
Submitted:
05 November 2023
Posted:
09 November 2023
You are already at the latest version
Abstract
The global shrimp farming industry is impacted by various infectious diseases, leading to marked production losses. Virulent Vibrio parahaemolyticus strains (VPAHPND) and other Vibrio species cause acute hepatopancreatic necrosis disease (AHPND) in shrimp. We isolated a binary toxin PirA/B-bearing V. campbellii strain (HJ-2023) from a Korean shrimp farm. The biochemical characteristics and antibiotic susceptibility of HJ-2023 were analyzed and compared to those of PirA/B-bearing VPAHPND. Non-AHPND-causing V. campbellii was the control. The pathogenicity of each strain was analyzed in white-leg shrimp. Phylogenetic analysis classified the isolated HJ-2023 strain as V. campbellii. Biochemical tests confirmed distinct properties of HJ-2023, compared to the genetically similar V. harveyi group. The AHPND-causing toxin gene of HJ-2023 showed 100% similarity to the rpoD gene of VCAHPND. An antibiotic disc susceptibility test indicated that HJ-2023 showed resistance to ampicillin, clindamycin, lincomycin, and erythromycin, which is effective against gram-positive bacteria, and showed weak sensitivity to gentamycin. Furthermore, HJ-2023 was 25% less toxic than VPAHPND but could be lethal during aquaculture-related outbreaks. These results highlight the complexity and virulence of AHPND-causing strains through multiple pathways and confirm the pathogenicity of V. campbellii, which increases the risk posed by AHPND to shrimp aquaculture. Our findings could help prevent AHPND.
Keywords:
Acute hepatopancreatic necrosis disease
; Vibrio parahaemolyticus
; Vibrio campbellii
; Litopenaeus vannamei
1. Introduction
Of the aquatic invertebrate species that are farmed worldwide, shrimp comprise a fast-growing species with a wide global market, with approximately 4 million tons being produced annually [1]. However, infectious diseases can impede productivity in shrimp farms; additionally, novel pathogens arising from mutations can emerge in farm environments. Acute hepatopancreatic necrosis disease (AHPND), a representative bacterial disease, has occurred frequently in shrimp farms over the past 12 years, causing enormous economic losses of more than $1 billion annually throughout Asia, where approximately 70% of global shrimp is produced [2,3,4]. Before being named AHPND, it was known as early mortality syndrome (EMS) [5]. AHPND was first discovered in China in 2009 [6] and has since spread to various countries in Southeast Asia and North America [7,8,9].
In 2013, the main causative agent of AHPND was identified to be Vibrio parahaemolyticus, a gram-negative, rod-shaped bacterium [10]. V. parahaemolyticus contains the pVA1 plasmid that expresses the fatal binary toxin PirA/B, which causes sudden death in shrimp [11]. This plasmid contains the toxin PirA/B genes of Photorhabdus and Xenorhabdus and is homologous to the insect toxin expression plasmid. When V. parahaemolyticus acquires a plasmid expressing the toxin genes PirA and PirB (15.75 and 56.12 kDa, respectively), it causes AHPND in shrimp [12,13,14,15].
V. parahaemolyticus AHPND is called EMS because it causes high mortality rates of 40–100% for approximately 8–30 d in giant tiger prawns (Penaeus monodon) and white-leg shrimp (Penaeus vannamei) in the post-larval stage after stocking [13,16]. However, AHPND has been shown to affect not only post-larval stages but also various growth stages, including the adult stage [17]. Additionally, shrimp affected by AHPND exhibit extensive necrosis and the detachment of tubular epithelial cells, empty digestive tracts, pale-to-white hepatopancreases, lethargy, anorexia, and slow growth [18,19].
Outbreaks of V. parahaemolyticus AHPND first occurred in China in 2009, followed by Vietnam (2010), Malaysia (2011), Thailand (2012), Mexico (2013), the Philippines (2015), and South America (2016) [11,14,19,20]. Initially, V. parahaemolyticus was identified as the only causative agent of AHPND; however, other Vibrio strains carrying a similar pVA1 plasmid containing PirA/B toxin genes have recently been reported in different parts of the world, including in Vibrio campbellii from Latin America [21], V. owensii from China [16], V. harveyi from Malaysia [22], and V. punensis from South America [23].
In South Korea, AHPND outbreaks caused by V. parahaemolyticus strains were reported in a shrimp farm on the west coast in 2016 and have continued to occur [24], but there have been no reports of AHPND outbreaks caused by other Vibrio spp. strains. In this study, we identified a novel Vibrio strain encoding an AHPND-causing toxin gene. The strain was isolated from a domestic vannamei shrimp farm, identified, and evaluated for its characteristics and toxicity.
2. Results
2.1. Bacterial identification via PCR
To identify the collected bacteria, PCR was performed on the HJ-2023 strain isolated from the hepatopancreases of white-leg shrimp and on the control bacteria (VPAHPND and VCnon-AHPND). PCR analysis was performed using primers targeting the 16S rRNA and rpoD genes that can detect the Vibrio strains HJ-2023, VPAHPND, and VCnon-AHPND. The toxR gene that detects V. parahaemolyticus was detected only in VPAHPND, and the vch gene sequence that detects V. campbellii was detected in the isolated HJ-2023 strain and VCnon-AHPND. Similarly, only HJ-2023 and VPAHPND were positive for gene sequences targeting the virulence genes PirA (AP3) and PirB, which were present in the pVA1 plasmid (Figure 1).
2.2. Phylogenetic tree based on pathogenic rpoD
Phylogenetic analysis of the rpoD gene grouped the isolated HJ-2023 strain with V. campbellii strains registered in GenBank and showed a close relationship between V. campbellii and V. harveyi (Figure 2).
2.3. Biochemical characteristics
The biochemical properties of the isolated HJ-2023 strain were determined using API Tests 20E and 20NE. Biochemical tests revealed differences in ODC, GEL, ARA, PNPG, MNE, and GNT between the AHPND and non-AHPND strains (Table 1). Furthermore, differences were observed in GLU, NAG, ADI, and CIT between the VPAHPND and HJ-2023 strains. The biochemical reactions of the isolated HJ-2023 strain were different from those of the existing strains; the HJ-2023 strain showed positive reactions for ONPG and ESC (Table 1 and Table 2).
2.4. Antibiotic susceptibility test
An antibiotic susceptibility test of the isolated HJ-2023 strain and the control bacteria VPAHPND and VCnon-AHPND revealed that the strains were resistant to streptomycin, erythromycin, cephalothin, kanamycin, vancomycin, ampicillin, clindamycin, and lincomycin. Furthermore, the strains exhibited sensitivity to norfloxacin, nalidixic acid, chloramphenicol, oxytetracycline, and tetracycline. The isolated HJ-2023 strain and VPAHPND showed an intermediate level of resistance to ciprofloxacin, whereas VCnon-AHPND showed sensitivity. The isolated HJ-2023 strain also showed an intermediate level of resistance to gentamicin, whereas the VPAHPND and VCnon-AHPND strains were gentamicin-resistant (Table 3).
2.5. Comparison of the virulence of the isolated HJ-2023 strain and that of the controls
After inoculating white-leg shrimp with the isolated HJ-2023 strain and the control strains, the cumulative mortality rate was observed for 5 d. In the high challenge concentration (2×106 CFU/50 µL) experiments, the average cumulative mortality rates were 90%, 95%, and 10% in the isolated HJ-2023 strain, VPAHPND, and VCnon-AHPND groups, respectively, compared to 0% mortality in the control group (no bacteria inoculated) at 12 h after infection. Comparing the virulence of the isolated HJ-2023 strain and VCnon-AHPND groups with that of the PBS control group further showed average cumulative mortality rates of 95% and 85%, respectively, in the high concentration challenge experiment and 30% and 0%, respectively, in the low concentration challenge experiment at 72 h. In the low challenge concentration (2×105 CFU/50 µL) experiments, the average cumulative mortality rates of the shrimp in the HJ-2023, VPAHPND, and VCnon-AHPND groups were 40%, 55%, and 0%, respectively. The cumulative mortality rate was maintained for 120 h (Figure 3).
3. Discussion
AHPND can cause serious economic losses by affecting major aquaculture species, such as Litopenaeus vannamei, Penaeus monodon, and M. rosenbergii [18], and has been listed by the World Organization for Animal Health in 2017 [10]. In South Korea, AHPND was designated as a statutory infectious disease for aquatic life in species subject to slaughter in 2021 [1]. White-leg shrimp, which is the most important species in the global shrimp farming industry and accounts for the majority of shrimp in South Korea, with high production and consumption rates, is one of the main species affected by AHPND [24].
We found that the bacteria from the white-leg shrimp farm were positive for the PirA/B toxin genes; however, the presence of V. parahaemolyticus could not be detected using PCR. Therefore, we assumed that it was the AHPND of another species and identified and characterized the isolated strain. In the case of Vibrio spp., it is difficult to distinguish between species because of their high genetic similarity. In particular, the 16S rRNA gene sequence of the Harveyi clade, which causes AHPND, shares more than 99% sequence homology with other species [25]. Therefore, we performed a PCR analysis using the rpoD and toxR gene sequences, which showed the minimum intraspecific similarity between V. parahaemolyticus and other Vibrio spp. [26]. It was confirmed that the isolated strain was not V. parahaemolyticus. The isolated strain was suspected to be V. campbellii among various Vibrio spp. Therefore, PCR was performed targeting the Vch gene, a V. campbellii detection gene, and the bacterium was identified as V. campbellii. The presence of the pVA1 plasmid containing the AHPND-inducing toxin genes (PirA/B) was confirmed using PCR followed by gene sequencing analysis. Most of the bacterial strains harboring pVA1 had 99.92–100% nucleotide sequence homology with each other [27]; the gene sequence from the isolated strain was 100% identical to the PirA/B toxin gene sequences of VPAHPND, the control strain used in this study (data not shown). The isolated strain was named V. campbellii AHPND (HJ-2023), and its characteristics and virulence were studied.
The PirA/B binary toxin is essential for the development of pathogenic lesions in AHPND.
According to reports on the PirA/B toxin genes present in pVA1, V. parahaemolyticus strains lacking PirA/B fail to cause AHPND; however, the strains transformed with plasmids containing PirA/B have been found to show clinical signs of AHPND and cause 100% mortality in shrimp larvae [28,29]. Additionally, pVA1 can be transferred among similar clusters and is capable of self-transfer using the pndA PSK gene system, which is a mobilizing gene associated with conjugative transfer [14]. Therefore, the pVA1 plasmid has the potential to be transferred among different Vibrio spp. or bacterial species through horizontal transfer [14,15,30]. Dong et al. [27] showed that the pVA1 plasmid can be transferred from AHPND-causing V. campbellii to non-AHPND-causing V. owensii in a controlled laboratory environment. The authors further provided direct evidence of horizontal plasmid transfer among Vibrio species. Accordingly, it is highly likely that the pVA1 plasmid from the V. parahaemolyticus was horizontally transferred to the V. campbellii strain.
Based on the phylogenetic analysis, the isolated HJ-2023 strain was classified into the V. campbellii group and confirmed to be biochemically different from the genetically similar V. harveyi group using biochemical tests [31]. The AHPND-causing gene from the isolated HJ-2023 strain also showed 100% similarity to the rpoD gene of VCAHPND, which was previously reported in China and Thailand [20,32]. To date, the Vibrio spp. that have been reported to cause AHPND are V. campbellii, V. harveyi, and V. owensii, all of which belong to the Harveyi clade of the Vibrionaceae family and are considered as the pathogenic clade of AHPND [33]. However, a recent study reported that V. punensis, a member of the Orientalis clade, was introduced with a toxin plasmid that causes AHPND [23]. Therefore, AHPND is not limited to one bacterial clade, and there is a possibility that the toxin plasmid can be transferred to various bacterial strains. This is likely to increase the virulence and spread of the disease via transfer of the toxin plasmid to potential bacterial pathogens via various routes.
Based on the biochemical characteristics determined using API Test 20E and 20NE and the percentage of identification (% ID), the isolated HJ-2023 strain showed 99.3% and 84.8% similarity to V. vulnificus and V. alginolyticus, respectively. VCnon-AHPND was identified as V. parahaemolyticus (81.8% similarity) and V. alginolyticus (97.9% similarity). Additionally, VPAHPND was found to be similar to V. parahaemolyticus (99.9%). A previous study reported that the classification of Vibrio species based on the biochemical characteristics of the API test is somewhat inappropriate [34]. As a result of the reading, however, the control VPAHPND was clearly classified as V. parahaemolyticus, and the isolated HJ-2023 and VCnon-AHPND strains were classified as V. alginolyticus and V. vulnificus. The isolated HJ-2023 strain was confirmed to be V. campbellii via gene sequence identification, but the same result was not obtained in the bacterial identification based on the biochemical characteristics determined using the API tests, as the information on V. campbellii was not registered in the bacterial identification reading software; therefore, it was likely confirmed as a similar Vibrio species. Because of this diversity in biochemical characteristics, when bacteria are isolated from diseased white-leg shrimp using TCBS medium, approximately 84.3% of them form green colonies [22]. V. campbellii also forms green colonies because it lacks a sucrose hydrolase gene, making it difficult to distinguish it from V. parahaemolyticus [20].
In the antibiotic disc susceptibility test, both the isolated HJ-2023 strain and the control bacteria VPAHPND and VCnon-AHPND showed strong resistance to ampicillin, clindamycin, and lincomycin, with a small inhibition area of 0 mm. Most Vibrio strains are resistant to ampicillin and lincomycin; therefore, they showed similar results [35]. The isolated HJ-2023 strain showed strong resistance against erythromycin, which is effective against gram-positive bacteria, in an area of 0 mm, whereas VPAHPND and VCnon-AHPND showed weak resistance. The isolated HJ-2023 strain further showed weak sensitivity to gentamycin, but VPAHPND and VCnon-AHPND showed resistance. Taken together, these results indicate that even within the same Vibrio genus, susceptibility and resistance to antibiotics differ depending on the species [36].
To compare the pathogenicity of the isolated HJ-2023 strain and that of the control bacteria VPAHPND and VCnon-AHPND, we observed the cumulative mortality rate after infection in white-leg shrimp. When the shrimp were artificially challenged with high concentrations of the isolated HJ-2023 strain and VPAHPND strains, more than 90% mortality was observed within 12 h. These results were similar to those of multiple studies that showed 100% mortality within 12–36 h in VPAHPND and VCAHPND artificial infection experiments [27,30]. In the low concentration experiments, when the concentration was 10-times lower than the high concentration, the cumulative mortality rates of the isolated HJ-2023 strain and VPAHPND were 30% and 55% within 72 h, respectively. Accordingly, the isolated HJ-2023 strain showed approximately 25% less toxicity than VPAHPND; however, it is undoubtedly a lethal strain when outbreaks occur in aquaculture fields.
V. campbellii is widely distributed in marine environments and is an important pathogen in marine shrimp and fish [37]. For a long time, V. campbellii was often misidentified because of its high similarity to V. harveyi and was considered non-pathogenic to shrimp. However, according to the results of the present study, although VCnon-AHPND itself causes mortality in shrimp, V. campbellii is conferred this ability by the pVA1 plasmid and appears to potentially cause stronger pathogenicity in shrimp. AHPND-causing V. harveyi is more virulent than the V. harveyi involved in non-AHPND [22]. The present study indicates that when devising measures to prevent the spread of AHPND, quarantine focusing on the V. parahaemolyticus strain, which is believed to be a unique strain of AHPND, is not a clear solution. In the present study, even the same Vibrio spp. showed various differences in their antibiotic susceptibility and biochemical characteristics. These results highlight the complexity of AHPND-causing strains and their virulence through multiple pathways, which may increase the risk of AHPND in the shrimp aquaculture industry.
Additional studies are needed to determine how the biochemical properties of VCnon-AHPND change as toxin plasmids are acquired. The present study may serve as a basis for attaining a better understanding of and preventing AHPND caused by various bacteria in shrimp aquaculture. Based on the results of this study, effective biosecurity measures should be considered to prevent the spread of AHPND caused by various bacterial strains.
4. Materials and methods
4.1. Sampling and polymerase chain reaction (PCR)
Penaeus vannamei was sampled from a shrimp farm in Chungcheongnam-do, Taean-gun, South Korea. The hepatopancreases of five randomly selected shrimp were aseptically obtained and plated on thiosulfate citrate bile salt sucrose (TCBS) (Difco, Franklin Lakes, NJ, USA). The plate was then incubated at 27 °C for 48 h, and a single colony was selected and subjected to PCR. The control strains used included V. parahaemolyticus [VPAHPND (ATCC 17802)], which causes AHPND, and V. campbellii [VCnon-AHPND (KCTC 2716)], which does not cause AHPND. Primer sets used to identify the Vibrio species are provided in Table 4. For the PCR, a 20-μL reaction was set up with 1 μL of each colony dilution as a template, 1 μL of each primer, and 10 μL of ExPrime Taq DNA Premix (Genetbio, Daejeon, South Korea). The thermal cycling conditions were as follows: initial denaturation at 95 °C for 10 min, followed by 30 cycles of 5 min denaturation at 95 °C, 30 s annealing at 42 °C (for PirA), 45 °C (for PirB), 50 °C (for rpoD and AP3), 52 °C (for vch), and 54 °C (for 16S rRNA and toxR), and extension at 72 °C for 1 min, with a final extension step at 72 °C for 7 min. The PCR products were electrophoresed on 1% agarose gels and analyzed using a Gel-Doc UV transilluminator (Bio-Rad, Hercules, CA, USA). We named the identified Vibrio campaberii AHPND strain as HJ-2023.
4.2. Phylogenetic analysis based on pathogenic rpoD
The rpoD gene amplified via PCR from the isolated HJ-2023 strain was cloned using pGEM®-T Easy Vector Systems (Promega, Madison, WI, USA). Sequencing was performed by COSMOgenetech (Seoul, Korea). The sequences were further compared with those of other available Vibrio core group member (V. campbellii, V. harveyi, V. rotiferianus, V. parahaemolyticus, and V. alginolyticus) rpoD gene sequences using a BLAST search in the National Center for Biotechnology Information GenBank database (www.ncbi.nlm.nih.gov/BLAST). The rpoD gene sequences were then aligned, and a phylogenetic tree was constructed by bootstrapping 1,000 replicates using the neighbor-joining method in Molecular Evolutionary Genetic Analysis 11 software (http://www.megasoftware.net/).
4.3. Biochemical tests
The biochemical identification of the isolated HJ-2023 strain and control bacteria, VPAHPND and VCnon-AHPND, was performed using commercially available API 20E and API 20NE TEST Kits (BioMérieux, Marcy-l'Étoile, France). Each bacterial suspension was prepared in 1.5% NaCl solution and analyzed at 37 ± 1 °C for 24 h according to the manufacturer's instructions. The results were interpreted on the APIWEBTM (BioMerieux) website.
4.4. Antimicrobial susceptibility test
For the antibiotic susceptibility test, the isolated HJ-2023 strain and the control bacteria VPAHPND and VCnon-AHPND, at 0.5 McFarland turbidity and in total volumes of 50 µL, were spread on Luria broth (LB) agar plates containing 1.5% NaCl. Fifteen types of antibiotic discs were then prepared and incubated at 27 °C for 24 h. After the incubation period, the diameters of the transparent area surrounding the antibiotic discs were measured in millimeters to calculate the inhibition area. The strains were classified into three grades—Resistant (R), Intermediate (I), and Sensitive (S)—based on the standards of the Clinical and Laboratory Standards Institute. The types and concentrations of the 15 antibiotic discs used for the strains are as follows: ciprofloxacin (CIP; 5 µg), norfloxacin (NoR; 10 µg), gentamicin (GM; 10 µg), nalidixic acid (NA; 30 µg), chloramphenicol (C; 30 µg), oxytetracycline (T; 30 µg), streptomycin (S; 10 µg), tetracycline (TE; 30 µg), erythromycin (E; 15 µg), cephalothin (CF; 30 µg), kanamycin (K; 30 µg), vancomycin (VA; 30 µg), ampicillin (AM; 10 µg), clindamycin (CC; 2 µg), and lincomycin (L; 2 µg).
4.5. In vivo experiments to assess pathogenicity
To compare the virulence of the isolated HJ-2023 strain and the control bacteria VPAHPND and VCnon-AHPND, in vivo tests were performed on white-leg shrimp. White-leg shrimp (average weight: 25 ± 5 g) were collected from a prawn farm located in Gunsan-si, Jeollabuk-do, placed in a breeding tank maintained at 25 °C for 1 week, and used for in vivo infection experiments. The water used for breeding the shrimp was maintained with 30% salinity using sea salt (KENT Marine Reef Salt, Franklin, WI, USA). To confirm the presence or absence of an AHPND infection, the hepatopancreases of five white-leg shrimp were isolated, and non-infection was confirmed via PCR. The strains (isolated HJ-2023 and the control bacteria VPAHPND and VCnon-AHPND) were cultured in LB supplemented with 1.5% NaCl and incubated at 27 °C with shaking until the absorbance at 600 nm (OD600) reached 1.1. The shrimp were then divided into two replicates, with seven groups in total (a control group and six experimental groups). Each group comprised 10 randomly assigned white-leg shrimp. Each group was inoculated via intramuscular injection of phosphate-buffered saline (control group) or 2×106 CFU/50 µL (high challenge concentration) and 2×105 CFU/50 µL (low challenge concentration) of bacteria. After inducing the infection, the cumulative mortality rate was observed for 5 d.
5. Patents
Author Contributions
Yue Jai Kang, Hee-Jae Choi, and Ji-Hoon Lee: Conceptualization and methodology. Hee-Jae Choi and Ji-Hoon Lee: Investigation. Da-Yeon Choi and Jun-Hwan Kim: Formal analysis. Ji-Hoon Lee and Yue Jai Kang: Writing—original draft preparation and reviewing. Yue Jai Kang: editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education [grant number NRF-2021R1I1A2045209].
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Han, J. E.; Lee, S. C.; Park, S. C.; Jeon, H. J.; Kim, K. Y.; Lee, Y. S.; Park, S.; Han, S.-H.; Kim, J. H.; Choi, S.-K. Molecular detection of Enterocytozoon hepatopenaei and vibrio parahaemolyticus-associated acute hepatopancreatic necrosis disease in southeast Asian penaeus vannamei shrimp imported into Korea. Aquaculture 2020, 517, 734812. [CrossRef]
- Jeon, H. J.; Song, J. W.; Lee, C.; Kim, B.; Park, S. Y.; Kim, J. H.; Han, J. E.; Park, J. H. Antibacterial activity of bacillus strains against acute hepatopancreatic necrosis disease-causing vibrio campbellii in Pacific white leg shrimp. Fishes 2022, 7, 287. [CrossRef]
- Rattanadilog Na Phuket, T.; Charoensapsri, W.; Amparyup, P.; Imjongjirak, C. Antibacterial activity and immunomodulatory role of a proline-rich antimicrobial peptide sppr-amp1 against vibrio campbellii infection in Shrimp Litopenaeus Vannamei. Fish & Shellfish Immunology 2023, 132, 108479.
- Yu, L. H.; Teh, C. S.; Yap, K. P.; Thong, K. L. Diagnostic approaches and contribution of next-generation sequencing technologies in genomic investigation of vibrio parahaemolyticus that caused acute hepatopancreatic necrosis disease (AHPND). Aquaculture International 2020, 28, 2547–2559. [CrossRef]
- Santos, H. M.; Tsai, C.-Y.; Maquiling, K. R.; Tayo, L. L.; Mariatulqabtiah, A. R.; Lee, C.-W.; Chuang, K. P. Diagnosis and potential treatments for acute hepatopancreatic necrosis disease (AHPND): A Review. Aquaculture International 2019, 28, 169–185. [CrossRef]
- Fao, A. Report of the FAO/MARD technical workshop on early mortality syndrome (EMS) or acute hepatopancreatic necrosis syndrome (AHPNS) of cultured shrimp (under TCP/VIE/3304). FAO, Rome, 2013, 54.
- de la Peña, L.; Cabillon, N.; Catedral, D.; Amar, E.; Usero, R.; Monotilla, W.; Calpe, A.; Fernandez, D.; Saloma, C. Acute hepatopancreatic necrosis disease (AHPND) outbreaks in penaeus vannamei and P. Monodon cultured in the Philippines. Diseases of Aquatic Organisms 2015, 116, 251–254. [CrossRef]
- Dhar, A.; Piamsomboon, P.; Aranguren Caro, L.; Kanrar, S.; Adami, R.; Juan, Y. First report of acute hepatopancreatic necrosis disease (AHPND) occurring in the USA. Diseases of Aquatic Organisms 2019, 132, 241–247. [CrossRef]
- Nunan, L.; Lightner, D.; Pantoja, C.; Gomez-Jimenez, S. Detection of acute hepatopancreatic necrosis disease (AHPND) in Mexico. Diseases of Aquatic Organisms 2014, 111, 81–86. [CrossRef]
- Yu, Y.-B.; Choi, J.-H.; Kang, J.-C.; Kim, H. J.; Kim, J.-H. Shrimp bacterial and parasitic disease listed in the OIE: A Review. Microbial Pathogenesis 2022, 166, 105545.
- Tran, L.; Nunan, L.; Redman, R.; Mohney, L.; Pantoja, C.; Fitzsimmons, K.; Lightner, D. Determination of the infectious nature of the agent of acute hepatopancreatic necrosis syndrome affecting penaeid shrimp. Diseases of Aquatic Organisms 2013, 105, 45–55. [CrossRef]
- Kumar, V.; Nguyen, D. V.; Baruah, K.; Bossier, P. Probing the mechanism of VPAHPND extracellular proteins toxicity purified from vibrio parahaemolyticus AHPND strain in germ-free artemia test system. Aquaculture 2019, 504, 414–419. [CrossRef]
- Kumar, V., Roy, S., Behera, B., Bossier, P., Das, B. Acute hepatopancreatic necrosis disease (Ahpnd): Virulence, pathogenesis and mitigation strategies in shrimp aquaculture. Toxins 2021, 13(8), 1-28. [CrossRef]
- Lee, C.-T.; Chen, I.-T.; Yang, Y.-T.; Ko, T.-P.; Huang, Y.-T.; Huang, J.-Y.; Huang, M.-F.; Lin, S.-J.; Chen, C.-Y.; Lin, S.-S.; Lightner, D. V.; Wang, H.-C.; Wang, A. H.-J.; Wang, H.-C.; Hor, L.-I.; Lo, C.-F. The opportunistic marine pathogen vibrio parahaemolyticus becomes virulent by acquiring a plasmid that expresses a deadly toxin. Proceedings of the National Academy of Sciences 2015, 112, 10798–10803. [CrossRef]
- Soto-Rodriguez, S. A.; Lozano-Olvera, R.; Ramos-Clamont Montfort, G.; Zenteno, E.; Sánchez-Salgado, J. L.; Vibanco-Pérez, N.; Aguilar Rendón, K. G. New insights into the mechanism of action of Pirab from vibrio parahaemolyticus. Toxins 2022, 14, 243. [CrossRef]
- Liu, L.; Xiao, J.; Zhang, M.; Zhu, W.; Xia, X.; Dai, X.; Pan, Y.; Yan, S.; Wang, Y. A vibrio owensii strain as the causative agent of AHPND in cultured shrimp, Litopenaeus Vannamei. Journal of Invertebrate Pathology 2018, 153, 156–164. [CrossRef]
- OIE. Chapter 2.2. 1. Acute Hepatopancreatic Necrosis Disease. World Organisation for Animal Health, Paris, France, 2019.
- Chang, Y.-H.; Kuo, W.-C.; Wang, H.-C.; Chen, Y.-M. Biocontrol of acute hepatopancreatic necrosis disease (AHPND) in shrimp using a microalgal-bacterial consortium. Aquaculture 2020, 521, 734990. [CrossRef]
- Kumar, R.; Ng, T. H.; Wang, H. Acute hepatopancreatic necrosis disease in penaeid shrimp. Reviews in Aquaculture 2020, 12, 1867–1880. [CrossRef]
- Dong, X.; Bi, D.; Wang, H.; Zou, P.; Xie, G.; Wan, X.; Yang, Q.; Zhu, Y.; Chen, M.; Guo, C.; Liu, Z.; Wang, W.; Huang, J. PIRABVP-bearing vibrio parahaemolyticus and vibrio campbellii pathogens isolated from the same ahpnd-affected pond possess highly similar pathogenic plasmids. Frontiers in Microbiology 2017, 8.
- Ahn, Y. S.; Piamsomboon, P.; Tang, K. F.; Han, J. E.; Kim, J. H. Complete genome sequence of acute hepatopancreatic necrosis disease-causing vibrio campbellii LA16-V1, isolated from Penaeus Vannamei cultured in a Latin American country. Genome Announcements 2017, 5. [CrossRef]
- Muthukrishnan, S.; Defoirdt, T.; Ina-Salwany, M. Y.; Yusoff, F. M.; Shariff, M.; Ismail, S. I.; Natrah, I. Vibrio parahaemolyticus and vibrio harveyi causing acute hepatopancreatic necrosis disease (AHPND) in Penaeus Vannamei (Boone, 1931) isolated from Malaysian shrimp ponds. Aquaculture 2019, 511, 734227. [CrossRef]
- Restrepo, L.; Bayot, B.; Arciniegas, S.; Bajaña, L.; Betancourt, I.; Panchana, F.; Reyes Muñoz, A. PIRVP genes causing AHPND identified in a new vibrio species (vibrio punensis) within the commensal orientalis clade. Scientific Reports 2018, 8. [CrossRef]
- Jang, G. I.; Park, J. I., Oh, E. G.; Kim, S. The relationship between acute hepatopancreatic necrosis disease (AHPND) in shrimp Litopenaeus vannamei and Vibrio parahaemolyticus Strains Isolated from shellfish and shrimp of the West Coast of Korea in 2019. Korean J Fish Aquat Sci 2020 53(5), 752–760.
- Cano-Gomez, A.; Høj, L.; Owens, L.; Andreakis, N. Multilocus sequence analysis provides basis for fast and reliable identification of vibrio harveyi-related species and reveals previous misidentification of important marine pathogens. Systematic and Applied Microbiology 2011, 34, 561–565. [CrossRef]
- Pascual, J.; Macián, M. C.; Arahal, D. R.; Garay, E.; Pujalte, M. J. Multilocus sequence analysis of the central clade of the genus vibrio by using the 16S rrna, recA, pyrh, rpod, gyrb, rctb and toxr genes. International Journal of Systematic and Evolutionary Microbiology 2010, 60, 154–165. [CrossRef]
- Dong, X.; Song, J.; Chen, J.; Bi, D.; Wang, W.; Ren, Y.; Wang, H.; Wang, G.; Tang, K. F.; Wang, X.; Huang, J. Conjugative transfer of the PVA1-type plasmid carrying the PIRABVP genes results in the formation of new AHPND-causing vibrio. Frontiers in Cellular and Infection Microbiology 2019, 9. [CrossRef]
- Muthukrishnan, S.; Defoirdt, T.; Shariff, M.; M. Y, I.-S.; Yusoff, F. M.; Natrah, I. Horizontal gene transfer of thepirabgenes responsible for acute hepatopancreatic necrosis disease (AHPND) turns a non-vibriostrain into an ahpnd-positive pathogen 2019.
- Tinwongger, S.; Nochiri, Y.; Thawonsuwan, J.; Nozaki, R.; Kondo, H.; Awasthi, S. P.; Hinenoya, A.; Yamasaki, S.; Hirono, I. Virulence of acute hepatopancreatic necrosis disease Pir ab-like relies on secreted proteins not on gene copy number. Journal of Applied Microbiology 2016, 121, 1755–1765. [CrossRef]
- Dong, X.; Wang, H.; Xie, G.; Zou, P.; Guo, C.; Liang, Y.; Huang, J. An isolate of vibrio campbellii carrying the PIR vp gene causes acute hepatopancreatic necrosis disease. Emerging Microbes & Infections 2017, 6, 1–3.
- San Luis, B. B.; Hedreyda, C. T. Analysis of a gene (VCH) encoding hemolysin isolated and sequenced from vibrio campbellii. The Journal of General and Applied Microbiology 2006, 52, 303–313. [CrossRef]
- Wangman, P.; Longyant, S.; Taengchaiyaphum, S.; Senapin, S.; Sithigorngul, P.; Chaivisuthangkura, P. Pira & B toxins discovered in archived shrimp pathogenic vibrio campbellii isolated long before EMS/AHPND outbreaks. Aquaculture 2018, 497, 494–502. [CrossRef]
- Xiao, J., Liu, L., Ke, Y., Li, X., Liu, Y., Pan, Y., & Wang, Y. Shrimp AHPND-causing plasmids encoding the PirAB toxins as mediated by pirAB-Tn903 are prevalent in various Vibrio species. Scientific reports 2017, 7(1), 42177. [CrossRef]
- Fabbro, C.; Cataletto, B.; Del Negro, P. Detection of pathogenic vibrio parahaemolyticus through biochemical and molecular-based methodologies in coastal waters of the Gulf of Trieste (North Adriatic Sea). FEMS Microbiology Letters 2010, 307, 158–164. [CrossRef]
- Elmahdi, S.; DaSilva, L. V.; Parveen, S. Antibiotic resistance of vibrio parahaemolyticus and vibrio vulnificus in various countries: A Review. Food Microbiology 2016, 57, 128–134. [CrossRef]
- Han, A.-R.; Yoon, Y.-J.; Kim, J.-W. Antibiotic resistance and plasmid profile of vibrio parahaemolyticus strains isolated from Kyunggi-Incheon Coastal Area. The Korean Journal of Microbiology 2012, 48, 22–28. [CrossRef]
- Dong, X.; Wang, H.; Zou, P.; Chen, J.; Liu, Z.; Wang, X.; Huang, J. Complete genome sequence of vibrio campbellii strain 20130629003s01 isolated from shrimp with acute hepatopancreatic necrosis disease. Gut Pathogens 2017, 9. [CrossRef]
- Kim, Y. B.; Okuda, J.; Matsumoto, C.; Takahashi, N.; Hashimoto, S.; Nishibuchi, M. Identification of vibrio parahaemolyticus strains at the species level by PCR targeted to the toxr gene. Journal of Clinical Microbiology 1999, 37, 1173–1177. [CrossRef]
- Sirikharin, R.; Taengchaiyaphum, S.; Sanguanrut, P.; Chi, T. D.; Mavichak, R.; Proespraiwong, P.; Nuangsaeng, B.; Thitamadee, S.; Flegel, T. W.; Sritunyalucksana, K. Characterization and PCR detection of binary, pir-like toxins from vibrio parahaemolyticus isolates that cause acute hepatopancreatic necrosis disease (AHPND) in shrimp. PLOS ONE 2015, 10. [CrossRef]
- Phiwsaiya, K.; Charoensapsri, W.; Taengphu, S.; Dong, H. T.; Sangsuriya, P.; Nguyen, G. T.; Pham, H. Q.; Amparyup, P.; Sritunyalucksana, K.; Taengchaiyaphum, S.; Chaivisuthangkura, P.; Longyant, S.; Sithigorngul, P.; Senapin, S. A natural vibrio parahaemolyticus Δ pira vp pirb vp+ mutant kills shrimp but produces neither PIR vp toxins nor acute hepatopancreatic necrosis disease lesions. Applied and Environmental Microbiology 2017, 83.
Figure 1.
PCR detection of the toxR, vch, AP3, rpoD, 16S, PirA, and PirB genes. Line M, 100-bp DNA ladder; Line 1, VPAHPND; Line 2, HJ-2023; Line 3, VCnon-AHPND; Line 4, Negative control.
Figure 1.
PCR detection of the toxR, vch, AP3, rpoD, 16S, PirA, and PirB genes. Line M, 100-bp DNA ladder; Line 1, VPAHPND; Line 2, HJ-2023; Line 3, VCnon-AHPND; Line 4, Negative control.
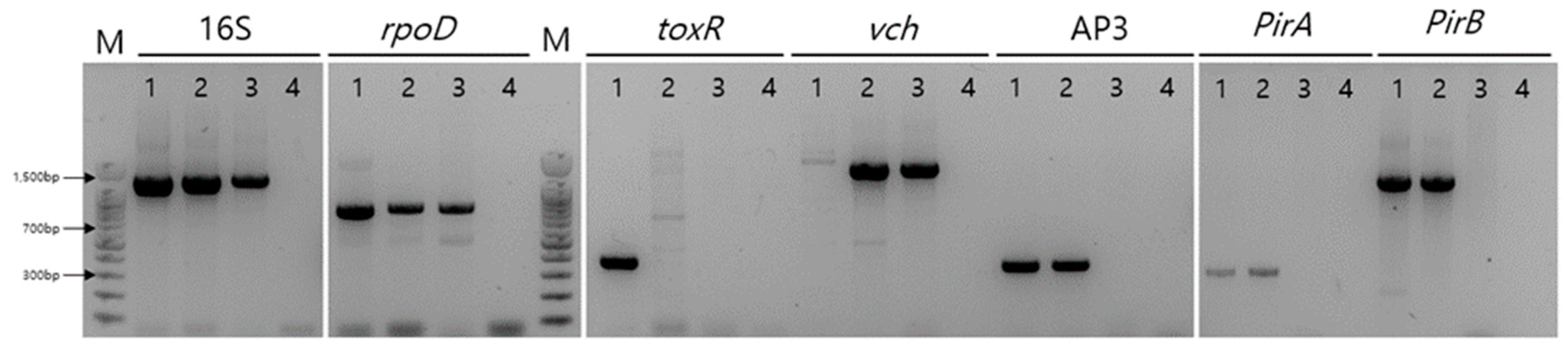
Figure 2.
Phylogenetic tree based on concatenated rpoD gene sequences. The analysis was performed using the neighbor-joining method. Bar = 0.01 expected nucleotide substitutions per site. The neighbor-joining tree reflects the percentage of trees based on 1,000 bootstrap sets possessing the branch (MEGA 7 software). Percentages are displayed for the results that were ≥ 10% at the internal nodes.
Figure 2.
Phylogenetic tree based on concatenated rpoD gene sequences. The analysis was performed using the neighbor-joining method. Bar = 0.01 expected nucleotide substitutions per site. The neighbor-joining tree reflects the percentage of trees based on 1,000 bootstrap sets possessing the branch (MEGA 7 software). Percentages are displayed for the results that were ≥ 10% at the internal nodes.
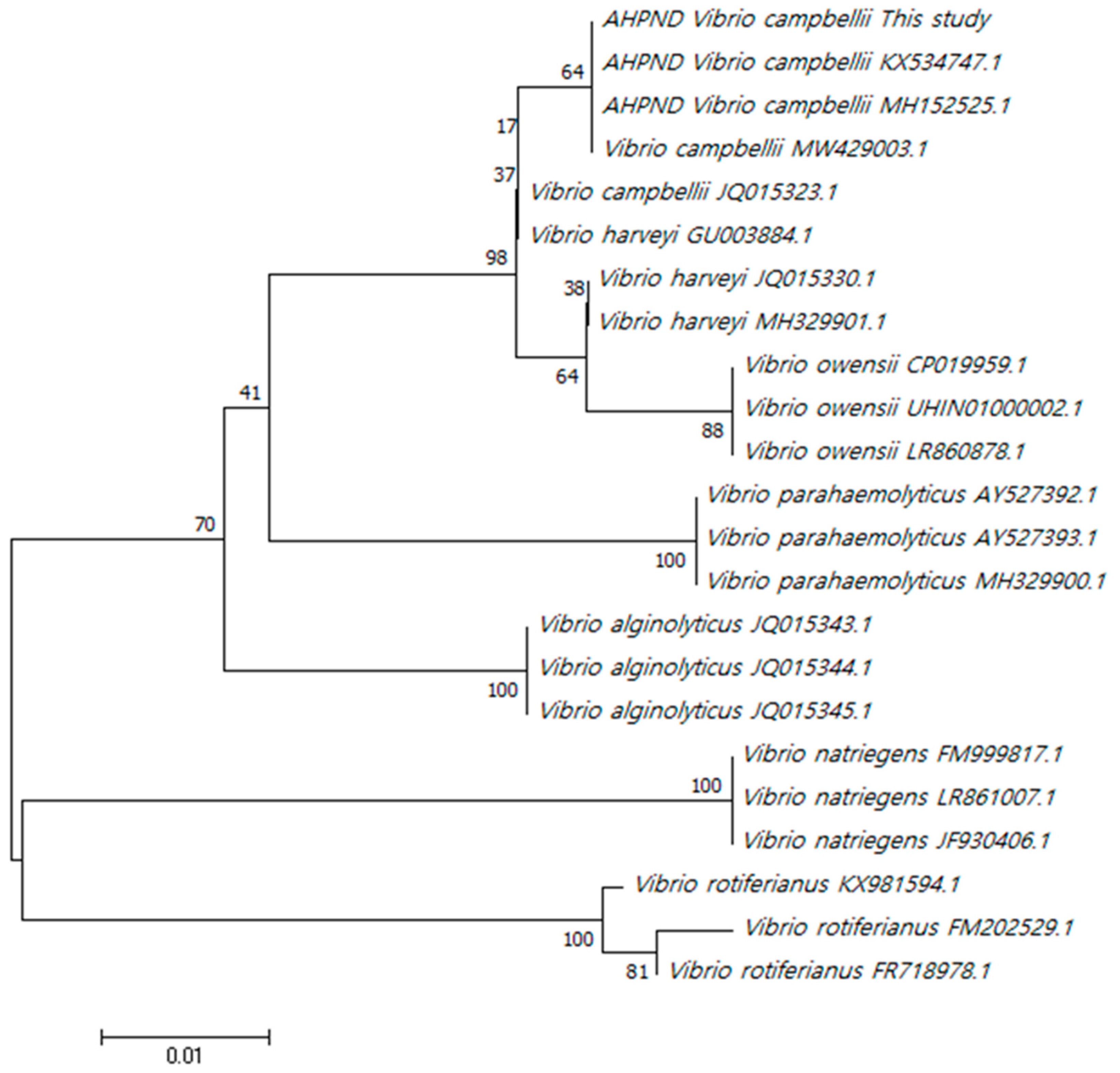
Figure 3.
Cumulative mortality rates of white-leg shrimp injected with HJ-2023, VPAHPND, or VCnon-AHPND (high concentration, 2×106 CFU/50 µL; and low concentration, 2×105 CFU/50 µL). The control groups were injected with phosphate-buffered saline. The groups were observed every 12 h for 5 d after the injection.
Figure 3.
Cumulative mortality rates of white-leg shrimp injected with HJ-2023, VPAHPND, or VCnon-AHPND (high concentration, 2×106 CFU/50 µL; and low concentration, 2×105 CFU/50 µL). The control groups were injected with phosphate-buffered saline. The groups were observed every 12 h for 5 d after the injection.
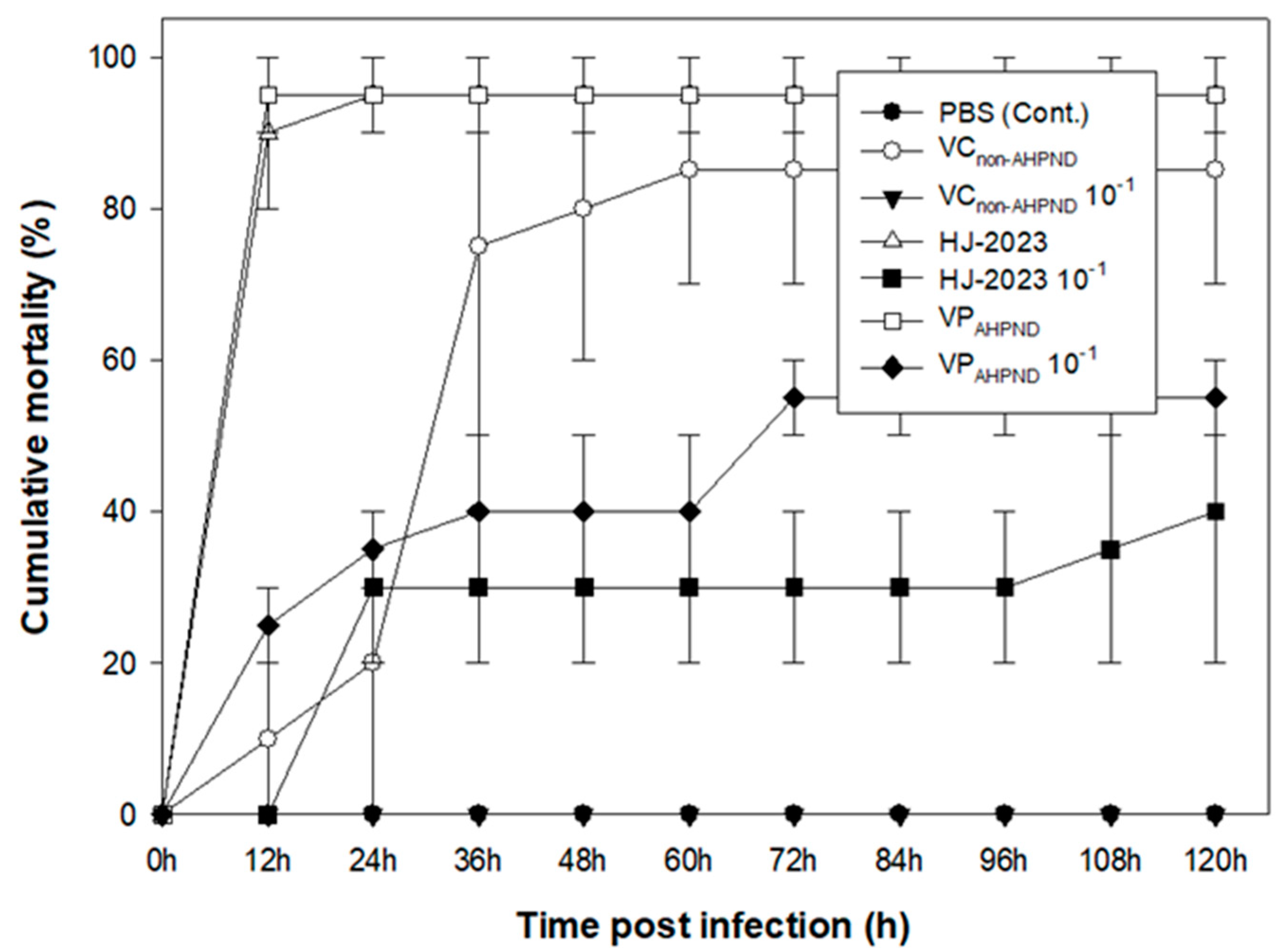

Table 1.
API test 20E.
| Isolates | VPAHPND | HJ-2023 | VCNon-AHPND | |
| Biochemical test | Abbreviation | |||
| β-galactosidase | ONPG | - | + | - |
| Arginine dihydrolase | ADH | - | - | - |
| Lysine décarboxylase | LDC | + | + | + |
| Ornithine décarboxylase | ODC | - | + | + |
| Citrate utilization | CIT | - | - | - |
| H2S production | H2S | - | - | - |
| Urease production | URE | - | - | - |
| Tryptophane désaminase | TDA | - | - | - |
| Indole production | IND | + | + | + |
| Acétoïne production (Vogues–Proskawer reaction) | VP | - | - | - |
| Gelatinase production | GEL | - | + | + |
| Glucose production | GLU | + | + | + |
| Manitol acidification | MAN | + | + | + |
| Inositol acidification | INO | - | - | - |
| Sorbitol acidification | SOR | - | - | - |
| Rhamose acidification | RHA | - | - | - |
| Saccharose acidification | SAC | - | - | - |
| Melobiose acidification | MEL | - | - | - |
| Amygdalin acidification | AMY | + | + | - |
| Arabinose acidification | ARA | - | - | + |
| Nitrite metabolism | NO2 | + | + | + |
| Oxidase test | OXY | + | + | + |
Table 2.
API test 20NE.
| Isolates | VPAHPND | HJ-2023 | VCNon-AHPND | |
| Biochemical test | Abbreviation | |||
| Nitrate reduction | NO3 | + | + | + |
| Indole production | TRP | + | + | + |
| Glucose fermentation | GLU | + | + | + |
| Arginine Dihydrolase | ADH | - | - | - |
| Urease production | URE | - | - | - |
| Hydrolysis (β-glucosidase) (ESCulin) | ESC | - | + | - |
| Hydrolysis (protease) (GELatin) | GEL | - | + | + |
| β-galactosidase (Para-Nitrophenyl-βD-Galactopyanosidase) | PNPG | - | + | + |
| Glucose assimilation | GLU | - | - | + |
| Arabinose assimilation | ARA | - | - | + |
| Mannose assimilation | MNE | + | + | + |
| Mannitol assimilation | MAN | - | - | + |
| N-acetyl-glucosamine assimilation | NAG | - | - | + |
| Maltose assimilation | MAL | + | + | + |
| Potassium gluconate assimilation | GNT | - | + | + |
| Capric acid assimilation | CAP | - | - | - |
| Adipic acid assimilation | ADI | - | - | + |
| Malate assimilation | MLT | + | + | + |
| Trisodium citrate assimilation | CIT | - | - | + |
| Phenylacetic acid assimilation | PAC | - | - | - |
| Oxidase test | OXY | + | + | + |
Table 3.
Mean and standard deviations of the diameters of the antibiotic inhibition zones (mm).
| Antibiotics(Conc.) | HJ-2023 | VPAHPND | VCnon-AHPND |
| CIP (5 µg) | 18.8 ± 0.2 (I) | 16.5 ± 0.1 (I) | 23.0 ± 0.1 (S) |
| NoR (10 µg) | 20.3 ± 0.3 | 20.0 ± 0.0 | 17.5 ± 0.1 |
| GM (10 µg) | 12.3 ± 0.1 (I) | 11.2 ± 0.2 (R) | 11.5 ± 0.1 (R) |
| NA (30 µg) | 21.0 ± 0.2 (S) | 23.2 ± 0.2 (S) | 28.2 ± 0.2 (S) |
| C (30 µg) | 23.3 ± 0.2 (S) | 19.7 ± 0.1 (S) | 32.5 ± 0.2 (S) |
| T (30 µg) | 22.3 ± 0.3 (S) | 20.5 ± 0.3 (S) | 31.2 ± 0.1 (S) |
| S (10 µg) | 8.3 ± 0.1 (R) | 9.5 ± 0.1 (R) | 0.0 ± 0.0 (R) |
| TE (30 µg) | 23.3 ± 0.1 (S) | 19.5 ± 0.3 (S) | 21.0 ± 0.1 (S) |
| E (15 µg) | 0.0 ± 0.0 (R) | 9.5 ± 0.1 (R) | 11.0 ± 0.1 (R) |
| CF (30 µg) | 9.8 ± 0.1 (R) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) |
| K (30 µg) | 12.3 ± 0.1 (R) | 12.0 ± 0.2 (R) | 10.5 ± 0.1 (R) |
| VA (30 µg) | 11.0 ± 0.0 (R) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) |
| AM (10 µg) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) |
| CC (2 µg) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) |
| L (2 µg) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) | 0.0 ± 0.0 (R) |
The non-observation of the inhibition zone around the disc was considered a 0-mm inhibition diameter. Resistant (R), sensitive (S), and intermediate (I). Ciprofloxacin (CIP; 5 µg), norfloxacin (NoR; 10 µg), gentamicin (GM; 10 µg), nalidixic acid (NA; 30 µg), chloramphenicol (C; 30 µg), oxytetracycline (T; 30 µg), streptomycin (S; 10 µg), tetracycline (TE; 30 µg), erythromycin (E; 15 µg), cephalothin (CF; 30 µg), kanamycin (K; 30 µg), vancomycin (VA; 30 µg), ampicillin (AM; 10 µg), clindamycin (CC; 2 µg), and lincomycin (L; 2 µg).
Table 4.
Primers used for bacterial identification.
| Target | Primer | Sequences (5′-3′) | Product size (bp) | Reference |
| 16S rRNA | V16S-F | GGG GAT AAC CAT TGG AAA CGA T | 1,313 | This study |
| V16S-R | AAA CTA CCT ACT TCT TTT GCA GC | |||
| rpoD | VRpoD-F | ATT GAA GAA GGT ATT AAC CAA G | 755 | This study |
| VRpoD-R | GAC ATR CGA CGG CTG ATG TCT TT | |||
| toxR | toxR-F | AAC CCG CTT TCT TCA GAC TC | 399 | [38] |
| toxR-R | AAC GAG TCT TCT GCA TGG TG | |||
| Vch | VCH-F | ATT AGA TAT TTG ACT GAA TGG CG | 1,359 | [31] |
| VCH-R | CTT AGA ATG GAT GAT TCG AAA GT | |||
| AP3 | AP3-F | ATG AGT AAC AAT ATA AAA CAT GAA AC | 333 | [39] |
| AP3-R | GTG GTA ATA GAT TGT ACA GAA | |||
| PirA | VpPirA-F | ATG AGT AAC AAT ATA AAA C | 336 | [40] |
| VpPirA-R | TTA GTG GTA ATA GAT TG | |||
| PirB | VpPirB-F | ATG ACT AAC GAA TAC GT | 1,317 | |
| VpPirB-R | CTA CTT TTC TGT ACC AA |
16S rRNA, 16S ribosomal RNA; AP3, AHPND primer 3; PirA and PirB, Photorhabdus insect-related (Pir) toxins A and B; rpoD, RNA polymerase sigma factor; toxR, Cholera toxin regulator; Vch, V. campbellii hemolysin.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



