
Submitted:
10 November 2023
Posted:
14 November 2023
You are already at the latest version
Abstract
When the zoonotic parasite of rodents that can cause human neuroangiostrongyliasis, i.e., Angiostrongylus cantonensis, is found in its natural definitive hosts, it is usually reported in isolation, as if the rat lungworm were the only component of the within-rat parasite community. In this study we report the coinfections found in rats naturally infected by A. cantonensis in urban populations of Rattus norvegicus and Rattus rattus in Valencia, Spain. In addition to the rat lungworms, which were found in 14 of the 125 rats studied (a prevalence of 11.20%), 18 other parasite infrapopulations (intestinal and tissular protists, microsporidia, and helminths) were found; some of them with high burdens. Fourteen of these 19 species found are potential zoonotic parasites, namely Blastocystis, Giardia duodenalis, Cryptosporidium spp., Enterocytozoon bieneusi, Encephalitozoon hellem, Toxoplasma gondii, Brachylaima spp., Hydatigera taeniaeformis larvae, Hymenolepis nana, Hymenolepis diminuta, Angiostrongylus cantonensis, Calodium hepaticum, Gongylonema neoplasticum and Moniliformis moniliformis. Despite the limited number of rats studied, the total predominance of coinfected rats as well as their high parasite loads seem to indicate a clear trend towards parasite tolerance.

Keywords:
Angiostrongylus cantonensis
; Rattus norvegicus
; Rattus rattus
; within-rat parasite community
; coinfections
; host tolerance
1. Introduction
Angiostrongylus cantonensis is a zoonotic parasite of rodents, including mainly the Norway rat, Rattus norvegicus, and the black rat, Rattus rattus, as their natural definitive hosts, which become infected through the ingestion of its intermediate hosts, snails and slugs, or its paratenic hosts, such as freshwater prawns, frogs, land crabs, among others [1]. The parasite is known as the rat lungworm, as the adults live in the pulmonary arteries of rats. When humans accidentally become infected – via the same route of transmission as rats – the parasite can cause neuroangiostrongyliasis due to the presence of the worms in the central nervous system. Therefore, its control must be established under the One Health concept as the control of zoonoses is an integral part of this approach.
Angiostrongylus cantonensis has been reported mainly in tropical and subtropical areas, limited by low temperatures. Until recently the parasite seemed to be far away from Europe However, it was found in rats and snails in Tenerife (Canary Islands, Spain) in 2010 [2]. Yet, although Tenerife is Europe in a political sense, it is Africa geographically speaking. Several years later, in 2019, the parasite was found in hedgehogs in Mallorca (Balearic Islands, Spain) which geographically is Europe [3]. Therefore, it was only a matter of time before it was found in continental Europe, which our research group did in 2022 in urban/periurban populations of R. norvegicus and R. rattus in the city of Valencia (Spain) [4,5].
When A. cantonensis is found in its natural definitive hosts, to our knowledge, it is systematically reported in isolation, as if the rat lungworm were the only component of the within-rat parasite community, thus ignoring possible relationships with other parasite populations (infrapopulations) present in the infected rats. In this context, we report the first data on the concomitant parasite populations, i.e., protists, microsporidia and helminth coinfections, found in rats naturally infected by A. cantonensis in an urban rat population in the city of Valencia (Spain). The results obtained allow us to discuss whether the host’s response to these coinfections leads to tolerance or resistance to their parasites.
2. Materials and Methods
Our research group signed an agreement with Valencia City Council allowing us to investigate the presence of zoonotic parasites in rats trapped by the pest control company Laboratorios Lokímica, as part of the municipal pest control campaign in the city.
The trapped rats were kept at -20º C until their parasitological examination. Once thawed, the rats were dissected to extract the adult helminths from the different organs. The helminths found were studied by conventional helminthological techniques [6]. The nematodes found in the pulmonary arteries were also identified by molecular techniques [4,5]. The study of protozoans in the large intestine content was made by means of the Midi Parasep® SF (Apacor Ltd., Wokingham, UK) concentration technique followed by a multiplex PCR (AllplexTM Gastrointestinal Panel-Parasite Assay) for the detection of protist parasites such as Giardia duodenalis, Entamoeba histolytica, Cryptosporidium spp., Blastocystis, Dientamoeba fragilis, and Cyclospora cayetanensis [7]. After DNA extraction, the microsporidia Enterocytozoon bieneusi, Encephalitozoon intestinalis, E. cuniculi and E. hellem were molecularly investigated by direct PCR [8]. Toxoplasma gondii was searched for in the rat brains by quantitative PCR (qPCR) [9] and the presence of Leishmania infantum in the spleens, ears and skin of the rats was also explored by qPCR [10].
The presence of Angiostrongylus cantonensis was investigated in 125 rats, 97 R. norvegicus (43 males, 49 females and 5 indeterminate; 63 adults, 32 juveniles and 2 indeterminate) and 28 R. rattus (9 males and 19 females; 20 adults and 8 juveniles) trapped between April 2021 and March 2023. We studied rats captured at three trapping sites located in 17 of the 19 districts into which Valencia is divided, namely, in the sewer system (55 individuals), in city parks and gardens (43) and in orchards located in a peri-urban area of Valencia (27). Most of the rats, 74, were trapped in spring, 20 in autumn and 31 in winter.
Rat species were identified based on the external morphometry according to J. Gosálbez [11]. Likewise, rats were considered juveniles or adults according to their body weight and external morphometry [11].
The comparison of the prevalences between intrinsic (age and sex) and extrinsic (site and season of capture) factors was made through the 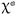 test. Statistical significance was established at p < 0.05.
test. Statistical significance was established at p < 0.05.
test. Statistical significance was established at p < 0.05. Statistical analysis was carried out using the IBM SPSS 26.0 for Windows [International Business Machines Corporation (IBM), Armonk, NY, USA] and StatView 5.0 [Statistical Analysis System (SAS) Institute Inc., Cary, NC, USA] software packages.
3. Results
3.1. Angiostrongylus cantonensis infected rats
The rat lungworm was identified in 14 of the 125 captured rats (11.20%), namely in 10 R. norvegicus (10.31%) and in 4 R. rattus (14.29%). It was found in the pulmonary arteries of 13 of the studied rats (10 R. norvegicus and 3 R. rattus). One black rat also harbored juvenile parasites in the brain and another one harbored the parasites exclusively in the brain. A total of 192 individuals of A. cantonensis were collected in the rats, with a mean intensity of 13.71 in the infected rats.
As for the sex of the rats, all R. norvegicus infected by A. cantonensis in the present study and two R. rattus [12/14 (85.71 %)] were male. Only the two juvenile black rats were female [2/14, 14.29%)]. The presence of A. cantonensis in R. norvegicus is sex-biased, with a higher prevalence of infection in males [10/43 (23.26%)] than in females (0/49); this difference being statistically significant (χ2=12.785, p=0.003). However, in the case of R. rattus, despite the low number of black rats parasitized (4/28), and although males are more highly parasitized than females [2/9 (22.22%) vs 2/19 (10.53%)], the presence of the rat lungworms in R. rattus females and its absence in R. norvegicus females, is statistically significant (χ2=5.314, p=0.0212).
Concerning the age of the rats, 11 rats infected by the nematode were adults [11/83 (13.25%)] and only 3 were juveniles [3/40 (7.5%)]. No statistically significant differences were found concerning the age of both rat species together. However, considering each rat species separately, juvenile black rats have a higher prevalence [2/8 (25.00%)] than juveniles of the Norwegian rats [1/32 (3.13%)]; this difference being statistically significant (χ2=4.414, p=0.0356).
Angiostrongylus cantonensis was found in rats trapped in 7 of the 17 surveyed districts of Valencia (41.18 %), as well as in the three trapping sites, i.e., in 6 rats of the 55 trapped in sewers (10.91 %), in 2 of the 43 caught in parks (4.65%) and in 6 of the 27 captured in the orchards (22.22%). Although the prevalences found are different according to the trapping sites, the results are not statistically significant when analyzed together due to the small sample sizes. However, there is a statistically significant difference between the prevalence found in parks compared to that found in orchards (χ2=5.509, p=0.0245).
Ten of the 74 rats trapped in spring were infected by A. cantonensis (13.51%), while 2 were found parasitized in autumn (10%) and 2 in winter (6.45%). Likewise, due to the small sample sizes, the results have no statistical significance.
3.2. Within-rat parasite community/coinfections
The parasite community in the studied organs of the 14 rats consisted of 19 different parasite infrapopulations. Table 1 shows, in addition to A. cantonensis, the 18 other parasite species found according to rat species. Specifically, the parasite community of the rats studied consisted of six protists, one trematode, three cestodes, eight nematodes and one acanthocephalan. Fourteen of these 19 species found are potentially zoonotic parasites, namely Blastocystis, Giardia duodenalis, Cryptosporidium spp., Enterocytozoon bieneusi, Encephalitozoon hellem, Toxoplasma gondii, Brachylaima spp., Hydatigera taeniaeformis larvae, Hymenolepis nana, Hymenolepis diminuta, Angiostrongylus cantonensis, Calodium hepaticum, Gongylonema neoplasticum and Moniliformis moniliformis. No rats were found infected by Leishmania infantum.
4. Discussion
4.1. Angiostrongylus cantonensis infected rats
Published prevalences of Angiostrongylus cantonensis in rats can vary from 3 to 100% depending on the endemic area [12]. In our study, we found an overall prevalence of A. cantonenesis of 11.20% in Valencia which - although not being very high - was obtained in highly populated zones. The fact that the highest A. cantonenesis prevalence was found in orchards is remarkable considering the high rate of consumption of raw vegetables in the Mediterranean diet, which poses a risk of acquiring the parasite larva through the ingestion of not sufficiently washed salads [13].
Previously, we obtained a prevalence of the rat lungworm of 8.51% when 94 rats were studied [4]. The only data on the prevalence of the nematode near Spain was obtained in Tenerife, where a prevalence of 19.19% was obtained after studying 297 rats, most of them from rural areas [14]. Therefore, although the greater the number of rats studied, the higher the prevalence, there are no statistically significant differences between these figures.
4.2. Within-rat parasite community/coinfections
The studied rats presented a rich and varied within-host parasite community, most remarkably being that 19 infrapopulations were found in only 14 rats, 14 of these parasite species being potentially zoonotic parasites, posing a possible risk of transmission to the human population with which the rats coexist.
The A. cantonensis-infected rats captured in the sewers presented the greatest parasite species richness, as only the stomach nematode Mastophorus muris and the microsporidian E. bieneusi were not found (Table 2 and Table 3).
All components of the parasite community were found in rats trapped in spring (10/14). Only two rats captured in autumn and two in winter were found to be infected by A. cantonensis, so the absence of certain parasites in these individuals cannot be discussed.
Except for the microsporidian Encephalitozoon hellem, all other parasite species, 18, were found in the 10 R. norvegicus infected by A. cantonensis and 12 different infrapopulations were found in only four individuals of R. rattus (Table 1).
Only the 10 A. cantonenesis-infected R. norvegicus analyzed in this study presented a richer parasite community than the 100 Norway rat individuals we previously studied in Barcelona [6,7,10], without even considering the microsporidians and T. gondii, which were not investigated in the Barcelona rats.
Concerning coinfections, there was no case of monoparasitism among the studied rats. Adult A. cantonensis-infected R. norvegicus harbored from 6 to 12 different infrapopulations in the same individual. The case of one adult R. norvegicus that harbored representatives of protists, trematodes, cestodes, nematodes as well as acanthocephalans in the intestine, most of the helminths with high parasite loads (Table 2, Rn IX), is remarkable. Also noteworthy is the case of one juvenile Norway rat harboring 57 individuals of A. cantonensis (26 males and 31 females) in the pulmonary arteries as well as high burdens of C. hepaticum infecting the liver (Table 2, Rn X). Both rats were trapped in the sewer system.
When analyzing the parasite community/coinfections found and the transmission routes, in the case of the monoxenous protists and microsporidia, rats become infected by the fecal/oral transmission route directly throughout the ingestion of cysts/oocysts/spores contaminating the environment, in particular the sewer system, and orchards which are not normally irrigated with safe or potable water.
The presence of T. gondii in the rats may also be related to contamination by oocysts from cat feces or by cannibalism, a common occurrence in cases of limited food supply. The absence of the usual amount of food on the streets, due to the lockdown and the closure of restaurants during the pandemic, could have led to an increase in cannibalism that favored the T. gondii life cycle.
Considering the helminth parasites, 7 worms presented an indirect or heteroxenous life cycle and 5 had a monoxenous or direct cycle (Table 1). In the case of H. nana, the parasite is able to complete its life cycle either with the intervention of an arthropod intermediate host harboring the larval stage (cysticercoid) (heteroxenous life cycle) or without the intervention of any intermediate host but directly inside the intestine of the definitive host (monoxenous life cycle). Only one R. norvegicus presented a high H. nana load (R.n. IX in Table 2), suggestive of the monoxenous-type cycle.
Among the monoxenous helminths, eggs shed in feces (or urine in the case of Trichosomoides crassicauda) are infective for the rats once the eggs embryonate in the soil. To become infected by C. hepaticum, a nematode that lives in the liver parenchyma, rats must also ingest the eggs that contaminate the environment. However, in this case, as the eggs are trapped in the liver, the rat must die in order to release the eggs which mature in the soil. It is noteworthy that all the 10 R. norvegicus were infected by C. hepaticum (Table 2). This could indicate an increase in rat mortality during the pandemic period that ultimately favored cannibalism, leading to the release of eggs into the environment enhancing the life cycle of C. hepaticum, as in the case of T. gondii.
In the case of Nippostrongylus brasiliensis (a murine model of Necator americanus), the larvae penetrate the skin of rats, or may also be ingested from the soil, and after molting and maturing in the lungs, reach the small intestine. The eggs are released in feces and hatch in the soil releasing the L1 larvae which become infective after molting. The nematode was found in almost all Norway rats at high burdens (Table 2). Exceptionally, two of them (VIII and IX) harbored hundreds and hundreds of N. brasiliensis. It is hard to believe that this life cycle does not include processes of autoinfection and that the extraordinary number of adults in the intestine is due to repeated infections.
For heteroxenous life cycles, rats must ingest the eggs of Hydatigera taeniaeformis shed in cat (definitive host) feces. Rats act as intermediate hosts harboring the metacestode (strobylocercus) in the liver parenchyma. Cats are the only predator that rats have in cities, completing the biological cycle. Several rats harbored both H. taeniaeformis and T. gondii, parasites that share a common infection route, i.e., cat feces.
To become infected by Brachylaima spp. and A. cantonensis, rats must ingest infected snails (also slugs or paratenic hosts in the case of the rat lungworms). Two Norway rats were coinfected by both helminths, leading to the hypothesis that these two parasites could have shared a snail as intermediate host.
For the rest of the heteroxenous helminths, intermediate hosts involve arthropods, mainly beetles for H. diminuta and cockroaches for Mastophorus muris, G. neoplasticum and M. moniliformis, with arthropods being an important element of the rat diet.
In terms of the host microhabitats for which the worms might compete, A. cantonenesis (adults, eggs and L1 larvae) and N. brasiliensis larvae (L3 and L4) share the same microhabitat, i.e., the lungs. In this regard, 8 of the 10 R. norvegicus and one R. rattus harbored both species (Table 2 and 3), so they do not appear to be competitors, at least in the studied rats. Likewise, 6 rats harbored A. cantonensis and T. gondii, parasites that share the brain as microhabitat at a particular time of their life cycles.
The liver was also coinfected by the tapeworm larvae of H. taeniaeformis and C. hepaticum in 3 R. norvegicus, while the small intestine presented the greatest species richness, namely up to 5 different ones (Table 1). N. brasiliensis always occupies the first part of the small intestine, the duodenum, while the remaining helminths (Hymenolepis spp., Brachylaima and M. moniliformis) are usually located in the jejunum and ileum.
As far as we are aware, only one case of coinfection has been published so far between a species of Angiostrongylus (A. chabaudii) and another parasite, specifically Dirofilaria immitis, in the definitive host, in this case in a wildcat [15]. Therefore, this study exposes, for the first time, the range of macro-, as well as micro-parasites (excluding bacteria and viruses), with which A. cantonensis can coexist in its definitive hosts in urban/periurban environments.
4.3. Host resistance or tolerance?
Hosts which are ubiquitous, like rats, are more likely to become coinfected, as are hosts that occupy different ecological niches in which several parasites are present [16]. Consequently, rats which flourish in a wide range of environmental conditions, like sewers, parks and gardens and orchards in this particular case, are exposed to a greater diversity of parasites. Once infected, hosts use two strategies to cope with their parasites: resistance or tolerance [17]. Hosts can, by different mechanisms, reduce parasite burdens (resistance) or they can minimize the damage caused by the parasite load (tolerance).
Angiostrongylus cantonensis was experimentally shown to cause a 10-20% mortality in R. norvegicus [18]. Also, an experimental study on parasite tolerance showed that mortality is related to the number of larvae of the rat lungworm used to infect rats [19]. However, it is difficult to know how to extrapolate these findings based on laboratory rats - not infected by any other parasite - to understand the consequences of coinfection in nature. Although we are aware of the limited number of rats studied, the total predominance of coinfected rats as well as their high parasite loads, seem to indicate a clear trend towards parasite tolerance.
In addition, coinfections can have negative effects on the host accelerating its mortality or, otherwise, coinfections can have positive effects on the host reducing its mortality [20]. In this context, it seems clear that if at some point coinfections led to an increase in the mortality rate of the urban rat populations in Valencia, those populations that survived, considering their high reproductive capacity, may have given rise to tolerant populations that justify these high prevalences, parasite loads, and coinfections found in this study.
Author Contributions
Conceptualization, M.T.G-P. and M.V.F.; methodology, M.T.G-P., C.G., M.T., M.G.-S., J.S., A.O., S.S.-D., M.V.F.; formal analysis, M.T.G-P. and M.V.F.; resources, M.T.G-P., C.G., M.T., M.G.-S., J.S., A.O., S.S.-D., R.B.-M., M.V.F. X.X.; writing—original draft preparation, M.T.G-P.; writing—review and editing, M.T.G-P., M.T., M.G.-S., J.S., A.O., M.V.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable. Ethics Committee approval is not necessary in our study, as all collections were conducted as part of the Municipal Pest Control Campaign. The use of snap-traps is one of the usual solutions for pest control. Following the Codes of Good Practice, these traps should be placed in areas or points not accessible to nontarget organisms (such as sewers or inside rodent safe stations in case of open areas) and checked frequently (every 4–5 days). We followed these criteria. There is no further legislation in our country on this issue.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge the Health Service of Valencia City Council for overseeing and promoting this research in the city.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cowie, R.H.; Ansdell, V.; Panosian Dunavan, C.; Rollins, R.L. Neuroangiostrongyliasis: Global Spread of an Emerging Disease. Am. J. Trop. Med. Hyg., 2022, 107, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Foronda, P.; López-González, M.; Miquel, J.; Torres, J.; Segovia, M.; Abreu-Acosta, N.; Casanova, J.C.; Valladares, B.; Mas-Coma, S.; Bargues, M.D.; et al. Finding of Parastrongylus cantonensis (Chen, 1935) in Rattus rattus in Tenerife, Canary Islands (Spain). Acta Trop. 2010, 114, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Esquivel, C; Sola, J; Delgado-Serra, S.; Puig Riera, M.; Negre, N.; Miranda, M.A.; Jurado-Rivera, J.A. Angiostrongylus cantonensis in North African hedgehogs as vertebrate hosts, Mallorca, Spain, October 2018. Euro Surveill. 2019, 24, 1900489. [Google Scholar] [CrossRef]
- Galán-Puchades, M.T.; Gómez-Samblás, M.; Osuna, A.; Sáez-Durán, S.; Bueno-Marí, R.; Fuentes, M.V. Autochthonous Angiostrongylus cantonensis Lungworms in Urban Rats, Valencia, Spain, 2021. Emerg. Infect. Dis. 2022, 28, 2564–2567. [Google Scholar] [CrossRef] [PubMed]
- Galán-Puchades, M.T.; Gómez-Samblás, M.; Osuna, A.; Sáez-Durán, S.; Bueno-Marí, R.; Fuentes, M.V. Update on the first finding of rat lungworm, Angiostrongylus cantonensis, in Rattus spp. in continental Europe, Valencia, Spain, 2022. Pathogens 2023, 12, 567. [Google Scholar] [CrossRef] [PubMed]
- Galán-Puchades, M.T.; Sanxis-Furió, J.; Pascual, J.; Bueno-Marí, R.; Franco, S.; Peracho, V.; Montalvo, T.; Fuentes, M.V. First survey on zoonotic helminthosis in urban brown rats (Rattus norvegicus) in Spain and associated public health considerations. Vet Parasitol. 2018, 259, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Galán-Puchades, M.T.; Trelis, M.; Sáez-Durán, S.; Cifre, S.; Gosálvez, C.; Sanxis-Furió, J.; Pascual, J.; Bueno-Marí, R.; Franco, S.; Peracho, V.; et al. One Health Approach to Zoonotic Parasites: Molecular Detection of Intestinal Protozoans in an Urban Population of Norway Rats, Rattus norvegicus, in Barcelona, Spain. Pathogens. 2021, 10, 311. [Google Scholar] [CrossRef] [PubMed]
- Haro, M.; Izquierdo, F.; Henriques-Gil, N.; Andrés, I.; Alonso, F.; Fenoy, S.; del Aguila, C. First detection and genotyping of human-associated microsporidia in pigeons from urban parks. Appl. Environ. Microbiol. 2005, 71, 3153–3157. [Google Scholar] [CrossRef] [PubMed]
- Reischl, U.; Bretagne, S; Krüger, D. ; Ernault, P.; Costa, J.M. Comparison of two DNA targets for the diagnosis of Toxoplasmosis by real-time PCR using fluorescence resonance energy transfer hybridization probes. BMC Infect Dis. 2003, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Galán-Puchades, M.T.; Gómez-Samblás, M.; Suárez-Morán, J.M.; Osuna, A.; Sanxis-Furió, J.; Pascual, J.; Bueno-Marí, R.; Franco, S.; Peracho, V.; Montalvo, T.; et al. Leishmaniasis in Norway Rats in Sewers, Barcelona, Spain. Emerg Infect Dis. 2019, 2019 25, 1222–1224. [Google Scholar] [CrossRef]
- Gosálbez, J. Insectivors i rosegadors de Catalunya. Metodologia d’estudi i cataleg faunistic; Ketres editora: Barcelona, Spain, 1987; p. 241. [Google Scholar]
- Wang, Q.P.; Lai, D.H.; Zhu, X.Q.; Chen, X.G.; Lun, Z.R. Human angiostrongyliasis. Lancet Infect. Dis. 2008, 8, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Prociv, P.; Spratt, D.M.; Carlisle, M.S. Neuroangistrongyliasis: unresolved issues. Int. J. Parasitol. 2000, 30, 1295–1303. [Google Scholar] [CrossRef] [PubMed]
- Martín-Carrillo, N.; Feliu, C.; Abreu-Acosta, N.; Izquierdo-Rodriguez, E.; Dorta-Guerra, R.; Miquel, J.; Abreu-Yanes, E.; Martin-Alonso, A.; García-Livia, K.; Quispe-Ricalde, M.A.; et al. A Peculiar Distribution of the Emerging Nematode Angiostrongylus cantonensis in the Canary Islands (Spain): Recent Introduction or Isolation Effect? Animals 2021, 11, 1267. [Google Scholar] [CrossRef] [PubMed]
- Gherman, C.M.; Ionica, A.M.; Deak, G.; Chisamera, G.B.; Mihalca, A.D. Coinfection with Angiostrongylus chabaudi and Dirofilaria immitis in a wildcat, Felis silvestris from Romania—a case report. Acta Vet. Brno 2019, 88, 303–306. [Google Scholar] [CrossRef]
- Morand, S. Wormy world: comparative tests of theoretical hypotheses on parasite species richness. In Evolutionary Biology of Host-Parasite Relationships: Theory Meets Reality; Poulin, R., Morand, S., Skorping, A., Eds.; Elsevier Science B.V.: Amsterdam, Netherlands, 2000; pp. 63–79. [Google Scholar]
- Rohr, J.R.; Raffel, T.R.; Hall, C.A. Developmental variation in resistance and tolerance in a multi-host–parasite system. Funct. Ecol. 2010, 24, 1110–1121. [Google Scholar] [CrossRef]
- Garcia, J.S.; Dos Santos Bonfim, T.C.; Junior, A.M.; Tunholi, V.M.; Tunholi-Alves, V.M.; Mota, E.M.; Simoes, R.D.; Santana, A.C.; Hooper, C.; Pinheiro, J.; et al. Hematological and histopathological changes in Rattus norvegicus (Wistar) experimentally infected by Angiostrongylus cantonensis (Chen, 1935). Parasitol. Int. 2014, 63, 631–637. [Google Scholar] [CrossRef]
- Ji, L.; Yiyue, X.; Xujin, H.; Minghui, Z.; Mengying, Z.; Yue, H.; Yanqi, W.; Langui, S.; Xin, Z.; Datao, L.; et al. Study on the tolerance and adaptation of rats to Angiostrongylus cantonensis infection. Parasitol Res. 2017, 116, 1937–1945. [Google Scholar] [CrossRef]
- Vaumourin, E.; Vourc’h, G.; Gasqui, P.; Vayssier-Taussat, M. The importance of multiparasitism: examining the consequences of co-infections for human and animal health. Parasites Vectors 2015, 8, 545. [Google Scholar] [CrossRef] [PubMed]

Table 1.
Parasitic infrapopulations found in 10 Rattus norvegicus and 4 Rattus rattus in urban and peri-urban areas of Valencia, Spain, 2022-2023.
Table 1.
Parasitic infrapopulations found in 10 Rattus norvegicus and 4 Rattus rattus in urban and peri-urban areas of Valencia, Spain, 2022-2023.
| Protists/Microsporidia species |
Microhabitat | Cycle | n (host) |
P (%) |
|---|---|---|---|---|
| Blastocystis | small intestine |
M | 8 (6 Rn, 2Rr) |
60 Rn 50 Rr |
| Giardia duodenalis | small intestine |
M | 9 (7 Rn, 2Rr) |
70 Rn 50 Rr |
| Cryptosporidium spp. | small intestine |
M | 1 (1 Rn) |
10 Rn |
| Enterocytoozoon bieneusi | small intestine |
M | 2 (1 Rn, 1 Rr) |
10 Rn 25 Rr |
| Encephalitozoon hellem | small intestine |
M | 1 (1 Rr) |
25 Rr |
| Toxoplasma gondii | brain | D | 6 (5 Rn, 1 Rr) |
50 Rn 25 Rr |
| Helminth species | ||||
|
Brachylaima spp. |
small intestine |
H | 1 (1 Rn) |
10 Rn |
|
Hydatigera taeniaeformis larvae |
liver | H | 5 (4 Rn,1 Rr) |
40 Rn 25 Rr |
|
Hymenolepis nana |
small intestine |
H/M | 3 (3 Rn) |
30 Rn |
|
Hymenolepis diminuta |
small intestine |
H | 4 (3 Rn,1 Rr) |
30 Rn 25 Rr |
| Angiostrongylus cantonensis | pulmonary arteries/brain | H | 14a (10 Rn, 4Rr) |
100 Rn 100 Rr |
|
Calodium hepaticumb |
liver | M | 11 (10 Rn,1 Rr) |
100 Rn 25 Rr |
|
Mastophorus muris |
stomach | H | 2 (1 Rn,1 Rr) |
10 Rn 25 Rr |
|
Eucoleus gastricus |
stomach | M | 3 (3 Rn) |
30 Rn |
|
Trichosomoides crassicauda |
urinary bladder | M | 3 (3 Rn) |
30 Rn |
|
Nippostrongylus brasiliensis |
small intestine/ lungs (larvae) |
M | 9 (8 Rn,1 Rr) |
80 Rn 25 Rr |
|
Heterakis spumosa |
large intestine |
M | 4 (3 Rn, 1Rr) |
30 Rn 25 Rr |
|
Gongylonema neoplasticum |
esophagus/ stomach |
H | 2 (2 Rn) |
20 Rn |
|
Moniliformis moniliformis |
small intestine |
H | 2 (2 Rn) |
20 Rn |
a In pulmonary arteries in 10 Rn and 3 Rr; in pulmonary arteries and brain in 1 Rn and only in brain in 1 Rr. bRange for C. hepaticum is not reported due to the difficulty in the reconstruction of dead parasites. Abbreviations: Rn, Rattus norvegicus; Rr, Rattus rattus; H, heteroxenous; M, monoxenous; n, number of parasitized hosts; P, prevalence. Potentially zoonotic species shaded in grey.
Table 2.
Concomitant parasite infrapopulations in naturally infected Rattus norvegicus (Rr) by Angiostrongylus cantonensis: numbers represent helminth loads and the total number of parasite species in individual rats is written in bold.
Table 2.
Concomitant parasite infrapopulations in naturally infected Rattus norvegicus (Rr) by Angiostrongylus cantonensis: numbers represent helminth loads and the total number of parasite species in individual rats is written in bold.
| Protists/Microsporidia species |
Rn Is |
Rn IIs |
Rn IIIo |
Rn IVo |
Rn Vo |
Rn VIo |
Rn VIIp |
Rn VIIIs |
Rn IXs |
Rn* Xs |
|---|---|---|---|---|---|---|---|---|---|---|
| Blastocystis | + | + | + | + | + | + | ||||
| G. duodenalis | + | + | + | + | + | + | + | |||
| Cryptosporidium spp. | + | |||||||||
| T. gondii | + | + | + | + | + | |||||
| E. bieneusi | + | |||||||||
| Helminth species | ||||||||||
| Brachylaima spp. | 4 | 4 | ||||||||
| H. taeniaeformis larvae | 1 | 1 | 1 | |||||||
| H. nana | 1 | 65 | ||||||||
| H. diminuta | 2 | 6 | 2 | 2 | 43 | |||||
| A. cantonensis | 2 | 4 | 7 | 13 | 7 | 3 | 33 | 4 | 19 | 57 |
| C. hepaticum | + | + | + | + | + | + | + | + | + | + |
| M. muris | 10 | 2 | ||||||||
| E. gastricus | 5 | 9 | 21 | |||||||
| T. crassicauda | 4 | 4 | 14 | |||||||
| N. brasiliensis | 80 | 65 | 31 | 6 | 2 | 25 | HH | HH | ||
| H. spumosa | 9 | 2 | 1 | |||||||
| G. neoplasticum | 1 | 3 | ||||||||
| M. moniliformis | 2 | 7 | ||||||||
| Total nº species | 8 | 6 | 6 | 10 | 7 | 8 | 7 | 8 | 12 | 3 |
s, sewers; o, orchards; p, parks. * juvenile Abbreviations: G. duodenalis, Giardia duodenalis; T. gondii, Toxoplasma gondii, E. bieneusi, Enterocytozoon bieneusi ; H. taeniaeformis larvae, Hydatigera taeniaeformis larvae; H. nana, Hymenolepis nana; H .diminuta, Hymenolepis diminuta; A. cantonensis, Angiostrongylus cantonensis; C. hepaticum, Calodium hepaticum; M. muris., Mastophorus muris; E. gastricus, Eucoleus gastricus; T. crassicauda, Trichosomoides crassicauda; N. brasiliensis, Nippostrongylus brasiliensis; H. spumosa, Heterakis spumosa; G. neoplasticum, Gongylonema neoplasticum; M. moniliformis, Moniliformis moniliformis; HH, hundreds. + Numbers are not reported due to the difficulty in the reconstruction of dead parasites. Potentially zoonotic species shaded in grey.
Table 3.
Concomitant parasite infrapopulations in naturally infected Rattus rattus (Rr) by Angiostrongylus cantonensis: numbers represent helminth loads and the total number of parasite species in individual rats is written in bold.
Table 3.
Concomitant parasite infrapopulations in naturally infected Rattus rattus (Rr) by Angiostrongylus cantonensis: numbers represent helminth loads and the total number of parasite species in individual rats is written in bold.
| Protists/Microsporidia species |
Rr* Is 
|
Rr IIp |
Rr IIIo |
Rr* IVo 
|
|---|---|---|---|---|
| Blastocystis | + | + | ||
| G. duodenalis | + | + | ||
| T. gondii | + | |||
| E. hellem | + | |||
| E. bieneusi | + | |||
| Helminth species | ||||
| H. taeniaeformis larvae | 1 | |||
| H. diminuta | 2 | |||
| A. cantonensis | 30 | 9 | 3 | 2 |
| C. hepaticum | + | |||
| M. muris | 2 | |||
| N. brasiliensis | 2 | |||
| H. spumosa | 7 | |||
| Total nº species | 4 | 3 | 6 | 4 |
s, sewers; o, orchards; p, parks. *Juveniles. Abbreviations: E. hellem, Encephalitozoon hellem. For the remaining abbreviations see Table 2. + Number is not reported due to the difficulty in the reconstruction of dead parasites. Potentially zoonotic species shaded in grey.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



