
Submitted:
06 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
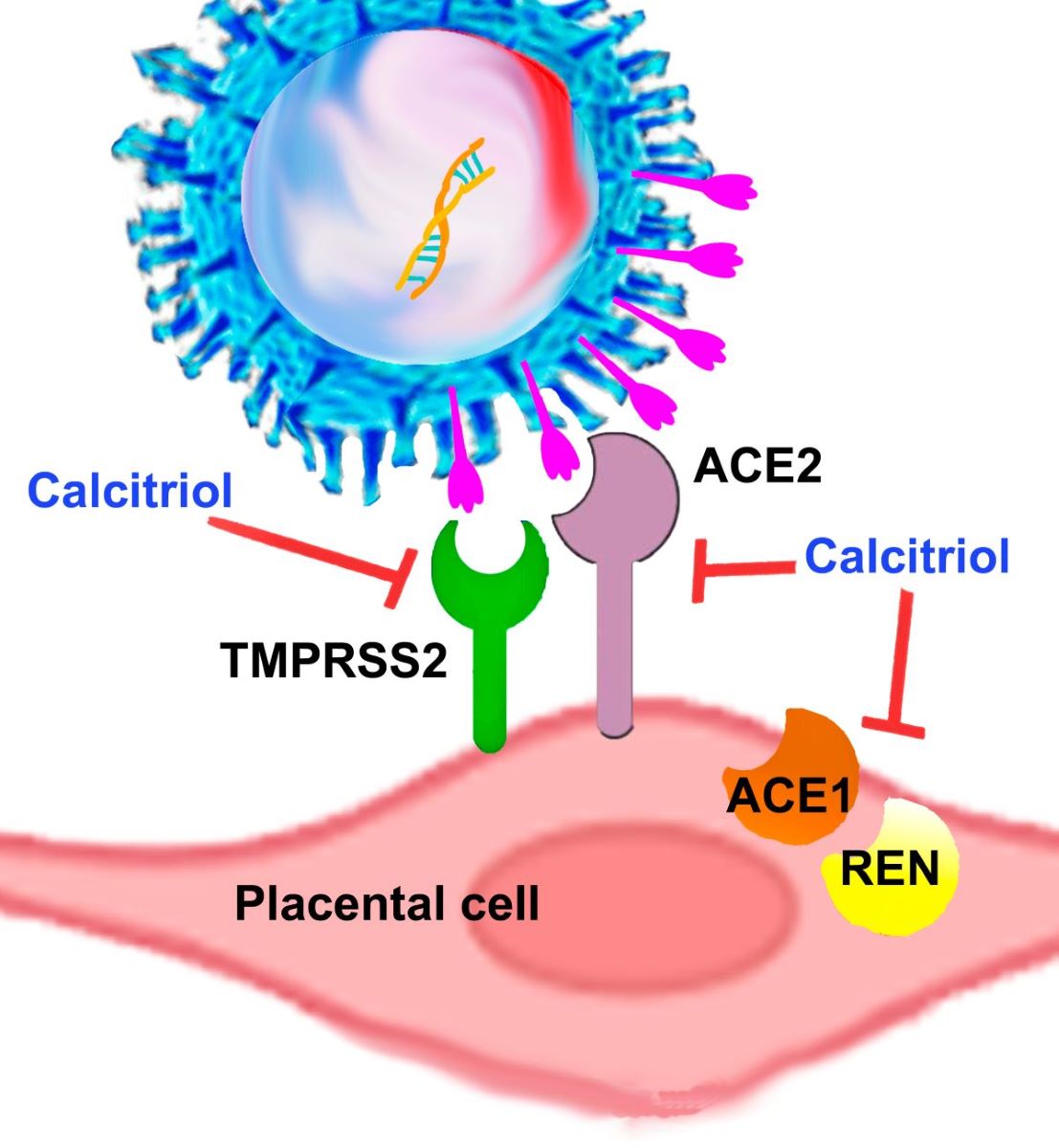
Angiotensin-converting enzyme (ACE)1, ACE2, and renin are components of the renin-angiotensin system (RAS), which regulates blood pressure. ACE2 also serves as a receptor for SARS-CoV-2 and together with the transmembrane serine protease 2 (TMPRSS-2), mediates viral cell-endocytosis. As the placenta expresses all these factors, it acts as a target for SARS-CoV-2 and also as a source of blood pressure modulators. An ACE1/ACE2 ratio imbalance can lead to RAS dysregulation and a bad prognosis in COVID-19 patients. Calcitriol, the vitamin D active metabolite, negatively regulates RAS, reduces inflammation, and enhances antiviral immunity, thereby playing a protective role against COVID-19 severity. Placental calcitriol has been inversely correlated with maternal blood pressure; however, its regulatory role in RAS components and SARS-CoV-2 receptors within the fetomaternal unit has been barely explored. Therefore, we investigated the effects of calcitriol on placental RAS components. Calcitriol downregulated ACE1, ACE2, TMPRSS-2, and renin gene expression in cultured syncytiotrophoblasts and the extravillous trophoblast cell line HTR-8/SVneo. The ACE1/ACE2 ratio was also downregulated by calcitriol. Similar results were obtained in syncytiotrophoblasts treated with calcidiol, the precursor of calcitriol. Altogether, these results support that vitamin D is essential in restricting SARS-CoV-2 placental infection while helping to regulate maternal blood pressure during pregnancy.
Keywords:
Syncytiotrophoblasts
; Renin-Angiotensin-Aldosterone
; ACE1
; ACE2
; TMPRSS2
; Calcitriol
; Vitamin D
; Blood pressure
; Pregnancy
1. Introduction
A substantial and consistent amount of information has accumulated in recent decades regarding the beneficial effects of vitamin D (VD) during the perinatal period. VD exerts its biological activity mainly at the transcriptional level, through the binding of its active metabolite, calcitriol, to the vitamin D receptor (VDR). The human placenta is a target and a source of calcitriol, as it expresses both the VDR and the vitamin D-activating cytochrome CYP27B1. Aside from its well-known calciotropic properties, the biological effects of VD during pregnancy can be categorized into four main domains. Firstly, VD exhibits anti-inflammatory properties by downregulating systemic and placental inflammatory factors, thereby maintaining a balanced immune response by limiting an exacerbation during inflammatory events [1,2,3]. Secondly, VD boosts the innate immune response by inducing antimicrobial peptides, aiding in defense against viral and bacterial invasions [3,4,5]. Thirdly, VD helps regulate the synthesis of placental steroid and protein hormones, contributing to a healthy pregnancy [6,7,8]. Lastly, VD regulates blood pressure, a crucial factor for pregnant women. In this regard, it is well established that calcitriol downregulates renin gene expression, as demonstrated in renal cells both in vitro and in vivo [9]. As a result, calcitriol inhibits the renin–angiotensin system (RAS), which is tightly involved in blood pressure modulation and electrolyte and volume homeostasis [10,11]. The RAS mediates its effects by generating angiotensin II (Ang II), a potent vasoconstrictor peptide. Ang II is synthesized after two sequential cleavage reactions, the first one upon angiotensinogen, catalyzed by renin, and the second one upon the resulting product, angiotensin I (Ang I), by the angiotensin-converting enzyme (ACE1). Since renin is the rate-limiting enzyme in Ang II synthesis, factors that modify its transcriptional regulation and secretion are of clinical importance in hypertensive disorders. In this context, and consistently with the inhibition of renin by calcitriol, pre-clinical, clinical and epidemiological studies have shown a negative association between calcitriol and blood pressure, as well as a reduction in blood pressure after patients treatment with calcitriol, its analogs or its precursor cholecalciferol [9,12,13,14,15,16,17]. Similarly, a negative correlation between calcitriol cord blood levels and maternal blood pressure has been described in a cohort of pregnant women affected by urinary tract infections [18], strongly suggesting that placental calcitriol is involved in lowering maternal blood pressure. Blood pressure during pregnancy must be closely monitored since hypertensive disorders of pregnancy can significantly increase perinatal morbidity and mortality [19]. In this sense, the natural counter-regulatory mechanism of RAS activation is represented by ACE2, which hydrolyzes Ang I and Ang II into Ang-(1-9) and Ang-(1-7), respectively [20]. Particularly, Ang-(1-7) acts as a vasodilator, anti-hypertensive, antioxidant and anti-inflammatory peptide [20]. The human placenta expresses all components of the RAS [21,22], being ACE2 mainly expressed in the syncytiotrophoblast layer [23]. Interestingly, ACE2 also acts as a receptor for SARS-CoV-2, the causal agent of COVID-19 disease, which produced the recent health contingency pandemic [24]. Accordingly, SARS-CoV-2 viral particles have been mainly localized in syncytiotrophoblast cells (STB) at the materno-fetal interface of the placenta [24,25]. In the aftermath, among the most vulnerable groups, the impact of COVID-19 was particularly significant among pregnant women, given that this disease was associated with increased risk of maternal death, cardiovascular disorders, obstetric hemorrhage, hypertension, and preeclampsia [26]. The mechanism of SARS-CoV-2 infection starts with the binding of the coronavirus spike proteins (S) receptor binding domain (RBD) to host cells membrane ACE2, followed by S cleavage by the transmembrane serine protease TMPRSS2. This process allows the viral envelope to fuse with the host cell membrane, after which the viral RNA is released into the cell for translation into structural and accessory viral proteins needed for genome replication. Thus, both ACE2 and TMPRSS2 are fundamental for the SARS-CoV-2 infection, representing potential therapeutic targets [27]. Notably, a significant amount of information has accumulated supporting a protective role of VD against COVID-19 severity and mortality [28,29], which is thought to be mediated through the downregulation of proinflammatory cytokines, the production of antiviral proteins and several other mechanisms [30,31,32]. However, there are contrasting results in the literature regarding the effects of calcitriol on ACE and TMPRSS2 expression in different tissues and conditions (Supplementary Table S1). At the same time, minimal information is available on the influence of VD in the components of RAS at the placental level. Therefore, in this study, we aimed to explore how calcitriol transcriptionally regulates RAS-related factors in cultured human trophoblast from term and first trimester placentas, to gain a deeper understanding on the potential role of placental VD in regulating COVID-19 susceptibility and maternal blood pressure during pregnancy.
2. Results
2.1. Calcitriol and Calcidiol Downregulate ACE1, ACE2, TMPRSS2 and Renin Gene Expression in Cultured Syncytiotrophoblast from Human Placentas
To investigate the effect of VD upon RAS components in the human placenta, we cultured primary trophoblast from healthy term placentas. These cells undergo spontaneous differentiation within 24–48 hours of culture, fusing into syncytiotrophoblasts (STB), which represent the endocrinologically most active placental cell lineage [33]. Cultured trophoblast cells from term placenta were allowed to differentiate into syncytiotrophoblasts within 24 hours of culture and further incubated for an additional 24 hours in the presence of increasing calcitriol concentrations (0.1–100 nM) or 0.1 % ethanol as its vehicle. Afterward, RNA was extracted for RT-qPCR assay. In cultured trophoblast cells, calcitriol significantly downregulated, in a concentration-dependent manner, the gene expression of ACE1, ACE2 and renin (REN). TMPRS2 gene expression was also reduced, although not significantly (Figure 1).
While calcitriol is the active hormonal VD form, its precursor 25-hydroxyvitamin D3 (calcidiol) must be biotransformed into calcitriol by CYP27B1 to activate the VDR and exert its biological effects. Since the placenta expresses both CYP27B1 and the VDR [7,34,35], using calcidiol provides additional information on the viability of cultured placentas cells by reflecting intracrine calcitriol production, as previously described [35]. Therefore, as a control, we cultivated trophoblasts with a single calcidiol concentration (1 μM), which was chosen as considered previously [36]. As shown in Figure 2, and similarly as observed with calcitriol, ACE1, ACE2 and REN gene expression was inhibited by calcidiol, whereas TMPRSS2 was not modified by calcidiol (Figure 2).
2.2. Calcitriol Inhibits RAS Components in the Placental Cell Line HTR-8/SVneo
In this study, we aimed to compare the potential differential effects of calcitriol in the placenta depending on the pregnancy trimester and cellular type. HTR-8/SVneo (HTR-8) is a human trophoblast cell line derived from a first-trimester placenta, with characteristic features of extravillous invasive trophoblast (EVT). Therefore, we used this cell line to compare EVTs and STB placental cells. As seen in Figure 3, and similarly to placental STB, calcitriol significantly downregulated ACE1, ACE2, and REN gene expression (Figure 3). Contrary to STBs, calcitriol robustly inhibited TMPRSS2 mRNA at all concentrations tested (0.1–10 nM). ACE1 was also robustly inhibited starting from the lowest dose tested (0.1 nM), while ACE2 was significantly inhibited from 1.0 nM. Therefore, EVTs were more sensitive to calcitriol inhibitory effects upon the RAS axis than STB.
2.3. Syncytiotrophoblast Cells Exibit Greater Expression of RAS Components than HTR8 EVT Cells, While ACE1/ACE2 Ratio is Higher in the Latter
It has been reported that ACE2 protein is predominantly found in the STB layer of chorionic villi, as compared to EVT or other placental cell types [23,37]. Therefore, we were interested in knowing if this difference was also valid at the mRNA level in our cultured STB and EVT cells. In addition, we compared the basal gene expression of the other RAS components. In line with a previously reported immunohistochemical study [37], we found that ACE2 gene expression was several folds more expressed in STB than in EVT HTR8 cells. This result was comparable to the other RAS components, except for ACE1, which was found within a similar range (Table 1). However, the ACE1/ACE2 ratio was higher in HTR8 cells (7.4 x 10−2) than that calculated for STB cells (9.4 x 10−5).
2.4. Calcitriol Reduces ACE1/ACE2 Ratio
An increased expression of ACE1 relative to ACE2 has been associated with inflammation and various pathologies’ morbidity. Indeed, an imbalance in the ACE1/ACE2 ratio may result in increased blood pressure, inflammation, and susceptibility to COVID-19 infection [38,39]. Therefore, we analyzed the ACE1/ACE2 ratio in function to the exposure of cells to different calcitriol concentrations. Figure 4 shows the calcitriol-dependent reduction of ACE1/ACE2 ratio in STB (Figure 4a) and HTR8 EVT cells (Figure 4b). As illustrated, the ACE1/ACE2 ratio in STB cultures was downregulated progressively with increasing calcitriol concentrations. Interestingly, although this ratio was also inhibited by calcitriol in HTR8 cells, there was a divergence in the trend’s pattern, as the highest reduction on ACE1/ACE2 ratio was achieved with the lowest calcitriol concentration.
3. Discussion
It is a well-known fact that SARS-CoV-2 uses ACE2 as a receptor in host cells and that TMPRSS2 primes S protein to facilitate viral RNA entry into the cells [27], highlighting the importance of these proteins in cell infection and COVID-19 therapeutic opportunities. The human placenta expresses ACE2 and TMPRSS2, whose levels vary depending on cell type and pregnancy trimester [40]. While conflicting information on the transmission of SARS-CoV-2 from mother to child exists, it is now well recognized that this virus can vertically infect the fetus by bypassing placental barriers, albeit at a low rate [41,42]. Importantly, STB is highly susceptible to SARS-CoV-2, representing the primary target of infection among placental cells, most probably due to the high levels of ACE2 and TMPRSS2; thus, supporting SARS-CoV-2 efficient entry and replication [41,43,44]. Accordingly, ACE2 knockdown in cultured placental cells abolished viral infection, drastically decreasing the SARS-CoV-2 genome presence in supernatants and cell lysates [43]. Similarly, the SARS-CoV-2 infection of STB cells was prevented by using anti-ACE2 antibodies [44]. This background strongly supports ACE2 and TMPRSS2-targeting as a strategy to prevent placental infection and vertical transmission of the virus. In the present study, we show that calcitriol and its precursor calcidiol significantly downregulated ACE2 gene expression both in primary STB and HTR8 cell cultures, firmly suggesting that intracrine and exogenous calcitriol can diminish the risk of viral entry into trophoblast cells, placental SARS-CoV-2 infection, and consequently, fetal infection. Our results are supported by an in silico analysis identifying the VDR as a putative repressor of ACE2 gene expression [45]. Moreover, calcitriol also diminished TMPRSS2 gene expression in first trimester HTR8 cells. Notably, in in vitro and experimental animal studies, TMPRSS2 KO contributed to reduced viral lung replication, low proinflammatory reaction, and mild lung pathology [46], while the functional inhibition of this protease blocked SARS-CoV-2 entry [27], demonstrating a positive defensive outcome through the suppression of TMPRSS2. Interestingly, TMPRSS2 is an androgen-stimulated gene, containing multiple androgen response elements upstream of its gene transcription start site [47,48], partially explaining the male bias in susceptibility to severe COVID-19 disease and mortality [49,50]. Accordingly, TMPRSS2 expression has been found significantly higher in placentas from male fetus compared to those from female [51]. Thus, its downregulation by calcitriol might have special positive implications for male offspring vulnerability, deserving further studies.
If our results are replicated in other cell types, they could contribute to explain the acknowledged preventive role of VD sufficiency upon COVID-19 infection and severity [28,29]. However, we did not anticipate the outcomes on TMPRSS2 and ACE2 gene expression, given the contrasting information reported in different tissues and conditions, with ACE2 being either upregulated or downregulated by VD derivatives (Supplementary Table S1). Nevertheless, our findings complement other studies postulating additional mechanisms by which active VD metabolites can impede viral entry. For instance, computational and functional analyses have demonstrated the potential for VD derivatives to physically bind ACE2 and TMPRSS2, thereby affecting their ability to recognize and prime the SARS-Cov2 S protein [52,53]. Similarly, molecular simulations have shown the feasibility of the bonding between VD derivatives and the spike protein, stabilizing it in the locked conformation, thus inhibiting its interaction with the host receptor [53,54]. Moreover, VD hydroxymetabolites have shown the potential to inhibit SARS-CoV-2 infection by restricting its replication cycle, through targeting SARS-CoV-2 replication enzymes [55]. Altogether, this information identifies VD active compounds as potential natural therapeutic agents for preventing placental SARS-CoV-2 infection.
On the other hand, it is well established that VD limits RAS activity by inhibiting renin expression [9,10,11]. Renin is released by the kidneys into the bloodstream when blood pressure drops. Then, renin catalytically cleaves circulating angiotensinogen to form Ang I, which ACE1 converts into Ang II, a potent vasoconstricting peptide hormone that efficiently raises blood pressure. Notably, experimental studies in mice have shown that renin from placental origin is released into maternal circulation and can cause hypertension [56]. In addition, it has been reported that the risk of developing hypertensive disorders of pregnancy in patients who tested positive for COVID-19 is over 70% higher than in those who did not [57], which is in line with the described association between severe COVID-19 and hypertension [58]. Therefore, an equilibrated placental RAS regulation, emphasizing renin expression, is crucial for an adequate pregnancy outcome, especially under the threat of COVID-19 infection. In this regard, we found in this study that REN gene expression was significantly downregulated by calcitriol in STBs and EVT cells. Both types of placental cells play distinct and important roles in the placenta, STBs primarily contributing to hormonogenesis, nutrient exchange, and immune regulation, while EVTs are involved in establishing the placental-uterine interface and maternal spiral arteries remodeling. The fact that calcitriol downregulated placental REN gene expression might be associated with its capacity to lower maternal blood pressure, which is in line with previous findings from our laboratory showing a negative correlation between cord blood calcitriol and maternal systolic and diastolic blood pressure in a cohort of patients with urinary tract infections [18]. While hypertension is associated with maternal mortality and perinatal morbidity, the causative mechanisms are not yet fully clarified. However, several conditions have been associated with this disorder, including placental ischemia, abnormal EVT invasion of spiral arteries, endothelial dysfunction, and increased production of placental vasoconstrictor factors [59]. Additional risk factors are placental and systemic inflammation, characterized by exacerbated inflammatory cytokines production [60]. Notably, the cytokine storm is a characteristic feature of COVID-19 infection, while placental inflammation and malperfusion have been reported in SARS-CoV-2-infected placentas [61]. Thus, the ability of calcitriol to restrain inflammation by downregulating placental inflammatory cytokines production [2] and to inhibit REN gene expression in trophoblast cells, as shown herein, may help to regulate blood pressure both under normal and COVID-19 infection conditions. Our results concur with Chen Chun-yan et al., who described that the treatment with calcitriol or calcidiol significantly reduced REN gene expression in cultured decidual epithelial cells [62].
An imbalance in RAS and its ACE1/ACE2 components has been proposed as a driver of COVID-19 pathobiology [38,63]. Indeed, SARS-CoV-2 infection causes inhibition of ACE2 in infected cells, increasing Ang II signaling [64]. In our study, the ratio ACE1/ACE2 was significantly reduced by calcitriol in placental cells, showing different trends depending on the hormone concentration. This result could reflect a positive outcome in vivo, given that a high ACE1/ACE2 ratio has been recognized as a critical predictor for COVID-19 severity [63]. Although calcitriol reduced this ratio in both cell types, the observed concentration-dependent differences might be consequential to the basal expression levels of both enzymes in each cell type, being ACE2 increased in STBs as compared to HTR8 cells, while ACE1 was within a similar expression rate. Also, the robust calcitriol-dependent inhibitory effect upon ACE1 in STBs compared to HTR8 might play a role, given that EVT cells required higher calcitriol concentrations for ACE1 downregulation compared to STBs. Based on the baseline expression levels of RAS components, it appears that STBs play a more preponderant role in the RAS placental system than EVT cells.
Previously, as an extrapolation of VD inhibiting the RAS, it had been postulated that VD restricts SARS-CoV-2 viral entry through the downregulation of ACE2 expression [32,65]. However, contradictory and limited information exists on ACE2 transcriptional regulation by calcitriol (Supplementary Table S1). At the same time, there is no information on the effects of VD derivatives on ACE2 and other RAS components’ expression at the feto-maternal unit, aside from one study regarding REN in decidua [62]. Thus, to our knowledge, this is the first report showing the transcriptional regulation of placental RAS components by VD natural derivatives in two different cell types.
While ACE2 helps to regulate blood pressure by shifting the balance towards vasodilators and anti-hypertensive factors such as Ang-(1-7), it also facilitates SARS-Cov-2 entry by binding to the viral S protein. These facts highlight the role of ACE2 as a double-edged sword in the context of COVID-19. In this scenario, our results showing ACE2 and TMPRSS2 downregulation by calcitriol suggest the reduction of the entry gate of SARS-CoV-2 to the placenta. At the same time, the inhibition of ACE1 and REN may account for blood pressure attenuation. The latter is supported by the reduced ACE1/ACE2 ratio in calcitriol-treated cells observed in this study. This outcome could translate into therapeutic opportunities for the prevention and mitigating effects of COVID-19.
Among the limitations of our study, the lack of RAS components protein analysis, Ang(1-7) quantification, and in vivo experiments, remain areas that warrant further exploration. In addition, since the RAS placental system is involved in placentation by participating in spiral artery remodeling, trophoblast invasion and placental angiogenesis [22], further studies are needed to understand the impact of VD as a modulator factor of placental RAS.
In summary, our results support the beneficial effect of VD sufficiency during pregnancy as a protective factor against hypertensive disorders and, possibly, COVID-19 vertical transmission.
4. Materials and Methods
4.1. Reagents
The RNA extraction solution Trizol was purchased from Life Technologies, Carlbad, USA. The LightCycler TaqMan Master reaction was bought from Roche (Roche Applied Science, IN, USA). Reverse transcription (RT) system (Maxima™ Reverse Transcriptase) was from Thermo (Thermo-Scientific, St. Louis, MO, USA). Calcidiol and calcitriol were from Sigma-Aldrich (St. Louis, MO).
4.2. Cell Cultures
The placentas from uncomplicated pregnancies were processed and cultured as previously described [33,36]. Briefly, villous tissue was enzymatically treated and trophoblasts were separated on Percoll gradients. After estimating cell viability (0.4% trypan-blue), the cells were seeded in supplemented medium (DMEM-HG, 100 U/mL penicillin, 100 mg/mL streptomycin, 10% fetal bovine serum) and maintained under standard culture conditions. Trophoblasts were incubated for 24 hours and then exposed to calcidiol (1 μM), calcitriol at different concentrations, or their vehicle (ethanol 0.1%), for one more day. The HTR8 cell line was from the American Type Culture Collection (ATCC, HTR-8/SVneo CRL-3271) and was maintained with supplemented medium. This cell line was derived from a first-trimester placenta, specifically from the cell population that grew out from chorionic villi explants. As described by ATCC, these cells express proteins typical of extravillous invasive trophoblast cells.
The study methodologies conformed to the standards set by the Declaration of Helsinki. The placentas used in this study were obtained from protocols approved by the applicable Medical Ethical Committee from the Instituto Mexicano del Seguro Social (R-2013-785-033) and the Ethical and Research Committees from Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán (REF 2221).
4.3. Reverse Transcription and Real-Time PCR Amplifications (qPCR)
The total RNA was extracted from cultured trophoblasts following instructions from the TRIzol reagent manufacturer (Invitrogen, Carlsbad, CA). RNA concentration was estimated spectrophotometrically at 260/280 nm, and 2 μg of RNA were used to synthesize cDNA using the Maxima First Strand cDNA Synthesis Kit (ThermoFisher Scientific). The cDNA was then submitted to standard qPCR conditions using the LightCycler 480 Instrument (Roche Diagnostics, Mannheim, Germany). The expression of the housekeeping gene GAPDH was used for normalization purposes. The oligonucleotides were synthesized by Integrated DNA Technologies (IDT), and their sequences and number of hydrolysis probes are described in Table 2.
4.5. Statistical Analysis
The statistical differences were calculated by one-way ANOVA, using a specialized software package (SigmaStat, Jandel Scientific). For the comparisons between the relative gene expression of RAS genes in STB and EVT, the statistical significance was calculated by the Rank Sum Test. Differences were considered statistically significant at P < 0.05.
5. Conclusions
Vitamin D metabolites inhibited the gene expression of RAS components and ACE1/ACE2 ratio in placental cells. These results have important implications for COVID-19 susceptibility and maternal blood pressure regulation, suggesting that VD supplementation may not only decrease the risk of SARS-CoV-2 infection, but also reduce the likelihood of COVID-19 worsening during pregnancy.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Vitamin D effect on RAS components in different tissues and/or conditions.
Author Contributions
Validation, methodology, investigation, formal analysis, writing-review and editing, R.V.-C.; validation, methodology, investigation, formal analysis, writing-review and editing, J.G.-Q. and A.O.-O.; methodology, formal analysis and visualization, writing-review and editing, E.A.; writing-review and editing, F.L.; conceptualization, validation, methodology, investigation, formal analysis, visualization, writing-original draft preparation, supervision, project administration and funding acquisition, L.D. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Annual Acquisition Program from the Departamento de Biología de la Reproducción Dr. Carlos Gual Castro, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, REF 2221 to L.D. The funders had no role in the study design, analysis and interpretation of the data, writing of the manuscript or the decision to submit the article for publication.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the article [and/or] its Supplementary Materials.
Acknowledgments
R.V.-C is a Master-degree student from Programa de Ciencias Bioquímicas, Universidad Nacional Autónoma de México (UNAM). The authors would like to thank Dr. David Barrera for donating the HTR8 cell line.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Colotta, F.; Jansson, B.; Bonelli, F. Modulation of inflammatory and immune responses by vitamin D. J Autoimmun 2017, 85, 78–97. [Google Scholar] [CrossRef] [PubMed]
- Diaz, L.; Noyola-Martinez, N.; Barrera, D.; Hernandez, G.; Avila, E.; Halhali, A.; Larrea, F. Calcitriol inhibits TNF-alpha-induced inflammatory cytokines in human trophoblasts. J Reprod Immunol 2009, 81, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.Q.; Kaplan, A.T.; Lagishetty, V.; Ouyang, Y.B.; Ouyang, Y.; Simmons, C.F.; Equils, O.; Hewison, M. Vitamin D and the regulation of placental inflammation. J Immunol 2011, 186, 5968–5974. [Google Scholar] [CrossRef]
- Aranow, C. Vitamin D and the immune system. J Investig Med 2011, 59, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Kaplan, A.T.; Low, J.; Nguyen, L.; Liu, G.Y.; Equils, O.; Hewison, M. Vitamin D induces innate antibacterial responses in human trophoblasts via an intracrine pathway. Biol Reprod 2009, 80, 398–406. [Google Scholar] [CrossRef]
- Barrera, D.; Avila, E.; Hernandez, G.; Halhali, A.; Biruete, B.; Larrea, F.; Diaz, L. Estradiol and progesterone synthesis in human placenta is stimulated by calcitriol. J Steroid Biochem Mol Biol 2007, 103, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Barrera, D.; Avila, E.; Hernandez, G.; Mendez, I.; Gonzalez, L.; Halhali, A.; Larrea, F.; Morales, A.; Diaz, L. Calcitriol affects hCG gene transcription in cultured human syncytiotrophoblasts. Reprod Biol Endocrinol 2008, 6, 3. [Google Scholar] [CrossRef]
- Stephanou, A.; Ross, R.; Handwerger, S. Regulation of human placental lactogen expression by 1,25-dihydroxyvitamin D3. Endocrinology 1994, 135, 2651–2656. [Google Scholar] [CrossRef]
- Li, Y.C.; Kong, J.; Wei, M.; Chen, Z.F.; Liu, S.Q.; Cao, L.P. 1,25-Dihydroxyvitamin D(3) is a negative endocrine regulator of the renin-angiotensin system. J Clin Invest 2002, 110, 229–238. [Google Scholar] [CrossRef]
- Li, Y.C. Vitamin D regulation of the renin-angiotensin system. J Cell Biochem 2003, 88, 327–331. [Google Scholar] [CrossRef]
- Li, Y.C.; Qiao, G.; Uskokovic, M.; Xiang, W.; Zheng, W.; Kong, J. Vitamin D: a negative endocrine regulator of the renin-angiotensin system and blood pressure. J Steroid Biochem Mol Biol 2004, 89-90, 387–392. [Google Scholar] [CrossRef]
- Forman, J.P.; Scott, J.B.; Ng, K.; Drake, B.F.; Suarez, E.G.; Hayden, D.L.; Bennett, G.G.; Chandler, P.D.; Hollis, B.W.; Emmons, K.M.; et al. Effect of vitamin D supplementation on blood pressure in blacks. Hypertension 2013, 61, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Kristal-Boneh, E.; Froom, P.; Harari, G.; Ribak, J. Association of calcitriol and blood pressure in normotensive men. Hypertension 1997, 30, 1289–1294. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Hanni, A.; Lithell, H.; Hvarfner, A.; Sorensen, O.H.; Ljunghall, S. Vitamin D is related to blood pressure and other cardiovascular risk factors in middle-aged men. Am J Hypertens 1995, 8, 894–901. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Lithell, H.; Skarfors, E.; Wide, L.; Ljunghall, S. Reduction of blood pressure by treatment with alphacalcidol. A double-blind, placebo-controlled study in subjects with impaired glucose tolerance. Acta Med Scand 1988, 223, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Wengle, B.; Wide, L.; Ljunghall, S. Reduction of blood pressure during long-term treatment with active vitamin D (alphacalcidol) is dependent on plasma renin activity and calcium status. A double-blind, placebo-controlled study. Am J Hypertens 1989, 2, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, M.; Begerow, B.; Minne, H.W.; Nachtigall, D.; Hansen, C. Effects of a short-term vitamin D(3) and calcium supplementation on blood pressure and parathyroid hormone levels in elderly women. J Clin Endocrinol Metab 2001, 86, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Olmos-Ortiz, A.; Olivares-Huerta, A.; Garcia-Quiroz, J.; Avila, E.; Halhali, A.; Quesada-Reyna, B.; Larrea, F.; Zaga-Clavellina, V.; Diaz, L. Cord Serum Calcitriol Inversely Correlates with Maternal Blood Pressure in Urinary Tract Infection-Affected Pregnancies: Sex-Dependent Immune Implications. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Grover, S.; Brandt, J.S.; Reddy, U.M.; Ananth, C.V. Chronic hypertension, perinatal mortality and the impact of preterm delivery: a population-based study. BJOG 2022, 129, 572–579. [Google Scholar] [CrossRef]
- Tamanna, S.; Lumbers, E.R.; Morosin, S.K.; Delforce, S.J.; Pringle, K.G. ACE2: a key modulator of the renin-angiotensin system and pregnancy. Am J Physiol Regul Integr Comp Physiol 2021, 321, R833–R843. [Google Scholar] [CrossRef]
- Marques, F.Z.; Pringle, K.G.; Conquest, A.; Hirst, J.J.; Markus, M.A.; Sarris, M.; Zakar, T.; Morris, B.J.; Lumbers, E.R. Molecular characterization of renin-angiotensin system components in human intrauterine tissues and fetal membranes from vaginal delivery and cesarean section. Placenta 2011, 32, 214–221. [Google Scholar] [CrossRef]
- Pringle, K.G.; Tadros, M.A.; Callister, R.J.; Lumbers, E.R. The expression and localization of the human placental prorenin/renin-angiotensin system throughout pregnancy: roles in trophoblast invasion and angiogenesis? Placenta 2011, 32, 956–962. [Google Scholar] [CrossRef] [PubMed]
- Taglauer, E.; Benarroch, Y.; Rop, K.; Barnett, E.; Sabharwal, V.; Yarrington, C.; Wachman, E.M. Consistent localization of SARS-CoV-2 spike glycoprotein and ACE2 over TMPRSS2 predominance in placental villi of 15 COVID-19 positive maternal-fetal dyads. Placenta 2020, 100, 69–74. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat Rev Mol Cell Biol 2022, 23, 3–20. [Google Scholar] [CrossRef]
- Hosier, H.; Farhadian, S.F.; Morotti, R.A.; Deshmukh, U.; Lu-Culligan, A.; Campbell, K.H.; Yasumoto, Y.; Vogels, C.B.; Casanovas-Massana, A.; Vijayakumar, P.; et al. SARS-CoV-2 infection of the placenta. J Clin Invest 2020, 130, 4947–4953. [Google Scholar] [CrossRef]
- Molina, R.L.; Tsai, T.C.; Dai, D.; Soto, M.; Rosenthal, N.; Orav, E.J.; Figueroa, J.F. Comparison of Pregnancy and Birth Outcomes Before vs During the COVID-19 Pandemic. JAMA Netw Open 2022, 5, e2226531. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280 e278. [Google Scholar] [CrossRef]
- Moukayed, M. A Narrative Review on the Potential Role of Vitamin D(3) in the Prevention, Protection, and Disease Mitigation of Acute and Long COVID-19. Curr Nutr Rep 2023, 12, 215–223. [Google Scholar] [CrossRef]
- Ashique, S.; Gupta, K.; Gupta, G.; Mishra, N.; Singh, S.K.; Wadhwa, S.; Gulati, M.; Dureja, H.; Zacconi, F.; Oliver, B.G.; et al. Vitamin D-A prominent immunomodulator to prevent COVID-19 infection. Int J Rheum Dis 2023, 26, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Gilani, S.J.; Bin-Jumah, M.N.; Nadeem, M.S.; Kazmi, I. Vitamin D attenuates COVID-19 complications via modulation of proinflammatory cytokines, antiviral proteins, and autophagy. Expert Rev Anti Infect Ther 2022, 20, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Vanegas-Cedillo, P.E.; Bello-Chavolla, O.Y.; Ramirez-Pedraza, N.; Rodriguez Encinas, B.; Perez Carrion, C.I.; Jasso-Avila, M.I.; Valladares-Garcia, J.C.; Hernandez-Juarez, D.; Vargas-Vazquez, A.; Antonio-Villa, N.E.; et al. Serum Vitamin D Levels Are Associated With Increased COVID-19 Severity and Mortality Independent of Whole-Body and Visceral Adiposity. Front Nutr 2022, 9, 813485. [Google Scholar] [CrossRef] [PubMed]
- Arboleda, J.F.; Urcuqui-Inchima, S. Vitamin D Supplementation: A Potential Approach for Coronavirus/COVID-19 Therapeutics? Front Immunol 2020, 11, 1523. [Google Scholar] [CrossRef] [PubMed]
- Kliman, H.J.; Nestler, J.E.; Sermasi, E.; Sanger, J.M.; Strauss, J.F., 3rd. Purification, characterization, and in vitro differentiation of cytotrophoblasts from human term placentae. Endocrinology 1986, 118, 1567–1582. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Gu, Y.; Zhao, S.; Sun, J.; Groome, L.J.; Wang, Y. Expressions of vitamin D metabolic components VDBP, CYP2R1, CYP27B1, CYP24A1, and VDR in placentas from normal and preeclamptic pregnancies. Am J Physiol Endocrinol Metab 2012, 303, E928–935. [Google Scholar] [CrossRef]
- Diaz, L.; Sanchez, I.; Avila, E.; Halhali, A.; Vilchis, F.; Larrea, F. Identification of a 25-hydroxyvitamin D3 1alpha-hydroxylase gene transcription product in cultures of human syncytiotrophoblast cells. J Clin Endocrinol Metab 2000, 85, 2543–2549. [Google Scholar] [CrossRef]
- Olmos-Ortiz, A.; Garcia-Quiroz, J.; Avila, E.; Caldino-Soto, F.; Halhali, A.; Larrea, F.; Diaz, L. Lipopolysaccharide and cAMP modify placental calcitriol biosynthesis reducing antimicrobial peptides gene expression. Am J Reprod Immunol 2018, 79, e12841. [Google Scholar] [CrossRef]
- Lye, P.; Dunk, C.E.; Zhang, J.; Wei, Y.; Nakpu, J.; Hamada, H.; Imperio, G.E.; Bloise, E.; Matthews, S.G.; Lye, S.J. ACE2 Is Expressed in Immune Cells That Infiltrate the Placenta in Infection-Associated Preterm Birth. Cells 2021, 10. [Google Scholar] [CrossRef]
- Pagliaro, P.; Penna, C. ACE/ACE2 Ratio: A Key Also in 2019 Coronavirus Disease (COVID-19)? Front Med (Lausanne) 2020, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Botto, L.; Lonati, E.; Russo, S.; Cazzaniga, E.; Bulbarelli, A.; Palestini, P. Effects of PM2.5 Exposure on the ACE/ACE2 Pathway: Possible Implication in COVID-19 Pandemic. Int J Environ Res Public Health 2023, 20. [Google Scholar] [CrossRef] [PubMed]
- Bloise, E.; Zhang, J.; Nakpu, J.; Hamada, H.; Dunk, C.E.; Li, S.; Imperio, G.E.; Nadeem, L.; Kibschull, M.; Lye, P.; et al. Expression of severe acute respiratory syndrome coronavirus 2 cell entry genes, angiotensin-converting enzyme 2 and transmembrane protease serine 2, in the placenta across gestation and at the maternal-fetal interface in pregnancies complicated by preterm birth or preeclampsia. Am J Obstet Gynecol 2021, 224, 298 e291–298 e298. [Google Scholar] [CrossRef]
- Valdespino-Vazquez, M.Y.; Helguera-Repetto, C.A.; Leon-Juarez, M.; Villavicencio-Carrisoza, O.; Flores-Pliego, A.; Moreno-Verduzco, E.R.; Diaz-Perez, D.L.; Villegas-Mota, I.; Carrasco-Ramirez, E.; Lopez-Martinez, I.E.; et al. Fetal and placental infection with SARS-CoV-2 in early pregnancy. J Med Virol 2021, 93, 4480–4487. [Google Scholar] [CrossRef] [PubMed]
- Patane, L.; Morotti, D.; Giunta, M.R.; Sigismondi, C.; Piccoli, M.G.; Frigerio, L.; Mangili, G.; Arosio, M.; Cornolti, G. Vertical transmission of coronavirus disease 2019: severe acute respiratory syndrome coronavirus 2 RNA on the fetal side of the placenta in pregnancies with coronavirus disease 2019-positive mothers and neonates at birth. Am J Obstet Gynecol MFM 2020, 2, 100145. [Google Scholar] [CrossRef] [PubMed]
- Ruan, D.; Ye, Z.W.; Yuan, S.; Li, Z.; Zhang, W.; Ong, C.P.; Tang, K.; Ka Ki Tam, T.T.; Guo, J.; Xuan, Y.; et al. Human early syncytiotrophoblasts are highly susceptible to SARS-CoV-2 infection. Cell Rep Med 2022, 3, 100849. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Neil, J.A.; Tan, J.P.; Rudraraju, R.; Mohenska, M.; Sun, Y.B.Y.; Walters, E.; Bediaga, N.G.; Sun, G.; Zhou, Y.; et al. A placental model of SARS-CoV-2 infection reveals ACE2-dependent susceptibility and differentiation impairment in syncytiotrophoblasts. Nat Cell Biol 2023, 25, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Glinsky, G.V. Tripartite Combination of Candidate Pandemic Mitigation Agents: Vitamin D, Quercetin, and Estradiol Manifest Properties of Medicinal Agents for Targeted Mitigation of the COVID-19 Pandemic Defined by Genomics-Guided Tracing of SARS-CoV-2 Targets in Human Cells. Biomedicines 2020, 8. [Google Scholar] [CrossRef]
- Iwata-Yoshikawa, N.; Okamura, T.; Shimizu, Y.; Hasegawa, H.; Takeda, M.; Nagata, N. TMPRSS2 Contributes to Virus Spread and Immunopathology in the Airways of Murine Models after Coronavirus Infection. J Virol 2019, 93. [Google Scholar] [CrossRef]
- Lin, B.; Ferguson, C.; White, J.T.; Wang, S.; Vessella, R.; True, L.D.; Hood, L.; Nelson, P.S. Prostate-localized and androgen-regulated expression of the membrane-bound serine protease TMPRSS2. Cancer Res 1999, 59, 4180–4184. [Google Scholar]
- Shen, L.W.; Mao, H.J.; Wu, Y.L.; Tanaka, Y.; Zhang, W. TMPRSS2: A potential target for treatment of influenza virus and coronavirus infections. Biochimie 2017, 142, 1–10. [Google Scholar] [CrossRef]
- Abate, B.B.; Kassie, A.M.; Kassaw, M.W.; Aragie, T.G.; Masresha, S.A. Sex difference in coronavirus disease (COVID-19): a systematic review and meta-analysis. BMJ Open 2020, 10, e040129. [Google Scholar] [CrossRef]
- Zhang, J.J.; Dong, X.; Liu, G.H.; Gao, Y.D. Risk and Protective Factors for COVID-19 Morbidity, Severity, and Mortality. Clin Rev Allergy Immunol 2023, 64, 90–107. [Google Scholar] [CrossRef]
- Shook, L.L.; Bordt, E.A.; Meinsohn, M.C.; Pepin, D.; De Guzman, R.M.; Brigida, S.; Yockey, L.J.; James, K.E.; Sullivan, M.W.; Bebell, L.M.; et al. Placental Expression of ACE2 and TMPRSS2 in Maternal Severe Acute Respiratory Syndrome Coronavirus 2 Infection: Are Placental Defenses Mediated by Fetal Sex? J Infect Dis 2021, 224, S647–S659. [Google Scholar] [CrossRef]
- Song, Y.; Qayyum, S.; Greer, R.A.; Slominski, R.M.; Raman, C.; Slominski, A.T.; Song, Y. Vitamin D3 and its hydroxyderivatives as promising drugs against COVID-19: a computational study. J Biomol Struct Dyn 2022, 40, 11594–11610. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, S.; Slominski, R.M.; Raman, C.; Slominski, A.T. Novel CYP11A1-Derived Vitamin D and Lumisterol Biometabolites for the Management of COVID-19. Nutrients 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Shoemark, D.K.; Colenso, C.K.; Toelzer, C.; Gupta, K.; Sessions, R.B.; Davidson, A.D.; Berger, I.; Schaffitzel, C.; Spencer, J.; Mulholland, A.J. Molecular Simulations suggest Vitamins, Retinoids and Steroids as Ligands of the Free Fatty Acid Pocket of the SARS-CoV-2 Spike Protein*. Angew Chem Int Ed Engl 2021, 60, 7098–7110. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, S.; Mohammad, T.; Slominski, R.M.; Hassan, M.I.; Tuckey, R.C.; Raman, C.; Slominski, A.T. Vitamin D and lumisterol novel metabolites can inhibit SARS-CoV-2 replication machinery enzymes. Am J Physiol Endocrinol Metab 2021, 321, E246–E251. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, E.; Ishida, J.; Sugiyama, F.; Horiguchi, H.; Murakami, K.; Fukamizu, A. Hypertension induced in pregnant mice by placental renin and maternal angiotensinogen. Science 1996, 274, 995–998. [Google Scholar] [CrossRef]
- Tamara Siblini, M.B., Sanjana Kulkarni, Muhammad Aslam. Coronavirus Disease 2019 (COVID-19) and Hypertensive Disorders of Pregnancy: Is the Placenta the Problem? 2023. [CrossRef]
- Gomez, J.; Albaiceta, G.M.; Garcia-Clemente, M.; Lopez-Larrea, C.; Amado-Rodriguez, L.; Lopez-Alonso, I.; Hermida, T.; Enriquez, A.I.; Herrero, P.; Melon, S.; et al. Angiotensin-converting enzymes (ACE, ACE2) gene variants and COVID-19 outcome. Gene 2020, 762, 145102. [Google Scholar] [CrossRef]
- Granger, J.P.; Alexander, B.T.; Bennett, W.A.; Khalil, R.A. Pathophysiology of pregnancy-induced hypertension. Am J Hypertens 2001, 14, 178S–185S. [Google Scholar] [CrossRef]
- Alexander, B.T.; Cockrell, K.L.; Massey, M.B.; Bennett, W.A.; Granger, J.P. Tumor necrosis factor-alpha-induced hypertension in pregnant rats results in decreased renal neuronal nitric oxide synthase expression. Am J Hypertens 2002, 15, 170–175. [Google Scholar] [CrossRef]
- Schwartz, D.A.; Baldewijns, M.; Benachi, A.; Bugatti, M.; Collins, R.R.J.; De Luca, D.; Facchetti, F.; Linn, R.L.; Marcelis, L.; Morotti, D.; et al. Chronic Histiocytic Intervillositis With Trophoblast Necrosis Is a Risk Factor Associated With Placental Infection From Coronavirus Disease 2019 (COVID-19) and Intrauterine Maternal-Fetal Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Transmission in Live-Born and Stillborn Infants. Arch Pathol Lab Med 2021, 145, 517–528. [Google Scholar] [CrossRef]
- Chen, C.Y.; Zheng, L.B.; Wang, Y.L.; Li, X.; Zhang, X.Q.; Cao, B. [Regulatory Effect of Vitamin D on Renin Expression at Maternal-Fetal Interface]. Sichuan Da Xue Xue Bao Yi Xue Ban 2022, 53, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Rysz, S.; Al-Saadi, J.; Sjostrom, A.; Farm, M.; Campoccia Jalde, F.; Platten, M.; Eriksson, H.; Klein, M.; Vargas-Paris, R.; Nyren, S.; et al. COVID-19 pathophysiology may be driven by an imbalance in the renin-angiotensin-aldosterone system. Nat Commun 2021, 12, 2417. [Google Scholar] [CrossRef] [PubMed]
- Sriram, K.; Insel, P.A. A hypothesis for pathobiology and treatment of COVID-19: The centrality of ACE1/ACE2 imbalance. Br J Pharmacol 2020, 177, 4825–4844. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.J.; Chang, H.S.; Yang, Y.P.; Lin, T.W.; Lai, W.Y.; Lin, Y.Y.; Chang, C.C. The role of micronutrient and immunomodulation effect in the vaccine era of COVID-19. J Chin Med Assoc 2021, 84, 821–826. [Google Scholar] [CrossRef]
Figure 1.
The gene expression of ACE1, ACE2, TMPRSS2 and REN is downregulated by calcitriol in a concentration-dependent manner in human placental syncytiotrophoblasts. Cultured trophoblast cells from term human placentas were allowed to differentiate into syncytiotrophoblasts within 24 hours of culture. They were further incubated for another 24 hours in the presence of increasing calcitriol concentrations or its vehicle (0.1% ethanol). Afterward, RNA was extracted and gene expression was analyzed by RT-qPCR using specific primers for the human genes (a) ACE1, (b) ACE2, (c) REN and (d) TMPRSS2. Results were normalized against the gene expression of GAPDH, and vehicle values were set to 1 in all cases for normalization. At least 4 different cell cultures from different placentas were used. Bars represent the mean ± SEM. * P < 0.05).
Figure 1.
The gene expression of ACE1, ACE2, TMPRSS2 and REN is downregulated by calcitriol in a concentration-dependent manner in human placental syncytiotrophoblasts. Cultured trophoblast cells from term human placentas were allowed to differentiate into syncytiotrophoblasts within 24 hours of culture. They were further incubated for another 24 hours in the presence of increasing calcitriol concentrations or its vehicle (0.1% ethanol). Afterward, RNA was extracted and gene expression was analyzed by RT-qPCR using specific primers for the human genes (a) ACE1, (b) ACE2, (c) REN and (d) TMPRSS2. Results were normalized against the gene expression of GAPDH, and vehicle values were set to 1 in all cases for normalization. At least 4 different cell cultures from different placentas were used. Bars represent the mean ± SEM. * P < 0.05).
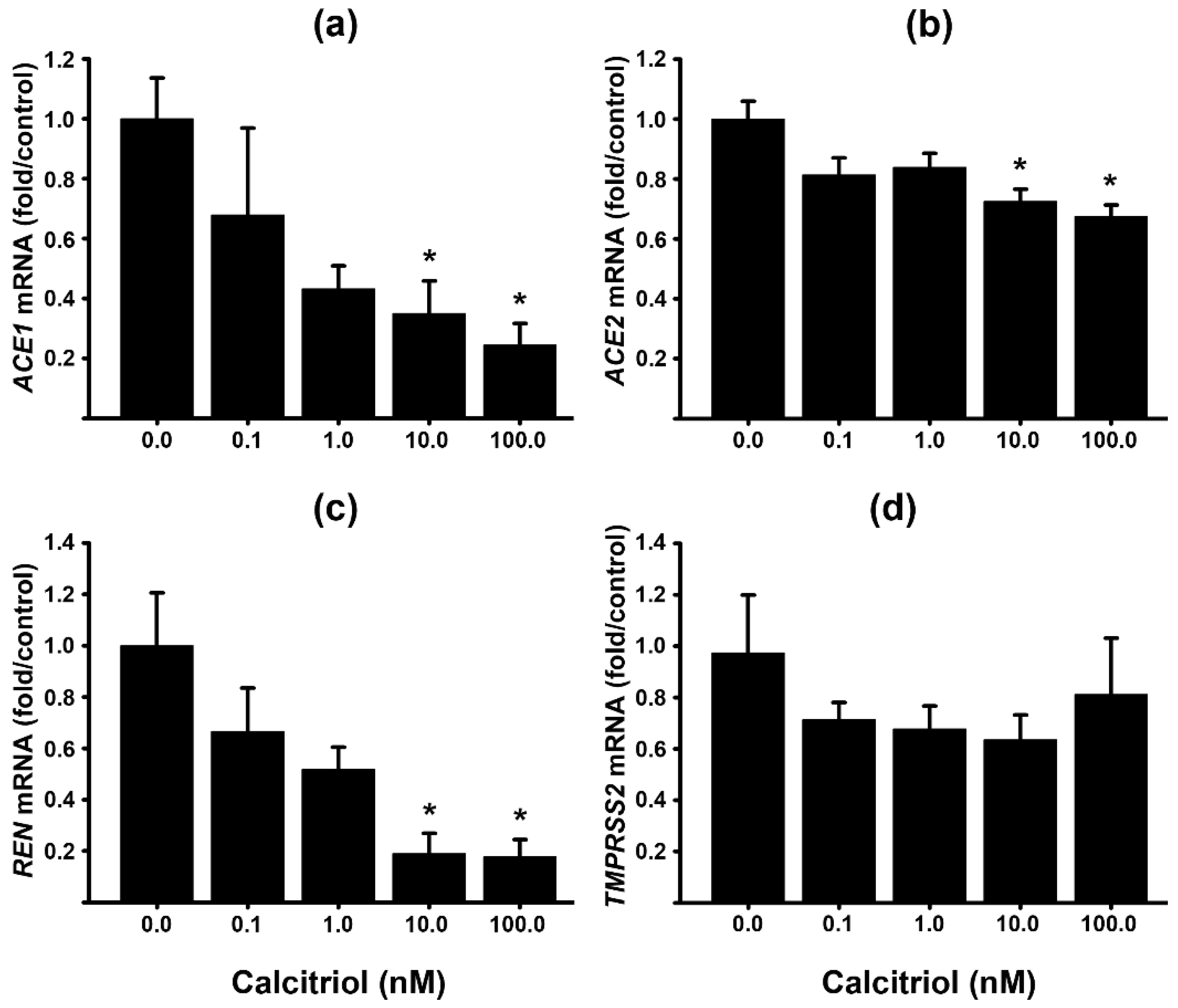
Figure 2.
Calcidiol, the precursor of calcitriol, reduces RAS components gene expression in cultured human syncytiotrophoblasts. Cultured trophoblasts were allowed to differentiate into syncytiotrophoblasts for 24 hours, and then were incubated in the presence of 1 calcidiol (μM) or its vehicle (0.1% ethanol) for additional 24 hours. The day after, RNA was extracted and gene expression was analyzed by RT-qPCR using specific primers for the RAS components. Results were normalized against the gene expression of GAPDH, and vehicle values were set to 1 (the dotted line representing control values). Three different cell cultures were used. Bars represent the mean ± SEM. * P < 0.05 vs. control.
Figure 2.
Calcidiol, the precursor of calcitriol, reduces RAS components gene expression in cultured human syncytiotrophoblasts. Cultured trophoblasts were allowed to differentiate into syncytiotrophoblasts for 24 hours, and then were incubated in the presence of 1 calcidiol (μM) or its vehicle (0.1% ethanol) for additional 24 hours. The day after, RNA was extracted and gene expression was analyzed by RT-qPCR using specific primers for the RAS components. Results were normalized against the gene expression of GAPDH, and vehicle values were set to 1 (the dotted line representing control values). Three different cell cultures were used. Bars represent the mean ± SEM. * P < 0.05 vs. control.
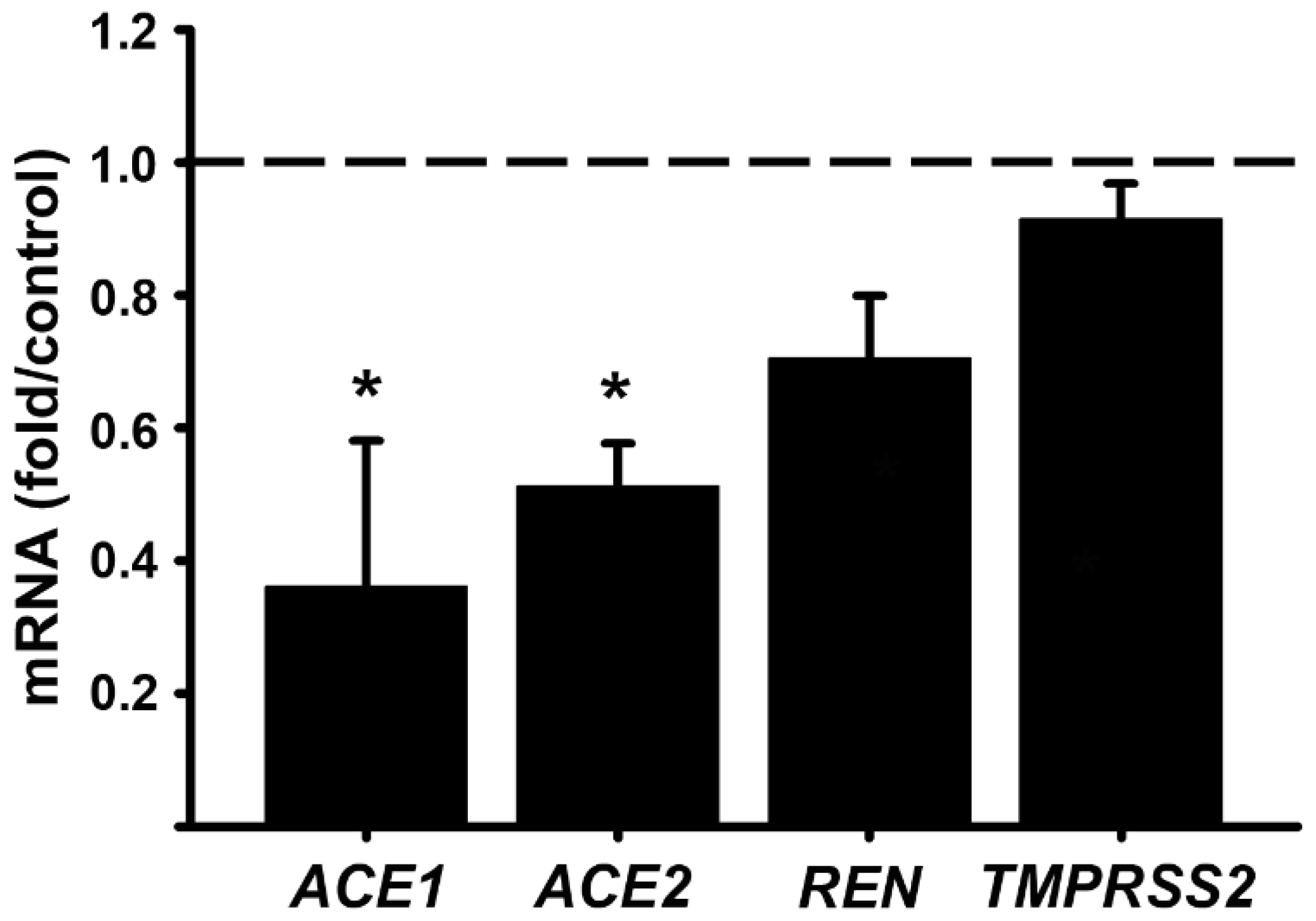
Figure 3.
Calcitriol inhibits the gene expression of RAS components in the placental EVT cell line HTR8. Cultured HTR8 cells were incubated with increasing calcitriol concentrations or its vehicle (0.1% ethanol) for 24 hours. The day after, RNA was extracted and gene expression was analyzed by RT-qPCR using specific primers for the human genes (a) ACE1, (b) ACE2, (c) REN, and (d) TMPRSS2. Results were normalized against GAPDH, used as a housekeeping gene, and vehicle values were set to 1 for normalization. N = 3. Bars represent the mean ± SEM. * P < 0.05.
Figure 3.
Calcitriol inhibits the gene expression of RAS components in the placental EVT cell line HTR8. Cultured HTR8 cells were incubated with increasing calcitriol concentrations or its vehicle (0.1% ethanol) for 24 hours. The day after, RNA was extracted and gene expression was analyzed by RT-qPCR using specific primers for the human genes (a) ACE1, (b) ACE2, (c) REN, and (d) TMPRSS2. Results were normalized against GAPDH, used as a housekeeping gene, and vehicle values were set to 1 for normalization. N = 3. Bars represent the mean ± SEM. * P < 0.05.
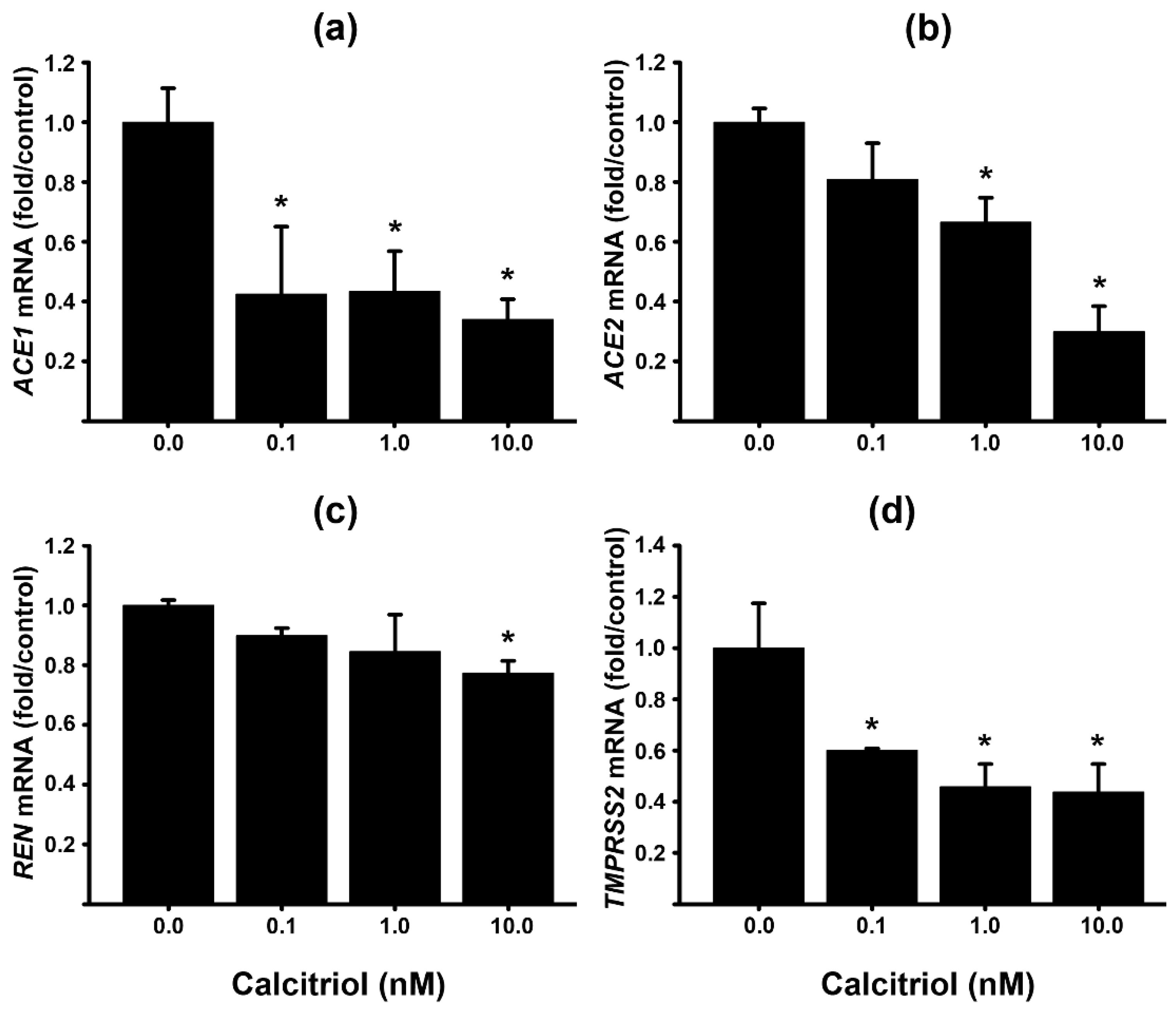
Figure 4.
ACE1/ACE2 ratio in human placental cells. The ACE1/ACE2 ratio, based on the gene expression of both enzymes, was calculated in syncytiotrophoblasts (STB) (a) or in HTR8 cells (b) treated with different calcitriol concentrations.
Figure 4.
ACE1/ACE2 ratio in human placental cells. The ACE1/ACE2 ratio, based on the gene expression of both enzymes, was calculated in syncytiotrophoblasts (STB) (a) or in HTR8 cells (b) treated with different calcitriol concentrations.
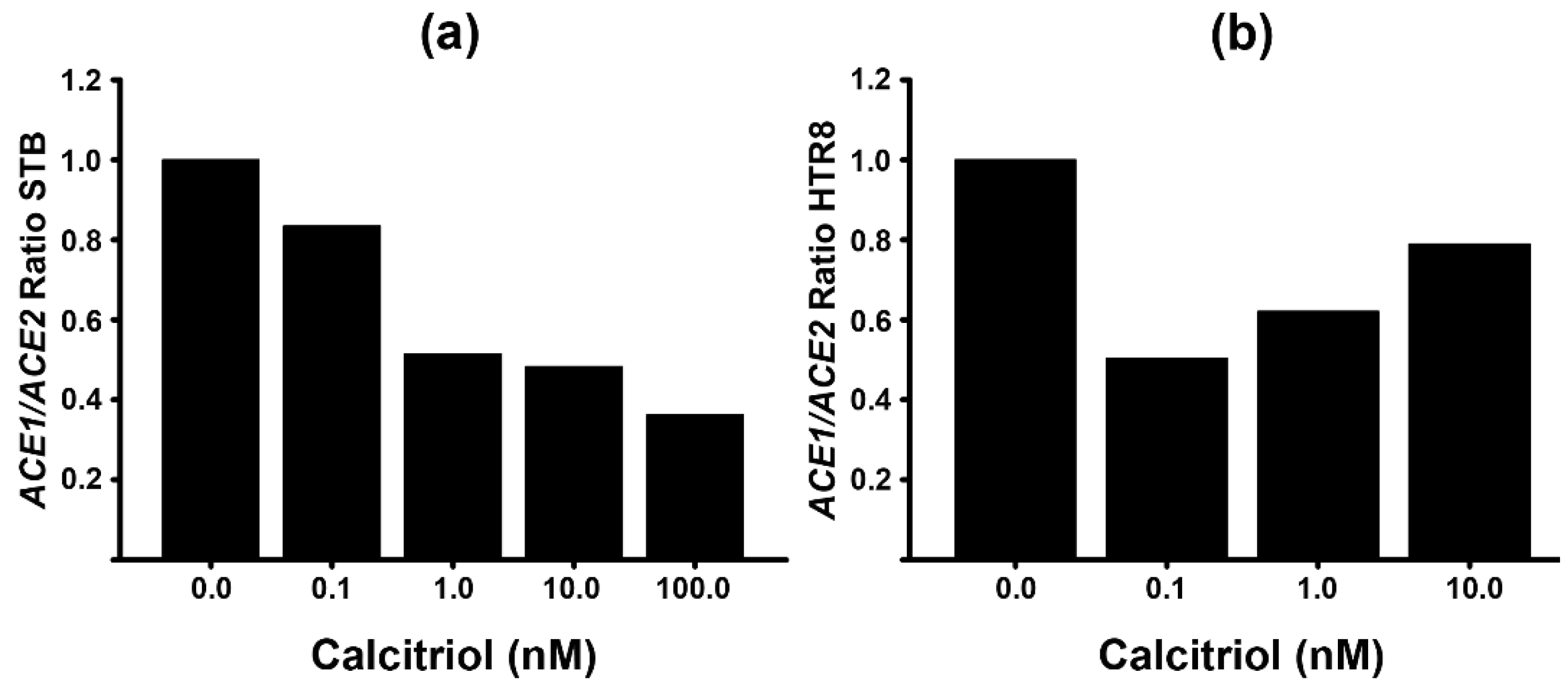

Table 1.
Basal gene expression of RAS genes in syncytiotrophoblast cells (STB) and EVT (HTR8).
| Gene | STB | HTR8 | P |
|---|---|---|---|
| ACE1 | 2.7 x 10−6 ± 7.2 x 10−7 | 1.7 x 10−6 ± 3.4 x 10−7 | 0.58 |
| ACE2 | 2.9 x 10−2 ± 4.6 x 10−3 | 2.3 x 10−5 ± 3.8 x 10−6 | <0.001 |
| TMPRSS2 | 1.8 x 10−2 ± 8.8 x 10−3 | 1.8 x 10−4 ± 6.5 x 10−5 | <0.001 |
| REN | 3.2 x 10−2 ± 5.3 x 10−3 | 8.7 x 10−5 ± 3.3 x 10−5 | <0.001 |
The gene expression is presented relative to that of GAPDH. Results are shown as the mean ± SEM from a minimum of 3 experiments, each with triplicate replicates. Statistical significance was determined using the Rank Sum Test, comparing the gene expression of each gene in STB and HTR8.
Table 2.
Primers sequence and probes for RT-qPCR.
| Gene | Upper Primer | Lower Primer | Probe Number |
|---|---|---|---|
| ACE1 | ctgctcatctgctgggagac | ttgtctgggaaaggcaccac | 33 |
| ACE2 | ttctgtcacccgattttcaa | tcccaacaatcgtgagtgc | 4 |
| TMPRSS2 | acctgatcacaccagccatg | tcaccctggcaagaatcgac | 4 |
| GAPDH | agccacatcgctgagacac | gcccaatacgaccaaatcc | 60 |
| REN | tacctttggtctcccgacag | ttgagggcattctcttgagg | 77 |
Hydrolysis probes are from the Universal Probe Library (Roche).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



