
Submitted:
13 November 2023
Posted:
14 November 2023
You are already at the latest version
Abstract
Declining agricultural productivity caused by soil salinization are becoming global dilemmas in recent years. Biofertilizers show great potential as a sustainable and environmentally friendly fertilization strategy for soil improvement, but their effectiveness for saline soil amendment and improving plant growth under saline stress is not well understood. Assess the effectiveness of biofertilizers in improving saline soils and enhancing crop growth under saline stress, as well as investigate its related potential mechanisms. Changes in soil physico-chemical properties, plant physiological parameters and soil microbial communities were analyzed through pot experiments. The results showed that the application of biofertilizer reduced total soluble salts (TSS) in soil by 30.8% and increased the Brassica rapa L. biomass by 8.4 times. Biofertilizer application increased soil organic matter (SOM), total nitrogen (TN) and available phosphorus (AP). Biofertilizer applications also increased the SOD, CAT, chl.a, chl.b, total soluble sugars, and Proline content. Biofertilizers present plant growth promotion and potential pathogenic fungi reduction by increasing the abundance of Bacillus and Planococcus and decreasing the abundance of Mortierella and Aspergillus. Overall, the results of this study demonstrated the excellent efficacy of biofertilizers in improving saline soils, and the application of biofertilizer strategies will greatly promote agricultural production.
Keywords:
saline soil
; biofertilizer
; soil microbial community
; Bacillus licheniformis
; Halobacillus profundi
; Brassica rapa L.
1. Introduction
Soil quality is a key determinant of land productivity, and global demand for food is exploding as the world's population grows [1]. Considering the poor soil ecology, saline soil, which means soils containing excessive saline and alkaline components, is a major obstacle to agricultural development and food security [2]. Globally, the area of saline soils was 935 million hectares in 2020 [3], which has been expanding at a rate of three hectares per minute because of unfavorable conditions, especially insufficient irrigation and longstanding aridity [4]. Soil salinization caused an annual economic loss of $27.3 billion owing to land degradation in irrigated regions [5]. Therefore, saline soils are in urgent need of restoration and improvement to sustain land productivity, guarantee food security, and avoid potential loss.
Improving saline soils is important for agricultural development and protecting food security [6]. Reclaiming salty land, building irrigation systems, and applying chemicals were the previously primary methods for saline soil amendment [7]. However, because of the necessitated initial investment as well as long-term upkeep, reclaiming salty land or constructing irrigation systems tends to be less cost-effective [8]. There were also numerous disadvantages of chemical agents, such as high prices, damaging soil structure, and impacting soil microbial activity [9]. Promoting an economically and environmentally friendly amendment method is therefore essential.
Biofertilizer is a widely used and safe fertilizer that combines organic fertilizer and plant growth promoting rhizobacteria (PGPR) [9,10]. The high level of organic matter can enhance the structure of soil, increase the activity of soil enzymes and promote the growth of plants [11]. In addition, biofertilizers can improve soils by promoting soil microbial community succession [12]. However, little attention was distributed to improving saline soils with biofertilizers.
In this study, it was hypothesized that biofertilizers have an excellent saline soil improvement effect. This study was aimed at determining the impact of biofertilizers in saline soil reclamation, and the associated potential mechanisms in this process were discussed.
2. Materials and Methods
2.1. Experimental design
The experimental soil was sampled at Shihezi, Xinjiang, China (44.32°N, 85.88°E), and then passed through a sieve fitted for 2 mm holes. The organic fertilizer applied in this study was produced by composting livestock and poultry manure [13,14]. Details of the experimental soils and organic fertilizer are shown in A Table S1. Bacillus licheniformis 4-2 (NCBI accession number: ON926979) strain or Halobacterium subtilis GT42 (NCBI accession number: ON926973). Strain inoculated with organic fertilizer to produce biofertilizer. The viable counts of B. licheniformis and H. profundi in biofertilizers were both greater than 2.0×108 CFU·g-1.
Pot trials were performed in a greenhouse during March to May 2021, which consisted of five treatments, with each performed in triplicate. Then, the sieved saline soil was randomly distributed and evenly loaded in 15 flowerpots (50 cm × 40 cm × 30 cm), each flowerpot containing 21 kg of soil. Five treatments were: CK (no fertilizer); OF (100% organic fertilizer, 300 g); BF (B. licheniformis biofertilizer, 300 g); HF (H. profundi biofertilizer, 300 g); OM (B. licheniformis biofertilizer, 150 g, with H. profundi biofertilizer, 150 g). All fertilizers were applied as base fertilizer without topdressing. The employed crop, Brassica rapa L., was first cultivated to seedlings elsewhere and then transplanted into experimental pots. In each pot, 12 plants were transplanted under identical growth conditions (four leaves, root length 6 cm). The average temperature in the greenhouse was maintained at 25 °C during the experiment.
2.2. Soil sampling and physicochemical properties analysis
In this experiment, the growth period of Brassica rapa L. was 30 days. After harvesting, soil samples are taken, and the soil in each pot is thoroughly mixed after removing vegetation and debris. Each pot's soil was sampled, and each sample was split into two sub-samples, one sieved through a 2mm sieve and cryopreserved for microbial DNA extraction, while the other stored in cold storage for physical and chemical soil testing. Determination methods of soil carbon and nitrogen indicators were based on a previous study [15]. The Soil pH, soil organic matter (SOM), available phosphorus (AP), and total soluble salts (TSS) content were evaluated in accordance with Chinese standards [16].
2.3. Soil DNA extraction and amplicon sequencing
Primer pairs 338F/806R and ITS1 /ITS2 were used to amplify the 16S ribosomal RNA gene for bacteria and the ITS gene for fungi, respectively [15]. Paired-end reads were generated after high-throughput sequencing on an Illumina MiSeq PE300 platform. Paired-end reads were Paired-end reads for sequence quality control and pre-processing through usearch 11 and Vsearch 2.22.1 [17,18]. The pre-processed reads were denoised through the unosie3 function to generate ASVs. Representative sequences were classified by the feature classifiers using the SILVA reference database (v.138.1) and UNITE (v.29.11.2022) in qiime2 (v.2022.8) [19]. Sequences corresponding to chimeras, chloroplasts, mitochondria and archaea were discarded. The NCBI database contains raw sequencing data with the accession numbers SRP371928 and SRP371945.
2.4. Plant sampling and assaying
At harvest, the yield of the plant Brassica rapa L. was assessed, and plant samples were taken. Plant samples were split in half, with one half dried at 72 °C for 24 h to obtain the dry matter content, and the other used to measure the levels of plant photosynthetic pigments, antioxidant substances, and stress resistance substances. Immediately after harvest, Plant chlorophyll was measured using landscape photometry [20]. Proline content was obtained by the ninhydrin method. [21]. Soluble sugar content and enzyme activity were measured after the pretreatment of fresh plants [22]. The extracts were measured at an absorbance of 485 nm for total soluble sugars [23]. Superoxide dismutase (SOD) efficacy was assessed using the nitroblue tetrazolium chloride method. [24]. The enhanced Beers & Sizer technique was utilized to measure the activity of the catalase (CAT) [25]. Thiobarbituric acid reaction was used to assess the amount of malondialdehyde (MDA) present [22]. The activities of superoxide anion (O2·-) in plant were quantitatively determined using Superoxide Anion Activity Assay Kit (BC1290, Solarbio).
2.5. Statistical analysis
The means and standard deviations in this study were analyzed and plotted using Origin 2021b software (Origin Lab, USA). The alterations of soil physicochemical characteristics and biomass were examined using ANOVA followed by the Tukey test. Differences in alpha diversity between fertilization treatments were calculated using the Wilcoxon rank sum test. The statistical significance threshold in SPSS (V26) for the aforementioned investigations was 0.05. The microbial community's degree of similarity using R software (Version 4.1.0) was, based on NMDS of Bray-Curtis distance matrices. Here we used the redundancy analysis (RDA) to investigate the effect of soil physiochemistry on microbial community structure, and the precise localization of microbiota based on linear discriminant (LEfSe) analysis [15]. Assessment of the relative contribution of different fertilization scenarios using a Partial Least Squares Path Model (PLS-PM). Soil physicochemical parameters (pH, TC, TN, AP, C/N), SOM, TSS content, soil microorganism (Shannon index and Chao index), and plant physiology (SOD, CAT, MDA, O2·-, chl. a, chl. b, Total soluble sugar, Proline) on plant biomass using the plspm package (v 0.4.7) in R software [26].
3. Results and Discussion
3.1. Amendments of saline soil by biofertilizer
The salinity and plant biomass of saline land are important indicators to reflect the salinity degree and the plant's resistance to saline stress. Fertilization reduced the TSS content of each treatment by 24.2%-30.8% (p < 0.05) (Figure 1a) as compared with CK, with an insignificant between-group difference. Organic fertilizer can effectively reduce the TSS content, while adding PGPR to saline soil presents a relatively weak effect. Therefore, its organic components may be primarily responsible for the biofertilizer's beneficial impact on saline soil. The application of organic matter improved soil aggregate structure, increased soil bulk density and porosity, and thus promoted the leaching of soil soluble cations [27]. In addition, organic fertilizer introduced large amounts of humus to saline soils, which could adsorb Na+ into the soil and chelate Ca2+ and Mg2+ in high pH environments [11]. Therefore, the application of organic fertilizer presented excellent sedative effects, thus contributing to the decrease of TSS content.
Compared with CK and OF, BF, HF, and OM, all significantly increased plant biomass (p < 0.05), among which OM exhibited the best growth promotion effect. In comparison to CK, plant height and dry weight in OM rose by 1.6 and 8.4 times, respectively, and 0.5 times and 0.7 times relative to OF (Figure 1b,c). Fertilization also significantly increased plant root length and underground biomass (p < 0.05) (Figure S1). Several factors may be primarily responsible for biofertilizer's ability to stimulate plant growth in saline soils. SOM reduced the TSS content, controlled the water potential of plant roots, and relieved ion stress for plants [28]. Organic fertilizer boosted soil plant nutrition by encouraging the production of soil aggregates and soil porosity. In addition, Bacillus and Halobacillus in biofertilizer under salt stress exhibited protease, amylase, nitrogen fixation, and phosphorylation functions, and may release indole-3-acetic (IAA) to promote photosynthesis and plant root development [29,30]. Notably, mixed plant growth-promoting bacteria produced microbial complementary effects [31], which could provide plausible reasoning for the above-mentioned outstanding performance of OM.
3.2. Response of plant physiological characteristics to fertilization under salt stress
To identify the correlation between the biofertilizers application and plant growth in saline soils, the physiological properties of plants, such as antioxidant activity, oxidant accumulation, photosynthetic pigment content, and osmotic regulators, were examined. The results showed that biofertilizers improved the overall plant physiology. For example, the use of biofertilizer activated the enzymatic activity of plant antioxidants and reduced oxide levels. SOD and CAT of OM increased by 60.2% and 89.4%, while MDA and O2·- decreased by 30.1% and 66.6% compared to CK (Figure 2a–d). SOD and CAT effectively removed O2·- and H2O2, and mitigated plant damage caused by the accumulation of reactive oxygen species (ROS). PGPR, including B. amyloliquefaciens and B. thuringiensis, induced SOD and CAT production in plants by producing signaling molecules that significantly reduced the accumulation of oxidative O2·- and MDA toxins in plant tissues [32]. Levels of antioxidant enzymes in the biofertilizer application group were higher than those treated with organic fertilizer alone (Figure 2a,b), suggesting that the added microorganisms might further promote the secretion of plant oxidase. In terms of reducing oxidative toxins, HF and OM groups presented the best performance, which may be related to the addition of Halobacillus. Using functional bacteria, particularly Halobacillus, biofertilizers are thought to reduce oxidative stress through the production of antioxidant enzymes and the removal of oxidative pollutants.
Fertilization obviously elevated the chlorophyll content of plants (p < 0.05) (Figure 2e,f). The single inoculum addition treatment and the OF treatment presented minor difference, but OM obtained the maximum chlorophyll content (p < 0.05). chl a and chl b levels in OM were higher by 1.4-fold and 1.1-fold, respectively, compared to the control group, which indicated that biofertilizers have the potential to increase photosynthesis in plants. The photosynthesis rate is highly sensitive to salt stress and closely related to plant development and biomass buildup [33]. Plant chloroplast ion poisoning and chlorophyll content decrease inhibited plant photosynthesis in saline soils [34]. Elevated chlorophyll content of plants can effectively increase plant photosynthesis to improve plant stress resistance [35]. The biofertilizer may encourage plants to use water and nutrients efficiently under drought stress, and thus restore plant photosynthesis [36], which seems to be further enhanced through interactions and cooperation of B. licheniformis and H. profundi in the present study.
Additionally, fertilization encouraged plants to accumulate total soluble sugar and Proline (Figure 2g,h). Proline content was substantially greater in the fertilization treatment groups than it was in CK (p <0.05). Notably, OM had a 1.9 and 1.3 times higher plant total soluble sugar and proline content than CK. Based on these results and previous reports, it was hypothesized that in the low water potential environment, biofertilizer could encourage plants to produce Proline and carbohydrates, and slow down their deterioration, thus, causing plants in saline soils to accumulate total soluble sugar and Proline [37,38]. Proline and sugars helped ease osmotic stress, balance the content of K+/Na+ in plants, and preserve plant cell membranes while fostering photosynthesis [39]. The use of biofertilizer helped plants accumulate total soluble sugar and Proline while also lowering osmotic stress.
3.3. Improvement of saline soil properties by biofertilizer
To evaluate how biofertilizer affects the properties of saline soils, physicochemical indicators were examined. The contents of SOM and TC in the soil were dramatically enhanced by fertilization, and OM showed the most obvious promotion (SOM and TC contents increased by 56.1% and 28.9%) (p < 0.05) (Figure 3a,b). Application of biofertilizer considerably decreased soil pH; however, the biofertilizer treatments did not statistically vary from one another, and OM's pH was much lower than that of CK and OF's (p < 0.05) (Figure 3b). SOM could improve the aggregate formation and soil structure. The stimulating effect of the biofertilizers and the manipulation of the microbial community can also regulate the mineralization of SOC [40]. The rise in soil TC may be related to the acidity in saline-alkali soils. The alkaline environment inhibits the reaction process of converting carbonate to carbon dioxide in the soil [41]. This suggested that using biofertilizers would be able to improve the carbon sink of saline soil. The added biofertilizers slightly reduced the C/N ratio of the saline soil, while increased the TN content (p < 0.05) (Figure 3d,e). In comparison to CK, TN in OM was elevated by 57%. Despite that the C/N ratio in the fertilization groups was much lower than that of CK, no discernible between-group difference was observed. Previous studies reported that biofertilizer inoculated with Bacillus sp. minimized ammonia volatilization (a crucial pathway of nitrogen loss) in alkaline soils [15]. Biofertilizers increase the content of inorganic nitrogen (NH4+-N, NO3--N, NO2--N) (Figure S2). The nutritional needs of plants growing on saline land could be met by solving the problem of nutrient deprivation in saline soil. Previous study have shown that soil microorganisms depend on the proper C/N ratio [42]. Microbial community succession in soils may be driven by the change in the C/N ratio during biofertilizer application.
Moreover, fertilizers application had a significant effect on soil AP (Figure 3f). Among them, HF and OM had the most significant effect on improving soil AP content compared with CK (p < 0.05), which increased by 3.1 times and 2.9 times, respectively. Due to the high pH of saline soil, most of the phosphorus elements existed in the form of ineffective phosphorus, such as Ca3(PO4)2, which has low bioavailability and results in the oligotrophy of saline soil [43]. Furthermore, H. profundi. has the potential to dissolve phosphate and release soluble phosphate ions, and the use of organic fertilizers could significantly raise the AP content of the soil [44,45,46]. Biofertilizers with the addition of PRPG might effectively improve the nutrient status of saline soils.
3.4. Alteration of microbial communities in saline soils by biofertilizer
Correlation between fertilization and microbial communities in saline soils were examined by amplified sequencing. Fertilization obviously improved bacterial Chao index (p < 0.05) while exerting little influence on fungal counterpart, and hardly affected the Shannon and Shannoneven indexes (Table 1), indicating fertilization may boost bacterial richness in saline soil. Similar enhancement limited to bacterial richness was observed in the synergistic remediation of saline land by plants and soil amendments [47]. The organic matter input increased microbial diversity in saline soils [48]. The improved soil physicochemical properties described in Section 3.3, especially abundant SOM, available nutrients and proper pH, could greatly favor bacterial thriving.
The results of NMDS research revealed that fertilization treatments' microbial community compositions considerably differed from CK (Figure 4). Previous studies had shown that SOM application significantly altered microbial community structure in saline soils [49]. Differences in microbial community composition between treatments may be driven by SOM.
There were striking phylum-level differences in the community composition under different fertilizations. Fertilization promoted the thriving of Bacteroidota, Firmicutes, Patescibacteria, Myxococcota, and Chloroflexi in OM compared to CK (68.2%, 246.3%, 27.8%, 188.9%, and 24.4%, respectively). The relative abundance of Actinobacteriota, Gemmatimonadota, Acidobacteriota, Cyanobacteria, and Desulfobacterota decreased in each fertilization treatment (p < 0.05) (Figure 5a). Previous research has shown that Bacteroidetes were normally capable of producing enzymes to break down starch and cellulose [50], Chloroflexi is also an important driver of SOM mineralization [51]. As a result, soil mineralization may have been aided by the use of biofertilizer and released more inorganic nutrients.
Fertilization promoted the thriving of Bacillus, Planococcus, and Salegentibacter compared to CK (2.2, 5.1, 46.6 times, respectively) (p < 0.05). Simultaneously, fertilization inhibited the growth of some genera, like Marinobacter, Arthrobacter, and Nitrolancea, with the relative abundance in OM 38.1%, 67.7%, 74.8% lower than CK (p < 0.05) (Figure 5c). Among them, Bacillus, a typical plant growth-promoting bacteria capable of producing signal molecules and secreting auxin [52], could assist in promoting plant development in saline soils. Meanwhile, Planococcus could dissolve phosphorus [53] and thus possibly account for the increased AP content in fertilized soils. Moreover, Planococcus may release ACC dehydrogenase, which can encourage microbial colonization and biofilm formation, enhancing plants' resilience to salt and alkali [54,55].
In terms of fungi, Ascomycota accounted for the relative abundance of over 80% in each group, suggesting that fertilization imposed an insignificant impact on the fungal community at the phylum level. Fertilization resulted in an abundance decline of 81.9%, 84.9%, and 82.6% in OM compared with CK for Mortierellomycota, Basidiomycota, and Chytridiomycota, respectively (p < 0.05) (Figure 5b). A number of plant pathogens originate from the Basidiomycota [56], which is a possible cause of reduced plant disease in saline soils.
At the level of the fungal genus, OM had considerably greater relative abundances of Acaulium, Sodiomyces, and Kernia than CK (92.7, 256.6, 9.6 times, respectively); Chaetomium, Mortierella, and Aspergillus in OM were significantly lower than that of CK (4.9, 5.7, and 1.3 times, respectively) (p < 0.05) (Figure 5d). Researchers considered Mortierella as a potential animal pathogen [57], and Aspergillus as a typical plant pathogenic bacteria [58]. According to this study, using biofertilizers reduced the number of these potentially dangerous bacteria, which may also lower the risk of plant diseases.
With the LDA threshold of 2.7, 32 bacterial taxa with statistically abundance differentials were identified by LEfSe analysis (p <0.05) (Figure 5e). Among them, OM had the most significantly different microorganisms, with a total of ten microorganisms, represented by Salegentibacter. Only six microbial genus were observed significant differences in CK, such as Marivirga. Previous studies have found that Luteimonas and Arenimonas have the potential to promote the aromatization and humification of SOM, which were significantly correlated with the growth of SOM, and were potential plant growth-promoting bacteria [59,60,61]. The application of biofertilizer caused significant differences in microorganisms of OM treatment. Therefore, there may be a deposit effect on SOM humification and increasing soil soluble organic matter.
The pH value, the TSS content, and the C/N ratio had a negative effect on the microbial community of the biofertilizer treatment, whereas the SOM, the TC, the TN, and the AP had a positive effect (Figure 6). The correlations between dominant microbial genera and physicochemical properties of saline soils were illustrated in Figure 7. In the bacterial community, SOM, TC, and TN were linked with Chryseolinea, Cellvibrio, Planococcus favorably, and are adversely associated to Nitrolancea, Limnobacter, Truepera, etc. Previous studies have suggested that Chryseolinea promoted plant growth by regulating nutrient uptake [62], and Cellvibrio may facilitate soil humification by producing amylase and cellulase [63].
The application of biofertilizer increased soil SOM, TC, TN, and the relative abundance of Cellvibrio and Chryseolinea increased the humification of soil cellulose and encouraged plants to absorb soil nutrients. TSS content, pH, and the C/N ratio were favorably connected with Sphingomonas and Vicingus, while negatively linked with Bacillus and Planococcus (Figure 7 a). This may suggest that biofertilizers improved the aforementioned soil qualities by reducing the stress of soil salinity, which may have favored the soil bacteria that benefits plant development.
For fungi, SOM, TC, and TN favored the thriving of Madurella, Sodiomyces, and Acaulium, while inhibiting the growth of Mortierella and Humicola. TSS content, pH, and C/N ratio were favorably connected with Mortierella, whereas it exerted the converse effect on Sodiomyces and Acaulium (Figure 7b). Sodiomyces is able to produce polysaccharides in an alkaline environment that help build extracellular polymers to support plant development [64]. Mortierella and Humicola, as pathogenic fungi, could induce animal and plant disease [65,66]. Biofertiliser application regulates changes in soil microbial diversity and encourages the development of Sodiomyces while decreasing the relative abundance of pathogenic bacteria, like Mortierella and Humicola, both directly and indirectly aided plant growth, enhanced plant stress resistance, and decreased the risk of plant diseases.
3.5. Response of plant physiological characteristics to fertilization under salt stress
The interactions between different factors and biomass during the application of Biofertilizer for the improvement of saline soils have been studied by means of PLS-PM (Figure 8). Through PLS-PM analysis, it was found that plant physiological factors were the most direct factors affecting the plant biomass in saline soils, followed by soil microorganisms. The fertilizer application significantly increased the content of SOM while reducing the TSS content. Changes in the soil environment could effectively drive microbial communities [67]. SOM caused changes in soil physicochemical indicators and microbial diversity that were significantly correlated with changes in plant physiological parameters. The presence of TSS content in saline soils led to negative physiological responses and stress on plants. Biofertilizer application promoted the succession of soil microbial communities mainly by increasing SOM. Soil microorganisms adjusted the physiological response of plants and improved the resistance of plants in saline soils. Through the above pathways, plant stress tolerance and plant biomass accumulation in saline soils were promoted.
4. Conclusions
This study demonstrated the efficacy of biofertilizers in ameliorating saline soil, as evidenced by an 8.4-fold increase in plant biomass and a 0.3-fold decrease in TSS. Significant increases in SOM, TC, TN and AP in saline soil were also observed, indicating significant improvements in soil structure and nutrient status. In terms of physiological traits, biofertilizers application increased the levels of SOD, CAT, chl.a, chl.b, total soluble sugars, and Proline, and reduced the concentrations of MDA and O2·-. These changes in physiological traits probably resulted in improved salt stress tolerance and higher plant biomass. Moreover, the application of biofertilizers altered the soil microbial community composition, such as an increase in the abundance of Bacillus and Planococcus, which may benefit soil quality and plant growth. Meanwhile, the abundance of phytopathogenic fungi such as Mortierella and Aspergillus declined. Altogether, these results indicated that biofertilizer was effective in improving saline soils and enhancing plant growth under saline stress. Consequently, biofertilizer could be a promising strategy to improve saline soils and enhance agricultural production.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Properties of saline-alkali soil and organic fertilizers; Figure S1: Plant root length (a) and dry weight of underground part (b) of plant; Figure S2: The content of NH4+-N, NO3--N, NO2--N in the saline-alkali soil.
Author Contributions
Conceptualization, Z.B.; Methodology, Z.B. and R.L.; Investigation, R.L., B.S., M.S. and G.Y.; Data curation, R.L., B.S., and M.S.; Visualization, R.L., B.S., M.S. and Q.H.; Writing - original draft, R.L., B.S., M.S. and Z.B.; Writing - review and editing, Q.H., X.Z. and J.Z.; Project administration, Z.B.; Funding acquisition, Z.B.; Supervision, Z.B., X.Z. and J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Weiqiao-UCAS Special Projects on Low-Carbon Technology Development (No. GYY-DTFZ-2022-004), the Strategic Priority Research Program of Chinese Academy of Sciences (No. XDA2301040103), the National Natural Science Foundation of China (No. 42377126) and the National Key Research & Development Program of China (2016YFC0501404).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw sequencing data were deposited into SRA database with an accession number of SRP371928 and SRP371945.
Acknowledgments
The authors are thankful to Binzhou Institute of Technology for providing consumables for the experimentation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Amirnia, R., Ghiyasi, M., Moghaddam, S.S., Rahimi, A., Damalas, C.A., Heydarzadeh, S., 2019. Nitrogen-Fixing soil bacteria plus mycorrhizal fungi improve seed yield and quality traits of lentil (Lens culinaris Medik). J. Soil Sci. Plant Nutr. 19, 592–602. [CrossRef]
- Backer, R., Rokem, J.S., Ilangumaran, G., Lamont, J., Praslickova, D., Ricci, E., Subramanian, S., Smith, D.L., 2018. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 9. [CrossRef]
- Barrón, C., Duarte, C.M., Frankignoulle, M., Borges, A.V., 2006. Organic carbon metabolism and carbonate dynamics in a Mediterranean seagrass (Posidonia oceanica), meadow. Estuaries Coasts 29, 417–426. [CrossRef]
- Bolyen, E., Rideout, J.R., Dillon, M.R., Bokulich, N.A., Abnet, C.C., Al-Ghalith, G.A., Alexander, H., Alm, E.J., Arumugam, M., Asnicar, F., Bai, Y., Bisanz, J.E., Bittinger, K., Brejnrod, A., Brislawn, C.J., Brown, C.T., Callahan, B.J., Caraballo-Rodríguez, A.M., Chase, J., Cope, E.K., Da Silva, R., Diener, C., Dorrestein, P.C., Douglas, G.M., Durall, D.M., Duvallet, C., Edwardson, C.F., Ernst, M., Estaki, M., Fouquier, J., Gauglitz, J.M., Gibbons, S.M., Gibson, D.L., Gonzalez, A., Gorlick, K., Guo, J., Hillmann, B., Holmes, S., Holste, H., Huttenhower, C., Huttley, G.A., Janssen, S., Jarmusch, A.K., Jiang, L., Kaehler, B.D., Kang, K.B., Keefe, C.R., Keim, P., Kelley, S.T., Knights, D., Koester, I., Kosciolek, T., Kreps, J., Langille, M.G.I., Lee, J., Ley, R., Liu, Y.-X., Loftfield, E., Lozupone, C., Maher, M., Marotz, C., Martin, B.D., McDonald, D., McIver, L.J., Melnik, A.V., Metcalf, J.L., Morgan, S.C., Morton, J.T., Naimey, A.T., Navas-Molina, J.A., Nothias, L.F., Orchanian, S.B., Pearson, T., Peoples, S.L., Petras, D., Preuss, M.L., Pruesse, E., Rasmussen, L.B., Rivers, A., Robeson, M.S., Rosenthal, P., Segata, N., Shaffer, M., Shiffer, A., Sinha, R., Song, S.J., Spear, J.R., Swafford, A.D., Thompson, L.R., Torres, P.J., Trinh, P., Tripathi, A., Turnbaugh, P.J., Ul-Hasan, S., van der Hooft, J.J.J., Vargas, F., Vázquez-Baeza, Y., Vogtmann, E., von Hippel, M., Walters, W., Wan, Y., Wang, M., Warren, J., Weber, K.C., Williamson, C.H.D., Willis, A.D., Xu, Z.Z., Zaneveld, J.R., Zhang, Y., Zhu, Q., Knight, R., Caporaso, J.G., 2019. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. [CrossRef]
- Bondarenko, S.A., Ianutsevich, E.A., Danilova, O.A., Grum-Grzhimaylo, A.A., Kotlova, E.R., Kamzolkina, O.V., Bilanenko, E.N., Tereshina, V.M., 2017. Membrane lipids and soluble sugars dynamics of the alkaliphilic fungus Sodiomyces tronii in response to ambient pH. Extremophiles 21, 743–754. [CrossRef]
- Chavoushi, M., Najafi, F., Salimi, A., Angaji, S.A., 2020. Effect of salicylic acid and sodium nitroprusside on growth parameters, photosynthetic pigments and secondary metabolites of safflower under drought stress. Sci. Hortic. 259, 108823. [CrossRef]
- de Azevedo Neto, A.D., Prisco, J.T., Enéas-Filho, J., Abreu, C.E.B. de, Gomes-Filho, E., 2006. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 56, 87–94. [CrossRef]
- Desale, P., Patel, B., Singh, S., Malhotra, A., Nawani, N., 2014. Plant growth promoting properties of Halobacillus sp. and Halomonas sp. in presence of salinity and heavy metals. J. Basic Microbiol. 54, 781–791. [CrossRef]
- Devkota, K.P., Devkota, M., Rezaei, M., Oosterbaan, R., 2022. Managing salinity for sustainable agricultural production in salt-affected soils of irrigated drylands. Agric. Syst. 198, 103390. [CrossRef]
- Ding, Z., Kheir, A.M.S., Ali, O.A.M., Hafez, E.M., ElShamey, E.A., Zhou, Z., Wang, B., Lin, X., Ge, Y., Fahmy, A.E., Seleiman, M.F., 2021. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manage. 277, 111388. [CrossRef]
- Edgar, R.C., 2018. Accuracy of taxonomy prediction for 16S rRNA and fungal ITS sequences. PeerJ 6, e4652. [CrossRef]
- FAO, 2021a. Global Map of Salt Affected Soils.
- FAO, 2021b. Economical losses due to soil salinization.
- Gadallah, M. a. A., 1999. Effects of proline and glycinebetaine on Vicia faba responses to salt stress. Biol. Plant. 42, 249–257. [CrossRef]
- Gu, Y., Dong, K., Geisen, S., Yang, W., Yan, Y., Gu, D., Liu, N., Borisjuk, N., Luo, Y., Friman, V.-P., 2020. The effect of microbial inoculant origin on the rhizosphere bacterial community composition and plant growth-promotion. Plant Soil 452, 105–117. [CrossRef]
- Jiang, Z., Lu, Y., Xu, J., Li, M., Shan, G., Li, Q., 2019. Exploring the characteristics of dissolved organic matter and succession of bacterial community during composting. Bioresour. Technol. 292, 121942. [CrossRef]
- Kean, T., Thanou, M., 2010. Biodegradation, biodistribution and toxicity of chitosan. Adv. Drug Deliv. Rev. 62, 3–11. [CrossRef]
- Kolton, M., Harel, Y.M., Pasternak, Z., Graber, E.R., Elad, Y., Cytryn, E., 2011. Impact of biochar application to soil on the root-associated bacterial community structure of fully developed greenhouse pepper plants. Appl. Environ. Microbiol. 77, 4924–4930. [CrossRef]
- Larsbrink, J., McKee, L.S., 2020. Chapter Two - Bacteroidetes bacteria in the soil: Glycan acquisition, enzyme secretion, and gliding motility, in: Gadd, G.M., Sariaslani, S. (Eds.), Advances in Applied Microbiology. Academic Press, pp. 63–98. [CrossRef]
- Leogrande, R., Vitti, C., 2019. Use of organic amendments to reclaim saline and sodic soils: a review. Arid Land Res. Manag. 33, 1–21. [CrossRef]
- Li, F., Chen, L., Zhang, J., Yin, J., Huang, S., 2017. Bacterial community structure after long-term organic and inorganic fertilization reveals important associations between soil nutrients and specific taxa involved in nutrient transformations. Front. Microbiol. 8. [CrossRef]
- Li, N., Shao, T., Zhou, Y., Cao, Y., Hu, H., Sun, Q., Long, X., Yue, Y., Gao, X., Rengel, Z., 2021. Effects of planting Melia azedarach L. on soil properties and microbial community in saline-alkali soil. Land Degrad. Dev. 32, 2951–2961. [CrossRef]
- Li, Y., Zhang, Jiejie, Zhang, Jianqiang, Xu, W., Mou, Z., 2019. Characteristics of inorganic phosphate-solubilizing bacteria from the sediments of a eutrophic Lake. Int. J. Environ. Res. Public. Health 16, 2141. [CrossRef]
- Liao, H., Zhao, Q., Cui, P., Chen, Z., Yu, Z., Geisen, S., Friman, V.-P., Zhou, S., 2019. Efficient reduction of antibiotic residues and associated resistance genes in tylosin antibiotic fermentation waste using hyperthermophilic composting. Environ. Int. 133, 105203.
- Lima, D.L.D., Santos, S.M., Scherer, H.W., Schneider, R.J., Duarte, A.C., Santos, E.B.H., Esteves, V.I., 2009. Effects of organic and inorganic amendments on soil organic matter properties. Geoderma 150, 38–45. [CrossRef]
- Liu, Q., Tang, J., Liu, X., Song, B., Zhen, M., Ashbolt, N.J., 2019. Vertical response of microbial community and degrading genes to petroleum hydrocarbon contamination in saline alkaline soil. J. Environ. Sci. 81, 80–92. [CrossRef]
- Liu, Z., Shang, H., Han, F., Zhang, M., Li, Q., Zhou, W., 2021. Improvement of nitrogen and phosphorus availability by Pseudoalteromonas sp. during salt-washing in saline-alkali soil. Appl. Soil Ecol. 168, 104117. [CrossRef]
- Mohammadi, K., Heidari, G., Khalesro, S., Sohrabi, Y., 2011. Soil management, microorganisms and organic matter interactions: A review. Afr. J. Biotechnol. 10, 19840–19849. [CrossRef]
- Mohanavelu, A., Naganna, S.R., Al-Ansari, N., 2021. Irrigation induced salinity and sodicity hazards on soil and groundwater: an overview of its causes, Impacts and mitigation strategies. Agriculture 11, 983. [CrossRef]
- Munchan, C., Kurata, O., Hatai, K., Hashiba, N., Nakaoka, N., Kawakami, H., 2006. Mass mortality of young striped jack pseudocaranx dentex caused by a fungus Ochroconis humicola. Fish Pathol. 41, 179–182. [CrossRef]
- Munns, R., Tester, M., 2008. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59, 651–681. [CrossRef]
- Orhan, F., 2021. Potential of Halophilic/Halotolerant bacteria in enhancing plant growth under salt stress. Curr. Microbiol. 78, 3708–3719. [CrossRef]
- Qi, R., Lin, W., Gong, K., Han, Z., Ma, H., Zhang, M., Zhang, Q., Gao, Y., Li, J., Zhang, X., 2021. Bacillus co-inoculation alleviated salt stress in seedlings cucumber. Agron.-Basel 11, 966. [CrossRef]
- Qurashi, A.W., Sabri, A.N., 2012. Bacterial exopolysaccharide and biofilm formation stimulate chickpea growth and soil aggregation under salt stress. Braz. J. Microbiol. 43, 1183–1191. [CrossRef]
- Radhakrishnan, R., Hashem, A., Abd_Allah, E.F., 2017. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 8. [CrossRef]
- Rady, M.M., Elrys, A.S., Selem, E., Mohsen, A.A.A., Arnaout, S.M.A.I., El-Sappah, A.H., El-Tarabily, K.A., Desoky, E.-S.M., 2022. Spirulina platensis extract improves the production and defenses of the common bean grown in a heavy metals-contaminated saline soil. J. Environ. Sci. [CrossRef]
- Rajput, L., Imran, A., Mubeen, F., Hafeez, F.Y., 2013. Salt-tolerant pgpr strain Planococcus Rifietoensis promotes the growth and yield of wheat (Triticum Aestivum L.) cultivated in saline soil. Pak. J. Bot. 45, 1955–1962. [CrossRef]
- Ribes, J.A., Vanover-Sams, C.L., Baker, D.J., 2000. Zygomycetes in human disease. Clin. Microbiol. Rev. 13, 236–301. [CrossRef]
- Rognes, T., Flouri, T., Nichols, B., Quince, C., Mahé, F., 2016. VSEARCH: a versatile open source tool for metagenomics. PeerJ 4, e2584. [CrossRef]
- Seyed Sharifi, R., Khalilzadeh, R., Jalilian, J., 2017. Effects of biofertilizers and cycocel on some physiological and biochemical traits of wheat (Triticum aestivum L.) under salinity stress. Arch. Agron. Soil Sci. 63, 308–318. [CrossRef]
- Shabala, S., Wu, H., Bose, J., 2015. Salt stress sensing and early signalling events in plant roots: Current knowledge and hypothesis. Plant Sci. 241, 109–119. [CrossRef]
- Shi, S., Tian, L., Nasir, F., Bahadur, A., Batool, A., Luo, S., Yang, F., Wang, Z., Tian, C., 2019. Response of microbial communities and enzyme activities to amendments in saline-alkaline soils. Appl. Soil Ecol. 135, 16–24. [CrossRef]
- Siddikee, Md.A., Chauhan, Puneet.S., Anandham, R., Han, G.-H., Sa, T., 2010. Isolation, characterization, and use for plant growth promotion under salt stress, of ACC deaminase-producing halotolerant bacteria derived from coastal soil. J. Microbiol. Biotechnol. 20, 1577–1584. [CrossRef]
- Singh, M., Awasthi, A., Soni, S.K., Singh, R., Verma, R.K., Kalra, A., 2015. Complementarity among plant growth promoting traits in rhizospheric bacterial communities promotes plant growth. Sci. Rep. 5, 15500. [CrossRef]
- Singh, S., Singh, M., 2000. Genotypic basis of response to saliniity stress in some crosses of spring wheat Triticum aestivum L. Euphytica 115, 209–214. [CrossRef]
- Sudhir, P., Murthy, S.D.S., 2004. Effects of salt stress on basic processes of photosynthesis. Photosynthetica 42, 481–486. [CrossRef]
- Sun, B., Bai, Z., Bao, L., Xue, L., Zhang, S., Wei, Y., Zhang, Z., Zhuang, G., Zhuang, X., 2020a. Bacillus subtilis biofertilizer mitigating agricultural ammonia emission and shifting soil nitrogen cycling microbiomes. Environ. Int. 144, 105989. [CrossRef]
- Sun, B., Bai, Z., Li, Y., Li, R., Song, M., Xu, S., Zhang, H., Zhuang, X., 2022a. Emission mitigation of CH4 and N2O during semi-permeable membrane covered hyperthermophilic aerobic composting of livestock manure. J. Clean. Prod. 379, 134850. [CrossRef]
- Sun, B., Gu, L., Bao, L., Zhang, S., Wei, Y., Bai, Z., Zhuang, G., Zhuang, X., 2020b. Application of biofertilizer containing Bacillus subtilis reduced the nitrogen loss in agricultural soil. Soil Biol. Biochem. 148, 107911. [CrossRef]
- Sun, B., Li, Y., Song, M., Li, R., Li, Z., Zhuang, G., Bai, Z., Zhuang, X., 2022b. Molecular characterization of the composition and transformation of dissolved organic matter during the semi-permeable membrane covered hyperthermophilic composting. J. Hazard. Mater. 425, 127496. [CrossRef]
- Szabados, L., Savouré, A., 2010. Proline: a multifunctional amino acid. Trends Plant Sci. 15, 89–97. [CrossRef]
- Tejada, M., Garcia, C., Gonzalez, J.L., Hernandez, M.T., 2006. Use of organic amendment as a strategy for saline soil remediation: Influence on the physical, chemical and biological properties of soil. Soil Biol. Biochem. 38, 1413–1421. [CrossRef]
- Trivedi, P., Singh, K., Pankaj, U., Verma, S.K., Verma, R.K., Patra, D.D., 2017. Effect of organic amendments and microbial application on sodic soil properties and growth of an aromatic crop. Ecol. Eng. 102, 127–136. [CrossRef]
- Tseng, M.J., Liu, C.-W., Yiu, J.-C., 2007. Enhanced tolerance to sulfur dioxide and salt stress of transgenic Chinese cabbage plants expressing both superoxide dismutase and catalase in chloroplasts. Plant Physiol. Biochem. 45, 822–833. [CrossRef]
- Velez, M.L., Marfetan, J.A., Salgado Salomon, M.E., Taccari, L.E., 2020. Mortierella species from declining Araucaria araucanatrees in Patagonia, Argentina. For. Pathol. 50, e12591. [CrossRef]
- Visioli, G., Sanangelantoni, A.M., Vamerali, T., Dal Cortivo, C., Blandino, M., 2018. 16S rDNA profiling to reveal the influence of seed-applied biostimulants on the rhizosphere of young maize plants. Molecules 23, 1461. [CrossRef]
- Volk, T.J., 2013. Fungi, in: Levin, S.A. (Ed.), Encyclopedia of Biodiversity (Second Edition). Academic Press, Waltham, pp. 624–640. [CrossRef]
- Wang, T., Chen, Y., Zhang, M., Chen, J., Liu, J., Han, H., Hua, X., 2017. Arabidopsis AMINO ACID PERMEASE1 contributes to salt stress-induced proline uptake from exogenous sources. Front. Plant Sci. 8, 2182. [CrossRef]
- Wang, X., Bian, Q., Jiang, Y., Zhu, L., Chen, Y., Liang, Y., Sun, B., 2021. Organic amendments drive shifts in microbial community structure and keystone taxa which increase C mineralization across aggregate size classes. Soil Biol. Biochem. 153, 108062. [CrossRef]
- Wang, X., Hu, M., Xia, Y., Wen, X., Ding, K., 2012. Pyrosequencing analysis of bacterial diversity in 14 wastewater treatment systems in China. Appl. Environ. Microbiol. 78, 7042–7047. [CrossRef]
- Wang, X., Xu, S., Wu, S., Feng, S., Bai, Z., Zhuang, G., Zhuang, X., 2018. Effect of Trichoderma viride biofertilizer on ammonia volatilization from an alkaline soil in Northern China. J. Environ. Sci. 66, 199–207. [CrossRef]
- Wu, S., Zhuang, G., Bai, Z., Cen, Y., Xu, S., Sun, H., Han, X., Zhuang, X., 2018. Mitigation of nitrous oxide emissions from acidic soils by Bacillus amyloliquefaciens, a plant growth-promoting bacterium. Glob. Change Biol. 24, 2352–2365. [CrossRef]
- Yang, W., Gong, T., Wang, J., Li, G., Liu, Y., Zhen, J., Ning, M., Yue, D., Du, Z., Chen, G., 2020. Effects of compound microbial fertilizer on soil characteristics and yield of wheat (Triticum aestivum L.). J. Soil Sci. Plant Nutr. 20, 2740–2748. [CrossRef]
- You, X., Yin, S., Suo, F., Xu, Z., Chu, D., Kong, Q., Zhang, C., Li, Y., Liu, L., 2021. Biochar and fertilizer improved the growth and quality of the ice plant (Mesembryanthemum crystallinum L.) shoots in a coastal soil of Yellow River Delta, China. Sci. Total Environ. 775, 144893. [CrossRef]
- Zhang, F., Xu, X., Wang, G., Wu, B., Xiao, Y., 2020. Medicago sativa and soil microbiome responses to Trichoderma as a biofertilizer in alkaline-saline soils. Appl. Soil Ecol. 153, 103573. [CrossRef]
- Zhang, X., Xu, S., Li, C., Zhao, L., Feng, H., Yue, G., Ren, Z., Cheng, G., 2014. The soil carbon/nitrogen ratio and moisture affect microbial community structures in alkaline permafrost-affected soils with different vegetation types on the Tibetan plateau. Res. Microbiol. 165, 128–139. [CrossRef]
- Zou, P., Tian, X., Dong, B., Zhang, C., 2017. Size effects of chitooligomers with certain degrees of polymerization on the chilling tolerance of wheat seedlings. Carbohydr. Polym. 160, 194–202. [CrossRef]
Figure 1.
The TSS content in the saline soil (a), Dry weight of aboveground part (b), and total height (c) of the plant. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi. P-values were analyzed using one-way ANOVA and Student's t-test (p < 0.05).
Figure 1.
The TSS content in the saline soil (a), Dry weight of aboveground part (b), and total height (c) of the plant. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi. P-values were analyzed using one-way ANOVA and Student's t-test (p < 0.05).
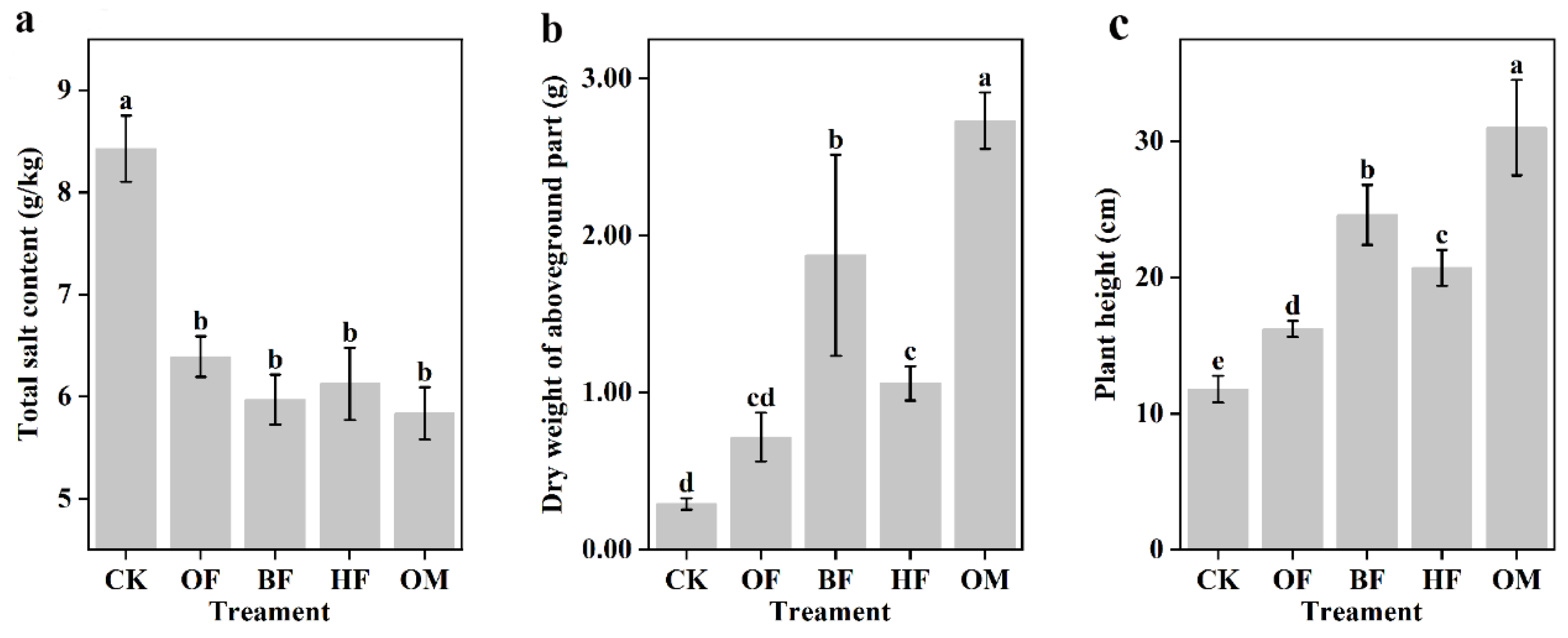
Figure 2.
The content of phytohormone SOD (a), O2·- (b), MDA (c), CAT (d), chl. a (e), chl. b (f), total soluble sugar (g), and Proline (h) inside the plant. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi. P-values were analyzed using one-way ANOVA and Student's t-test (p < 0.05).
Figure 2.
The content of phytohormone SOD (a), O2·- (b), MDA (c), CAT (d), chl. a (e), chl. b (f), total soluble sugar (g), and Proline (h) inside the plant. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi. P-values were analyzed using one-way ANOVA and Student's t-test (p < 0.05).
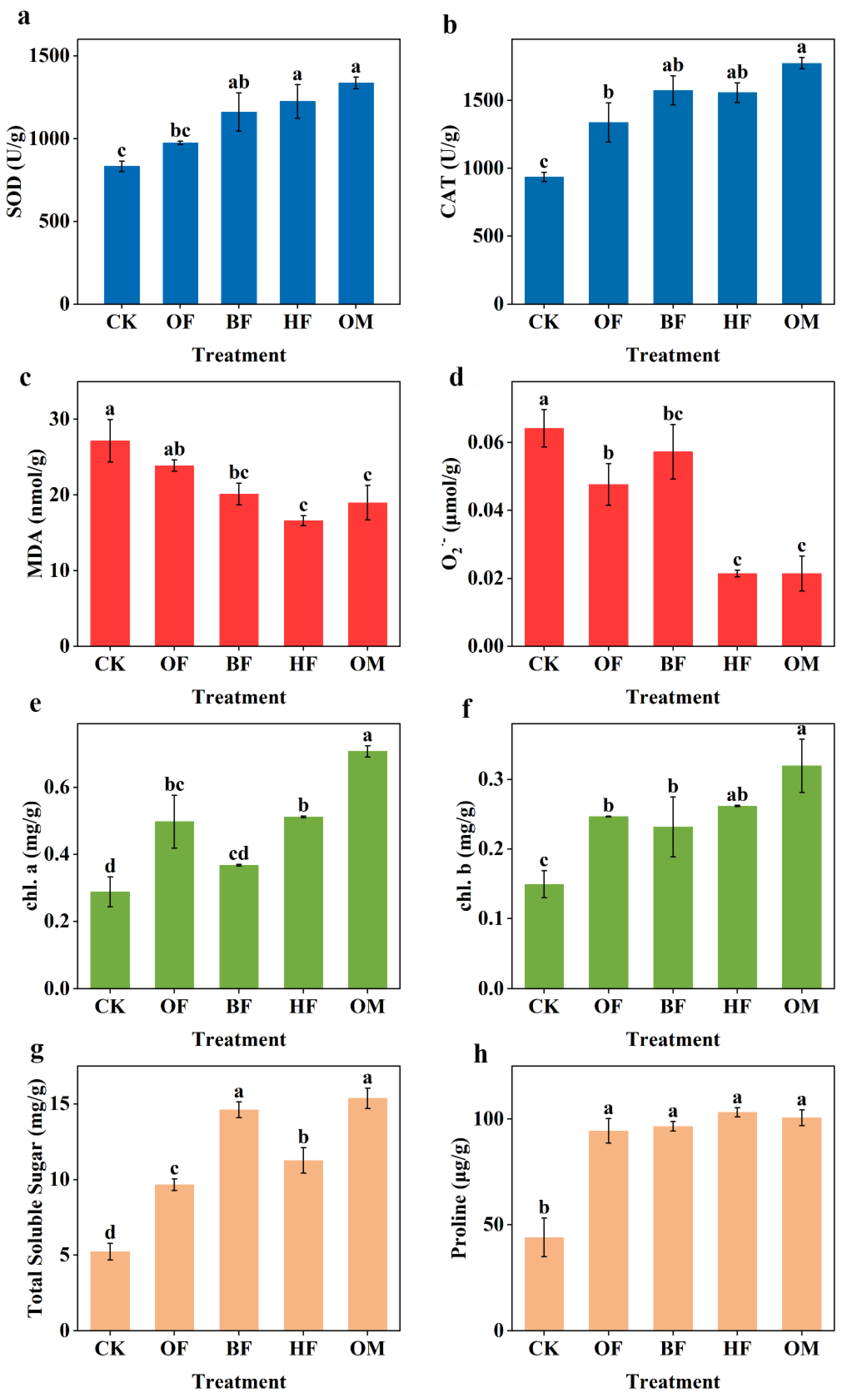
Figure 3.
The content of SOM (a), TC (b), pH (e), TN (d), C/N (e), and AP (f) in the saline soil. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi. P-values were analyzed using one-way ANOVA and Student's t-test (p < 0.05).
Figure 3.
The content of SOM (a), TC (b), pH (e), TN (d), C/N (e), and AP (f) in the saline soil. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi. P-values were analyzed using one-way ANOVA and Student's t-test (p < 0.05).
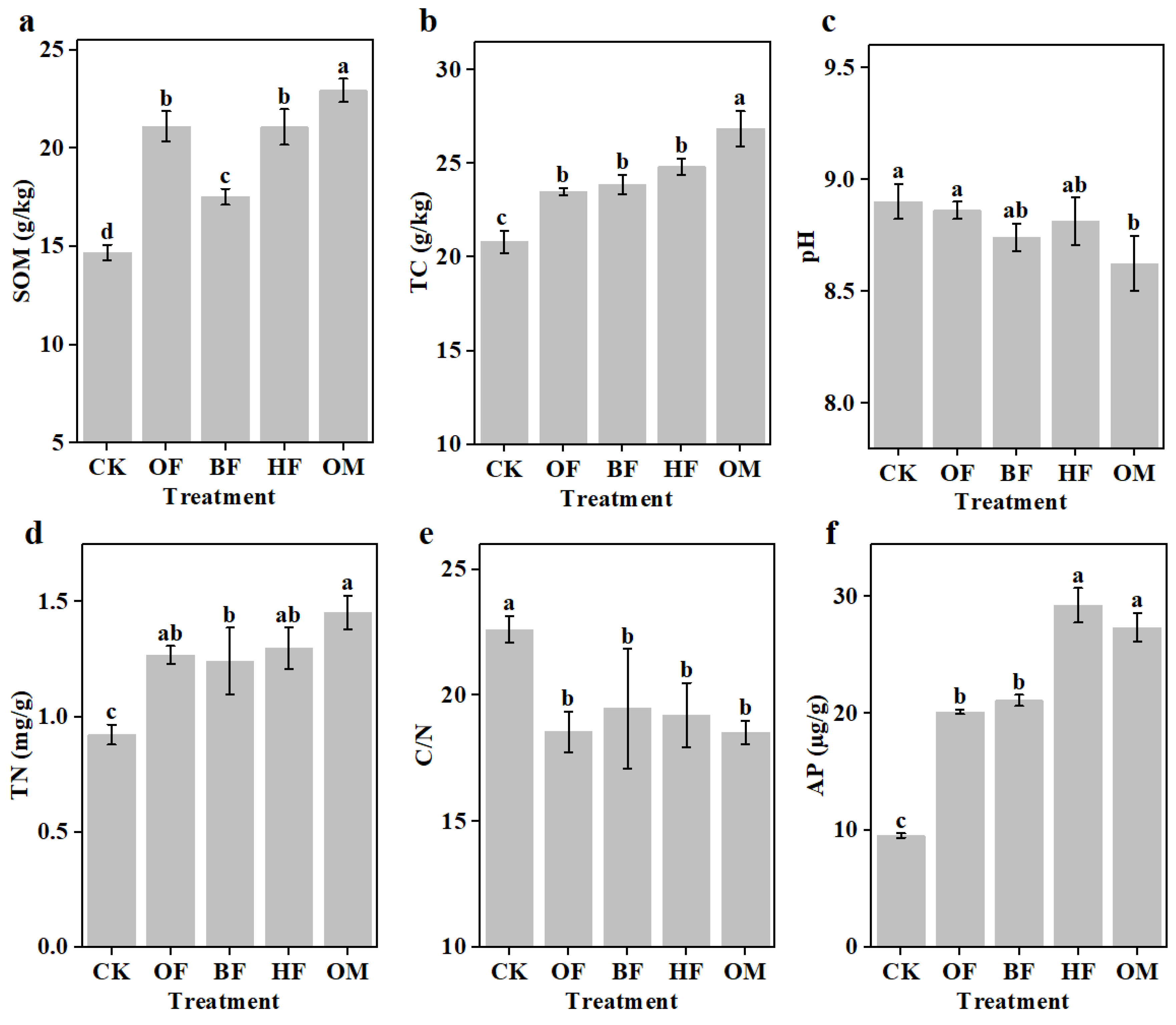
Figure 4.
Nonmetric multidimensional scaling (NMDS) ordination plots for bacterial (a) and fungal (b) community composition at genus level based on the Bray-Curtis distance similarity. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi.
Figure 4.
Nonmetric multidimensional scaling (NMDS) ordination plots for bacterial (a) and fungal (b) community composition at genus level based on the Bray-Curtis distance similarity. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi.
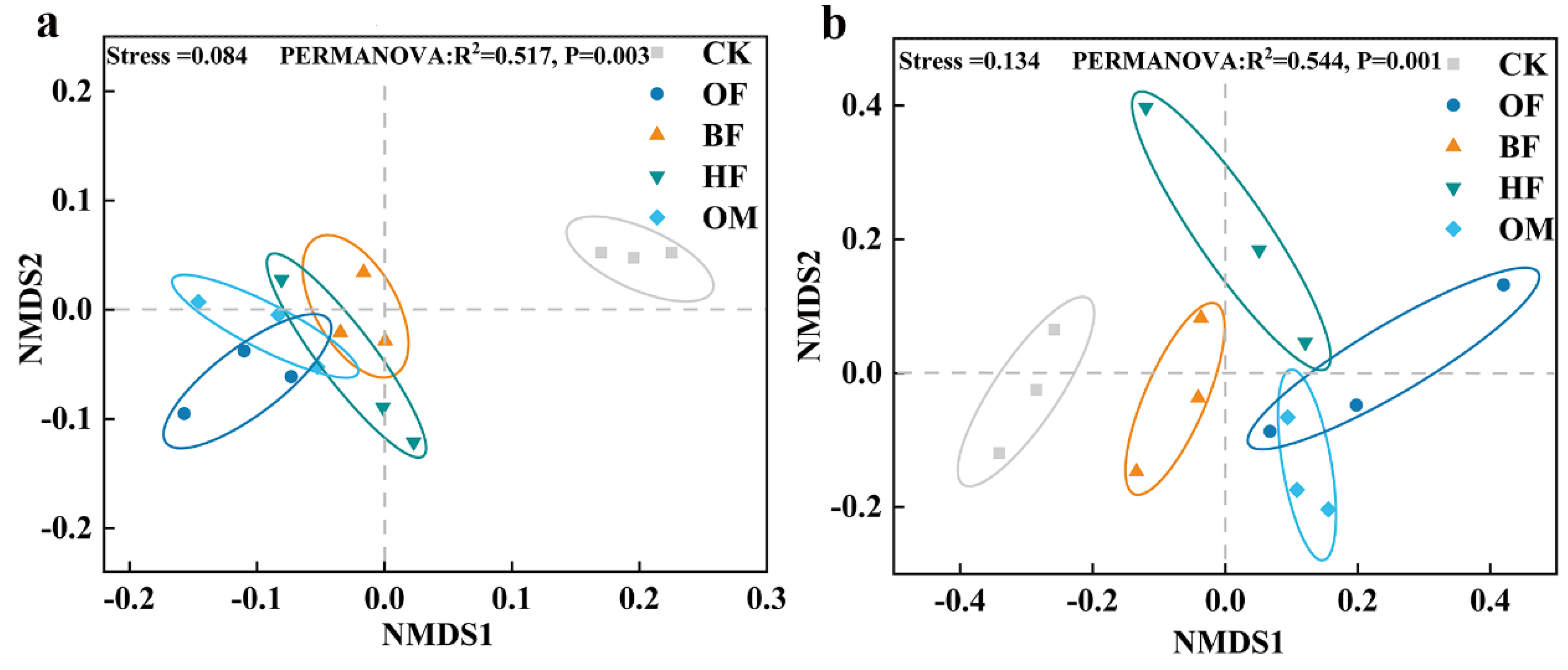
Figure 5.
Bacterial community composition at phylum level (>1%) (a) and at genus level (>1%) (c). Fungal community composition at phylum level (>1%) (b) and at genus level (>1%) (d). With a threshold value of 2.2, LEfSe determined the degree of divergence between various fertilizer treatments (e). CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi.
Figure 5.
Bacterial community composition at phylum level (>1%) (a) and at genus level (>1%) (c). Fungal community composition at phylum level (>1%) (b) and at genus level (>1%) (d). With a threshold value of 2.2, LEfSe determined the degree of divergence between various fertilizer treatments (e). CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi.
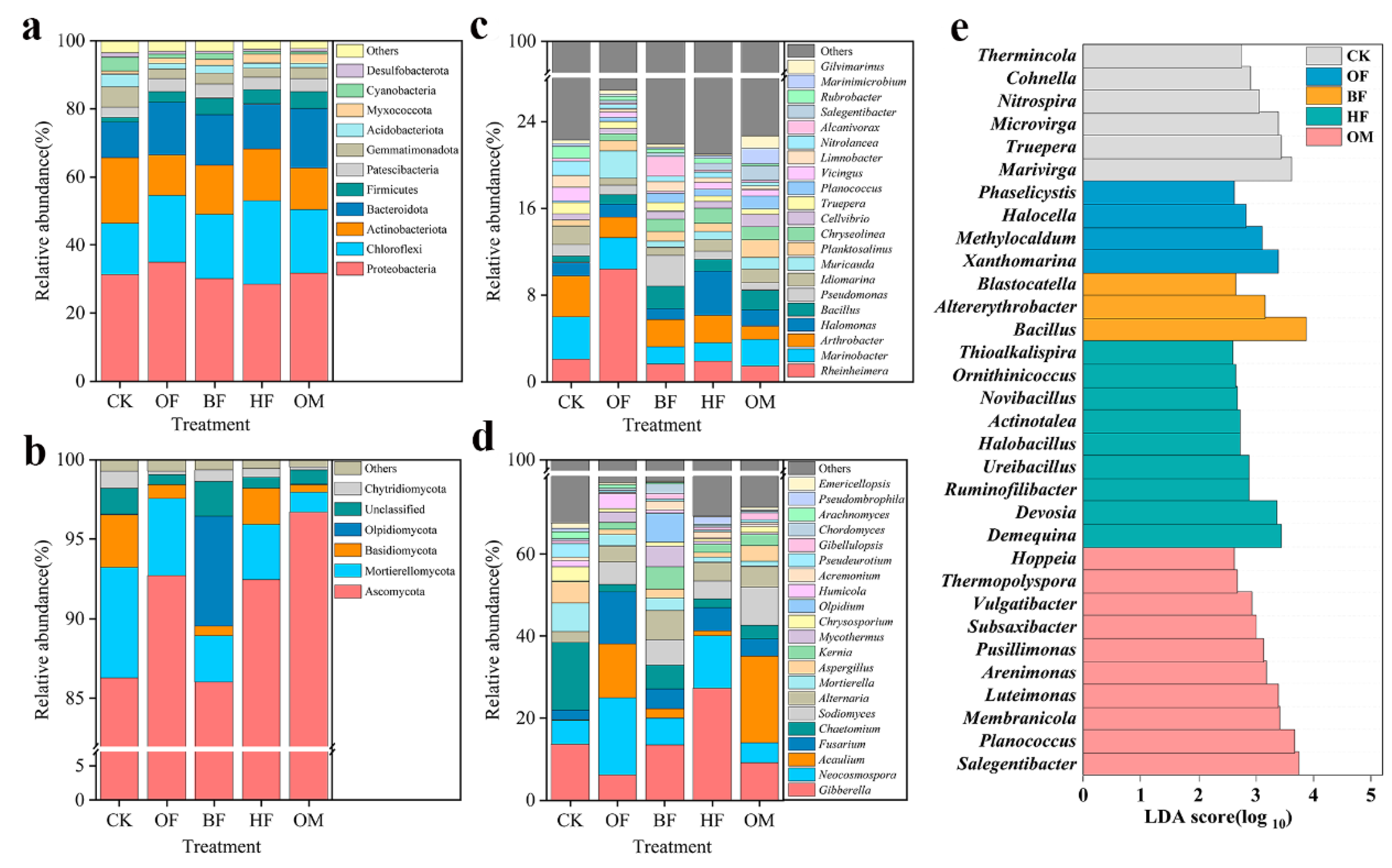
Figure 6.
Phylum level redundancy analysis (RDA) of bacterial (a) and fungal (b) community with environmental factors. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi.
Figure 6.
Phylum level redundancy analysis (RDA) of bacterial (a) and fungal (b) community with environmental factors. CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi.
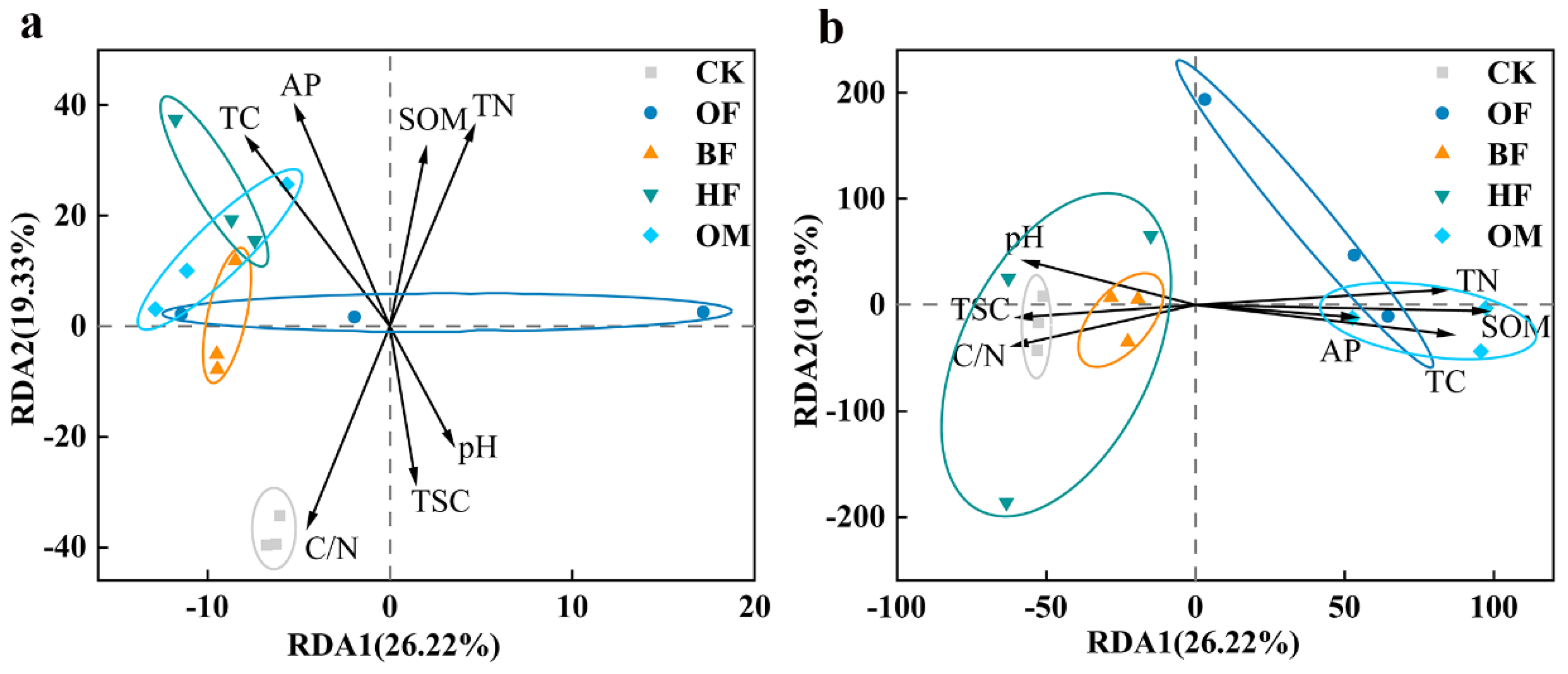
Figure 7.
Spearman correlation heatmaps between the top 25 most prevalent genera and the bacterial (a) and fungal (b) community compositions in soil at the genus level (* 0.01 < p ≤ 0.05, ** 0.001 < p ≤ 0.01, *** p ≤ 0.001).
Figure 7.
Spearman correlation heatmaps between the top 25 most prevalent genera and the bacterial (a) and fungal (b) community compositions in soil at the genus level (* 0.01 < p ≤ 0.05, ** 0.001 < p ≤ 0.01, *** p ≤ 0.001).
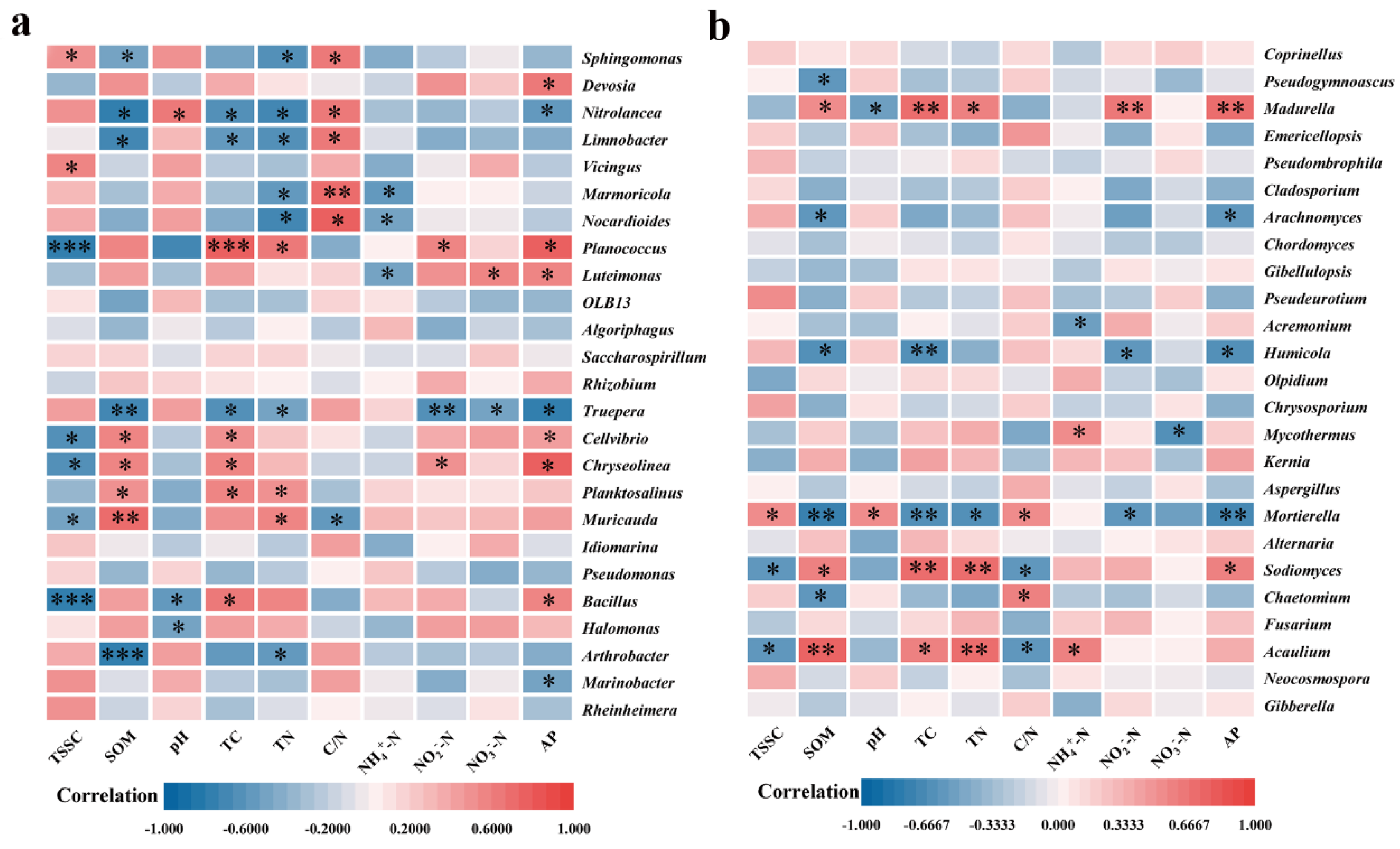
Figure 8.
Path Partial Least Squares Model (PLS-PM) showed the effects of several key factors (SOM, Physicochemical parameters, TSS content, Soil microorganism, Plant physiology) on the phytomass in saline soil (a). The route coefficients are shown by the numbers next to the arrows, and the blue and red arrows indicate adverse and favorable effects, respectively. The size of the route coefficient is also shown by the thickness of the arrows. Path coefficients and determination coefficients (R2) were generated after 999 bootstrap repeats were computed. Significance levels are denoted by * (p < 0.05), ** (p < 0.01), and *** (p < 0.001), respectively. Consequences, both direct and indirect, are standardized and obtained from the PLS-PM (b).
Figure 8.
Path Partial Least Squares Model (PLS-PM) showed the effects of several key factors (SOM, Physicochemical parameters, TSS content, Soil microorganism, Plant physiology) on the phytomass in saline soil (a). The route coefficients are shown by the numbers next to the arrows, and the blue and red arrows indicate adverse and favorable effects, respectively. The size of the route coefficient is also shown by the thickness of the arrows. Path coefficients and determination coefficients (R2) were generated after 999 bootstrap repeats were computed. Significance levels are denoted by * (p < 0.05), ** (p < 0.01), and *** (p < 0.001), respectively. Consequences, both direct and indirect, are standardized and obtained from the PLS-PM (b).
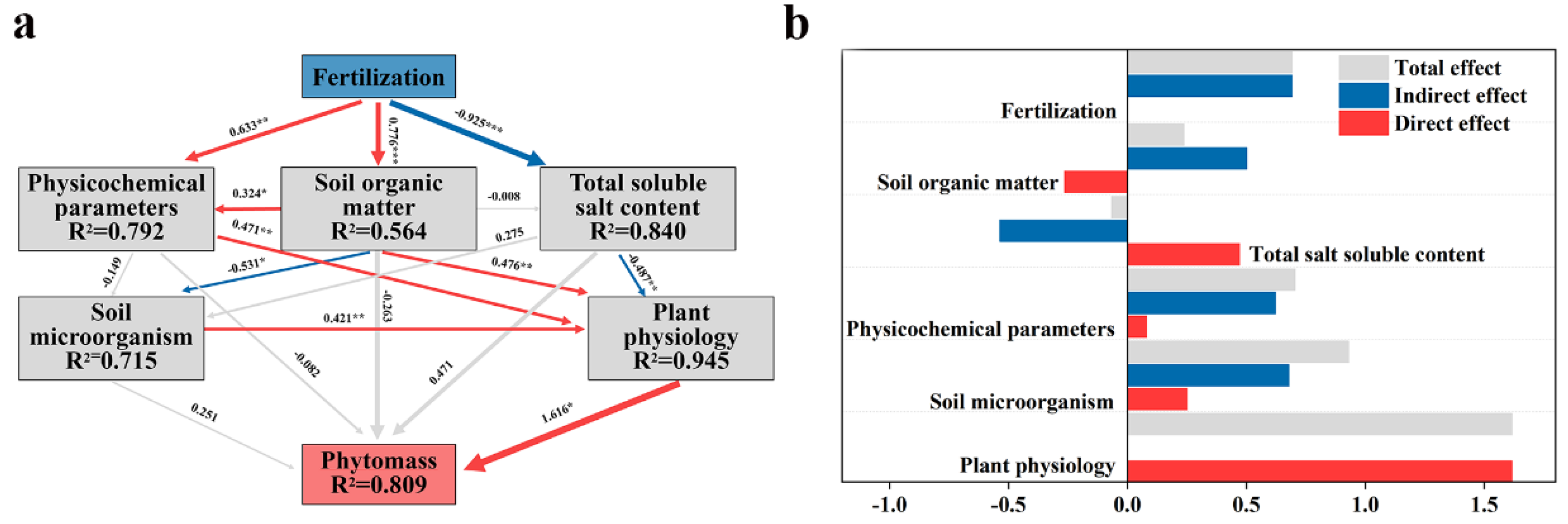

Table 1.
Microbial Diversity Index Table.
| Treatment | Chao index | Shannon index | Shannoneven index | |||
| Bacteria | Fungi | Bacteria | Fungi | Bacteria | Fungi | |
| CK | 2378±100 c | 415±7 a | 6.23±0.04 a | 4.09±0.14 a | 0.83±0.01 a | 0.06±0.01 a |
| OF | 2729±13 a | 394±25 a | 6.11±0.07 a | 3.07±0.48 a | 0.76±0.02 a | 0.03±0.01 a |
| BF | 2609±131 a | 458±24 a | 6.13±0.27 a | 3.65±0.10 a | 0.81±0.03 a | 0.04±0.01 a |
| HF | 2563±56 b | 410±75 a | 5.89±0.62 a | 2.90±0.73 a | 0.78±0.01 a | 0.02±0.01 a |
| OM | 2555±68 b | 407±23 a | 6.01±0.64 a | 3.23±0.13 a | 0.79±0.02 a | 0.03±0.01 a |
Notes: CK, control; OF, organic fertilizer; BF, organic fertilizer with B. licheniformis; HF, organic fertilizer with H. profundi. OM organic fertilizer with B. licheniformis and H. profundi. P-values were analyzed using Wilcoxon rank sum test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



