
Submitted:
14 November 2023
Posted:
14 November 2023
You are already at the latest version
Abstract
The regulation of intracellular pH in yeast (Saccharomyces cerevisiae) cells is critical for cell function and viability. In yeast, protons (H+) can be excreted from cell by the plasma membrane ATPase Pma1 and pumped into vacuoles by the vacuolar H+-ATPase. Because Pma1 is critical to the survival of yeast cells, it is unknown whether other compensatory components are involved in the pH homeostasis in the absence of Pma1. To elucidate how intracellular pH is regulated independently of Pma1, we employed a screening approach by exposing the yeast haploid deletion mutant library (ver 4.0) to the selective plant plasma membrane H+-ATPase inhibitor PS-1, which we previously reported. After repeated screenings and verification, we identified two proteins, ALY1 and ALY2, that play a role in the regulation of intracellular pH when Pma1 is deficient. Our research uncovers a new perspective on the regulation of intracellular pH related to Pma1 and also preliminarily reveals a role for ALY1 and ALY2 in the regulation of intracellular pH.
Keywords:
Saccharomyces cerevisiae
; plasma membrane H+-ATPase
; PMA1
; suppressor
; ALY1/2
1. Introduction
Proper cytosolic pH homeostasis is crucial for yeast survival, since protons (H+) in the cytosol act not only as facilitators for a series of cellular activities but also as potent second messengers in a variety of physiological programs, such as growth, development, and stress responses [1]. pH homeostasis is tightly related to nutrient signaling, and nutrient supply can regulate cytosolic pH homeostasis [2]. For instance, when glucose is added to the cultures with carbon-starved budding yeast (Saccharomyces cerevisiae) cells, the cytosol initially acidification before becoming more alkaline, which might be caused by glucose glycolysis leading to the release of H+ (acidification), followed by H+ extrusion by the plasma membrane H+-ATPase (Pma1) (alkalinization) [3]. Phosphate metabolism in yeast cells is interconnected with intracellular pH, since the uptake of inorganic phosphate, its subsequent intracellular distribution, and its conversion into polyphosphates (PolyP) are dependent on intracellular pH homeostasis; moreover, polyphosphate metabolism contributes to the regulation of cytosolic pH [4]. In addition to functions in nutrient signaling, cytosolic pH homeostasis also plays a role in protein trafficking along the yeast secretory pathway: aberrant cytosolic pH can lead to the mislocalization of plasma membrane proteins, such as Pma1 [5]. The yeast strain Debaryomyces hansenii is better able to regulate pH and exhibits a better ability to cope with NaCl stress than Saccharomyces cerevisiae [6].
Pma1 localized to the plasma membrane is the primary determinant of cytosolic pH by pumping cytoplasmic H+ out of the cell while also creating the necessary electrochemical gradient for H+-coupled nutrient solute uptake, making Pma1 an essential component of multiple cellular physiological activities in yeast cells [7]. Glucose stimulates Pma1 activity and is a carbon source for yeast growth [8]. Budding yeast grown in glucose-limited conditions displays decreased Pma1 activity and depolarized membrane potential with an acidic cytosolic environment, suggesting that Pma1 has an essential role in the coordination of cellular pH and metabolism [2, 9]. Pma1-mediated regulation of cytosolic pH also generates a potential second messenger of glucose availability in the form of H+ to conserve energy in the face of environmental stress [2]. In response to glucose supply, the C-terminal tail of Pma1 is a major site of regulation via phosphorylation on Ser/Thr residues, thereby alleviating the autoinhibition imposed by this region on Pma1 activity in glucose-starved yeast cells; the identity of the protein kinases that phosphorylate Pma1 is still not clear [2, 10]. Pma1 is also a key element involved in promoting target of rapamycin complex 1 (TORC1) activation in response to H+-coupled nutrient uptake [11], and plays an essential role in PolyP metabolism [12]. Besides nutrient signaling, Pma1 in budding yeast is also involved in various stress responses, such as exposure to weak acids, reactive oxygen species production, and ethanol treatment [13]. Studies of the replicative lifespan of budding yeast showed that endocytic genes such as vacuolar protein sorting 8 (Vps8), Vps9, and Vps21 are important for Pma1 accumulation in mother cells during cell division, although Pma1 accumulation in mother cells does not reflect the age of mother cells [14]. As Pma1 function is essential for yeast growth, pharmacological drugs have been designed for use in humans using Pma1 as an antifungal target [15].
Vacuolar H+-ATPase (V-ATPase) usually acts in close coordination with Pma1 to regulate cellular pH [1]. V-ATPase is a highly conserved H+ pump that translocates H+ from the cytosol to the vacuolar compartment, thereby regulating the luminal acidification of multiple biosynthetic and endocytic organelles in various cellular responses, such as vesicle trafficking and pH homeostasis [16]. V-ATPase acts interdependently with Pma1 in pH regulation and yeast growth [17-19]. Upon addition of glucose to glucose-deprived yeast cells, the transient cytosolic acidification and a subsequent alkanization of cytosolic pH is delayed in the mutant strains vacuolar membrane atpase3 (vma3Δ) and vacuolar pH1 (vph1Δ) lacking vacuolar H+-ATPase activity, which are defective in the regulation of vacuolar pH [20]. The ubiquitin ligase reverses Spt-phenotype5 (Rsp5) and the arrestin-related adaptor protein regulator of IME2 8 (Rim8) mediate the ubiquitination, internalization, and degradation of Pma1 in vma mutants to balance overall pH [18]. The cross-talk between V-ATPase and Pma1 offered a functional basis for exploring the compensatory downregulation of Pma1 in V-ATPase mutant strains, resulting in the isolation of the two phosphatases calcium-responsive calcineurin and glucose-sensitive phosphatase glycogen 7 (Glc7), which is essential for ubiquitination and endocytic downregulation of Pma1 when V-ATPase activity is compromised [19].
Although the compensatory ubiquitination, internalization, and degradation of Pma1 have been uncovered and largely studied in yeast cells when V-ATPase is defective, whether a similar compensatory mechanism exists in yeast cells lacking Pma1 activity is unknown. Here, we report on the identification of the two arrestin genes arrestin-like yeast protein1 (Aly1) and Aly2 based on their negative synthetic genetic interactions with Pma1 deficient mutants. We demonstrate that Aly1 and Aly2 serve as modulators of cytoplasmic pH when Pma1 is deficient in yeast.
2. Results
2.1. Screening for new components possibly involved in cytosolic pH homeostasis
Multiple physiological activities in yeast cells rely on the maintenance of cytosolic pH, in which Pma1 plays a major role by excreting H+ out of the cell. To identify components that might compensate for the loss of Pma1 function in cytosolic pH homeostasis, we looked for mutants that can sustain growth in the absence of Pma1 activity. We previously identified PS-1 as a small molecule inhibitor of plant plasma membrane (PM) H+-ATPase [22]. PS-1 might inhibit yeast Pma1, as the heterogeneous expression of a plant PM H+-ATPase gene in yeast cells was able to rescue growth of a yeast pma1 mutant [22]. As shown in Figure 1A, the control yeast strain SPJ1929 showed vigorous growth on glucose-containing medium; however, the addition of PS-1 to the medium gradually suppressed the growth of yeast cells in a dose-dependent manner, with 30 μM PS-1 having a substantial negative effect on yeast growth. This result indicates that the activity of yeast Pma1 can also be inhibited by PS-1, thus offering a means to screen for additional components involved in pH homeostasis in the absence of Pma1.
We performed a genome-wide rescue screen on a collection of single gene deletion yeast strains in the S. pombe haploid deletion mutant set ver 4.0 (M-4030H), which consists of 3,400 mutants. Specifically, we looked for mutants that could still grow when 30 μM PS-1 was added to the culture medium. We carried out a first screen by spotting 95 mutant strains and 1 control strain on one plate with a toothpick, each strain being spotted four times, for a total of 36 plates to screen the entire collection. In the first screen (Figure 1B), the growth of most of yeast mutants was suppressed by PS-1 to the same extent as the control strain SPJ1929 (indicated in the white square). However, some mutants appeared to show better growth than the control strain, suggesting that negative components might exist to regulate pH in the absence of Pma1 activity.
From the first screen, we obtained 115 mutants with some degree of rescue compared to the control strain SPJ1929 on medium containing 30 µM PS-1. To reduce the errors and narrow the screen range, we rescreened all 115 mutants in a second screen with 26 mutants screened simultaneously on one plate as eight spots from a toothpick. Some mutant strains showed better growth compared to the control strain SPJ1929 (indicated in the white square), although most mutant strains were comparable to the control strain, reducing the number of candidates to 33 mutants (Figure 1C). To quantitatively compare the growth rate of the yeast cells, we performed a third screen by spotting serial dilutions from liquid cultures onto medium containing 30 µM PS-1, yielding 24 mutant strains with reproducible better growth than the control strain (Figure 1D). PCR analysis of each mutant strain revealed that 3 mutant strains did not carry a deletion in their intended target gene, while the remaining 21 mutants did.
2.2. The loss of Aly1 and Aly2 function can rescue growth of yeast cells with reduced Pma1 activity
Pma1 is essential for yeast survival. To further verify the function of these screened genes in the regulation of pH homeostasis is independent of Pma1, we tested the function of these 21 genes in the budding yeast strain RS-72, which can grow on galactose-containing medium but not on glucose-containing medium, because expression of the endogenous Pma1 gene is placed under the control of the galactose-inducible galactokinase1 (GAL1) promoter (ProGAL:Pma1), providing an efficient means to selectively shut down Pma1 expression [23]. As the initial mutant screen was performed in Schizosaccharomyces pombe (fission yeast), we searched the RS-72 genome for genes homologous to the 21 candidate genes. We detected putative orthologs for 16 genes in Saccharomyces cerevisiae (budding yeast) (Table 1); the remaining 5 genes from fission yeast seemed to lack orthologs in budding yeast.
We deleted each of these 16 genes in the RS-72 background and assessed the resulting growth phenotypes when grown on glucose- or galactose-containing medium. We determined that one mutant with a deletion in Aly1 (aly1Δ) could grow well both on glucose- and on galactose-containing media, in contrast to RS-72, which only grew on medium containing galactose. By contrast, as a control which can only use glucose as a carbon source, the Saccharomyces cerevisiae strain AH109 only grew on glucose-containing medium (Figure 2A). This result suggests that the loss of Aly1 function can rescue yeast growth when Pma1 activity is diminished.
We also assessed the contribution of the Aly1 paralog Aly2 to pH regulation by deleting the gene in the RS-72 background. The aly2Δ yeast strain showed the same phenotype as the aly1Δ deletion strain, growing both on glucose- and on galactose-containing media (Figure 2B). We conclude that Aly1 and Aly2 participate in the regulation of yeast growth in a non-redundant manner.
We also individually deleted Aly1 or Aly2 in the yeast strain AH109 to investigate whether carbon source use was disturbed in aly1Δ and aly2Δ mutants. The aly1Δ and aly2Δ mutants grew on glucose-containing medium but not on galactose-containing medium (Figure 2C). This result suggests that the loss of Aly1 or Aly2 function does not disturb the use of carbon source, indicating that the rescued growth seen for aly1Δ and aly2Δ on glucose-containing medium in the RS-72 background was due to decreased Pma1 activity.
2.3. The loss of Aly1 or Aly2 function promotes H+ extrusion in yeast cells with reduced Pma1 activity
We looked for evidence of H+ extrusion in yeast cells using the pH indicator bromocresol purple, which changes color from purple to yellow upon acidification. We added the pH indicator to galactose- or glucose-containing medium, which turned purple at the beginning of the growth period (Figure 3A, B). When provided galactose, RS-72, aly1Δ, and aly2Δ spots acquired yellow halos around them, indicative of medium acidification and thus H+ extrusion. By contrast, RS-72 cells failed to grow on glucose-containing medium, whereas both aly1Δ and aly2Δ mutants displayed strong growth and H+ extrusion, as evidenced by the yellow color of the growth medium. This result suggests that the loss of Aly1 or Aly2 may helpful to alleviate the repression of other components participating in H+ extrusion besides Pma1.
As a more quantitative method, we measured growth medium pH over time in RS-72 and the aly1Δ and aly2Δ mutants grown in the presence of glucose or galactose. The pH of RS-72 cultures dropped by about 1.5 pH units within 12 h when grown in galactose-containing medium, with a more modest decrease of 0.6 pH units when grown in glucose-containing medium (Figure 3C, D). In contrast to RS-72, aly1Δ and aly2Δ cultures showed a comparable rate of medium acidification of 1–1.3 pH units regardless of carbon source, indicating that the loss of Aly1 and Aly2 function can promote H+ extrusion even in the absence of functional Pma1. The slower acidification rate of aly1Δ and aly2Δ mutants compared with RS-72 in galactose-containing medium might reflect their slower growth rate. These results suggest that Pma1-mediated H+ extrusion is essential for yeast growth; Aly1 and Aly2 play a negative role in H+ extrusion in Pma1-defective yeast strains, and the loss of Aly1 or Aly2 function can rescue H+ exclusion to promote yeast growth.
2.4. The loss of Aly1 or Aly2 function recovers cytosolic pH in yeast cells with reduced Pma1 activity
Although the loss of Aly1 and Aly2 function promotes H+ extrusion when Pma1 is absent, whether cytosolic pH changes in these mutants is unknown. We used the fluorescent-based cytosolic pH dye SNARF-AM to evaluate changes in intracellular pH by measuring the fluorescence intensity ratio between 580 nm and 640 nm in the cytosol. When indicated by a pseudocolor scale, a more acidified environment in the cell produces a greener color, while a more alkaline environment produces a bluer color (Figure 4A). The fluorescence intensity ratio of 580 nm/640 nm showed a linear relationship with cytosolic pH (Figure 4B).
We thus incubated RS-72, aly1Δ, and aly2Δ yeast cells grown in galactose-containing medium or glucose-containing medium with SNARF-AM. We established that cytosolic pH in RS-72 cells was lower when grown in glucose-containing medium than in galactose-containing medium. This result indicates an acidified environment when Pma1 is expressed and supports the idea that Pma1 mediates H+ extrusion during yeast growth. By contrast, aly1Δ and aly2Δ cells showed a more alkaline pH when grown in either glucose-containing medium or galactose-containing medium. This result also supports the idea that the regulation of cytosolic pH consists of other components besides Pma1, with Aly1 and Aly2 acting as negative regulators of cytosolic pH in the absence of Pma1.
3. Discussion
Aly1 and Aly2 belong to the α-arrestin family of proteins, which comprises 14 members in budding yeast. This family serves as trafficking adaptor proteins in the regulation of signal-induced PM protein endocytosis and intracellular sorting of nutrient permeases [24, 25]. α-arrestins are also named arrestin-related trafficking adaptors (ARTs); Aly1 is also called Art6, and Aly2 is also called Art3. ARTs possess PY motifs that can interact with the WW domain of the ubiquitin ligase Rsp5 to form various ART-Rsp5 complexes [26, 27]. ART-Rsp5 complexes play key roles in the regulation of the composition of PM proteins through endocytosis and recycling. For example, the resistance to O-dinitrobenzene 1 (Rod1)-Rsp5 complex regulates endocytosis and recycling of Jen1 (a yeast monocarboxylate transporter) in response to glucose addition and glucose removal, respectively [28]. The regulation of the ART-Rsp5 is complex and coordinated, as is that of the endocytosis of amino acid transporters (AATs), which is activated by the Art2-Rsp5 complex through interaction between the basic patch of Art2 and the C-terminal acidic sorting motifs of AATs during amino acid starvation and also activated by the Art1-Rsp5 complex through the interaction of Art1 and the N-terminal acidic sorting motifs of the same AATs under amino acid-replete conditions [29].
Aly2 localizes to both the PM and the internal compartments of endosomes and trans-Golgi network (TGN), and its paralog Aly1 localizes to the internal compartments. Aly1 and Aly2 regulate PM endocytosis and intracellular trafficking in response to various cues. Both proteins can bind directly to the y-subunit of the clathrin adaptor AP-1, Apl4, and regulate the intracellular sorting of Gap1, a general amino acid permease, thus mediating Gap1 trafficking from the TGN to the endosome/vacuole under nitrogen-replete conditions and Gap1 recycling from endosomal compartments back to the TGN and/or the PM to under nitrogen-starvation conditions [27]. Aly1 and Aly2 mediate the endocytosis of Dip5 (dicarboxylic amino acid permease 5), an aspartic acid/glutamic acid transporter [24]. The dephosphorylation of Aly1 by calcineurin is required for the internalization of Dip5 from the PM to the vacuole but not for the intracellular sorting of Gap1 [25]. Besides nutrient transporters, Aly2 is also involved in the endocytosis of Acr3, an arsenite and antimonite transporter, to promote Acr3 proteolysis in the vacuole [30]. Aly1 and Aly2 can also promote K+ inwardly rectifying channel in mammals (Kir2.1) trafficking to the PM and enhance intracellular potassium levels in a yeast model [31]. In our study, the loss of Aly1 or Aly2 function caused a rise in intracellular pH and rescued yeast growth when Pma1 was deficient. The protein composition at the PM is tightly regulated in response to various physiological environments to maintain intracellular homeostasis of ions, nutrients, and pH. Although the regulatory role of Aly1 and Aly2 was partially uncovered here, their detailed function and mechanism in pH homeostasis regulation is still unknown. Whether the increased endocytosis of nutrient transporters from the PM or the decreased recycling of these nutrient transporters back to the PM is affected by the loss of Aly1 and Aly2 needs investigation.
Lower intracellular pH can regulate phospholipid metabolism when yeast cells are cultured in glucose-starvation conditions, which is accomplished by the binding of a pH biosensor, phosphatidic acid (PA), to overproducer of inositol 1 (Opi1) to further repress phospholipid metabolic genes [32]. Besides intracellular pH, Aly1 and Aly2 may also participate in the regulation of lipid metabolism. Aly1 and Aly2 regulate the trafficking of glycerophosphoinositol 1 (Git1), a transporter for glycerophosphoinositol (GPI), and the loss of Aly1 and Aly2 function might influence the phosphoinositol (PI) balance, such as by increasing phosphatidylinositol-3-phosphate (PI3P) on the limiting membrane of the vacuole [33]. A change in the composition of membrane lipids might influence the trafficking of membrane proteins. Some nutrient proteins are clustered at the PM in sphingolipid- and ergosterol-rich membrane compartments occupied by Can1 (MCCs) and respond to their substrates by conformational changes and lateral movement from the MCCs to surrounding lipid domains, leading to their endocytosis [26]. How the composition of lipids might have changed following the loss of Aly1 or Aly2 function in the absence of Pma1, and whether the loss of Aly1 or Aly2 function rescues intracellular pH homeostasis through regulating the lipid balance, need further study.
Since Pma1 and V-ATPase act interdependently in the regulation of intracellular pH, V-ATPase is thought to be regulated to balance intracellular pH. When V-ATPase is deficient, Pma1 is degraded through the Rim8-Rsp5 complex to balance intracellular pH [18]. However, whether and how vacuolar H+-ATPase is regulated to balance intracellular pH when Pma1 is defective has not been reported yet. Pma1 and V-ATPase act coordinately in response to various cues, such as glucose-replete or -deprivation conditions. When glucose supply is sufficient, intracellular pH decreases, which leads to the activation of Pma1 to pump protons out of the cell but also leads to the activation of V-ATPase through the assembly of the V1 and Vo subunits to pump protons into the vacuole [7]. However, when glucose is depleted, Pma1 becomes inactive and the V1 and Vo subunits of V-ATPase separate to decrease V-ATPase activity [7]. The activation of V-ATPase is dependent on external pH, as rapid vacuole acidification can be observed at external pH 5.0 while alkalinization at external pH 7.0 [9]. In our study, the deficient function of Pma1 resulted in a lower pH in the cytosol; whether V-ATPase activity can be stimulated by the observed lower intracellular pH in the absence of Pma1 function is unknown. 3,5-bisphosphate (PI(3,5)P2) plays a critical role in activating V-ATPase activity in response to salt stress [34], and the loss of Aly1 or Aly2 function results in increased accumulation of PI3P [33], which can be converted to PI(3,5)P2. These studies offer a clue that the loss of Aly1 or Aly2 function in cells lacking Pma1 activity might partially rescue the intracellular pH homeostasis by stimulating V-ATPase activity through the modulation of lipid composition.
4. Materials and Methods
4.1. Yeast strains
The library of haploid deletion mutants (ver 4.0) and SPJ1929 (h+, leu1-32, ura4-D18, ade6-216) strains are in fission yeast (Schizosaccharomyces pombe). The AH109 and RS-72 (MATa, ade1-100, his4-519, leu2-3, 112 and ura3-52) strains are Saccharomyces cerevisiae strains. In yeast strain RS-72, the gene encoding endogenous yeast H+ pump, Pma1, is under the control of the GAL1 promoter, making yeast viable only when grown on a medium with galactose as a carbon source, as previously reported [21]. Strains were grown in rich medium (YPD) containing 1% (w/v) yeast extract, 2% (w/v) peptone, and 2% (w/v) glucose (or 2% [w/v] galactose where specifically indicated).
4.2. Aly1 and Aly2 deletion assay
The full-length coding sequences of Aly1 and Aly2 genes in the yeast strains RS-72 and AH109 were replaced by the coding sequence of the hygromycin resistance gene (HygR).
4.3. Suppressor screening assay
In the first screen, there are 95 mutant strains and 1 control strain on one plate were dipped with a toothpick, each strain was spotted for four times, for a total of 36 plates to screen the entire collection. In the second screen, each mutant strain and 1 control strain were dipped on the medium for eight times. In the third screen, each strain was spotted serial dilutions from liquid cultures onto the medium.
4.4. SNARF-AM staining assay
Yeast cells were collected by centrifugation and washed in ddH2O twice before being incubated with 10 μM SNARF-AM for 1 h. The yeast cells in Figure 4A were incubated with 0.2% (v/v) Triton X100 and 10 μM SNARF-AM in a buffer of the indicated pH. The fluorescent signals were visualized with a Nikon CSU W1 confocal microscope at 488 nm excitation with 580 nm and 640 nm emission individually. The SNARF-AM dye was purchased from Thermo Fisher, more details can visit. https://www.thermofisher.cn/cn/zh/home.html.
Author Contributions
GY.L. and YQ.Y. designed the study and analyzed the data. GY.L. performed the experiments. GY.L. and XL.H. wrote the paper.
Funding
This work was supported by the National Key R&D Program of China (2022YFF1001601), and supported by the National Natural Science Foundation of China (32070301).
Declaration of interests
The authors declare no competing interests.
References
- Deprez, M.A., Eskes, E., Wilms, T., Ludovico, P., and Winderickx, J. (2018). pH homeostasis links the nutrient sensing PKA/TORC1/Sch9 ménage-à-trois to stress tolerance and longevity. Microbial cell 5, 119-136.
- Isom, D.G., Page, S.C., Collins, L.B., Kapolka, N.J., Taghon, G.J., and Dohlman, H.G. (2018). Coordinated regulation of intracellular pH by two glucose-sensing pathways in yeast. J BIOL CHEM 293, 2318-2329. [CrossRef]
- Ramos, S., Balbín, M., Raposo, M., Valle, E., and Pardo, L.A. (1989). The mechanism of intracellular acidification induced by glucose in Saccharomyces cerevisiae. Journal of general microbiology 135, 2413-2422. [CrossRef]
- Eskes, E., Deprez, M.A., Wilms, T., and Winderickx, J. (2018). pH homeostasis in yeast; the phosphate perspective. CURR GENET 64, 155-161. [CrossRef]
- Huang, C., and Chang, A. (2011). pH-dependent cargo sorting from the Golgi. The J BIOL CHEM 286, 10058-10065.
- Mortensen, H.D., Gori, K., Siegumfeldt, H., Nissen, P., Jespersen, L., and Arneborg, N. (2006). Intracellular pH homeostasis plays a role in the NaCl tolerance of Debaryomyces hansenii strains. Appl. Microbiol. Biotechnol. 71, 713-719. [CrossRef]
- Kane, P.M. (2016). Proton Transport and pH Control in Fungi. Adv. Exp. Med. Biol. 892, 33-68.
- Serrano, R. (1983). In vivo glucose activation of the yeast plasma membrane ATPase. FEBS LETT 156, 11-14. [CrossRef]
- Ma, T.Y., Deprez, M.A., Callewaert, G., and Winderickx, J. (2021). Coordinated glucose-induced Ca2+ and pH responses in yeast Saccharomyces cerevisiae. Cell calcium 100, 102479. [CrossRef]
- Lecchi, S., Nelson, C.J., Allen, K.E., Swaney, D.L., Thompson, K.L., Coon, J.J., Sussman, M.R., and Slayman, C.W. (2007). Tandem phosphorylation of Ser-911 and Thr-912 at the C terminus of yeast plasma membrane H+-ATPase leads to glucose-dependent activation. J BIOL CHEM 282, 35471-35481. [CrossRef]
- Saliba, E., Evangelinos, M., Gournas, C., Corrillon, F., Georis, I., and André, B. (2018). The yeast H+-ATPase Pma1 promotes Rag/Gtr-dependent TORC1 activation in response to H+-coupled nutrient uptake. ELIFE 7.
- Tomashevsky, A.A., and Petrov, V.V. (2022). Point mutations in the different domains of the Saccharomyces cerevisiae plasma membrane PMA1 ATPase cause redistribution among fractions of inorganic polyphosphates. J Biomol Structure Dyn 40, 635-647. [CrossRef]
- Lee, Y., Nasution, O., Lee, Y.M., Kim, E., Choi, W., and Kim, W. (2017). Overexpression of PMA1 enhances tolerance to various types of stress and constitutively activates the SAPK pathways in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 101, 229-239. [CrossRef]
- Yoon, S.Y., Jang, E., Ko, N., Kim, M., Kim, S.Y., Moon, Y., Nam, J.S., Lee, S., and Jun, Y. (2022). A Genome-Wide Screen Reveals That Endocytic Genes Are Important for Pma1p Asymmetry during Cell Division in Saccharomyces cerevisiae. Int. J. Mol. Sci. 23. [CrossRef]
- Tung, T.T., and Nielsen, J. (2022). Drug Discovery and Development on Pma1, Where Are We Now? A Critical Review from 1995 to 2022. CHEMMEDCHEM 17, e202200356.
- Wilms, T., Swinnen, E., Eskes, E., Dolz-Edo, L., Uwineza, A., Van Essche, R., Rosseels, J., Zabrocki, P., Cameroni, E., Franssens, V., et al. (2017). The yeast protein kinase Sch9 adjusts V-ATPase assembly/disassembly to control pH homeostasis and longevity in response to glucose availability. PLoS Genet. 13, e1006835. [CrossRef]
- Martínez-Muñoz, G.A., and Kane, P. (2008). Vacuolar and plasma membrane proton pumps collaborate to achieve cytosolic pH homeostasis in yeast. J BIOL CHEM 283, 20309-20319. [CrossRef]
- Smardon, A.M., and Kane, P.M. (2014). Loss of vacuolar H+-ATPase activity in organelles signals ubiquitination and endocytosis of the yeast plasma membrane proton pump Pma1p. J BIOL CHEM 289, 32316-32326. [CrossRef]
- Velivela, S.D., and Kane, P.M. (2018). Compensatory Internalization of Pma1 in V-ATPase Mutants in Saccharomyces cerevisiae Requires Calcium- and Glucose-Sensitive Phosphatases. GENETICS 208, 655-672. [CrossRef]
- Tarsio, M., Zheng, H., Smardon, A.M., Martínez-Muñoz, G.A., and Kane, P.M. (2011). Consequences of loss of Vph1 protein-containing vacuolar ATPases (V-ATPases) for overall cellular pH homeostasis. J BIOL CHEM 286, 28089-28096. [CrossRef]
- Cid, A., Perona, R., and Serrano, R. (1987). Replacement of the promoter of the yeast plasma membrane ATPase gene by a galactose-dependent promoter and its physiological consequences. CURR GENET 12, 105-110. [CrossRef]
- Yang, Y., Liu, X., Guo, W., Liu, W., Shao, W., Zhao, J., Li, J., Dong, Q., Ma, L., He, Q., et al. (2022). Testing the polar auxin transport model with a selective plasma membrane H+-ATPase inhibitor. J Integr Plant Biol 64, 1229-1245. [CrossRef]
- Palmgren, M.G., and Christensen, G. (1994). Functional comparisons between plant plasma membrane H+-ATPase isoforms expressed in yeast. J BIOL CHEM 269, 3027-3033. [CrossRef]
- Hatakeyama, R., Kamiya, M., Takahara, T., and Maeda, T. (2010). Endocytosis of the aspartic acid/glutamic acid transporter Dip5 is triggered by substrate-dependent recruitment of the Rsp5 ubiquitin ligase via the arrestin-like protein Aly2. Mol. Cell. Biol. 30, 5598-5607. [CrossRef]
- O'Donnell, A.F., Huang, L., Thorner, J., and Cyert, M.S. (2013). A calcineurin-dependent switch controls the trafficking function of α-arrestin Aly1/Art6. J BIOL CHEM 288, 24063-24080.
- Kahlhofer, J., Leon, S., Teis, D., and Schmidt, O. (2021). The α-arrestin family of ubiquitin ligase adaptors links metabolism with selective endocytosis. Biol. Cell. 113, 183-219. [CrossRef]
- O'Donnell, A.F., Apffel, A., Gardner, R.G., and Cyert, M.S. (2010). Alpha-arrestins Aly1 and Aly2 regulate intracellular trafficking in response to nutrient signaling. Mol. Biol. Cell 21, 3552-3566.
- Becuwe, M., and Léon, S. (2014). Integrated control of transporter endocytosis and recycling by the arrestin-related protein Rod1 and the ubiquitin ligase Rsp5. ELIFE 3.
- Ivashov, V., Zimmer, J., Schwabl, S., Kahlhofer, J., Weys, S., Gstir, R., Jakschitz, T., Kremser, L., Bonn, G.K., Lindner, H., et al. (2020). Complementary α-arrestin-ubiquitin ligase complexes control nutrient transporter endocytosis in response to amino acids. ELIFE 9.
- Wawrzycka, D., Sadlak, J., Maciaszczyk-Dziubinska, E., and Wysocki, R. (2019). Rsp5-dependent endocytosis and degradation of the arsenite transporter Acr3 requires its N-terminal acidic tail as an endocytic sorting signal and arrestin-related ubiquitin-ligase adaptors. BBA-BIOMEMBRANES 1861, 916-925. [CrossRef]
- Hager, N.A., Krasowski, C.J., Mackie, T.D., Kolb, A.R., Needham, P.G., Augustine, A.A., Dempsey, A., Szent-Gyorgyi, C., Bruchez, M.P., Bain, D.J., et al. (2018). Select α-arrestins control cell-surface abundance of the mammalian Kir2.1 potassium channel in a yeast model. J BIOL CHEM 293, 11006-11021.
- Young, B.P., Shin, J.J., Orij, R., Chao, J.T., Li, S.C., Guan, X.L., Khong, A., Jan, E., Wenk, M.R., Prinz, W.A., et al. (2010). Phosphatidic acid is a pH biosensor that links membrane biogenesis to metabolism. Science 329, 1085-1088. [CrossRef]
- Robinson, B.P., Hawbaker, S., Chiang, A., Jordahl, E.M., Anaokar, S., Nikiforov, A., Bowman, R.W., 2nd, Ziegler, P., McAtee, C.K., Patton-Vogt, J., et al. (2022). Alpha-arrestins Aly1/Art6 and Aly2/Art3 regulate trafficking of the glycerophosphoinositol transporter Git1 and impact phospholipid homeostasis. Biol. Cell 114, 3-31.
- Li, S.C., Diakov, T.T., Xu, T., Tarsio, M., Zhu, W., Couoh-Cardel, S., Weisman, L.S., and Kane, P.M. (2014). The signaling lipid PI(3,5)P₂ stabilizes V₁-V(o) sector interactions and activates the V-ATPase. Mol. Biol. Cell 25, 1251-1262.
Figure 1.
Screening for suppressors that can grow on medium containing PS-1. (A) Growth of the fission yeast strain SPJ1929 is inhibited by PS-1. (B) Representative photograph of a plate from the first screen for suppressors with better growth than the control on medium containing PS-1. (C) Representative photograph of a plate from the second screen for suppressors. (D) Representative photograph of a plate from the third screen for suppressors.
Figure 1.
Screening for suppressors that can grow on medium containing PS-1. (A) Growth of the fission yeast strain SPJ1929 is inhibited by PS-1. (B) Representative photograph of a plate from the first screen for suppressors with better growth than the control on medium containing PS-1. (C) Representative photograph of a plate from the second screen for suppressors. (D) Representative photograph of a plate from the third screen for suppressors.
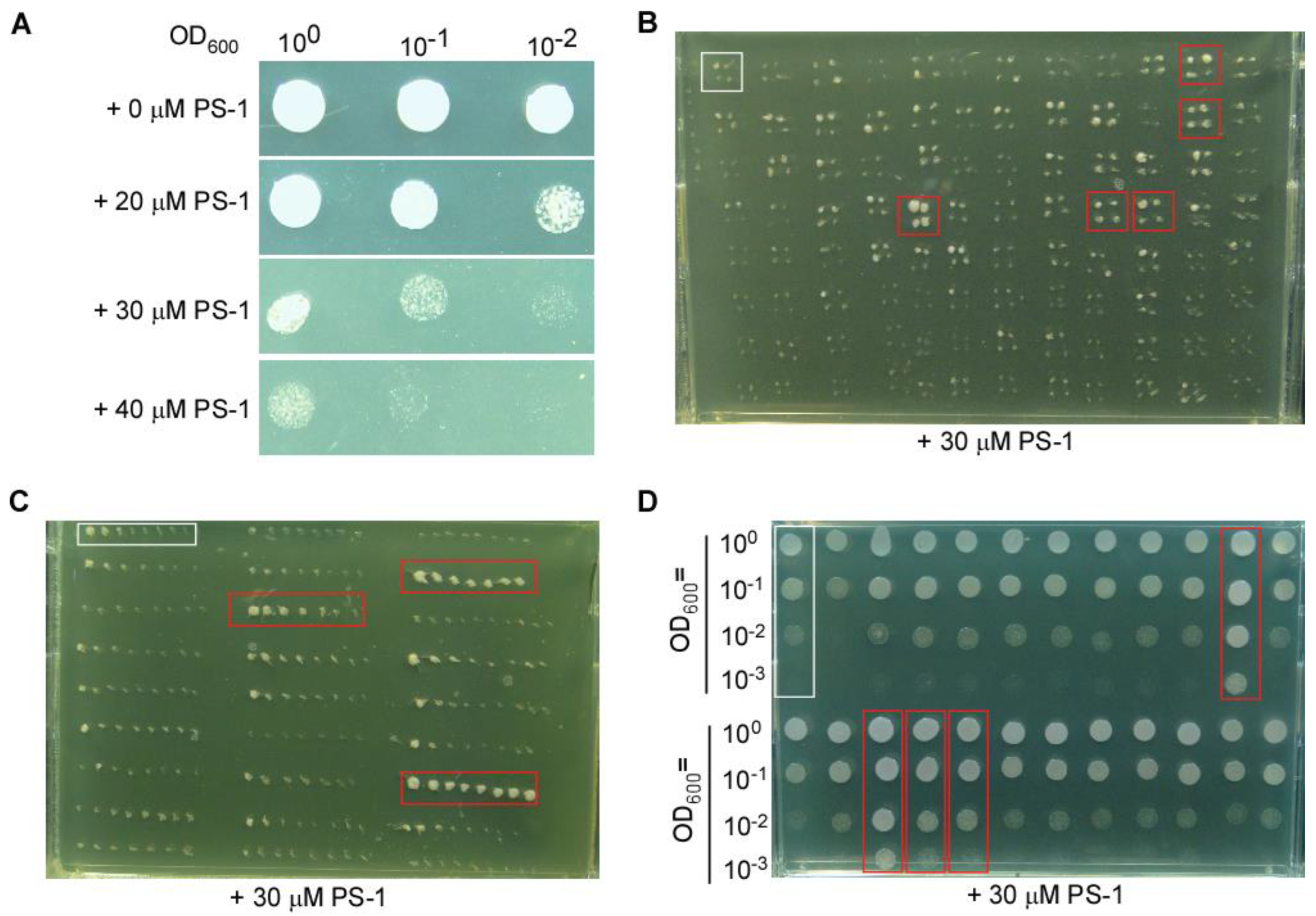
Figure 2.
The loss of Aly1 or Aly2 function can rescue yeast growth in Pma1-deficient strains. (A) Deletion of Aly1 in the budding yeast strain RS-72 can rescue growth when a reduction in Pma1 expression due to grow on the medium with glucose as the sole carbon source. (B) Deletion of Aly2 in the budding yeast strain RS-72 can rescue growth when a reduction in Pma1 expression due to grow on the medium with glucose as the sole carbon source. (C) Deletion of Aly1 or Aly2 in the budding yeast strain AH109 strain cannot rescue yeast growth on medium with galactose as the sole carbon source.
Figure 2.
The loss of Aly1 or Aly2 function can rescue yeast growth in Pma1-deficient strains. (A) Deletion of Aly1 in the budding yeast strain RS-72 can rescue growth when a reduction in Pma1 expression due to grow on the medium with glucose as the sole carbon source. (B) Deletion of Aly2 in the budding yeast strain RS-72 can rescue growth when a reduction in Pma1 expression due to grow on the medium with glucose as the sole carbon source. (C) Deletion of Aly1 or Aly2 in the budding yeast strain AH109 strain cannot rescue yeast growth on medium with galactose as the sole carbon source.
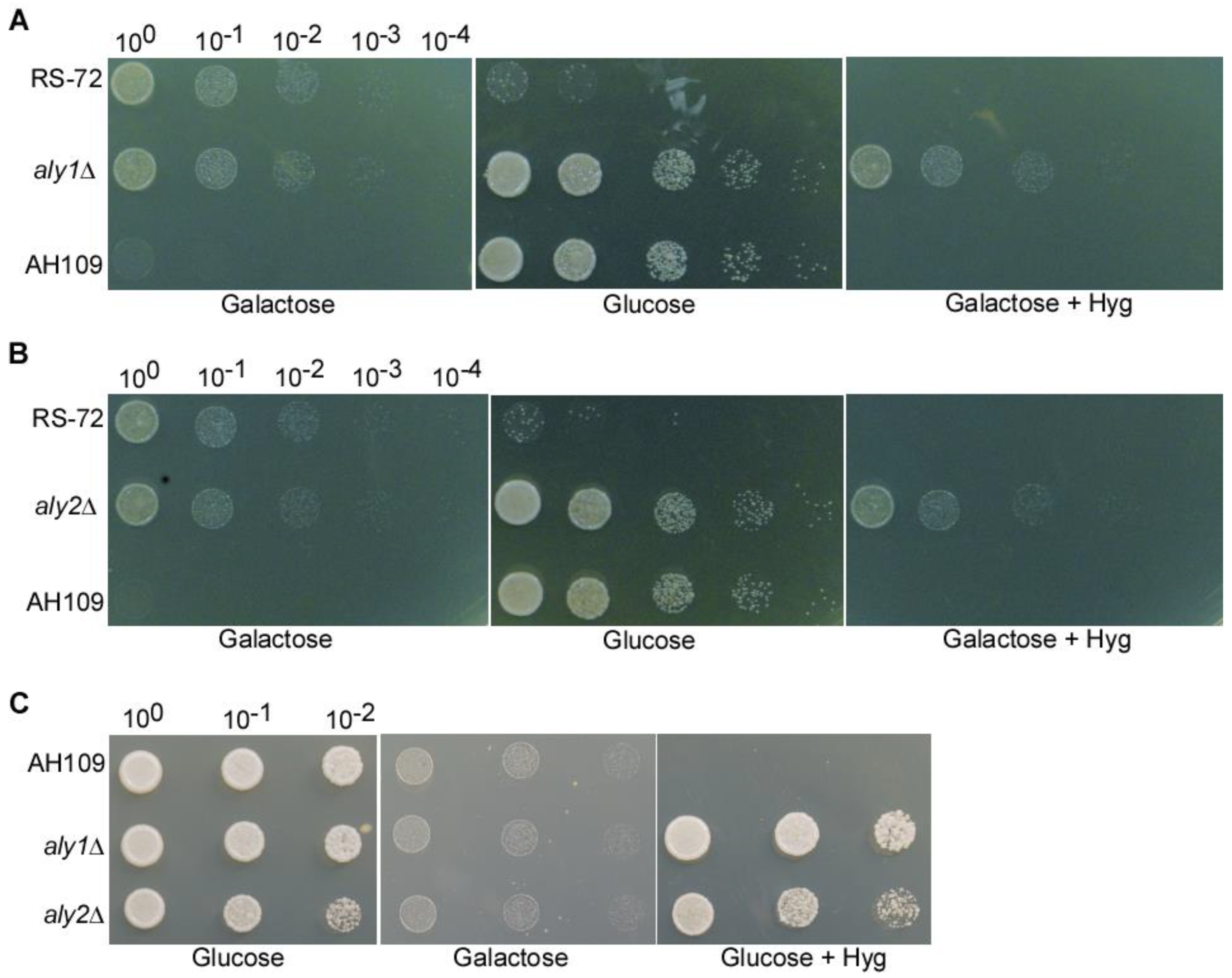
Figure 3.
The loss of Aly1 or Aly2 function promotes H+ extrusion in yeast cells with reduced Pma1 activity. (A and B) Visualization of the acidification of growth medium using the pH-sensitive dye bromocresol purple. (C and D) Acidification of the yeast growth medium over time in aly1 (C) or aly2 (D) deletion strains, values were taken from 3 independent repeated experiments, Error bars, means ± SD.
Figure 3.
The loss of Aly1 or Aly2 function promotes H+ extrusion in yeast cells with reduced Pma1 activity. (A and B) Visualization of the acidification of growth medium using the pH-sensitive dye bromocresol purple. (C and D) Acidification of the yeast growth medium over time in aly1 (C) or aly2 (D) deletion strains, values were taken from 3 independent repeated experiments, Error bars, means ± SD.
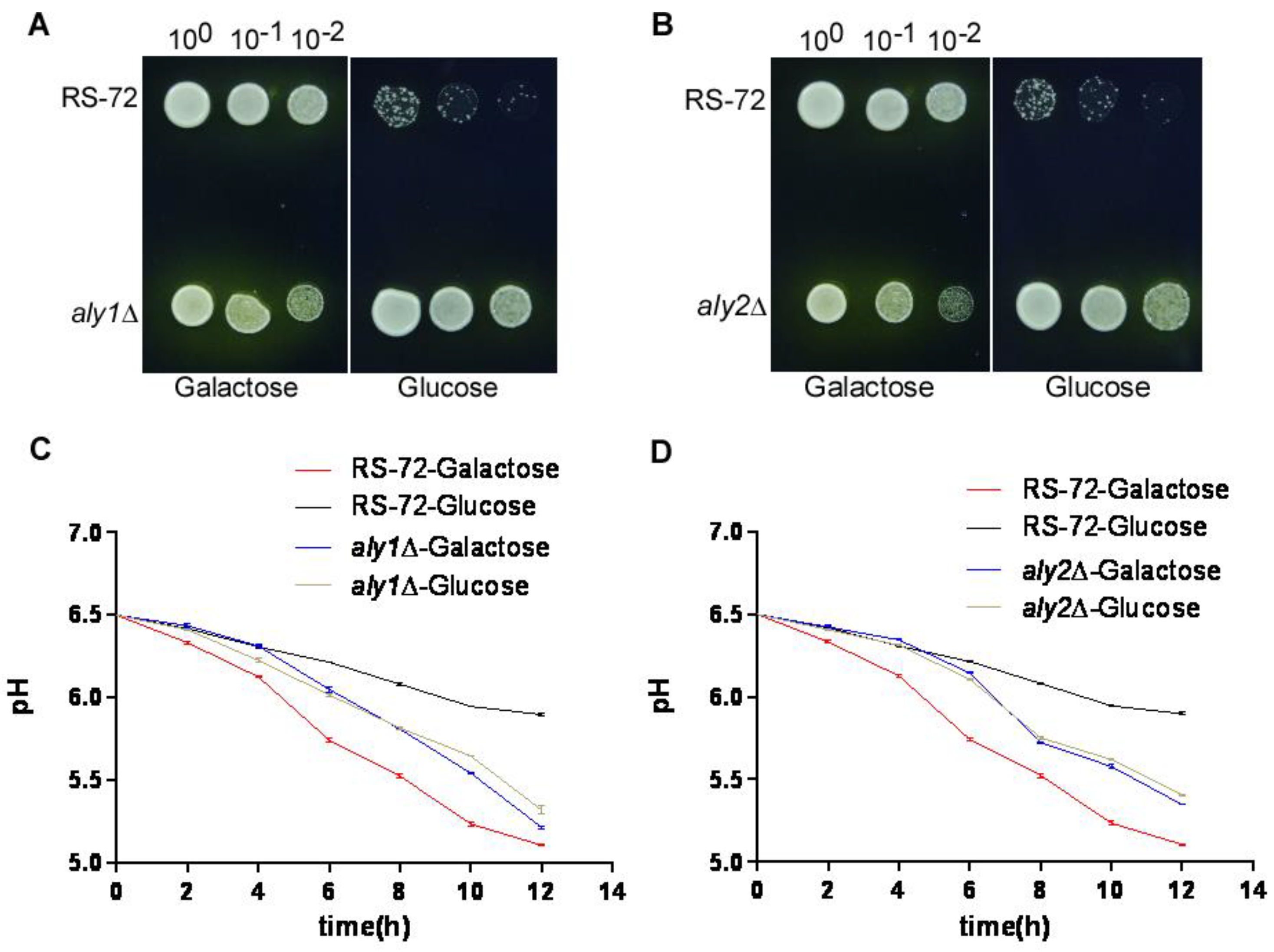
Figure 4.
The loss of Aly1 or Aly2 function recovers cytosolic pH in yeast cells with reduced Pma1 activity. (A) Representative confocal images of yeast cells stained with SNARF-AM in a buffer containing Triton X100 with different pH values. (B) Standard curve of fluorescence intensity ratio of 580 nm/640 nm as a function of pH for the cells shown in (A), values were taken from 3 independent repeated experiments. Error bars, means ± SD. (C) Representative confocal images indicating the pH value in the cytosol of RS-72, aly1 and aly2 yeast cells.table.
Figure 4.
The loss of Aly1 or Aly2 function recovers cytosolic pH in yeast cells with reduced Pma1 activity. (A) Representative confocal images of yeast cells stained with SNARF-AM in a buffer containing Triton X100 with different pH values. (B) Standard curve of fluorescence intensity ratio of 580 nm/640 nm as a function of pH for the cells shown in (A), values were taken from 3 independent repeated experiments. Error bars, means ± SD. (C) Representative confocal images indicating the pH value in the cytosol of RS-72, aly1 and aly2 yeast cells.table.
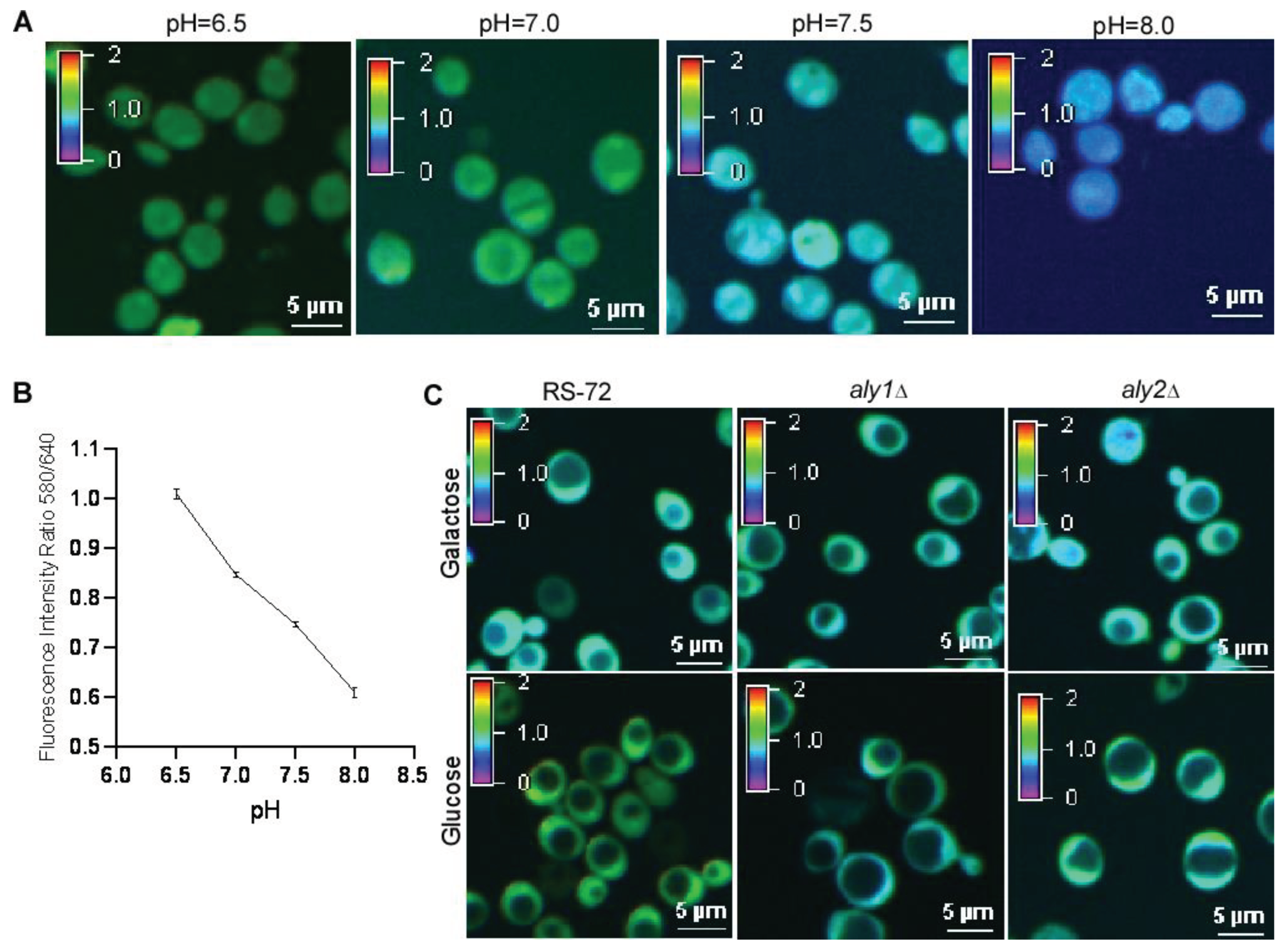

Table 1.
Suppressor of Pma1 in Schizosaccharomyces pombe (fission yeast) and the ortholog in Saccharomyces cerevisiae (budding yeast).
Table 1.
Suppressor of Pma1 in Schizosaccharomyces pombe (fission yeast) and the ortholog in Saccharomyces cerevisiae (budding yeast).
| NO. | Ver4.0 position | Systemic ID | ortholog in budding yeast | Gene description |
|---|---|---|---|---|
| 1 | V4-P02-12(A12) | SPAC12G12.13c | YNL299W|YOL115W | poly(A) polymerase Cid14 |
| 2 | V4-P02-54(E6) | SPAC13G7.11 | YBR185C | mitochondrial respiratory complex assembly protein (predicted) |
| 3 | V4-P04-33(C9) | SPAC17G8.13c | YBL052C | histone acetyltransferase Mst2 |
| 4 | V4-P04-46(D10) | SPAC1805.01c | YAL017W|YOL045W | serine/threonine protein kinase Ppk6 (predicted) |
| 5 | V4-P06-75(G3) | SPAC22H10.09 | NONE | sequence orphan |
| 6 | V4-P07-62(F2) | SPAC25B8.18 | YKR049C | mitochondrial electron carrier (predicted) |
| 7 | V4-P12-04(A4) | SPAC6G10.08 | YDL066W|YLR174W|YNL009W | isocitrate dehydrogenase Idp1 (predicted) |
| 8 | V4-P13-47(D11) | SPACUNK4.12c | YLR389C | metallopeptidase (predicted) |
| 9 | V4-P17-74(G2) | SPBC18H10.06c | YKL018W | Set1C complex subunit Swd2.1 |
| 10 | V4-P18-15(B3) | SPBC19G7.04 | NONE | HMG box protein |
| 11 | V4-P19-85(H1) | SPBC2D10.04 | YJL084C|YKR021W | arrestin Aly1 related, implicated in endocytosis |
| 12 | V4-P22-41(D5) | SPBC609.02 | YNL128W | phosphatidylinositol-3,4,5-trisphosphate3-phosphatasePtn1 |
| 13 | V4-P25-06(A6) | SPCC1235.09 | YBR103W | Set3 complex subunit Hif2 |
| 14 | V4-P26-63(F3) | SPCC1906.02c | YGL110C | CUE domain protein Cue3 (predicted) |
| 15 | V4-P27-17(B5) | SPCC31H12.05c | YER133W | serine/threonine protein phosphatase Sds21 |
| 16 | V4-P28-81(G9) | SPCC794.10 | YHL012W|YKL035W | UTP-glucose-1-phosphate uridylyltransferase (predicted) |
| 17 | V4-P30-05(A5) | SPAC186.02c | NONE | hydroxyacid dehydrogenase (predicted) |
| 18 | V4-P33-74(G2) | SPBC29A3.21 | NONE | sequence orphan |
| 19 | V4-P34-95(H11) | SPBPB10D8.07c | YPL092W | membrane transporter (predicted) |
| 20 | V4-P35-39(D3) | SPCC162.11c | YNR012W | uridine kinase/uracil phosphoribosyltransferase (predicted) |
| 21 | V4-P35-50(E2) | SPCC16C4.17 | NONE | meiotically upregulated gene Mug123 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



