
Submitted:
14 November 2023
Posted:
15 November 2023
You are already at the latest version
Abstract
Parkinson's disease (PD) stands as the most prevalent degenerative movement disorder, characterized by the loss of dopaminergic neurons in the substantia nigra of the midbrain. In this study, we assessed the transcriptome by analyzing post-mortem mRNA extracted from the substantia nigra of individuals with PD and healthy controls. A total of 16,148 transcripts were identified, with 92 mRNAs displaying differential expression between PD and control groups. Specifically, 33 mRNAs were significantly upregulated, while 59 mRNAs were downregulated in PD compared to controls. The identification of statistically significant signaling pathways, with an adjusted p-value threshold of 0.05, unveiled noteworthy insights. Particularly, enriched categories included cardiac muscle contraction (involving genes such as ATPase Na+/K+ transporting subunit beta 2 (ATP1B2), solute carrier family 8 member A1 (SLC8A1), and cytochrome c oxidase subunit II (COX2)), GABAergic synapse (involving GABA type A receptor-associated protein like 1 (GABARAPL1), G protein subunit beta 5 (GNB5), and solute carrier family 38 member 2 (SLC38A2)), autophagy (involving GABARAPL1 and tumor protein p53-inducible nuclear protein 2 (TP53INP2)), and Fc gamma R-mediated phagocytosis (involving amphiphysin (AMPH)). These findings uncover new pathophysiological dimensions underlying PD, including the involvement of cardiac muscle contraction and specific mitochondrial activity. This knowledge not only contributes to better diagnostic precision but also paves the way for the development of new targeted therapies.Keywords: mRNAs; RNA sequencing; Parkinson’s disease; transcriptome analysis; substantia nigra
Keywords:
mRNAs
; RNA sequencing
; Parkinson’s disease
; transcriptome analysis
; substantia nigra
1. Introduction
Parkinson's disease (PD) stands out as the most prevalent movement disorder and neurodegenerative disease after Alzheimer’s dementia, affecting approximately seven million people globally [1,2]. Clinically, PD is a heterogeneous condition primarily characterized by resting tremor, bradykinesia, and rigidity. Additionally, numerous non-motor symptoms, equally debilitating and often preceding motor manifestations, contribute to the complexity of the disease [3,4,5,6]. Despite considerable research efforts, encompassing both preclinical and clinical studies, PD remains incurable. The progressive loss of dopaminergic (DA) neurons in the substantia nigra pars compacta (SNpc), a crucial part of the midbrain regulating movement tone and velocity, persists without effective intervention, and the complete pathomechanisms behind this degeneration remain incompletely elucidated [1,7].
Pathophysiologically, PD has a multifactorial origin involving complex interactions between various genetic and environmental factors. Some factors affect the elderly in general, while others seem more specifically tied to this disorder. It's noteworthy that post-mortem studies consistently report that over 60% of DA neurons in the SNc are already degenerated when overt clinical signs manifest. This indicates that PD is neuropathologically evident long before its clinical onset, suggesting a period during which the human brain can compensate for dopaminergic loss until reaching a "clinical threshold" for PD [8,9,10].
Histological studies reveal that PD is characterized by the abnormal deposition of the insoluble alpha-synuclein protein, forming aggregates known as Lewy bodies [11]. These protein aggregates progressively accumulate throughout the brainstem and various neocortical and limbic regions, reflecting the progressive degeneration of the entire central nervous system (CNS) [12]. More recently, a neuroinflammatory state has been observed in the brains of PD patients, particularly evident in the SNc [13,14]. Both neuroinflammation and dysfunctional activation of the immune system within the CNS significantly contribute to PD pathology and pathophysiology [15]. The inflammasome, a crucial complex of immune-modulating receptors and sensors, plays a role in recruiting proteins associated with apoptotic mechanisms through caspase-1 activation [16,17]. Caspase-1, in turn, activates the pro-inflammatory cytokines interleukin (IL)-1β and IL-18, perpetuating the neuroinflammatory state in PD brains [17,19,20,21]. A recent proposed model suggests that alpha-synuclein activates the inflammasome in the SN, leading to IL-activated pro-inflammatory profiles, neuronal death, and clinical symptoms [22,23,24].
Genetically, several genes or gene variants, including leucine-rich repeat kinase 2 (LRRK2), synuclein alpha (SNCA), glucosylceramidase beta-1 (GBA1), Parkin RBR E3 ubiquitin protein ligase (PARKIN), and PTEN-induced kinase 1 (PINK1), have been implicated in causing PD [25,26,27,28]. Molecular profiling studies of post-mortem SNpc samples, aimed at identifying differential molecular expression changes specific to PD compared to controls, have been conducted [29]. For instance, Simunovic et al. [30] used RNA microarrays to analyze SNpc gene expression in PD samples, identifying dysregulation of known molecular regulatory pathways in PD, including dysfunction in mitochondrial and oxidative stress-induced cellular responses [31,32]. In a recent comparative gene expression analysis on laser-dissected neurons from SNpc, Zaccaria et al. [33] revealed 52 dysregulated genes in PD samples compared to controls.
In our study, we conducted mRNA analysis and subsequent enrichment using the Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) to assess mRNAs extracted postmortem from the SN of subjects with PD and healthy controls.
2. Results
In the examination of mRNA deregulation in the SNc of PD patients, we conducted gene expression profiling on 8 post-mortem SNpc samples from PD patients and 6 from healthy control subjects (CTRL) using next-generation sequencing (RNA-Seq). The aim was to identify specific and differential changes in molecular expression. Following the removal of low-quality reads and adapter sequences, the high-quality reads were aligned against the human genome reference (hg38). In detail, we identified a total of 16,148 transcripts (Supplementary Table 1, sheet A), with 92 mRNAs differentially expressed (DEGs) between the two groups (PD vs. CTRL). Among these, 33 mRNAs were significantly upregulated (Table 1; Figure 1), while 59 mRNAs were significantly downregulated in PD compared to CTRL (Table 2; Figure 1). The normalized count of mRNAs is available at ArrayExpress (E-MTAB-13295).
The heatmap (Figure 1A) illustrates statistically significant differences in mRNA expression profiles between PD and CTRL. The volcano plot (Figure 1B) depicts the distribution of differentially expressed transcripts by their fold change and p-values. The most upregulated genes are toward the right, the most downregulated toward the left, and the most statistically significant genes at the top.
We employed the pathfindR tool to analyze significant DEGs in PD based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway database. Enrichment analysis was performed to explore functional variations between the two groups and investigate pathways potentially associated with PD. Statistically significant signaling pathways included (hsa04260) cardiac muscle contraction, (hsa04727) GABAergic synapse, (hsa04140) Autophagy, and (hsa04666) Fc gamma R-mediated phagocytosis (Figure 2A and Supplementary Table 1).
For Gene Ontology (GO) enrichment, we identified both down-regulated and up-regulated genes involved in various Molecular Functions or Biological Processes (Figure 2B and Supplementary Table 2). Notable enrichments encompassed chemical synaptic transmission, cellular response to hypoxia, autophagosome assembly, GABA receptor binding, and cell communication by electrical coupling in both heart and brain activity.
To explore Diseases and Biological Functions significantly enriched in DEGs and assess potential associations with PD susceptibility in the SNc, we employed Ingenuity Pathway Analysis (IPA). The analysis revealed significant enrichment in Neurological Disease, with multiple annotations related to disorders of basal ganglia, movement disorders, neuromuscular disease, dyskinesia, progressive motor neuropathy, familial neurological disorder, Parkinson's disease, progressive neurological disorder, abnormal morphology of the nervous system, and tauopathy (Figure 3 and Supplementary Table 3). The DEGs IPA Network Analysis identified 7 networks with nodes and interactions associated with the top Diseases or Functions Annotations, such as Neurological Disease, Organismal Injury and Abnormalities, and Psychological Disorders (Figure 4 and Table 3).
Among the genes of interest, GABARAPL1 was found to be overexpressed, as indicated by both KEGG and GO analyses, involving a distinct set of pathways (5 and 3, respectively): GABAergic synapse, autophagy – animal, NOD-like receptor signaling pathway, autophagy – other, and mitophagy – animal (according to KEGG), as well as autophagosome, GABA receptor binding, and Tat protein binding (according to GO). Autophagy, a highly conserved cellular degradation process regulated by specific autophagy-related (Atg) factors, entails the formation of double-membrane autophagosomes that engulf cytoplasmic components for degradation. In mammals, this process is complex due to the presence of six Atg8 homologues, categorized into the GABA type A receptor-associated protein (GABARAP) and microtubule-associated protein 1 light chain 3 (MAP1LC3) subfamilies [39]. GABARAPL1/GEC1, a member of the GABARAP subfamily, exhibits the highest mRNA expression among Atg8 homologues in the CNS. Notably, GABARAPL1 brain expression is observable as early as embryonic day 11, increasing progressively to peak in adulthood. Significantly, GABARAPL1 expression in the adult brain is particularly intense in neurons involved in motor and neuroendocrine functions, notably in the SNc [40]. Dysregulation of Atg8 homologues has been observed in other synucleinopathies, such as Lewy-body dementia and multiple system atrophy [41]. In PD, alterations in autophagic mechanisms are evident, as demonstrated by transcript levels of several autophagy genes in blood cells. A recent study found overexpression of autophagy-related genes, including MAP1LC3B, GABARAP, GABARAPL1, GABARAPL2, and sequestosome 1 (P62/SQSTM1) in PD patients, with potential implications for predicting markers and therapeutic responses [42].
Similarly, the SLC38A2 gene was overexpressed and associated with two relevant pathways according to KEGG: GABAergic synapse and protein digestion and absorption. This finding strengthens the connection between GABA overexpression, PD pathology, and neural degeneration. In a recent study on a rotenone-induced PD rat model [43], nardosinone, a biochemical compound enhancing NGF-mediated neurite outgrowth and synaptogenesis, demonstrated anti-PD efficacy. Transcriptome and proteome analysis suggested that the anti-PD target of nardosinone is the SLC38A2 gene, potentially involving the GABAergic synaptic pathway. This underscores the SLC38A2 gene as a potential target for PD treatment and the modulatory effects of this gene as anti-PD agents through the GABA system [43].
Diseases and Function analysis in the IPA software highlighted that DEGs correlated with Neurological Disease (Figure 3, Figure 4A, Figure 4B). Network IPA Analysis (Figure 4B) revealed downregulation of many mitochondrial genes in the SN of PD subjects, including MT-CO1, MT-CO2, MT-CO3, MT-CYB, and Cytochrome-oxidase. This further supports mitochondrial dysfunction in PD, where mitochondria, involved in crucial functions, primarily energy generation, are essential for nearly all cellular activities. Alterations in mitochondrial functioning lead to insufficient energy production, particularly affecting the CNS [45]. Evidence indicates that mitochondrial respiratory chain dysfunction plays a primary role in various neurodegenerative diseases, including PD [45,46]. Impaired elements of the respiratory chain, such as defects in complex I, have been associated with PD and frontal cortex dysfunction [47,48,49,50]. Damage to the electron transport chain increases oxidative stress and neuronal dysfunction, potentially contributing to the onset and progression of PD. Progressive mitochondrial damage results in the accumulation of non-functional mitochondria, further contributing to neuronal degeneration [53].
Looking at the downregulated genes in the SN of PD patients, a group of genes involved in maintaining the structure and function of glial cells was noted. These genes include MBP (myelin basic protein), CPN (myelin protein cyclic nucleotide phosphodiesterase), CTNNA3 (alpha-T-catenin), AQP1 (aquaporin 1), and GFAP (glial fibrillary acidic protein). Among the upregulated genes, SLC8A1 (solute carrier family 8 member A1) is involved in linking trans-plasmalemmal gradients of sodium and calcium ions to the membrane potential of astrocytes. This outcome suggests the emerging important role of glial cells in neurodegeneration and PD pathogenesis [59].
A limitation of this study is the small number of PD and control subjects recruited; however, the ability to sample human brains and extract mRNA sequences directly from the SN of both patients and controls is a significant strength of this report.
3. Materials and Methods
3.1. Human post-mortem midbrain samples
Human midbrain samples were generously provided by the Parkinson's UK Brain Bank (Imperial College London, London, UK). A total of 8 PD and 6 CTRL samples were acquired, and the specimens were histologically sectioned at the midbrain, encompassing the human SN in all slides. Each section had a thickness of 4 µm. Supplementary Table 4 provides a summary of the clinical features of the included subjects. The study adhered to the principles of the Declaration of Helsinki of 1964 and its subsequent amendments. The Ethics Committee of the Oasi Research Institute—IRCCS of Troina (Italy) granted approval for the protocol on April 5th, 2022 (approval code: 2022/04/05/CE-IRCCS-OASI/52).
3.2. RNA isolation from human midbrain samples
RNA was extracted from 4 µm FFPE slide-mounted sections using the RecoverAll Total Nucleic Acid Isolation Protocol (ThermoFisher Scientific), following the manufacturer's instructions. Subsequently, the RNA was stored at -80°C until further processing.
3.3. RNA Sequencing and Functional Analysis
RNA sequencing and subsequent data analysis were conducted by Genomix4Life Srl (Baronissi, Italy). The quality and quantity of RNA were assessed using a Qubit fluorometer (Thermo Fisher Scientific) and a TapeStation 4200 (Agilent Technologies, 5301 Stevens Creek Blvd, Santa Clara, CA, USA), respectively.
Indexed libraries were prepared from 50 ng of purified RNA each, employing the Illumina Stranded Total RNA with Ribo-Zero Plus Kit (Illumina), following the manufacturer's guidelines. Library quantification was performed using the TapeStation 4200 (Agilent Technologies) and Qubit (Thermo Fisher). Subsequently, the indexed libraries were pooled in equimolar amounts, resulting in a final concentration of 2 nM. The Illumina NovaSeq 6000 System was utilized to sequence the pooled samples in a 2 × 75 paired-end format.
Quality control analysis was executed on the raw fastq files generated earlier using the FastQC tool [http://www.bioinformatics.babraham.ac.uk/projects/fastqc/]. Trim-ming of short reads (<25 bp) and removal of adapter sequences were carried out with cutadapt (v.2.8) [60]. The trimmed fastq files were mapped to the reference genome (GenCode (HG38-Release 37 (GRCh38.p13)) [https://www.gencodegenes.org/human/]) using STAR (version 2.7.3a) [61] with standard parameters. Gene quantification per sample was accomplished using featureCount (version 2.0) [https://bioweb.pasteur.fr/docs/modules/subread/1.4.6-p3/SubreadUsersGuide], and a custom R script was employed for data normalization and differential expression analysis using the Bioconductor DESeq2 [62] package. The threshold for considering genes as differentially expressed was set at Fold-Change ≥ 1.50 or ≤ -1.50 (|FC| ≥ 1.50) with adjusted p-values ≤ 0.05 (padj). Volcano plots and heatmaps were generated using the EnhancedVolcano (10.18129/B9.bioc.EnhancedVolcano) and ComplexHeatmap [63] packages in R.
For functional analysis, KEGG pathway and GO database analyses were conducted using the R package pathfinder [https://doi.org/10.3389/fgene.2019.00858]. Additionally, to gain a deeper understanding of the complex transcriptomics data, Ingenuity Pathway Analysis (IPA) [64] was performed, specifically for investigating diseases and function analysis.
The raw data (.fastq files) and the normalized count of identified mRNAs are available on ArrayExpress (E-MTAB-13295).
4. Conclusions
Despite the limited sample size, the comprehensive analysis conducted on the transcriptome of these samples, along with KEGG and GO enrichment analyses, revealed statistically significant findings associated with the following signaling pathways: cardiac muscle contraction, GABAergic synapse, autophagy, Fc gamma R-mediated phagocytosis signaling pathway, cellular response to chemical synaptic transmission to hypoxia, autophagosome assembly, GABA receptor binding, and cellular communication via electrical coupling involved in cardiac conduction.
Upon examining the genes that were either upregulated or downregulated, these results contribute to a translational perspective and unveil novel pathophysiological insights into PD. This includes implications in cell communication via electrical coupling, specific mitochondrial activity, and neuronal-glia interaction. These findings may pave the way for enhanced diagnostic precision and the development of novel targeted therapies.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Table 1: KEGG pathway database and enrichment analysis; Supplementary Table 2: GO pathway database and enrichment analysis; Supplementary Table 3: KEGG pathway database and enrichment analysis; Supplementary Table 3: Diseases and Biological Functions; Supplementary Table 4: main clinical features on the subjects included in the study.
Author Contributions
Concept and Design, M.S., G.C., and R.F.; performed Next generation sequencing analysis, M.R., G.M., and G.M.V.; acquisition of data or analysis, M.R., G.M., and G.M.V. and M.S.; writing original draft preparation, M.S., G.L., M.R., G.M., F.A.S., M.G.S., G.C., S.M., and R.F.; Final approval, M.S., G.L., M.R., and R.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Italian Ministry of Health, grant number RC- 2779777.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki of 1964 and its later amendments and the Ethics Committee of the Oasi Research Institute-IRCCS of Troina (Italy) approved the protocol on April 5th, 2022 (approval code: 2022/04/05/CE-IRCCS-OASI/52).
Informed Consent Statement
The Centre for Blast Injury Studies fully complies with The Human Tissue Act 2004 (which replaced the Human Tissue Act 1961, the Anatomy Act 1984 and the Human Organ Transplants Act 1989), governed by the Human Tissue Authority, which outlines the use of human tissue for scientific purposes in the UK. Although the Human Tissue Act does not cover the use of tissues that are sourced outside the UK, the Centre also complies with its standards in its dealings with human tissue from overseas. Imperial College London also holds an institutional license from the Human Tissue Authority to collect, store and use human tissue. As provided for in the Human Tissue Act, and depending on the nature of the research, organizations other than the Human Tissue Authority regulate the actual research on human tissue and in the case of this study this has been via Ethics Committee of the Oasi Research Institute-IRCCS of Troina (Italy). .
Data Availability Statement
https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (E-MTAB-13295).
Acknowledgments
Thanks to Parkinson's UK Brain Bank (Imperial College London, London, UK) for providing human midbrain samples.
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study, collection, analyses, or interpretation of data, as well as in the writing of the manuscript or in the decision to publish the results.
References
- Kalia, L.V.; Lang, A.E. Parkinson’s Disease. Lancet 2015, 386, 896–912. [Google Scholar] [CrossRef] [PubMed]
- Quik, M. Smoking, Nicotine and Parkinson’s Disease. Trends Neurosci 2004, 27, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.E.; Obeso, J.A. Time to Move beyond Nigrostriatal Dopamine Deficiency in Parkinson’s Disease. Ann Neurol 2004, 55, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Obeso, J.A.; Stamelou, M.; Goetz, C.G.; Poewe, W.; Lang, A.E.; Weintraub, D.; Burn, D.; Halliday, G.M.; Bezard, E.; Przedborski, S.; et al. Past, Present, and Future of Parkinson’s Disease: A Special Essay on the 200th Anniversary of the Shaking Palsy. Mov Disord 2017, 32, 1264–1310. [Google Scholar] [CrossRef]
- Figorilli, M.; Lanza, G.; Congiu, P.; Lecca, R.; Casaglia, E.; Mogavero, M.P.; Puligheddu, M.; Ferri, R. Neurophysiological Aspects of REM Sleep Behavior Disorder (RBD): A Narrative Review. Brain Sci 2021, 11, 1588. [Google Scholar] [CrossRef] [PubMed]
- Joza, S.; Hu, M.T.; Jung, K.-Y.; Kunz, D.; Stefani, A.; Dušek, P.; Terzaghi, M.; Arnaldi, D.; Videnovic, A.; Schiess, M.C.; et al. Progression of Clinical Markers in Prodromal Parkinson’s Disease and Dementia with Lewy Bodies: A Multicentre Study. Brain 2023, 146, 3258–3272. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.E. A Critical Appraisal of the Premotor Symptoms of Parkinson’s Disease: Potential Usefulness in Early Diagnosis and Design of Neuroprotective Trials. Mov Disord 2011, 26, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-C.; Ulane, C.M.; Burke, R.E. Clinical Progression in Parkinson Disease and the Neurobiology of Axons. Ann Neurol 2010, 67, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.E.; O’Malley, K. Axon Degeneration in Parkinson’s Disease. Exp Neurol 2013, 246, 72–83. [Google Scholar] [CrossRef]
- Dauer, W.; Przedborski, S. Parkinson’s Disease: Mechanisms and Models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef]
- Braak, H.; Sandmann-Keil, D.; Gai, W.; Braak, E. Extensive Axonal Lewy Neurites in Parkinson’s Disease: A Novel Pathological Feature Revealed by Alpha-Synuclein Immunocytochemistry. Neurosci Lett 1999, 265, 67–69. [Google Scholar] [CrossRef]
- Braak, H.; Del Tredici, K.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of Brain Pathology Related to Sporadic Parkinson’s Disease. Neurobiol Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Fearnley, J.M.; Lees, A.J. Ageing and Parkinson’s Disease: Substantia Nigra Regional Selectivity. Brain 1991, 114 ( Pt 5), 2283–2301. [Google Scholar] [CrossRef]
- Hirsch, E.C.; Breidert, T.; Rousselet, E.; Hunot, S.; Hartmann, A.; Michel, P.P. The Role of Glial Reaction and Inflammation in Parkinson’s Disease. Ann N Y Acad Sci 2003, 991, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.E.; Akther, M.; Jakaria, M.; Kim, I.-S.; Azam, S.; Choi, D.-K. Targeting the Microglial NLRP3 Inflammasome and Its Role in Parkinson’s Disease. Mov Disord 2020, 35, 20–33. [Google Scholar] [CrossRef] [PubMed]
- de Rivero Vaccari, J.P.; Dietrich, W.D.; Keane, R.W. Therapeutics Targeting the Inflammasome after Central Nervous System Injury. Transl Res 2016, 167, 35–45. [Google Scholar] [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The Inflammasome: A Molecular Platform Triggering Activation of Inflammatory Caspases and Processing of proIL-Beta. Mol Cell 2002, 10, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of Assembly, Regulation and Signalling. Nat Rev Immunol 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Malik, A.; Kanneganti, T.-D. Inflammasome Activation and Assembly at a Glance. J Cell Sci 2017, 130, 3955–3963. [Google Scholar] [CrossRef]
- Van Opdenbosch, N.; Lamkanfi, M. Caspases in Cell Death, Inflammation, and Disease. Immunity 2019, 50, 1352–1364. [Google Scholar] [CrossRef]
- Franklin, B.S.; Bossaller, L.; De Nardo, D.; Ratter, J.M.; Stutz, A.; Engels, G.; Brenker, C.; Nordhoff, M.; Mirandola, S.R.; Al-Amoudi, A.; et al. The Adaptor ASC Has Extracellular and “prionoid” Activities That Propagate Inflammation. Nat Immunol 2014, 15, 727–737. [Google Scholar] [CrossRef]
- Cabrera Ranaldi, E.D.L.R.M.; Nuytemans, K.; Martinez, A.; Luca, C.C.; Keane, R.W.; de Rivero Vaccari, J.P. Proof-of-Principle Study of Inflammasome Signaling Proteins as Diagnostic Biomarkers of the Inflammatory Response in Parkinson’s Disease. Pharmaceuticals (Basel) 2023, 16, 883. [Google Scholar] [CrossRef]
- Codolo, G.; Plotegher, N.; Pozzobon, T.; Brucale, M.; Tessari, I.; Bubacco, L.; de Bernard, M. Triggering of Inflammasome by Aggregated α-Synuclein, an Inflammatory Response in Synucleinopathies. PLoS One 2013, 8, e553752013–8. [Google Scholar] [CrossRef] [PubMed]
- de Rivero Vaccari, J.P.; Dietrich, W.D.; Keane, R.W. Activation and Regulation of Cellular Inflammasomes: Gaps in Our Knowledge for Central Nervous System Injury. J Cereb Blood Flow Metab 2014, 34, 369–375. [Google Scholar] [CrossRef]
- Funayama, M.; Nishioka, K.; Li, Y.; Hattori, N. Molecular Genetics of Parkinson’s Disease: Contributions and Global Trends. J Hum Genet 2023, 68, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Nalls, M.A.; Pankratz, N.; Lill, C.M.; Do, C.B.; Hernandez, D.G.; Saad, M.; DeStefano, A.L.; Kara, E.; Bras, J.; Sharma, M.; et al. Large-Scale Meta-Analysis of Genome-Wide Association Data Identifies Six New Risk Loci for Parkinson’s Disease. Nat Genet 2014, 46, 989–993. [Google Scholar] [CrossRef]
- Bandres-Ciga, S.; Diez-Fairen, M.; Kim, J.J.; Singleton, A.B. Genetics of Parkinson’s Disease: An Introspection of Its Journey towards Precision Medicine. Neurobiol Dis 2020, 137, 104782. [Google Scholar] [CrossRef] [PubMed]
- Dulski, J.; Uitti, R.J.; Ross, O.A.; Wszolek, Z.K. Genetic Architecture of Parkinson’s Disease Subtypes - Review of the Literature. Front Aging Neurosci 2022, 14, 1023574. [Google Scholar] [CrossRef]
- Redenšek, S.; Dolžan, V.; Kunej, T. From Genomics to Omics Landscapes of Parkinson’s Disease: Revealing the Molecular Mechanisms. OMICS 2018, 22, 1–16. [Google Scholar] [CrossRef]
- Simunovic, F.; Yi, M.; Wang, Y.; Macey, L.; Brown, L.T.; Krichevsky, A.M.; Andersen, S.L.; Stephens, R.M.; Benes, F.M.; Sonntag, K.C. Gene Expression Profiling of Substantia Nigra Dopamine Neurons: Further Insights into Parkinson’s Disease Pathology. Brain 2009, 132, 1795–1809. [Google Scholar] [CrossRef]
- Salemi, M.; Cosentino, F.; Lanza, G.; Cantone, M.; Salluzzo, M.G.; Giurato, G.; Borgione, E.; Marchese, G.; Santa Paola, S.; Lanuzza, B.; et al. MRNA Expression Profiling of Mitochondrial Subunits in Subjects with Parkinson’s Disease. Arch Med Sci 2023, 19, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Salemi, M.; Lanza, G.; Mogavero, M.P.; Cosentino, F.I.I.; Borgione, E.; Iorio, R.; Ventola, G.M.; Marchese, G.; Salluzzo, M.G.; Ravo, M.; et al. A Transcriptome Analysis of MRNAs and Long Non-Coding RNAs in Patients with Parkinson’s Disease. Int J Mol Sci 2022, 23, 1535. [Google Scholar] [CrossRef] [PubMed]
- Zaccaria, A.; Antinori, P.; Licker, V.; Kövari, E.; Lobrinus, J.A.; Burkhard, P.R. Multiomic Analyses of Dopaminergic Neurons Isolated from Human Substantia Nigra in Parkinson’s Disease: A Descriptive and Exploratory Study. Cell Mol Neurobiol 2022, 42, 2805–2818. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, J.; Li, S.; Zhang, J.; Zheng, J.; Hou, W.; Zhao, H.; Guo, Y.; Liu, X.; Dou, K.; et al. N-Myc Downstream-Regulated Gene 2, a Novel Estrogen-Targeted Gene, Is Involved in the Regulation of Na+/K+-ATPase. J Biol Chem 2011, 286, 32289–32299. [Google Scholar] [CrossRef] [PubMed]
- Mauri, N.; Kleiter, M.; Dietschi, E.; Leschnik, M.; Högler, S.; Wiedmer, M.; Dietrich, J.; Henke, D.; Steffen, F.; Schuller, S.; et al. A SINE Insertion in ATP1B2 in Belgian Shepherd Dogs Affected by Spongy Degeneration with Cerebellar Ataxia (SDCA2). G3 (Bethesda) 2017, 7, 2729–2737. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Daniels, S.; Kim, Y.; Chu, H.-Y. Cell Type-Specific Decrease of the Intrinsic Excitability of Motor Cortical Pyramidal Neurons in Parkinsonism. J Neurosci 2021, 41, 5553–5565. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Arias, P.; Einholm, A.P.; Mamsa, H.; Concheiro, C.; Gutiérrez-de-Terán, H.; Romero, J.; Toustrup-Jensen, M.S.; Carracedo, A.; Jen, J.C.; Vilsen, B.; et al. A C-Terminal Mutation of ATP1A3 Underscores the Crucial Role of Sodium Affinity in the Pathophysiology of Rapid-Onset Dystonia-Parkinsonism. Hum Mol Genet 2009, 18, 2370–2377. [Google Scholar] [CrossRef] [PubMed]
- Bardutz, H.; Singh, J.; Rehman, Z.; Bernat, P. Parkinson’s Disease and the Cardiac Cycle: A Rapid Literature Review and Case Series. Life (Basel) 2023, 13, 1003. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, M.B.E.; Keulers, T.G.; Vooijs, M.A.; Rouschop, K.M.A. LC3/GABARAP Family Proteins: Autophagy-(Un)Related Functions. FASEB J 2016, 30, 3961–3978. [Google Scholar] [CrossRef]
- Le Grand, J.N.; Bon, K.; Fraichard, A.; Zhang, J.; Jouvenot, M.; Risold, P.-Y.; Boyer-Guittaut, M.; Delage-Mourroux, R. Specific Distribution of the Autophagic Protein GABARAPL1/GEC1 in the Developing and Adult Mouse Brain and Identification of Neuronal Populations Expressing GABARAPL1/GEC1. PLoS One 2013, 8, e63133. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Tanji, K. [Multiple system atrophy and autophagy]. Rinsho Shinkeigaku 2014, 54, 966–968. [Google Scholar] [CrossRef]
- El Haddad, S.; Serrano, A.; Moal, F.; Normand, T.; Robin, C.; Charpentier, S.; Valery, A.; Brulé-Morabito, F.; Auzou, P.; Mollet, L.; et al. Disturbed Expression of Autophagy Genes in Blood of Parkinson’s Disease Patients. Gene 2020, 738, 144454. [Google Scholar] [CrossRef]
- Bian, L.-H.; Yao, Z.-W.; Wang, Z.-Y.; Wang, X.-M.; Li, Q.-Y.; Yang, X.; Li, J.-Y.; Wei, X.-J.; Wan, G.-H.; Wang, Y.-Q.; et al. Nardosinone Regulates the Slc38a2 Gene to Alleviate Parkinson’s Symptoms in Rats through the GABAergic Synaptic and cAMP Pathways. Biomed Pharmacother 2022, 153, 113269. [Google Scholar] [CrossRef]
- Li, P.; Matsunaga, K.; Yamamoto, K.; Yoshikawa, R.; Kawashima, K.; Ohizumi, Y. Nardosinone, a Novel Enhancer of Nerve Growth Factor in Neurite Outgrowth from PC12D Cells. Neurosci Lett 1999, 273, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Lezi, E.; Swerdlow, R.H. Mitochondria in Neurodegeneration. Adv Exp Med Biol 2012, 942, 269–286. [Google Scholar] [CrossRef]
- Power, J.H.T.; Barnes, O.L.; Chegini, F. Lewy Bodies and the Mechanisms of Neuronal Cell Death in Parkinson’s Disease and Dementia with Lewy Bodies. Brain Pathol 2017, 27, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Parker, W.D.; Boyson, S.J.; Parks, J.K. Abnormalities of the Electron Transport Chain in Idiopathic Parkinson’s Disease. Ann Neurol 1989, 26, 719–723. [Google Scholar] [CrossRef]
- Schapira, A.H.; Cooper, J.M.; Dexter, D.; Jenner, P.; Clark, J.B.; Marsden, C.D. Mitochondrial Complex I Deficiency in Parkinson’s Disease. Lancet 1989, 1, 1269. [Google Scholar] [CrossRef] [PubMed]
- Bindoff, L.A.; Birch-Machin, M.; Cartlidge, N.E.; Parker, W.D.; Turnbull, D.M. Mitochondrial Function in Parkinson’s Disease. Lancet 1989, 2, 49. [Google Scholar] [CrossRef]
- Parker, W.D.; Parks, J.K.; Swerdlow, R.H. Complex I Deficiency in Parkinson’s Disease Frontal Cortex. Brain Res 2008, 1189, 215–218. [Google Scholar] [CrossRef]
- Houshmand, M.; Kasraie, S.; Etemad Ahari, S.; Moin, M.; Bahar, M.; Zamani, A. Investigation of tRNA and ATPase 6/8 Gene Mutations in Iranian Ataxia Telangiectasia Patients. Arch Med Sci 2011, 7, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Koyano, F.; Okatsu, K.; Ishigaki, S.; Fujioka, Y.; Kimura, M.; Sobue, G.; Tanaka, K.; Matsuda, N. The Principal PINK1 and Parkin Cellular Events Triggered in Response to Dissipation of Mitochondrial Membrane Potential Occur in Primary Neurons. Genes Cells 2013, 18, 672–681. [Google Scholar] [CrossRef]
- McCoy, M.K.; Cookson, M.R. Mitochondrial Quality Control and Dynamics in Parkinson’s Disease. Antioxid Redox Signal 2012, 16, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.-A.; Trapp, B.D. Axon-Glial Signaling and the Glial Support of Axon Function. Annu Rev Neurosci 2008, 31, 535–561. [Google Scholar] [CrossRef] [PubMed]
- Weng, A.; Rabin, E.E.; Flozak, A.S.; Chiarella, S.E.; Aillon, R.P.; Gottardi, C.J. Alpha-T-Catenin Is Expressed in Peripheral Nerves as a Constituent of Schwann Cell Adherens Junctions. Biol Open 2022, 11, bio059634. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhan, J.; Cai, Q.; Xu, F.; Chai, R.; Lam, K.; Luan, Z.; Zhou, G.; Tsang, S.; Kipp, M.; et al. The Water Transport System in Astrocytes-Aquaporins. Cells 2022, 11, 2564. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.L.; Faridounnia, M.; Armao, D.; Snider, N.T. Stability Dynamics of Neurofilament and GFAP Networks and Protein Fragments. Curr Opin Cell Biol 2023, 85, 102266. [Google Scholar] [CrossRef] [PubMed]
- Rose, C.R.; Ziemens, D.; Verkhratsky, A. On the Special Role of NCX in Astrocytes: Translating Na+-Transients into Intracellular Ca2+ Signals. Cell Calcium 2020, 86, 102154. [Google Scholar] [CrossRef] [PubMed]
- Valori, C.F.; Guidotti, G.; Brambilla, L.; Rossi, D. Astrocytes: Emerging Therapeutic Targets in Neurological Disorders. Trends Mol Med 2019, 25, 750–759. [Google Scholar] [CrossRef]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J Comput Biol 2017, 24, 1138–1143. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Eils, R.; Schlesner, M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal Analysis Approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Visualization of differentially expressed genes (DEGs). (A) Heatmap of significant DEGs in patients with Parkinson’s disease (PD) and healthy controls (CTRL). In yellow, the genes with up normalized expression level, whereas in blue the down genes. The log2 (foldChange) bar indicates in red and in green the up and down-regulated genes, respectively. (B) Volcano plot of significant DEGs based on fold changes and p-values. The green color shows the down-regulates genes, whereas the red color shows the up-regulates genes.
Figure 1.
Visualization of differentially expressed genes (DEGs). (A) Heatmap of significant DEGs in patients with Parkinson’s disease (PD) and healthy controls (CTRL). In yellow, the genes with up normalized expression level, whereas in blue the down genes. The log2 (foldChange) bar indicates in red and in green the up and down-regulated genes, respectively. (B) Volcano plot of significant DEGs based on fold changes and p-values. The green color shows the down-regulates genes, whereas the red color shows the up-regulates genes.
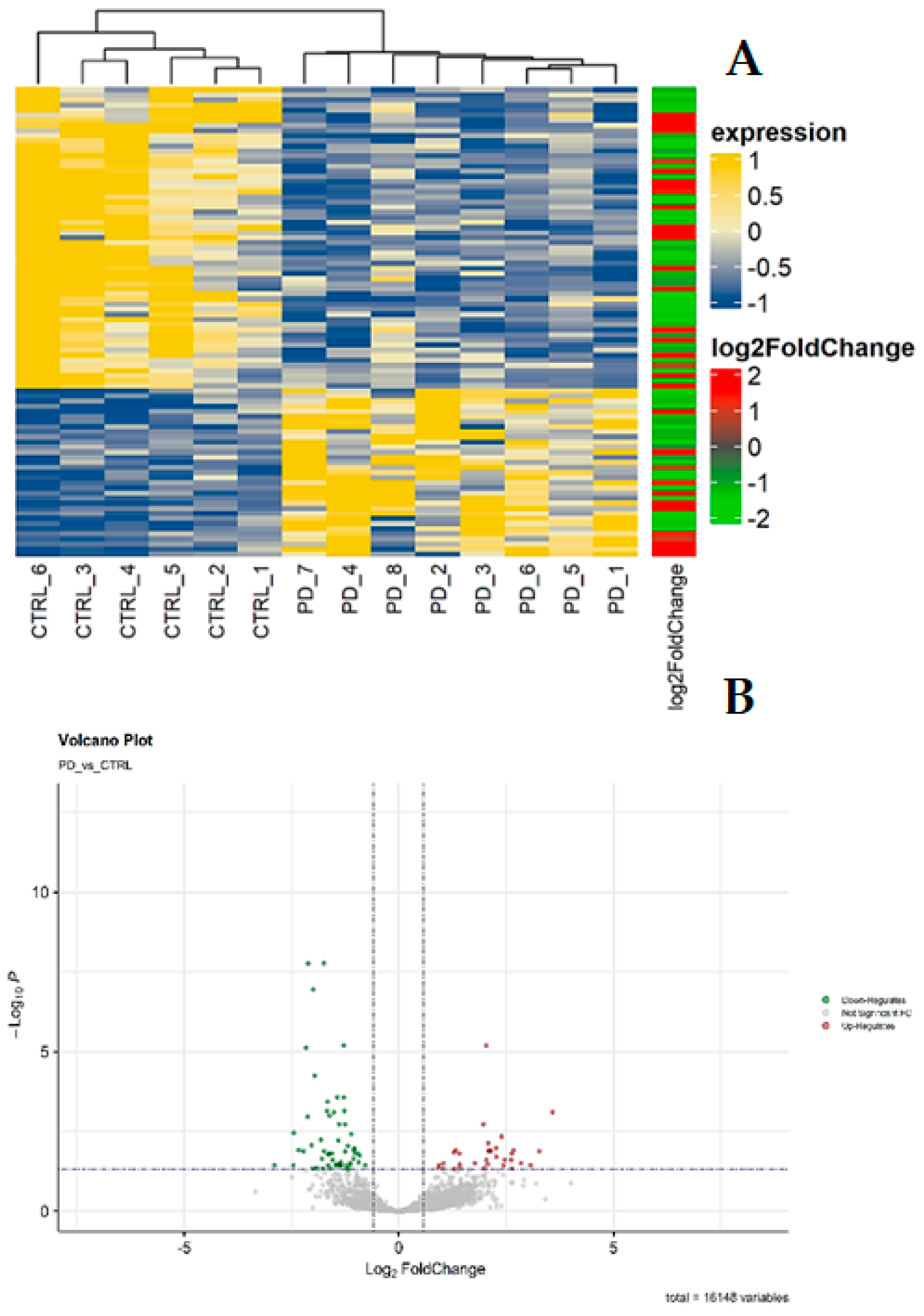
Figure 2.
KEGG and GO enrichment analysis of differentially expressed genes. (A) UpSet Plot shown the intersections of significant genes and top 30 enriched KEGG pathway and (B) GO terms, with the Log2FoldChange scale.
Figure 2.
KEGG and GO enrichment analysis of differentially expressed genes. (A) UpSet Plot shown the intersections of significant genes and top 30 enriched KEGG pathway and (B) GO terms, with the Log2FoldChange scale.
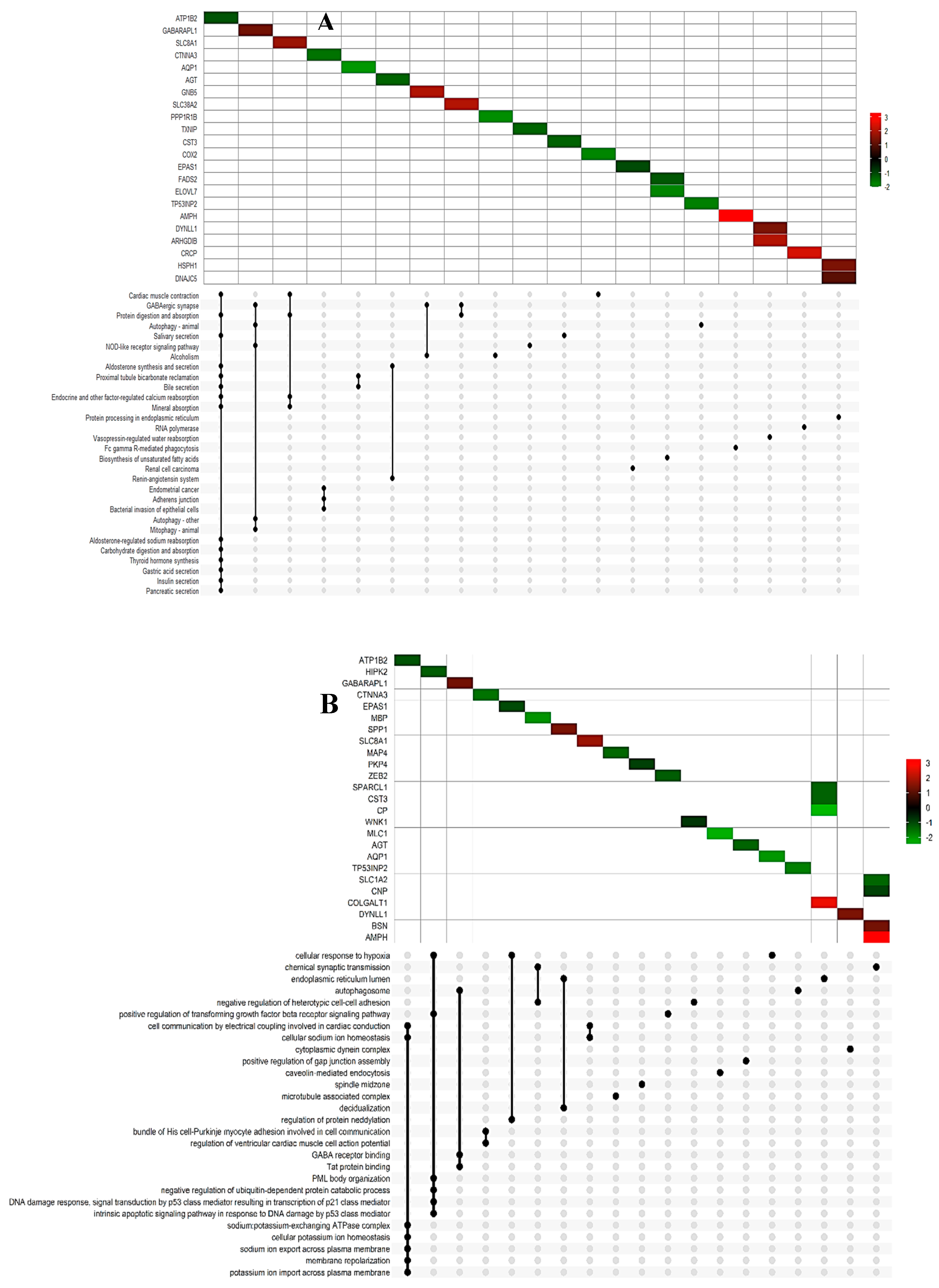
Figure 3.
Ingenuity Pathway Analysis (IPA). Diseases and Function analysis of differentially expressed genes in IPA software with Log2 (p-value) scale.
Figure 3.
Ingenuity Pathway Analysis (IPA). Diseases and Function analysis of differentially expressed genes in IPA software with Log2 (p-value) scale.
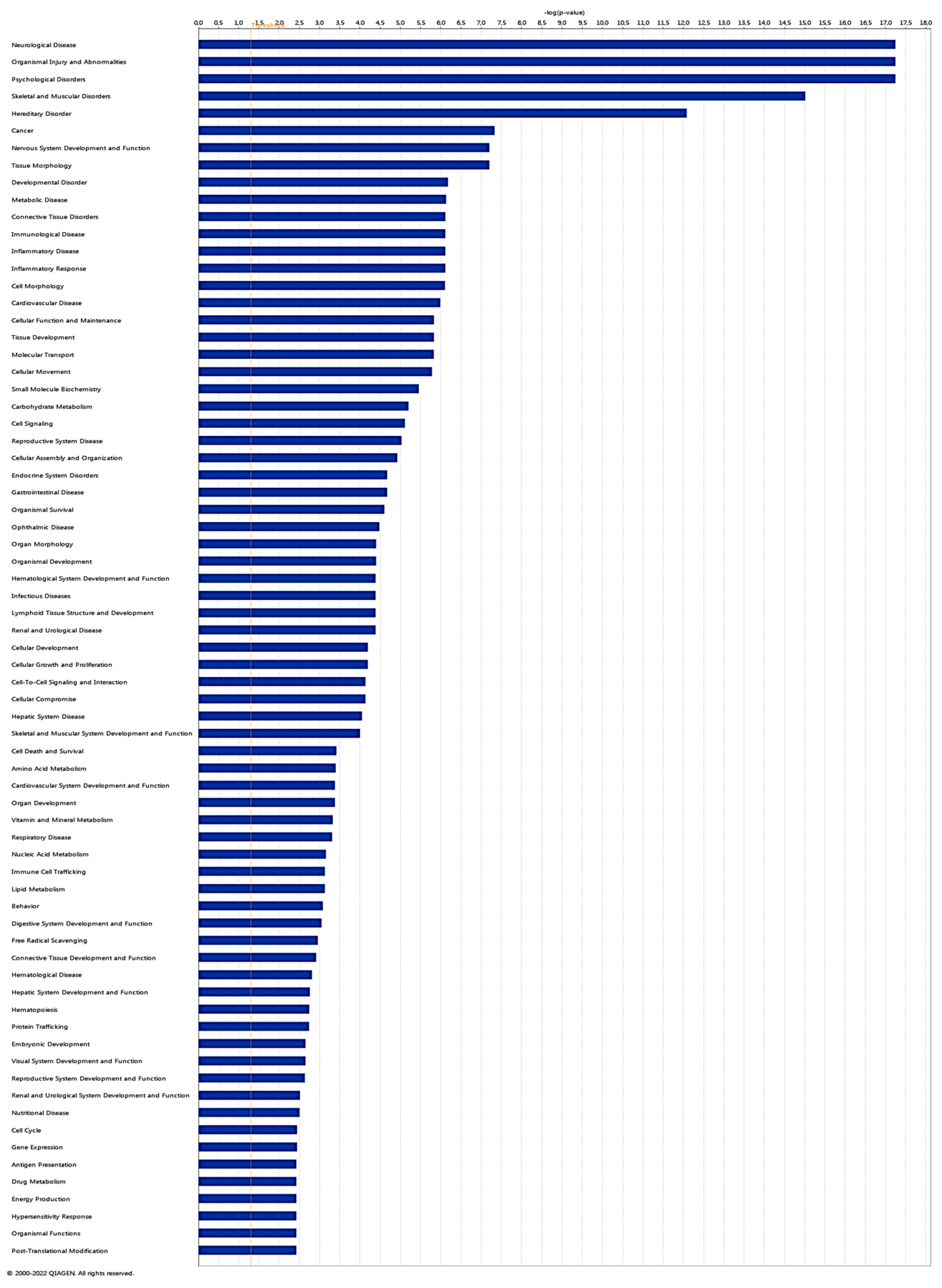
Figure 4.
Network Ingenuity Pathway Analysis (IPA). Neurological Disease, Organismal Injury and Abnormalities, Psychological Disorders. Red indicates the upregulated transcripts, whereas green the downregulated transcripts.
Figure 4.
Network Ingenuity Pathway Analysis (IPA). Neurological Disease, Organismal Injury and Abnormalities, Psychological Disorders. Red indicates the upregulated transcripts, whereas green the downregulated transcripts.
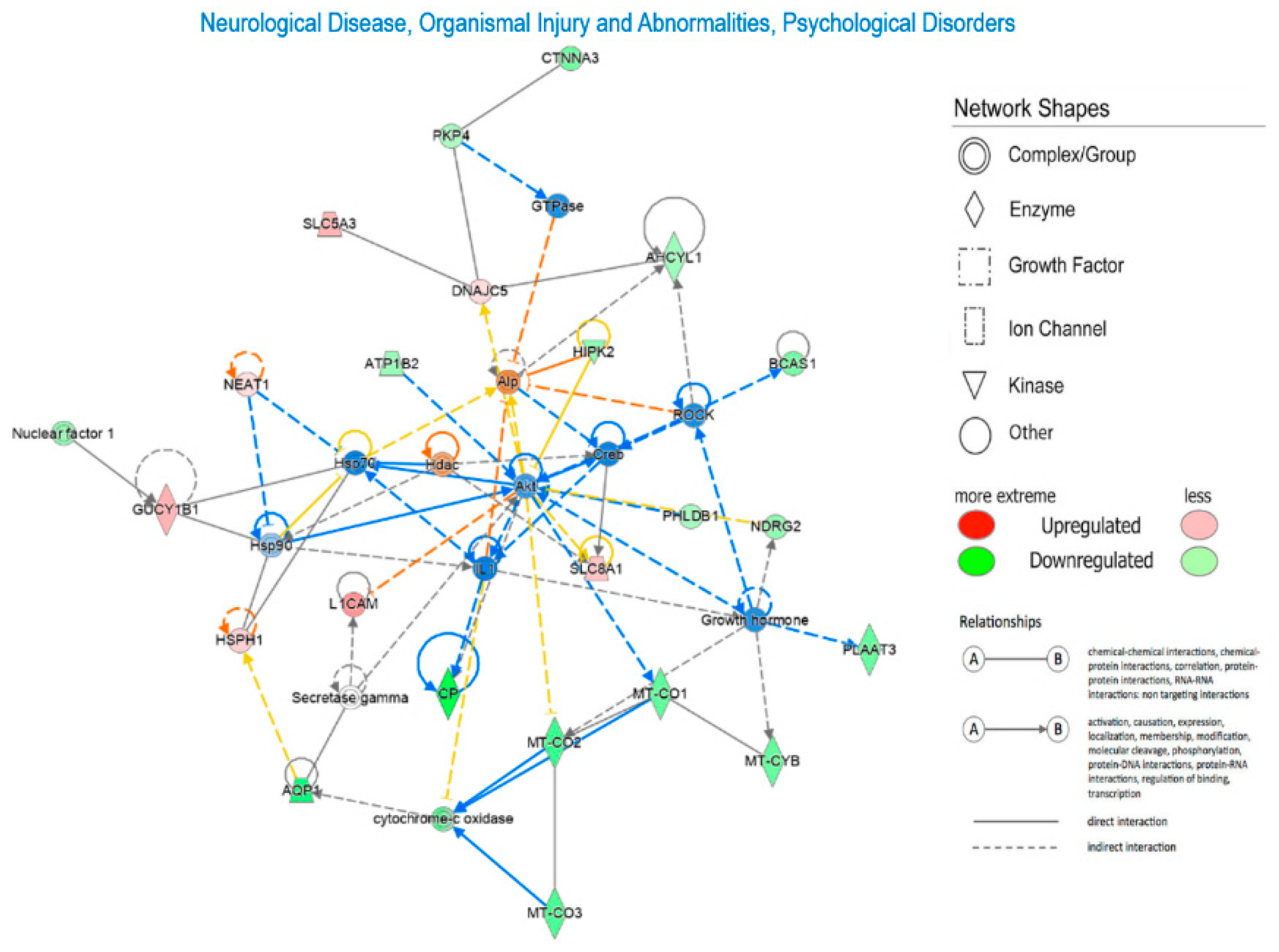

Table 1.
mRNAs down-expressed in PD subjects compared to controls (padj ≤0.05 and |FC| ≥1.5).
| Gene ID | Fold change | Gene ID | Fold change |
|---|---|---|---|
| ETNPP1 | -7.435 | AC093330.1 | -2.718 |
| MIND4P12 | -5.475 | MAP4 | -2.693 |
| CP | -5.444 | S1C1A2 | -2.651 |
| M1C1 | -5.02 | TXNIP | -2.625 |
| PPDPF | -4.631 | AGf | -2.61 |
| PAQR6 | -4.469 | RHOBTB3 | -2.604 |
| ACBD7 | -4.345 | CST3 | -2.562 |
| MOBP | -4.315 | SPARC11 | -2.55 |
| AQP1 | -4.069 | MIND2P28 | -2.544 |
| TAG1N | -4.001 | HIPK2 | -2.427 |
| MBP | -3.977 | ZEB2 | -2.425 |
| PAJP2B | -3.892 | NDRG2 | -2.408 |
| PPP1R1B | -3.766 | MIURN | -2.382 |
| MTATP6P1 | -3.736 | DAAM2 | -2.381 |
| MT-C02 | -3.5 | NFIX | -2.343 |
| E1OV17 | -3.456 | FADS2 | -2.325 |
| SCARA3 | -3.419 | ATP1B2 | -2.268 |
| MT-C03 | -3.338 | HEPACAM | -2.263 |
| TP53INP2 | -3.336 | SHIN1 | -2.203 |
| FAM107A | -3.168 | FAR1 | -2.182 |
| SEPTIN4 | -3.158 | AHCYU | -2.157 |
| A1AD | -3.122 | EPAS1 | -2.075 |
| MT-CO1 | -3.116 | PH1DB1 | -2.058 |
| IRAG1 | -3.089 | GFAP | -2.043 |
| P1AAT3 | -3.056 | MAP4K4 | -2.041 |
| MT-CYB | -3.045 | PAD12 | -1.938 |
| CINNA3 | -2.949 | CNP | -1.916 |
| FAT3 | -2.906 | PKP4 | -1.874 |
| BCAS1 | -2.857 | WNK1 | -1.714 |
| TSC2204 | -2.767 |
Table 2.
mRNAs over-expressed in PD subjects compared to controls (padj ≤0.05 and |FC| ≥1.5).
| Gene ID | Fold change | Gene ID | Fold change |
|---|---|---|---|
| I110RA | 12.026 | FYB1 | 4.204 |
| AMPH | 9.664 | ARHGDIB | 4.111 |
| HS6ST3 | 8.405 | S1C38A2 | 4.104 |
| VSN11 | 7.195 | GNB5 | 3.998 |
| CO1GA1T1 | 6.373 | NAA30 | 3.930 |
| PRDM11 | 6.234 | S1C8A1 | 3.409 |
| 11CAM | 6.208 | DYNLL1 | 2.680 |
| GPR34 | 6.141 | HSPH1 | 2.666 |
| ZNF618 | 5.543 | SPP1 | 2.660 |
| RCSD1 | 5.419 | BSN | 2.518 |
| CRCP | 5.257 | GABARAP11 | 2.471 |
| 1NA | 4.831 | YWHAG | 2.443 |
| PIPRT | 4.812 | QDPR | 2.060 |
| GUCY1B1 | 4.422 | DNAjC5 | 2.042 |
| S1CSA3 | 4.338 | NEAT1 | 1.926 |
| SRGN | 4.250 | TNPO1 | 1.890 |
| SCN8A | 4.236 |
Table 3.
Networks obtained by IPA Core Analysis.
| Molecules in network | Score | Focus molecules | Diseases and functions |
|---|---|---|---|
| AHCYL1, Akt, Alp, AQP1, ATP1B2, BCAS1, CP, Creb, CTNNA3, cytochrome-c oxidase, DNAJCS, Growth hormone, GTPase, GUCY1B1, Hdac, HIPK2, Hsp70, Hsp90, HSPH1, IL1, L1CAM, MT-C01, MTC02, MT-C03, MT-CYB, NDRG2, NEAT1, Nuclear factor M1, PHLDB1, PKP4, PLAAT3, ROCK, Secretase gamma, SLCSA3, SLC8A1 | 49 | 22 | Neurological Disease, Organismal Injury and Abnormalities, Psychological Disorders |
| 14-3-3, 20s proteasome, 26s Pro teasome, ALAD, BSN, Calmodulin, calpain, CG, CNP, COLGALT1, Collagen Alpha1, Collagen type I (complex), Collagen type IV, EPAS1, ERK1/2, FAR1, Focal adhesion kinase, GFAP, HEPACAM, IN A, insulin, MAP4, MBP, MLC1, PDGF BB, Pka, PP2A, SEPTIN4, SLC1A2, SRGN, TAGLN, Tgf beta, transglutaminase, VSNL1, WNK1 | 38 | 18 | Cellular Function and Maintenance, Nervous System Development and Function, Tissue Development |
| AGT, AMPH, Ap1, ARHGDIB, Calcineurin protein(s), CD3, collagen type I (family), cytokine, ELOVL7, FYB1, GNBS, Gsk3, IKK (complex), IL12 (complex), integrin, integrin alpha L beta 2, Jnk, LDL, MAP4K4, Mek, MTURN, NFAT (complex), Nfat (family), NFIX, NFkB (complex), Nrlh, P38 MAPK, PAD12, Pkc(s), PPP1R1B, Rac, SPARCL1, SPP1, TCR, voltage-gated calcium channel Act in, AMPK, Ck2, C LEC9A, CST3, DYNLL1, | 25 | 13 | Cardiovascular Disease, Cell-To-Cell Signaling and Interaction, Organismal Injury and Abnormalities |
| Actin, AMPK, Ck2, C LEC9A, CST3, DYNLL1, ERK, F Actin, FADS2, FAM 107 A, GABARAPL1, G PR34 , hemoglobin, Histone h3, Histone h4, IgG, IL1ORA, IL12 (family), immunoglobulin, interferon alpha, Mapk, MHC Class II (complex), Notch, P13K (complex), RNA polymerase 11 , SHTN1, Siglech, SRC (family), trypsin, tubulin, TXNIP, Ubiquitin, Vegf, YWHAG, ZEB2 | 20 | 11 | Cell-To-Cell Signaling and Interaction, Infectious Diseases, Organismal Injury and Abnormalities |
| ACOD1, CARD16, CASP8, Cd24a, COL2A1, cytokine receptor, Dglucose, DAAM2, FAT3, GBPS, HCAR2, HEPACAM2, IFNG, ligp1, IL10RB, IL17RE, IL18BP, IL2RA, IRAGl , LGALS1, LTC4S, MLKL, NAA30, NLRCS, PARVG, PLAAT3, PRDM11, QDPR, REL, RHOBTB3, SCARA3, Tlr11, TNFRSF10B, TP531NP2, ZBP1 | 18 | 10 | Gastrointestinal Disease, Inflammatory Response, Organismal Injury and Abnormalities |
| CFB, CHADL, CNTLN, CSNK1A1, EP300, ETNPPL, FAM110D, FAM83G, FRMD4A, FRY, HDAC4, HDAC5, IKZF2, IL15RA, importin alpha, MECOM, miR-129-Sp (and other miRNAs w/seed UUUUUGC), MOBP, NRBP2, PAIP2B, PAQR6, PDCD1LG2, PPDPF, PRMT1, RCSD1, SCN8A, SMARCB 1, SNX22, SNX24, SOX2, SOX9, TNP01, TSC22D4, ZDBF2, ZNF 618 | 18 | 10 | Carbohydrate Metabolism, Cell Cycle, Cellular Assembly and Organization |
| ACBD7, betaestradiol, CA2, CALCRL, Clathrin, CRCP, DEFB116, DNPH1, DOCK3, ERBB, FMOS, HS6ST3, HTR4, INP P5F, L-histidine, L1CAM, Ly6a (includes others), MAL, NOS1, OGDHL, OGN, Pplc, PROTEASE, PTEN, PTPRT, PYGL, SEMA3A, sGC, SLC38A2, SLC02B1, SRC, sulfotransferase, SULT1C2, TBC1D24, Wap | 10 | 6 | Cellular Development, Connective Tissue Development and Function, Skeletal and Muscular System Development and Function |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



