
Submitted:
03 November 2023
Posted:
06 November 2023
You are already at the latest version
Abstract
Carbohydrates such as fructooligosaccharides (FOS) are associated with improved gastrointestinal health and prevention of excess body fat. We evaluated the long-term effects of high amounts of FOS on metabolic parameters, non-alcoholic fatty liver disease (NAFLD) and short-chain fatty acids (SCFA). Sixty C57BL/6 mice received diets for four months: control (C), normolipid rich in fiber (F), normolipid supplemented with FOS (FOS), high fat (HL), high fat with high fiber (HLF) and high fat with FOS (HLFOS). We analyzed: animal weight; body composition; food intake; fasting blood glucose; serum and liver lipid profile; liver and intestinal histologies; malondialdehyde (MDA), hepatic retinol and α-tocopherol; SCFA in the feces. Supplementation with FOS in a high-fat diet promoted less body weight gain and reduced liver and retroperitoneal adipose tissue weights compared to HL and HF. FOS prevented NASH and decreased alanine aminotransferase and serum cholesterol levels in experimental animal models of obesity and metabolic syndrome (MS). There was found statistical differences in the dosages of the three main SCFAs in feces (acetic, isobutyric and isovaleric acids). Long-term supplementation with high doses of FOS was effective in reducing weight, adiposity, NAFLD and serum cholesterol in C57BL mice with obesity and MS induced by a high-fat diet.
Keywords:
Fructooligosaccharides (FOS)
; Hyperlipidic Diet
; Non-alcoholic fatty liver disease (NAFLD)
; prebiotics.
1. Introduction
Non-alcoholic fatty liver disease (NAFLD) is the most common cause of chronic liver disease of the 21s century, with a prevalence rates range between 23% and 32% depending on the geographical region [1,2]. NAFLD is seen in 47·3–63·7% of people with type 2 diabetes and up to 80% of people with obesity, because of its close association with the metabolic syndrome (MS) [3,4], and with lipid accumulation, inflammation, excessive calorie intake, genetic susceptibility, and insulin resistance [5]. Despite not being included as one of the criteria for diagnosing MS, NAFLD, the pro-inflammatory state and endothelial dysfunction are known to be associated with the metabolic, physiological and biochemical changes of this syndrome [6,7].
The intestinal microbiota establishes a symbiotic relationship with its host, contributing nutrients and energy by metabolizing dietary components in the large intestine, including cholesterol [8]. Several studies suggest that the intestinal microbiome represents an environmental factor that contributes to the development of NAFLD [9]. The intestinal microbiota may vary according to the stage of NAFLD, and more studies are needed to confirm the influence of specific bacteria on liver diseases [10].
Fructooligosaccharides (FOS) are widely used prebiotics, being a source of energy and an essential nutrient for intestinal bacteria, which carry out their fermentation and promote the colonization and activities of beneficial bacteria, improve the metabolism of the intestinal microbiota, improve host immunity and reduce inflammation [11]. Many probiotics and prebiotics have been linked to maintaining intestinal microbiota homeostasis and reducing NAFLD-associated dysregulation of hepatic carbohydrate and lipid metabolism. Therefore, microbiota-based treatments are beneficial for the prevention and treatment of NAFLD, however, more studies are needed to understand the mechanisms used by FOSs [10].
To elucidate the potential benefits of FOS in improving gastrointestinal health, systemic inflammation and metabolic parameters, it is necessary to establish an animal experimental protocol that evaluates a dietary intervention with this prebiotic in the long term. It is expected that the provision of high doses of FOS will contribute to the consolidation of benefits, as well as to the investigation of possible harms, in experimental animal models. Although the use of 25% of FOS was verified in the study by Mao et al (2018), this amount is too high if we consider the tolerance limit in humans. The present work invested in methodological differences by analyzing high doses of FOS supplemented in diets with two types of lipid composition and offered in the long term, which is little described in similar studies in the literature. Thus, the effects of 15% FOS supplementation for four months on metabolic parameters and the production of short-chain fatty acids in the feces of C57BL mice fed both a normolipid and hyperlipid diet rich in fiber were evaluated.
2. Materials and Methods
2.1. Animals and Diet
Sixty male C57BL mice weighing 20 g at the beginning of the experiment were obtained from the Central Animal House of the Faculty of Medicine of Ribeirão Preto (FMRP), University of São Paulo (USP), and maintained under controlled conditions of temperature (24 ± 2 °C) and of humidity and on a light (7:00 am–7:00 pm)/dark (7:00 pm–7:00 am) cycle. Water and food were supplied ad libitum. Animals were handled according to Brazilian College of Animal Experimentation recommendations and all procedures were approved by the Ethics Committee of FMRP (protocol no. 10/2016).
The animals underwent a period of adaptation to the environment and diet for 7 days. During this period, experimental diets were gradually introduced. Animals were randomly assigned to six experimental groups of 6 mices each: control (C), normolipid rich in fiber (F), normolipid supplemented with FOS (FOS), high fat (HL), high fat with high fiber (HLF) and high fat with FOS (HLFOS). For FOS supplementation in the diets, the product Orafti® SIPX (2016) from Beneo Animal Nutrition was used, which consists of powdered chicory inulin containing a mixture of oligosaccharides and polysaccharides composed of fructose units. The soybean oil (®Liza) was purchased at the local market. Vitamin, mineral mix, choline, and L-cystine were purchased from Rhoster (Araçoiaba da Serra, Brazil). All diets were based on the AIN-93 and are described in Table 1 [12].
Food intake and weight were determined per cage (4 animals/cage) over a period of 18 weeks and are reported as mean food intake and weight in g/day. Weight was measured weekly in the morning, between 8:00 and 9:00. The food intake was measured twice a week, always leaving sufficient quantity available.
At the end of experiment, animals were starved for 12 hours and then anesthetized with ketamine and xylazine diluted in saline at the proportion of 1 : 1:2 ml. It was administrated dose applications of 10 µl/g weight each. Blood was immediately collected by cardiac puncture, left to rest at room temperature for 30 minutes, and centrifuged at 3500 rpm at 4°C for serum separation afterward. Serum was stored at −80°C for later analysis. Liver and colon were weighed and frozen in aluminum parts for further analysis. The animals’ feces were extracted from the final portion of the intestine after euthanasia and stored in eppendorf tubes and frozen at -20°C.
2.2. Histopathological Analysis of the Intestine
Colon fragments were sectioned in an annular cross-section (proximal, middle and distal thirds), and the material was immersed in buffered formalin. The fragments were analyzed together, choosing well-oriented villi, with apparent and continuous basal, medial and apical portions [13]. For this analysis, a conventional light microscope with 20x magnification was used. Subsequently, the images were analyzed using the Image J software to quantify the intestinal muscle thickness and the total diameter of the intestinal lumen.
2.3. Histopathological Analysis of the Liver
Liver fragments were fixed in 10% buffered formalin for 24 hours and embedded in paraffin. Histological preparations containing 4 µm thick sections were stained with hematoxylin and eosin (H&E). Hepatic steatosis was evaluated semi-quantitatively and classified in crosses, according to Oh et al. (1998) [14] with some modifications. The steatosis score is associated with the morphological location of the liver (zone 1, 2 and 3), with crosses being assigned according to the degree of steatosis: without steatosis (0Y); 1-25% in zone 3 only (1Y); 25-50% only in zone 3 (2Y); 50-75% encompassing zones 2 and 3 (3Y) and 75-100% involving zones 1, 2 and 3 (4Y). In addition, the presence of inflammatory infiltrate (L: mild; M: moderate and I: intense) and Mallory's bodies (A: absent; FC: few corpuscles and MC: many corpuscles) were also evaluated. For these analyses, a conventional light microscope with magnifications of 20 and 40 times was used.
2.4. Biochemical and Hepatic Analyses
Protein determination in liver and serum was performed using a commercial kit using the Biuret method (Labtest Diagnóstica S.A., Brazil). Total cholesterol (TC) and serum triacylglycerides (TAG), as well as in the liver, were determined using commercial kits from Labtest (Labtest Diagnóstica S.A., Brazil). Aspartate aminotransferase (AST) and alanine aminotransferase (ALT) transaminases were quantitatively determined using a commercial Labtest kit (Labtest Diagnóstica S.A., Brazil) in a serum sample with continuous UV kinetic reaction. For the determination and quantification of total fat in the liver, the method proposed by Bligh and Dyer (1959) [15] was used.
2.5. Analysis of Glycemia
At the end of experiment, animals glycemia was determined using obtained samples from animal's tail and the freestyle lite Abbot®A glucometer.
2.6. Analysis of Lipid Peroxidation and Antioxidant Parameters
The analysis of the Total Antioxidant Capacity (TAC) was performed based on the methodology described by Erel (2004) [16]. The Gerard-Monnier et al. method [17] with some modifications, as it is thoroughly described in S1 text section, was used to determine the hepatic malondialdehyde (MDA). Vitamin A (α-tocopherol) was determined by adapted Arnaud et al. method [18]. Complete methodology is detailed and described in S1 text section.
2.7. Determination of Short Chain Fatty Acids in Feces
The determination of short-chain fatty acids in feces was performed based on the methodology of Zhao, Nyman and Jonsson (2006) [19] with adaptations. Complete methodology is detailed and described in S1 text section.
2.8. Statistical Analysis
One-way analysis of variance (ANOVA) was applied to the data of the various groups, with the Tukey post-test, using the GraphPad Prism software, version 5.00 for Windows (GraphPad Software, San Diego, CA, USA), with the level of significance set up at p < 0.05. Data are reported as mean ± standard deviation.
3. Results
3.1. Effects of FOS on Body Weight Gain and Energy Intake
It was verified, in the first month, higher caloric intake by the animals in the three groups that received a high-fat diet compared to the normolipid control group, and supplementation with fiber or FOS did not promote changes in caloric intake within the groups with similar lipid content. In the other months, no statistically significant difference was found between the monthly caloric intake of each experimental group (Table 2). In the first experimental month, the higher caloric intake of the animals that received a high-fat diet promoted greater weight gain in the HL group compared to the other groups, and supplementation with fiber and FOS reduced the weight gain of the animals that consumed a high-fat diet. Although no statistically significant difference was verified in the caloric intake of the animals in the second, third and fourth months, it was verified in the weight evolution (Figure 1) that from the fifth experimental week onwards, the HL and HLF groups presented significantly higher weights than the other groups, and on the last day of the experiment the weight of the HLFOS group did not show statistical difference in relation to the three groups that received a normolipid diet (Table 2).
3.2. Liver Weight, Epididymal and Retroperitoneal Adipose Tissue Weight
Although the FOS-enriched high-fat diet provided lower body weight and reduction of retroperitoneal adipose tissue compared to the HL and HLF groups, there was no statistically significant difference in relation to the weight of epididymal adipose tissue and the sum of adipose tissues analyzed between these groups (Table 3). Although no statistically significant difference was verified between the groups in relation to the liver dosage of total fat or cholesterol (Table 3), it was verified that the supply of FOS in the high-fat diet promoted lower liver weight (Table 3) and reduction the degree of hepatic steatosis (Figure 2). The presence of inflammatory infiltrate or Mallory's bodies was not identified in the histopathological analysis of liver slides.
3.3. Effects of FOS on Triacylglycerol, Cholesterol, VLDL, HDL-C, Glycemia, and Triacylglycerol/HDL-Cholesterol Ratio
Regarding serum levels, there was no statistically significant difference in AST and triglyceride values, as well as in the AST/ALT ratio. It was found that the high-fat diet promoted higher fasting glycemia compared to normolipid diets, with no statistical differences being observed with fiber or FOS supplementation. The ALT dosage was significantly higher in the HL and HLF groups compared to the C and F groups, and the supplementation with FOS in the high fat diet promoted a reduction in the ALT value so that there was no statistical difference when compared to the animals that received normolipid diet. Finally, serum cholesterol dosages were also significantly higher in the HL and HLF groups, and supplementation of the high-fat diet with FOS reduced serum cholesterol so that there was no difference between this and the F group (Table 4).
3.4. Effects of FOS on Oxidative Stress Parameters
There was a significant increase in MDA in the HLF group when compared to the other groups that received a normolipid diet. Regarding hepatic antioxidant parameters, no statistically significant difference was observed in retinol dosages, while in α-tocopherol dosages it was verified that the high-fat diet promoted a reduction of this antioxidant in the liver, and the supply of FOS promoted a tendency to increase this parameter in animals that ate a high fat diet, but without statistical significance. Finally, supplementation with microcrystalline cellulose and FOS significantly increased hepatic α-tocopherol in the groups that received a normolipid diet, with the highest value observed in the F group compared to the other experimental groups. There was no statistically significant difference in TAC (Table 5).
3.5. Effects on Enteric Muscle Thickness and Intestinal Lumen Diameter
The histological analysis of the colon verified that the largest total diameter of the intestinal lumen occurred in the HL group, followed by the HLF group and then by the HLFOS, and the smallest diameter value was found in the animals of the C and FOS groups, which did not present statistical difference each other. Intestinal muscle thickness, on the other hand, showed a statistically significant difference between all groups, with the smallest thickness being found in the HL group, followed by the HLF and then by the F; FOS supplementation increased the thickness of the intestinal musculature in animals that received a high-fat diet, but reduced it in those that consumed a normolipid diet (Table 6).
3.6. Effects of FOS on Short-chain Fatty Acids in Feces
The analysis of short-chain fatty acids in feces did not find statistically significant differences in the analyzes of propionic, butyric and valeric acids. However, it was found that microcrystalline cellulose supplementation had opposite effects on acetic acid concentration depending on the lipid content of the diet. Regarding isobutyric and isovaleric acids, it was observed that the HLFOS group had significantly lower concentrations compared to group F, and in the case of isobutyric this difference extended to group C (Table 7).
4. Discussion
Although NAFLD, which has no specific medical treatment, is not part of the diagnostic criteria for MS, its presence increases the risk of a cardiovascular event in these individuals [20,21]. Many studies have suggested that NAFLD progression can be inhibited with the use of prebiotics, probiotics and symbiotics [22,23,24], but the mechanisms of the effects of FOS on body composition, metabolic parameters, production of short-chain fatty acids in feces and NAFLD need further understanding. The present study evaluated the long-term effects of 15% FOS supplementation compared to 15% microcrystalline cellulose in experimental animal models with obesity and metabolic syndrome induced by a high-fat diet, as well as in animals receiving a normolipid diet. The findings indicate that FOSs promoted a reduction in body weight, retroperitoneal adipose tissue and serum cholesterol and prevented NAFLD in C57BL mice with obesity and metabolic syndrome induced by a high-fat diet.
Despite the fact that supplementation with FOS did not interfere with the monthly caloric intake regardless of the lipid content of the diet, from the fifth experimental week onwards, a lower weight of the animals that received a high-fat diet with FOS (HLFOS) was verified, and at the end of the eighteen weeks the weight of the HLFOS group was equal to that of the groups that received a normolipid diet, which was not observed with the diet enriched with microcrystalline cellulose. In addition, there was a reduction in the weight of the retroperitoneal adipose tissue in the HLFOS group compared to the other groups that received a high-fat diet. Such results are similar to the study by Mao et al.[25], who verified a reduction in body weight in mice receiving a normolipid diet rich in FOS (25%), but the same was not observed in animals that received only 5% of this prebiotic.
The benefits of FOS in reducing weight gain may be due to the production of SCFA from intestinal bacteria, as Lu et al (2016)[26] found that the long-term offer of a high-fat diet supplemented with 5% of the main SCFA, alone or mixed, stimulated beige adipogenesis, increasing fat oxidation and energy expenditure mediated by GPR43 and GPR41 activation. In addition, SCFA are also related to increased plasma concentration of PYY (Peptide YY), which is an intestinal peptide hormone secreted in the postprandial period and which decreases intestinal motility and exerts anorexigenic effects in eutrophic and with obesity individuals. [27,28]. The divergence of findings regarding the effects of FOS on food intake and body composition can be explained by the different methodologies found, whether in relation to the duration of the experiment, the type of diet or the amount of FOS.
Animals that received a high-fat diet had higher liver weight, presence of hepatic steatosis and increased ALT, so that long-term FOS supply prevented the development of NAFLD and reduced liver weight and ALT levels, while the same was not observed in animals that received a high-fat diet with high doses of cellulose. These results were consistent with those found by Matsumoto et al. (2017) [29] in C57BL mice with NAFLD induced by a choline-deficient diet, in which the supply of 5% FOS for 3 weeks improved liver parameters with a reduction in ALT levels, as well as in the degree of steatosis, in the presence inflammation and hepatic ballooning. Prevention of NAFLD was associated with decreased intestinal permeability and SCFA production by intestinal bacteria.
In our study, it was expected that the offer of FOS would improve fasting glycemia and serum triglycerides in experimental animal models of obesity and MS, but there was no statistically significant difference in these analyses, with only a reduction in serum cholesterol being observed in the HLFOS group compared to to the HL and the HLF. These findings were consistent with Mao et al. (2018) [25], whose supply of FOS in a normolipid diet did not interfere with the analyzed serum biochemical parameters, even with the use of high amounts of this prebiotic.
Considering that the oxidative stress promoted by the increase of fatty acids in the liver is an important factor in the pathogenesis of non-alcoholic steatohepatitis resulting from NASH, it is necessary to develop therapeutic targets that can act in this way [30]. The present study did not find alteration in the total antioxidant capacity between the groups, and the supply of FOS did not interfere in the dosages of hepatic MDA and retinol, regardless of the lipid content of the diet. Based on these findings, it cannot be concluded that the supply of 15% FOS, in the long term, improved the antioxidant system in experimental animal models of obesity and MS induced by a high-fat diet.
Histological analysis of the colon showed that the high-fat diet increased the diameter of the intestinal lumen and reduced the enteric muscle thickness, whereas the supplementation with fructooligosaccharides was able to reverse these changes. Although the supply of FOS in the normolipid diet did not change the diameter of the intestinal lumen compared to group C, a reduction in the enteric musculature was observed. Wedel et al (2006)[31] demonstrated that severe colorectal motility disorders, such as idiopathic megacolon and slow-transit constipation, are associated with deficient expression of proteins linked to intestinal smooth muscle contraction. In a review study, it was found that both an increase and a decrease in smooth muscle contractility was present in intestinal inflammation, and the functional deficiency of smooth muscle cells can occur due to changes in the activities of muscarinic receptors and ion channels [32]. Thus, the evidence that fermentable dietary fibers influence enteric muscle thickness and intestinal lumen diameter suggests that changes in gastrointestinal health do not only involve the modulation of intestinal microbiota and mucosal permeability.
The prebiotic effect of FOS is responsible for the modulation of the intestinal microbiota and the modifications in SCFA, and currently the modulation of the intestinal microbiota appears to be a promising direction for the treatment of NASH[33,34,35]. The production of SCFA by intestinal bacteria plays an important role in liver health, as on the one hand it contributes to increased intestinal absorption and, consequently, caloric intake, and on the other hand it suppresses colon inflammation via activation of GPR43, protecting the liver against toxic components coming from the portal vein [36,37]. Furthermore, the benefits of FOS in non-alcoholic hepatic steatosis would be related to its prebiotic properties, which can be confirmed with the results of the present study, since NASH induced by the high-fat diet was only prevented by offering FOS, while that supplementation with microcrystalline cellulose, which does not present prebiotic characteristics in monogastric animals, did not have the same effects.
In the present study, the supply of high doses of FOS showed significant benefits in experimental animal models of obesity and MS induced by a high-fat diet, but the same was not observed in the groups that received a normolipid diet with FOS. Furthermore, no signs of liver toxicity or worsening of metabolic parameters resulting from the use of high doses of fructooligosaccharides were found, indicating that the amount used in the long term does not seem to trigger possible harm to the health of the animal models in this experiment. Thus, the present study contributes to the development of new research with FOS, either to explore the mechanisms by which this prebiotic exerts different effects according to the lipid content of the diet, or to invest in experimental protocols that define therapeutic amounts of fructooligosaccharides in prevention and treatment of obesity, MS and NAFLD.
5. Conclusions
In summary, the long-term supply of 15% FOS promoted a reduction in body weight, retroperitoneal adipose tissue and serum cholesterol and prevented NAFLD in C57BL mice with obesity and MS induced by a high-fat diet. The alterations found in the intestinal histology suggest that the benefits of FOS in gastrointestinal health are not restricted to the modulation of the microbiota, reinforcing the importance of new studies that investigate the effects of this prebiotic at the local level, correlating them with peripheral alterations.
Data Availability Statement
The data used to support this study are available from the corresponding author upon request.
Acknowledgments
This study was supported by grant from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), No. 88887.878134/2023-00.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Younossi Z: Anstee QM, Marietti M, Hardy T, Henry L, Eslam M, et al. Global burden of NAFLD and NASH: trends, predictions, risk factors and prevention. Nat Rev Gastroenterol Hepatol [Internet]. 2018 Jan;15(1):11–20. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28930295.
- Younossi ZM, Koenig AB, Abdelatif D, Fazel Y, Henry L, Wymer M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology [Internet]. 2016 Jul;64(1):73–84. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26707365.
- Polyzos SA, Kountouras J, Mantzoros CS. Obesity and nonalcoholic fatty liver disease: From pathophysiology to therapeutics. Metabolism [Internet]. 2019 Mar;92:82–97. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0026049518302531. [CrossRef]
- Younossi ZM, Golabi P, de Avila L, Paik JM, Srishord M, Fukui N, et al. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: A systematic review and meta-analysis. J Hepatol [Internet]. 2019 Oct;71(4):793–801. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0168827819303939. [CrossRef]
- Arab JP, Karpen SJ, Dawson PA, Arrese M, Trauner M. Bile acids and nonalcoholic fatty liver disease: Molecular insights and therapeutic perspectives. Hepatology [Internet]. 2017 Jan 4;65(1):350–62. Available from: https://onlinelibrary.wiley.com/doi/10.1002/hep.28709. [CrossRef]
- Sociedade Brasileira de Cardiologia. I Diretriz Brasileira de Diagnóstico e Tratamento da Síndrome Metabólica. Vol. 84, Arquivos Brasileiros de Cardiologia. Brasil; 2005.
- Alberti KGMM, Eckel RH, Grundy SM, Zimmet PZ, Cleeman JI, Donato KA, et al. Harmonizing the metabolic syndrome: a joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International. Circulation [Internet]. 2009 Oct 20;120(16):1640–5. Available from: http://www.ncbi.nlm.nih.gov/pubmed/19805654.
- Canfora EE, Meex RCR, Venema K, Blaak EE. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat Rev Endocrinol [Internet]. 2019 May;15(5):261–73. Available from: http://www.ncbi.nlm.nih.gov/pubmed/30670819. [CrossRef]
- Aron-Wisnewsky J, Vigliotti C, Witjes J, Le P, Holleboom AG, Verheij J, et al. Gut microbiota and human NAFLD: disentangling microbial signatures from metabolic disorders. Nat Rev Gastroenterol Hepatol [Internet]. 2020 May;17(5):279–97. Available from: http://www.ncbi.nlm.nih.gov/pubmed/32152478. [CrossRef]
- Huang X, Chen Q, Fan Y, Yang R, Gong G, Yan C, et al. Fructooligosaccharides attenuate non-alcoholic fatty liver disease by remodeling gut microbiota and association with lipid metabolism. Biomed Pharmacother [Internet]. 2023 Mar;159:114300. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0753332223000884. [CrossRef]
- Guarino M, Altomare A, Emerenziani S, Di Rosa C, Ribolsi M, Balestrieri P, et al. Mechanisms of Action of Prebiotics and Their Effects on Gastro-Intestinal Disorders in Adults. Nutrients [Internet]. 2020 Apr 9;12(4):1037. Available from: https://www.mdpi.com/2072-6643/12/4/1037. [CrossRef]
- Reeves PG, Nielsen FH, Fahey GC. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J Nutr [Internet]. 1993 Nov;123(11):1939–51. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0022316623056341. [CrossRef]
- Gurmini J, Cecílio WAC, Schuler SL, Olandoski M, Noronha L de. Desnutrição intra-uterina e suas alterações no intestino delgado de ratos Wistar ao nascimento e após a lactação. J Bras Patol e Med Lab [Internet]. 2005 Aug;41(4):271–8. Available from: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1676-24442005000400009&lng=pt&nrm=iso&tlng=pt.
- Oh SI, Kim CI, Chun HJ, Park SC. Chronic Ethanol Consumption Affects Glutathione Status in Rat Liver , ,. J Nutr [Internet]. 1998 Apr;128(4):758–63. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0022316623017996. [CrossRef]
- Bligh EG, Dyer WJ. A RAPID METHOD OF TOTAL LIPID EXTRACTION AND PURIFICATION. Can J Biochem Physiol [Internet]. 1959 Aug 1;37(8):911–7. Available from: http://www.nrcresearchpress.com/doi/10.1139/o59-099. [CrossRef]
- Erel O. A novel automated direct measurement method for total antioxidant capacity using a new generation, more stable ABTS radical cation. Clin Biochem [Internet]. 2004 Apr;37(4):277–85. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0009912003002315.
- Gérard-Monnier D, Erdelmeier I, Régnard K, Moze-Henry N, Yadan JC, Chaudière J. Reactions of 1-Methyl-2-phenylindole with Malondialdehyde and 4-Hydroxyalkenals. Analytical Applications to a Colorimetric Assay of Lipid Peroxidation. Chem Res Toxicol [Internet]. 1998 Oct 1;11(10):1176–83. Available from: https://pubs.acs.org/doi/10.1021/tx9701790. [CrossRef]
- Arnaud J, Fortis I, Blachier S, Kia D, Favier A. Simultaneous determination of retinol, α-tocopherol and β-carotene in serum by isocratic high-performance liquid chromatography. J Chromatogr B Biomed Sci Appl [Internet]. 1991 Dec;572(1–2):103–16. Available from: https://linkinghub.elsevier.com/retrieve/pii/037843479180476S.
- Zhao G, Nyman M, Åke Jönsson J. Rapid determination of short-chain fatty acids in colonic contents and faeces of humans and rats by acidified water-extraction and direct-injection gas chromatography. Biomed Chromatogr [Internet]. 2006 Aug;20(8):674–82. Available from: https://onlinelibrary.wiley.com/doi/10.1002/bmc.580. [CrossRef]
- Du F, Huang R, Lin D, Wang Y, Yang X, Huang X, et al. Resveratrol Improves Liver Steatosis and Insulin Resistance in Non-alcoholic Fatty Liver Disease in Association With the Gut Microbiota. Front Microbiol [Internet]. 2021 Feb 23;12. Available from: https://www.frontiersin.org/articles/10.3389/fmicb.2021.611323/full. [CrossRef]
- Luna-Luna M, Medina-Urrutia A, Vargas-Alarcón G, Coss-Rovirosa F, Vargas-Barrón J, Pérez-Méndez Ó. Adipose Tissue in Metabolic Syndrome: Onset and Progression of Atherosclerosis. Arch Med Res [Internet]. 2015 Jul;46(5):392–407. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0188440915001253. [CrossRef]
- Carpi RZ, Barbalho SM, Sloan KP, Laurindo LF, Gonzaga HF, Grippa PC, et al. The Effects of Probiotics, Prebiotics and Synbiotics in Non-Alcoholic Fat Liver Disease (NAFLD) and Non-Alcoholic Steatohepatitis (NASH): A Systematic Review. Int J Mol Sci [Internet]. 2022 Aug 8;23(15):8805. Available from: https://www.mdpi.com/1422-0067/23/15/8805. [CrossRef]
- Scorletti E, Afolabi PR, Miles EA, Smith DE, Almehmadi A, Alshathry A, et al. Synbiotics Alter Fecal Microbiomes, But Not Liver Fat or Fibrosis, in a Randomized Trial of Patients With Nonalcoholic Fatty Liver Disease. Gastroenterology [Internet]. 2020 May;158(6):1597-1610.e7. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0016508520301244. [CrossRef]
- Hernández-Pérez S, Oliart-Ros RM, Casas-Godoy L, Sandoval G, Guarner-Lans V, Castrejón-Téllez V, et al. Beneficial Effects of Fructooligosaccharides Esterified with Lauric Acid in a Metabolic Syndrome Model Induced by a High-Fat and High-Carbohydrate Diet in Wistar Rats. J Med Food [Internet]. 2022 Aug 1;25(8):828–35. Available from: https://www.liebertpub.com/doi/10.1089/jmf.2021.0109. [CrossRef]
- Mao B, Gu J, Li D, Cui S, Zhao J, Zhang H, et al. Effects of Different Doses of Fructooligosaccharides (FOS) on the Composition of Mice Fecal Microbiota, Especially the Bifidobacterium Composition. Nutrients [Internet]. 2018 Aug 16;10(8):1105. Available from: http://www.mdpi.com/2072-6643/10/8/1105. [CrossRef]
- Lu Y, Fan C, Li P, Lu Y, Chang X, Qi K. Short Chain Fatty Acids Prevent High-fat-diet-induced Obesity in Mice by Regulating G Protein-coupled Receptors and Gut Microbiota. Sci Rep [Internet]. 2016 Nov 28;6(1):37589. Available from: https://www.nature.com/articles/srep37589. [CrossRef]
- Wang SZ, Yu YJ, Adeli K. Role of Gut Microbiota in Neuroendocrine Regulation of Carbohydrate and Lipid Metabolism via the Microbiota-Gut-Brain-Liver Axis. Microorganisms [Internet]. 2020 Apr 7;8(4). Available from: http://www.ncbi.nlm.nih.gov/pubmed/32272588. [CrossRef]
- Nishida A, Miyamoto J, Shimizu H, Kimura I. Gut microbial short-chain fatty acids-mediated olfactory receptor 78 stimulation promotes anorexigenic gut hormone peptide YY secretion in mice. Biochem Biophys Res Commun [Internet]. 2021 Jun;557:48–54. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0006291X21005805. [CrossRef]
- Matsumoto K, Ichimura M, Tsuneyama K, Moritoki Y, Tsunashima H, Omagari K, et al. Fructo-oligosaccharides and intestinal barrier function in a methionine–choline-deficient mouse model of nonalcoholic steatohepatitis. Strnad P, editor. PLoS One [Internet]. 2017 Jun 20;12(6):e0175406. Available from:. [CrossRef]
- Powell EE, Wong VWS, Rinella M. Non-alcoholic fatty liver disease. Lancet [Internet]. 2021 Jun;397(10290):2212–24. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0140673620325113.
- Wedel T, Van Eys GJ, Waltregny D, Glénisson W, Castronovo V VJ. Novel smooth muscle markers reveal abnormalities of the intestinal musculature in severe colorectal motility disorders. Neurogastroenterol Motil. 2006;18(7):526–38. [CrossRef]
- Ohama T, Hori M, Ozaki H. Mechanism of abnormal intestinal motility in inflammatory bowel disease: how smooth muscle contraction is reduced? J Smooth Muscle Res [Internet]. 2007;43(2):43–54. Available from: http://www.jstage.jst.go.jp/article/jsmr/43/2/43_2_43/_article.
- Tilg H, Adolph TE, Dudek M, Knolle P. Non-alcoholic fatty liver disease: the interplay between metabolism, microbes and immunity. Nat Metab [Internet]. 2021 Dec 20;3(12):1596–607. Available from: https://www.nature.com/articles/s42255-021-00501-9. [CrossRef]
- Jensen T, Abdelmalek MF, Sullivan S, Nadeau KJ, Green M, Roncal C, et al. Fructose and sugar: A major mediator of non-alcoholic fatty liver disease. J Hepatol [Internet]. 2018 May;68(5):1063–75. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0168827818300667. [CrossRef]
- Ji Y, Yin Y, Li Z, Zhang W. Gut Microbiota-Derived Components and Metabolites in the Progression of Non-Alcoholic Fatty Liver Disease (NAFLD). Nutrients [Internet]. 2019 Jul 25;11(8):1712. Available from: https://www.mdpi.com/2072-6643/11/8/1712. [CrossRef]
- Ziętek M, Celewicz Z, Szczuko M. Short-Chain Fatty Acids, Maternal Microbiota and Metabolism in Pregnancy. Nutrients [Internet]. 2021 Apr 9;13(4):1244. Available from: https://www.mdpi.com/2072-6643/13/4/1244. [CrossRef]
- Zhu L, Baker RD, Baker SS. Gut microbiome and nonalcoholic fatty liver diseases. Pediatr Res [Internet]. 2015 Jan 13;77(1–2):245–51. Available from: https://www.nature.com/articles/pr2014157. [CrossRef]
Figure 1.
Weight evolution of the experimental groups expressed as mean and standard deviation over the weeks. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat dietary group with FOS. FOS – fructooligosaccharides.
Figure 1.
Weight evolution of the experimental groups expressed as mean and standard deviation over the weeks. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat dietary group with FOS. FOS – fructooligosaccharides.
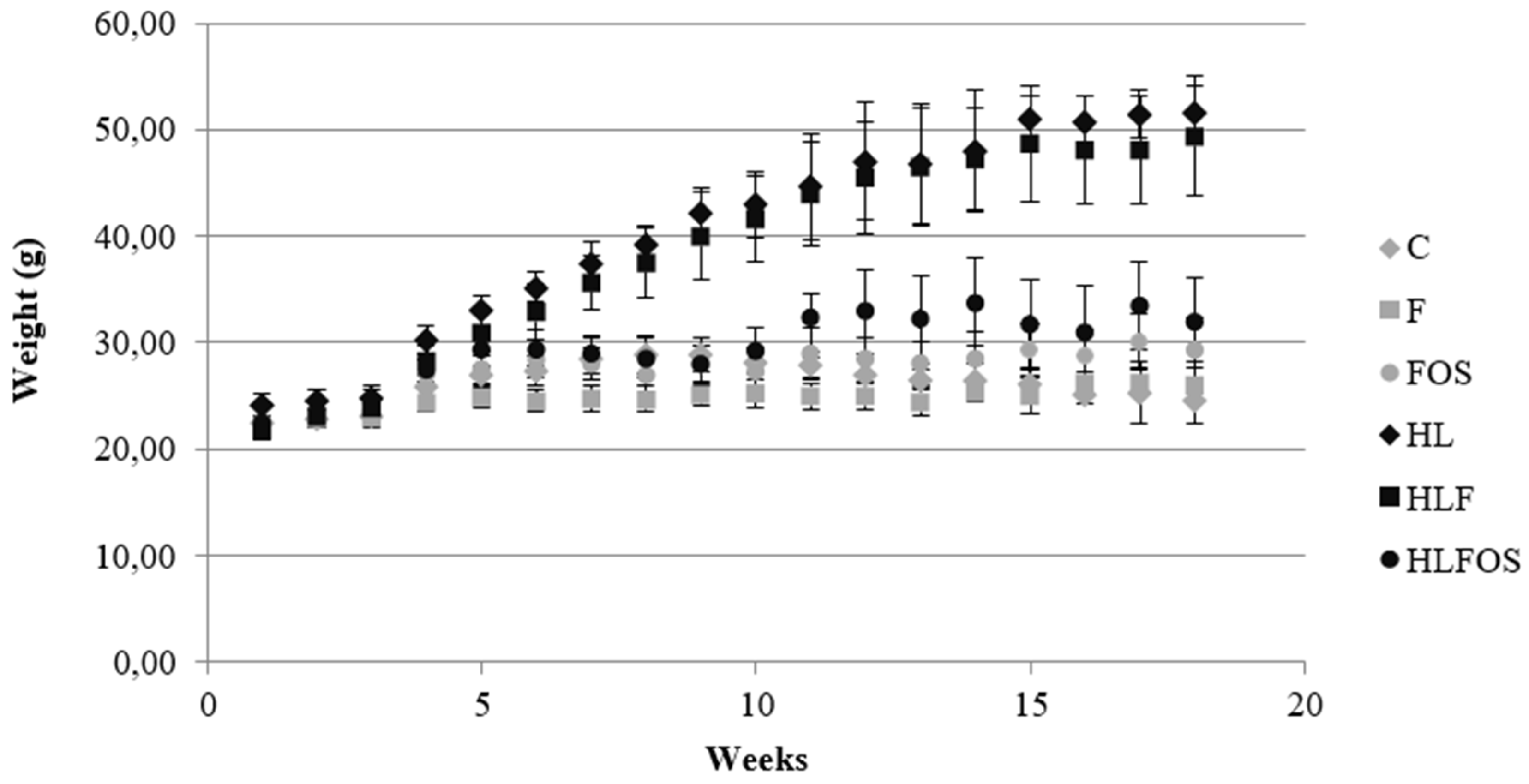
Figure 2.
Analysis of hepatic parameters. Steatosis score (a). Liver photomicrograph (b). A = Control; B = High fiber control; C = Control supplemented with fructooligosaccharides; D = Hyperlipidic control; E = Hyperlipidic rich in fiber; F = Hyperlipid supplemented with fructooligosaccharides. Data are reported as mean ± standard deviation for a period of 18 weeks. *p < 0.0001.
Figure 2.
Analysis of hepatic parameters. Steatosis score (a). Liver photomicrograph (b). A = Control; B = High fiber control; C = Control supplemented with fructooligosaccharides; D = Hyperlipidic control; E = Hyperlipidic rich in fiber; F = Hyperlipid supplemented with fructooligosaccharides. Data are reported as mean ± standard deviation for a period of 18 weeks. *p < 0.0001.
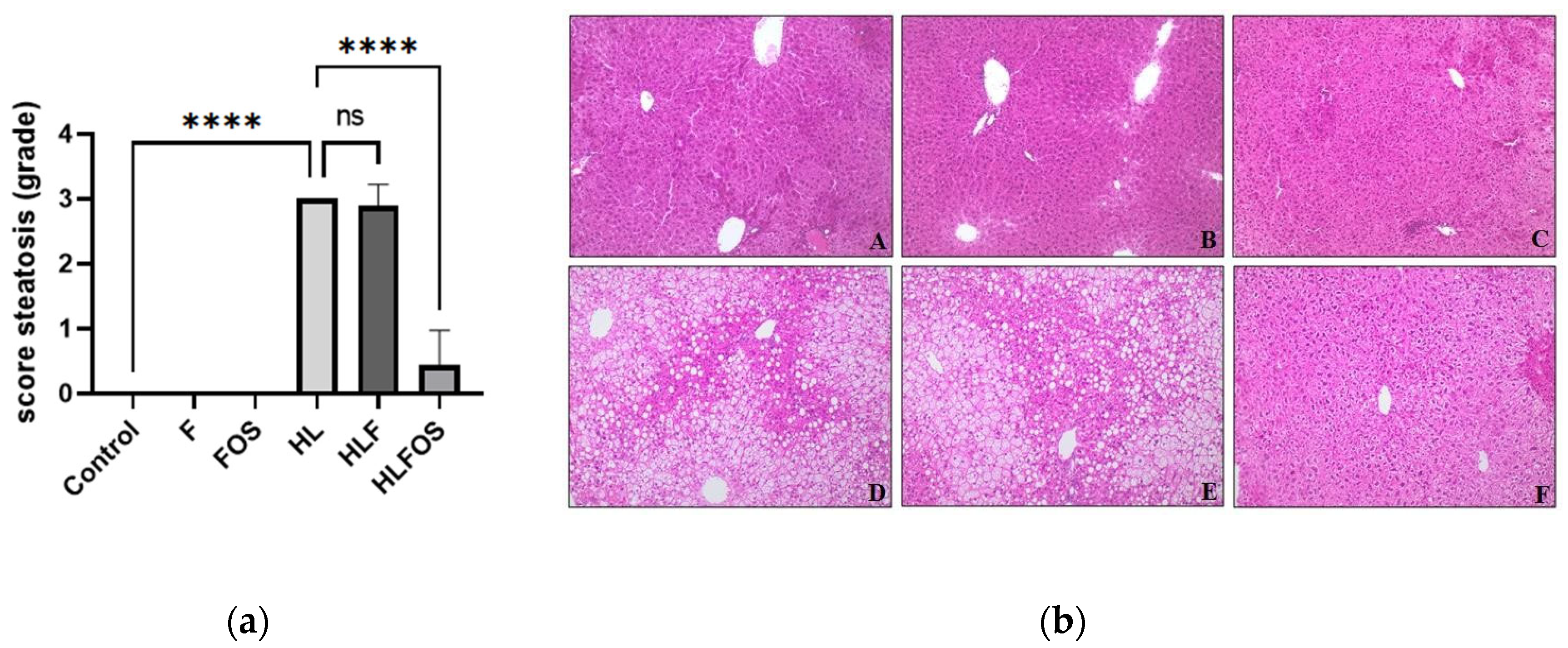

Table 1.
Composition of experimental diets per 100 g.
| Ingredients | C | F | FOS | HL | HLF | HLFOS |
|---|---|---|---|---|---|---|
| Casein (g) | 20 | 20 | 20 | 20 | 20 | 20 |
| Corn starch (g) | 63 | 53 | 53 | 25 | 15 | 15 |
| Soybean oil (g) | 7 | 7 | 7 | 0 | 0 | 0 |
| Lard | 0 | 0 | 0 | 45 | 45 | 45 |
| FOS (g) | 0 | 0 | 15 | 0 | 0 | 15 |
| Fiber (g) | 5 | 15 | 0 | 5 | 15 | 0 |
| Mineral mix (g) | 3,5 | 3,5 | 3,5 | 3,5 | 3,5 | 3,5 |
| Vitamins (g) | 1 | 1 | 1 | 1 | 1 | 1 |
| L-cysteine (g) | 0,3 | 0,3 | 0,3 | 0,3 | 0,3 | 0,3 |
| Choline (g) | 0,25 | 0,25 | 0,25 | 0,25 | 0,25 | 0,25 |
| BHT (mg) | 1,4 | 1,4 | 1,4 | 1,4 | 1,4 | 1,4 |
| Calories | 415 | 415 | 415 | 605 | 605 | 605 |
C – control group; CF – normolipid rich in fiber group; FOS – normolipid supplemented with FOS group; HL – high fat group; HLF – high fat with high fiber group; HLFOS – high fat with FOS group. FOS – fructooligosaccharides. BHT - butylated hydroxytoluene.
Table 2.
Comparison of the average monthly caloric intake per animal of each experimental group and comparison of the weight of each experimental group during the four months.
Table 2.
Comparison of the average monthly caloric intake per animal of each experimental group and comparison of the weight of each experimental group during the four months.
| C | F | FOS | HL | HLF | HLFOS | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Weight (g) | Caloric intake (kcal) | Weight (g) | Caloric intake (kcal) | Weight (g) | Caloric intake (kcal) | Weight (g) | Caloric intake (kcal) | Weight (g) | Caloric intake (kcal) | Weight (g) | Caloric intake (kcal) | |
| 1st month | 24,72 ± 0,67 a,c | 83,71 ± 16,55 a | 23,77 ± 0,68 a | 94,31 ± 18,39 a,b | 25,66 ± 1,20 c,d | 111,91 ± 30,46 a,b,c | 28,17 ± 0,72 b | 134,23 ± 32,50 b,c | 26,55 ± 0,86 d | 139,53 ± 29,28 c | 26,03 ± 1,03 c,d | 131,02 ± 31,00 b,c |
| 2nd month | 28,41 ± 1,60 a | 109,54 ± 20,84 | 24,77 ± 1,00 b | 123,68 ± 20,06 | 27,92 ± 1,67 a | 107,73 ± 23,69 | 38,48 ± 1,85 c | 110,98 ± 34,09 | 36,50 ± 3,08 c | 119,87 ± 38,00 | 28,72 ± 1,74 a | 86,59 ± 6,94 |
| 3rd month | 27,38 ± 1,15 a,c | 89,83 ± 25,01 | 24,92 ± 1,19 a | 95,87 ± 20,99 | 28,23 ± 2,12 a,c | 111,09 ± 26,50 | 45,38 ± 4,70 b | 106,44 ± 32,00 | 44,41 ± 4,84 b | 110,41 ± 14,86 | 31,72 ± 3,38 c | 104,36 ± 23,14 |
| 4th month | 25,72 ± 1,44 a | 87,49 ± 18,05 | 25,73 ± 1,17 a | 89,94 ± 18,44 | 29,17 ± 2,49 a,c | 96,60 ± 39,55 | 48,78 ± 5,92 b | 99,07 ± 29,38 | 48,05 ± 5,07 b | 97,25 ± 16,10 | 33,14 ± 4,36 c | 97,00 ± 10,51 |
a,b,c Values followed by different letters in the same line indicate a statistically significant difference with p<0.05. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat diet with FOS group. FOS – fructooligosaccharides.
Table 3.
Comparison of liver and adipose tissue weight of each experimental group and liver lipid profile.
Table 3.
Comparison of liver and adipose tissue weight of each experimental group and liver lipid profile.
| C | F | FOS | HL | HLF | HLFOS | |
|---|---|---|---|---|---|---|
| Liver weight (g) | 0,93 ± 0,23 a | 0,96 ± 0,28 a | 1,14 ± 0,29 a | 2,24 ± 0,71 b | 1,84 ± 0,58 b | 1,12 ± 0,15 a |
| Epididymal adipose tissue (g) | 0,27 ± 0,13 a | 0,36 ± 0,12 a | 0,54 ± 0,27 a | 1,46 ± 0,33 b | 1,51 ± 0,46 b | 1,34 ± 0,67 b |
| Retroperitoneal adipose tissue (g) | 0,07 ± 0,05 a | 0,10 ± 0,06 a | 0,17 ± 0,14 a,c | 1,02 ± 0,42 b | 1,08 ± 0,41 b | 0,54 ± 0,35 c |
| Liver weight/Body weight Ratio | 3,50 ± 0,55 a | 3,49 ± 0,68 a | 3,73 ± 0,77 a,b | 4,53 ± 1,07 b | 3,86 ± 0,77 a,b | 3,52 ± 0,50 a,b |
| Adipose tissue sum (g) | 0,34 ± 0,18 a | 0,46 ± 0,16 a | 0,71 ± 0,41 a | 2,48 ± 0,63 b | 2,59 ± 0,71 b | 1,88 ± 1,02 b |
| Ratio of the sum of adipose tissues/Body weight | 1,32 ± 0,74 a | 1,73 ± 0,58 a | 2,26 ± 1,08 a | 5,14 ± 1,14 b | 5,55 ± 1,35 b | 5,88 ± 3,04 b |
| Total fat (g/g Tissue) | 0,37 ± 0,04 | 0,43 ± 0,20 | 0,36 ± 0,05 | 0,53 ± 0,22 | 0,39 ± 0,06 | 0,39 ± 0,03 |
| Total cholesterol (mg/g total fat) | 9,47 ± 1,53 | 7,70 ± 2,36 | 8,18 ± 2,05 | 8,62 ± 0,88 | 8,41 ± 1,26 | 7,70 ± 1,06 |
| Triglycerides (mg/g total fat) | 72,81 ± 54,06 a,b | 33,05 ± 43,16 a | 39,14 ± 20,44 a | 184,76 ± 104,33 b | 130,24 ± 85,93 a,b | 77,59 ± 36,89 a,b |
a,b,c Values followed by different letters in the same line indicate a statistically significant difference with p<0.05. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat diet with FOS group. FOS – fructooligosaccharides.ferent letters in the same line indicate a statistically significant difference with p<0.05. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat diet with FOS group. FOS – fructooligosaccharides.
Table 4.
Comparison of serum levels of fasting glucose, aspartate aminotransferase (AST), alanine aminotransferase (ALT), cholesterol and triglycerides according to the experimental group.
Table 4.
Comparison of serum levels of fasting glucose, aspartate aminotransferase (AST), alanine aminotransferase (ALT), cholesterol and triglycerides according to the experimental group.
| C | F | FOS | HL | HLF | HLFOS | |
|---|---|---|---|---|---|---|
| Fasting glycemia (mg/dL) | 124,36 ± 21,17 a | 117,00 ± 15,42 a | 131,00 ± 27,16 a | 189,89 ± 29,42 b | 177,90 ± 22,06 b | 170,00 ± 35,00 b |
| AST (U/mL) | 84,73 ± 32,48 | 70,49 ± 46,16 | 81,57 ± 25,44 | 80,71 ± 18,27 | 96,17 ± 15,92 | 75,82 ± 24,78 |
| ALT (U/mL) | 23,66 ± 5,74 a | 22,08 ± 4,72 a | 24,60 ± 4,44 a,c | 38,80 ± 15,87 b | 36,23 ± 12,88 b,c | 25,08 ± 6,27 a,c |
| AST/ALT ratio | 3,61 ± 1,17 | 3,13 ± 1,70 | 3,34 ± 0,90 | 2,34 ± 0,87 | 2,91 ± 0,94 | 3,03 ± 0,59 |
| Serum cholesterol (mg/dL) | 125,03 ± 28,90 a | 133,33 ± 11,05 a,c | 126,70 ± 30,02 a | 284,88 ± 96,44 b | 272,29 ± 62,57 b | 202,17 ± 24,76 c |
| Serum triacylglycerol (mg/dL) | 85,24 ± 27,67 | 96,77 ± 34,71 | 82,95 ± 14,58 | 92,10 ± 29,84 | 99,09 ± 18,63 | 99,48 ± 18,44 |
a,b,c Values followed by different letters in the same line indicate a statistically significant difference with p<0.05. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat diet with FOS group. FOS – fructooligosaccharides.
Table 5.
Liver analysis of malondialdehyde (MDA), retinol, α-tocopherol and total antioxidant capacity (TAC) according to the experimental group.
Table 5.
Liver analysis of malondialdehyde (MDA), retinol, α-tocopherol and total antioxidant capacity (TAC) according to the experimental group.
| C | F | FOS | HL | HLF | HLFOS | |
|---|---|---|---|---|---|---|
| MDA (µmol/g protein) | 1,65 ± 0,56 a | 1,65 ± 0,25 a | 1,42 ± 0,34 a | 2,10 ± 0,69 a,b | 2,65 ± 0,82 b | 1,90 ± 0,57 a,b |
| α-Tocopherol (nmol/ g tissue) | 101,69 ± 46,16 a | 218,97 ± 42,65 b | 160,13 ± 57,04 c | 34,33 ± 31,63 d | 28,81 ± 24,29 d | 90,60 ± 18,48 a,d |
| Retinol (nmol/ g tissue) | 71,35 ± 41,99 | 58,30 ± 28,19 | 86,77 ± 55,02 | 42,05 ± 10,98 | 44,90 ± 16,57 | 63,37 ± 20,04 |
| α-Tocopherol/ MDA ratio | 0,33 ± 0,13 a,c | 1,05 ± 0,51 b | 0,59 ± 0,23 a | 0,12 ± 0,12 c | 0,11 ± 0,10 c | 0,35 ± 0,13 a,c |
| TAC (µM) | 2,26 ± 0,12 | 2,19 ± 0,05 | 2,25 ± 0,08 | 2,24 ± 0,09 | 2,22 ± 0,11 | 2,20 ± 0,13 |
a,b,c Values followed by different letters in the same line indicate a statistically significant difference with p<0.05. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat diet with FOS group. FOS – fructooligosaccharides.
Table 6.
Comparison of the total diameter of the intestinal lumen and intestinal muscle thickness according to the experimental group.
Table 6.
Comparison of the total diameter of the intestinal lumen and intestinal muscle thickness according to the experimental group.
| C | F | FOS | HL | HLF | HLFOS | |
|---|---|---|---|---|---|---|
| Total diameter of the intestinal lumen (µm) | 555,22 ± 16,94 a | 618,04 ± 20,43 b | 554,67 ± 8,48 a | 846,81 ± 7,82 c | 718,46 ± 6,78 d | 668,37 ± 10,60 e |
| Intestinal muscle thickness (µm) | 59,04 ± 1,80 a | 28,58 ± 1,33 b | 54,76 ± 1,46 c | 8,51 ± 0,51 d | 23,32 ± 1,44 e | 41,70 ± 0,72 f |
a,b,c,d,e,f Values followed by different letters in the same line indicate a statistically significant difference with p<0.05. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat diet with FOS group. FOS – fructooligosaccharides.
Table 7.
Comparison of concentrations of short-chain fatty acids analyzed in feces according to the experimental group.
Table 7.
Comparison of concentrations of short-chain fatty acids analyzed in feces according to the experimental group.
| C | F | FOS | HL | HLF | HLFOS | |
|---|---|---|---|---|---|---|
| Acetic acid | 61,66 ± 7,75 a,b | 52,03 ± 7,59 a | 57,33 ± 6,11 a,b | 60,87 ± 6,80 a,b | 63,96 ± 3,53 b | 56,77 ± 12,43 a,b |
| Propionic acid | 15,10 ± 2,85 | 14,42 ± 2,14 | 17,27 ± 3,24 | 14,48 ± 3,37 | 14,57 ± 3,13 | 13,46 ± 4,27 |
| Isobutyric acid | 2,07 ± 0,77 a | 2,03 ± 0,79 a | 1,77 ± 0,67 a,b | 1,87 ± 0,64 a,b | 1,63 ± 0,57 a,b | 1,00 ± 0,42 b |
| Butyric acid | 15,83 ± 9,83 | 23,82 ± 9,36 | 17,89 ± 11,40 | 15,92 ± 5,82 | 13,57 ± 4,28 | 23,76 ± 9,57 |
| Isovaleric acid | 3,08 ± 1,24 a,b | 4,25 ± 2,02 a | 3,13 ± 1,30 a,b | 3,80 ± 1,04 a,b | 3,60 ± 0,53 a,b | 2,30 ± 1,03 b |
| Valeric acid | 2,25 ± 0,60 | 3,43 ± 1,44 | 2,60 ± 0,87 | 3,05 ± 0,74 | 2,67 ± 0,54 | 2,70 ± 0,97 |
a,b Values followed by different letters in the same line indicate a statistically significant difference with p<0.05. C – control group; FC – high-fiber normolipid diet group; FOS – normolipid diet with FOS group; HL – high-fat diet group; HLF – fiber-rich high-fat diet group; HLFOS – high-fat diet with FOS group. FOS – fructooligosaccharides.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



