
Submitted:
06 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
Perilla frutense var. acuta (Lamiaceae) has been used to treat indigestion, asthma, and allergies in traditional medicine. In this study, luteolin 7-O-diglucuronide (1), apigenin 7-O-diglucuronide (2), and rosmarinic acid (3) were isolated from the leaves of P. frutescens var. acuta. Various approaches were performed to investigate the anti-inflammatory activity of the isolated compounds (1-3) and their working mechanisms. In silico docking simulation demonstrated that 1-3 would work as a PPAR-α/δ/γ agonist, and in vitro PPAR-α/δ/γ transcriptional assay showed that the Perilla extract and 3 increased PPAR-α luciferase activity [1.71 and 1.61 times of the control (PPAR-α + PPRE, p < 0.001)]. In the NF-κB luciferase assay, 1 suppressed NF-κB activity the most (56.83 % at 5 µM; 74.96 % at 10 µM; 79.86 % at 50 µM). In addition, 1 and 2 inhibited the mRNA expression of NF-κB target genes, including Il6, Mcp1, and Tnfa, at 50 µM, and 3 suppressed the genes at the mRNA level in a dose-dependent manner. We report that 1 and 2 exert anti-inflammatory effects through NF-κB inhibition, and PPAR-α/NF-κB signaling pathway is related to the anti-inflammatory activity of 3.
Keywords:
Perilla frutescens var. acuta
; Anti-inflammation
; Peroxisome proliferator-activated receptor
; Nu-clear factor kappa B
; Flavonoid diglucuronide
1. Introduction
Perilla frutescens (L.) Britton var. acuta Kudo, which belongs to Lamiaceae family, is native to South and East Asia and widely cultivated in Korea, China, India, Japan, and Vietnam as a functional food, a spice, and the main source of perilla oil [1,2]. While P. frutescens var. acuta has been known to treat indigestion, fever, asthma, and anxiety in traditional medicine [2], modern scientists have also reported anti-allergic, antimicrobial, antioxidant, and cytotoxic activities of P. frutescens var. acuta [3]. This plant contains various classes of compounds, including fatty acids, flavonoids, phenolic acids, triterpenoids, and volatile oils [3]. Although previous studies have reported that P. frutescens shows anti-inflammatory effects through inhibition of tumor necrosis factor-α (TNF-α), cyclooxygenase-2 (COX-2), interleukin-6 (IL-6), IL-8, and inducible nitric oxide synthase (iNOS), active ingredients and their mechanisms of action are not well understood yet [4,5].
Inflammation is a coordination of multiple signaling pathways to regulate the inflammatory mediators from the blood when harmful stimuli, including pathogens, damaged cells, or irritants, invade the body [6]. As peroxisome proliferator-activated receptor (PPAR) and nuclear factor kappa B (NF-κB) are transcription factors involved in the inflammatory responses [7], PPARs inhibit NF-κB binding to DNA promoter regions and then its target gene expression [8]. Thus, PPAR agonists can suppress the release of pro-inflammatory mediators, including IL-6, monocyte chemoattractant protein-1 (MCP-1), and TNF-α [9].
In this study, luteolin 7-O-diglucuronide (1), apigenin 7-O-diglucuronide (2), and rosmarinic acid (3) were isolated from the leaves of P. frutescens var. acuta. The anti-inflammatory activity of 1-3 was investigated through in silico docking simulation against PPAR-α/δ/γ, in vitro PPAR-α/δ/γ transcriptional luciferase assay, NF-κB luciferase assay, and measurement of NF-κB target gene expression.
2. Results
2.1. Isolation of the compounds
Compounds 1-3 were isolated from the Perilla water extract using various column chromatography techniques. Identification of structures was conducted based on 1D/2D NMR data (1H, 13C, DEPT 135, HSQC, COSY, HMBC, and NOESY spectra), optical rotation, UV, and HR-MS analyses. The isolated compounds were thus confirmed as luteolin 7-O-diglucuronide (1), apigenin 7-O-diglucuronide (2), and rosmarinic acid (3) (Figure 1). 1 and 2 have been first isolated from the Elodea Species [10], and later they have been also found in P. frutescens var. acuta [11,12]. Flavonoids are one of the abundant secondary metabolites in P. frutescens and apigenin and its derivatives have been known as one of the main flavonoids in this species [13]. 3 has been isolated from Rosmarinus officinalis L. for the first time, and then also found in the leaves of P. frutescens var. acuta in 1982 [14]. 3 is generally found in the species of the Boraginaceae and the Lamiaceae families [15].
2.2. In silico docking simulation
In silico molecular docking simulation shows the interaction of the receptor and ligand in a preferred position with a minimum free binding energy for a stable complex and helps to predict the action of ligand as agonist or antagonist [16]. In this docking study were shown the non-covalent interactions between the isolated compounds and PPAR-α/δ/γ. The complexes with lower energies mean to be more stable in simulation work [17].
The docking energies of 1-3 with PPAR-α were lower than that of control (eicosapentaenoic acid, EPA) or equal in Autodock Vina, Autodock 4, and Dock 6 (Table 1). Especially, 1 (-13.2 Kcal/mol) showed the highest affinity for the binding site in Autodock 4 and 3 consistently showed modest binding affinity in three systems. We also investigated the pharmacophores contributing to the interactions of the ligands and PPAR-α. Figure 2a demonstrates molecular docking models of 1-3 and EPA (a pan-activator of PPARs) [18]. The green color arrow indicates the hydrogen bond donor, the red color the hydrogen bond acceptor, and the yellow color the hydrophobic interaction or Van der Walls force. 1 and 2 formed several hydrophobic interactions and hydrogen bonds with the key amino acids such as Ala333A, Cys276, and Thr279 of PPAR-α, leading to a strong binding relationship. 3 also showed a strong binding affinity caused by several hydrogen bonds and hydrophobic interactions. On the other hand, EPA formed a hydrogen bond with Cys276 and weak hydrophobic interactions induced by the alkyl chain in the ligand. Based on the in silico docking energy and pharmacophore studies, 1-3 would act as a PPAR-α agonist.
The interactions between 1-3 and PPAR-δ were more stabilized than that of EPA in Autodock Vina, Autodock 4, and Dock 6 (Table 1). In addition, the best docking poses of the ligands with PPAR-δ were investigated (Figure 2b). Although there were a few hydrogen bonds between PPAR-δ and 1/2, a stable binding affinity was found due to the hydrophobic interactions with the amino acids including Phe282, Thr289, and Met453 in PPAR-δ. On the other hand, 3 formed an ionic interaction with Lys367, several hydrophobic interactions, and hydrogen bonds, leading to a strong binding affinity with PPAR-δ. Therefore, 1-3 are expected to act as PPAR-δ agonists.
1 and 2 showed more stable binding affinities with PPAR-γ than the control (EPA) in Autodock 4 and Dock6 (Table 1). Moreover, 3 demonstrated the higher docking energies to the PPAR-γ receptor than the control in all three systems. In pharmacophore analysis (Figure 2c), 1-3 owned more than five hydrogen bonds, which is generally considered a high number in docking simulation, and 2 and 3 also formed an ionic interaction with Arg288 of the receptor. As a result, 1-3 are potential PPAR-γ agonists, and thus can show pharmacological activities in the related biological systems.
2.3. Cell viability
As cytotoxicity is a critical obstacle for activity study, the cytotoxicity of the compounds was examined before performing the in vitro assays. Raw 264.7 cells were treated with the extract (0, 10, 50, 100 µg/mL) and the compounds (0, 10, 50, 100 µM) for 24 hours and the cell viability was determined using an MTT assay (Figure 3 and Table S2). Although the extract showed 80.3 and 81.8 % cell viability at high concentrations (50 and 100 µg/mL), the cell viability of 1-3 exceeded 94 % in all concentrations against Raw 264.7 cells.
2.4. PPAR-α/δ/γ transcriptional activity
To verify the validity of the results from docking simulation, 1-3 (10 µM each) were screened for their in vitro PPAR-α/δ/γ transactivation potency using Ac2F cells. As shown in Figure 4a and Table S3, WY14643 (a well-known PPAR-α agonist), the Perilla water extract, and 3 increased PPAR-α promoter luciferase reporter activity (1.58, 1.71, and 1.61 times, respectively; p < 0.001) compared to the control (PPAR-α + PPRE group), and the extract and 3 showed higher PPAR-α agonistic activity than WY14643. The in silico docking analysis also suggested that 3 owns the strongest binding affinity as an PPAR-α agonist in Autodock Vina and Dock6. On the other hand, as shown in Figure 4b and Table S4, PPAR-δ transcriptional activity of the extract and compounds was compared with GW501516 (a widely used PPAR-δ agonist), but none of them showed higher potency than GW501516 [1.43 times stronger than the control (PPAR-δ + PPRE), p < 0.05], and the order of in silico binding energies between the ligands and PPAR-δ did not exactly match the in vitro PPAR-δ transcriptional potency. PPAR-γ transactivation activity was also measured with the water extract and compounds using Ac2F cells transiently transfected with pcDNA/PPAR-γ and PPRE (Figure 4c and Table S5). Although PPAR-γ agonistic potency of 2 was 1.17 times stronger than the control (PPAR-γ+PPRE), it did not exceed the activity of rosiglitazone (a well-known PPAR-γ agonist, 1.54 times stronger than the control).
2.5. NF-κB transcriptional activity
Previous study has reported that PPAR-α/δ/γ inhibit NF-κB activation involved in inflammatory responses [19,20,21]. Therefore, we investigated whether the compounds could inhibit inflammation through the NF-κB signaling pathway. As shown in Figure 5 and Table S6, NF-κB-driven luciferase assay was performed using HEK293T cells and the NF-κB transcriptional activity remarkably increased after 1 µg/mL of lipopolysaccharide (LPS) induction for 6 hours. On the other hand, compound treatment (5, 10, 50 µM) suppressed the NF-κB transcriptional activity in a dose-dependent manner and 1 (% inhibition: 56.8 % at 5 µM; 75.0 % at 10 µM; 79.9 % at 50 µM) showed the strongest activity, followed by 3 (% inhibition: 43.3 % at 5 µM; 63.6 % at 10 µM; 71.1 % at 50 µM) and 2 (% inhibition: 55.9 % at 5 µM; 61.2 % at 10 µM; 75.3 % at 50 µM). 3 has previously shown to down-regulate PPAR-γ/NF-κB-mediated signaling pathway in a rat myocardial tissue [22].
2.6. NF-κB target gene expression
Based on the PPAR-α/δ/γ and NF-κB luciferase assays of 1-3, the expression levels of NF-κB target genes were measured for Il6, Mcp1, and Tnfa in Raw 264.7 macrophages. As shown in Figure 6a and Table S7, the Perilla water extract significantly reduced the mRNA levels of Il6, Mcp1, and Tnfa in a dose-dependent manner. In case of 1 and 2, they notably inhibited the mRNA levels of Il6 (% inhibition: 1, 79.0 %; 2, 81.0 %) and Mcp1 (% inhibition: 1, 67.9 %; 2, 44.7 %) at 50 µM (Figure 6b,c, and Table S7). The mRNA levels of Mcp1 and Tnfa treated with 3 were strongly inhibited at a concentration of 10 µM (% inhibition: Mcp1, 53.9 %; Tnfa, 39.6 %).
3. Discussion
The anti-inflammatory effects of P. frutescens using in vivo model has been conducted in several studies. Yuan and coworkers have studied the effects of the P. frutescens extract against chronic obstructive pulmonary disease (COPD) airway inflammation in cigarette smoke/LPS-induced COPD mice [23]. They have observed significant decreases in inflammatory cell infiltration in lung tissue and production of inflammatory cytokines of the bronchoalveolar lavage fluid. Oh, et al. have also shown the protective effects of the P. frutescens var. acuta extract (EPPF) and 3 (rosmarinic acid, RA) against allergic inflammations in ovalbumin (OVA)-sensitized mouse model [24]. In the OVA-sensitized mice, the number of nasal rubbing and concentrations of IgE and histamine have been decreased by EPPF or RA administration. In addition, the expression of IL-1β, IL-6, and TNF-α has been decreased at both the mRNA and the protein levels by introducing EPPF or RA in the nasal mucosa of the OVA-sensitized mice. Along with 3, various compounds have also been tested for in vivo anti-inflammatory models. For example, tormentic acid has shown comparable activity with hydrocortisone (ID50 = 0.03 mg/ear) in its inhibitory effect on pro-inflammatory responses induced by 12-O-tetradecanoylphorbol-13-acetate (1 µg/ear) in the mouse model experiment [25].
To the best of our knowledge, this study reports the in silico simulations as a PPAR-α/δ/γ agonist and in vitro PPAR-α/δ/γ, and NF-κB transcription activities of 1 and 2 for the first time. Although they did not increase PPAR-α/δ/γ promoter luciferase reporter activity, the NF-κB transcriptional activity was inhibited by 1 and 2 in a dose-dependent manner, and 1 showed the strongest activity among the tested compounds. In addition, the mRNA expression levels of Il6, Mcp1, and Tnfa, were suppressed by 1 and 2 in Raw 264.7 cells. Rosmarinic acid (3) also inhibited NF-κB activation as a PPAR-α agonist, which is revealed for the first time in this study. Although Rajagopal and coworkers have reported that 3 acts as a PPAR-γ agonist in the docking simulation study [26], the potential of 3 as a PPAR-α/δ agonist is firstly shown in this study. It has been reported that 3 have alleviated inflammation by suppressing the TGF-β/IL-17A signaling in human adipocytes [27], and also attenuated inflammation of cardiomyocyte by initiating PPAR-γ/NF-κB signaling pathway [22].
As we demonstrated in this study, the activation of PPARs inhibits NF-κB-dependent inflammation and AMP kinase (AMPK), sirtuin 1 (SIRT1), and p300 are involved in the process [28,29,30]. In particular, PPAR activators promote AMPK, which increases SIRT1 expression and p300 phosphorylation. The activated AMPK-SIRT1/p300 signal leads to decreased acetylation of p65 subunit in the NF-κB complex and translocation into the nucleus, concluding the reduced expression of NF-kB p65 target genes, such as Il6, Il1b, and Tnfa. In addition, PPAR-α/γ agonists potentiate the IκBα expression, which is an inhibitory protein against NF-κB, and thus induce anti-inflammatory activity [31,32]. Activation of PPAR-δ also inhibits the assembly of TAK1, TAB1, and HSP27, consequently interfering with the function of p65 NF-κB [33].
A limitation of this study is that the in silico simulations of 1-3 as PPAR-α/δ/γ agonists showed inconsistency with the results from the in vitro PPAR transcriptional assay. Indeed, major limitations may be caused by limited conformations of ligand-receptor in pose prediction, effects of the solvents, or approximated scoring system [34]. Nevertheless, docking simulation helps to investigate the potential of compounds for therapeutic activities and predict ligand/target relationships at a molecular level. A better description of the behavior for ligand-receptor and refinement of docking procedures will lead to a better correlation with experimental data.
4. Materials and Methods
4.1. Plant material
The leaves of P. frutescens var. acuta were purchased from Megabiosoop, in April 2019. A voucher specimen (No. EA387) has been deposited at the Natural Product Chemistry Laboratory, College of Pharmacy, Ewha Womans University.
4.2. General experimental procedures
Optical rotation was obtained on a P-1010 polarimeter (Jasco, Tokyo, Japan) and UV spectrum was recorded on a U-3000 spectrophotometer (Hitachi, Tokyo, Japan). NMR spectrum was run on a Varian Unity Inova 400 MHz FT-NMR instrument (Agilent Technologies, CA, USA) with TMS as an internal standard and data was processed in MestReNova 9.0 (Mestrelab Research SL, Santiago de Compostela, Spain). HRESIMS was performed on Agilent 6230 Accurate-Mass TOF LC/MS system (Agilent). For column chromatography, Diaion HP-20 and Kieselgel 60 F254 (silica gel, 0.25 mm layer thickness) were purchased from Mitsubishi Chemical Co. (Tokyo, Japan) and Merck & Co. (NJ, USA), respectively. MPLC was performed using CombiFlash (Teledyne Isco Inc., NE, USA), equipped with RediSep Rf C18 column (130 g, Teledyne Isco Inc.) and RediSep Rf normal phase silica column (40g and 220g). Preparative HPLC purification was conducted using Acme 9000 system (Young Lin, Seoul, Korea) equipped with a YMC-Pack Pro C18 column (5 μm, 250 mm × 20 mm i.d., YMC Co., Kyoto, Japan).
4.3. Extraction and isolation
The dried leaves of P. frutescens var. acuta (2 kg) were extracted with water (20 L) for 15 hours at room temperature and then the extract was evaporated in vacuo at 40 °C to afford a concentrated water extract (352.8 g). The water extract was chromatographed over Diaion HP-20 using a gradient mixture (MeOH-H2O, 0:100 to 100:0) to afford the pooled fractions (Fr.1-Fr.7). Fr.4 (37.7 g) was subjected to RP-MPLC with a mixture of MeOH-H2O gradient system to give seven subfractions (Fr.4.1-Fr.4.7). 1 (2.1 g) was precipitated from Fr.4.2. A part (2.0 g) of Fr.6 (7.5 g) was subjected to RP-MPLC with a gradient mixture (MeOH-H2O, 5:95 to 100:0) and the subfraction Fr. 6.3 (125.9 mg) was purified using a preparative HPLC instrument with an isocratic solvent system (30 % MeOH, 8 mL/min) to afford 2 (tR 85.3 min. 49.4 mg). Fr. 7 (7.9 g) was subjected to MPLC with a solvent mixture (CH2Cl2-MeOH, 100:0 to 0:100) to acquire 9 subfractions (Fr.7.1-Fr.7.9). Subfraction Fr.7.4.4.5.5 (252.6 mg) was purified on a preparative HPLC instrument using an isocratic solvent system (50 % MeOH, 5mL/min) to yield 3 (tR 13.3 min. 27.6 mg).
Luteolin 7-O-diglucuronide (1): Yellow amorphous solid; [α]D20 -34.6 (c 0.1, MeOH); UV (MeOH) λmax (log ε) 254 (4.68), 347 (4.67); HRESIMS m/z 639.1194 [M+H]+ (calcd for C27H27O18); 1H NMR (pyridine-d5, Figure S1) δH 7.86 (d, H-2', J = 2.3 Hz), 7.46 (dd, H-6', J = 2.3, 8.2 Hz), 7.23 (d, H-5', J = 8.2 Hz), 7.18 (d, H-8, J = 2.0 Hz), 7.15 (d, H-6, J = 2.0 Hz), 6.83 (s, H-3), 6.04 (d ,H-1'', J = 6. 8 Hz), 5.57 (d, H-1''', J = 8.2 Hz), 4.92 (d, H-5'', J = 9.6 Hz), 4.75 (m, H-4'', H-5'''), 4.61 (m, H-2'', H-3'', H-4'''), 4.40 (t, H-3''', J =9.0 Hz), 4.27 (t, H-2''', J = 8.2 Hz); 13C NMR (pyridine-d5, Figure S2) δC 182.8 (C-4), 172.6 (C-6'''), 172.0 (C-6''), 165.3 (C-2), 163.7 (C-7), 162.7 (C-5), 157.8 (C-9), 151.8 (C-4'), 147.7 (C-3'), 122.7 (C-1'), 119.7 (C-6'), 116.8 (C-5'), 114.7 (C-2'), 107.0 (C-1'''), 106.8 (C-10), 104.0 (C-3), 100.9 (C-6), 100.3 (C-1''), 95.9 (C-8), 84.2 (C-2''), 78.2 (C-5'''), 77.9 (C-3'''), 77.6 (C-5''), 77.0 (C-3''), 76.2 (C-2'''), 73.8 (C-4'''), 72.7 (C-4'') [35].
Apigenin 7-O-diglucuronide (2): White amorphous solid; [α]D20 -62.7 (c 0.1, MeOH); UV (MeOH) λmax (log ε) 268 (4.60), 334 (4.66); HRESIMS m/z 623.1243 [M+H]+ (calcd for C27H27O17); 1H NMR (pyridine-d5, Figure S3) δH 7.83 (d, H-2', H-6', J = 7.4 Hz), 7.29 (d, H-8, J = 2 Hz), 7.20 (d, H-3', H-5', J = 7.4 Hz), 7.16 (d, H-6, J = 2 Hz), 6.81 (s, H-3), 6.09 (d, H-1'', J = 7.6 Hz), 5.57 (d, H-1''', J = 8.4 Hz), 4.94 (d, H-5'', J = 9.5 Hz), 4.74 (t , H-4'',H-5''', J = 9.5 Hz), 4.60 (m, H-2'', H-3'', H-4''', J = 7.6 Hz), 4.40 (t, H-3''', J = 9.1 Hz), 4.26 (m, H-2''', J = 9.1,8.4 Hz); 13C NMR (pyridine-d5, Figure S4) δC 182.9 (C-4), 172.6 (C-6'''), 172.1 (C-6''), 164.9 (C-2), 163.8 (C-7), 162.8 (C-4'), 162.7 (C-5), 157.8 (C-9), 129.0 (C-2' and C-6'), 116.8 (C-3' and C-5'), 107.0 (C-1'''), 106.8 (C-10), 103.9 (C-3), 101.0 (C-6), 100.3 (C-1''), 95.9 (C-8), 84.2 (C-2''), 78.2 (C-5'''), 77.8 (C-3'''), 77.6 (C-5''), 77.1 (C-3''), 76.2 (C-2'''), 73.4 (C-4'''), 72.7 (C-4'') [35].
Rosmarinic acid (3): Yellow amorphous solid; [α]D20 101.3 (c 0.07, MeOH); UV (MeOH) λmax (log ε) 328 (4.40); HRESIMS m/z 359.0767 [M-H]- (calcd for C18H17O8, 360.0764); 1H NMR (methanol-d4, Figure S5) δH 7.54 (d, H-7, J = 15.8Hz) , 7.04 (d, H-2, J = 2.7 Hz), 6.94 (dd, H-6, J = 2.7,8.4 Hz), 6.77 (d, H-5, J =8.4 Hz), 6.75 (d, H-2', J =1.8 Hz), 6.69 (d, H-5', J = 8.2 Hz), 6.62 (d, H-6', J = 1.8,8.2 Hz), 6.26 (d, H-8, J = 15.8 Hz), 5.17 (q, H-8', J = 4.1, 8.8 Hz ), 3.10 (dd, H-7', J = 4.1, 14.3 Hz), 2.99 (dd, H-7', J = 8.8, 14.3 Hz); 13C NMR (methanol-d4 , Figure S6) δC 168.6 (C-9), 149.7 (C-4), 147.5 (C-7), 146.9 (C-3), 146.2 (C-3'), 145.2 (C-4'), 129.7 (C-1'), 127.8 (C-1), 123.1 (C-6), 121.8 (C-6'), 117.6 (C-2'), 116.5 (C-5), 116.3 (C-5'), 115.2 (C-2), 114.8 (C-8), 75.3 (C-8'), 38.2(C-7') [36].
4.4. Molecular docking
Crystal structures of PPAR-α/δ/γ were obtained from RCSB PDB website [PDB ID: 1K71 (PPAR-α); 1GWX (PPAR-δ); and 3DZY (PPAR-γ)] (https://www.rcsb.org/, accessed on 16 March 2023). 3D structures of 1-3 and EPA (a positive control) were acquired from PubChem website (https://pubchem.ncbi.nlm.nih.gov/, accessed on 16 March 2023). Three programs were used for docking simulation: Autodock Vina 1.1.2 (Scripps Research, CA, USA), Autodock 4.2.6 (Scripps Research, CA, USA), and Dock6 (UCSF, CA, USA). Docking preparation was conducted to add hydrogens and assign charges of the compounds in UCSF Chimera (UCSF, CA, USA). Pharmacophore analysis was conducted using LigandScout 4.0 (inte:Ligand, Vienna, Austria) to explore possible interactions of the receptors and ligands.
4.5. Cell viability
The cell viability of Raw 264.7 macrophages was determined by 3-(4,5-dimethylthiazolyl-2)-2,5-diphenyl tetrazolium bromide (MTT) assay. At the time of 70% confluence, Raw 264.7 cells were treated with P. frutescens var. acuta extract (0, 10, 50, 100 µg/mL) or 1-3 (0, 5, 10, 50 µM) for 24 hours. After aspirating the cell culture medium, cells were incubated in DMEM with 10% FBS and 5 mg/mL MTT solution. After 1 hour of incubation, the concentration of formazan, a purple product converted from a tetrazolium salt by the viable cells, was measured using a spectrophotometer at 595 nm.
4.6. PPAR and NF-κB transcriptional activity
Luciferase assays were performed to determine the transcriptional activity of PPAR transcription factors in the Ac2F cell. Briefly, Ac2F cells were transfected with the PPRE-X3-TK-LUC plasmid (0.2 µg) with PPAR-α, PPAR-δ, or PPAR-γ expression vectors (0.1 µg) using Lipofectamine 3000 reagent (Invitrogen, CA, USA.). The cells were further treated with 1-3 or WY14643 (a known PPAR-α agonist), GW501516 (a known PPAR-δ agonist), and rosiglitazone (a known PPAR-γ agonist), respectively. The luciferase activity was measured using One-Glo Luciferase Assay System (Promega, WI, USA). After adding the luciferase substrate, the luminescence was measured using a luminescence plate reader (Berthold Technologies GmbH & Co., Bad Wildbad, Germany).
Luciferase assays were also performed to determine the transcriptional activity of NF-κB in the HEK293T cells. The cells were transfected with the NF-κB promoter-Luc plasmid (0.1 µg) for 24 hours, and co-treated with test compounds 1-3 and LPS (1 µg/mL) for 6 hours, and lysed using One-Glo Luciferase Assay System and a luminescence plate reader. The results are presented as mean ± S.E. (n=5) and each measurement was performed in triplicates. Statistical significance was tested using the one-way ANOVA/post hoc test.
4.7. NF-κB target gene expression
Raw 264.7 macrophages (Korea Cell Line Bank, Seoul, Korea) were grown in DMEM containing 4500 mg/L glucose and L-glutamine (Sigma-Aldrich, MO, USA) supplemented with 10% FBS (Sigma-Aldrich) and 1% antibiotic antimycotic solution at 37 °C and 5% CO2. Two days after confluence, P. frutescens var. acuta extract (0, 10, 50, 100 µg/mL) or 1-3 (0, 5, 10, 50 µM) dissolved in DMSO (Sigma-Aldrich) were treated for 24 hours, and 100 ng/mL of LPS (Sigma-Aldrich) for 6 hours. Then, total RNA was extracted from Raw 264.7 cells using Trizol reagent (Invitrogen, CA, USA) and cDNA was synthesized from 1 μg of total RNA using the PrimeScript II 1st strand cDNA synthesis kit (Takara, Japan). The mRNA levels of Il6, Mcp1, and Tnfa were quantified using StepOnePlus Real-time PCR System (Applied Biosystems, MA, USA) and SYBR Green PCR Master Mix (Applied Biosystems), and then normalized relative to 18S rRNA. Fold changes of gene expression were calculated by the ΔΔCt method. Specific primer sequences used are shown in Table S1.
5. Conclusions
In this study, two flavonoid diglucuronides, luteolin 7-O-diglucuronide (1) and apigenin 7-O-diglucuronide (2), and rosmarinic acid (3) were isolated from the leaves of P. frutescens var. acuta. In silico PPAR docking simulation unlocked the potential of 1-3 as PPAR agonists, and these results were evaluated using in vitro PPAR luciferase assay. In addition, the Perilla water extract and 1-3 suppressed LPS-induced upregulation of Il6, Mcp1, and Tnfa in Raw 264.7 cells, which may be mediated through the PPAR/NF-κB signaling pathway. This study provides scientific backgrounds of P. frutescens var. acuta as a functional food, which are supported by its biologically active ingredients and anti-inflammatory mechanism of action to prevent chronic inflammatory diseases.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
I.Y.: Investigation, Writing-Original Draft. S.H.: Investigation, Writing-Original Draft. H.J.J.: Investigation, Writing-Original Draft. S.G.N.: Formal analysis, Writing-Original Draft. H.Y.C.: Conceptualization, Supervision, Writing-Reviewing and Editing. Y.K.K.: Resources. S.S.: Investigation, Supervision, Writing-Original Draft. E.K.S.: Conceptualization, Supervision, Writing-Reviewing and Editing.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request.
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MIST) (No. 2021R1A2C1003350) and by Basic Science Research Program through the NRF funded by the Ministry of Education (RS-2023-00243759). This work was also supported by a research grant from Seoul Women’s University (2023-0057). The authors want to thank Ewha Drug Development Research Core Center for letting us use Agilent 6230 ToF LC/MS (Agilent, CA, USA).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Plants of the World Online. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:190343-2 (accessed on 23 August, 2023).
- Ahmed, H.M. Ethnomedicinal, phytochemical, and pharmacological investigations of Perilla frutescens (L.) Britt. Molecules, 2018; 24 , 102. [Google Scholar]
- Yu, H.; Qiu, J.-F.; Ma, L.-J.; Hu, Y.-J.; Li, P.; Wan, J.-B. Phytochemical and phytopharmacological review of Perilla frutescens L.(Labiatae), a traditional edible-medicinal herb in China. Food Chem. Toxicol. 2017; 108, 375–391. [Google Scholar]
- Ueda, H.; Yamazaki, M. Inhibition of tumor necrosis factor-α production by orally administering a perilla leaf extract. Biosci. Biotechnol. Biochem. 1997, 61, 1292–1295. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.P.; Lin, C.H.; Chen, Y.C.; Kao, S.H. Anti-inflammatory effects of Perilla frutescens leaf extract on lipopolysaccharide-stimulated Raw 264.7 cells. Mol. Med. Rep. 2014, 10, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Korbecki, J.; Bobiński, R.; Dutka, M. Self-regulation of the inflammatory response by peroxisome proliferator-activated receptors. Inflamm. Res. 2019, 68, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Ricote, M.; Li, A.C.; Willson, T.M.; Kelly, C.J.; Glass, C.K. The peroxisome proliferator-activated receptor-γ is a negative regulator of macrophage activation. Nature 1998, 391, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Mues, R. Species specific flavone glucuronides in Elodea species. Biochem. Syst. Ecol. 1983, 11, 261–265. [Google Scholar] [CrossRef]
- Aritomi, M.; Kumori, T.; Kawasaki, T. Cyanogenic glycosides in leaves of Perilla frutescens var. acuta. Phytochemistry 1985, 24, 2438–2439. [Google Scholar] [CrossRef]
- Makino, T.; Furuta, Y.; Fujii, H.; Nakagawa, T.; Wakushima, H.; Saito, K.-i.; Kano, Y. Effect of oral treatment of Perilla frutescens and its constituents on Type-I allergy in mice. Biol. Pharm. Bull. 2001, 24, 1206–1209. [Google Scholar] [CrossRef]
- Jiang, T.; Guo, K.; Liu, L.; Tian, W.; Xie, X.; Wen, S.; Wen, C. Integrated transcriptomic and metabolomic data reveal the flavonoid biosynthesis metabolic pathway in Perilla frutescens (L.) leaves. Sci. Rep. 2020; 10, 16207. [Google Scholar]
- Aritomi, M. Chemical studies on the constituents of edible plants (Part 1). Phenolic compounds in leaves of Perilla frutescens BRITTON var. acuta KUDO f. viridis MAKINO. J. Home Econ. 1982, 33, 353–359. [Google Scholar]
- Petersen, M.; Simmonds, M.S. Rosmarinic acid. Phytochemistry 2003, 62, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.H.; Stephens, E.; O'Brien, D.P.; Zhou, M. Understanding noncovalent interactions: Ligand binding energy and catalytic efficiency from ligand-induced reductions in motion within receptors and enzymes. Angew. Chem.-Int. Ed. 2004; 43, 6596–6616. [Google Scholar]
- Tanaka, N.; Zhang, X.; Sugiyama, E.; Kono, H.; Horiuchi, A.; Nakajima, T.; Kanbe, H.; Tanaka, E.; Gonzalez, F.J.; Aoyama, T. Eicosapentaenoic acid improves hepatic steatosis independent of PPAR-α activation through inhibition of SREBP-1 maturation in mice. Biochem. Pharmacol. 2010, 80, 1601–1612. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, W.; Shi, M.; Yang, Z.; Zhang, X.; Gong, P. Docosahexaenoic acid attenuates LPS-stimulated inflammatory response by regulating the PPAR-γ/NF-κB pathways in primary bovine mammary epithelial cells. Res. Vet. Sci. 2017, 112, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Barroso, E.; Eyre, E.; Palomer, X.; Vázquez-Carrera, M. The peroxisome proliferator-activated receptor β/δ (PPAR-β/δ) agonist GW501516 prevents TNF-α-induced NF-κB activation in human HaCaT cells by reducing p65 acetylation through AMPK and SIRT1. Biochem. Pharmacol. 2011, 81, 534–543. [Google Scholar] [CrossRef]
- Ramanan, S.; Kooshki, M.; Zhao, W.; Hsu, F.-C.; Robbins, M.E. PPAR-α ligands inhibit radiation-induced microglial inflammatory responses by negatively regulating NF-κB and AP-1 pathways. Free Radical Biol. Med. 2008, 45, 1695–1704. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wang, D.; Ye, L.; Li, P.; Hao, W.; Chen, X.; Ma, J.; Wang, B.; Shang, J.; Li, D. Rosmarinic acid protects against inflammation and cardiomyocyte apoptosis during myocardial ischemia/reperfusion injury by activating peroxisome proliferator-activated receptor gamma. Front. Pharmacol. 2017, 8, 456. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, X.; Fang, N.; Li, P.; Zhang, Z.; Lin, M.; Hou, Q. Perilla leaf extract (PLE) attenuates COPD airway inflammation via the TLR4/Syk/PKC/NF-κB pathway in vivo and in vitro. Front. Pharmacol. 2022, 12, 763624. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-A.; Park, C.-S.; Ahn, H.-J.; Park, Y.S.; Kim, H.-M. Effect of Perilla frutescens var. acuta Kudo and rosmarinic acid on allergic inflammatory reactions. Exp. Biol. Med. 2011, 236, 99–106. [Google Scholar]
- Banno, N.; Akihisa, T.; Tokuda, H.; Yasukawa, K.; Higashihara, H.; Ukiya, M.; Watanabe, K.; Kimura, Y.; Hasegawa, J.-i.; Nishino, H. Triterpene acids from the leaves of Perilla frutescens and their anti-inflammatory and antitumor-promoting effects. Biosci. Biotechnol. Biochem. 2004, 68, 85–90. [Google Scholar] [CrossRef]
- Rajagopal, P.; Jayaraman, S.; Jh, S.F.; Radhakrishnan, S.; Laxman, P.A.; Muthaiah, V.P.K.; Tripathi, S.C.; Gugapriya, T.; Tarnekar, A.M.; Muthiyan, G.G. Molecular docking analysis of PPAR-γ with compounds from Ocimum tenuiflorum. Bioinformation 2021, 17, 928. [Google Scholar] [CrossRef]
- Vasileva, L.V.; Savova, M.S.; Tews, D.; Wabitsch, M.; Georgiev, M.I. Rosmarinic acid attenuates obesity and obesity-related inflammation in human adipocytes. Food Chem. Toxicol. 2021, 149, 112002. [Google Scholar] [CrossRef]
- Yeung, F.; Hoberg, J.E.; Ramsey, C.S.; Keller, M.D.; Jones, D.R.; Frye, R.A.; Mayo, M.W. Modulation of NFBdependent transcription and cell survival by the SIRT1 deacetylase. EMBO J. 2004, 23, 2369–2380. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lin, Q.; Lin, R.; Zhang, J.; Ren, F.; Zhang, J.; Ji, M.; Li, Y. PPAR-α agonist fenofibrate attenuates TNF-α-induced CD40 expression in 3T3-L1 adipocytes via the SIRT1-dependent signaling pathway. Exp. Cell Res. 2013, 319, 1523–1533. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Xiao, F.; Liu, Y.; Wang, J.; Gao, H.; Rong, S.; Yao, Y.; Li, J.; Xu, G. The peroxisome proliferator-activated receptor-γ agonist pioglitazone prevents NF-κB activation in cisplatin nephrotoxicity through the reduction of p65 acetylation via the AMPK-SIRT1/p300 pathway. Biochem. Pharmacol. 2016, 101, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Delerive, P.; Gervois, P.; Fruchart, J.-C.; Staels, B. Induction of IκBα expression as a mechanism contributing to the anti-inflammatory activities of peroxisome proliferator-activated receptor-α activators. J. Biol. Chem. 2000, 275, 36703–36707. [Google Scholar]
- Scirpo, R.; Fiorotto, R.; Villani, A.; Amenduni, M.; Spirli, C.; Strazzabosco, M. Stimulation of nuclear receptor peroxisome proliferatoractivated receptor- limits NFBdependent inflammation in mouse cystic fibrosis biliary epithelium. Hepatology 2015, 62, 1551–1562. [Google Scholar] [CrossRef]
- Stockert, J.; Wolf, A.; Kaddatz, K.; Schnitzer, E.; Finkernagel, F.; Meissner, W.; Müller-Brüsselbach, S.; Kracht, M.; Müller, R. Regulation of TAK1/TAB1-mediated IL-1β signaling by cytoplasmic PPAR-β/δ. PloS One 2013, 8, e63011. [Google Scholar] [CrossRef] [PubMed]
- Pantsar, T.; Poso, A. Binding affinity via docking: Fact and fiction. Molecules 2018, 23, 1899. [Google Scholar] [CrossRef]
- Kim, J.; Kang, H.; Choi, H.; Jo, A.; Oh, D.-R.; Kim, Y.; Im, S.; Lee, S.-G.; Jeong, K.-I.; Ryu, G.-C. Aqueous extract of Perilla frutescens var. acuta relaxes the ciliary smooth muscle by increasing NO/cGMP content in vitro and in vivo. Molecules, 2018; 23, 1777. [Google Scholar]
- Aung, H.T.; Nikai, T.; Niwa, M.; Takaya, Y. Rosmarinic acid in Argusia argentea inhibits snake venom-induced hemorrhage. J. Nat. Med. 2010, 64, 482–486. [Google Scholar] [CrossRef]
Figure 1.
Structures of the isolated compounds (1-3) in this study.
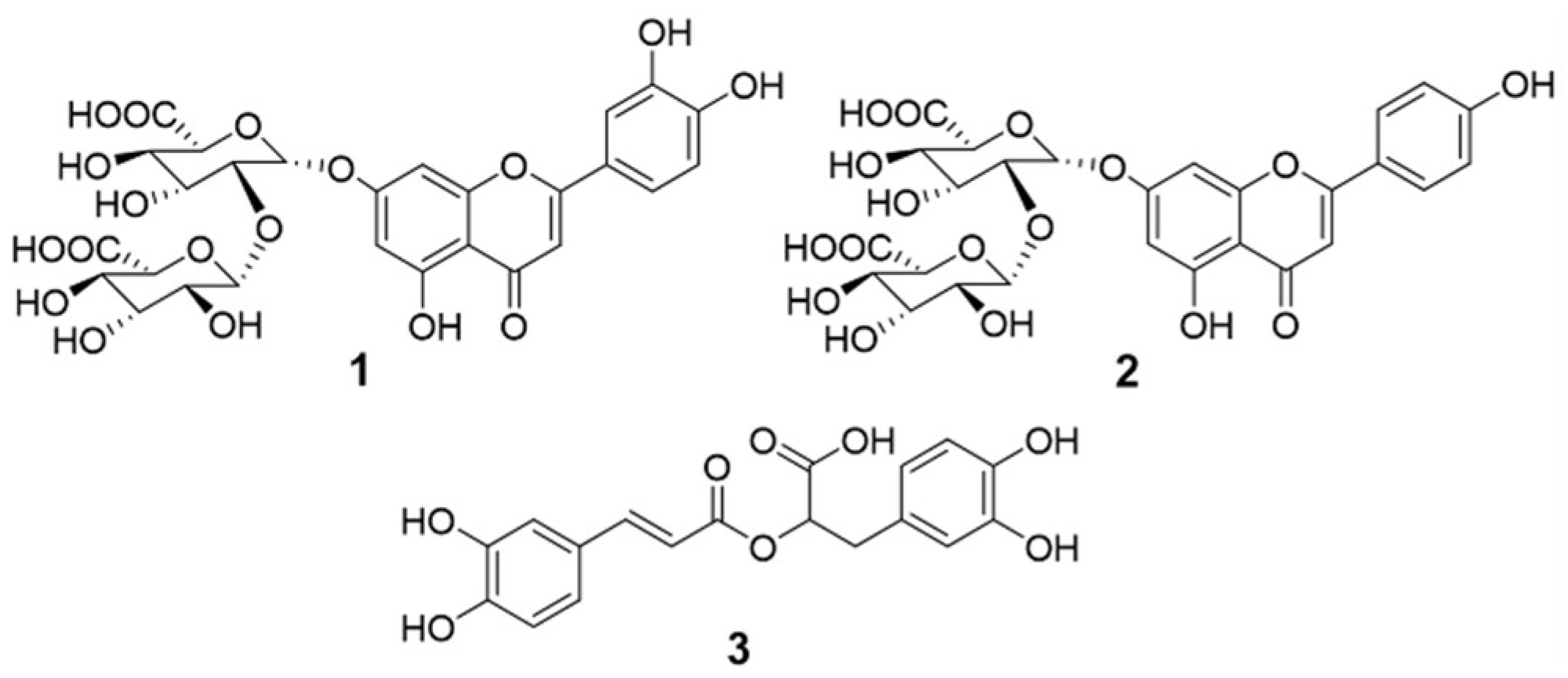
Figure 2.
Docking simulations of 1, 2, 3, and EPA against (a) PPAR-α, (b) PPAR-δ, and (c) PPAR-γ in AutoDock 4.2. The green arrow indicates the hydrogen bond donor, the red arrow the hydrogen bond acceptor, and the yellow color the hydrophobic interaction or Van der Walls force. (ALA, Alanine; ARG, Arginine; CYS, Cysteine; GLU, Glutamic acid; ILE, Isoleucine; LEU, Leucine; LYS, Lysine; MET, Methionine; PHE, Phenylalanine; SER, Serine; THR, Threonine; TRP, Tryptophan; TYR, Tyrosine; VAL, Valine.).
Figure 2.
Docking simulations of 1, 2, 3, and EPA against (a) PPAR-α, (b) PPAR-δ, and (c) PPAR-γ in AutoDock 4.2. The green arrow indicates the hydrogen bond donor, the red arrow the hydrogen bond acceptor, and the yellow color the hydrophobic interaction or Van der Walls force. (ALA, Alanine; ARG, Arginine; CYS, Cysteine; GLU, Glutamic acid; ILE, Isoleucine; LEU, Leucine; LYS, Lysine; MET, Methionine; PHE, Phenylalanine; SER, Serine; THR, Threonine; TRP, Tryptophan; TYR, Tyrosine; VAL, Valine.).
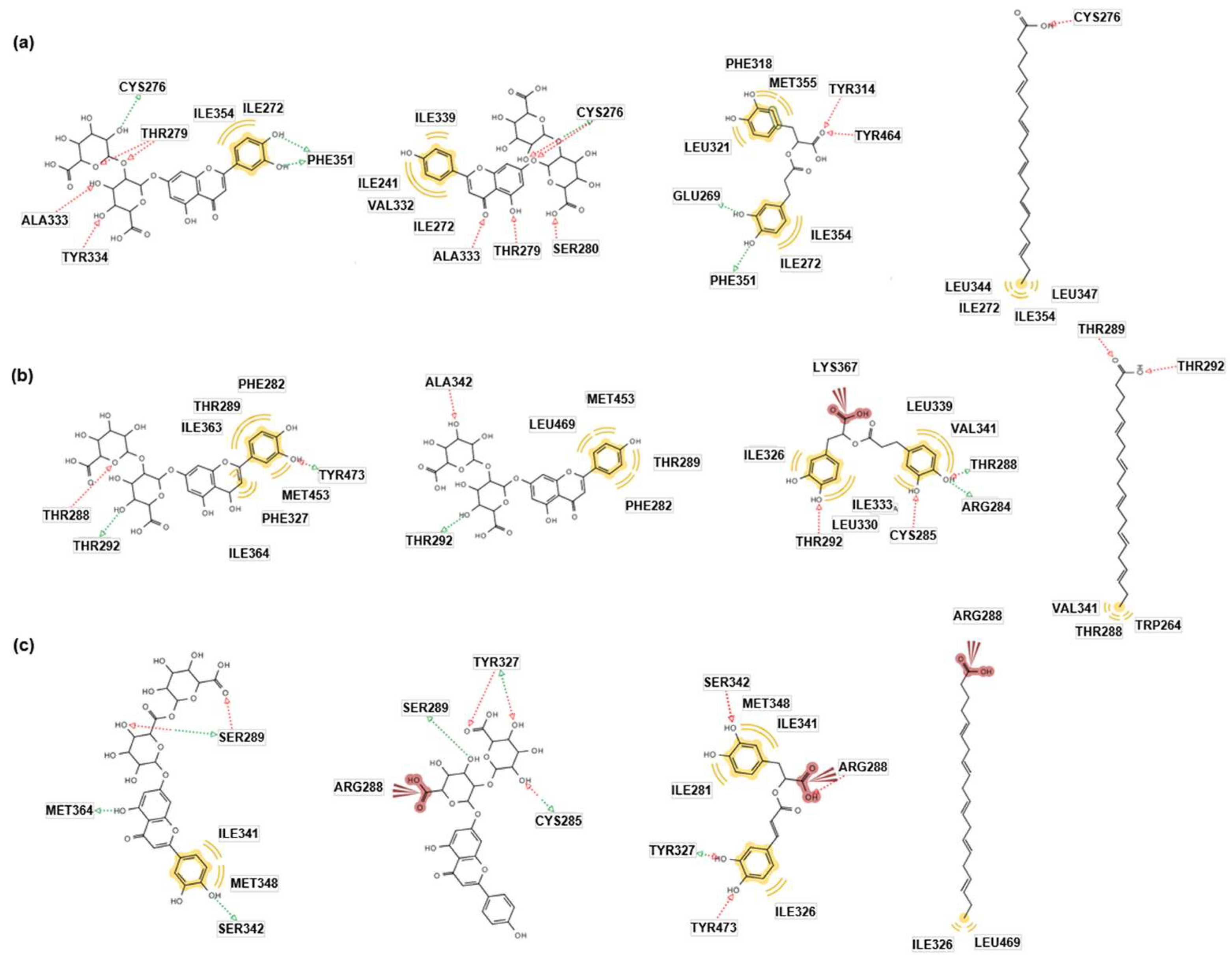
Figure 3.
Cell viability of the P. frutense var. acuta extract and the isolated compounds 1-3. (a) The Perilla water extract (0, 10, 50, 100 µg/mL). (b)-(d) 1-3 (0, 5, 10, 50 µM). Raw 264.7 cells were treated with the indicated doses of the drugs for 24 hours and cell viability was measured using MTT assay. A,B Different superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different.
Figure 3.
Cell viability of the P. frutense var. acuta extract and the isolated compounds 1-3. (a) The Perilla water extract (0, 10, 50, 100 µg/mL). (b)-(d) 1-3 (0, 5, 10, 50 µM). Raw 264.7 cells were treated with the indicated doses of the drugs for 24 hours and cell viability was measured using MTT assay. A,B Different superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different.
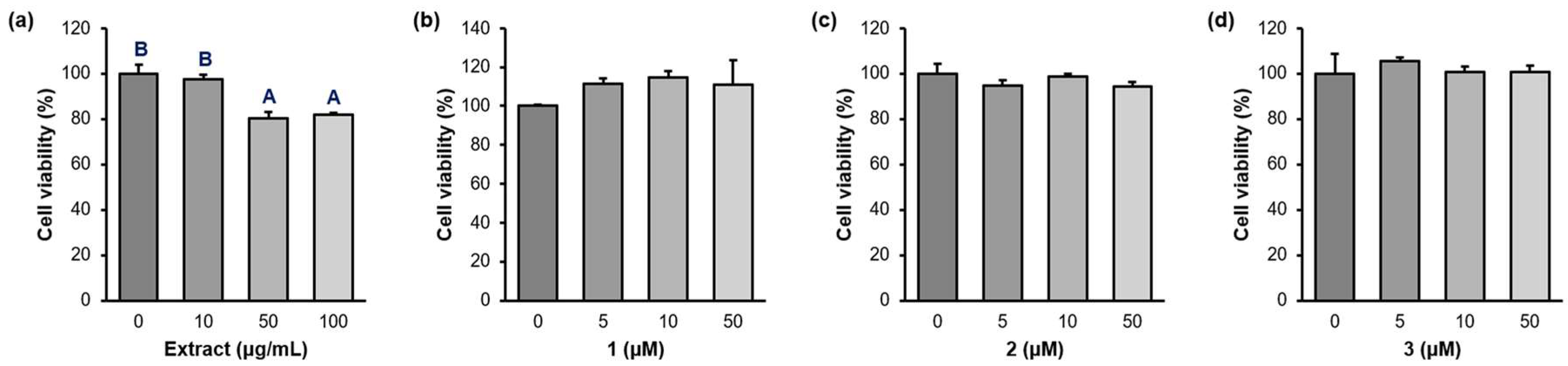
Figure 4.
PPAR-α/δ/γ transcriptional activities of the extract and 1-3 from P. frutescens var. acuta. (a) The effect of the extract and 1-3 on PPAR-α transcriptional activity was examined using the PPRE luciferase system. ###p < 0.001 vs. pcDNA + PPRE group, **p < 0.001 vs. PPAR-α + PPRE group. (b) The effect of the extract and 1-3 on PPAR-δ transcriptional activity was measured using the PPRE luciferase system. ###p < 0.001 vs. pcDNA + PPRE group. *p < 0.05 vs. PPAR-δ + PPRE group. (c) The effect of the extract and 1-3 on PPAR-γ transcriptional activity was measured using the PPRE luciferase system. ###p < 0.001 vs. pcDNA + PPRE group. **p < 0.005 vs. PPAR-γ + PPRE group.
Figure 4.
PPAR-α/δ/γ transcriptional activities of the extract and 1-3 from P. frutescens var. acuta. (a) The effect of the extract and 1-3 on PPAR-α transcriptional activity was examined using the PPRE luciferase system. ###p < 0.001 vs. pcDNA + PPRE group, **p < 0.001 vs. PPAR-α + PPRE group. (b) The effect of the extract and 1-3 on PPAR-δ transcriptional activity was measured using the PPRE luciferase system. ###p < 0.001 vs. pcDNA + PPRE group. *p < 0.05 vs. PPAR-δ + PPRE group. (c) The effect of the extract and 1-3 on PPAR-γ transcriptional activity was measured using the PPRE luciferase system. ###p < 0.001 vs. pcDNA + PPRE group. **p < 0.005 vs. PPAR-γ + PPRE group.
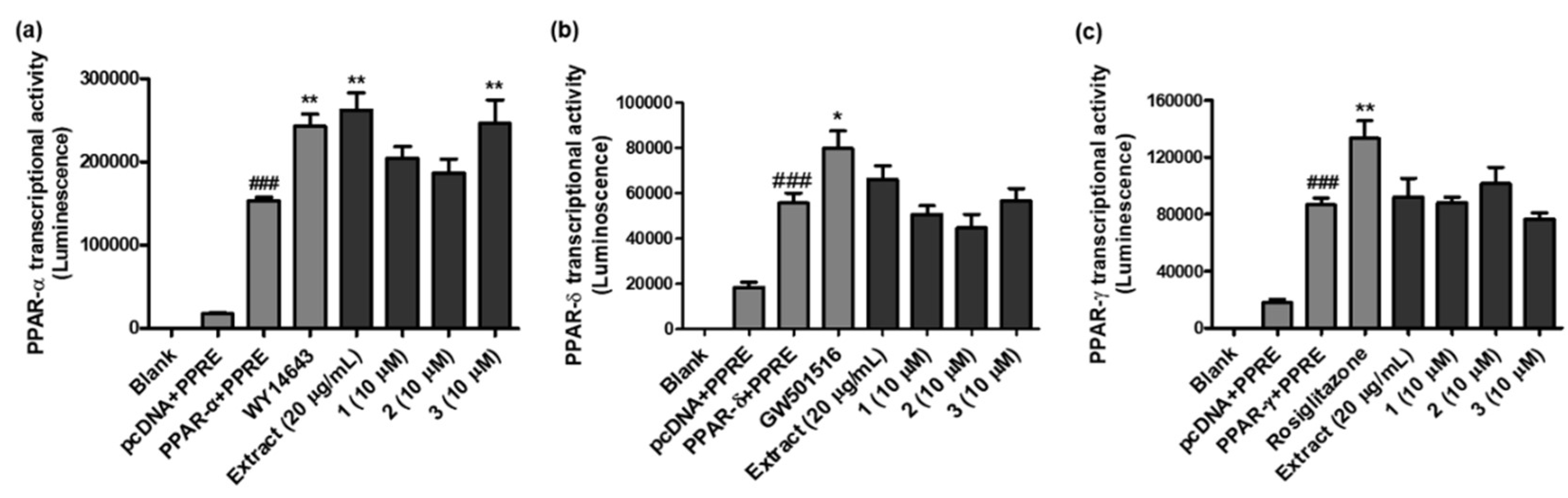
Figure 5.
NF-κB transcriptional activity of 1-3 (5, 10, 50 µM) from P. frutense var. acuta. NF-κB transcriptional activity was measured using LPS-treated HEK293T cells with or without 1-3 treatment. ###p < 0.001 vs. control, ***p < 0.001 and **p < 0.01 vs. LPS-treated group.
Figure 5.
NF-κB transcriptional activity of 1-3 (5, 10, 50 µM) from P. frutense var. acuta. NF-κB transcriptional activity was measured using LPS-treated HEK293T cells with or without 1-3 treatment. ###p < 0.001 vs. control, ***p < 0.001 and **p < 0.01 vs. LPS-treated group.
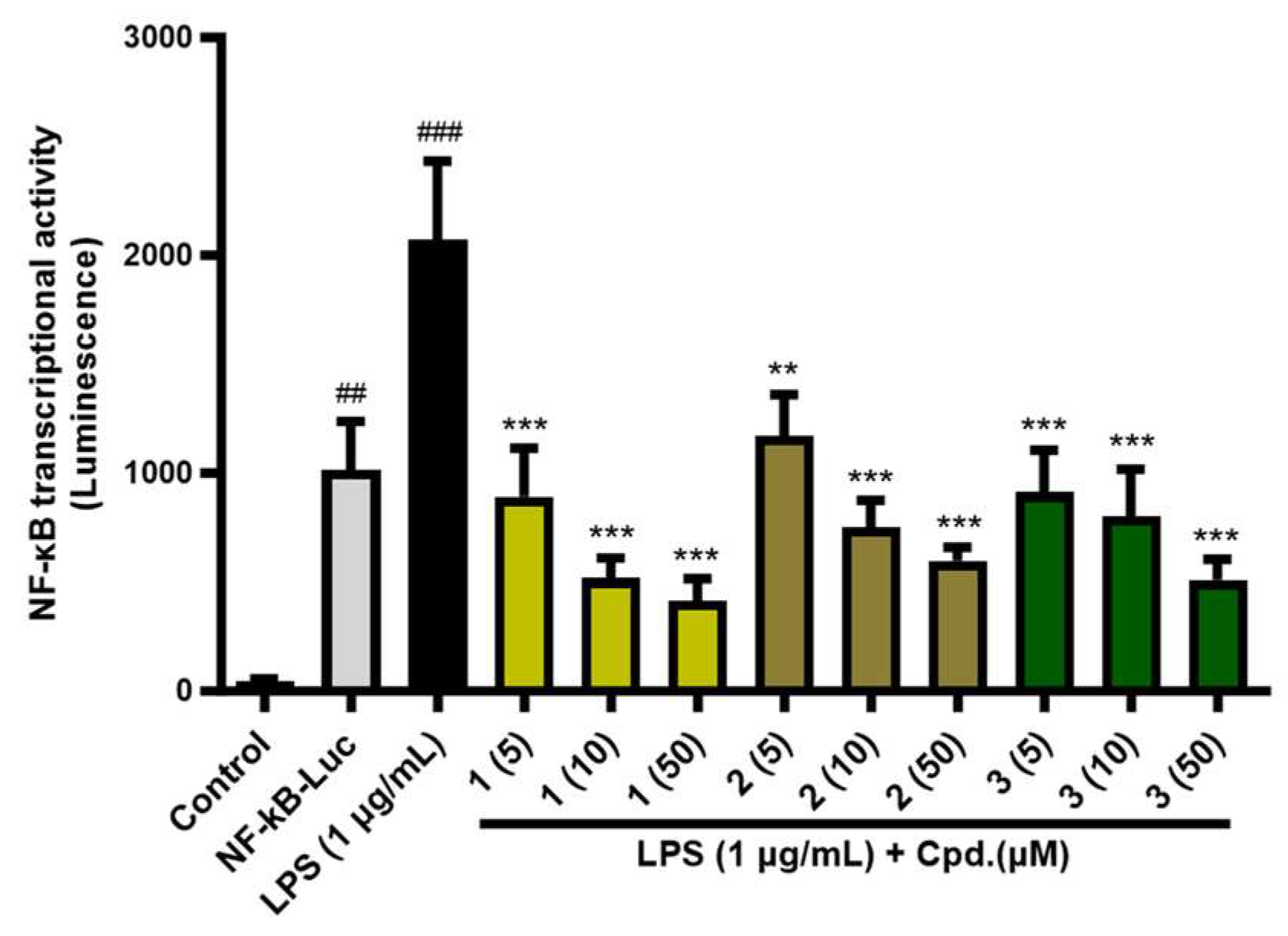
Figure 6.
The effects of the P. frutescens var. acuta extract and 1-3 on the mRNA levels of pro-inflammatory cytokines in Raw 264.7 cells pretreated with LPS. (a) The water extract (0, 10, 50, 100 µg/mL). (b)-(d) 1-3 (0, 5, 10, 50 µM). Raw 264.7 cells were pretreated with each component dissolved in DMSO for 24 hours, and then 100 ng/mL of LPS for 6 hours. The mRNA levels were determined by quantitative real-time polymerase chain reaction with normalization relative to 18s rRNA. Data are presented as fold changes compared to Veh-treated control. A-D Different superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different. (Veh-treated control = 1; means ± standard error of mean; n=3). (Veh, vehicle; Il6, interleukin 6; Mcp1, monocyte chemoattractant protein 1; Tnfa, tumor necrosis factor α.).
Figure 6.
The effects of the P. frutescens var. acuta extract and 1-3 on the mRNA levels of pro-inflammatory cytokines in Raw 264.7 cells pretreated with LPS. (a) The water extract (0, 10, 50, 100 µg/mL). (b)-(d) 1-3 (0, 5, 10, 50 µM). Raw 264.7 cells were pretreated with each component dissolved in DMSO for 24 hours, and then 100 ng/mL of LPS for 6 hours. The mRNA levels were determined by quantitative real-time polymerase chain reaction with normalization relative to 18s rRNA. Data are presented as fold changes compared to Veh-treated control. A-D Different superscripts indicate significant differences at P < 0.05 by Duncan’s multiple comparison test. If a group has a common superscript with another group, it means they are not statistically different. (Veh-treated control = 1; means ± standard error of mean; n=3). (Veh, vehicle; Il6, interleukin 6; Mcp1, monocyte chemoattractant protein 1; Tnfa, tumor necrosis factor α.).
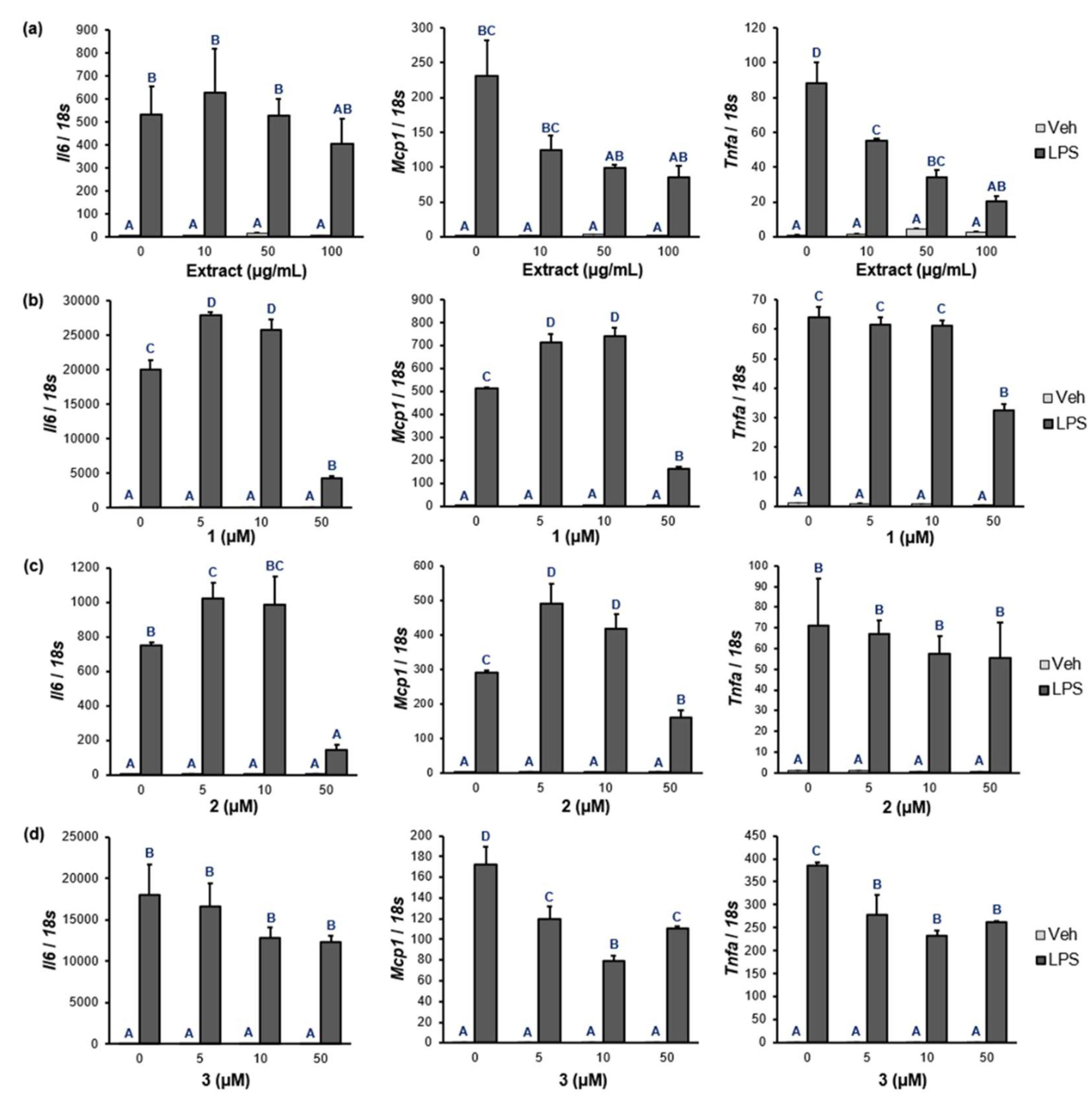

Table 1.
Docking energy (Kcal/mol) of 1-3 and EPA with PPAR-α/δ/γ.
| Compound | Autodock Vina | Autodock 4 | Dock6 | |
| PPAR-α | EPA * | -6.7 | -7.8 | -35.5 |
| 1 | -7.2 | -13.2 | -41.5 | |
| 2 | -6.5 | -9.1 | -41.7 | |
| 3 | -8.6 | -9.7 | -43.0 | |
| PPAR-δ | EPA | -7.8 | -7.4 | -41.0 |
| 1 | -9.7 | -14.7 | -58.1 | |
| 2 | -9.4 | -13.2 | -43.3 | |
| 3 | -8.3 | -9.3 | -41.5 | |
| PPAR-γ | EPA | -6.8 | -8.1 | -37.2 |
| 1 | -5.8 | -9.7 | -57.1 | |
| 2 | -5.6 | -13.4 | -51.6 | |
| 3 | -7.6 | -8.9 | -40.1 |
* A positive control.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



