
Submitted:
03 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
Legumes are a diverse and important group of plants that play a vital role in agriculture, food security, and environmental sustainability. Rhizobia are symbiotic bacteria that form nitrogen-fixing nodules on legume roots, providing the plant with a valuable source of nitrogen. Phenolic acids are a group of secondary metabolites produced by plants that have a wide range of biological functions, including defense against pests and diseases, tolerance to abiotic stresses, and nutrient uptake. In the context of climate change and the imperative for sustainable agriculture, this study delves into the dynamic responses of legume species to varying light intensities and their intricate interactions with soil microorganisms. We investigated the impact of light intensity and rhizobial inoculation on the biomass, nitrate reductase, acid phosphatase, and production of phenolic acids in the roots of four legume species, Trifolium repens, Vicia sativa, Ornithopus compressus, and Coronilla juncea. Plants were grown under three light intensity regimes (low, medium, and high) and inoculated with either rhizobia or a non-inoculated control. The results highlights that shaded-light-adapted species, T. repens and V. sativa, increased root exudate production when exposed to high light intensity. This response aligns with their mining strategy, effectively allocating resources to optimize nutrient acquisition under varying conditions. In contrast, species hailing from well- illuminated environments, O. compressus and C. juncea, displayed distinct strategies by significantly increasing biomass under high irradiance, capitalizing on the available light and nutrients. The mining strategy of legumes emerged as a central theme, influencing biomass production, nitrogen dynamics, and enzymatic activities. The strong correlations between biomass and total nitrogen accumulation underscore the role of the mining strategy in efficient nutrient acquisition. Inoculated plants, which rely more on biological nitrogen fixation (BNF), exhibited lower δ15N values, indicative of a successful mining strategy to acquire and utilize atmospheric nitrogen. Enzymatic activities and phenolic acids exhibited significant interspecies variations, reflecting the adaptability of legumes to different light conditions. The findings of this study could be used to develop new strategies for improving legume stress tolerance, nutrient uptake capacity, and rhizosphere health.
Keywords:
legumes
; light availability
; symbionts
; nitrate reductase
; acid phosphatase
; phenolic acid
1. Introduction
Light is a fundamental factor influencing plant performance and growth. It serves as the primary energy source driving photosynthesis, the vital process through which plants convert light energy into chemical energy stored in the form of sugars and other organic compounds [1]. This process is the foundation of a plant's energy supply, allowing it to synthesize the necessary molecules for growth and development. Moreover, light plays a pivotal role in various stages of a plant's life cycle. From seed germination to seedling development, flowering, and seed production, light cues regulate the timing and progression of these critical events [2]. The availability of light signals to seeds when it's time to germinate, and it guides the young seedlings towards the light source, a phenomenon known as phototropism. This ensures that plants can maximize their exposure to light, thereby optimizing photosynthesis. The quality and quantity of light, as well as its duration, can influence a plant's structure, shape, and size. For instance, the direction and intensity of light affect the orientation and elongation of stems, leaves, and roots [3]. This light-mediated growth adaptation is critical for plants to thrive in diverse environments, from densely shaded forests to open, sun-exposed fields.
Light, often an overlooked environmental factor in soil ecosystems, holds substantial significance in shaping the behavior, physiology, and ecology of soil-borne microorganisms. The influence of light extends to several critical aspects concerning these microorganisms, from their metabolic activities to their community dynamics and their ability to colonize various niches within the soil matrix. Although historically overshadowed by other factors such as moisture and nutrient availability, emerging research has unveiled the profound impact of light on the microbial world beneath our feet [4]. One of the primary facets in which light exerts its influence is through its effects on microbial metabolism. Certain soil microorganisms, particularly cyanobacteria, are adept at photosynthesis, harnessing solar energy for the synthesis of organic compounds [5]. This process not only sustains their own growth but also enriches the surrounding soil environment, which in turn can support the broader microbial community [6].
One way that light influences soilborne microorganisms is through its effect on plant growth. The relationship between light, plant growth, and soilborne microorganisms is a complex and essential aspect of ecosystem functioning. Light is the primary source of energy for photosynthesis that enables plants to convert light energy into chemical energy, producing sugars and other organic compounds. These compounds serve as the primary carbon source for plants, and excess carbon is secreted into the rhizosphere as root exudates [7]. Healthy and actively growing plants are capable of harnessing more light energy and converting it into higher photosynthetic rates, leading to an increased production of organic carbon compounds. These root exudates, released by plants into the soil, consist of various substances, including sugars, amino acids, organic acids, and secondary metabolites [8]. This rich mixture of compounds acts as a nutritional buffet for soilborne microorganisms. It provides the necessary carbon and energy sources for various bacterial, fungal, and archaeal populations inhabiting the rhizosphere [9].
The availability of root exudates can profoundly impact the structure and diversity of soilborne microbial communities. Different plant species release distinct exudate profiles, shaping the composition of microorganisms that colonize plant roots [10]. Additionally, the quality and quantity of root exudates can change in response to light conditions. For example, light intensity and spectral composition affect photosynthesis and subsequently alter root exudate composition [11]. This is in clearly in line with the nutrient mining hypothesis. When resources, such as nutrients, are available in a limited quantity, plants maximize their nutrient uptake to support growth and reproduction [12]. This strategy also implies that under nutrient scarcity, some plants increase the production of enzymes that facilitate the breakdown of organic matter in the soil. By doing so, these plants release nutrients trapped in organic compounds, making them available for uptake by the plant and thus overcoming the lack of nutrients. When light is abundant, plants allocate more carbon to growth, increasing their demand for nutrients to support this growth. Consequently, under conditions of high light, plants tend to extract nutrients from the soil at an accelerated rate, promoting nutrient mining [13].
Light intensity also affects the translocation of photosynthetic products to the roots. Under high light conditions, a greater proportion of assimilated carbon is allocated belowground, fueling root exudation, which contains various compounds, including enzymes, organic acids and sugars [14]. These root exudates serve as an energy source for soil microbial communities, promoting nutrient mineralization and nutrient availability in the rhizosphere [15] as well as inducing the establishment of mutualistic interactions [16]. This, in turn, exacerbates the nutrient mining strategy of plants, as nutrient uptake is facilitated by the enhanced nutrient cycling in the rhizosphere [17]. In contrast, under low light conditions, carbon allocation to growth is limited, leading to reduced nutrient mining and increased resource conservation. Here, plants may prioritize the storage of carbon and nutrients for future growth when light conditions become more favorable [18].
In this study, we aimed to explore the impact of varying light intensities on the behavior of both inoculated and non-inoculated plants. Specifically, we postulated that plants originating from shaded environments (Trifolium repens L. and Vicia sativa L.) will exhibit an increase in root exudate production when subjected to high light intensity within a controlled setting, owing to their adaptation to such conditions [19]. Conversely, our second hypothesis suggests that plants originating from shaded environments (Ornithopus compressus L. and Coronilla juncea L.) will not display a significant increase in root exudate production when exposed to high light intensity [20]. By examining these hypotheses, we aim to gain a deeper understanding of how plants respond to changes in light conditions and the potential implications for soilborne microorganisms in their immediate vicinity.
2. Materials and Methods
2.1. Experimental Setting
The experimental design was the same as in [4]. In brief, a total of 24 tanks (45 cm x 32 cm x 40 cm) were utilized for the study. Eight tanks were placed in a growth chamber with a 12-hour light-dark cycle at 900 µmol.m-2.s-1 (intermediate light intensity). Another eight tanks were situated in a separate chamber with the same light-dark cycle but a light intensity of 1800 µmol.m-2.s-1 (high light intensity). The remaining eight tanks were placed on a lab bench in front of a window to receive natural light, measured at 450 µmol.m-2.s-1. Light was provided by lamps covering the entire photosynthetically active range.
The lab and the two chambers, maintained temperatures of 24±1 °C during the light period and 17±1 °C during the night, mimicking the average spring temperatures in the Iberian Peninsula. Each treatment consisted of two tanks, with four plant species under study: Vicia sativa L., Trifolium repens L., Coronilla juncea L., and Ornithopus compressus L. Seeds were sterilized with NaClO at 50% for 5 minutes and germinated in the dark on agar plates at 22 °C. After 33 days, seedlings were placed in each tank, suspended in a water-culture solution with a sponge rubber seedling holder. The four plant species are native to the Mediterranean region. The former ones, Trifolium repens and Vicia sativa, are usually found in shaded areas whereas the later ones, Coronilla juncea and Ornithopus compressus, have their natural range of distribution in well-irradiated areas.
Each tank was filled with a Hoagland nutrient solution at 5% in distilled water, containing low nitrogen levels to encourage nodule formation and N2-fixing symbioses. Groups of 60 seedlings each were subjected to different radiation levels: high radiation (1800 µmol.m-2.s-1), intermediate radiation (900 µmol.m-2.s-1), and low natural radiation (450 µmol.m-2.s-1) using different lightbulbs. Half of each tank (30 seedlings) was inoculated with the appropriate rhizobial strain, and the other half remained non-inoculated as controls.
2.2. Calculation of δ15N
The isotopic ratio of δ15N was calculated as δ = 1000 ‰ (Rsample/Rstandard), where R is the molar ratio of the heavier to the lighter isotope of the samples and standards according to [23].
%NDFA was calculated according to [24]:
where B represents the δ15N natural abundance of the nitrogen derived from BNF of the above-ground tissue of Trifolium repens, Vicia sativa, Coronilla juncea and Ornithopus compressus grown in a nitrogen-free culture. The B value of the four species was determined in this study as −2.38 ‰, -2.11 ‰, -2.96 ‰ and -3.01 ‰ respectively.
where R is the molar ratio of 15N and 14N of the samples and standards.
2.3. Enzime Activities Measurements
The extraction and assay of root nitrate reductase was performed following the methods by [25]. In short, duplicated samples of 0.1 g of fresh tissues were incubated for 1 hour at 30 °C in darkness after vacuum infiltration. The buffer used contained 0.1M KNO3 and 0.1M phosphate buffered at pH 7.5. Nitrite was measured colorimetrically at 540 nm after reacting with N-(1-naphthyl)-ethylene-diamine dihydrochloride. Nitrate reductase activity was estimated out of the amount of nitrite produced. Total nitrate reductase activity in roots was estimated by multiplying the nitrate reductase estimated as above, multiplied by the dry weigh of roots.
Acid phosphatase activity was assayed using the colorimetric method described by [26]. The reaction mixture contained 100 μL of 50 mM sodium acetate buffer (pH 5.0), 20 μL of enzyme extract, and 50 μL of 5 mM p-nitrophenyl phosphate. The reaction mixture was incubated at 37 °C for 30 minutes, and the reaction was stopped by adding 100 μL of 1M sodium hydroxide. The absorbance of the reaction mixture was measured at 405 nm, and the acid phosphatase activity was calculated using the extinction coefficient of p-nitrophenol (18 mM-1cm-1).
where, A405 is the absorbance of the reaction mixture at 405 nm; V is the total volume of the reaction mixture (μL); ε is the extinction coefficient of p-nitrophenol (18 mM-1cm-1); t is the incubation time (min; and m is the amount of protein in the enzyme extract (mg).
Phenolic acid content was determined using the Folin-Ciocalteu (FC) colorimetric method, following the procedure established for total phenolic acid estimation in plant extracts. This method is based on the reaction of the FC reagent with phenolic compounds, forming a blue complex, which is quantified spectrophotometrically at 760 nm [27]. In short, 50 grams of roots were frozen in liquid nitrogen and quickly ground using a mortar and pestle. One gram of the powdered plant material was transfer to a 50 ml centrifuge tube were 10 mL of 80% methanol was added. After thorough mixing the mixture was centrifuged at 10,000 rpm for 10 minutes. The supernatant was transferred to a clean test tube. The reaction mix consisted of 1 mL of the extract, 1 mL of 10% FC and 4 mL of a 7.5% sodium carbonate. This mix was incubated at room temperature for 30 minutes and then measured the absorbance of the reaction mixture at 760 nm using a spectrophotometer.
The total phenolic acid content in the root extracts was calculated as:
The results are expressed in milligrams of gallic acid equivalents (GAE) per gram of plant material.
2.4. Statistical Analysis
IBM SPSS Statistics 25 software [28] (Chicago, USA) was used for all data analysis. Analysis of variance (ANOVA) were used to compare means of all the plant variables obtained in the four different soils. Values were considered significant when p ≤ 0.05. Where the ANOVA showed significant differences between treatments, a Tuckey’s post hoc test was used to separate the means. Where two-way ANOVA showed significant effects at 95% (P < 0:05), Fisher’s least significant differences were determined using a one-way ANOVA for all replicates showing some germination. Normality and homoscedasticity of data were tested by means of David’s and Cochran’s tests.
3. Results
3.1. Biomass Production
The response of the plants to inoculation under three regimes of light intensity in terms of biomass production was different between treatments and among species with a minimum of 17.01 mg produced by non-inoculated plants of T. repens under low radiation; and a maximum of 4304 mg produced by inoculated plants of O. compressus under the intermediate radiation (Figure 1). The biomass production of the species originating from areas with high irradiance was generally lower than that of species from shaded areas. The former produced significantly more biomass under the intermediate radiation regimen after inoculation, whereas the latter produced significantly more biomass under the high and intermediate irradiance treatments after inoculation (Figure 1). As a general rule, inoculation enhanced biomass production regardless of the light treatment, with the exception of C. juncea, this species did not show as high an enhancement of biomass production under inoculation as that observed in the other species.
3.2. Nitrogen Concentrantions and Rhizobia Symbiotic Efficiency
The nitrogen content of plants grown under the three regimes of light intensity, inoculated and non-inoculated varied over a wide range of 10.65 to 40.2 mg.g–1 (Figure 2). Non-inoculated plants of V. sativa under low radiation, accumulated significantly (p= 0.0347) lower quantities of nitrogen, whereas plants of O. compressus under intermediate radiation accumulated significantly (p= 0.0065) larger quantities of nitrogen. There was a significantly (R2= 0.7048) correlation between biomass production and total N accumulation (Figure 2), with inoculated plants of O. compressus under the intermediate irradiance holding the greatest amount of both biomass and total N.
Isotopic δ15N natural abundance was analyzed in the shoots of the four species under all the light and inoculation treatments at harvesting. There was a clear correlation between total N accumulation and δ15N. The greater the BNF the higher the N content in plants (Figure 3). The isotopic analysis of plant material showed values of δ15N close to or below zero for all the inoculated plants regardless the light treatment. The lowest values of δ15N (i.e. the greatest BNF) were achieved by inoculated plants of V. sativa at intermediate light intensity, followed by inoculated plants of O. compressus in the low radiation treatment. The highest values of δ15N were observed in inoculated plants of O. compressus under the intermediate and high radiation treatments respectively. Inoculated plants of T. repens under the high and low radiation treatments achieve values of δ15N close to cero. Inoculated plants of V. sativa under the low radiation treatments did not show BNF.
3.3. Enzymatic Activities
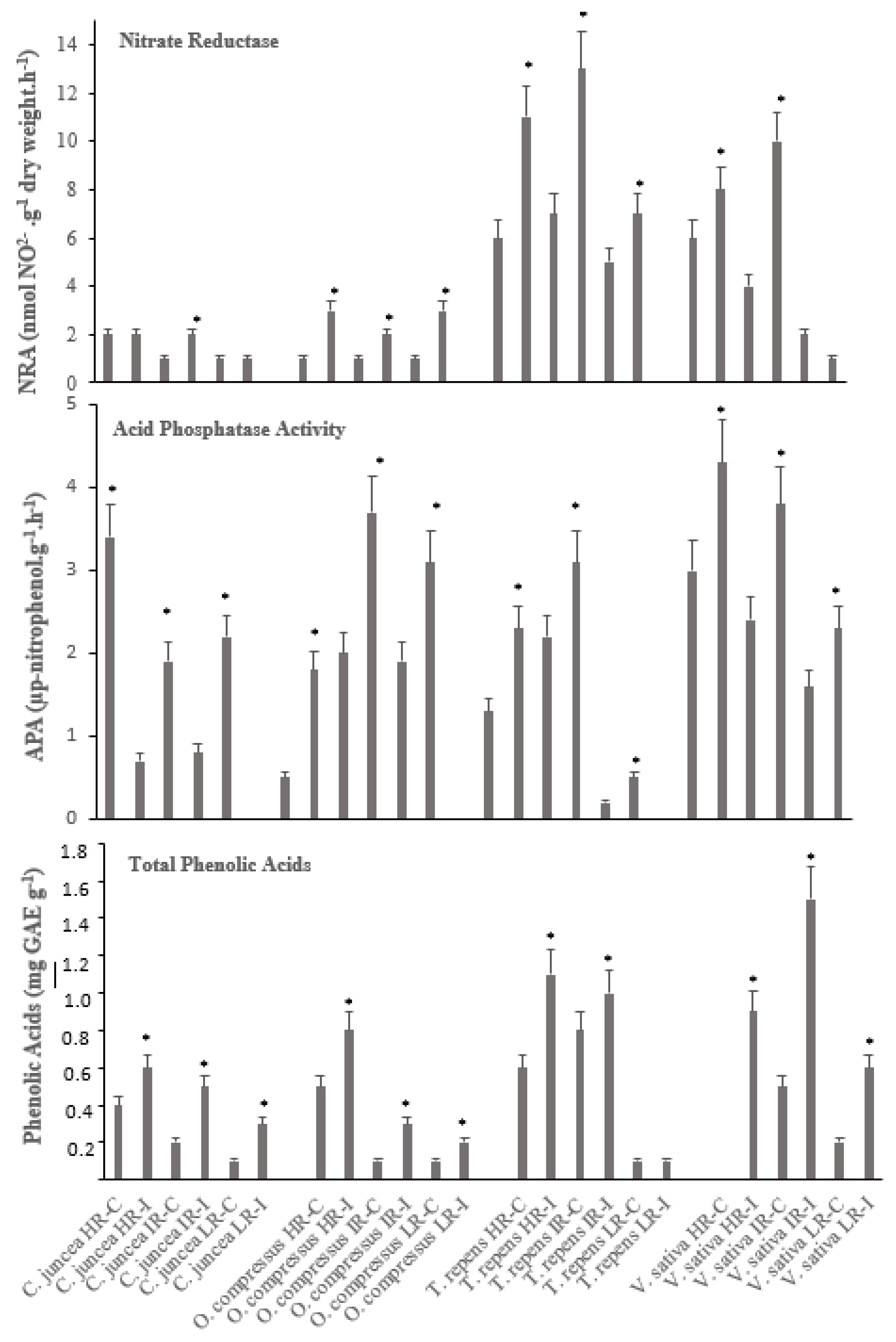
In general, the four legume species exhibited high capacity for nitrate reductase activity (NRA) within their root systems when subjected to rhizobial inoculation and challenging light conditions. The observed NRA response displayed significant interspecies variation, likely attributable to the distinct environmental histories of these plant species. Notably, NRA exhibited a remarkable increase in the presence of rhizobia across all species (Figure 4). Nevertheless, a striking divergence in NRA levels was evident when comparing the four species. Among the species examined, T. repens displayed the highest significant (p= 0.014) NRA levels (5-13 nmol NO2-. g-1. h-1) under conditions of moderate and high irradiance with the highest levels occurring in the presence of rhizobial inoculation. This was closely followed by V. sativa (1-10 nmol NO2-. g-1. h-1), underscoring the adaptability of plants originating from shaded-illuminated habitats in responding to external stimuli, ultimately enhancing NRA activity. Conversely, O. compressus (1-3 nmol NO2-. g-1. h-1) and C. juncea (1-2 nmol NO2-. g-1. h-1), both hailing from well-irradiated environments, exhibited the significantly lowest NRA levels (p= 0.0022).
The acid phosphatase activity (APA) in the roots of the four legume species exhibited a narrow range, with values ranging from 3.4 to 4.2 (µg p-nitrophenol.g-1.h-1). Despite this overall similarity, several differences were observed. In all species, the roots of the inoculated plants consistently displayed significantly higher APA (p< 0.005), except for C. juncea under high radiation treatment, where the highest APA was recorded in non-inoculated plants (p= 0.0233). Similarly, this species was the one that showed the lower values of APA. For T. repens the lowest APA levels were observed in plants under the low radiation treatment regardless the inoculation status (Figure 4).
All four species have similar total phenolic acid content, ranging from 0.4 to 1.2 mg GAE/g-1 (Figure 4) suggesting that all four species have a similar antioxidant capacity and may be similarly resistant to pests and diseases. Inoculated plants of V. sativa in the intermediate radiation treatment had the highest total phenolic acid content (1.5 mg GAEg-1) a value significantly different from that of the non-inoculated plants.
Surprisingly, non-inoculated plants of V. sativa in the high radiation treatment did not produce phenolic acids. The inoculated vs. non-inoculated pattern was observed consistently across all species, with the exception of T. repens under low radiation treatment. On the other hand, O. compressus, and C. juncea were the two species with the significantly lowest total phenolic acids contents (p< 0.005).
4. Discussion
In this study, we aimed to explore the impact of varying light intensities on the behavior of both inoculated and non-inoculated leguminous plants. Specifically, we postulated that plants originating from shaded environments (T. repens and V. sativa) will exhibit an increase in root exudate production when subjected to high light intensity within a controlled setting, owing to their adaptation to such conditions [19]. Conversely, our second hypothesis suggests that plants originating from high-light environments (O. compressus and C. juncea) will not display a significant increase in root exudate production when exposed to high light intensity [20]. By examining these hypotheses, we aim to gain insight into how plants respond to changes in light conditions and the potential implications for soilborne microorganisms in their immediate vicinity and its application in agriculture.
The response of the plants to inoculation under three regimes of light intensity in terms of biomass production was different between treatments and among species, with significant variations in biomass production (Figure 1). Notably, biomass production was generally lower in species originating from high-irradiance areas when compared to those from shaded environments, highlighting potential trade-offs in resource allocation and environmental adaptation. Inoculation positively influenced biomass production across species, except for Coronilla juncea, which showed a less pronounced response.
The nitrogen content of plants grown under the three regimes of light intensity, inoculated and non-inoculated varied over a wide range of 10.65 to 40.2 mg.g–1 (Figure 2). Non-inoculated plants of V. sativa under low radiation, accumulated significantly (p= 0.0347) lower quantities of nitrogen, whereas plants of O. compressus under intermediate radiation accumulated significantly (p= 0.0065) larger quantities of nitrogen. There was a significantly (R2= 0.7048) correlation between biomass production and total N accumulation (Figure 2), with inoculated plants of O. compressus under the intermediate irradiance holding the greatest amount of both biomass and total N. This range of values under different light treatments, reflects the plasticity of legumes in adapting to varying light conditions as also observed in plants of Glycine max L. [29,30] and Medicago sativa L. [31]. There was a strong correlation between biomass production and total nitrogen accumulation, indicating the interplay between light intensity, nitrogen uptake, and biomass.
Isotopic analysis further revealed a connection between total nitrogen accumulation and δ15N values, suggesting that greater biological nitrogen fixation (BNF) resulted in higher nitrogen content (Figure 3). [32] found that light intensity affected the rhizosphere microbiome composition and nitrogen fixation in soybean-rhizobia symbiosis. They found that high light intensity increased the abundance of nitrogen-fixing bacteria in the rhizosphere, which led to increased nitrogen fixation, as happens in our study, with the exception of V. sativa. The contrasting responses of plants to similar light intensities is not novel but still controversial. [31] and [30] both found that light intensity and rhizobial infection as well as plant growth in M. sativa and G. max were enhanced at high light intensity. However, [33] found that high light intensity decreased the root exudation of soybean plants, with led to decreased nutrient uptake and growth. Consequently, the nature of the plant species might have a role to play in their response to light, still to be elucidated.
The isotopic analysis of plant material showed values of δ15N close to or below zero for all the inoculated plants regardless the light treatment. The lowest values of δ15N (i.e. the greatest BNF) were achieved by inoculated plants of V. sativa at intermediate light intensity, followed by inoculated plants of O. compressus in the low radiation treatment. The highest values of δ15N were observed in inoculated plants of O. compressus under the intermediate and high radiation treatments respectively. Inoculated plants of T. repens under the high and low radiation treatments achieve values of δ15N close to cero. Inoculated plants of V. sativa under the low radiation treatments did not show BNF. These results let us think that specific environmental factors, as exemplified by V. sativa in the high radiation treatment, add complexity to nitrogen dynamics in legume species [34].
Nitrate reductase activity (NRA) within root systems exhibited a high capacity across all four legume species, with a substantial increase in response to rhizobial inoculation. However, NRA levels varied significantly among species, with T. repens and V. sativa, originating from shaded-illuminated habitats, showing the highest NRA levels under moderate and high irradiance. This in concordance with observations of other legumes in stressful conditions, were, for example drought or salinity induce higher production of NR to enhance nodule formation and BHF [35,36,37]. Conversely, O. compressus and C. juncea, from well-illuminated environments, exhibited significantly lower NRA levels. Acid phosphatase activity (APA) showed a relatively narrow range across the species, with consistently higher APA levels in inoculated plants. Yet, specific responses, as observed in V. sativa under high radiation, highlighted complex interactions between light conditions, inoculation, and enzymatic activities. As it occurred with NRA, stressed legumes like Medicago sativa or Glycine max [33,31,38], they increase the production of APA when the environment imposes stress on the plant, thus aiding the legumes to acquire P, that together with N is one of the most limiting nutrients for plants and that are crucial in nodule formation. These responses can be shown by plants that were formed under stressful conditions and behavior is maintained is next generations [39,40,41]. The variations in enzymatic activities, particularly nitrate reductase and acid phosphatase, underscore the complex relationships between plants and their associated microorganisms. These enzymes play crucial roles in nutrient acquisition and nutrient cycling in the rhizosphere. The observed interspecies differences in enzymatic responses suggest that the selection of legume species for specific agricultural contexts should consider their interactions with soil microorganisms and nutrient cycling.
All four species displayed similar total phenolic acid content, indicating comparable antioxidant capacities and potential resistance to pests and diseases. The response of V. sativa in the high radiation treatment, not producing phenolic acids, suggested intricate responses to high radiation conditions that cannot be depicted just by looking at our results and further research is required. Inoculated plants generally exhibited higher total phenolic acid content, consistently across species, with T. repens in low radiation as an exception. O. compressus and C. juncea exhibited the lowest total phenolic acid contents, possibly reflecting their adaptation to well-irradiated environments. All these findings are in accordance with those for G. max [42]. The consistent total phenolic acid content across the four legume species, with specific responses to different light conditions and inoculation, highlights the potential of these compounds in plant defense mechanisms. The findings suggest that legume species originating from shaded-light environments may allocate more resources to phenolic acid production as part of their adaptive strategy. This information can be valuable for breeding programs aiming to enhance the resistance of legume crops to pests and diseases.
Our results offer valuable insights into the mining strategy of legume plants in the context of changing light conditions [43]. The mining strategy of plants theory states that plants have evolved different strategies to acquire nutrients from the soil. Some plants are "exploiters", meaning that they have a high nutrient demand and are able to rapidly acquire nutrients from the soil. Other plants are "conservators," meaning that they have a low nutrient demand and are able to efficiently acquire nutrients from the soil even when they are scarce [19,20]. In the face of climate change, the mining strategy becomes particularly relevant, as plants must adapt to shifting environmental factors while efficiently utilizing available resources [14]. We have demonstrated that plants originating from high-light environments may be more exploitive, while plants originating from shaded environments may be more conservative. Additionally, rhizobial inoculation may enhance the ability of plants to acquire nutrients from the soil, even under challenging light conditions [44]. The species from shaded environments, T. repens and V. sativa, displayed lower biomass production under high irradiance, indicating potential resource allocation towards other essential processes like root exudate production as previously demonstrated [4]. This response aligns with the mining strategy of optimizing resource use under varying conditions. In contrast, species from well-illuminated environments, O. compressus and C. juncea, exhibited significant increases in biomass under high irradiance, potentially as a strategy to capture available light and nutrients more efficiently. The strong correlations between biomass production and total nitrogen accumulation emphasize the role of the mining strategy in nutrient acquisition. Legumes, particularly those adapted to high-light environments, appear to invest in nitrogen uptake and biological nitrogen fixation (BNF) as part of their mining strategy. Inoculated plants, which rely more on BNF, exhibited lower δ15N values, indicating a successful mining strategy to acquire and utilize atmospheric nitrogen [45,34]. Enzymatic activities, such as nitrate reductase and acid phosphatase, play crucial roles in nutrient mining [36,38]. The significant interspecies variations in nitrate reductase activity highlight the adaptability of legumes to different light conditions and their resource allocation strategies. Shaded-light-adapted species (T. repens and V. sativa) demonstrated higher nitrate reductase levels under moderate to high irradiance, aligning with the mining strategy of optimizing nutrient uptake in well-illuminated environments. The consistent total phenolic acid content across species, with specific responses to different light conditions and inoculation, underscores the multifaceted nature of the mining strategy [19,43]. Legume species appear to allocate resources to phenolic acid production, potentially as part of their defense mechanism against pests and diseases, while adapting to changing light conditions [31,38].
The findings of this study have practical implications for the cultivation of legume crops in diverse environmental settings. Understanding how legumes respond to varying light conditions, nitrogen dynamics, and enzymatic activities can inform strategies to optimize legume-based agriculture. For example, the observed positive impact of rhizobial inoculation on biomass production in most species suggests that this agricultural practice may be particularly beneficial in environments with limited nitrogen availability. Farmers could select crop varieties that are adapted to different light environments and that have different mining strategies. Additionally, farmers could use rhizobial inoculants to improve nutrient acquisition in crops under a variety of light conditions [46],[47]. However, careful consideration of the specific legume species and their adaptation to light conditions is necessary to maximize the benefits of inoculation.
In conclusion, this study provides compelling evidence that varying light intensities indeed influence the behavior of legume species, with different responses observed among those adapted to high-light and shaded environments. Our findings support the first hypothesis, indicating that high-light-adapted species such as O. compressus and C. juncea exhibit increased root exudate production in response to high light intensity. Conversely, the second hypothesis is upheld as legumes originating from well-illuminated environments, including T. repens and V. sativa, do not significantly increase root exudate production under high light intensity. The study also highlights the intricate interplay between nitrogen dynamics, enzymatic activities, and phenolic acid content in legumes, shedding light on their adaptation strategies and potential applications in agriculture. These insights offer a valuable foundation for the development of tailored agricultural practices and crop breeding strategies to address the challenges of a changing climate and ensure sustainable food production.
Author Contributions
All authors have contributed equally to all parts of the study, from the design, sample collection and analyses, data analyses and interpretation and writing-up the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data will be made available on request.
Acknowledgments
The authors acknowledge Jesús del Gran Poder for his assistance during the lab work, particularly in the maintenance of the irradiance conditions through the entire length of the experiment.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kloepper, J. W.; & Ryu, C. M. J. Sustainable agriculture: The emerging role of rhizosphere bacteria. Nature Reviews Microbiology 2006, 4, 866–879. [Google Scholar]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. (2017) Plant Physiology and Development. 6th Edition, Artmed, Porto Alegre, 858 p.
- Gazolla-Neto, A.; Aumonde, T. Z.; Pedó, T.; Olsen, D. Ação de níveis de luminosidade sobre o crescimento de plantas de maria-pretinha (Solanum americanum Mill). 2013; 11. [Google Scholar]
- De Lara-Del Rey, I.A.; Pérez-Fernández, M.A. Regulatory Effect of Light and Rhizobial Inoculation on the Root Architecture and Plant Performance of Pasture Legumes. Agronomy 2023, 13, 2058. [Google Scholar] [CrossRef]
- Domínguez-Martín, M. A.; Sauer, P. V.; Kirst, H.; Sutter, M.; Bína, D.; Greber, B. J.; Nogales, E.; Polívka, T.; Kerfeld, C. A. Structures of a Phycobilisome in Light-Harvesting and Photoprotected States. Nature 2022, 609, 835–845. [Google Scholar] [CrossRef]
- Alsanius, B.; Karlsson, M.; Rosberg, A.; Dorais, M.; Naznin, M.; Khalil, S.; Bergstrand, K.-J. Light and Microbial Lifestyle: The Impact of Light Quality on Plant–Microbe Interactions in Horticultural Production Systems—A Review. Horticulturae 2019, 5, 41. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Domanski, G. Carbon Input by Plants into the Soil. Review. J. Plant Nutr. Soil Sci. 2000, 163, 421–431. [Google Scholar] [CrossRef]
- Badri, D. V.; Vivanco, J. M. Regulation and Function of Root Exudates. Plant Cell & Environment 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Haichar, F. E. Z.; Santaella, C.; Heulin, T.; Achouak, W. Root Exudates Mediated Interactions Belowground. Soil Biology and Biochemistry 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Bais, H. P.; Weir, T. L.; Perry, L. G.; Gilroy, S.; Vivanco, J. M. THE ROLE OF ROOT EXUDATES IN RHIZOSPHERE INTERACTIONS WITH PLANTS AND OTHER ORGANISMS. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Robson, T. M.; Klem, K.; Urban, O.; Jansen, M. A. K. Re-interpreting plant morphological responses to UV-B radiation. Plant, Cell & Environment, 2015; 38, 856–866. [Google Scholar]
- Bloom, A. J.; Chapin III, F. S.; & Mooney, H. A. (1985). Resource limitation in plants an economic analogy. Annual Review of Ecology and Systematics 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Milla, R.; Reich, P. B.; & Castro-Díez, P. Tri-trophic interactions in the context of climate change: Effects of psyllid herbivory on nitrogen acquisition and resource allocation in a fast-growing herb. Oikos 2008, 117, 1865–1877. [Google Scholar]
- Lambers, H.; Raven, J.; Shaver, G.; Smith, S. Plant Nutrient-Acquisition Strategies Change with Soil Age. Trends in Ecology & Evolution 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R. P.; Finzi, A. C.; Bernhardt, E. S. Enhanced Root Exudation Induces Microbial Feedbacks to N Cycling in a Pine Forest under Long-term CO 2 Fumigation. Ecology Letters 2011, 14, 187–194. [Google Scholar] [CrossRef]
- Mandal, S. M.; Chakraborty, D.; Dey, S. Phenolic Acids Act as Signaling Molecules in Plant-Microbe Symbioses. Plant Signaling & Behavior 2010, 5, 359–368. [Google Scholar] [CrossRef]
- Bainard, L. D.; Klironomos, J. N.; Hart, M. M. Differential Effect of Sample Preservation Methods on Plant and Arbuscular Mycorrhizal Fungal DNA. Journal of Microbiological Methods 2010, 82, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Mougel, C.; Jaillard, B.; Hinsinger, P. Plant-Microbe-Soil Interactions in the Rhizosphere: An Evolutionary Perspective. Plant Soil 2009, 321, 83–115. [Google Scholar] [CrossRef]
- Smith, A. Adaptation of Plants to High-Light Environments. Plant Ecology 2019, 214, 215–231. [Google Scholar]
- Brown, C. Shade as a Driver of Plant Adaptations to Low-Light Environments. Environmental Biology 2020, 145, 1611–1621. [Google Scholar]
- Lachica M., Aguilar A., Yánez J. Análisis foliar. Métodos utilizados en la Estación Experimental del Zaidín CSIC (II). An. Edaf. Agrobiol. 1973, 32, 1033–1047.
- Bouat, A. , Crouzet C. Notes techniques sur un appareil semiautomatique de clorage de l’azote et de certains composes volatiles. Ann. Agric. 1965, 16, 107–118. [Google Scholar]
- Farquhar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annual Review of Physiology Plant and Molecular Biology 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Shearer. G.; Kohl, D.H. N2-fixation in field settings: Estimations based on natural 15N abundance. Functional Plant Biology 1986, 13, 699–756. [Google Scholar]
- Langelaan, J. G.; Troelstra, S. R. Growth, Chemical Composition, and Nitrate Reductase Activity of Rumex Species in Relation to Form and Level of N Supply. Plant Soil 1992, 145, 215–229. [Google Scholar] [CrossRef]
- Streeter, J. G. Acid and alkaline phosphatase activities in roots of legumes. Plant Physiology 1985, 77, 610–614. [Google Scholar]
- Singleton, V. L.; Orthofer, R.; & Lamuela-Raventós, R. M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods in Enzymology 1999, 299, 152–178. [Google Scholar]
- IBM SPSS Software. Available online: https://www.ibm.com/analytics/spss-statistics-software (accessed on 1 Nov 2023).
- Liu, Y.; Liu, M.; Liu, S.; Wang, X.; & Li, X. Light intensity and rhizobial inoculation affect root morphology and exudation in soybean (Glycine max L.). Journal of Plant Physiology, 2021; 259, 153333.
- Li, Y.; Sun, W.; Zhang, X.; Li, F.; &; Wang, Y. Light intensity-dependent effects of rhizobial inoculation on growth, photosynthesis, and nitrogen metabolism in soybean (Glycine max L.). Frontiers in Plant Science 2022, 13, 873180. [Google Scholar]
- Cui, J.; Zhao, J.; Wang, Y.; Liu, S.; Li, J.; & Zhang, F. Light intensity and rhizobial inoculation affect the growth, photosynthesis, and nitrogen metabolism of alfalfa (Medicago sativa L.). Journal of Integrative Agriculture, 2022; 21, 617–628. [Google Scholar]
- Chen, Y.; Wang, Z.; Zhang, Z.; Sun, Y.; & Li, X. Light intensity affects the rhizosphere microbiome composition and nitrogen fixation in soybean-rhizobia symbiosis. Frontiers in Microbiology 2021, 12, 746538. [Google Scholar]
- Wang, C.; Han, J.; Sun, J.; &; Gao, Q. Light intensity affects the growth, physiological traits, and root exudates of soybean plants (Glycine max L.) inoculated with rhizobia. Plant Physiology and Biochemistry, 2020; 152, 129–141. [Google Scholar]
- Zhao, J.; Cui, J.; Chen, J.; Liu, S.; & Zhang, F. Rhizobial inoculation enhances the tolerance of alfalfa (Medicago sativa L.) to salt stress under different light conditions. Frontiers in Plant Science, 2023; 14, 1063596.
- Hussain, H. A.; Farooq, M.; Ashraf, M.; & Khan, M. I. R. Physiological and molecular mechanisms of drought tolerance in legumes: A review. Agronomy 2020, 10, 695. [Google Scholar]
- Khan, A.; Ali, S.; Hameed, A.; Farooq, M.; & Iqbal, N. Role of nitrate reductase activity in drought tolerance of legumes: A review. Plants 2022, 11, 348. [Google Scholar]
- Wang, C.; Zhang, H.; Zhang, W.; Wang, L.; Zhang, Y.; Li, G.; & Yang, J. Nitrate reductase and its regulation in plants under abiotic stress. Frontiers in Plant Science 2022, 13, 858372. [Google Scholar]
- Hu, J.; Lu, L.; Chen, X.; &; Li, Z. Acid phosphatase activity and physiological responses of alfalfa (Medicago sativa L.) to salt stress and arbuscular mycorrhizal fungus inoculation. Environmental Science and Pollution Research 2022, 29, 43338–43353. [Google Scholar]
- Güsewell, S. N : P Ratios in Terrestrial Plants: Variation and Functional Significance. New Phytologist 2004, 164, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F. S. ; III, & Bloom, A. J. Phosphate absorption: A major component of nutrient acquisition for some wild plant communities. Oecologia 1981, 51, 11–21. [Google Scholar]
- Míguez-Montero, M.A.; Valentine, A.; Pérez-Fernández, M.A. Regulatory Effect of Phosphorus and Nitrogen on Nodulation and Plant Performance of Leguminous Shrubs. AoB Plants 2019, 12, plz047. [Google Scholar] [CrossRef]
- Meng, X.; Li, H.; Wang, Z.; Ma, Z.; Wang, J.; Wang, X.; &; Li, X. Rhizobial inoculation enhances the tolerance of soybean (Glycine max L.) to high light stress through modulating the antioxidant system and nitrogen metabolism. Frontiers in Plant Science 2021, 12, 657643. [Google Scholar]
- Baxter, I.; & Dilkes, B. J. The plant mining strategy theory and its implications for plant evolution and nutrient cycling. Journal of Ecology 2018, 106, 1696–1704. [Google Scholar]
- Hussain, H. A. , Farooq, M., Ashraf, M., & Khan, M. I. R. Physiological and molecular mechanisms of drought tolerance in legumes: A review. Agronomy 2020, 10, 695. [Google Scholar]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Chapin, F. S. The Mineral Nutrition of Wild Plants. Annu. Rev. Ecol. Syst. 1980, 11, 233–260. [Google Scholar] [CrossRef]
Figure 1.
Plant dry weights of V. sativa, T. repens, C. juncea and C. compressus grown under three regimes of light irradiance (HR-High irradiance; IR- Intermediate irradiance; and LR- Low irradiance) with (I) and without (C) bacterial inoculation. Values are presented as means (n = 15) with standard error bars and letters indicate significant differences among treatments using the post hoc Fisher’s LSD, multiple range test (P ≤ 0.05). Color code: Orange: Species from shaded environments; Green: species from well-irradiated environments.
Figure 1.
Plant dry weights of V. sativa, T. repens, C. juncea and C. compressus grown under three regimes of light irradiance (HR-High irradiance; IR- Intermediate irradiance; and LR- Low irradiance) with (I) and without (C) bacterial inoculation. Values are presented as means (n = 15) with standard error bars and letters indicate significant differences among treatments using the post hoc Fisher’s LSD, multiple range test (P ≤ 0.05). Color code: Orange: Species from shaded environments; Green: species from well-irradiated environments.
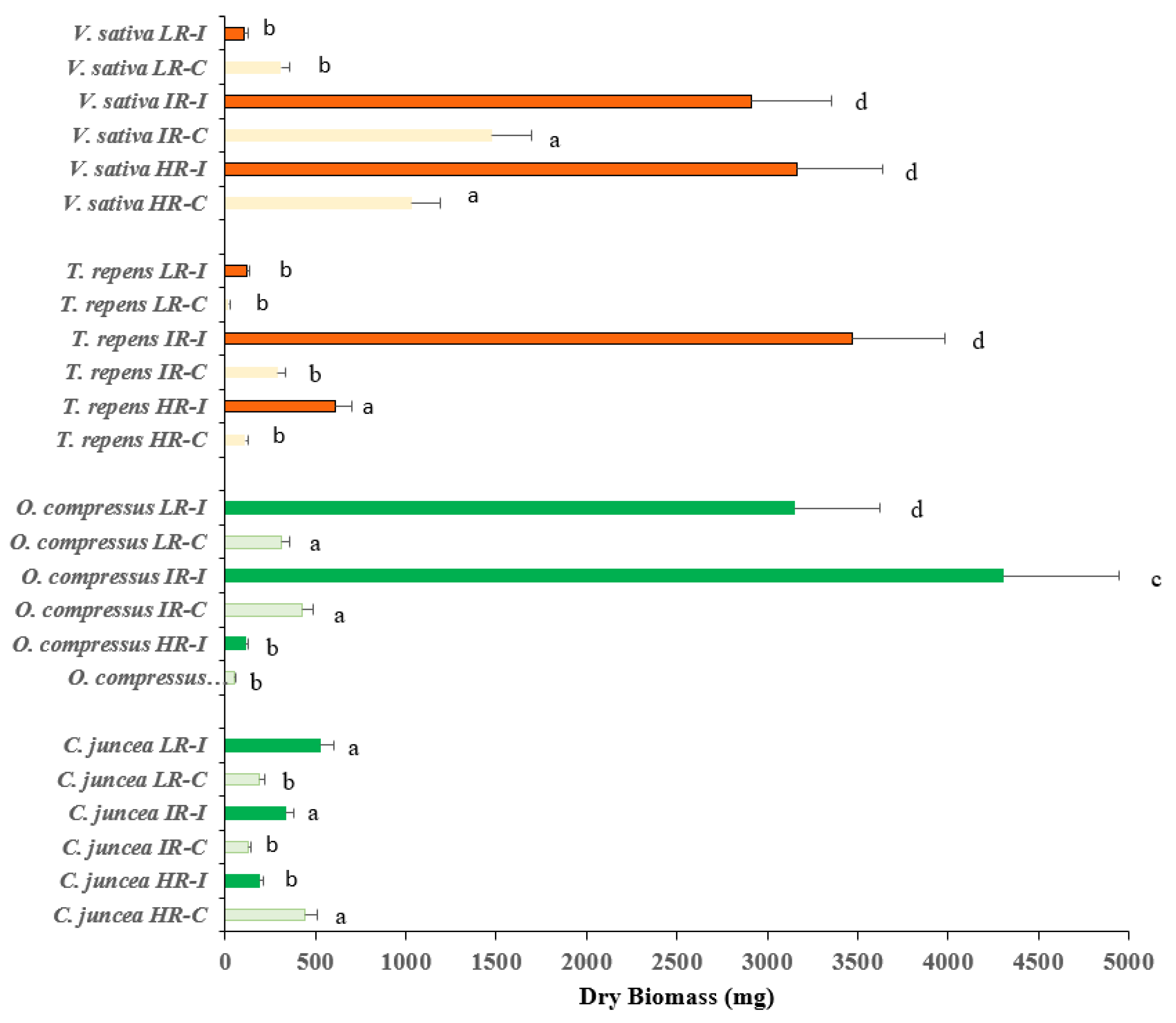
Figure 2.
Plant dry mass vs. total plant nitrogen content of V. sativa, T. repens, C. juncea and O. compressus grown under three regimes of light irradiance (HR-High irradiance; IR- Intermediate irradiance; and LR- Low irradiance) with (I) and without (C) bacterial inoculation. Each point corresponds to the mean value for a plant species, light regime, and inoculation treatment.
Figure 2.
Plant dry mass vs. total plant nitrogen content of V. sativa, T. repens, C. juncea and O. compressus grown under three regimes of light irradiance (HR-High irradiance; IR- Intermediate irradiance; and LR- Low irradiance) with (I) and without (C) bacterial inoculation. Each point corresponds to the mean value for a plant species, light regime, and inoculation treatment.
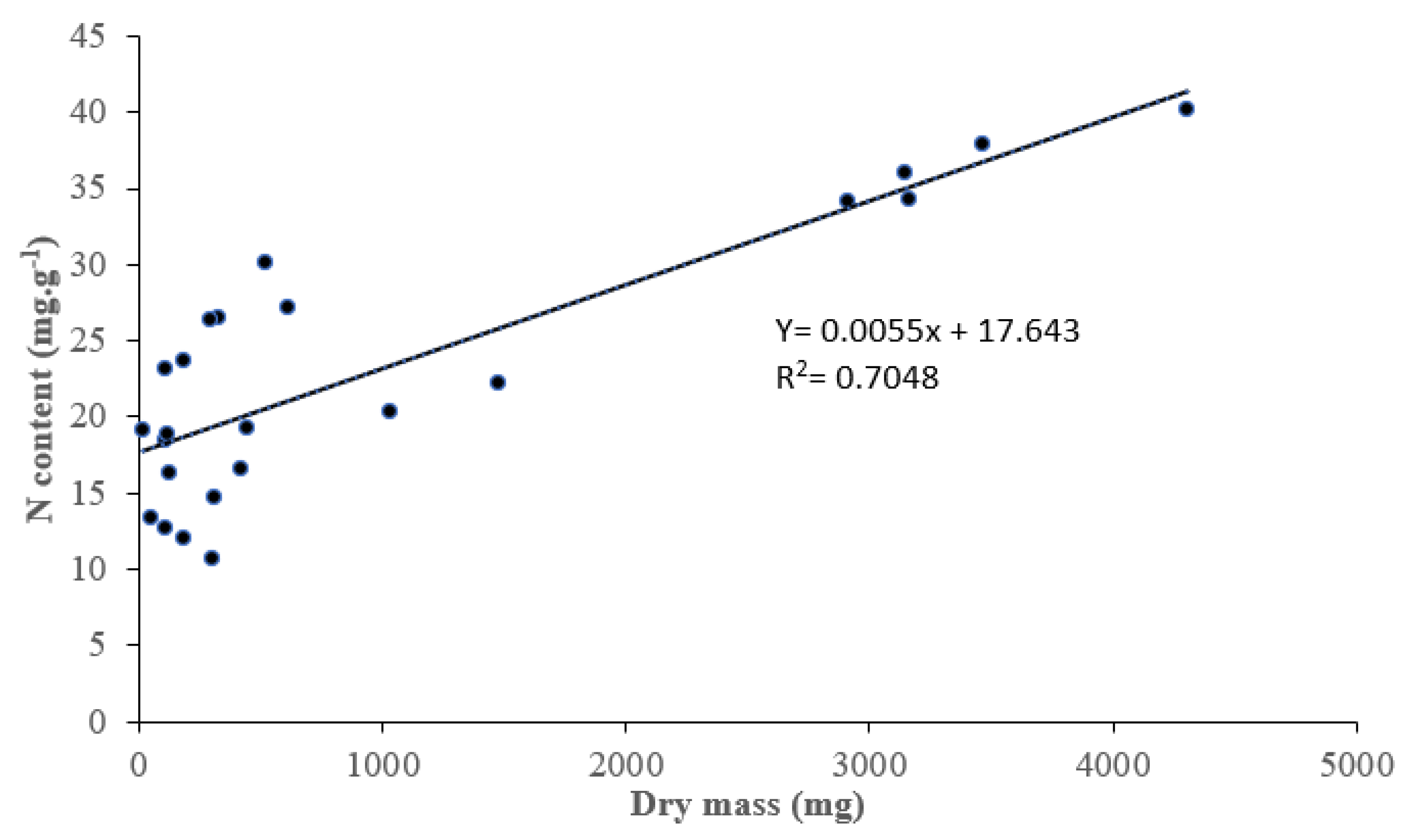
Figure 3.
δ15N plant content vs. total plant nitrogen content of V. sativa, T. repens, C. juncea and O. compressus grown under three regimes of light irradiance (HR-High irradiance; IR- Intermediate irradiance; and LR- Low irradiance) with (I) and without (C) bacterial inoculation. Each point corresponds to the mean value for a plant species, light regime, and inoculation treatment.
Figure 3.
δ15N plant content vs. total plant nitrogen content of V. sativa, T. repens, C. juncea and O. compressus grown under three regimes of light irradiance (HR-High irradiance; IR- Intermediate irradiance; and LR- Low irradiance) with (I) and without (C) bacterial inoculation. Each point corresponds to the mean value for a plant species, light regime, and inoculation treatment.
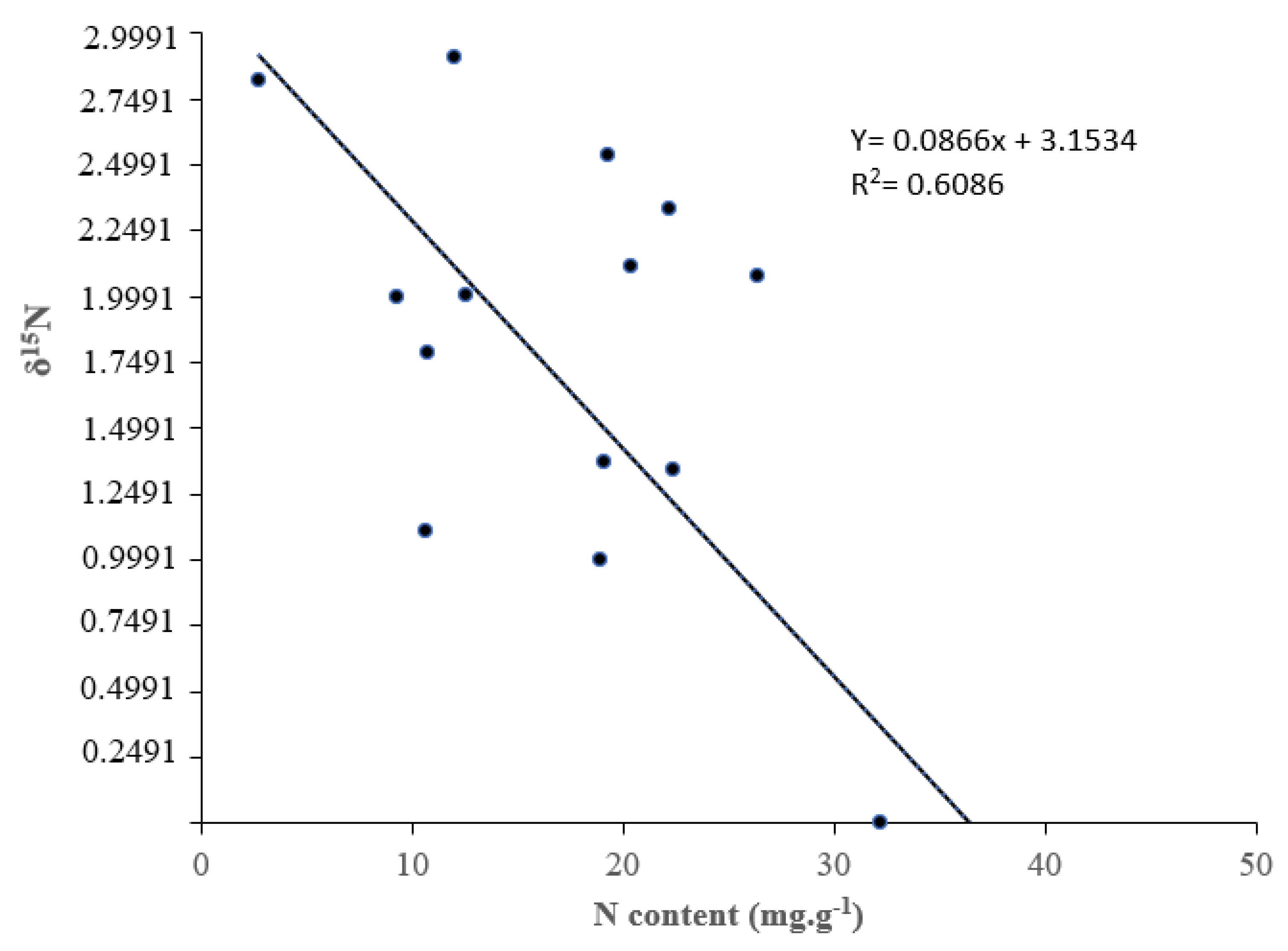
Figure 4.
Values of Nitrate reductase and Acid Phosphatase activities and total Phenolic acids measured in plants of V. sativa, T. repens, C. juncea and O. compressus grown under three regimes of light irradiance and inoculation (as in Figure 1). Values are means (n = 15±SD). Asterisks indicate significant differences between control after post hoc Fisher’s LSD, multiple range test (p ≤ 0.05).
Figure 4.
Values of Nitrate reductase and Acid Phosphatase activities and total Phenolic acids measured in plants of V. sativa, T. repens, C. juncea and O. compressus grown under three regimes of light irradiance and inoculation (as in Figure 1). Values are means (n = 15±SD). Asterisks indicate significant differences between control after post hoc Fisher’s LSD, multiple range test (p ≤ 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



