
Submitted:
07 November 2023
Posted:
07 November 2023
You are already at the latest version
Abstract
OsNRAMP5—a member of the same family as the iron, manganese (Mn), and cadmium (Cd) transporter OsNRAMP1—is responsible for the transport of Mn and Cd from soil in rice. Knockout of OsNRAMP5 markedly reduces both Cd and Mn absorption; therefore, in low -Mn environments, such plants would exhibit Mn deficiency and suppressed growth. Amino acid mutations in IRT1 alter its metal selectivity. We generated random mutations in OsNRAMP5 via error-prone PCR, and used yeast to screen for the retention of Mn absorption and inhibition of Cd absorption. The results showed ththat alanine 512th is the most important amino acid residue for Cd absorption, and that its substitution resulted in the absorption of Mn but not Cd.
Keywords:
OsNRAMP5
; cadmium
; manganese
; rice
; transporter
; random mutation
1. Introduction
The global population surpassed 8 billion in 2023 and is increasing; it is projected to exceed 11.2 billion in 2100 [1]. Worldwide, 750 million people suffer from hunger and undernourishment, a number projected to exceed 840 million in 2030 and 2 billion in 2050 [2]. The total global cultivable area has decreased since 1961 as a result of urbanization [3]. The improvements in crop varieties and techniques resulting from the ‘Green Revolution’ have increased yields per unit area; however, further increasing yields is problematic. To provide sufficient food for the increasing global population, there is a need to develop plants that are tolerant to poor environments.
Soil contamination by toxic heavy metals precludes its use for agricultural purposes. When crops absorb nutrients such as trace elements from soil, they also take up harmful heavy metals. Among these harmful heavy metals, cadmium (Cd) is an atypical transition heavy metal readily absorbed in conjunction with other minerals required for plant growth (e.g., iron [Fe], zinc [Zn], and manganese [Mn]) [4]. It has a long biological half-life; high mobility, solubility, fluidity, and bioaccumulation; and long-lasting toxicity, irrespective of concentration [5]. Cd is not essential for plant growth or the biological functions of humans and animals. In plants, excess Cd causes growth disorders. Cd contamination is a severe and ubiquitous environmental problem, and Cd enters food chains by being absorbed by plants, and then subsequently accumulating in animals and humans. Lifelong intake of Cd, which has a biological half-life of around 10 years, can damage the lungs, kidneys, bones, and reproductive system. In Japan, Itai-Itai disease was first reported in the 1910s, and Cd-exposed miners in Europe suffered lung damage in the 1930s; in both cases, the damage was induced by chronic Cd intoxication [6].
Cd is produced by natural activities (volcanic activity, weathering, and erosion), anthropological activities (smoking, smelting, and fossil fuel combustion), and remobilization of historical sources, including watercourse contamination. Those industrial activities including mining and smelter could influence paddy fields to a large extent [7]. As a result, dietary intake accounts for approximately 90% of all Cd intake in the non-smoking population; other sources include drinking water and exposure to inexpensive jewelry, toys, and plastics [8]. According to the national food survey, and estimation of total diet, the Cd intake worldwide is within the range from 0.1-0.51 µg/kg of body weight per day, but comparing the intake of different countries, Asian nations, such as China (0.21-0.51 µg/kg) and Japan (0.31-0.36 µg/kg), showed a higher level of intake than those of the United States (0.13-0.15 µg/kg) and European nations (0.16 µg/kg in Finland, 0.18 µg/kg in Germany, etc.) [9], which could be contributed to the larger consumption of rice in Asian nations [10]. Specifically in China, which is the largest rice producer [11], although the National Standard of the People’s Republic of China limits the Cd content in rice to 0.2 mg/kg [12], but 10.3% of rice on the Chinese market exceeds this limit [13]. The independent market surveys carried in 6 administrative regions in those three major cropping regions showed tested samples from all administrative regions are Cd-contaminated to different extents: the average Cd content ranged from 0.12 to 0.46 mg/kg and 14%-100% exceeded the standard limit [14,15,16,17,18,19]
In rice, Cd is transported within the plant via the apoplastic and symplastic pathways, and both pathways involve transporters of other metallic elements essential for plant growth. Because Cd shares similar chemical properties with Fe, they are closely associated together in plants [20]. The mechanisms of the uptake and accumulation of Fe and Cd are somehow common as a result of similar entry routes within rice. During the vegetative stage, Fe and Cd are absorbed by specific root transporter and transported to aerial parts via xylem-to-phloem transfer system, and at grain-filling, grain Fe and Cd are both derived from phloem [21]. With the presence of Cd, Fe deficiency symptoms could be induced because Cd inhibits not only the absorption of Fe [22], but the transportation of Fe from root to shoot [23]. On the other hand, the addition of Fe could also reduce Cd content in rice [24] and enhance rice growth and yield [25], which suggests that Cd translocation into rice might occur via Fe metabolic pathways [26]. The interaction between Mn and Cd has also been identified, since the accumulation of Cd is reduced in both root and shoot in the Mn sufficiency environment compared to the Mn deficiency environment [27]. Fe and Mn alleviated Cd toxicity by preventing Cd from being absorbed by forming an Fe plaque on the surface of rice root [28]. Meanwhile, Fe and Mn could also protect plants from damage induced by Cd on root growth and photosynthesis [29].
Several genes in rice have been reported to take part in xylem loading and phloem redistribution of Fe, Mn and Cd at different locations in plants [30]. For example, members of the heavy metal-associated protein (HMA) metal -transporter family transport Cd to the root vascular bundle. Similar to AtHMA4 and AhHMA4, OsHMA2 has also been identified as a transporter of both Zn and Cd, and in the OsHMA2-suppressed rice, the concentration of both Cd and Zn decreased in leaves and seeds, which suggested that OsHMA2 played a role in Cd loading to the xylem and participated in root-to-shoot translocation of Cd apart from Zn [31]. Different from OsHMA2, OsHMA3 reportedly does not transport other metals such as Zn [32]. To be specific, OsHMA3, a regulator for Cd transport in xylem in rice, has the function of mediating vacuolar sequestration of Cd in root cells [33]. The expression of OsHMA3 was directly proportional to Cd concentration in the environment [34], but with excessive Fe treatment, the expression of OsHMA3 would significantly increase [35]. RNAi-mediated knock-down of OsHMA3 increased root-to-shoot Cd translocation, and on the contrary, the overexpression of OsHMA3 reduced shoot Cd accumulation, which indicated that OsHMA3 has the function in vacuolar compartmentation of Cd in root, which decreased the xylem loading of Cd and subsequent shoot Cd accumulation [36]. Cd is also transported to seeds via the phloem in a manner involving the product of OsLCT; the phloem and seeds of OsLCT1 mutants generated by RNA interference had low levels of Cd [37]. Because Cd is toxic, it is detoxified by inclusion in complexes with thiol compounds such as phytochelatin (PC) and glutathione (GS, a synthetic substrate for PC). In rice, such thiol compounds are synthesized by OsGS and OsPCS, resulting in the extracellular transport of some Cd [38]. Therefore, it is necessary to modify steps in the plant Cd transport pathway—for instance, Cd absorption from soil, transportation from root to leaf, and sequestration into the vacuole—to enhance its detoxification. Doing so would enable the development of low-Cd foods in which Cd is not stored in seeds.
The natural resistance-associated macrophage protein (NRAMP) family is involved in the absorption of metal elements in diverse taxa. Human NRAMP1 transports divalent metals (e.g., Mn, Fe, and cobalt), as does divalent metal transporter 1 (DMT1; alternatively, NRAMP2, DCT), which is a transporter of Cd and Fe [39]. Rice has seven NRAMP transporters, among which OsNRAMP1 is responsible for the uptake and transport of Cd in plants [40]. Transformation with OsNRAMP1 reduced the Cd tolerance of yeast [41]. However, OsNARMP1 also transports Mn and Fe. Similar interaction between Cd and Fe was also found in both the ferrous Fe transporter iron-regulated transporter 1 (IRT1) and IRT2 in rice. Both OsIRT1 and OsIRT2 are related to Fe uptake in root, and also showed an influx activity of Cd as well as Fe in yeast, showing that OsIRT1 and OsIRT2 are important transporters in root with the function of uptake of Cd [42,43]. OsIRTs may contribute to the uptake of Cd in aerobic conditions when water was released. Meanwhile, Cd is absorbed in roots through the OsNRAMP5 transporter, and OsNRAMP5, which has been identified as a transporter of Mn and Cd, is responsible for the absorption of Mn and Cd from soil [44]. The reason that rice accumulates more Cd than other cereal crops may also be considered that OsNRAMP5 gene has a higher expression in rice [45]. Interestingly, Fe absorption by OsNRAMP5 in root and shoot tissues did not differ significantly between the wild type and an OsNRAMP5 mutant [46]. Furthermore, knockout of OsNRAMP5 markedly reduced the amount of Cd in rice by abolishing its uptake from soil. Therefore, knockout of OsNRAMP5 is a promising trait for producing low-Cd rice. Because OsNRAMP5 transports both Mn and Cd, OsNRAMP5 knockout also reduced Mn absorption by about 90% [47]; therefore, in low-Mn environments, such plants would exhibit Mn deficiency and suppressed growth.
Mutations in OsIRT1 alter its metal selectivity [48]. Among the 538 amino acid residues comprising OsNRAMP5, one or more may mediate its transport of Mn, Cd, or both. Therefore, substitution of a specific amino acid residue may affect Mn and/or Cd transport in a manner that does not alter the Mn uptake while suppressing Cd uptake. Rice with such a mutation could maintain Mn uptake while avoiding Cd accumulation when grown in Cd-contaminated soil with low Mn, and that there would be no negative influence on the growth. The development of rice varieties that can absorb Mn but not Cd would enable the cultivation of soil with a greater range of Cd contamination levels than would rice varieties with OsNRAMP5 knockout, and enlarge the production of sufficient crops with low Cd concentration. To this end, in the present study, we introduced mutations into OsNRAMP5 and evaluated their effects on Mn and Cd transport.
2. Results
2.1. OsNRAMP5 transported both Mn and Cd
The transport of Cd and Mn by OsNRAMP5 was evaluated in yeast. In the absence of Mn, the growth of the negative control (VC) was inhibited, but OsNRAMP5-expressing N5 showed good growth as a result of transport of Mn by OsNRAMP5 (Figure 1a). In the presence of Cd, the growth of N5 was impaired but that of VC was unaffected (Figure 1b). These results indicate that, when expressed in yeast, OsNRAMP5 transports both Mn and Cd.
2.2. Patterns of OsNRAMP5 mutations
We selected 100 colonies from ten -Mn/ +Cd plates in the first screening (200,000 colonies in total), and 20 colonies in the second screening (Table 1). The nucleotide sequences of 20 mutants were classified into four patterns: pattern 1, two nucleotide mutations corresponding to one amino acid mutation; pattern 2, four nucleotide mutations corresponding to three amino acid mutations; pattern 3, two nucleotide mutations corresponding to one amino acid mutation; and pattern 4, 3 nucleotide mutations corresponding to three amino acid mutations. These four patterns were divided into two types according to their amino acid mutations: patterns 1 and 3 and patterns 2 and 4. The G to A substitution at position 1534 (alanine to threonine at residue 512) was present in all four patterns.
2.3. Mutants absorb Mn but not Cd
The absorption of Cd and Mn by the 20 mutants was compared with those of VC and N5. Under Mn-deficient conditions, all mutants showed growth similar to N5, indicative of similar levels of Mn absorption (Figure 2a). In the presence of Cd, N5 showed little growth, but the mutants—particularly those with pattern 3 (M35, M36, M70, and M82)—showed improved growth compared to other mutants (Figure 2b). In the presence of Cd but not Mn, the growth of VC and N5 was inhibited by Mn deficiency and Cd toxicity, respectively. However, all mutants showed improved growth, indicating that they could absorb Mn but not Cd in the environment of -Mn/ +Cd (Figure 2c).
The growth rates of M6 (mutant from pattern 1) and M9 (mutant from pattern 2) were compared with those of VC and N5 in liquid medium. Under Mn-deficient conditions, M6 and M9 grew slightly slower than N5, but much faster than VC (Figure 3a). In the presence of Cd, the growth of N5 was suppressed, and the growth of the two mutants was similar to VC (Figure 3b). In the presence of Cd but not Mn, the growth rates of the two mutants were higher than those of VC and N5 (Figure 3c).
2.4. Mutants show reduced absorption of Cd, but similar absorption of Mn compared to N5
M6 and M9 showed Cd absorption similar to that of VC but significantly different from that of N5 in the presence of 20 μM CdCl2 for 72 h (Figure 4a). In the presence of Cd, M6, M9, VC, and N5 showed similar Mn absorption. In the absence of Cd, M6 showed greater Mn absorption than N5, and M9 showed Mn absorption lower than N5 but similar to VC (Figure 4b).
M6 showed similar Fe absorption to N5 in the absence of Cd, which was significantly higher than in the presence of Cd. However, M9 showed similar absorption of Fe to VC irrespective of the presence of Cd. Meanwhile, the absorption of Fe in both M9 and VC showed lower levels in the absence of Cd, but M6, M9, VC, and N5 showed similar levels of Fe absorption in the presence of Cd (Figure 4c). There is no significant difference in the absorption of Cu compared to M6, M9, VC, and N5 irrespective of the presence of Cd. Therefore, neither pattern 1 mutants nor the pattern 2 mutants showed an influence in the absorption of Cu. Moreover, M6, M9, VC, and N5 showed similar Zn absorption in the presence of Cd, although in the absence of Cd, only M6 showed elevated Zn absorption (Figure 4e).
2.5. Alanine 512 is essential for Cd absorption
The 20 plasmids harbored a substitution of alanine for threonine at residue 512. To evaluate its importance, we mutated alanine 512 to methionine (A512M), isoleucine (A512I), and aspartic acid (A512D). The metal-transport activities of the mutants were compared with those of VC, N5, and A512T. All mutants had growth rates similar to A512T (Figure 5).
3. Discussion
The growth of the mutants in the presence of Cd and the absence of Mn is likely attributable to the mutations (Table 1, Figure 2a,2b, Figure 3a,3b). In the medium containing Cd but not Mn, VC and N5 did not grow as a result of Mn deficiency and Cd toxicity, respectively (Figure 1, Figure 2), but obtained OsNRAMP5 mutants showed good growth (Figure 2). In liquid medium, the growth rates of the M6 and M9 mutants were similar to N5 in the absence of Mn (Figure 3a), and both mutants showed similar growth rates under all conditions (Figure 3). These findings indicated that M6 and M9 had reduced absorption of Cd, but similar or increased absorption of Mn compared to N5, suggesting that the OsNRAMP5 mutants could mediate the absorption of Mn while suppressing that of Cd (Figure 2c, Figure 3, Figure 4a, 4b). Also, alanine 512, which was common to all mutations, is likely involved in the absorption of Cd (Figure 5). However, even though there was a significant growth improvement of M6 and N5 with the absence of Mn, the absorption of Mn still showed similar performance with presence of both Mn and Cd in M6, M9, VC and N5 (Figure 4b) due to competitivity of Mn to Cd [49]. Furthermore, since the previous research on OsIRTs indicated that although Cd accumulation in roots and shoots of OsIRT1 overexpression plants was increased under MS medium with excessive Cd, such a phenotype was not shown in the paddy field, which demonstrates that the contribution of the transporters is also affected by the external environment conditions [50].
In the presence of Cd, M6 showed lower Cd absorption and higher Mn and Zn absorption compared to N5, suggesting that the altered Cd and Mn absorption of M6 resulted from mutation of alanine 512 (Figure 5). Alanine 512 is also important for the absorption of Zn. In the absence of Cd, Zn absorption by N5 was markedly lower than that of M6, and slightly but non-significantly lower than that of M9 (Figure 4e). However, because Cd is more competitive and easier to be absorbed than Zn [49], the Zn absorption by M6 and M9 decreased to a level similar to those of VC and N5 in the presence of Cd. Cd is less competitive to Cu [49], which might lead to no significant change in absorption of Cu with both presence of Cd and absence of Cd in all M6, M9, VC and N5 (Figure 4d). Moreover, mutations of serine 8, cysteine 111, or both may impede the absorption of Zn and Fe, possibly explaining the similar Zn absorption of N5 and M9, and lower Fe absorption in M9 than N5 and M6 in the absence of Cd (Figure 4c, 4e). For all metals investigated, M9 had similar absorption rates as VC (Figure 4), suggesting that serine 8, cysteine 111, or both are important for metal transport by OsNRAMP5. In M9, the A512T mutation non-significantly enhanced Mn transport compared to VC (Figure 4b). This may explain why the mutations of patterns 2 and 4 were obtained by screening in the absence of Mn.
Particular attention should be paid to the change at nucleotide 507, because yeast with pattern 3 grew better than pattern 1 on a Cd-containing medium with the same construction of amino acid (Table 1, Figure 2b). The changes from C to T at nucleotide 21 (pattern 1) and T to C at nucleotide 507 (pattern 3) (Table 1) could alter transcriptional efficiency, RNA stability, and transfer RNA levels even though the amino acid was kept the same [51]. Whether these are silent mutations that affect absorption could be evaluated by creating a plasmid with the only a mutation at nucleotide 21 and a plasmid with the only a mutation at nucleotide 507, and evaluating the effect on metal absorption compared to M6, A512T, and yeast with pattern 3 mutant introduced in future study.
Regarding structural prediction, OsNRAMP5-Q337K, in which glutamine residue in the 8th transmembrane domain was substituted by lysine residue, reduced the grain Cd concentration without causing severe Mn deficiency in rice [52]. The three mutations in this study were predicted to be cytoplasmic (Figure 6). It is necessary to investigate how these residues, which are not extracellular and so cannot interact directly with extracellular metal ions, affect the absorption of metals.
4. Materials and methods
4.1. Mn and Cd absorption assays
The plasmids pDR195 and pDR195 containing OsNRAMP5 were transformed into the Mn-absorbing transporter-deficient mutant yeast strain Δsmf1 (MATalpha, his3Δ1; leu2Δ0; meta15Δ0; ura3Δ0; YOL122c:: kanMX4), generating VC and N5, respectively. Metal transport assays were carried out in synthetic defined (SD) medium (2% glucose, 0.5% yeast nitrate base without amino acids, and 2% agar) containing Cd (100 μM CdCl2) but not Mn (20 mM EGTA, pH 5.9) [44,46]. The media were spotted with 8 µL of yeast suspension (OD600=0.1, 0.01, 0.001), incubated statically at 30°C for 2 days, and the growth of the yeast strains was monitored.
4.2. Error-prone PCR
To ligate OsNRAMP5 into pDR195, HindIII and EcoRI sites were introduced (Ligation Mighty Mix (TaKaRa)), and the vector was digested by BamHI and XholI. Mutations were introduced into OsNRAMP5 via error-prone PCR, which we employed previously [55] in ten tubes (using 50×Titanium Taq DNA Polymerase (TaKaRa)). The PCR conditions were denaturation at 95℃ for 15s, annealing at 55℃ for 15 s, and extension at 68℃ for 2 min for 30 cycles, during which the Mn concentration was changed, and random mutations were introduced. OsNRAMP5 has 1614 nucleotides; we used 300 μM Mn to introduce two to five mutations.
4.3. Screening
OsNRAMP5 fragments with random mutations were ligated into pDR195. The vectors were transformed into Δsmf1 and cultured on -Mn/+Cd SD medium (20 mM EGTA, 100 μM CdCl2, pH=5.9) for 1st48 h at 30°C for the first screening (ten plates). Next, colonies were diluted and cultured in fresh -Mn/+Cd SD medium for the second screening (100 plates) and the colonies were sequenced.
4.4. Growth assay
Plasmids harboring OsNRAMP5 mutants were transformed into Δsmf1, and cultured on solid -Mn, +Cd, and -Mn/+Cd SD media. The growth rates of the mutants were analyzed in liquid -Mn, +Cd, and -Mn/+Cd SD media and compared with those of VC and N5.
4.5. Uptake of metals
VC, N5, and two OsNRAMP5 mutant yeast strains (M6 and M9) were cultured in 50 mL of liquid SD medium (20 µM CdCl2, 2.5 μM MnSO4, 0.75 μM FeCl3, 1.5 μM ZnSO4, 0.15 μM CuSO4) for 72 h at 30˚C. The yeasts were centrifuged at 8000 xg for 5 min, washed with 50 mL of distilled H2O at 8000 xg for 5 min for 2 times, and washed with 30 mL of distilled H2O at 8000 xg for 5 min. The pellets were dried in a warmer at 60˚C for 48 h. The dried pellets were weighed, wet-ashed diluted with 2% HNO3, and the Fe, Mn, Cu, Zn, and Cd concentrations were analyzed by inductively coupled plasma optical emission spectrometry (ISPS-3500, Seiko Instruments Inc.).
4.6. Amino acid substitution
The candidate amino acid residue in pDR195 (digested by SalI) was substituted for other amino acid residues to evaluate its function in Mn and Cd transport. The resulting plasmids were transformed into Δsmf1, and cultured on solid -Mn, +Cd, and -Mn/+Cd SD media.
4.7. Statistical analysis
The significance of differences was evaluated by one-way analysis of variance followed by Tukey’s multiple comparison test P< 0.05 was considered to indicate significance.
5. Conclusion
The results in this study indicate that alanine 512 mediates, at least in part, Cd transport by OsNRAMP5; its substitution significantly decreased Cd transport but increased Mn transport. Furthermore, it is also necessary to consider the two amino acid residue changes, S8R and C111Y, other than the 512th alanine for the metal selectivity of OsNRAMP5 protein. Rice harboring metal-selective OsNRAMP5 could grow in low-Mn soil and/or Cd-contaminated soil, thereby increasing the cultivable land area in the future.
Author Contributions
Conceptualization, H.N. and Z.Q.; methodology, H.N. and Z.Q.; data curation, Z.Q. and H.N.; writing-original draft preparation, Z.Q.; writing-review and editing, H.N.; project administration, H.N; funding acquisition, H.N. and Z.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This work was partially supported by a grant from The Tojuro Iijima Foundation for Food Science and Technology.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- United Nations Department of Economic and Social Affairs Population Division. World Population Prospects; Key findings & advance tables 2015 REVISION.; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Food and Agriculture Organization of the United Nations. The State of Food Security and Nutrition in the World; THE STATE OF FOOD SECURITY AND NUTRITION IN THE WORLD, TRANSFORMING FOOD SYSTEMS FOR AFFORDABLE HEALTHY DIETS.; Rome, Italy, 2020.
- Ritchie, H.; Roser, M. OurWorldInData; Land Use; Oxford, England, 2013.
- Haider, F.U.; Cai, L.; Coulter, J.A.; Cheema, SA.; Wu, J.; Zhang, R.; Ma, W.; Muhammad, F. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol Environ Saf. 2021, Mar 15, 211:111887.
- Khan, M.A.; Khan, S.; Khan, A.; Alam M. Soil contamination with cadmium, consequences and remediation using organic amendments. Sci Total Environ. 2017, Dec 1;601-602:1591-1605.
- Rafati Rahimzadeh, M.; Rafati Rahimzadeh M.; Kazemi, S.; Moghadamnia, A.A. Rafati Cadmium toxicity and treatment: An update. Caspian J Intern Med. 2017, Summer;8(3):135-145.
- Uraguchi S, Fujiwara T. Cadmium transport and tolerance in rice: perspectives for reducing grain cadmium accumulation. Rice (N Y). 2012 Feb 27;5(1):5.
- Department of Public Health, Environmental and Social Determinants of Health World Health Organization. EXPOSURE TO CADMIUM: A MAJOR PUBLIC HEALTH CONCERN.; Geneva, Switzerland, 2019.
- Bellinger, D.; Bolger, M.; Goyer, R.; Barraj, L.; Baines, J. WHO food additives series: 46, cadmium. World Health Organization: Geneva, Switzerland. 2001.
- China Food and Drug Administration. National food safety standard Contamination Limit in Food. Beijing, China. 2017. 9.
- Scientific Opinion of the Panel on Contaminants in the Food Chain. Cadmium in food. The EFSA Journal 2009, 980, 1–139. [Google Scholar]
- Skorbiansky, S.R.; Childs, N.; Hansen, J. Rice in Asia’s Feed Markets. RCS-18L-01. USDA-ER: Washington DC, SW, USA 2018.
- Liu, B.; Li, T.; Cai, Y. Brief introduction to status quo, hazards and repair methods of cadmium rice. Modern Food. 2018. 21:86-89.
- Li, Q.; Shi, L.; Chen, J.; Tan, H.; Zhou, H.; Yao, L.; Xu, M.; Shang-guan, J.; Lu, K. Investigation on cadmium pollution in foods in country of Jiangxi Province. Chinese Journal of Food Hygiene. 2008, 20(4):330-331.
- Lei, M.; Zeng, M.; Wang, L.; Williams Paul, N.; Sun, G. Arsenic, lead and cadmium pollution in rice from Hunan markets and contaminated areas and their health risk assessment. Acta Scientiae Circumstantiae. 2010, 30(11):2314-2320.
- Wu, D.; Yang, X.; Li, C.; Zhou, C.; Qin, F. Contrentations and health risk assessment of heavy metals in soil and rice and zinc-lead mining area in Guizhou province, China. Journal of Agro-Enviroment Science. 2013,32(10): 1992-1998.
- Cai, W.; Su, Z.; Hu, S.; Huang, W.; Xu, X.; Huang. X. Assessment of the content and exposure of lead and cadmium in the major food of Guangdong residents. Chin J Health Lab Tec. 2015, 25(14):2388-2392.
- Ren, R.; Gong, L.; Wang, S.; Jin, Q. Survey of heavy metal contamination and risk assessment of exposure in Hangzhou indigenous rice. Chin J Health Lab Tec. 2020, 30(12):1516-1520.
- Jiang, Y.; Ma, N.; Meng, H.; Shi, M.; Zhao, P. Risk assessment on the dietary exposure of cadmium in Guangxi residents. Chinese Journal of Food Hygiene. 2021, 30(12):191-195.
- Liu, H.; Zhang, C.; Wang, J.; Zhou, C.; Feng, H.; Mahajan, MD.; Han, X. Influence and interaction of iron and cadmium on photosynthesis and antioxidative enzymes in two rice cultivars. Chemosphere. 2017, Mar; 171:240-247. 20.
- Gao, L.; Chang, J.; Chen, R.; Li, H.; Lu, H.; Tao, L.; Xiong, J. Comparison on cellular mechanisms of iron and cadmium accumulation in rice: prospects for cultivating Fe-rich but Cd-free rice. Rice (N Y). 2016, Dec;9(1):39.
- Sharma, S.S.; Kaul, S.; Metwally, A.; Goyal, K.C.; Finkemeier, I.; Dietz, K.J. Cadmium toxicity to barley (Hordeum vulgare) as affected by varying Fe nutritional status. Plant Science. 2004, 166(5): 1287-1295.
- Xu, S.S.; Lin, S.; Lai, Z. Cadmium impairs iron homeostasis in Arabidopsis thaliana by increasing the polysaccharide contents and the iron-binding capacity of root cell walls. Plant and Soil. 2015, 392, 71–85. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Wang, J.; Zhou, C.; Feng, H.; Mahajan, MD.; Han, X. Influence and interaction of iron and cadmium on photosynthesis and antioxidative enzymes in two rice cultivars. Chemosphere. 2017,171: 240–247.
- Liu, HJ.; Zhang, JL.; Zhang, FS. Role of iron plaque in Cd uptake by and translocation within rice (Oryza sativa L.) seedlings grown in solution culture. Environmental and Experimental Botany. 2007, 59: 314–320.
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, JF. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell. 2012; 24: 2155–2167.
- Wang, M.; Ma, W.; Chaney, RL.; Green, CE.; Chen W. Effects of Mn2+ on Cd accumulation and ionome in rice and spinach. J Environ Qual. 2022 Sep;51(5):890-898.
- Liu, WJ.; Zhu, YG.; Smith, FA. Effects of iron and manganese plaques on arsenic uptake by rice seedlings (Oryza sativa L.) grown in solution culture supplied with arsenate and arsenite. Plant and Soil. 2005, 277: 127–138.
- Sebastian, A.; Prasad, M. Iron-and manganese-assisted cadmium tolerance in Oryza sativa L.: lowering of rhizotoxicity next to functional photosynthesis. Planta. 2015, 241: 1519–1528.
- Hussain, B.; Li, J.; Ma, Y.; Tahir, N.; Ullah, A. Effects of Fe and Mn cations on Cd uptake by rice plant in hydroponic culture experiment. PLoS ONE. 2020, 15(12): e0243174.
- Uraguchi, S.; Fujiwara, T. Cadmium transport and tolerance in rice: perspectives for reducing grain cadmium accumulation. Rice (N Y). 2012, Feb 27;5(1):5.
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, NK.; Nakanishi H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012 Nov;35(11):1948-57. Uraguchi S, Fujiwara T. Cadmium transport and tolerance in rice: perspectives for reducing grain cadmium accumulation. Rice (N Y). 2012, Feb 27;5(1):5.
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H.; Satoh-Nagasawa, N.; Watanabe, A.; Fujimura, T.; Akagi, H. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011,5:190–199.
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, CF.; Ando, T.; Yano, M.; Ma, JF. Gene limiting cadmium accumulation in rice. PNAS. 2010, 5:16500–16505.
- Cui, J.; Liu, T.; Li, Y.; Li, F. Selenium reduces cadmium uptake into rice suspension cells by regulating the expression of lignin synthesis and cadmium-related genes. Science of the Total Environment. 2018, 644: 602–610.
- Chen, Z.; Tang, YT.; Yao, AJ.; Cao, J.; Wu, ZH.; Peng, ZR.; Wang, SZ.; Xiao, S.; Baker, A.J.M.; Qiu, RL. Mitigation of Cd accumulation in paddy rice (Oryza sativa L.) by Fe fertilization. Environmental Pollution. 2017, 231: 549–559.
- Uraguchi, S.; Kamiya, T.; Sakamoto, T.; Kasai, K.; Sato, Y.; Nagamura, Y.; Yoshida, A.; Kyozuka, J.; Ishikawa, S.; Fujiwara, T. Low-affinity cation transporter (OsLCT1) regulates cadmium transport into rice grains. Proc Natl Acad Sci USA. 2011, 108: 20959–20964.
- Zhong, S.; Li, X.; Li, F.; Huang, Y.; Liu, T.; Yin, H.; Qiao, J.; Chen, G.; Huang, F. Cadmium uptake and transport processes in rice revealed by stable isotope fractionation and Cd-related gene expression. Science of The Total Environment. 2021, 806(2):150633.
- Nevo, Y.; Nelson, N. The NRAMP family of metal-ion transporters. Biochim Biophys Acta. 2006, Jul;1763(7):609-20.
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, NK. Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant Signal Behav. 2011 Nov;6(11):1813-6.
- Takahashi, R.; Ishimaru, Y.; Senoura, T.; Shimo, H.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. The OsNRAMP1 iron transporter is involved in Cd accumulation in rice. Journal of Experimental Botany. 2011, 62, 4843–4850. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Suzuki, M.; Tsukamoto, T.; Suzuki, K.; Nakazono, M.; Kobayashi, T.; Wada, Y.; Watanabe, S.; Matsuhashi, S.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, NK. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+ Plant J. 2006, 5:335–346.
- Nakanishi, H.; Ogawa, I.; Ishimaru, Y.; Mori, S.; Nishizawa, NK. Iron deficiency enhances cadmium uptake and translocation mediated by the Fe2+ transporters OsIRT1 and OsIRT2 in rice. Soil Sci Plant Nutr. 2006. 5:464–469.
- Chang, JD.; Huang, S.; Yamaji, N.; Zhang, W.; Ma, JF.; Zhao FJ. OsNRAMP1 transporter contributes to cadmium and manganese uptake in rice. Plant Cell Environ. 2020, Oct;43(10):2476-2491.
- Wang, J.; Wang, PM.; Gu, Y.; Kopittke, PM.; Zhao, FJ.; Wang, P. Iron-manganese (oxyhydro) oxides, rather than oxidation of sulfides, determine mobilization of Cd during soil drainage in paddy soil systems. Environmental Science & Technology. 2019, 53: 2500–2508. 45.
- Ishimaru, Y.; Bashir, K.; Nakanishi, H.; Nishizawa, N.K. OsNRAMP5, a major player for constitutive iron and manganese uptake in rice. Plant Signal Behav. 2012, Jul;7(7):763-6.
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T.; Nakanishi, H.; Nishizawa, N.K. Characterizing the role of rice NRAMP5 in Manganese, Iron and Cadmium Transport. Sci Rep. 2012, 2:286.
- Rogers, E.E.; Eide, D.J.; Guerinot, M.L. Altered selectivity in an Arabidopsis metal transporter. Proc Natl Acad Sci USA. 2000, 97(22):12356–12360.
- Jalali, M.; Moradi, F. Competitive sorption of Cd, Cu, Mn, Ni, Pb and Zn in polluted and unpolluted calcareous soils. Environ Monit Assess. 2013, Nov;185(11):8831-46. Epub 2013 May 17. [CrossRef] [PubMed]
- Lee, S.; An, G. Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ. 2009, 5:408–416.
- Chamary, JV.; Hurst, LD.The PRICE of SILENT MUTATIONS. Scientific American. 2009, 300(6): 46–53.
- Kuramata, M.; Abe, T.; Tanikawa, H.; Sugimoto, K.; Ishikawa, S. A weak allele of OsNRAMP5 confers moderate cadmium uptake while avoiding manganese deficiency in rice. Journal of Experimental Botany. 2022, 73(18):6475–6489.
- Oki, H.; Yamaguchi, H.; Nakanishi, H.; Mori, S. (1999) Introduction of the reconstructed yeast ferric reductase gene, refre1, into tobacco. Plant and Soil. 1999, 215: 211–220.
Figure 1.
OsNRAMP5 transport activity in yeast. Growth of two individual manganese-absorbing transporter-deficient mutant Δsmf1 harboring empty pDR195 vector (VC 1 and 2) and OsNRAMP5 (N5 4 and 6) in synthetic defined medium containing (a) 20 mM EGTA and (b) 100 μM CdCl2.
Figure 1.
OsNRAMP5 transport activity in yeast. Growth of two individual manganese-absorbing transporter-deficient mutant Δsmf1 harboring empty pDR195 vector (VC 1 and 2) and OsNRAMP5 (N5 4 and 6) in synthetic defined medium containing (a) 20 mM EGTA and (b) 100 μM CdCl2.
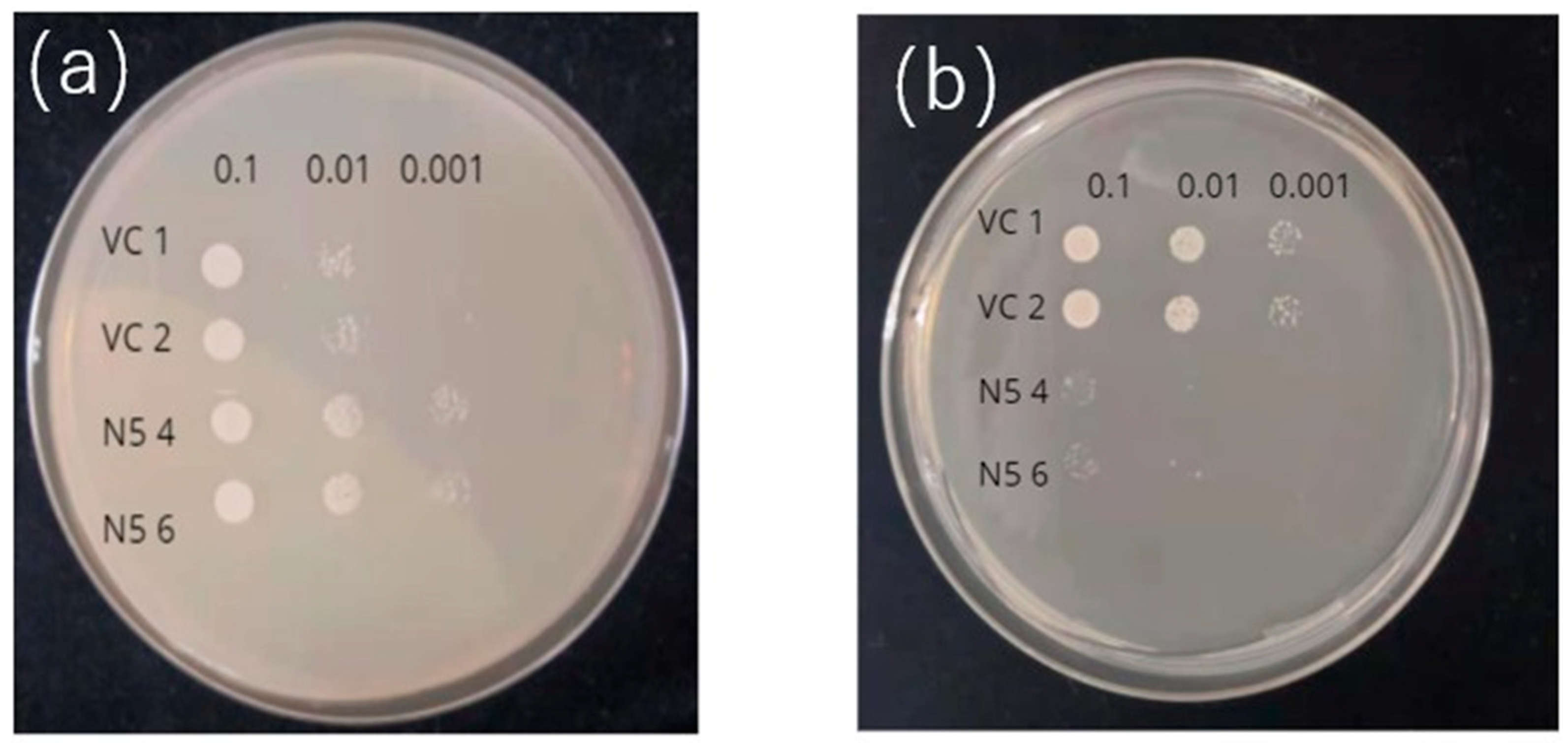
Figure 2.
Transport activities of OsNRAMP5 mutants in Δsmf1 yeast expressing the OsNRAMP5 mutants (M), empty pDR195 vector (VC), and wild-type OsNRAMP5 (N5) in solid synthetic defined medium containing (a) 20 mM EGTA, (b) 100 μM CdCl2, and (c) 20 mM EGTA and 100 μM CdCl2.
Figure 2.
Transport activities of OsNRAMP5 mutants in Δsmf1 yeast expressing the OsNRAMP5 mutants (M), empty pDR195 vector (VC), and wild-type OsNRAMP5 (N5) in solid synthetic defined medium containing (a) 20 mM EGTA, (b) 100 μM CdCl2, and (c) 20 mM EGTA and 100 μM CdCl2.

Figure 3.
Transport activities of OsNRAMP5 mutants in Δsmf1 yeast expressing the OsNRAMP5 mutants (M6 and M9), empty pDR195 vector (VC), and wild-type OsNRAMP5 (N5) in liquid synthetic defined medium containing (a) 20 mM EGTA, (b) 100 μM CdCl2, and (c) 20 mM EGTA and 100 μM CdCl2.
Figure 3.
Transport activities of OsNRAMP5 mutants in Δsmf1 yeast expressing the OsNRAMP5 mutants (M6 and M9), empty pDR195 vector (VC), and wild-type OsNRAMP5 (N5) in liquid synthetic defined medium containing (a) 20 mM EGTA, (b) 100 μM CdCl2, and (c) 20 mM EGTA and 100 μM CdCl2.
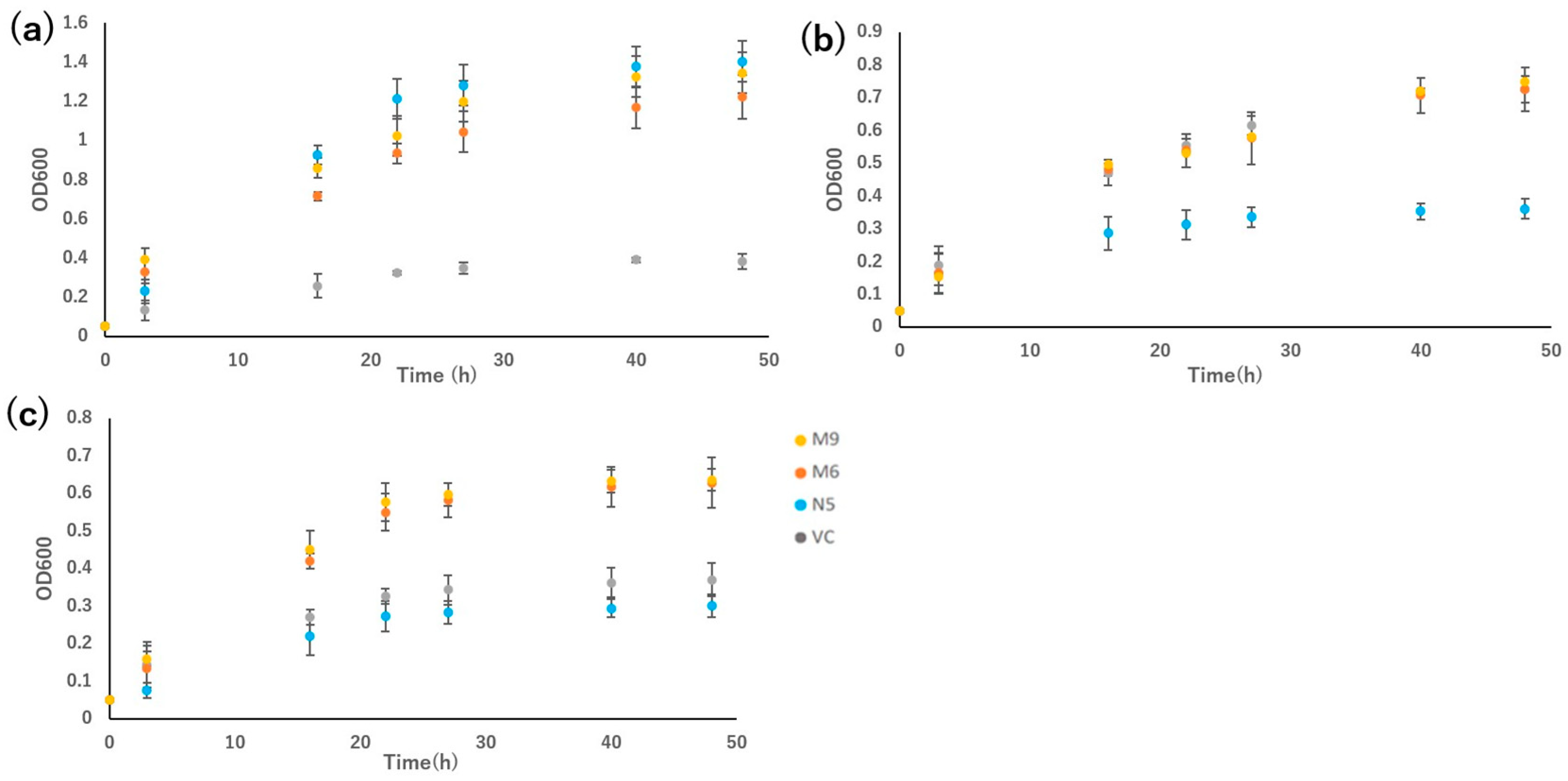
Figure 4.
Metal concentrations in empty pDR195 vector (VC), wild-type OsNRAMP5 (N5), and mutants (M6 and M9) in the presence or absence of 20 μM CdCl2 for 72 h followed by drying for 24 h: (a) cadmium, (b) manganese, (c) iron, (d) copper, and (e) zinc. Data are the means ± standard deviation (n = 3). Different letters indicate significant differences (P < 0.05).
Figure 4.
Metal concentrations in empty pDR195 vector (VC), wild-type OsNRAMP5 (N5), and mutants (M6 and M9) in the presence or absence of 20 μM CdCl2 for 72 h followed by drying for 24 h: (a) cadmium, (b) manganese, (c) iron, (d) copper, and (e) zinc. Data are the means ± standard deviation (n = 3). Different letters indicate significant differences (P < 0.05).
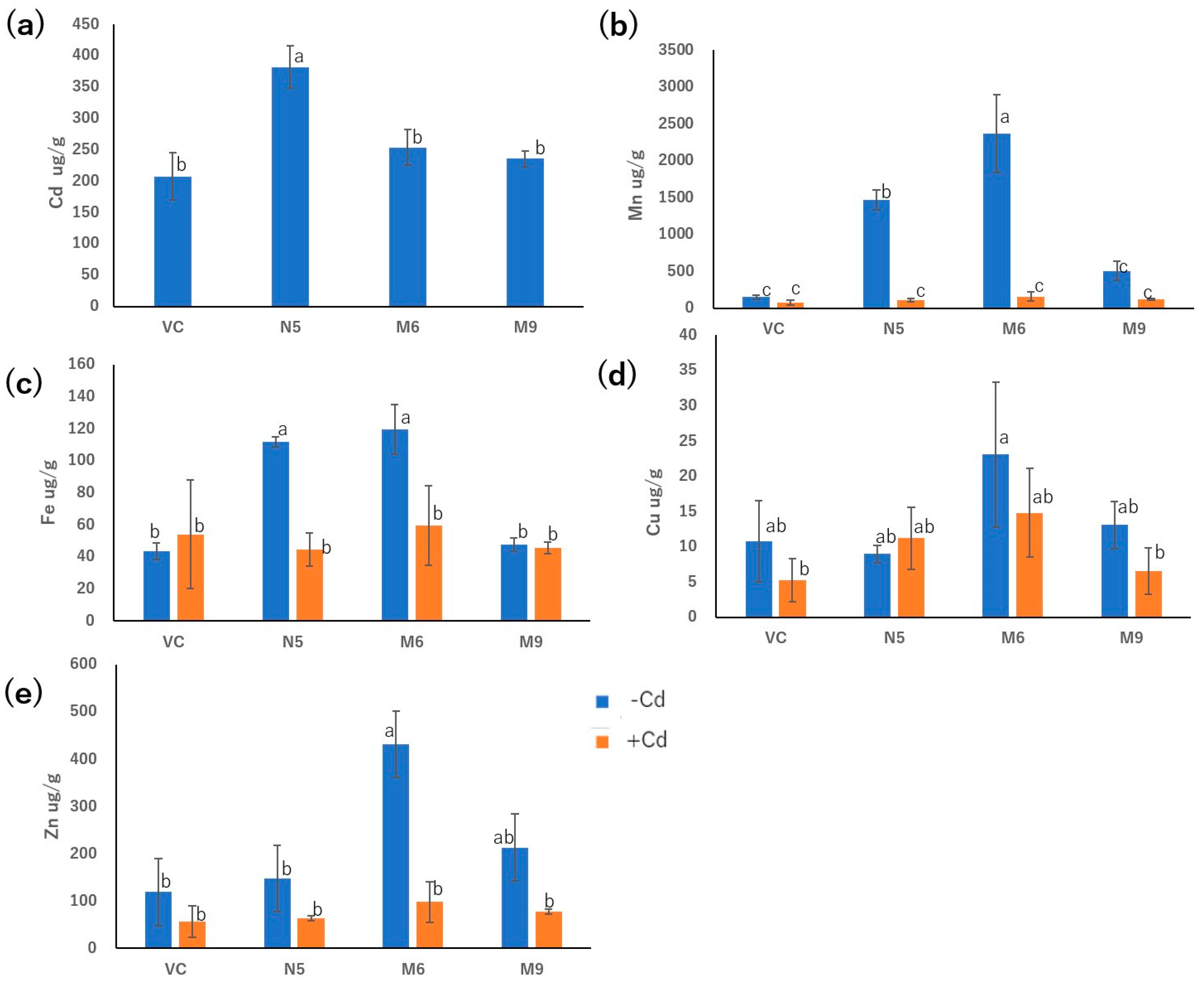
Figure 5.
Transport activities of OsNRAMP5 mutants. Growth of empty pDR195 vector (VC), wild-type OsNRAMP5 (N5), and amino-acid-512 mutants in synthetic defined medium containing (a) 20 mM EGTA, (b) 100 μM CdCl2, and (c) 20 mM EGTA and 100 μM CdCl2.
Figure 5.
Transport activities of OsNRAMP5 mutants. Growth of empty pDR195 vector (VC), wild-type OsNRAMP5 (N5), and amino-acid-512 mutants in synthetic defined medium containing (a) 20 mM EGTA, (b) 100 μM CdCl2, and (c) 20 mM EGTA and 100 μM CdCl2.

Figure 6.
Structural prediction. Alanine 512 (red) is important for cadmium and manganese transport and affects the absorption of zinc. Serine 8 and cysteine 111 (blue) also influence metal absorption.
Figure 6.
Structural prediction. Alanine 512 (red) is important for cadmium and manganese transport and affects the absorption of zinc. Serine 8 and cysteine 111 (blue) also influence metal absorption.
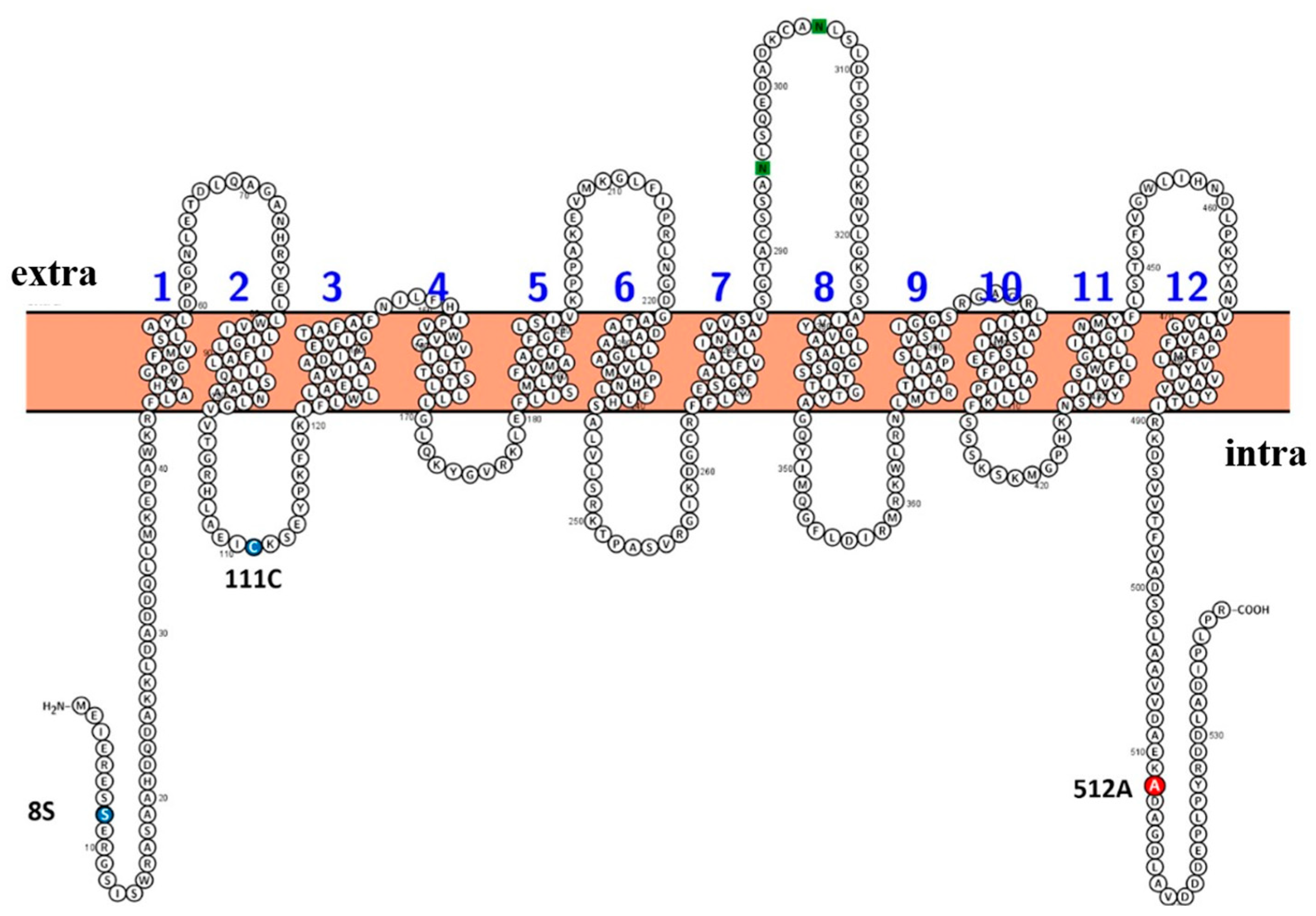

Table 1.
Sequences of mutant OsNRAMP5, and the locations of the mutations from the start codon of OsNRAMP5.
Table 1.
Sequences of mutant OsNRAMP5, and the locations of the mutations from the start codon of OsNRAMP5.
| Pattern | Plasmid No. | Mutation Location | Change in base | Chang in a.a. |
|---|---|---|---|---|
| 1 | 6, 69, 75, 76, 77, 78, 79, 80, 81, 87, 95, 97, 98 | #21 #1534 |
C->T G->A |
− A->T |
| 2 | 9, 73 | #22 #291 #332 #1534 |
A->C T->C G->A G->A |
S->R − C->Y A->T |
| 3 | 35, 36, 70, 82 | #507 #1534 |
T->C G->A |
− A->T |
| 4 | 89 | #22 #332 #1534 |
A->C G->A G->A |
S->R C->Y A->T |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



