
Submitted:
09 November 2023
Posted:
14 November 2023
You are already at the latest version
Abstract
Black Knot (BK) is a deadly disease of European (Prunus domestica) and Japanese (Prunus salicina) plums caused by the hemibiotrophic fungus Apiosporina morbosa. Generally, phytopathogens hamper the balance of primary defense phytohormones such as salicylic acid (SA)-jasmonic acid (JA) balance for disease progression. Thus, we quantified the important phytohormones titers in tissues of susceptible and resistant genotypes belonging to European and Japanese plums at five different time points. The results suggested auxin-cytokinins interplay driven by A. morbosa appeared to be vital in disease progression by hampering the plant defense system via perturbance in SA, JA, and gibberellic acid. The results further point out that SA and JA in plant defense are not necessarily antagonistic always, especially in woody perennials. Results also suggest that the changes in SA and JA content following BK disease infection and progression in plums could be utilized as phytohormonal markers in the identification of BK-resistant cultivars.
Keywords:
Apiosporina morbosa
; black knot
; phytohormones
; plant growth regulators
; plant immunity
; plant-pathogen interactions
1. Introduction
Plant-invading pathogens based on nutrient acquisition are classified as necrotrophs, biotrophs, and hemibiotrophs [1]. Hemibiotrophs show a biotrophic phase of lifestyle in the beginning and later enter the necrotrophic phase of lifestyle [2]. Plant defense responses often adapt to the lifestyle of infecting pathogens, with salicylic acid (SA) dependent defense system acting against biotrophs and hemibiotrophs while jasmonic acid (JA) and ethylene (ET) dependent defense system working against necrotrophs [3,4]. Further, increased SA enhances resistance while increased JA could make the plant susceptible to biotrophs and hemibiotrophs [5,6]. The complex defense network of plants is further manipulated by other hormones including auxins, gibberellic acid (GA), cytokinins, ET, and abscisic acid (ABA) [7]. Pathogens also use different strategies to manipulate the plant defense system for entering and colonizing the plant, thus establishing the disease [8]. A common strategy used by pathogens is the manipulation of hormone signaling to promote disease [9]. Phytopathogens exploit plant defense network by either directly or indirectly producing, and/or manipulating signaling pathway(s) of plant defense system-related hormones. Many plant pathogens produce hormones that contribute to virulence and thus hormones are often considered virulence factors [10]
Both the European (Prunus domestica) and Japanese (Prunus salicina) plums are affected by the fungus Apiosporina morbosa, which causes a tumor-like disease known as ‘Black knot’ (BK) [11]. Agrios [2] considered A. morbosa as hemibiotrophic fungi because of its biotrophic behavior at the early stage of infection and then conversion into necrotrophic phage at the later stage. Warty, black knots that can range in size from half an inch to over a foot in length are a sign of the disease [11]. Once established, the disease advances and gets worse. Old knots occasionally develop a powdery pink or white saprophytic fungal growth, and they are frequently attacked by various insects, which causes additional harm to the trees. Multiple infections cause trees to lose vigor, blossom poorly, become less productive overall, and become more vulnerable to additional harm, such as winter injury. [12]. BK causes significant financial losses in North America [13,14]. Besides, BK has not received enough attention from the scientific community which could be because of the challenges in testing under controlled conditions, unlike other fungal diseases.
The knot-forming nature of the BK disease points out the possible involvement of phytohormones like auxins and/or cytokinins in symptom development. We have previously reported that hormones such as auxin and cytokinins seem to be highly responsible for enhancing the plum’s susceptibility to BK [15]. Here we show that SA and JA act differently during BK disease progression than previously reported. Understanding the changes in SA and JA content after BK disease infection and progression in plums might help in the identification of BK-resistant cultivars using phytohormones as markers.
2. Results
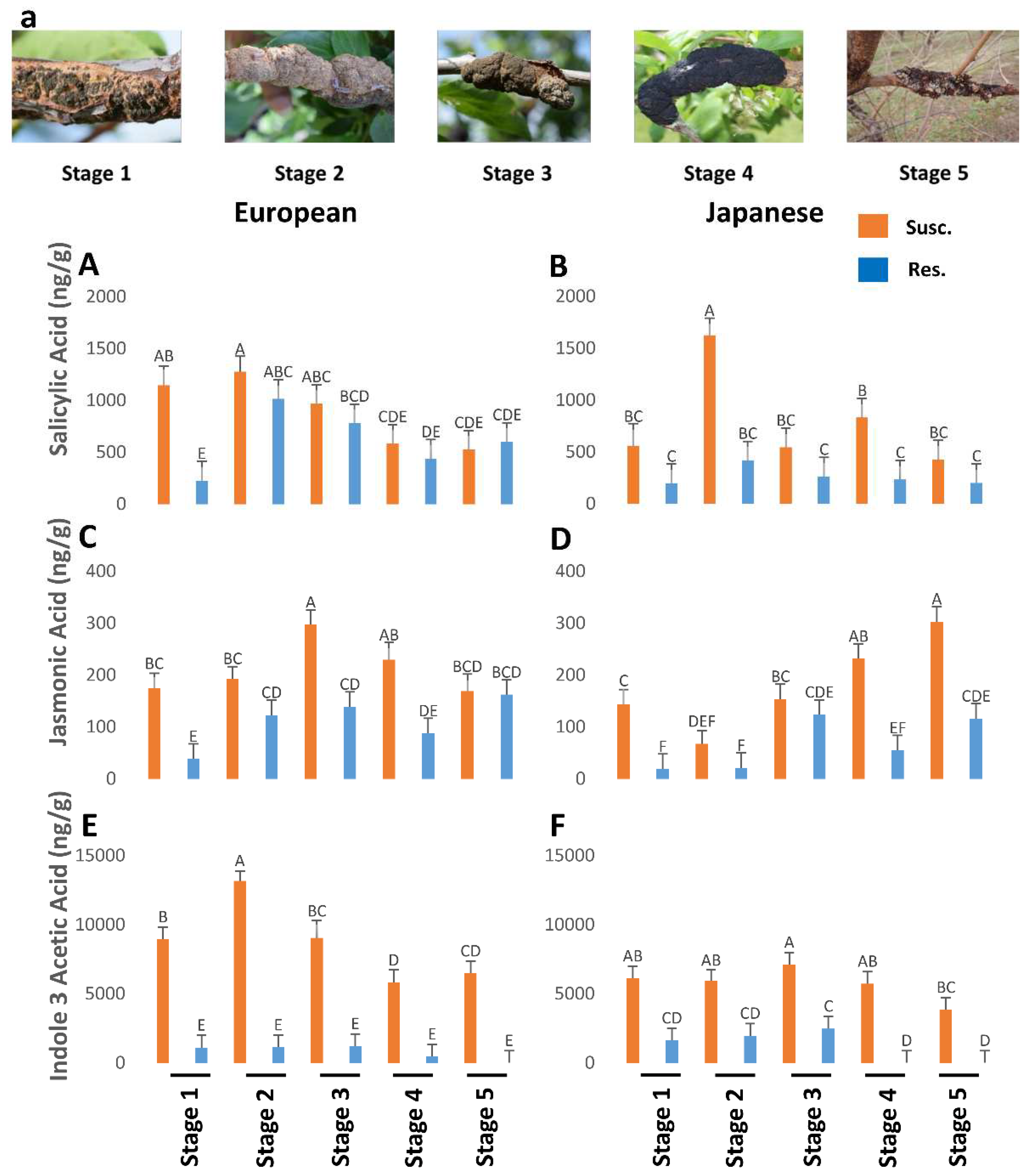
Susceptible genotypes had generally higher levels of both SA and JA throughout the black knot progression. The levels of SA were generally higher in susceptible genotypes of European and Japanese plums as compared to resistant ones throughout the five BK developmental stages, but statistically insignificant (Figure 1A,B). Jasmonic acid was significantly higher in susceptible genotypes than resistant ones at all the stages in both the plums except for the last stage in the Japanese plum, where it was not statistically significant (Figure 1C,D). Overall, the European plum possessed higher (approximately 1.3-2 fold) SA (Figure 1A,B) and JA (Figure 1C,D) titers compared to the Japanese plum. It is interesting to note that both JA and SA are high following BK infection while the conventional observations always pit them as antagonistic hormones. The concomitant rise in JA and SA triggered a deeper look at other hormones.
Salicylic Acid
SA levels were quite high at the onset of BK (stage 1) and peaked in stage 2, in the susceptible genotypes of both plums. Thereafter it gradually declined, barring an unexplained increase in stage 4 of Japanese plums. Interestingly even the resistant genotypes followed the same trend, although the SA levels in resistant genotypes were always lower than in the susceptible genotypes. Only in stage 5 of European plums, SA content between susceptible and resistant genotypes was very similar (Figure 1A,B). Overall, the European plum had significantly higher (1.42 times) amounts of SA than the Japanese plum (F1,144=7.66; P=0.0064), and susceptible genotypes contained significantly higher (1.93-fold) amounts of SA than resistant genotypes (F1,144=25.33; P=0.0001).
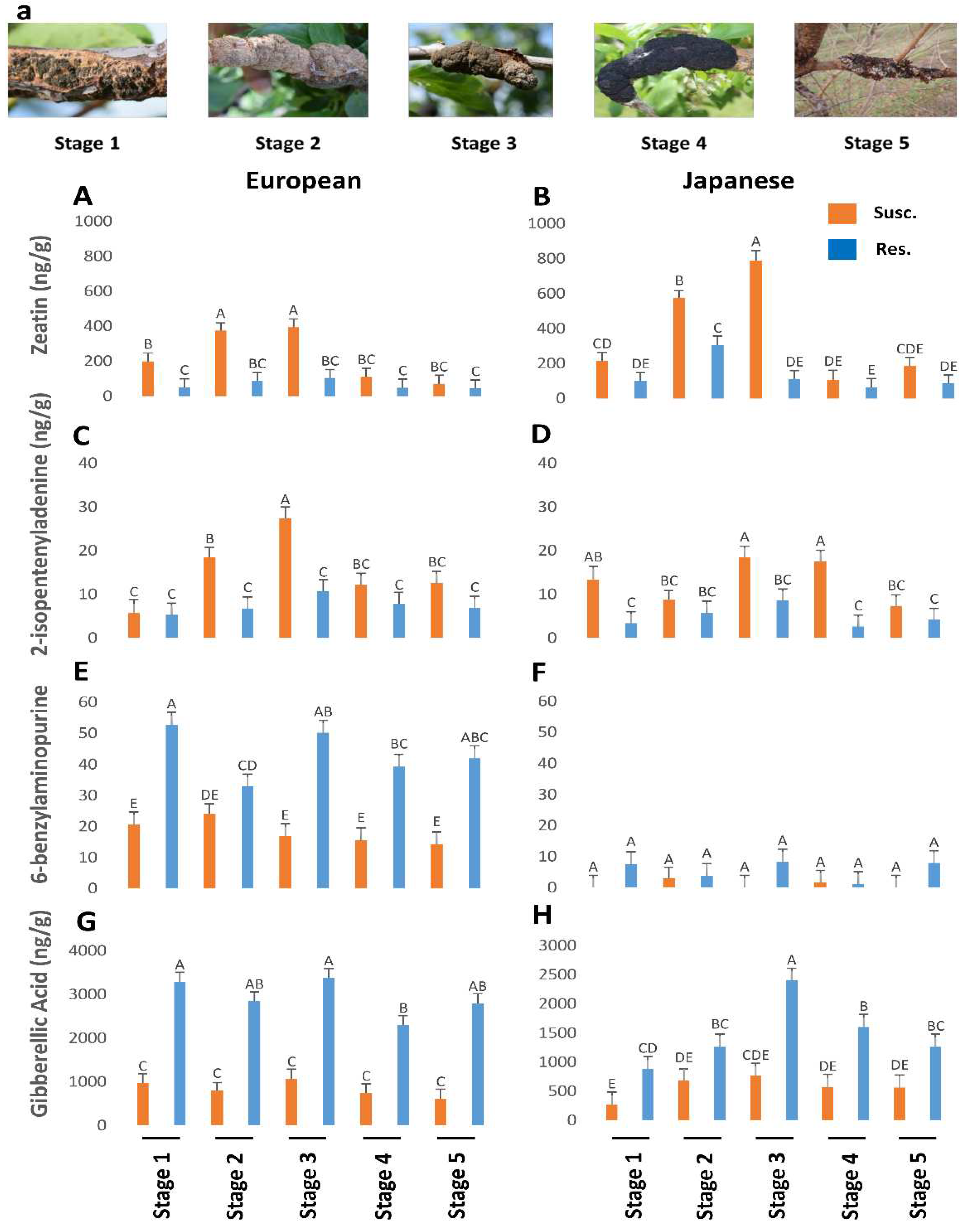
Figure 2.
The titers of zeatin (P<0.05), 2-isopentenyladenine (P<0.05), 6-benzylaminopurine (P=0.47), gibberellic acid (P=0.19) in resistant and susceptible genotypes of European and Japanese plums to Black Knot (BK) at five different BK developmental stages (1-5) (Figure 2a). Stage 1 is the appearance of visual symptoms of BK while stage 5 is the most developed knot. Different letters denote statistical significance and error bars represent means ± SEM (ng/g DW) for all the phytohormones.
Figure 2.
The titers of zeatin (P<0.05), 2-isopentenyladenine (P<0.05), 6-benzylaminopurine (P=0.47), gibberellic acid (P=0.19) in resistant and susceptible genotypes of European and Japanese plums to Black Knot (BK) at five different BK developmental stages (1-5) (Figure 2a). Stage 1 is the appearance of visual symptoms of BK while stage 5 is the most developed knot. Different letters denote statistical significance and error bars represent means ± SEM (ng/g DW) for all the phytohormones.
Jasmonic Acid
The European plum contained significantly higher (1.3 times) amounts of JA than the Japanese plum (F1,142=8.71; P=0.0037). When the data from European and Japanese plums were analyzed together, the susceptible genotypes had significantly higher (2.21 times) amounts of JA than the resistant ones (F1,142=70.33; P=0.0001). Separately, European and Japanese susceptible genotypes had 1.93 times and 2.68 times higher amounts of JA than resistant ones, respectively but the differences were statistically insignificant. Both in European and Japanese plums, at all BK developmental stages, JA was higher in susceptible genotypes than resistant genotypes, but the difference was not statistically significant at 2nd and 5th stages in the European plum and 2nd and 3rd stages in Japanese plum (Figure 1C,D). In susceptible European plums, JA levels peaked at stage 3 and then dropped till stage 5 while in susceptible Japanese plums, after a slight dip in stage 2 the JA levels increased exponentially till stage 5 (Figure 1C,D).
Principal Component Analysis
In PCA of susceptible genotypes of European plum, approximately 61-67% variability was explained by PCA I and approximately 20-26% by PCA II (Figure 3). However, in the resistant genotypes, approximately 49-67% variability was explained by PCAI and 30% by PCA II. In PCA of susceptible genotypes of Japanese plum, approximately 44-84% variability was explained by PCA I and approximately 10-39% by PCA II. However, in the resistant genotypes, approximately 70% of the variability was explained by PCAI and 25% by PCA II. At stage 1, in European susceptible genotypes GA and BA were positively correlated, while other compounds were negatively correlated with GA and BA. However, zeatin was unrelated to any of the groups. It is noteworthy the strong correlation between IAA and JA at stage 1 (>0.8). At stage 5, JA, SA, ip, and zeatin appeared to be highly correlated (>0.9). Zeatin and ip were moderately negatively correlated with GA (>0.8). Interestingly, this was not the case in resistant genotypes of the European plum. Here, at stages 1 and 5, all the compounds were positively correlated. Intriguingly, at stage 1 and stage 5 of resistant European genotypes, zeatin and ip, and zeatin and SA were unrelated to other compounds, respectively. At stage 1, zeatin and ip, IAA and JA, and GA, BA, and SA were highly correlated (Figure 3). And, these results are in agreement with the phytohormonal responses at specific stages (Fig 1, 2).
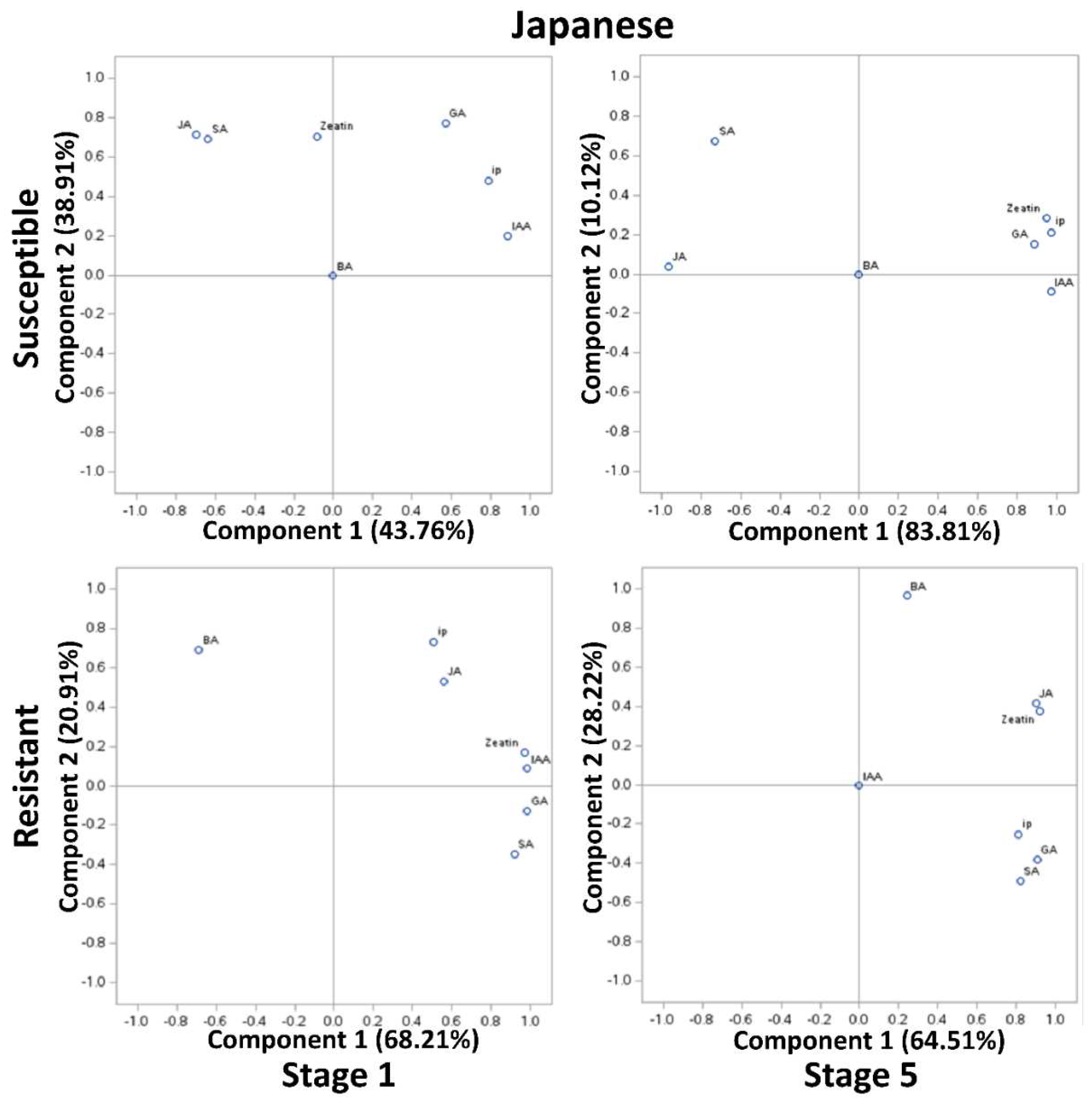
In Japanese susceptible plum genotypes, at stages 1 and 5, JA and SA (>0.7), and zeatin, ip, GA, and IAA were clustered (>0.8) but negatively correlated with each other (Figure 3). In the resistant Japanese genotypes, all compounds except BA were highly positively correlated at stage 1 and 5 (>0.7) (Figure 3). Surprisingly, at stage 5, except IAA, all compounds were positively correlated whereas BA was exceptionally poorly correlated to other compounds (<0.3) (Figure 3). It is noteworthy of undetected and unrelated IAA in both the resistant genotypes at stage 5 (Figure 3).
Overall, our results suggest that reduced GA and increased auxin, cytokinins, SA and JA appeared during BK disease progression.
Figure 3.
Principal component analysis (PCA) of phytohormonal contents of the Black Knot (BK) resistant and susceptible genotypes of European plums at the extreme ends of BK progression i.e. 1st (beginning of the infection) and the 5th (highly developed knot) stages. Phytohormones tryptophan (TRP), N-acetylserotonin (NAS), serotonin, indole-3-acetic acid (IAA), zeatin, 2-isopentenyladenine (IP), 6-benzylaminopurine (BA), salicylic acid (SA), jasmonic acid (JA) and gibberellic acid (GA) are labeled at data points. The distance of the data point from the center is directly proportional to the variability explained by PCA I and II, and the angle of a line passing through the center connecting two data points is inversely proportional to the correlation between phytohormones.
Figure 3.
Principal component analysis (PCA) of phytohormonal contents of the Black Knot (BK) resistant and susceptible genotypes of European plums at the extreme ends of BK progression i.e. 1st (beginning of the infection) and the 5th (highly developed knot) stages. Phytohormones tryptophan (TRP), N-acetylserotonin (NAS), serotonin, indole-3-acetic acid (IAA), zeatin, 2-isopentenyladenine (IP), 6-benzylaminopurine (BA), salicylic acid (SA), jasmonic acid (JA) and gibberellic acid (GA) are labeled at data points. The distance of the data point from the center is directly proportional to the variability explained by PCA I and II, and the angle of a line passing through the center connecting two data points is inversely proportional to the correlation between phytohormones.
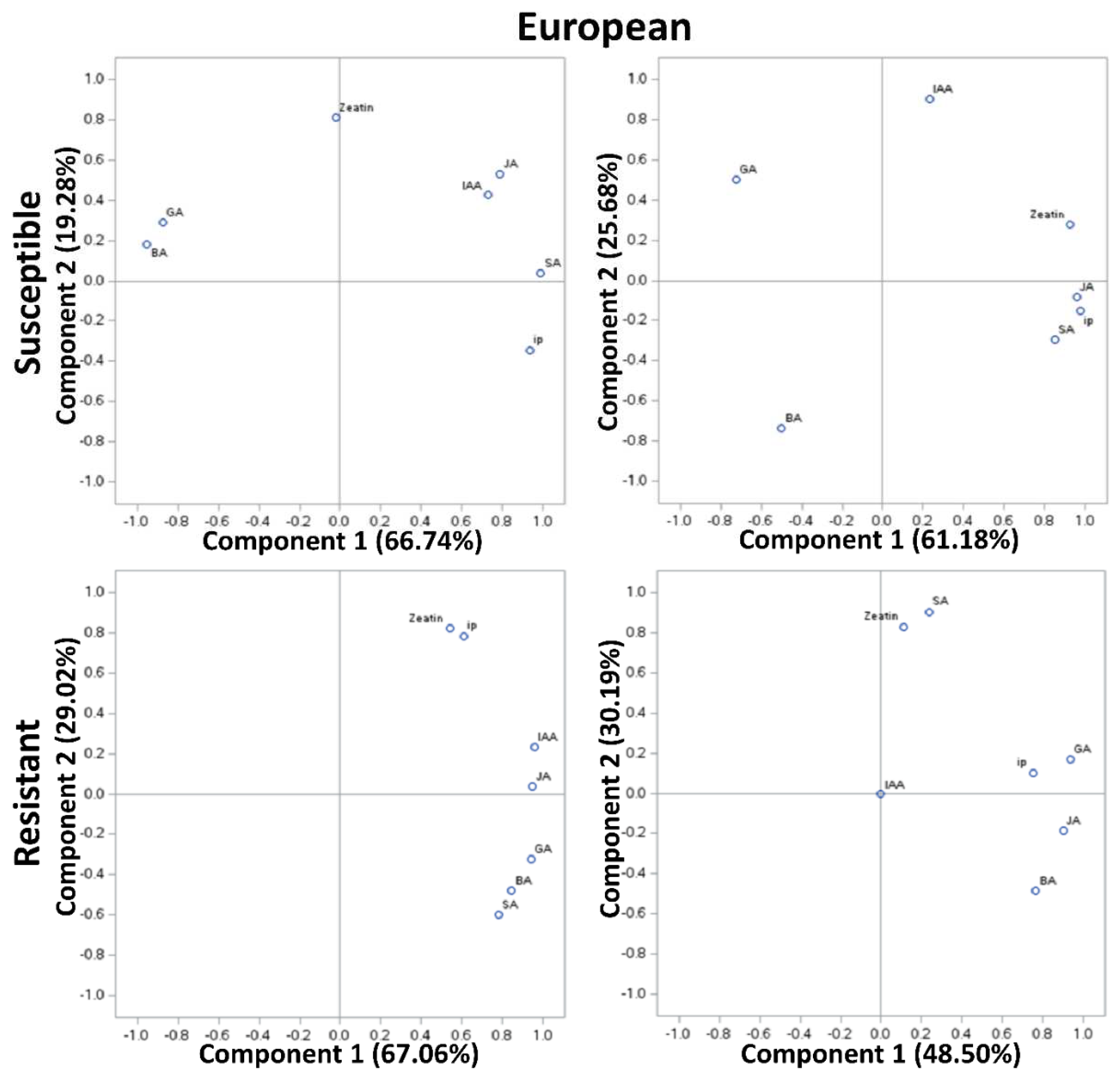
Figure 4.
Principal component analysis (PCA) of phytohormonal contents of the Black Knot (BK) resistant and susceptible genotypes of Japanese plums at the extreme ends of BK progression i.e. 1st (beginning of the infection) and the 5th (highly developed knot) stages. Phytohormones tryptophan (TRP), N-acetylserotonin (NAS), serotonin, indole-3-acetic acid (IAA), zeatin, 2-isopentenyladenine (IP), 6-benzylaminopurine (BA), salicylic acid (SA), jasmonic acid (JA) and gibberellic acid (GA) are labeled at data points. The distance of the data point from the center is directly proportional to the variability explained by PCA I and II, and the angle of a line passing through the center connecting two data points is inversely proportional to the correlation between phytohormones.
Figure 4.
Principal component analysis (PCA) of phytohormonal contents of the Black Knot (BK) resistant and susceptible genotypes of Japanese plums at the extreme ends of BK progression i.e. 1st (beginning of the infection) and the 5th (highly developed knot) stages. Phytohormones tryptophan (TRP), N-acetylserotonin (NAS), serotonin, indole-3-acetic acid (IAA), zeatin, 2-isopentenyladenine (IP), 6-benzylaminopurine (BA), salicylic acid (SA), jasmonic acid (JA) and gibberellic acid (GA) are labeled at data points. The distance of the data point from the center is directly proportional to the variability explained by PCA I and II, and the angle of a line passing through the center connecting two data points is inversely proportional to the correlation between phytohormones.
3. Discussion
BK disease advances very slowly; the full development of the knots takes almost two years. Consistently controlled infection is a failure due to its incredibly slow nature and environment-dependent infection process. So, before a phenotypic determination of resistant genotypes can be made, one must rely on the natural infection of susceptible genotypes in a genetically heterogeneous population over an extensive period of at least 7-8 years. This is possibly the most significant factor in the disease's understudied status. We created a scoring system for disease progression based on years of observations, showing 5 distinct stages of BK development in plums (Figure 1a) [17]. For the current study samples from different stages were collected as described previously, and concomitant branches from resistant genotypes were analyzed for changes in SA and JA titers, to see if SA and/or JA can be a marker to identify BK resistance in plums and to check the possibility of using SA and/or JA in priming of plum seedling/trees to control the BK disease in an environment-friendly way.
The discussion of IAA, zeatin, ip, BA, and GA titers is published elsewhere [15]. The higher titers of SA and JA in the European plum as compared to the Japanese plum might be related to the hexaploidy nature of the European plum. Due to the hexaploid nature, the European plum might have multiple copies of specific genes involved in SA and JA synthesis which might lead to the overexpression of those genes resulting in higher titers of a specific compound in the European plum than in the Japanese. Also, in a study conducted on Populus spp., as compared to diploid Populus spp., 87 genes were up-regulated in allotriploid Populus spp. Further, in allotetraploid Populus spp., 259 upregulated genes were observed as compared to diploid [18]. Besides, we compared the most cultivated European and Japanese plums; the European plum is not a polyploid developed from the Japanese plum but an allohexaploid developed naturally by an interspecific cross between Prunus cerasifera and Prunus spinosa [19]. Thus, another possible reason behind the different titers of SA and JA in European and Japanese plums could lie in the phylogenic difference between these two species.
- Auxin vs SA-JA in BK Disease Progression
IAA plays a key role in BK disease development most probably through promoted JA, increased fungal virulence, and increased hypertrophy and hyperplasia leading to knot formation which serves as a shelter to the fungus. Boosted auxin signaling can increase disease symptoms, development of galls/knots and feeding sites, and/or suppress SA-mediated defense responses [20]. Enhanced JA levels while no significant difference in SA levels of susceptible genotypes of European and Japanese plums was observed as compared to corresponding resistant genotypes after BK infection. In biotrophic and hemibiotrophic pathogens-plant interaction, the mode of auxin action is the antagonistic behavior of auxin and SA signaling [21]. Further, recent studies showed that the virulence-promoting behavior of increased auxin involves the suppression of SA-mediated defenses [22,23]. Fusarium oxysporum needs auxin signaling and transport for colonizing the host plant effectively may be through SA-dependent defense suppression [24]. Recent evidence suggested that in plant defense, SA and auxin pathways act antagonistic while JA and auxin pathways share many similarities [25]. SA accumulation suppresses IAA and JA biosynthesis [26]. Arabidopsis NahG plants, which were unable to accumulate SA showed 25 folds higher accumulation of JA after Pseudomonas syringae pv tomato DC3000 infection indicating pathogen-induced SA accumulation is responsible for JA suppression [27]. Auxin signaling showed deciding importance in activating the JA pathway after Rice black-streaked dwarf virus infection in rice [28]. Infecting citrus flower petals by Colletotrichum acutatum enhanced the accumulation of IAA and JA, supporting the synergism between IAA and JA [29]. Vinutha et al. (2020) observed the synergetic effect between IAA and JA during virus infection on tomato plants and mentioned the strong interplay between biosynthesis pathways.
- 2.
- Cytokinins vs SA-JA in BK Disease Progression
SA has a negative regulatory effect on cytokinin signaling [31]. Thus, plants accumulating high levels of SA may have reduced cytokinin content and/or signaling. Unexpectedly, in susceptible genotypes of European and Japanese plums as compared to corresponding resistant genotypes, there was a trend of a lower amount of BA at all BK developmental stages except the 4th stage in Japanese plums. It suggests the dissimilarity between BA and other cytokinin titers showing the inability of BK fungus in BA synthesis. The trend of increased SA levels in susceptible genotypes of European and Japanese plums might be suppressing BA synthesis in infected plum tissues as an antagonism between SA and cytokinin synthesis is observed previously. But increased SA levels did not suppress the zeatin and ip as they could be synthesized by A. morbosa and not plum tissues.
Cytokinins enhance SA-mediated defense and expression of PR genes [32]. Some studies reported that high levels of cytokinins in plants are linked to resistance against viruses [33,34] and nematodes [35]. Exogenous application of cytokinins initiates SA-mediated defense response which explains the higher susceptibility of ahk mutants to Pseudomonas syringae pv. tomato and Hyaloperonospora arabidopsidis like foliar pathogens [31,36]. Besides, cytokinins-induced pathogen resistance requires the SA pathway as well as may need additional signaling mechanisms [37].
Thus, depending upon plant-pathogen interactions, cytokinins induce either SA-mediated defense or promote plant susceptibility. In contrast to cytokinin-mediated immunity, cytokinin-induced susceptibility is activated at sub-micromolar cytokinin levels [38,39]. The effect of cytokinins on plant immunity has been shown to work based on a dose-dependent manner in different pathosystems [31,40]. It was noted that lower concentrations of cytokinins help pathogen success [31]. Exogenous application of low concentrations of cytokinin BA (<1 µM) to Arabidopsis increased the establishment of oomycete Hpa on wild-types as compared to the mock treatment. Similarly, a moderate increase in cytokinins in wheat leaves increased the powdery mildew growth rather than resistance [41]. Based on the information available on the involvement of cytokinins in plant immunity, we propose that delayed tissue senescence due to low to moderate levels of cytokinins enhances biotrophic and hemibiotrophic diseases but resists necrotrophic diseases while high levels of cytokinins inhibit biotrophic and hemibiotrophic diseases but support necrotrophs because of the activation of SA-mediated defense.
In addition, the concentration of other hormones with cytokinins also should be considered in the defense activation of plants against pathogens [42,43] as all plant hormones work together in a complex system. It is the interaction between multiple hormones that regulate the defense response and not a single hormone [39]. High levels of cytokinins in A. morbosa susceptible genotypes of plum could not promote SA accumulation and thus SA-mediated defense. The possible ground might be that it is not only cytokinins but the complex action of several hormones as mentioned earlier. More specifically, higher levels of IAA and JA in susceptible genotypes of plum to A. morbosa might be suppressing SA synthesis as it is well known. It was noted that auxins work in an antagonistic way to cytokinins in the plant immunity system [44]. Another possible explanation for not induction of SA-mediated defense in susceptible genotypes of plum is that turning on SA-mediated defense may need higher levels of cytokinins than available levels.
- 3.
- GA vs SA-JA in BK Disease Progression
The trend of reduced levels of GA in susceptible genotypes of European and Japanese plums points out its suppression due to high levels of A. morbosa-induced cytokinins especially zeatin and ip [15]. Moreover, GA promotes the degradation of DELLA proteins [45]. In contrast, stabilized DELLA proteins enhance JA signaling while attenuating SA signaling [46]. Therefore, DELLA proteins promote resistance to necrotrophs and susceptibility of biotrophs and hemibiotrophs by suppressing SA and promoting JA signaling [47]. Thus, enhanced GA might promote resistance to biotrophs and hemibiotrophs, and susceptibility to necrotrophs through the degradation of DELLAs. Reduced GA levels in BK-infected tissues of susceptible genotypes of plum might promote stabilized DELLA proteins contributing to enhanced JA and suppressed SA signaling that might help in the establishment and development of the BK disease. In addition, the exogenous application of GA to Allium sativum plants increased resistance to the hemibiotrophic fungus Fusarium verticillioides [48] probably through the activation of SA-mediated defense. Necrotrophic fungus Gibberella fujikuroi causing the foolish-seedling disease of rice synthesizes GA to promote disease may be through the degradation of DELLAs to suppress JA-mediated resistance to necrotrophs [47] and to promote SA-mediated HR and thus cell death.
- 4.
- JA and SA in BK Disease Progression
Generally, SA is involved in resistance against biotrophs and hemibiotrophs while JA is involved in resistance against necrotrophs [49]. Often, these two hormones work antagonistically in response to a specific pathogen, with the induction of one leading to the suppression of the other [46]. However, the contradiction between JA and SA is not conserved in plants [50]. So far, several studies showed the synergetic or neutral relationship between SA and JA pathways in several plant species [51,52,53,54,55,56]. But, unfortunately, the synergistic action of these two hormones has not received enough attention from the scientific community. It was reported that the synergistic action of SA and JA in multiple genotypes of woody perennial Populus spp. after infection with the biotrophic rust fungus Melampsora larici-populina [56]. Exogenously applied SA and SA hyperaccumulating lines enhanced JA levels, and exogenously applied JA increased SA accumulation in Poplar [56]. The accumulation of SA was followed by JA after infection of hemibiotrophic fungus Ophiostoma novo-ulmi to woody perennial Ulmus americana tissues disclosing synergistic action in between SA and JA in woody trees [57]. In the same study, the exogenous application of SA enhanced the resistance of U. americana to O. novo-ulmi suggesting the role of SA in resistance to hemibiotrophic pathogens. In woody plants, SA and JA pathways are not necessarily antagonistic [56]. Furthermore, woody perennials can store a large reserve of carbon and thus might have evolved SA-JA mediated co-defense system, especially since such long-living plants are subjected to simultaneous attack by multiple insects, pathogens, and herbivores. In contrast, short-living plants have a limited reserve of carbon to defend themselves against multiple attackers thus their defense system is designed to turn on against a single attacker at a time [56]. In our study, we observed the synergism between JA and SA after the infection of European and Japanese plums with A. morbosa, a hemibiotrophic fungus. Besides, an increase in cytokinins like ip and zeatin might have contributed to the increase in SA as well, as referred earlier. But the increase in SA was insignificant, unlike the increase in JA. An insignificant increase in SA could not promote resistance and restrict A. morbosa infection in plums, though enhanced SA is well known to induce resistance against hemibiotrophs. The inability of SA to induce resistance to BK might be related to the insufficient difference in SA content before and after the pathogen attack as SA is already high in plums. It is crucial to understand that in plants like Arabidopsis and Tobacco high levels are SA are produced immediately after biotrophic and hemibiotrophic pathogen attack. But, similarly to plums, in plants like rice, the SA levels are already high and did not increase significantly after the pathogen attack making the plant susceptible to hemibiotroph like Magnaporthe oryzae [58]. Thus, the noted resistance/susceptibility differences in different plants to different pathogens might be related to the differences in SA content and signaling [39].
Similar to the antagonism between SA and JA, the conflict between auxin and SA and the collegial between auxin and JA is well known [25]. As an example, auxin-dependent suppression of SA-mediated defense and activation of JA-mediated defense was observed after infection of Arabidopsis with a hemibiotrophic fungus Fusarium oxysporum [24]. A. morbosa-driven increase in IAA might be responsible for the hike in JA after the pathogen attack in plums. Also, as mentioned earlier, reduced GA through the stabilization of DELLAs could have contributed to the enhanced JA content. Taking together, significantly promoted JA might be responsible for the increased susceptibility of European and Japanese plums to A. morbosa. Additionally, in Cucumber Mosaic Virus infected Arabidopsis, it has been noted that auxin and SA systemically co-increased indicating antagonism between SA and auxin cannot be anticipated in all the cases [59]. Similarly, we observed the co-increase in IAA and SA supporting that antagonism between SA and auxin is not conserved.
Botrytis cinerea secretes a virulence factorβ-(1,3)(1,6)-D-glucan which activates the SA pathway that works antagonistically to the JA pathway, and thus, increases disease severity in Tomato. Therefore, B. cinerea manipulates the SA pathway to establish disease in Tomato [60]. In contrast, the Hemibiotrophic fungus Ustilago maydis secrets Cmu1 effectors to inhibit SA biosynthesis to suppress SA-mediated immunity [61]. In addition, Topless proteins are involved in the suppression of SA signaling [62]. A few of the Tips like Tip1 and Tip2 [63] and other effectors like Jsi1 and Nkd1 [62] induce strong SA-related defense responses indicating antagonism between Topless and effectors in SA-induced defense. Likewise, A. morbosa might be driving auxin to suppress SA-mediated plant resistance and promote JA-mediated susceptibility.
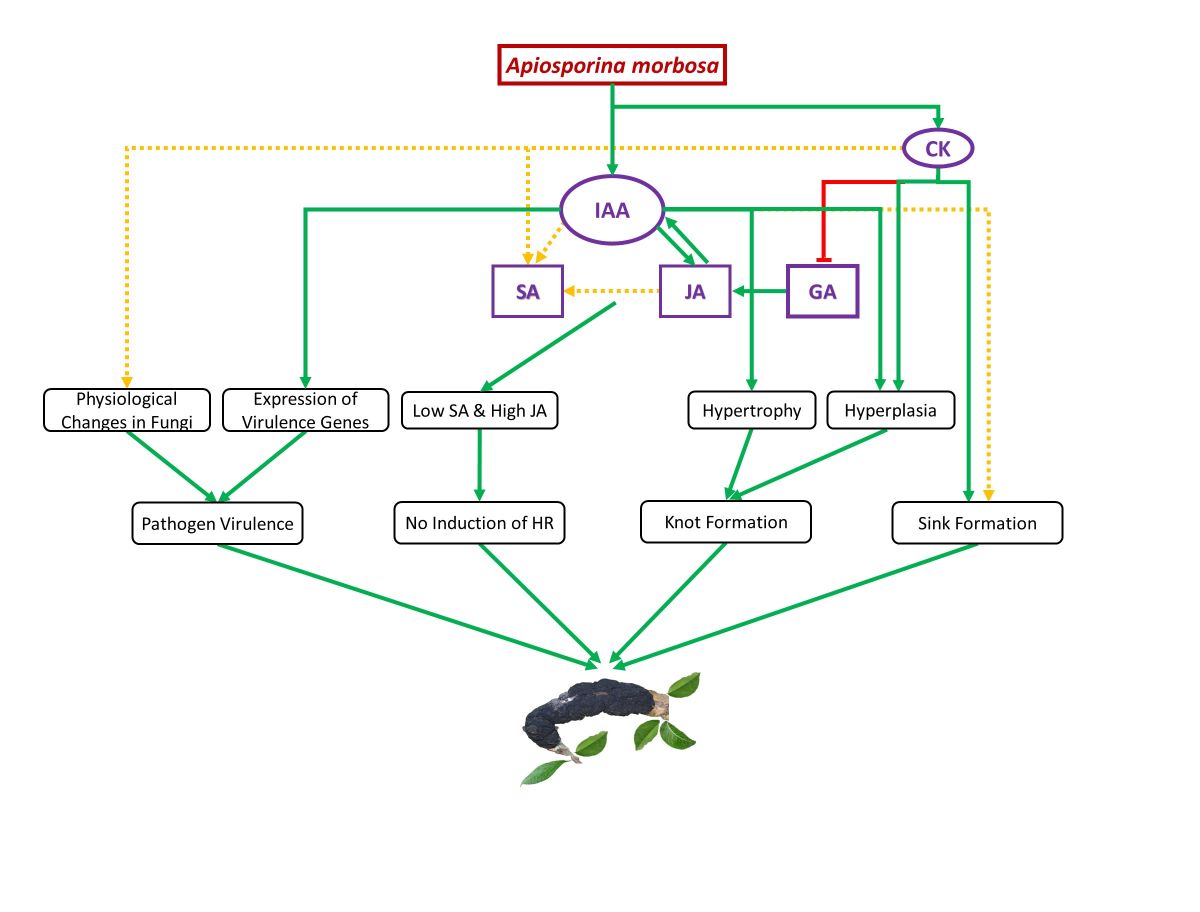
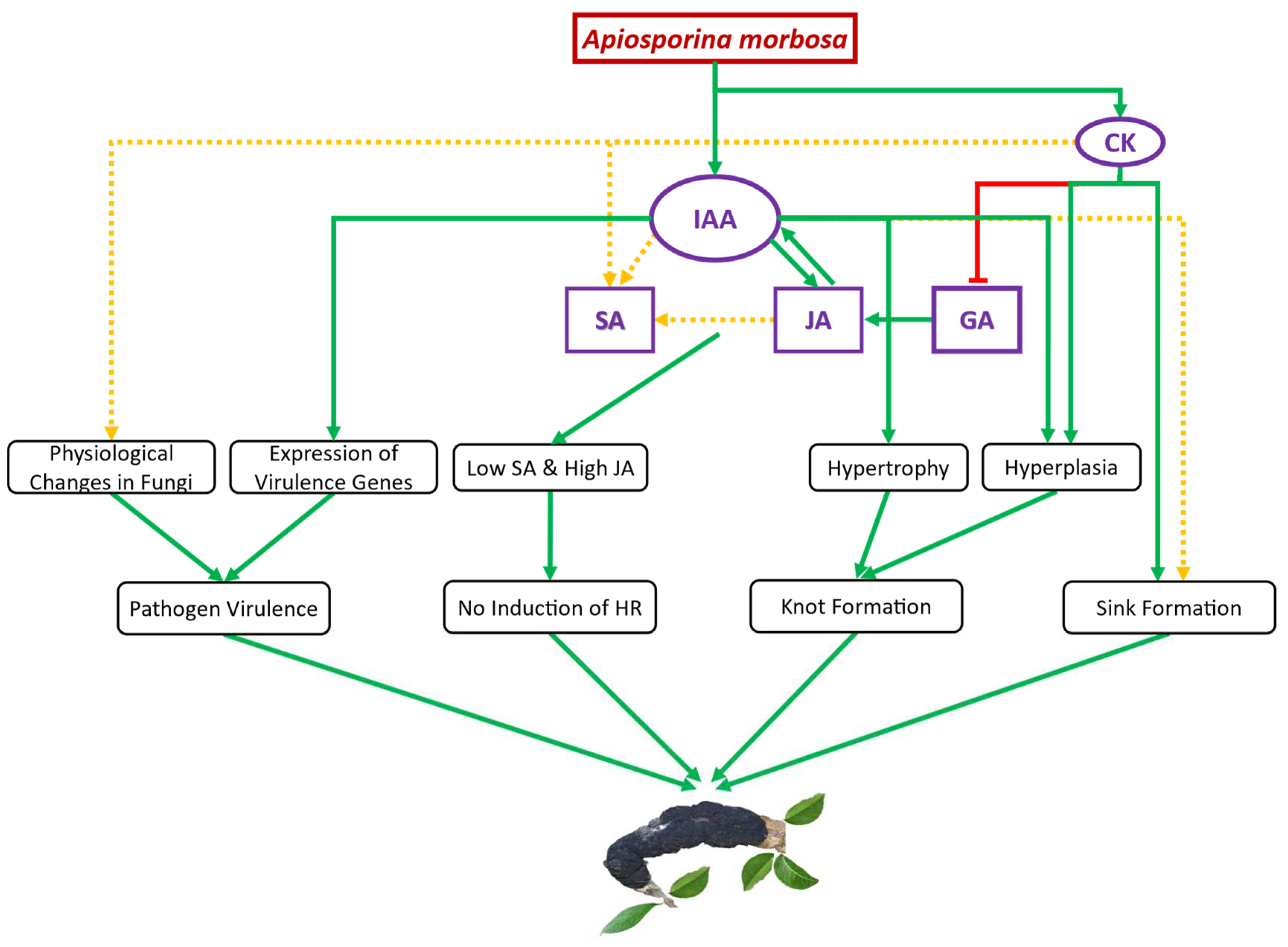
Figure 5.
Schematic representation of the proposed auxin and cytokinins (CK) mediated Black Knot (BK) establishment and development. Immediately following infection, Apiosporina morbosa-driven indole-3-acetic acid (IAA) promotes jasmonic acid (JA) synthesis and enhanced JA stimulates endogenous IAA synthesis in plum tissues. IAA together with JA seems to promote salicylic acid (SA) synthesis, while cytokinins may also be responsible for increased SA. Besides, auxin causes hypertrophy and hyperplasia of infected plum cells leading to knot formation. Cytokinin induced suppression of gibberellic acid (GA) because elevated cytokinins might help in promoting JA synthesis in infected plum cells. In the figure, the arrow and green color indicate a positive effect while the blunt end and red color indicate a negative effect. A solid green line denotes strong proof of disease progression, and a dotted yellow line suggests inconclusive evidence towards disease progression. .
Figure 5.
Schematic representation of the proposed auxin and cytokinins (CK) mediated Black Knot (BK) establishment and development. Immediately following infection, Apiosporina morbosa-driven indole-3-acetic acid (IAA) promotes jasmonic acid (JA) synthesis and enhanced JA stimulates endogenous IAA synthesis in plum tissues. IAA together with JA seems to promote salicylic acid (SA) synthesis, while cytokinins may also be responsible for increased SA. Besides, auxin causes hypertrophy and hyperplasia of infected plum cells leading to knot formation. Cytokinin induced suppression of gibberellic acid (GA) because elevated cytokinins might help in promoting JA synthesis in infected plum cells. In the figure, the arrow and green color indicate a positive effect while the blunt end and red color indicate a negative effect. A solid green line denotes strong proof of disease progression, and a dotted yellow line suggests inconclusive evidence towards disease progression. .
4. Materials and Methods
4.1. Sample Collection
From the University of Guelph’s plum breeding program, 15-year-old European (P. domestica) and Japanese (P. salicina) plum trees were selected for the current study. The phytohormonal analysis was performed using susceptible genotypes ‘Vision’ and ‘Veeblue’ from the European plum and ‘Vampire’ and ‘Shiro’ from the Japanese plum while resistant genotypes V982014 and V911415 from the European plum and ‘Underwood’ and ‘Redcoat’ from the Japanese plum to the BK disease. The resistance and susceptibility of these genotypes were established based on the assessment of BK incidence and progression over 10 years, with limited disease control in a diverse population consisting of ~150 genotypes of each Japanese and European plum (Shinde et al. Submitted). Samples from black knots at five different developmental stages (based on the stage of the infection, from the end of May till mid-October on approximately 3-week intervals) were collected from susceptible genotypes and stored at -80°C after flash freezing with liquid nitrogen (Figure 1a). In the case of resistant genotypes, at the same time points mentioned above, the branches of comparable age and size were used for the sample collection.
4.2. Freeze Drying and Grinding
Freeze drying of all the woody samples was done for 48 hours with FreeZone 4.5L -50°C Benchtop Freeze Dryer (Labconco Corporation, 8811 Prospect Avenue, Kansas City, Missouri, 64132, USA) immediately after the sample collection from the field. Then freeze-dried samples were stored at -80°C until they were ground using IKA® A 11 basic Analytical mill (IKA Works, Inc., 2635, Northchase Parkway SE, Wilmington, North Carolina, 28405, USA) with liquid nitrogen.
4.3. Hormone Extraction, Identification, and Quantification
SA and JA from ground woody samples were extracted using the methanol double extraction method. Briefly, 100 mg of freeze-dried, powdered woody samples were extracted with a solvent mixture (methanol: formic acid: milli-Q water = 15:1:4). Samples were then held at -20 oC for 30 min and spun down (15 min, 4 oC, 14000 rpm) and the supernatant removed. A second extraction was performed on the same sample using similar conditions described above and the supernatants were pooled. Solid phase extraction (Oasis® HLB 1cc (30 mg), Waters Canada, Mississauga, ON, Canada) was deployed to concentrate the samples before eluting them in 200 μL methanol. Later, the eluant was filtered through a 0.22 μM centrifuge filter (Millipore; 1 min, 13000 rpm). All standards were analytical grade and purchased from Sigma Aldrich, Canada. SA and JA were separated by reverse phase liquid chromatography (ultra-performance liquid chromatography system (UPLC); LC-40D XS, Shimadzu, Japan) by injection of a 5 μL aliquot of sample onto Shim-pack Scepter LC column (2.1x50mm, 1.9 μm; Mandel Scientific Company, Guelph, ON, Canada). Metabolites were separated with a gradient of solvents A (0.1% formic acid) and B (100% methanol) with initial conditions at 95% A (5% B) increased to 5% A (95% B) over 4 minutes using a curve of 0. The column temperature was 40°C and the flow rate was 0.2 mL/min. Metabolite peaks were identified by comparison to standards and quantified by a standard curve generated using a similar separation method and gradient conditions. SA and JA were detected using a single quadrupole mass spectrometer (LCMS 2020, Shimadzu, Japan) in single ion recording mode (SIR). JA and SA were detected in negative mode with a cone voltage of 10 for mass to charge (m/z) of 209 and 137, respectively. In both cases, the probe temperature was set to 250 °C with a gain of 5; the capillary voltage (negative) was set to 0.5 kV. The linear range for both compounds was 1.53 ng/mL-6.25 μg/mL.
4.4. Statistical Analysis
The study was performed using four biological replicates from each genotype and two genotypes from each group, thus eight technical replicates from each group. Hormonal data of European and Japanese plums were analyzed together using general linear mixed models (proc GLIMMIX) in SAS v9.4 (SAS Institute Inc., 111, Hampton Woods Ln, Raleigh, North Carolina, 27607, USA). Shapiro-Wilk normality tests and studentized residual plots were used to test error assumptions of variance analysis including random, homogenous, and normal distributions of error. Outliers were removed using Lund’s test. Means were calculated using the LSMEANS statement, and significant differences between the treatments were determined using a post-hoc LSD test α ≤ 0.05 and are mentioned in each figure. The methodology for indole 3 acetic acid (IAA), zeatin, 2-isopentenyladenine (ip), 6-benzylaminopurine (BA), and gibberellic acid-3 (GA) is described elsewhere [15]. The graphical presentation of these compounds was adapted from elsewhere [15]. Further, to underpin SA-JA interplay, at the 1st and 5th stages (extreme stages of BK infection), previously reported compounds along with SA and JA data were reanalyzed in Principal Component Analysis (PCA). Here, PCA was conducted separately on phytohormonal data from resistant, and susceptible genotypes of European (Fig.13A) and Japanese plums (Figure 3), using PROC PRINCOMP in SAS v9.4 (SAS Institute Inc., 111, Hampton Woods Ln, Raleigh, North Carolina, 27607, USA).
5. Conclusions
BK disease establishment and development in European and Japanese plums seem to be achieved by hemibiotrophic fungus A. morbosa-driven auxin (IAA) and cytokinins (zeatin and ip) through unbalancing other phytohormones like GA, JA, and SA to suppress the plant defense system, changing plant physiology such as induction of knots and formation of the nutrient sink at the point of infection, and enhancing fungal virulence (Figure 4). SA, JA, and IAA content increased at the point of infection synergistically after BK fungal infection in multiple genotypes of both of the commercially cultivated plum species indicating antagonism between SA and JA, and SA and IAA cannot be anticipated in all plant-pathogen interactions especially involving woody perennials. One or more than one out of JA, IAA, zeatin, and ip might be responsible for the enhanced accumulation of SA in A. morbosa-infected plum tissues but further research is needed to point out the exact reason and phenomenon. An insignificant increase in SA content after A. morbosa infection in susceptible plum trees could not elicit HR suggesting an ample increase in the amount of SA is needed for HR induction. As the role of SA and JA in disease resistance is well understood, priming seeds or seedlings or trees with SA or JA or both depending on pathogen lifestyle (biotrophic, hemibiotrophic, and necrotrophic) could come up as an environmentally friendly approach to combat different diseases. Besides, SA levels in the plums could be used as a phytohormonal marker to select cultivars in breeding for BK disease resistance.
Author Contributions
All authors contributed essentially to the project. RS executed the experiments and collected and analyzed the samples for biochemicals. MA developed a protocol for biochemical analysis and performed the statistical analysis. WEK, MS, and PS participated in the conception and design of the study. JS conceived the project and supervised the work. RS prepared the manuscript and all authors read and approved the final manuscript.
Funding
This study was funded by the Ontario Ministry of Agriculture, Food and Rural Affairs and by Ontario Tender Fruit Marketing Board through a grant to JS. The support of the Niagara Peninsula Fruit and Vegetable Growers Association is also acknowledged.
Data Availability Statement
Data supporting the findings of this study are available within the paper. Further information can be obtained from the corresponding author.
Acknowledgments
We thank Glen Alm for his input in the research. We are also thankful to the number of summer students for their help in the project.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Mengiste, T. Plant Immunity to Necrotrophs. Annual Review of Phytopathology, 2012, 50. [CrossRef]
- Agrios, G.N. Plant Pathology; Elsevier, 2005.
- Glazebrook, J. Contrasting Mechanisms of Defense against Biotrophic and Necrotrophic Pathogens. Annual Review of Phytopathology, 2005, 43. [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature, 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Ghozlan, M.H.; EL-Argawy, E.; Tokgöz, S.; Lakshman, D.K.; Mitra, A. Plant Defense against Necrotrophic Pathogens. Am J Plant Sci, 2020, 11. [CrossRef]
- Katagiri, F.; Tsuda, K. Understanding the Plant Immune System. Molecular Plant-Microbe Interactions, 2010, 23. [CrossRef]
- Ma, K.W.; Ma, W. Phytohormone Pathways as Targets of Pathogens to Facilitate Infection. Plant Mol Biol, 2016, 91, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Dou, D.; Zhou, J.M. Phytopathogen Effectors Subverting Host Immunity: Different Foes, Similar Battleground. Cell Host and Microbe, 2012, 12. [CrossRef]
- Kunkel, B.N.; Johnson, J.M.B. Auxin Plays Multiple Roles during Plant-Pathogen Interactions. Cold Spring Harbor perspectives in biology, 2021, 13. [CrossRef]
- Laird, T.S.; Flores, N.; Leveau, J.H.J. Bacterial Catabolism of Indole-3-Acetic Acid. [CrossRef]
- Kayal, W. el; Chamas, Z.; El-Sharkawy, I.; Subramanian, J. Comparative Anatomical Responses of Tolerant and Susceptible European Plum Varieties to Black Knot Disease. Plant Dis, 2021, 105. [CrossRef]
- Wilcox, W.F. Black Knot of Plums. 1992.
- Stewart, S.A.; Weber, D.J. Environmental Site Characteristics and Incidence of Chokecherry Black Knot in Utah. Great Basin Naturalist, 1984, 44, 6. [Google Scholar]
- Scorza, R.; Demuth, M. Black Knot [Apiosporina Morbosa (Schw.)] Resistance in Imported and Domestic Prunus domestica L. Germplasm and Cultivars. Journal of the American Pomological Society. 2015, 69, 45–50. [Google Scholar]
- Shinde, R.; Ayyanath, M.-M.; Shukla, M.; El Kayal, W.; Saxena, P.; Subramanian, J. Hormonal Interplay Leading to Black Knot Disease Establishment and Progression in Plums. Plants, 2023, 12. [CrossRef]
- Shinde, R.; Shum, C.; Gill, R. ; Subramanian Jayasankar. Phenotyping Black Knot Resistance in Plum Germplasm.; Guelph, 2023.
- Kayal, W. el; Chamas, Z.; El-Sharkawy, I.; Subramanian, J. Comparative Anatomical Responses of Tolerant and Susceptible European Plum Varieties to Black Knot Disease. Plant Dis, 2021, 105. [CrossRef]
- Liqin, G.; Jianguo, Z.; Xiaoxia, L.; Guodong, R. Polyploidy-Related Differential Gene Expression between Diploid and Synthesized Allotriploid and Allotetraploid Hybrids of Populus. Molecular Breeding, 2019, 39. [CrossRef]
- Zhebentyayeva, T.; Shankar, V.; Scorza, R.; Callahan, A.; Ravelonandro, M.; Castro, S.; DeJong, T.; Saski, C.A.; Dardick, C. Genetic Characterization of Worldwide Prunus domestica (Plum) Germplasm Using Sequence-Based Genotyping. Hortic Res, 2019, 6. [CrossRef]
- Cui, F.; Wu, S.; Sun, W.; Coaker, G.; Kunkel, B.; He, P.; Shan, L. The Pseudomonas syringae Type III Effector AvrRpt2 Promotes Pathogen Virulence via Stimulating Arabidopsis Auxin/Indole Acetic Acid Protein Turnover. Plant Physiol, 2013, 162, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pajerowska-Mukhtar, K.; Culler, A.H.; Dong, X. Salicylic Acid Inhibits Pathogen Growth in Plants through Repression of the Auxin Signaling Pathway. Current Biology, 2007, 17, 1784–1790. [Google Scholar] [CrossRef] [PubMed]
- McClerklin, S.A.; Goo Lee, S.; Harper, C.P.; Nwumeh, R.; Jez, J.M.; Kunkel, B.N. Indole-3-Acetaldehyde Dehydrogenase-Dependent Auxin Synthesis Contributes to Virulence of Pseudomonas Syringae Strain DC3000. 2018. [CrossRef]
- Djami-Tchatchou, A.T.; Harrison, G.A.; Harper, C.P.; Wang, R.; Prigge, M.J.; Estelle, M.; Kunkel, B.N. Dual Role of Auxin in Regulating Plant Defense and Bacterial Virulence Gene Expression During Pseudomonas syringae PtoDC3000 Pathogenesis. 2020, 33. [CrossRef]
- Kidd, B.N.; Kadoo, N.Y.; Dombrecht, B.; Tekeoğlu, M.; Gardiner, D.M.; Thatcher, L.F.; Aitken, E.A.B.; Schenk, P.M.; Manners, J.M.; Kazan, K. Auxin Signaling and Transport Promote Susceptibility to the Root-Infecting Fungal Pathogen Fusarium oxysporum in Arabidopsis. / 733 MPMI, 2011, 24, 733–748. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Manners, J.M. Linking Development to Defense: Auxin in Plant-Pathogen Interactions. Trends in Plant Science, 2009, 14. [CrossRef]
- Yuan, H.M.; Liu, W.C.; Lu, Y.T. CATALASE2 Coordinates SA-Mediated Repression of Both Auxin Accumulation and JA Biosynthesis in Plant Defenses. Cell Host Microbe, 2017, 21. [CrossRef]
- Spoel, S.H.; Koornneef, A.; Claessens, S.M.C.; Korzelius, J.P.; van Pelt, J.A.; Mueller, M.J.; Buchala, A.J.; Métraux, J.P.; Brown, R.; Kazan, K.; van Loon, L.C.; Dong, X.; Pieterse, C.M.J. NPR1 Modulates Cross-Talk between Salicylate- and Jasmonate-Dependent Defense Pathways through a Novel Function in the Cytosol. Plant Cell, 2003, 15. [CrossRef]
- Zhang, H.; Tan, X.; Li, L.; He, Y.; Hong, G.; Li, J.; Lin, L.; Cheng, Y.; Yan, F.; Chen, J.; Sun, Z. Suppression of Auxin Signalling Promotes Rice Susceptibility to Rice Black Streaked Dwarf Virus Infection. Mol Plant Pathol, 2019, 20. [CrossRef]
- Lahey, K.A.; Yuan, R.; Burns, J.K.; Ueng, P.P.; Timmer, L.W.; Chung, K.R. Induction of Phytohormones and Differential Gene Expression in Citrus Flowers Infected by the Fungus Colletotrichum acutatum. Molecular Plant-Microbe Interactions, 2004, 17. [CrossRef]
- Vinutha, T.; Vanchinathan, S.; Bansal, N.; Kumar, G.; Permar, V.; Watts, A.; Ramesh, S. v.; Praveen, S. Tomato Auxin Biosynthesis/Signaling Is Reprogrammed by the Geminivirus to Enhance Its Pathogenicity. Planta, 2020, 252. [CrossRef]
- Argueso, C.T.; Ferreira, F.J.; Epple, P.; To, J.P.C.; Hutchison, C.E.; Schaller, G.E.; Dangl, J.L.; Kieber, J.J. Two-Component Elements Mediate Interactions between Cytokinin and Salicylic Acid in Plant Immunity. PLoS Genet, 2012, 8. [CrossRef]
- Reusche, M.; Klásková, J.; Thole, K.; Truskina, J.; Novák, O.; Janz, D.; Strnad, M.; Spíchal, L.; Lipka, V.; Teichmann, T. Stabilization of Cytokinin Levels Enhances Arabidopsis Resistance against Verticillium longisporum. Molecular Plant-Microbe Interactions, 2013, 26. [CrossRef]
- Pogány, M.; Koehl, J.; Heiser, I.; Elstner, E.F.; Barna, B. Juvenility of Tobacco Induced by Cytokinin Gene Introduction Decreases Susceptibility to Tobacco Necrosis Virus and Confers Tolerance to Oxidative Stress. Physiol Mol Plant Pathol, 2004, 65. [CrossRef]
- Darke, S.F.; Burritt, D.J.; Jameson, P.E.; Guy, P.L. Effects of Plant Hormones on White Clover Mosaic Potexvirus Double-Stranded RNA. Plant Pathol, 2000, 49. [CrossRef]
- Shanks, C.M.; Rice, J.H.; Zubo, Y.; Schaller, G.E.; Hewezi, T.; Kieber, J.J. The Role of Cytokinin during Infection of Arabidopsis Thaliana by the Cyst Nematode Heterodera schachtii. Molecular Plant-Microbe Interactions, 2016, 29. [CrossRef]
- Choi, J.; Huh, S.U.; Kojima, M.; Sakakibara, H.; Paek, K.H.; Hwang, I. The Cytokinin-Activated Transcription Factor ARR2 Promotes Plant Immunity via TGA3/NPR1-Dependent Salicylic Acid Signaling in Arabidopsis. Dev Cell, 2010, 19. [CrossRef]
- Gupta, R.; Pizarro, L.; Leibman-Markus, M.; Marash, I.; Bar, M. Cytokinin Response Induces Immunity and Fungal Pathogen Resistance, and Modulates Trafficking of the PRR LeEIX2 in Tomato. Mol Plant Pathol, 2020, 21. [CrossRef]
- Spallek, T.; Gan, P.; Kadota, Y.; Shirasu, K. Same Tune, Different Song — Cytokinins as Virulence Factors in Plant–Pathogen Interactions? Current Opinion in Plant Biology, 2018, 44, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, T.; Argueso, C.T. Should I Fight or Should I Grow Now? The Role of Cytokinins in Plant Growth and Immunity and in the Growth-Defence Trade-Off. Annals of Botany, 2017, 119. [CrossRef]
- Großkinsky, D.K.; Naseem, M.; Abdelmohsen, U.R.; Plickert, N.; Engelke, T.; Griebel, T.; Zeier, J.; Novák, O.; Strnad, M.; Pfeifhofer, H.; van der Graaff, E.; Simon, U.; Roitsch, T. Cytokinins Mediate Resistance against Pseudomonas syringae in Tobacco through Increased Antimicrobial Phytoalexin Synthesis Independent of Salicylic Acid Signaling. Plant Physiol, 2011, 157. [CrossRef]
- Babosha, A. v. Regulation of Resistance and Susceptibility in Wheat–Powdery Mildew Pathosystem with Exogenous Cytokinins. J Plant Physiol, 2009, 166, 1892–1903. [Google Scholar] [CrossRef] [PubMed]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth-Defense Tradeoffs in Plants: A Balancing Act to Optimize Fitness. Molecular Plant, 2014, 7. [CrossRef]
- Belkhadir, Y.; Yang, L.; Hetzel, J.; Dangl, J.L.; Chory, J. The Growth-Defense Pivot: Crisis Management in Plants Mediated by LRR-RK Surface Receptors. Trends in Biochemical Sciences, 2014, 39. [CrossRef]
- Naseem, M.; Philippi, N.; Hussain, A.; Wangorsch, G.; Ahmed, N.; Dandekara, T. Integrated Systems View on Networking by Hormones in Arabidopsis Immunity Reveals Multiple Crosstalk for Cytokinin. Plant Cell, 2012, 24. [CrossRef]
- Sun, T.P.; Gubler, F. Molecular Mechanism of Gibberellin Signaling in Plants. Annual Review of Plant Biology, 2004, 55. [CrossRef]
- Grant, M.R.; Jones, J.D.G. Hormone (Dis)Harmony Moulds Plant Health and Disease. Science, 2009, 324. [CrossRef]
- Navarro, L.; Bari, R.; Achard, P.; Lisón, P.; Nemri, A.; Harberd, N.P.; Jones, J.D.G. DELLAs Control Plant Immune Responses by Modulating the Balance of Jasmonic Acid and Salicylic Acid Signaling. Current Biology, 2008, 18, 650–655. [Google Scholar] [CrossRef] [PubMed]
- Saloua, N.; Mourad, S.M.; Ammar, O. In Vitro and in Vivo Evaluating the Efficacy of Salicylic Acid and Gibberellic Acid Against Fusariumverticillioides in Garlic (Allium sativum L). World Journal of Environmental Biosciences, 2018, 7, 1–8. [Google Scholar]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; Dong, X. NPR3 and NPR4 Are Receptors for the Immune Signal Salicylic Acid in Plants. Nature, 2012, 486. [CrossRef]
- de Vleesschauwer, D.; Gheysen, G.; Höfte, M. Hormone Defense Networking in Rice: Tales from a Different World. Trends in Plant Science, 2013, 18. [CrossRef]
- Rostás, M.; Winter, T.R.; Borkowski, L.; Zeier, J. Copper and Herbivory Lead to Priming and Synergism in Phytohormones and Plant Volatiles in the Absence of Salicylate-Jasmonate Antagonism. Plant Signal Behav, 2013, 8. [CrossRef]
- Sun, N.; Kong, X.; Liu, Y.; Gong, T.; Gu, X.; Liu, L. The THO/TREX Complex Active in Alternative Splicing Mediates Plant Responses to Salicylic Acid and Jasmonic Acid. Int J Mol Sci, 2021, 22. [CrossRef]
- Betsuyaku, S.; Katou, S.; Takebayashi, Y.; Sakakibara, H.; Nomura, N.; Fukuda, H. Salicylic Acid and Jasmonic Acid Pathways Are Activated in Spatially Different Domains around the Infection Site during Effector-Triggered Immunity in Arabidopsis thaliana. Plant Cell Physiol, 2018, 59. [CrossRef]
- Lemarié, S.; Robert-Seilaniantz, A.; Lariagon, C.; Lemoine, J.; Marnet, N.; Jubault, M.; Manzanares-Dauleux, M.J.; Gravot, A. Both the Jasmonic Acid and the Salicylic Acid Pathways Contribute to Resistance to the Biotrophic Clubroot Agent Plasmodiophora brassicae in Arabidopsis. Plant Cell Physiol, 2015, 56. [CrossRef]
- Chen, Q.; Zhang, R.; Li, D.; Wang, F. Integrating Transcriptome and Coexpression Network Analyses to Characterize Salicylic Acid- and Jasmonic Acid-Related Genes in Tolerant Poplars Infected with Rust. Int J Mol Sci, 2021, 22, 5001. [Google Scholar] [CrossRef] [PubMed]
- Ullah, C.; Schmidt, A.; Reichelt, M.; Tsai, C.J.; Gershenzon, J. Lack of Antagonism between Salicylic Acid and Jasmonate Signalling Pathways in Poplar. New Phytologist, 2022, 235, 701–717. [Google Scholar] [CrossRef] [PubMed]
- Sherif, S.M.; Shukla, M.R.; Murch, S.J.; Bernier, L.; Saxena, P.K. Simultaneous Induction of Jasmonic Acid and Disease-Responsive Genes Signifies Tolerance of American Elm to Dutch Elm Disease. Sci Rep, 2016, 6. [CrossRef]
- Silverman, P.; Seskar, M.; Kanter, D.; Schweizer, P.; Métraux, J.P.; Raskin, I. Salicylic Acid in Rice: Biosynthesis, Conjugation, and Possible Role. Plant Physiol, 1995, 108. [CrossRef]
- Likić, S.; Šola, I.; Ludwig-Müller, J.; Rusak, G. Involvement of Kaempferol in the Defence Response of Virus Infected Arabidopsis Thaliana. Eur J Plant Pathol, 2014, 138. [CrossRef]
- Caarls, L.; Pieterse, C.M.J.; van Wees, S.C.M. How Salicylic Acid Takes Transcriptional Control over Jasmonic Acid Signaling. Front Plant Sci, 2015, 6. [CrossRef]
- Djamei, A.; Schipper, K.; Rabe, F.; Ghosh, A.; Vincon, V.; Kahnt, J.; Osorio, S.; Tohge, T.; Fernie, A.R.; Feussner, I.; Feussner, K.; Meinicke, P.; Stierhof, Y.D.; Schwarz, H.; MacEk, B.; Mann, M.; Kahmann, R. Metabolic Priming by a Secreted Fungal Effector. Nature, 2011, 478. [CrossRef]
- Altmann, M.; Altmann, S.; Rodriguez, P.A.; Weller, B.; Elorduy Vergara, L.; Palme, J.; Marín-de la Rosa, N.; Sauer, M.; Wenig, M.; Villaécija-Aguilar, J.A.; Sales, J.; Lin, C.W.; Pandiarajan, R.; Young, V.; Strobel, A.; Gross, L.; Carbonnel, S.; Kugler, K.G.; Garcia-Molina, A.; Bassel, G.W.; Falter, C.; Mayer, K.F.X.; Gutjahr, C.; Vlot, A.C.; Grill, E.; Falter-Braun, P. Extensive Signal Integration by the Phytohormone Protein Network. Nature, 2020, 583. [CrossRef]
- Navarrete, F.; Gallei, M.; Kornienko, A.E.; Saado, I.; Khan, M.; Chia, K.S.; Darino, M.A.; Bindics, J.; Djamei, A. TOPLESS Promotes Plant Immunity by Repressing Auxin Signaling and Is Targeted by the Fungal Effector Naked1. Plant Commun, 2022, 3. [CrossRef]
Figure 1.
The titers of salicylic acid (P=0.06), jasmonic acid (P<0.05), and indole-3-acetic acid (P<0.05) in resistant and susceptible genotypes of European and Japanese plums to Black Knot (BK) at five different BK developmental stages (1-5) (Figure 1a). Stage 1 is the appearance of visual symptoms of BK while stage 5 is the most developed knot. Different letters denote statistical significance and error bars represent means ± SEM (ng/g DW) for all the phytohormones.
Figure 1.
The titers of salicylic acid (P=0.06), jasmonic acid (P<0.05), and indole-3-acetic acid (P<0.05) in resistant and susceptible genotypes of European and Japanese plums to Black Knot (BK) at five different BK developmental stages (1-5) (Figure 1a). Stage 1 is the appearance of visual symptoms of BK while stage 5 is the most developed knot. Different letters denote statistical significance and error bars represent means ± SEM (ng/g DW) for all the phytohormones.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



