0; Nichirei Biosciences, Inc., Tokyo, Japan) for 20 min for antigen retrieval. After blocking with SuperBlock T20 (Thermo Fisher Scientific, Inc.), the sections were incubated with C44Mab-108 (10 μg/mL) in the presence or absence of the CD44v4 peptides (10 μg/mL) and then treated with the EnVision+ Kit for mouse (Agilent Technologies Inc.) for 30 min. The color was developed using 3,3′-diaminobenzidine tetrahydrochloride (DAB; Agilent Technologies Inc.). Counterstaining was performed with hematoxylin (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). Leica DMD108 (Leica Microsystems GmbH, Wetzlar, Germany) was used to examine the sections and obtain images.
3. Results
3.1. Epitope mapping of C44Mab-108 with alanine (or glycine)-substituted CD44v4 peptide
We previously established an anti-CD44v4 mAb (C44Mab-108) by peptide immunization of CD44v4 region (273-DHTKQNQDWTQWNPSHSNP-291) [19]. We confirmed that C44Mab-108 recognizes with only the variant 4-encoded region peptide (aa 271-290), but not other regions of CD44v3-10 extracellular domain [19]. To identify the binding epitope of C44Mab-108, we synthesized 20 alanine (or glycine)-substituted peptides of the CD44v4 (Table 1). C44Mab-108 exhibited reaction with A271G, F272A, D273A, H274A, T275A, K276A, Q277A, N278A, Q279A, T282A, Q283A, W284A, N285A, P286A, S287A, H288A, S289A, N290A, and wild-type (WT) (Figure 1A). In contrast, C44Mab-108 did not react with D280A and W281A (Figure 1A). This result indicated that Asp280 and Trp281 are included in the critical epitope of C44Mab-108. The results are summarized in Table 1. Figure 1B shows the schematic illustration of CD44s, CD44v3-10, and the critical aa (Asp280 and Trp281) recognized by C44Mab-108.
3.2. Flow cytometry using C44Mab-108 with alanine-substituted CD44v4 peptides
We next performed a peptide-blocking assay using flow cytometry to confirm the importance of the C44Mab-108 epitope. As shown in Figure 2, C44Mab-108 reacted with the CHO/CD44v3-10 cells. This reaction was completely neutralized by WT. In contrast, D280A and W281A did not block the reaction of C44Mab-108 with CHO/CD44v3-10. The result confirmed that Asp280 and Trp281 of CD44v3-10 are critical for detection by C44Mab-108 using flow cytometry.
3.3. Immunohistochemistry using C44Mab-108 with alanine-substituted CD44v4 peptides
We also performed a peptide-blocking assay using immunohistochemical analysis. As shown in Figure 3, C44Mab-108 stained the FFPE section of OSCC, which was completely neutralized by WT. In contrast, D280A and W281A did not neutralize the reaction. These results were corresponding to that of Figure 2.
4. Discussion
In the present study, the critical epitope of C44Mab-108 was determined as Asp280 and Trp281 in the CD44v4 region. Since the reactivity of C44Mab-108 to T282A and W284A was also reduced (Figure 1A), Thr282 and Trp284 may contribute to the recognition partially. Figure 1B shows the homology of the v4 region among human, mouse, Chinese hamster, and rat sequences. The Asp280 and Trp281 are conserved in human, mouse, Chinese hamster, but not in rat. Furthermore, Thr282 and Trp284 are also conserved in human and mouse. Although the result suggests that C44Mab-108 might recognize both human and mouse CD44v4, it did not react with mouse CD44v4 using flow cytometry (data not shown).
Since C44Mab-108 was established by the peptide immunization, C44Mab-108 can recognize the definite peptide structure of the variant 4-encoded region. In contrast, CD44 is predicted to carry 146 O-glycan sites in the variant region. Among them, 41 of these sites have already been experimentally confirmed [25]. Thr282 is also a confirmed O-glycan site [26]. Further studies are required to reveal whether the O-glycan at Thr282 affects the recognition by C44Mab-108.
In our previous study, C44Mab-108 could detect CD44v3-10-overexpressed cells such as CHO/CD44v3-10, but not detect endogenous CD44v4 in several cancer cell lines in flow cytometry [19]. In contrast, C44Mab-108 could detect endogenous and membranous CD44v4 in immunohistochemistry [19]. These results suggest that the variant 4-encoded region is folded into the inside of protein in living cells, but exposed by antigen retrieval in immunohistochemistry. Recently, we provided a potential strategy for developing cancer-specific antibodies that target locally misfolded cell surface receptors such as human epidermal growth factor receptor 2 [27,28]. Further studies are needed to investigate whether C44Mab-108 is involved in the recognition of specific CD44v4 type and/or specific condition of cells.
Funding
This research was supported in part by Japan Agency for Medical Research and Development (AMED) under Grant Numbers: JP23ama121008 (to Y.K.), JP23am0401013 (to Y.K.), 23bm1123027h0001 (to Y.K.), and JP23ck0106730 (to Y.K.), and by the Japan Society for the Promotion of Science (JSPS) Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 22K06995 (to H.S.), 21K07168 (to M.K.K.), and 22K07224 (to Y.K.).
Conflicts of Interest
The authors have no conflict of interest.
References
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: from adhesion molecules to signalling regulators. Nat Rev Mol Cell Biol 2003, 4, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Zuo, X.; Wei, D. Concise Review: Emerging Role of CD44 in Cancer Stem Cells: A Promising Biomarker and Therapeutic Target. Stem Cells Transl Med 2015, 4, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: therapeutic implications. J Hematol Oncol 2018, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Wang, Q.; An, J.; Chen, J.; Li, X.; Long, Q.; Xiao, L.; Guan, X.; Liu, J. CD44 Glycosylation as a Therapeutic Target in Oncology. Front Oncol 2022, 12, 883831. [Google Scholar] [CrossRef] [PubMed]
- Mishra, M.N.; Chandavarkar, V.; Sharma, R.; Bhargava, D. Structure, function and role of CD44 in neoplasia. J Oral Maxillofac Pathol 2019, 23, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Zöller, M. CD44: can a cancer-initiating cell profit from an abundantly expressed molecule? Nat Rev Cancer 2011, 11, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.L.; Jackson, D.G.; Simon, J.C.; Tanczos, E.; Peach, R.; Modrell, B.; Stamenkovic, I.; Plowman, G.; Aruffo, A. CD44 isoforms containing exon V3 are responsible for the presentation of heparin-binding growth factor. J Cell Biol 1995, 128, 687–698. [Google Scholar] [CrossRef]
- Jackson, D.G.; Bell, J.I.; Dickinson, R.; Timans, J.; Shields, J.; Whittle, N. Proteoglycan forms of the lymphocyte homing receptor CD44 are alternatively spliced variants containing the v3 exon. J Cell Biol 1995, 128, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Matzke, A.; Sargsyan, V.; Holtmann, B.; Aramuni, G.; Asan, E.; Sendtner, M.; Pace, G.; Howells, N.; Zhang, W.; Ponta, H.; et al. Haploinsufficiency of c-Met in cd44-/- mice identifies a collaboration of CD44 and c-Met in vivo. Mol Cell Biol 2007, 27, 8797–8806. [Google Scholar] [CrossRef] [PubMed]
- Orian-Rousseau, V.; Chen, L.; Sleeman, J.P.; Herrlich, P.; Ponta, H. CD44 is required for two consecutive steps in HGF/c-Met signaling. Genes Dev 2002, 16, 3074–3086. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Itai, S.; Nakamura, T.; Yanaka, M.; Kaneko, M.K.; Kato, Y. Detection of high CD44 expression in oral cancers using the novel monoclonal antibody, C(44)Mab-5. Biochem Biophys Rep 2018, 14, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Goto, N.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Monoclonal Antibody for Multiple Applications against Esophageal Squamous Cell Carcinomas. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Ejima, R.; Suzuki, H.; Tanaka, T.; Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 6 Monoclonal Antibody C(44)Mab-9 for Multiple Applications against Colorectal Carcinomas. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Establishment of a Novel Anti-CD44 Variant 10 Monoclonal Antibody C(44)Mab-18 for Immunohistochemical Analysis against Oral Squamous Cell Carcinomas. Curr Issues Mol Biol 2023, 45, 5248–5262. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Suzuki, H.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 5 Monoclonal Antibody C(44)Mab-3 for Multiple Applications against Pancreatic Carcinomas. Antibodies (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Suzuki, H.; Goto, N.; Tanaka, T.; Ouchida, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 8 Monoclonal Antibody C(44)Mab-94 against Gastric Carcinomas. Antibodies (Basel) 2023, 12. [Google Scholar] [CrossRef]
- Suzuki, H.; Kitamura, K.; Goto, N.; Ishikawa, K.; Ouchida, T.; Tanaka, T.; Kaneko, M.K.; Kato, Y. A Novel Anti-CD44 Variant 3 Monoclonal Antibody C(44)Mab-6 Was Established for Multiple Applications. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ozawa, K.; Tanaka, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 7/8 Monoclonal Antibody, C(44)Mab-34, for Multiple Applications against Oral Carcinomas. Biomedicines 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Tanaka, T.; Goto, N.; Kaneko, M.K.; Kato, Y. Development of a Novel Anti-CD44 Variant 4 Monoclonal Antibody C(44)Mab-108 for Immunohistochemistry. Curr Issues Mol Biol 2023, 45, 1875–1888. [Google Scholar] [CrossRef] [PubMed]
- Tawara, M.; Suzuki, H.; Goto, N.; Tanaka, T.; Kaneko, M.K.; Kato, Y. A Novel Anti-CD44 Variant 9 Monoclonal Antibody C(44)Mab-1 Was Developed for Immunohistochemical Analyses against Colorectal Cancers. Curr Issues Mol Biol 2023, 45, 3658–3673. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Kaneko, M.K.; Kato, Y. Development of a Novel Epitope Mapping System: RIEDL Insertion for Epitope Mapping Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Kaneko, M.K.; Takei, J.; Tateyama, N.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C(44)Mab-46) Using the REMAP Method. Monoclon Antib Immunodiagn Immunother 2021, 40, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Takei, J.; Asano, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of the Anti-CD44 Monoclonal Antibody (C(44)Mab-46) Using Alanine-Scanning Mutagenesis and Surface Plasmon Resonance. Monoclon Antib Immunodiagn Immunother 2021, 40, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Itai, S.; Ohishi, T.; Kaneko, M.K.; Yamada, S.; Abe, S.; Nakamura, T.; Yanaka, M.; Chang, Y.W.; Ohba, S.I.; Nishioka, Y.; et al. Anti-podocalyxin antibody exerts antitumor effects via antibody-dependent cellular cytotoxicity in mouse xenograft models of oral squamous cell carcinoma. Oncotarget 2018, 9, 22480–22497. [Google Scholar] [CrossRef] [PubMed]
- Steentoft, C.; Vakhrushev, S.Y.; Joshi, H.J.; Kong, Y.; Vester-Christensen, M.B.; Schjoldager, K.T.; Lavrsen, K.; Dabelsteen, S.; Pedersen, N.B.; Marcos-Silva, L.; et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. Embo j 2013, 32, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Mereiter, S.; Martins Á, M.; Gomes, C.; Balmaña, M.; Macedo, J.A.; Polom, K.; Roviello, F.; Magalhães, A.; Reis, C.A. O-glycan truncation enhances cancer-related functions of CD44 in gastric cancer. FEBS Lett 2019, 593, 1675–1689. [Google Scholar] [CrossRef] [PubMed]
- Arimori, T.; Mihara, E.; Suzuki, H.; Ohishi, T.; Tanaka, T.; Kaneko, M.K.; Takagi, J.; Kato, Y. Locally misfolded HER2 expressed on cancer cells is a promising target for development of cancer-specific antibodies Cell Press Community Review 2023. [CrossRef]
- Suzuki, H.; Kaneko, M.K.; Kato, Y. A cancer-specific monoclonal antibody against HER2 for breast cancers Preprint 2023. [CrossRef]
Figure 1.
Determination of the C44Mab-108 epitope by ELISA using alanine (or glycine)-substituted CD44v4 peptides. (A) The alanine (or glycine)-substituted CD44v4 peptides in PBS or PBS alone were immobilized on immunoplates. The plates were incubated with C44Mab-108 (10 µg/mL), followed by peroxidase-conjugated anti-mouse immunoglobulins. Error bars represent means ± SDs. N.C., negative control (PBS). (B) Schematic illustration of CD44s, CD44v3-10, and the C44Mab-108 epitope. The C44Mab-108 epitope includes Asp280 and Trp281 of CD44v3-10. The epitope is conserved in mouse and Chinese hamster (Cham), but not in rat.
Figure 1.
Determination of the C44Mab-108 epitope by ELISA using alanine (or glycine)-substituted CD44v4 peptides. (A) The alanine (or glycine)-substituted CD44v4 peptides in PBS or PBS alone were immobilized on immunoplates. The plates were incubated with C44Mab-108 (10 µg/mL), followed by peroxidase-conjugated anti-mouse immunoglobulins. Error bars represent means ± SDs. N.C., negative control (PBS). (B) Schematic illustration of CD44s, CD44v3-10, and the C44Mab-108 epitope. The C44Mab-108 epitope includes Asp280 and Trp281 of CD44v3-10. The epitope is conserved in mouse and Chinese hamster (Cham), but not in rat.
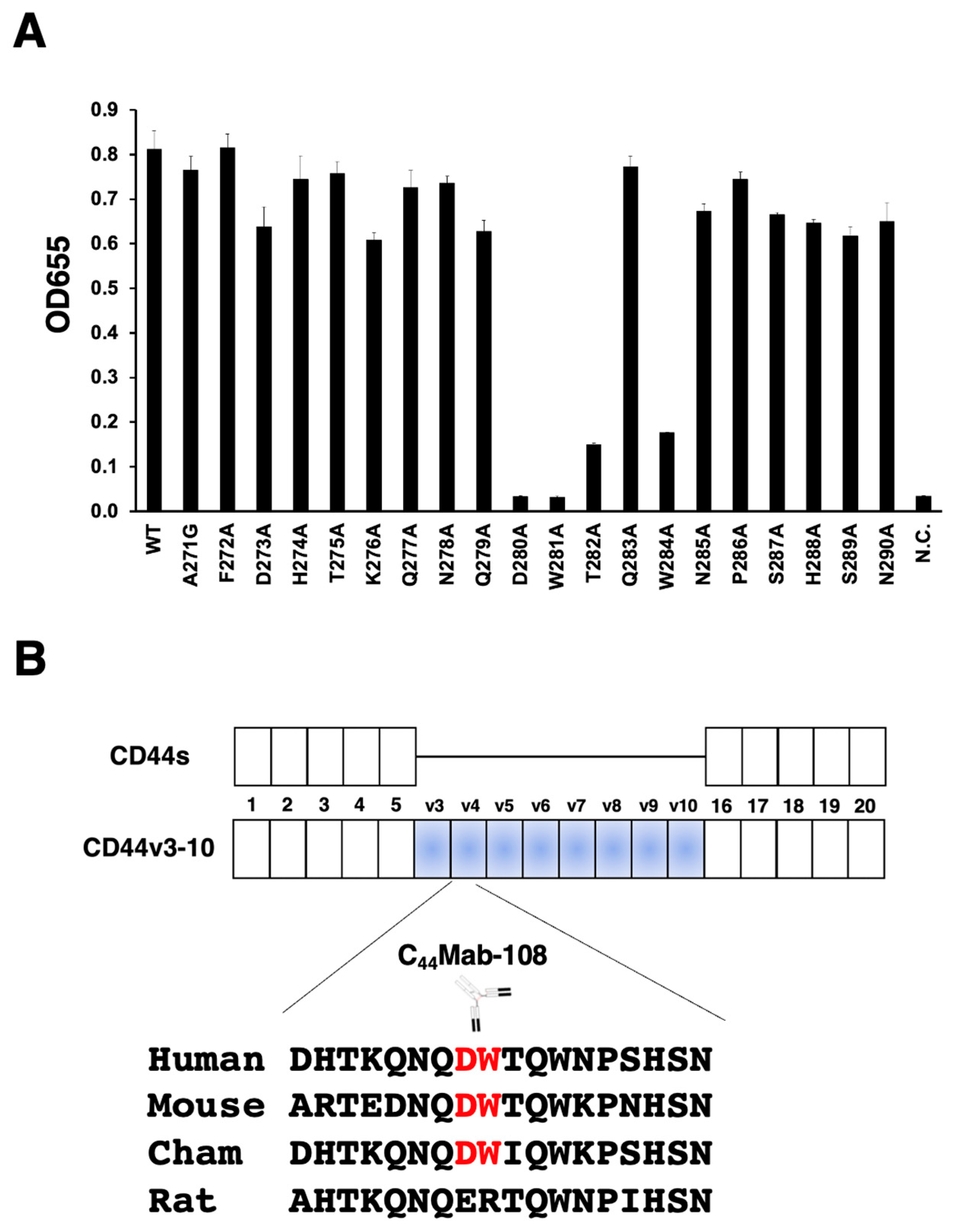
Figure 2.
Flow cytometry using C44Mab-108 and CD44v4 peptides. C44Mab-108 (10 µg/mL) plus WT, the alanine-substituted peptides (10 μg/ml), or control (0.1% DMSO in blocking buffer, –peptide) were reacted with CHO/CD44v3-10 cells for 30 min at 4°C, followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. The black line represents the control. WT, wild type; DMSO, dimethyl sulfoxide.
Figure 2.
Flow cytometry using C44Mab-108 and CD44v4 peptides. C44Mab-108 (10 µg/mL) plus WT, the alanine-substituted peptides (10 μg/ml), or control (0.1% DMSO in blocking buffer, –peptide) were reacted with CHO/CD44v3-10 cells for 30 min at 4°C, followed by treatment with Alexa Fluor 488-conjugated anti-mouse IgG. The black line represents the control. WT, wild type; DMSO, dimethyl sulfoxide.
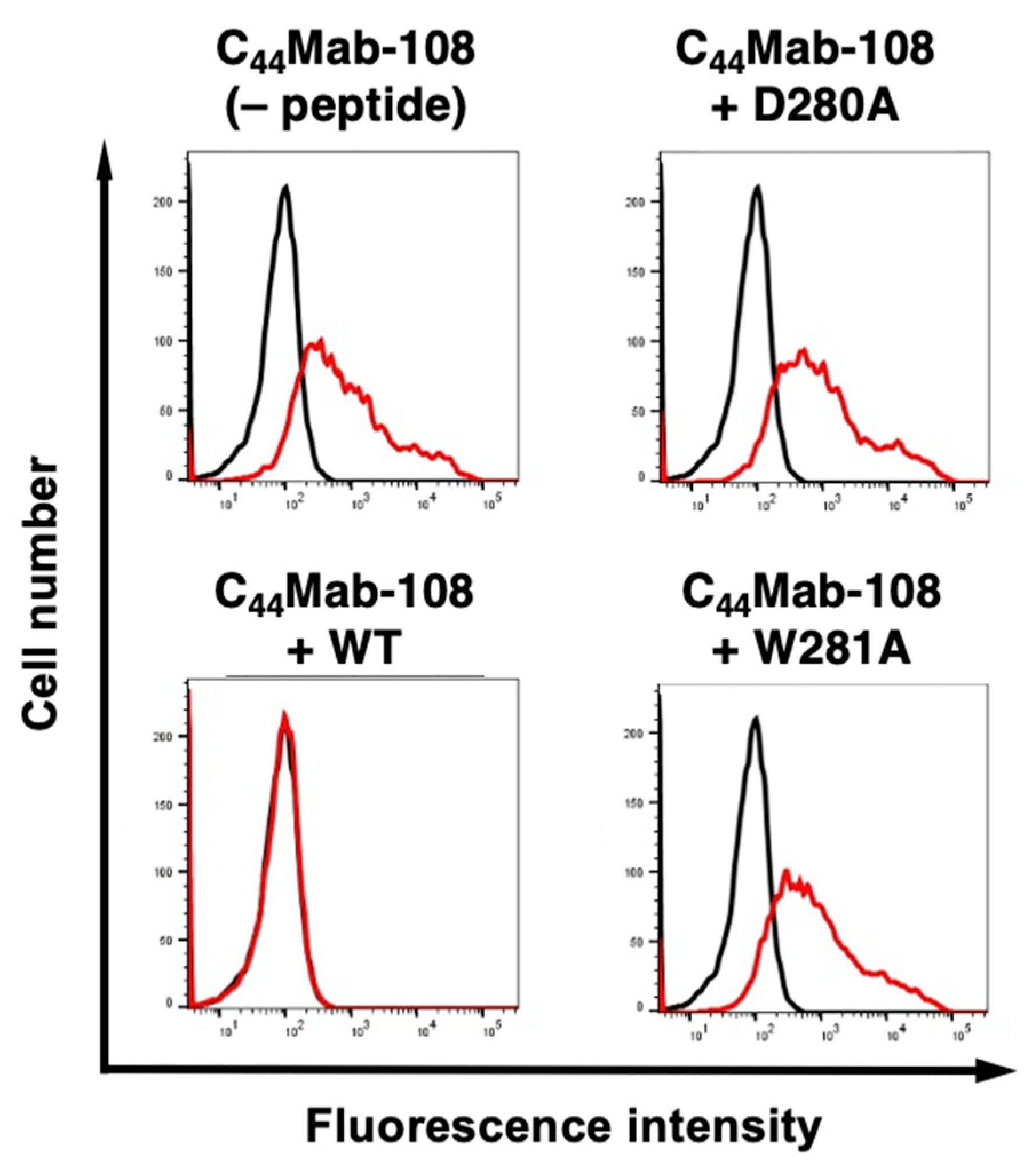
Figure 3.
Immunohistochemistry using C44Mab-108 and CD44v4 peptides. The FFPE sections of OSCC were incubated with C44Mab-108 (10 µg/mL) plus WT, the alanine-substituted peptides (10 μg/ml), or control (0.1% DMSO in blocking buffer, –peptide), followed by that with the Envision + Kit. Scale bar = 100 μm. FFPE, formalin-fixed paraffin-embedded; OSCC, oral squamous cell carcinoma; WT, wild type; DMSO, dimethyl sulfoxide.
Figure 3.
Immunohistochemistry using C44Mab-108 and CD44v4 peptides. The FFPE sections of OSCC were incubated with C44Mab-108 (10 µg/mL) plus WT, the alanine-substituted peptides (10 μg/ml), or control (0.1% DMSO in blocking buffer, –peptide), followed by that with the Envision + Kit. Scale bar = 100 μm. FFPE, formalin-fixed paraffin-embedded; OSCC, oral squamous cell carcinoma; WT, wild type; DMSO, dimethyl sulfoxide.
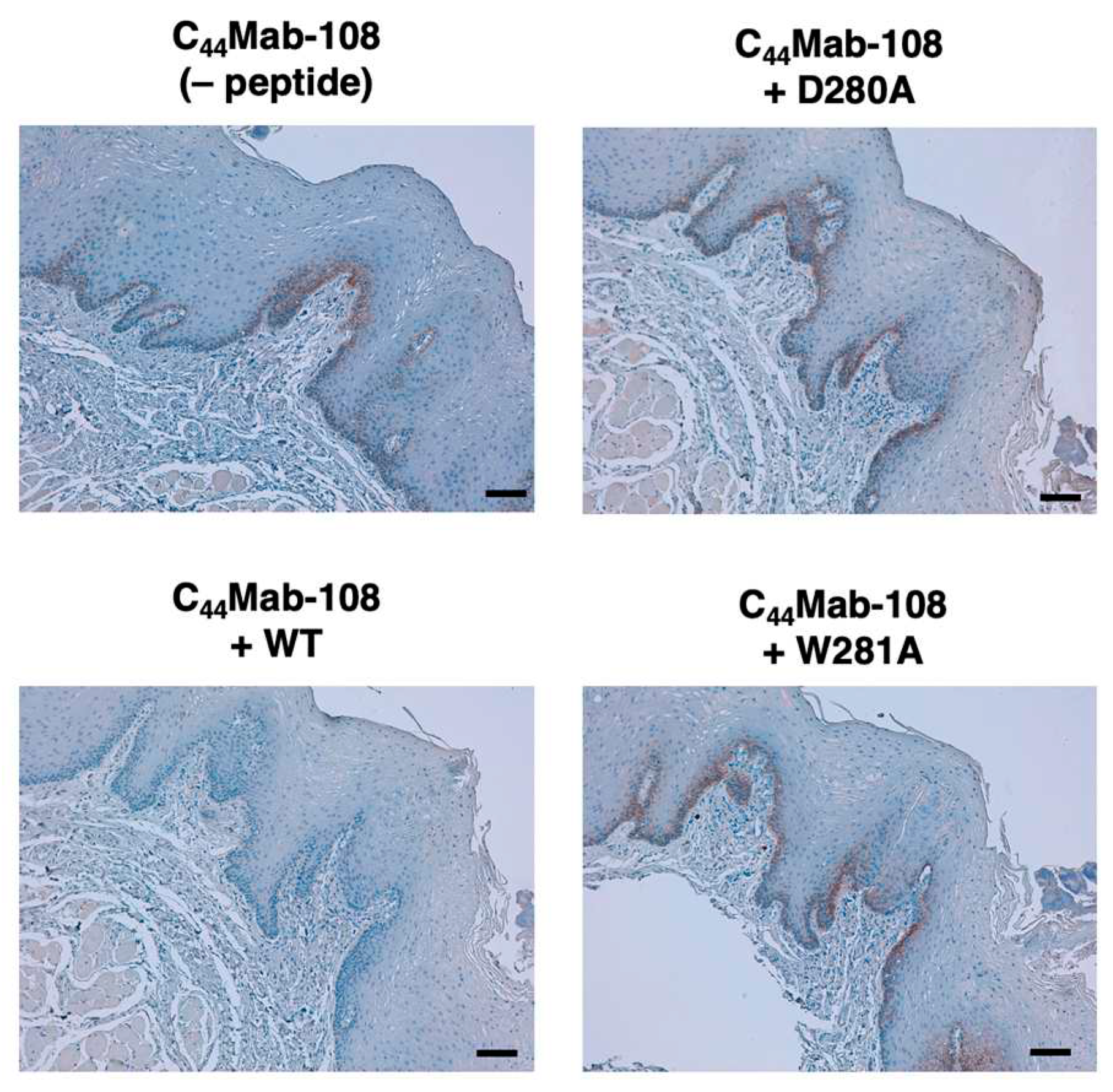

Table 1.
Identification of the C44Mab-108 epitope using alanine (or glycine)-substituted CD44v4 peptides.
Table 1.
Identification of the C44Mab-108 epitope using alanine (or glycine)-substituted CD44v4 peptides.
| Peptides | Sequences | C44Mab-108 |
| WT (271-290) | AFDHTKQNQDWTQWNPSHSN | +++ |
| A271G | GFDHTKQNQDWTQWNPSHSN | +++ |
| F272A | AADHTKQNQDWTQWNPSHSN | +++ |
| D273A | AFAHTKQNQDWTQWNPSHSN | +++ |
| H274A | AFDATKQNQDWTQWNPSHSN | +++ |
| T275A | AFDHAKQNQDWTQWNPSHSN | +++ |
| K276A | AFDHTAQNQDWTQWNPSHSN | +++ |
| Q277A | AFDHTKANQDWTQWNPSHSN | +++ |
| N278A | AFDHTKQAQDWTQWNPSHSN | +++ |
| Q279A | AFDHTKQNADWTQWNPSHSN | +++ |
| D280A | AFDHTKQNQAWTQWNPSHSN | – |
| W281A | AFDHTKQNQDATQWNPSHSN | – |
| T282A | AFDHTKQNQDWAQWNPSHSN | + |
| Q283A | AFDHTKQNQDWTAWNPSHSN | +++ |
| W284A | AFDHTKQNQDWTQANPSHSN | + |
| N285A | AFDHTKQNQDWTQWAPSHSN | +++ |
| P286A | AFDHTKQNQDWTQWNASHSN | +++ |
| S287A | AFDHTKQNQDWTQWNPAHSN | +++ |
| H288A | AFDHTKQNQDWTQWNPSASN | +++ |
| S289A | AFDHTKQNQDWTQWNPSHAN | +++ |
| N290A | AFDHTKQNQDWTQWNPSHSA | +++ |
+++, OD655≧0.6; ++, 0.3≦OD655<0.6; +, 0.1≦OD655<0.3; –, OD655<0.1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



