
Submitted:
01 November 2023
Posted:
03 November 2023
You are already at the latest version
Abstract
Rapid analysis and characterization of compounds by mass spectrometry may overlook trace compounds, although targeted analysis methods can significantly improve detection sensitivity, it is difficult to discover novel scaffold compounds. This study developed a strategy for discovering trace compounds in the aging process of traditional Chinese medicine based on MS fragmentation and known metabolic pathway. Specifically, we found that the characteristic component of C. reticulata 'Chachi', methyl N-methylanthranilate (MMA), fragmented in ESI-CID to produce rearrangement ion 3-hydroxyindole, which has been proven to exist in trace amounts in C. reticulata 'Chachi' comparing with the reference substance by LC-MS/MS. By combining the known metabolic pathways of 3-hydroxyindole and the possible methylation reactions that may occur during aging, 10 possible indole derivatives were untargeted predicted. These compounds were confirmed to originate from MMA by purchasing or synthesizing reference substances and all of them were detected in C. reticulata 'Chachi' through LC-MS/MS analysis, achieving trace compound analysis from untargeted to targeted. It may contribute to explain the aging mechanism of C. reticulata 'Chachi' and the strategy of CID-induced special rearrangement ion binding metabolic pathway has potential application value in discovering trace compounds.
Keywords:
Citrus reticulata 'Chachi'
; methyl N-methylanthranilate
; mass spectrometry
; 3-hydroxyindole
; indole derivatives
1. Introduction
In recent years, LC-MS/MS have played an extremely important role in pharmaceutical [1,2] and foods [3,4,5] analysis. Untargeted metabolomics [6,7,8] based on LC-MS is a widely unbiased method for detecting metabolites in biological samples [9]. By screening differential metabolites and analyzing their metabolic pathways, potential biomarkers are identified. However, data processing is complex, sensitivity is limited, and some low abundance compounds cannot be detected, limiting its application in discovering new biomarkers and understanding disease mechanisms. Targeted metabolomics [6,10,11] uses reference substances to detect specific metabolites that have already been noticed, which can effectively improve sensitivity and more accurately elucidate the relationship between metabolites and diseases. The analysis strategy [12] by integrating the univariate and multivariate correlation analysis approach was developed to discover tumor tissue derived (TTD) metabolites in plasma samples, improving the accuracy of untargeted metabolomics screening for biomarkers. A novel integrated strategy [13] that employs chemical derivatization, liquid chromatography–mass spectrometry, and molecular networking (MN) technique to enable the rapid recognition and identification of carboxylated gut microbiota–host co-metabolites with high sensitivity and selectivity, which 261 carboxylated metabolites from mouse feces were detected. Comprehensive two-dimensional gas chromatography/time-of-flight mass spectrometry (GC×GC/TOFMS) were applied for the characterization of the acidic fraction and the complex hydrocarbons in the cigarette smoke condensates. The method can be more widely applied to the analysis of compounds present in mixtures with comparable complexity [14,15]. Targeted data-dependent acquisition (DDA) based on the inclusion list of differential and preidentified ions (dpDDA) was developed to improve the data quality of untargeted metabolomics [16]. The transition from untargeted to targeted analysis strategies can significantly increase the sensitivity of detection, but it is difficult to discover novel scaffold compounds.
We have previously established a method [17] to simulate and predict the degradation products and metabolites of special drugs based on electrospray ionization-collision-induced dissociation-mass spectrometry (ESI-CID-MS) The acidic microdroplets [18] formed in ESI can accelerate degradation [19] or metabolism of pharmaceuticals, specifically, when the analyte is ionized and sprayed, it is charged and activated to become an excited species, the excited ions enter the subsequent triple quadrupole and collide with Ar in the collision cell to undergo non-specific free radical or rearrangement reactions, thereby enabling an accelerated process in degradation and metabolism of drugs. Two low-abundance rearrangement ions, (aza) carbazole and (aza) biphenylene were successfully identified by ESI-CID-MS of a series of commercially available kinase inhibitor drugs containing diphenylene-like structures. Therefore, CID-induced rearrangement ions were crucial to discover low concentration compounds.
Citri Reticulatae Pericarpium (CRP) is the dried pericarp derived from mature Citrus reticulata Blanco and its cultivars, which mainly include the C. reticulata 'Chachi' of Guangdong, the C. reticulata 'Dahongpao' of Sichuan, the C. reticulata 'Tangerina' of Fujian and the C. reticulata 'Unshiu' of Zhejiang [20,21]. Among them, C. reticulata 'Chachi' is also known as 'Xinhuichenpi' produced in Xinhui District, Guangdong, is considered geoherbs due to its superior quality, which contains bioactive substances such as flavonoids [22,23], alkaloids [24], volatile oils [25], polysaccharides [26] along with greatest antioxidant and anti-inflammatory activities [22,27]. In addition, C. reticulata 'Chachi' is applied in both clinical and food accessories at the same time which has a long history as a traditional spice and seasoning among the people. C. reticulata 'Chachi' needs to be stored for at least 3 to 10 years or more, and the longer the aging time, the darker the color and the better the quality. The process of pericarp turning from yellow to brown or even black was attributed to the Maillard reaction in a number of studies [22]. During the aging of CRP, the substrates of this reaction, namely amino acids, saccharides have a decreasing trend, and the intermediate 5-hydroxymethylfurfural is forming. In the final stage of Maillard reaction, various active intermediates conduct condensation and other reactions with amino acids, and then form brown nitrogen-containing polymers or copolymer melanin, which eventually leads to color browning [28]. The investigation showed that the increase of nitrogenous compounds in the C. reticulata 'Chachi' preserved for 10 years was related to the Maillard browning product [29] but there is relatively little research on which trace amounts of nitrogen-containing compounds are generated during the aging process.
MMA, a characteristic component of C. reticulata 'Chachi', is an important basis for identifying the authenticity of C. reticulata 'Chachi' [30]. As mentioned earlier, the aging and browning of C. reticulata 'Chachi' are related to an increase in nitrogen-containing compounds, while current research on MMA mainly focuses on changes in its content during the storage [31]. So, during the continuous aging and drying process of C. reticulata 'Chachi', the conversion products produced by this characteristic component have attracted our attention, which may be conducive to explaining the aging mechanism. A fragment ion m/z 134 was noticed in the MS/MS spectrum of MMA (Figure S2), we speculated that the structure of m/z 134 may be 3-hydroxyindole according to the chemical principles and properties of ESI-CID-MS induced rearrangement. This study confirmed that the production of 3-hydroxyindole in the degradation of MMA under LC-MS/MS analysis.
3-hydroxyindole was believed to play an important role besides indican in the synthesis of indole alkaloids in Isatis indigotica (Chinese woad), which is a species with an ancient and well-documented history as an indigo dye and medicinal plant [32] and similar plants also included Polygonum tinctorium [33,34], Isatis tinctoria (European woad, a species in the same genus as I. indigotica) [35,36,37]. Currently, two biosynthetic pathways of 3-hydroxyindole had been reported [38]. One is to be produced by indole (an intermediate metabolite of the tryptophan biosynthetic pathway) under the catalysis of a monooxygenase [39,40]. In addition, it is released by hydrolysis from indican. However, 3-hydroxyindole is unstable and it can be oxidized to isatin, which is further dimerized to indigo and indirubin [41]. Shintaro Inoue et al [42]. cloned a Flavin-containing monooxygenase (PtFMO) gene from P. tinctorium and found that indigo was produced in the presence of tryptophan when recombinant PtFMO was expressed in E. coli. The co-expression of PtFMO with indoxyl β-D-glucoside synthase (PtIGS), which catalyzes the glucosylation of 3-hydroxyindole, brought about the formation of indican in E. coli. In a word, as an indigo plant, P. tinctorium hydrolyzed the released indican to generate 3-hydroxyindole [43], which spontaneously converted to indigo in the presence of oxygen or undergone glycosylation reaction to recover indican. The dry roots and leaves of I. indigotica, named ‘Ban-Lan-Gen’ (BLG, Isatidis Radix) and ‘Da-Qing-Ye’ (DQY, Isatidis Folium), respectively [44], Zou et al [41] reported a LC-MS/MS quantitative method firstly to determine indican, isatin, indirubin and indigo in BLG and DQY. And the differences of chemical components of BLG and DQY lead to differences in pharmacological effects, indigo is often used for coloring and protecting liver and antibiosis. Indirubin has physiological properties that include anti-inflammatory [45] and anti-tumor activities [46], as well as strengthening the immune system [47].
In this study, we focused on the rearrangement ions generated from MMA induced by ESI-CID and predicted its structure to be 3-hydroxyindole. Derived from the previously reported pathway of 3-hydroxyindole, namely its oxidation and coupling and possible methylation reactions during the aging process of C. reticulata 'Chachi', 10 compounds derived from MMA were untargeted predicted and derived. Then the existence of these trace compounds was targeted verified in C. reticulata 'Chachi' and the changes during the aging process were observed through LC-MS/MS. In summary, we developed a strategy for discovering trace compounds in the aging process of C. reticulata 'Chachi' based on mass spectrometry fragmentation and known metabolic pathway, which provided reference value for the analysis of trace compounds from untargeted speculation to targeted verification. It is of great significance for explaining the superiority and 'aging is good' of C. reticulata 'Chachi'.
2. Materials and Methods
2.1. Reagents and materials
MMA was purchased from Zesheng Technology Co. Ltd (Anhui, China), 3-hydroxyindole was obtained from J&K Scientific (Shanghai, China)., Isatin, Indigo, Indirubin were purchased from Beijing Innochem Technology Co., Ltd (Beijing, China). MDA was purchased from TCI (Shanghai, China). N-methylisatin was purchased from Bide Pharmatech Ltd. (Shanghai, China). Dimethyl sulfoxide (DMSO) was purchased from Solarbio (Beijing, China). LC–MS grade methanol, acetonitrile, and ethanol were obtained from Merck (Darmstadt, Germany). water was purified with a Milli-Q water purification system (Millipore Corp., Bedford, MA, USA).
2.2. Synthesis of compounds 7, 9, 11 and 12
N, N', O, O'-Tetramethyl-leuko-indigo [48,49], N-methyl-3-hydroxyindole [50], N, N'-demethylindigo and N, N'-demethylindirubin [51,52] were synthesized according to the reported schemes. The specific synthesis steps or diagrams, the results of NMR and liquid chromatography-high resolution tandem mass spectrometry (LC-HR-MS) were all showed in the supplementary information (Figure S1).
2.3. Collection of C. reticulata 'Chachi' and C. reticulata 'Dahongpao'
C. reticulata 'Chachi' aged for different years were obtained from Xinhui District, Jiangmen City, Guangdong Province and authenticated by the National Institutes for Food and Drug Control. C. reticulata 'Dahongpao' samples were obtained from Sichuan Province. Detailed samples information is listed in the Table S1.
2.4. Samples extraction
Each C. reticulata 'Chachi' and C. reticulata 'Dahongpao' samples were ground powder with the aid of machine manufactured by Shanghai one bio Technology Co., LTD (Shanghai, China). An aliquot (1.0 g) of each sample was added to 20 mL of methanol with ultrasonication (SB-25-12DTD, Ningbo Scientz Biotechnology Co. Ltd., Ningbo, China) for 40 min at room temperature, followed by centrifugation at 8000 rpm for 5 min. The supernatants were stored at −80 °C until analysis and filtered through a 0.22 μm membrane filter before direct injection into the LC-MS/MS system.
2.5. Preparation of standard solutions
MMA and MDA standard stock solutions were prepared in ethanol. Isatin and N-methylisatin standard stock solutions in acetonitrile were prepared. Standard stock solutions of 3-hydroxyindole, indirubin, indigo and their N, N-methylated derivatives were dissolved in DMSO, to make a 1 mM stock solution and stored at −20 °C in the dark. The working standard solutions were prepared by diluting their stock solution with acetonitrile, a concentration of 100 μg∙mL-1 for LC-HR-MS analysis, 1 μg∙mL-1 for ESI-CID-MS analysis.
2.6. LC-HR-MS analysis
The 1260 HPLC system and quadrupole time-of-flight mass spectrometer equipped with an ESI interface (6538, Agilent) were performed. Agilent MassHunter Workstation software was used to control LC-MS, and to process the data. Isocratic elution of acetonitrile and water (80:20, v/v, 0.1% formic acid in both acetonitrile and water) was used for 2 min. The LC solvent flow rate was 0.3 mL/min, and injection volumes were 3 μL. The mass spectrometer was operated in positive mode. The ESI parameters were optimized as follows: capillary voltage 3000 V, drying gas (N2) 5.0 L/min, gas temperature 300 °C, nebulizer gas pressure 40 psi, sheath gas (N2) 10 L/min, fragmentor: 150V.
2.7. LC-ESI-CID-MS analysis
LC-ESI-CID-MS analysis was performed on a Waters Xevo TQ-S (Waters Corp., Milford, MA, USA), chromatographic separation was performed at 25 °C on an Thermo Scientific™ Hypersil GOLD™ Phenyl reversed phase HPLC columns. (1.9 μm, 2.1 × 100 mm). The data were analyzed with MassLynx. A gradient elution of 0.1% formic acid in water (A) and acetonitrile (B) was used: 0-0.75 min, 5% B; 0.75-1 min, 5-25% B; 1-3 min, 25-50% B; 3-9 min, 50-95% B; 9-11min, 95% B; 11-13 min, 95-5% B; 13-15 min, 5% B. The injection volume was 3 μL, flow rate: 0.2 mL/min. The operating parameters were optimized as follows: capillary voltages: 3kV, cone voltages: 65 kV, desolvation temperature: 350 °C, desolvation gas flow: 550L/Hr, collision gas flow: 0.15 mL/min.
3. Results
3.1. Prediction of indole derivatives based on rearrangement ions and metabolic pathways
Based on previous speculation, the m/z 134 produced by MMA (compound 1) may be the 3-hydroxyindole (compound 2), and it is known that isatin (compound 3) is produced after the oxidation of 3-hydroxyindole, further generating indigo (compound 4) and indirubin (compound 5). Therefore, we reasonably proposed the pathway from compound 1 to compound 5.
In plant, modifying enzymes of different families act on many alkaloids to produce a diverse array of biologically important alkaloid derivatives having altered physical, chemical, and biological properties, chemical modification reactions catalyzed by these enzymes, including mainly methylation, glycosylation, oxidation, reduction, hydroxylation, and acylation [53]. Methylations play a central role in the functionalization of specialized metabolites, and the methyl transferase (MT) gene family enzymes catalyze this type of reaction [54], which O-methyltransferase (OMT) and N-methyltransferase (NMT) are well-known classes of MTs catalyzing the methylation of alkaloids. OMT represents the largest class of enzymes catalyzing the methyl transfer reaction at the hydroxyl position of the alkaloidal substrate. Unlike OMT, NMT acts on nitrogen atom of substrate adding methyl group [55,56]. For example, 6-O-methylated was derivative from coclaurine through 1-benzylisoquinoline 6-O-methyltransferase (6OMT), then N-methylcellulose can be formed under the catalysis of N-methyltransferase in benzylisoquinoline alkaloid biosynthesis in sacred lotus (Nelumbo nucifera) [56]. Therefore, we speculated that the characteristic component MMA in C. reticulata 'Chachi' may undergo methylation during the ripening and drying process of Citrus reticulata Blanco, resulting in MDA (compound 8), which is based on MMA with an additional methyl atom on the N atom. Similarly, MDA may also produce a structure of m/z 148, namely N-methyl-3-hydroxyindole (compound 9), which was inferred to also undergo oxidation and coupling reactions, generating N-methylisatin (compound 10), N, N'-demethylindigo (compound 11) and N, N'-demethylindirubin (compound 12). A production pathway from compound 8 to compound 12 may exist in C. reticulata 'Chachi' or other plants, corresponding to the pathway of compound 1-5.
In addition, compound 9 is an N-methylated derivative of compound 2, O-methylation reactions may also occur during the drying process of C. reticulata 'Chachi', so we considered that compound 6 can be derived from compound 2 and coupled by 2-site. The speculated coupling product is N, N', O, O'-Tetramethyl-leuko-indigo (compound 7). Based on the above speculation, we have summarized a production pathway of indole derivatives, as shown in Figure 1.
3.2. Feasibility verification of predicted compounds by the reference substance
Firstly, we validated that fragment ion 134 generated by MMA is 3-hydroxyindole. The LC-HR-MS accurately reflected the consistency between the 134.06 secondary fragment ions of MMA and 3-hydroxyindole (Figure 2A). According to the compound 1 to compound 2 in C. reticulata 'Chachi' during aging process, N-methyl-3-hydroxyindole should be found by the same fragmentation mode. Similarly, we compared 148.07 secondary fragment ions of MDA and compound 9 reference substances in LC-HR-MS (Figure 2B).
The powder of 3-hydroxyindole was exposed to ultraviolet light for 48 hours, and the oxidized substance was analyzed by LC-HR-MS. The full scan spectra of LC-HR-MS revealed that m/z 148.0393 and m/z 263.0817 were produced after exposure to oxygen and UV irradiation, consistent with the reported pathways, matching with the [M+H] + of isatin, indigo, and indirubin, respectively, as shown in Table 1. The extracted ion chromatograms (EIC) and secondary fragment ions of oxidized 3-hydroxyindole for protonated molecular weights at m/z 148.0, m/z 263.1 further confirmed the production of known pathway products. The experimental phenomenon in synthesis process showed that N-methyl-3-hydroxyindole (under argon protection) was light green, once it was exposed to oxygen, it would be oxidized, and the color changed to dark green, which indicated other transformations had also occurred. Therefore, the m/z 148.0757, N-methylisatin at m/z 162.0551 and N, N'-demethylindigo or N, N'-demethylindirubin at m/z 291.1128 were detected in the LC-HR-MS spectrum of synthetic solution. The retention time of EIC at m/z 162.1 and m/z 291.1 and secondary fragment ions were also consistent with compounds 10, 11 and 12. As we speculated, N-methyl-3-hydroxyindole spontaneously oxidized and dimerized after contacting with air, similar to the oxidative dimerization pathway of 3-hydroxyindole.
In the LC-HR-MS spectrum of 3-hydroxyindole methylation, the m/z 291.1128 were found. Although 3-methoxy-1-methylindole at m/z 162.0913 was not found, m/z 321.1592 was found, which was consistent with the prediction. In other words, after methylation at both N and O sites of 3-hydroxyindole, a 2-site coupling occurred, forming N, N', O, O' -Tetramethyl-leuko-indigo.
Since MMA is a characteristic component in C. reticulata 'Chachi', the predicted compounds originating from MMA should be found in C. reticulata 'Chachi'. This section examined the existence of the 11 compounds mentioned above in C. reticulata 'Chachi' by multiple reaction monitoring (MRM) mode, except for 3-methoxy-1-methylindole, which was difficult to obtain a reference substance. MMA was known to be a characteristic component in C. reticulata 'Chachi', with an ions peak at 6.20 minutes (Figure 3A). The m/z 134→106 ion pairs of 3-hydroxyindole were selected for MRM analysis, an ion peak at 3.42 minutes was observed. In the chromatogram of C. reticulata 'Chachi', although the main peak was mainly at the peak position of MMA (Figure S3), an ion peak at 3.42 minutes can also be observed, indicating the presence of 3-hydroxyindole (Figure 3B). In this study, it was found for the first time in C. reticulata 'Chachi' that MMA can be converted into 3-hydroxyindole. The spectra of C. reticulata 'Chachi' were compared with isatin (m/z 148→92), indigo (m/z 263→219) and indirubin (m/z 263→219). The results showed that C. reticulata 'Chachi' had an ion peak with the same retention time (3.39 minutes) as isatin, as shown in Figure 3C, and C. reticulata 'Chachi' exhibited a low abundance peak at 5.70 min and 6.08 min, with the same RT as indigo and indirubin (Figure 3D), respectively. This was also the first time for the conversion of 3-hydroxyindole to indigo or indirubin has been verified in C. reticulata 'Chachi'.
As shown in Figure 3F. the MRM chromatogram of selected ion pairs m/z 180→120 showed that C. reticulata 'Chachi' had an ion peak at 2.77 minutes with the same retention time as MDA, which indicating the presence of MDA in C. reticulata 'Chachi'. Rapid LC-MS/MS analysis was performed on the N-methyl-3-hydroxyindole synthesized solution, chromatogram of m/z 148→105 ion pairs showed that the retention time of N-methyl-3-hydroxyindole was 3.99 minutes, which ion peaks were observed in C. reticulata 'Chachi' at the same time (Figure 3G). The MRM spectra of C. reticulata 'Chachi' were compared with N-methylisatin (m/z 162→116), N, N'-demethylindigo (m/z 291→146) and N, N'-demethylindirubin (m/z 291→146). The results showed that C. reticulata 'Chachi' had the same retention time with N-methylisatin, N, N'-demethylindigo and N, N'-demethylindirubin at 3.83 minutes, 6.15 min and 6.53 min (Figure 3H, 3I), respectively. In addition, C. reticulata 'Chachi' and N, N', O, O'-Tetramethyl-leuko-indigo also had a consistent ions peak at 8.11 minutes (Figure 3E). The retention time of extracted ion chromatogram and secondary fragment ions between C. reticulata 'Chachi' and compounds 1-12 were basically consistent, as shown in the Table S2. The experimental results confirmed that our speculated production pathway of indole derivatives can be found not only in the reference substance, but also in C. reticulata 'Chachi', which demonstrated the unique role of MMA in C. reticulata 'Chachi'.
The synthesis solution of N-methyl-3-hydroxyindole turned dark green after oxidation, and after 48 h, the color changed to yellow, which the MRM chromatogram of m/z 291→146 ion pairs showed that it yielded four ion peaks except for the peaks of N, N'-demethylindigo and N, N'-demethylindirubin (Figure 3J). Interestingly, the chromatogram of C. reticulata 'Chachi' showed that not only compounds 11 and 12, but also two other ion peaks with corresponding RT of 6.34 min and 6.66 min, indicating that there may also be isomers of compounds 11 or 12 in C. reticulata 'Chachi', which requires further confirmation.
C. reticulata 'Chachi' of different years were collected and the above compounds were analyzed successively, summarized in Table 2. MMA, MDA, isatin, indigo and indirubin were all detected, regardless of the storage time of C. reticulata 'Chachi'. However, methylated products and their subsequent coupling products were not detected in some samples, possibly due to low content. During the storage period of 5-9 years, all compounds in the pathway were detected, indicating that the indole derivatives of C. reticulata 'Chachi' may be relatively stable during the aging time. When stored for more than 10 years, these components may undergo other transformations, which was meaningful for exploring the relationship between the efficacy and aging time of C. reticulata 'Chachi'. Interestingly, MMA and 3-hydroxyindole were not found in C. reticulata 'Dahongpao', while a small amount of indirubin, MDA and N-methyl-3-hydroxyindole were found in it (Figure S4). Therefore, we considered that this type of indole derivatives was related to the color of these two different types of CRP. The color of the peel changed from orange to dark green, then to brown, finally to dark brown even to black during the drying and aging process. The appearance of C. reticulata 'Dahongpao' appears reddish in color, and the scent is fresher compared to the thick taste of C. reticulata 'Chachi'.
4. Conclusion
A strategy for discovering trace compounds in the aging process of C. reticulata 'Chachi' has been developed based on the occurrence of non-specific free radical reaction or rearrangement reaction in the mass spectrometry and known metabolic pathway. A total of 11 degradation products and metabolites were predicted and derived. In detail, the component 3-hydroxyindole was found in the fragment ions of the unique component MMA in C. reticulata 'Chachi', and the same fragmented mode was found in the methylated product MDA of MMA, which produced N-methyl-3-hydroxyindole. In addition, the oxidative dimerized pathway of 3-hydroxyindole can also occur in N-methyl-3-hydroxyindole, producing isatin, indigo, indirubin, and their N-methyl or N, N-dimethyl products, respectively. Eleven compounds were verified the presence of C. reticulata 'Chachi' and C. reticulata 'Dahongpao'. MMA, 3-hydroxyindole, indigo and N, N-dimethylindigo as well as N, N', O, O'- tetramethylindigo were only found in C. reticulata 'Chachi', MDA was not a characteristic compound of C. reticulata 'Chachi', but also existed in the C. reticulata 'Dahongpao' cultivar. Indirubin can also be found in reddish C. reticulata 'Dahongpao'. It needs further study about the reasons for the differences between the two cultivars. Moreover, with the increase of C. reticulata 'Chachi' age, the content of these indole derivatives was relatively stable during a storage period of 5-9 years, which was conductive to the analysis of C. reticulata 'Chachi', such as identification of origin or age, explanation of browning color, etc. In addition, there may also be isomers in the C. reticulata 'Chachi', except for N, N'-demethylindigo and N, N'-demethylindirubin. The presence of these indole derivatives was reported for the first time in C. reticulata 'Chachi', which is of great significance for explaining the aging process of C. reticulata 'Chachi and may be applied to distinguish species. This strategy expands the method of discovering low abundance compounds in drugs and more comprehensively explores trace compounds in drugs to explain their relationship with pharmacological activity.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Tian Li: Investigation, Methodology, Writing – original draft. Ke Chen & Xiaoming Wang: Synthesis of target compounds. Ying Wang: Providing samples and identifing authenticity. Yinlong Guo: Conceptualization, Funding acquisition, Methodology, Writing – review & editing. Yue Su: Funding acquisition, Investigation, Validation.
Funding
This work was supported by the National Key R & D Program of China (Grant No. 2021YFF0701900), Science and Technology Innovation Action Plan of Shanghai (Grant No. 22DZ2204300).
Data Availability Statement
The data used to support the findings of this study can be made available by the corresponding author upon request.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Onorato, J.; Henion, J.D. Evaluation of Triterpene Glycoside Estrogenic Activity Using LC/MS and Immunoaffinity Extraction. Anal. Chem. 2001, 73, 4704–4710. [Google Scholar] [CrossRef] [PubMed]
- Visconti, G.; de Figueiredo, M.; Strassel, O.; Boccard, J.; Vuilleumier, N.; Jaques, D.; Ponte, B.; Rudaz, S. Multitargeted Internal Calibration for the Quantification of Chronic Kidney Disease-Related Endogenous Metabolites Using Liquid Chromatography–Mass Spectrometry. Anal. Chem. 2023, 95, 13546–13554. [Google Scholar] [CrossRef]
- Zhang, X.; Su, M.; Du, J.; Zhou, H.; Li, X.; Zhang, M.; Hu, Y.; Ye, Z. Profiling of naturally occurring proanthocyanidins and other phenolic compounds in a diverse peach germplasm by LC-MS/MS. Food Chem. 2023, 403, 134471. [Google Scholar] [CrossRef]
- Xue, G.; Su, S.; Yan, P.; Shang, J.; Wang, J.; Yan, C.; Li, J.; Wang, Q.; Xiong, X.; Xu, H. Integrative analyses of widely targeted metabolomic profiling and derivatization-based LC-MS/MS reveals metabolic changes of Zingiberis Rhizoma and its processed products. Food Chem. 2022, 389, 133068. [Google Scholar] [CrossRef]
- Custodio-Mendoza, J.A.; Sendón, R.; de Quirós, A.R.-B.; Lorenzo, R.A.; Carro, A.M. Development of a QuEChERS method for simultaneous analysis of 3-Monochloropropane-1,2-diol monoesters and Glycidyl esters in edible oils and margarine by LC-APCI-MS/MS. Analytica Chimica Acta 2023, 1239, 340712. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics--the link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar] [CrossRef]
- Wang, L.; Xing, X.; Chen, L.; Yang, L.; Su, X.; Rabitz, H.; Lu, W.; Rabinowitz, J.D. Peak Annotation and Verification Engine for Untargeted LC–MS Metabolomics. Anal. Chem. 2019, 91, 1838–1846. [Google Scholar] [CrossRef]
- Qiu, J.; Li, T.; Zhu, Z.-J. Multi-dimensional characterization and identification of sterols in untargeted LC-MS analysis using all ion fragmentation technology. Analytica Chimica Acta 2021, 1142, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Wu, H.-Y.; Wu, W.-S.; Hsu, J.-Y.; Chang, C.-W.; Lee, Y.-H.; Liao, P.-C. Identification of Xenobiotic Biotransformation Products Using Mass Spectrometry-Based Metabolomics Integrated with a Structural Elucidation Strategy by Assembling Fragment Signatures. Anal. Chem. 2023, 95, 14279–14287. [Google Scholar] [CrossRef]
- Dudley, E.; Yousef, M.; Wang, Y.; Griffiths, W.J. Chapter 2 - Targeted metabolomics and mass spectrometry. In Adv. Protein Chem. Struct. Biol.; Donev, R., Ed.; Academic Press, 2010; Volume 80, pp. 45–83. [Google Scholar]
- Kell, D.B. Metabolomics and systems biology: making sense of the soup. Curr. Opin. Microbiol. 2004, 7, 296–307. [Google Scholar] [CrossRef]
- Wang, Z.; Cui, B.; Zhang, F.; Yang, Y.; Shen, X.; Li, Z.; Zhao, W.; Zhang, Y.; Deng, K.; Rong, Z.; et al. Development of a Correlative Strategy To Discover Colorectal Tumor Tissue Derived Metabolite Biomarkers in Plasma Using Untargeted Metabolomics. Anal. Chem. 2019, 91, 2401–2408. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Z.; Chen, Y.-Y.; Wu, X.-Z.; Bai, P.-R.; An, N.; Liu, X.-L.; Zhu, Q.-F.; Feng, Y.-Q. Uncovering the Carboxylated Metabolome in Gut Microbiota–Host Co-metabolism: A Chemical Derivatization-Molecular Networking Approach. Anal. Chem. 2023, 95, 11550–11557. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Cai, J.; Kong, H.; Wu, M.; Hua, R.; Zhao, M.; Liu, J.; Xu, G. Analysis of Cigarette Smoke Condensates by Comprehensive Two-Dimensional Gas Chromatography/Time-of-Flight Mass Spectrometry I Acidic Fraction. Anal. Chem. 2003, 75, 4441–4451. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhao, M.; Kong, H.; Cai, J.; Wu, J.; Wu, M.; Hua, R.; Liu, J.; Xu, G. Characterization of complex hydrocarbons in cigarette smoke condensate by gas chromatography–mass spectrometry and comprehensive two-dimensional gas chromatography–time-of-flight mass spectrometry. J. Chromatogr. A 2004, 1043, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liao, J.; Le, W.; Wu, G.; Zhang, W. Improving the Data Quality of Untargeted Metabolomics through a Targeted Data-Dependent Acquisition Based on an Inclusion List of Differential and Preidentified Ions. Anal. Chem. 2023, 95, 12964–12973. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-L.; Zhou, B.-W.; Cheng, J.; Zhang, F.; Zhang, J.; Zhang, L.; Guo, Y.-L. Mass Spectrometry-Based Discovery of New Chemical Scaffold Rearrangement Ions: Aza-biphenylene as a Novel Potent Biradical Agent in Cancer Chemotherapy. Anal. Chem. 2020, 92, 14517–14527. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, S.; Zhang, X.; Wang, H.; Zhang, F.; Hou, Y.; Su, Y.; Guo, Y. An unexpected acid-catalyzed decomposition reaction of cilnidipine and pranidipine to the decarboxylative bridged tricyclic products via cascade rearrangements. Org. Chem. Front. 2017, 4, 2163–2166. [Google Scholar] [CrossRef]
- Lu, H.; Yin, Y.; Sun, J.; Li, W.; Shen, X.; Feng, X.; Ouyang, J.; Na, N. Accelerated plasma degradation of organic pollutants in milliseconds and examinations by mass spectrometry. Chin. Chem. Lett. 2021, 32, 3457–3462. [Google Scholar] [CrossRef]
- Liu, N.; Li, X.; Zhao, P.; Zhang, X.; Qiao, O.; Huang, L.; Guo, L.; Gao, W. A review of chemical constituents and health-promoting effects of citrus peels. Food Chem. 2021, 365, 130585. [Google Scholar] [CrossRef]
- Yu, X.; Sun, S.; Guo, Y.; Liu, Y.; Yang, D.; Li, G.; Lü, S. Citri Reticulatae Pericarpium (Chenpi): Botany, ethnopharmacology, phytochemistry, and pharmacology of a frequently used traditional Chinese medicine. J. Ethnopharmacol. 2018, 220, 265–282. [Google Scholar] [CrossRef]
- Fu, M.; Xu, Y.; Chen, Y.; Wu, J.; Yu, Y.; Zou, B.; An, K.; Xiao, G. Evaluation of bioactive flavonoids and antioxidant activity in Pericarpium Citri Reticulatae (Citrus reticulata ‘Chachi’) during storage. Food Chem. 2017, 230, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-J.; Wu, X.; Li, M.-M.; Li, G.-Q.; Yang, Y.-T.; Luo, H.-J.; Huang, W.-H.; Chung, H.Y.; Ye, W.-C.; Wang, G.-C.; et al. Antiviral Activity of Polymethoxylated Flavones from “Guangchenpi”, the Edible and Medicinal Pericarps of Citrus reticulata ‘Chachi’. J. Agric. Food. Chem. 2014, 62, 2182–2189. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Zou, B.; An, K.; Yu, Y.; Tang, D.; Wu, J.; Xu, Y.; Ti, H. Anti-asthmatic activity of alkaloid compounds from Pericarpium Citri Reticulatae (Citrus reticulata ‘Chachi’). Food Funct. 2019, 10, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Chen, X.; Li, Y.; Li, L. Effect of Drying Methods on Volatile Compounds of Citrus reticulata Ponkan and Chachi Peels as Characterized by GC-MS and GC-IMS. Foods (Basel, Switzerland) 2022, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Jiang, Y.; Wen, L.; Yang, B. Characterization of polysaccharide structure in Citrus reticulate ‘Chachi’ peel during storage and their bioactivity. Carbohydr. Res. 2021, 508, 108398. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.-L.; Chen, X.-L.; Gong, M.-J.; Xu, Y.; Tu, H.-S.; Zhang, L.; Liao, B.-s.; Qiu, X.-H.; Zhang, J.; Huang, Z.-H.; et al. Guang Chen Pi (the pericarp of Citrus reticulata Blanco's cultivars ‘Chachi’) inhibits macrophage-derived foam cell formation. J. Ethnopharmacol. 2022, 293, 115328. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, X.; Gao, Y.; Han, Y.; Wu, X. Path analysis of non-enzymatic browning in Dongbei Suancai during storage caused by different fermentation conditions. Food Chem. 2021, 335, 127620. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-N.; Xie, S.-M.; Dai, Y.-T. Study on the change of compositions and Maillard Browning Reaction in Guang Citri Reticulatae Pericarpium during ageing. Journal of Guangdong Pharmaceutical University 2023, 39, 73–81. [Google Scholar]
- Li, S.-Z.; Guan, X.-M.; Gao, Z.; Lan, H.-C.; Yin, Q.; Chu, C.; Yang, D.-P.; Liu, E.H.; Zhou, P. A simple method to discriminate Guangchenpi and Chenpi by high-performance thin-layer chromatography and high-performance liquid chromatography based on analysis of dimethyl anthranilate. J. Chromatogr. B 2019, 1126-1127, 121736. [Google Scholar] [CrossRef]
- Chao, Y.; Tan, E.y.; Ma, S.; Chen, B.; Liu, M.; Wang, K.; Yang, W.; Wei, M.; Zheng, G. Dynamic variation of the phytochemical and volatile compounds in the pericarp of Citrus reticulata ‘‘Chachi’’ (Rutaceae) during 2 years of storage. J. Food Sci. 2022, 87, 153–164. [Google Scholar] [CrossRef]
- Feng, J.; Huang, D.; Yang, Y.; Chen, J.; Qiu, S.; Lv, Z.; Ma, X.; Li, Y.; Li, R.; Xiao, Y.; et al. Isatis indigotica: from (ethno) botany, biochemistry to synthetic biology. Mol Horticulture 2021, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Angelini, L.G.; Campeol, E.; Tozzi, S.; Gilbert, K.G.; Cooke, D.T.; John, P. A New HPLC-ELSD Method To Quantify Indican in Polygonum tinctorium L. and To Evaluate β-Glucosidase Hydrolysis of Indican for Indigo Production. Biotechnol. Progr. 2003, 19, 1792–1797. [Google Scholar] [CrossRef]
- Gilbert, K.G.; Maule, H.G.; Rudolph, B.; Lewis, M.; Vandenburg, H.; Sales, E.; Tozzi, S.; Cooke, D.T. Quantitative Analysis of Indigo and Indigo Precursors in Leaves of Isatis spp. and Polygonum tinctorium. Biotechnol. Progr. 2004, 20, 1289–1292. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.-K.-O.; Marcelo, P.; Gontier, E.; Dauwe, R. Metabolic markers for the yield of lipophilic indole alkaloids in dried woad leaves (Isatis tinctoria L.). Phytochemistry 2019, 163, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Oberthür, C.; Graf, H.; Hamburger, M. The content of indigo precursors in Isatis tinctoria leaves--a comparative study of selected accessions and post-harvest treatments. Phytochemistry 2004, 65, 3261–3268. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Chen, M.-H.; Wang, X.-L.; Guo, Q.-L.; Zhu, C.-G.; Lin, S.; Xu, C.-B.; Jiang, Y.-P.; Li, Y.-H.; Jiang, J.-D.; et al. Antiviral enantiomers of a bisindole alkaloid with a new carbon skeleton from the roots of Isatis indigotica. Chin. Chem. Lett. 2015, 26, 931–936. [Google Scholar] [CrossRef]
- Chen, J.; Dong, X.; Li, Q.; Zhou, X.; Gao, S.; Chen, R.; Sun, L.; Zhang, L.; Chen, W. Biosynthesis of the active compounds of Isatis indigoticabased on transcriptome sequencing and metabolites profiling. BMC Genomics 2013, 14, 857. [Google Scholar] [CrossRef]
- Maier, W.; Schumann, B.; Gröger, D. Biosynthesis of indoxyl derivatives in Isatis tinctoria and Polygonum tinctorium. Phytochemistry 1990, 29, 817–819. [Google Scholar] [CrossRef]
- Xia, Z.-Q.; Zenk, M.H. Biosynthesis of indigo precursors in higher plants. Phytochemistry 1992, 31, 2695–2697. [Google Scholar] [CrossRef]
- Zou, P.; Koh, H.L. Determination of indican, isatin, indirubin and indigotin in Isatis indigotica by liquid chromatography/electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 1239–1246. [Google Scholar] [CrossRef]
- Inoue, S.; Morita, R.; Minami, Y. An indigo-producing plant, Polygonum tinctorium, possesses a flavin-containing monooxygenase capable of oxidizing indole. Biochem. Biophys. Res. Commun. 2021, 534, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Minami, Y.; Kanafuji, T.; Miura, K. Purification and Characterization of a β-Glucosidase from Polygonum tinctorium, Which Catalyzes Preferentially the Hydrolysis of Indican. Biosci., Biotechnol., Biochem. 1996, 60, 147–149. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, Z.; Gao, T.; Chen, Y.; Yang, Q.; Fu, C.; Zhu, Y.; Wang, F.; Liao, W. Isatidis Radix and Isatidis Folium: A systematic review on ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2022, 283, 114648. [Google Scholar] [CrossRef]
- Xie, J.; Tian, S.; Liu, J.; Huang, S.; Yang, M.; Yang, X.; Xu, R.; Lin, J.; Han, L.; Zhang, D. Combination Therapy with Indigo and Indirubin for Ulcerative Colitis via Reinforcing Intestinal Barrier Function. Oxid. Med. Cell. Longev. 2023, 2023, 2894695. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Peng, R.; Min, Q.; Hui, S.; Chen, X.; Yang, G.; Qin, S. Bisindole natural products: A vital source for the development of new anticancer drugs. Eur. J. Med. Chem. 2022, 243, 114748. [Google Scholar] [CrossRef] [PubMed]
- Shao, K.; Wang, T.; Li, T.; Zhang, A.; Cai, M.; Zhao, G.; Fu, Q.; Wang, Q.; Liu, X.; Hou, M. Indirubin regulates MPL and TNF expression in peripheral blood mononuclear cells from patients with primary immune thrombocytopenia. Exp. Hematol. 2019, 73, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Wei, H.; Duan, Y.; Ning, H.; Yu, J.; Zhu, Y.; Hou, W.; Li, Y. An Improved Synthesis of the Triethylene Glycol-Substituted 4-(N-Methyl-N-Boc-Amino)Styrylpyridine. ACS Omega 2020, 5, 19446–19452. [Google Scholar] [CrossRef] [PubMed]
- Banjare, S.K.; Nanda, T.; Ravikumar, P.C. Cobalt-Catalyzed Regioselective Direct C-4 Alkenylation of 3-Acetylindole with Michael Acceptors Using a Weakly Coordinating Functional Group. Org. Lett. 2019, 21, 8138–8143. [Google Scholar] [CrossRef]
- Liu, Q.; Li, G.; Yi, H.; Wu, P.; Liu, J.; Lei, A. Pd-Catalyzed Direct and Selective C-H Functionalization: C3-Acetoxylation of Indoles. Chemistry – A European Journal 2011, 17, 2353–2357. [Google Scholar] [CrossRef]
- McCosker, P.M.; Butler, N.M.; Shakoori, A.; Volland, M.K.; Perry, M.J.; Mullen, J.W.; Willis, A.C.; Clark, T.; Bremner, J.B.; Guldi, D.M.; et al. The Cascade Reactions of Indigo with Propargyl Substrates for Heterocyclic and Photophysical Diversity. Chemistry – A European Journal 2021, 27, 3708–3721. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Bonasera, A.; Hristov, L.; Garmshausen, Y.; Schmidt, B.M.; Jacquemin, D.; Hecht, S. N,N′-Disubstituted Indigos as Readily Available Red-Light Photoswitches with Tunable Thermal Half-Lives. J. Am. Chem. Soc. 2017, 139, 15205–15211. [Google Scholar] [CrossRef] [PubMed]
- Bhambhani, S.; Kondhare, K.R.; Giri, A.P. Diversity in Chemical Structures and Biological Properties of Plant Alkaloids. Molecules (Basel, Switzerland) 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Rahikainen, M.; Alegre, S.; Trotta, A.; Pascual, J.; Kangasjärvi, S. Trans-methylation reactions in plants: focus on the activated methyl cycle. Physiol. Plant. 2018, 162, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Liscombe, D.K.; Facchini, P.J. Evolutionary and cellular webs in benzylisoquinoline alkaloid biosynthesis. Curr. Opin. Biotechnol. 2008, 19, 173–180. [Google Scholar] [CrossRef]
- Menéndez-Perdomo, I.M.; Facchini, P.J. Isolation and characterization of two O-methyltransferases involved in benzylisoquinoline alkaloid biosynthesis in sacred lotus (Nelumbo nucifera). J. Biol. Chem. 2020, 295, 1598–1612. [Google Scholar] [CrossRef]
Figure 1.
Putative diagram of indole derivatives.
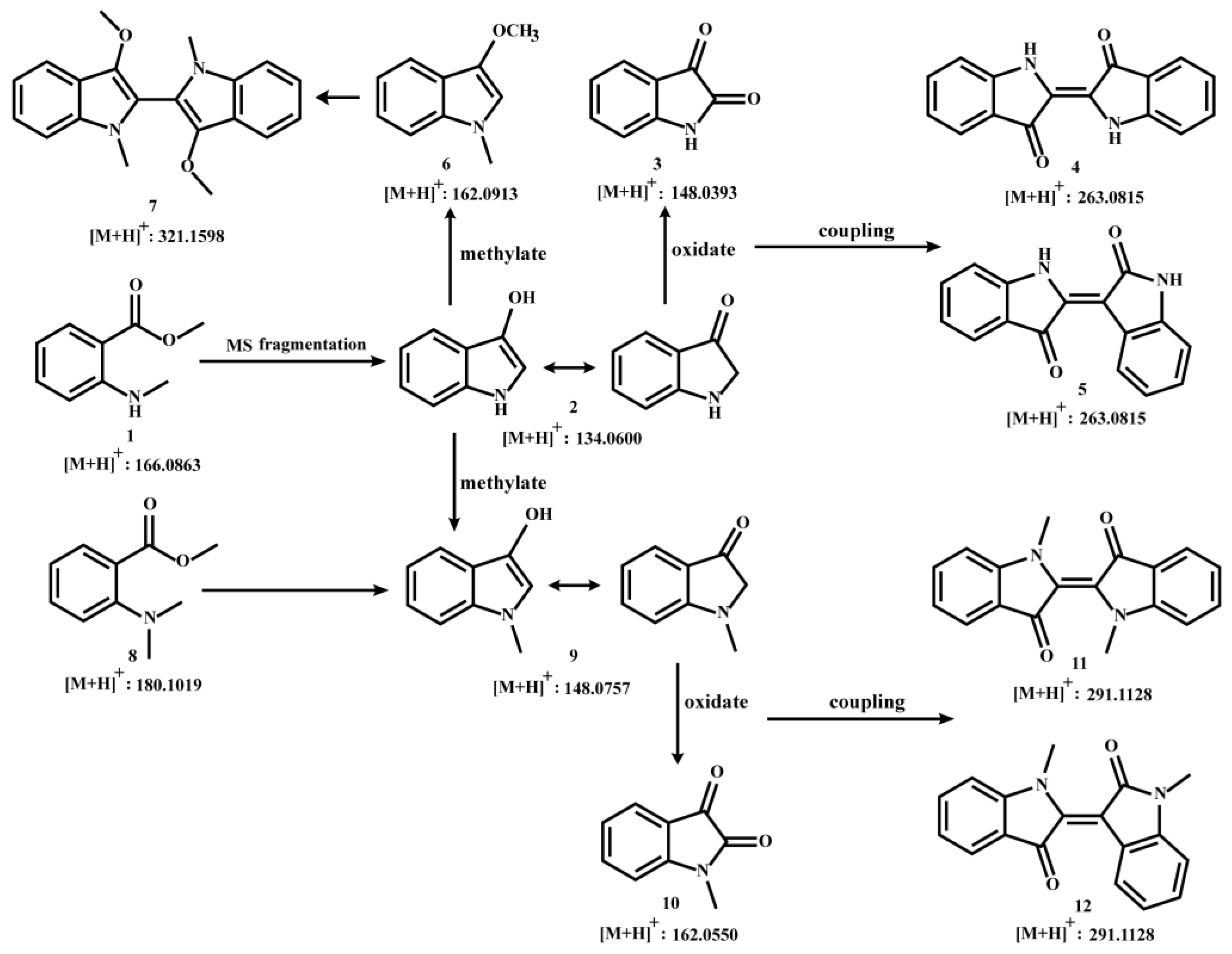
Figure 2.
(A): MS/MS spectra of MMA and 3-hydroxyindole at m/z 134.06 in LC-HR-MS. (B): MS/MS spectra of MDA and N-methyl-3-hydroxyindole at m/z 148.0757 in LC-HR-MS.
Figure 2.
(A): MS/MS spectra of MMA and 3-hydroxyindole at m/z 134.06 in LC-HR-MS. (B): MS/MS spectra of MDA and N-methyl-3-hydroxyindole at m/z 148.0757 in LC-HR-MS.
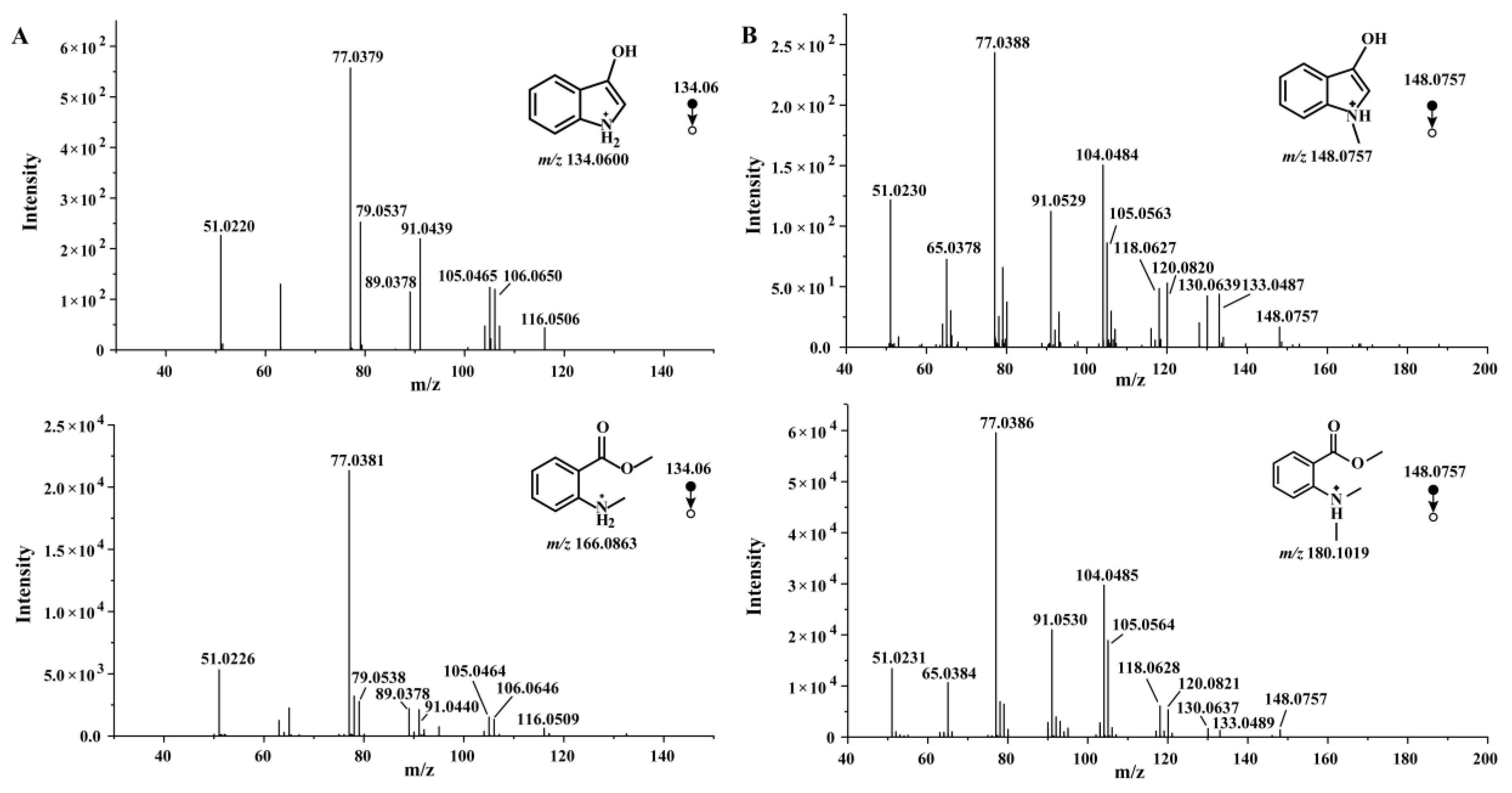
Figure 3.
Comparison of MRM chromatogram between compounds 1-12 and C. reticulata 'Chachi'. (A) MMA; (B) 3-hydroxyindole; (C) isatin; (D) indigo and indirubin; (E) N, N', O, O'-Tetramethyl-leuko-indigo; (F) MDA; (G) N-methyl-3-hydroxyindole; (H) N-methylisatin; (I) N, N'-demethylindigo and N, N'-demethylindirubin; (J) synthesis fluid of N-methyl-3-hydroxyindole.
Figure 3.
Comparison of MRM chromatogram between compounds 1-12 and C. reticulata 'Chachi'. (A) MMA; (B) 3-hydroxyindole; (C) isatin; (D) indigo and indirubin; (E) N, N', O, O'-Tetramethyl-leuko-indigo; (F) MDA; (G) N-methyl-3-hydroxyindole; (H) N-methylisatin; (I) N, N'-demethylindigo and N, N'-demethylindirubin; (J) synthesis fluid of N-methyl-3-hydroxyindole.

Table 1.
Feasibility verification of predicted compounds by the reference substance.
| Substrates | Reaction | Products | Structure | Retention time/min | [M+H] + | Secondary fragment ions (m/z) |
|
|---|---|---|---|---|---|---|---|
| Theoretical mass (m/z) |
Experimental mass (m/z) |
||||||
| 3-hydroxyindole | oxidation | Isatin | 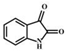 |
3.39 | 148.0393 | 148.0393 | 65.0, 77.0, 92.0, 102.0, 120.0, 130.0 |
| Indigo | 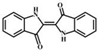 |
5.70 | 263.0815 | 263.0817 | 132.0, 206.0, 180.0, 190.0, 235.0, 219.0 | ||
| Indirubin | 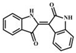 |
6.08 | 263.0815 | 263.0817 | 190.0, 219.0, 165.0, 132.0, 235.0, 206.0 | ||
| methylation | N, N', O, O'-Tetramethyl-leuko-indigo | 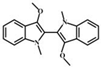 |
8.11 | 321.1598 | 321.1592 | 306.1, 291.1, 275.1, 247.1, 175.1, 160.1 | |
| N-methyl-3-hydroxyindole | oxidation | N-methylisatin | 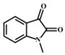 |
3.83 | 162.0550 | 162.0551 | 116.0, 106.0, 79.0, 65.0, 77.0, 134.0, 144.0 |
| N, N'-demethylindigo | 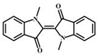 |
6.15 | 291.1128 | 291.1128 | 132.0, 276.0, 146.1, 247.1, 120.1, 172.1 | ||
| N, N'-demethylindirubin | 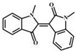 |
6.53 | 291.1128 | 291.1128 | 247.1, 276.1, 146.1, 132.0, 263.1, 158.1 | ||
Note, the retention time, accurate mass and secondary fragment ions were consistent with the reference substances3.3. Analysis and validation of compounds in C. reticulata 'Chachi'.
Table 2.
Differences in 12 compounds of C. reticulata 'Chachi' in different years and C. reticulata 'Dahongpao'.
Table 2.
Differences in 12 compounds of C. reticulata 'Chachi' in different years and C. reticulata 'Dahongpao'.
| Samples Number | Aging Time | MMA | 3-hydroxyindole | isatin | indigo | indirubin | N, N', O, O'-Tetramethyl-leuko-indigo | MDA | N-methyl-3-hydroxyindole | N-methylisatin | N, N'-demethylindigo | N, N'-demethylindirubin |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1 | +++ | − | ++ | + | + | + | ++ | ++ | ++ | + | + |
| 2 | 2 | +++ | ++ | ++ | + | + | − | ++ | ++ | − | + | + |
| 3 | 2 | +++ | − | ++ | ++ | + | + | ++ | − | ++ | + | + |
| 4 | 3 | +++ | ++ | ++ | + | + | ++ | ++ | ++ | − | + | + |
| 5 | 3 | +++ | − | ++ | + | + | − | ++ | +++ | ++ | − | + |
| 6 | 3 | +++ | ++ | ++ | + | + | + | ++ | ++ | ++ | + | − |
| 7 | 5 | +++ | ++ | ++ | + | + | + | ++ | ++ | ++ | + | + |
| 8 | 5 | +++ | ++ | ++ | + | + | + | ++ | ++ | ++ | + | + |
| 9 | 6 | +++ | ++ | ++ | + | + | + | ++ | ++ | ++ | + | + |
| 10 | 9 | +++ | ++ | ++ | + | + | + | ++ | ++ | ++ | + | + |
| 11 | 10 | +++ | ++ | ++ | + | + | − | ++ | ++ | ++ | ++ | + |
| 12 | 10 | +++ | ++ | ++ | + | + | + | ++ | ++ | − | − | + |
| 13 | >10 | +++ | ++ | ++ | + | + | + | ++ | ++ | − | + | + |
| 14 | >10 | +++ | ++ | ++ | + | + | + | ++ | ++ | − | − | + |
| 15 | >10 | +++ | ++ | ++ | + | + | − | ++ | +++ | ++ | + | + |
| 16 | >10 | +++ | ++ | ++ | ++ | + | ++ | ++ | ++ | ++ | + | + |
| 17 | >10 | +++ | − | ++ | + | + | + | +++ | ++ | ++ | + | − |
| 1' | 1 | − | − | ++ | − | + | − | ++ | ++ | ++ | − | − |
| 2' | 2 | − | − | ++ | − | + | − | ++ | ++ | ++ | − | − |
| 3' | 3 | − | − | ++ | − | + | − | ++ | ++ | ++ | − | − |
Note, Samples Number 1-17: C. reticulata 'Chachi'; 1'-3': C. reticulata 'Dahongpao'; +: the MS signals intensity of samples; −: undetected.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



