
Submitted:
15 November 2023
Posted:
16 November 2023
You are already at the latest version
Abstract
Quercetin (QUA), a flavonoid compound, is ubiquitously found in plants and has demonstrated a diverse range of biological activities. The primary objective of the present study is to assess the potential antiemetic properties of QUA using an in vivo and in silico approach. In this experiment, emesis was induced in 4-day-old chicks by orally administering copper sulfate pentahydrate (CuSO4.5H2O) at a dose of 50 mg/kg (orally). Domperidone (6 mg/kg), ondansetron (5 mg/kg), and hyoscine (21 mg/kg) were treated as positive controls (PCs), and distilled water was employed as a negative control (NC). QUA was administered orally at two distinct doses (25 and 50 mg/kg). Additionally, QUA (50 mg/kg) and PCs were administered separately or in combination to assess their antagonistic or synergistic effects on the chicks. The binding affinity of QUA and referral ligands towards 5HT3, D2, D3, and M1–M5 receptors was estimated, and ligand-receptor interactions were visualized through various computational tools. In vivo findings indicate that QUA (25 and 50 mg/kg) has a significant effect on reducing the number of retches (16.50 ± 4.65 and 10.00 ± 4.19 times) and increasing the chick latency period (59.25 ± 4.75 and 94.25 ± 4.01 seconds), respectively. Additionally, QUA (50 mg/kg) in combination with domperidone and ondansetron exhibited superior antiemetic effects, reducing the number of retches and increasing the onset of emesis-inducing time. Furthermore, it is worth noting that QUA demonstrated the strongest binding affinity towards the D2 receptor with a value of -9.7 kcal/mol through the formation of hydrogen and hydrophobic bonds. In summary, the study found that QUA exhibited antiemetic activity in chicks, potentially by interacting with the D2 receptor pathway.
Keywords:
Quercetin
; antiemetic
; molecular docking
; D2 receptor
; toxicity
1. Introduction
Emesis, commonly referred to as vomiting, is a well-recognized physiological reflex that can be triggered by various peripheral and central stimuli [1]. These stimuli encompass gastrointestinal irritation or inflammation, the presence of cancer chemotherapy, circulating toxins and other pharmacological agents, pain, anxiety, and motion that provokes a response [2]. The defensive reflex is a commonly observed phenomenon in both animals and humans. In contrast, nausea is a subjective experience characterized by a disagreeable sensation closely linked to the act of vomiting [3].
The pathophysiology of vomiting is characterized by a significant degree of complexity. The reticular formation encompasses a vomiting center (VC) which can be stimulated by afferent stimuli originating from the gastrointestinal tract or by the chemoreceptor trigger zone (CTZ) [4,5]. The latter is commonly regarded as the primary site for the induction of emesis resulting from the administration of various cancer chemotherapeutic agents and toxic compounds [6,7]. Some pharmaceutical agents that are incapable of traversing the blood-brain barrier can be employed to specifically target it [8]. Besides the CTZ, various anatomical sites such as the gastrointestinal tract (GIT), higher cortical centers, the vestibular system, and the thalamus have been recognized as consistent stimuli for the initiation of emesis [9]. The ventral VC located in the reticular formation can be stimulated through the convergence of afferent stimuli originating from the GIT or by the CTZ [10,11]. Moreover, the VC assumes a crucial function in orchestrating the contractions of smooth muscles and skeletal activities linked to the emetic process [12,13]. The close proximity between the nucleus tractus solitaries (NTS) and the CTZ may elicit a reaction at the CTZ in the presence of emetogenic chemicals in the bloodstream or cerebrospinal fluid (CSF) [14,15]. Furthermore, the CTZ plays a regulatory role in various physiological processes, including the control of food intake, conditioned taste aversion, and GIT motility [16,17]. The initiation of emesis occurs when the CTZ detects emetogenic toxins in the bloodstream and CSF [18]. This detection is facilitated by various receptors within the CTZ, such as, serotonin receptor (5HT3), muscarinic acetylcholine receptors (M1-M4), dopamine receptors (D2, D3), histamine (H1), neurokinin 1 receptor (NK1) for substance P and opioid receptors [19,20,21]. Once these receptors detect the emetogenic toxins, they transmit a signal to the nearby NTS [22,23]. The NTS functions as the principal mechanism by which all emetic stimuli elicit the physiological response of vomiting, providing a comprehensive final pathway. During the act of emesis, the abdominal muscles undergo a retrograde contraction while the lower esophageal sphincter relaxes, facilitating the expulsion of stomach contents through the oral cavity and inducing retching [24,25].
Dopamine receptors are vital to the performance of several critical processes in everyday life. There are five distinct subtypes of dopamine receptors [26]. The D2 receptor has a role in several cognitive and behavioral processes, including movement, attention, sleep, memory, and learning [27]. The D2 receptor plays a vital role in the physiology of eliciting emesis [28]. The precise signaling processes behind the induction of vomiting by the D2 receptor have yet to be elucidated. The research evidence suggests that the activation of extracellular signal-regulated kinase (ERK), phosphatidylinositol-3 kinases (PI3K), and protein kinase C (PKC)-related signaling cascades via the D2 receptor might possibly lead to the occurrence of emesis [29,30].
At present, a diverse range of antiemetic medications that are approved for clinical use are utilized in the treatment and control of symptoms associated with nausea and vomiting. The medications can be classified into various categories, such as anti-dopaminergic drugs, serotonin antagonists, antihistamines, NK1-receptor inhibitors, anticholinergic drugs, and receptor agonists for corticosteroids, GABAB, and cannabinoid receptor type 1 (CB1) [31,32]. The extended utilization of synthetic antiemetic medications is also associated with various adverse effects, such as muscle weakness, spasms, or convulsions [33,34]. Hence, natural products have become a crucial requirement in contemporary society owing to their minimal adverse effects and economic advantages [10,35]. Current efforts in the exploration of new antiemetic drugs derived from natural sources are primarily centered around mechanism-based strategies that target specific cellular and molecular mechanisms. Polysaccharides, flavonoids, alkaloids, diterpenes, glucosides, cannabinoids, diarylheptanoids, chalcones, saponins, hydroxycinnamic acids, phenylpropanoids, terpenes, and lignans encompass a wide range of bioactive compounds that are of interest in the exploration of potential candidates for the development of antiemetic drugs [10,17].
The compound known as Quercetin (QUA), with its IUPAC name being 2-(3, 4-dihydroxy phenyl)-3,5,7-trihydroxychromen-4-one, is a polyphenolic flavonoid that exhibits a broad distribution. A diverse array of fruits, vegetables, grains, and leaves, such as grapes, green tea, apples, citrus fruits, berries, cherries, capers, leafy greens, kale, and red onions, are known to contain compounds that exhibit a broad spectrum of health-promoting effects in relation to various diseases [36,37,38]. QUA possess a wide range of applications in the production of nutritional supplements, beverages, and food products [39]. According to numerous studies, QUA has been found to possess various pharmacological activities, including but not limited to anticancer [40], antioxidant [41], anti-obesity [42], anti-aging [43], anti-inflammatory [44], antidiabetic [45], antiallergic [46], cardioprotective [47], immune modulatory [48], neuroprotective [49], antiviral, and antimicrobial effects [50]. In a study conducted by Lee et al. (2005), the researchers examined the impact of QUA on the activity of the mouse 5HT3 channel, a receptor known to be involved in the physiological process of vomiting [51].
On the other hand, the utilization of computational methodologies in the field of drug discovery and development facilitates the efficient examination of a large collection of compounds and the anticipation of potential binding agents through the application of modeling, simulation, and visualization techniques. Furthermore, this approach aids in the anticipation of pharmacokinetics, toxicity, and binding sites, all of which play a pivotal role in elucidating mechanistic processes and identifying and cultivating promising drug candidates [52,53]. The primary objective of this study is to evaluate the potential antiemetic effect of QUA in chick (in vivo). Additionally, we also carried out an in silico study to identify potential molecular interactions with the emesis inducing receptors underlying the observed effect.
2. Materials and Methods
2.1. Chemical reagents and standards
The compound QUA [2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one], with a purity of 98%, extrapure grade yellow color, and a CAS number of 6151-25-3, was acquired from Sisco Research Laboratories Pvt. Ltd (India). Copper sulfate pentahydrate (CuSO4.5H2O) was obtained from Merck (India). The reference/standard drugs ondansetron (OND), domperidone (DOM), and hyoscine butyl bromide (HYS) were obtained from Incepta Pharma. Ltd., Beximxo Pharma. Ltd., and Opsonin Pharma. Ltd., respectively, all located in Bangladesh.
2.2. Animals
Young chickens (Gallus gallus domesticus) of both genders, aged 3 days, with an average weight ranging from 40-48 gm (Grade-A), were obtained from Nourish Poultry & Hatchery Ltd., located on Sonargaon Janapath Road, Uttara, Dhaka-1230, Bangladesh. The chickens were housed in group in stainless steel enclosures that featured an upper hood opening. The enclosures were maintained at room temperature and subjected to a twelve-hour light and dark cycle. The individuals were granted unrestricted access to regular provisions of food and water. An additional day was allocated for preparatory activities prior to the commencement of the experiment, following the completion of data collection from suppliers. Following 12 hours of fasting, the antiemetic test was carried out. This study was approved and funded by the Bangabandhu Sheikh Mujibur Rahman Science and Technology University Research Center (BSMRSTU-RC) (Approval No. 2023-33).
2.3. In vivo investigation
2.3.1. CuSO4.5H2O-induced emesis model in broiler chicks
The Investigation was carried out with slight modifications to the methodologies outlined by Akita et al. (1998) [54]. All the chicks were allocated by dividing them into nine groups, each consisting of five individuals. Before receiving the treatments, each chick specimen was housed in a spacious, transparent receptacle made of plastic for 10 minutes. Two different doses (25 and 50 mg/kg) of the test sample QUA were prepared by dissolving it in distilled water (DW) and subsequently administered orally based on information obtained from a literature review. The positive control (PC) drugs, namely DOM, OND, and HYS were administered orally at doses of 6, 5, and 21 mg/kg body weight (b.w.), respectively. Three doses of the reference drugs were prepared by combining them with QUA at 50 mg/kg. These combined doses were then administered orally to animals to assess any potential synergistic effects. This study designated the compound DW as the negative control (NC). It was administered orally at 150 mg/kg b.w. Following a treatment duration of 30 minutes, emesis was induced in each chick specimen through the oral administration of CuSO4.5H2O at a dosage of 50 mg/kg of b.w. Subsequently, the latency period, defined as the time interval between the administration of CuSO4.5H2O treatment and the occurrence of the first retch, as well as the total number of retches within a 10-minute timeframe following the administration of CuSO4.5H2O treatment, was meticulously documented. The equations used to calculate the percentage increase in latency and decrease in retches inspection of NC were as follows:
Where, B = Mean of latency in seconds in standard and test groups, C = Mean of latency in seconds in NC group, X = Mean of retches in NC group, Y = Mean of retches in standard and test groups
2.4. Statistical analysis
The antiemetic activity values are presented as the mean value accompanied by the standard error of the mean (SEM). The statistical significance of the difference was calculated using Graph Pad Prism (version 6.0), a statistical software program. A 95% confidence interval was employed for this determination. Significance was attributed to p-values less than 0.05, while p-values less than 0.0001 were deemed highly significant.
2.5. In silico investigation
2.5.1. Receptors selection and preparation
The literature review had identified 8 receptors that were implicated in the induction of emesis. The three-dimensional (3D) structures of the targeted receptors, namely 5HT3 (PDB ID: 6Y5B), D2 (PDB ID: 6CM4), D3 (PDB ID: 3PBL), M1 (PDB ID: 6WJC), M2 (PDB ID: 5ZK8), M3 (PDB ID: 4U15), M4 (PDB ID: 7V6A), M5 (PDB ID: 6OL9) were obtained from the Protein Data Bank (https://www.rcsb.org/). Following the collection process, the receptors were subjected to optimization procedures aimed at mitigating potential docking interference. This was carried out by eliminating extraneous molecules, such as lipids, water molecules, and heteroatoms, from the protein sequence using the PyMol software package (version 2.4.1) [55]. The energy minimization and geometry optimization of the receptors were conducted using the Swiss-PDB Viewer software package, employing the GROMOS96 force field [56]. The resulting Protein Data Bank (PDB) file was saved for subsequent molecular docking analysis.
2.5.2. Ligand collection and preparation
The 3D conformers of DOM (Compound CID: 3151), OND (Compound CID: 4595), HYS (Compound CID: 3000322), and QUA (Compound CID: 5280343) were obtained in structure-data file (SDF) format from the PubChem chemical database (https://pubchem.ncbi.nlm.nih.gov/). Subsequently, the 3D conformers of the chemical agents were optimized to minimize internal energies using the Chem3D 16.0 program package, which is commonly employed for molecular docking studies [57,58]. Figure 1 exhibits the visual representations of the chemical agents in a two-dimensional (2D) format.
2.5.3. Molecular docking protocol
Molecular docking is a computational tool employed in the field of medicinal chemistry for the purpose of drug design. The estimation of the drugs' active binding potential against the active sites of receptors was conducted through the utilization of the PyRx software package in the process of molecular docking. To facilitate the docking process, the dimensions of the grid box were adjusted to their maximum values along the x-, y-, and z-axes. The calculation was then executed over a span of 2000 steps [59,60]. The outcome of the docking potential is stored in the comma-separated values ('csv') file format, while the ligand-protein complex is obtained in PDB format in order to gather the ligand in PDBQT format. The ligand-receptor interactions and their corresponding active sites were examined using the Discovery Studio Visualizer (v21.1.020298) and PyMol (v2.4.1) software packages. Subsequently, the amino acid (AA) residues, bond types, number of hydrogen bond (HB), length of HBs and other bonds type for each ligand-receptor interaction were documented [61,62].
2.5.4. Toxicity prediction
The ProTox-II web servers were employed to predict toxicity parameters. The objective of the Protox II web server was to analyze a range of parameters associated with hepatotoxicity, carcinogenicity, immunotoxicity, mutagenicity, and cytotoxicity [63,64]. The chemical agent’s data obtained from PubChem, including SMILES (Simplified Molecular Input Line-Entry System) representations, was inputted into the search bars of Protox II for analysis.
3. Results
3.1. In vivo investigation
3.2. CuSO4.5H2O-induced emesis test
In the in vivo experiment, it was observed that administering two distinct doses of QUA resulted in a significant dose-dependent elevation in the onset of retching in the experimental chickens, as compared to the NC group. The onset periods for NC group were 8.25 ± 2.88 seconds. Simultaneously, the latency of QUA-25 and QUA-50 groups were 59.25 ± 4.75 and 94.25 ± 4.01 seconds, respectively. On the contrary, the different PC groups (DOM, OND, and HYS) administering to the animals, demonstrated a higher latency period comparing to the NC but reduced than the highest dose of test group (QUA-50) and the latency periods are 61.00 ± 3.34, 16.17±3.64, and 11.50 ± 2.38 seconds, for the standard groups DOM, OND, and HYS, respectively. It is noteworthy that combining QUA with PC resulted in a higher latent period than administering PC or the different test doses alone. The highest latency period was observed in the DOM + QUA-50 group and the animal in this group delayed initiating the retching by 156.67 ± 2.40 seconds. The onset of retching for the other two combined groups was noted at 146.33 ± 4.09, and 128.20+4.15 seconds, for the OND + QUA-50 and HYS + QUA-50 groups respectively. Figure 2 illustrates the latency observed across all treatment groups.
The NC group exhibited the highest number of retches, with a mean value of 64.25 ± 3.69. The frequency of retching in the experimental group decreased progressively as the dosage of the test sample (QUA) increased. For instance, QUA-25 and QUA-50 demonstrated 16.50 ± 4.65 and 10.00 ± 4.19 retches, respectively, this is a dose-dependent response. On the other hand, the animal taken standard drugs also revealed a significant diminishing in the frequency of retching and we observed the mean of retching of 11.80 ± 2.09, 30.80 ± 4.04, and 48.83 ± 4.17 for DOM, OND, and HYS respectively. The administration of combination drug therapy remarkably decreased the frequency of retches compared to the administration of the drug alone, both the test compound and standard. The DOM + QUA-50 manifested the lowest number of retches, with a mean value of 6.00 ± 2.52. The other two combination therapies also revealed a lower number of retches with the mean value of 8.33 ± 1.20 and 12.2 ± 2.15 for OND + QUA-50 and HYS + QUA-50, respectively. Figure 3 displays the total count of retches observed across all treatment groups.
In comparison of the results among the different treatment groups, we found that the highest percentage increase in latency was 94.73% for the DOM + QUA-50 group. A dose-dependent elevation of the increase in latency was observed in the test groups (QUA-25 and QUA-50). The fasted onset of retching was observed in the HYS group, and animal treatment with HYS revealed a 28% increase in latency, indicating the most vital effect of CuSO4.5H2O among the different PC and test groups. As with latency, we also calculated the highest protection from the effect of retching in the DOM + QUA-50, and animals in this group revealed a 90.66% decrease in retches. The test sample demonstrated a dose-dependent percentage decrease in retching. QUA-25 and QUA-50 expressed 74.32% and 84.43% percentage decreases in retches, respectively. The percentage increase in latency and percentage decrease in retching of all the treatment groups are provided in Table 1.
3.3. In silico investigation
3.3.1. QUA and DOM with dopaminergic (D2 and D3) receptor interaction
Both QUA and standard DOM exhibited higher binding affinity toward the D2 receptor (Table 2). The binding affinities of QUA for the D2 and D3 receptors were -9.7 and -8.5 kcal/mol, respectively. The interaction between QUA and the D2 receptor involved the formation of four HBs with specific AA residues, namely SER193 (1.85 Å), TYR416 (2.20 Å), THR119 (2.14 Å), and CYS118 (2.18 Å). Additionally, QUA formed three hydrophobic (HP) bonds with ASP114, TRP386, and PHE390. In contrast, it is worth noting that DOM exhibited binding affinities with D2 and D3 receptors at energy levels of -9.8 and -9.4 kcal/mol, respectively. The DOM molecule interacted with the D2 receptor by forming a single HB with the AA residue SER409 (1.89 Å). Furthermore, DOM formed six HP interactions with ASP114, PHE389, VAL91, LEU94, TYR408, and TRP413. Figure 4 illustrates the 2D and 3D structures of the non-bond interactions between QUA and DOM with the D2 receptor.
3.3.2. QUA and OND with serotoninergic (5HT3) receptor interaction
For the 5-HT3 receptor, QUA exhibited a binding affinity of -7.9 kcal/mol, while the standard OND compound showed a binding affinity of -8.1 kcal/mol. QUA's binding interaction with the 5-HT3 receptor was facilitated by two HBs with specific AA residues, namely ILE267 (2.51 Å) and ASP271 (2.95 Å), in addition to HP bonding interactions with ILE268, LEU266, LEU234, and PRO230. On the other hand, OND's binding interaction with the 5-HT3 receptor was facilitated by two HP bonds with specific AA residues, namely LEU234 and VAL264 (Table 3). Figure 5 illustrates the 2D and 3D structures of the non-bond interactions between QUA and OND with the 5HT3 receptor.
3.3.3. QUA and HYS with muscarinic (M1, M2, M3, M4, and M5) receptor interaction
Both QUA and standard HYS exhibited higher affinity with the M4 receptor (Table 4). The binding affinities of QUA for the M1, M2, M3, M4, and M5 receptors were -7.8, -8.2, -9.0, -9.2, and -8.0 kcal/mol, respectively. The interaction between QUA and the M4 receptor involved the formation of two HBs with specific AA residues, namely TYR416 (2.54 Å) and SER436 (2.06 Å). Additionally, QUA formed two HP bonds with PHE186 and TRP435. In contrast, it is worth noting that HYS exhibited binding affinities with M1, M2, M3, M4, and M5 receptors at energy levels of -7.1, -7.5, -8.6, -8.9, and -7.7 kcal/mol, respectively. The HYS compound interacted with the M4 receptor by forming three HBs with TYR92 (2.81 Å), ASP432 (2.71 Å), and ASN423 (3.70 Å). Furthermore, HYS formed three HP interactions with TYR439, PHE186, and TRP435. Figure 6 illustrates the 2D and 3D structures of the non-bond interactions between QUA and HYS with the M4 receptor.
3.3.4. In silico toxicity predictions
The web server ProTox-II was utilized to predict toxicity parameters. In our in silico toxicity study, QUA exhibited an LD50 of 159 mg/kg body weight with toxicity class 3. However, QUA showed no toxicity effects on hepatotoxicity, immunotoxicity, and cytotoxicity. Nevertheless, QUA showed toxicity effects in terms of carcinogenicity and mutagenicity. In contrast, it has been observed that standard compounds, namely DOM and OND, exhibited immunotoxicity and mutagenicity toxicity, respectively. On the other hand, it has been determined that HYS does not demonstrate toxicity across all assessed parameters.

Table 5.
The toxicity prediction of four compounds, namely QUA, DOM.OND, and HYS, using the ProTox-II model.
Table 5.
The toxicity prediction of four compounds, namely QUA, DOM.OND, and HYS, using the ProTox-II model.
| Properties | Parameters | QUA | DOM | OND | HYS |
|---|---|---|---|---|---|
| Toxicity | LD50 | 159 mg/kg | 715 mg/kg | 95 mg/kg | 1,275 mg/kg |
| Toxicity class | 3 | 4 | 3 | 4 | |
| Hepatotoxicity | Inactive | Inactive | Inactive | Inactive | |
| Carcinogenicity | Active | Inactive | Inactive | Inactive | |
| Immunotoxicity | Inactive | Active | Inactive | Inactive | |
| Mutagenicity | Active | Inactive | Active | Inactive | |
| Cytotoxicity | Inactive | Inactive | Inactive | Inactive |
4. Discussion
Inadequate management of chemotherapy-induced nausea and vomiting (CINV) can significantly impact functional capabilities and may compromise treatment compliance [65]. The oral ingestion of toxic CuSO4.5H2O can lead to a distinct vagal emetic response due to its properties as an oxidizing agent and its corrosive effects on the gastrointestinal mucous membranes [66,67,68]. The induction of emesis occurs as a result of peripheral mechanisms, which involve the stimulation of visceral afferent nerve fibers in the gastrointestinal tract [69]. These stimuli are then transmitted to the vomiting center [70,71]. Recent findings demonstrated that 5HT3, D2, D3, H1, and M1-M5 receptors are present at the stimulation site and contribute to the induction of emesis. Antagonists of the 5HT3, D2, D3, H1, and M1-M5 receptors have the ability to prevent or significantly decrease emesis induced by chemotherapeutic agents [72,73].
The chosen standard drug, DOM, exhibited peripheral selectivity as an antagonist for D2 and D3 receptors [74]. Its mechanism of action involves the inhibition or antagonism of these receptors at the CTZ in the brain, thereby facilitating the desired therapeutic effects [75]. During our investigation, the group DOM ingestion displayed a mean value of 11.80 ± 2.09 retches in chicks, while the mean value of retches in the NC group was 64.25 ± 3.69. The administration of OND and HYS decreased the frequency of retching episodes in the chick group compared to the control group receiving the vehicle. Based on empirical findings, it is possible to postulate that QUA demonstrated a safeguarding influence against toxicity by mitigating or preventing neural signals responsible for eliciting emetic responses. The significant reduction in the frequency of retches observed in both QUA groups compared to the NC group supports this notion, with the obtained mean values being 16.50 ± 4.65 and 10.00 ± 4.19 for the QUA-25 and QUA-50 groups, respectively. These values are close to or better than the standard groups. On the contrary, the different PC groups (DOM, OND, and HYS) administered to the animals demonstrated longer latency periods compared to the NC group. Specifically, the latency periods were 61.00 ± 3.34, 16.17 ± 3.64, and 11.50 ± 2.38 seconds for DOM, OND, and HYS, respectively. Interestingly, the treatment group (QUA-50) exhibited the highest latency period among all PC the groups. The result shows that QUA-50 is more effective in reducing retches and prolonging latency periods compared to the standard DOM, OND, and HYS groups in CuSO4.5H2O-mediated emesis.
In the field of pharmacology, the phenomenon where the combined impact of multiple medications surpasses the individual effects observed when each drug is administered in isolation is referred to as a synergistic effect or synergism [76]. Our experimental study demonstrated that using a combined drug therapy resulted in a reduction in the occurrence of retches and an increase in the duration of the latency period in chicks, indicating the presence of a synergistic effect. Previous research has shown that administering antiemetic drugs effectively delays the occurrence of nausea or vomiting in response to emetic stimuli induced by cancer chemotherapy or acute toxicity [77]. Our study observed that the test combined group (DOM + QUA 50) displayed a significantly prolonged latency period of (156.67 ± 2.40) seconds compared to the NC group.
In a study conducted by Wang and Borison (1951) and Niijima et al. (1987), it was observed that CuSO4.5H2O does not exhibit the expected response to vagal nerve stimulation. Specifically, the researchers observed that vagotomy could not prevent emesis, a procedure that involves severing the distal portion of the vagus nerve in the gastrointestinal tract [78,79]. This suggests that the emetic response to CuSO4.5H2O may involve chemoreceptor signaling. Figure 7 illustrates a possible antiemetic mechanism of QUA and standard drugs.
Molecular docking is a computational methodology employed to investigate the compatibility of a ligand with a receptor binding site, taking into consideration both geometric and energetic factors [80,81,82]. The estimation of the interaction level between a ligand and a receptor is accomplished by assessing their binding affinity [83]. The binding interactions between QUA and the D2 receptor were higher compared to other receptors involved in the induction of emesis. The binding energy of QUA with D2 was -9.7 kcal/mol, whereas the binding energy of standard DOM was -9.8 kcal/mol. The drug-receptor interaction visualization indicates that the binding sites for QUA and DOM were SER193, TYR416, THR119, CYS118, ASP114, TRP386, and PHE390 for QUA, and SER409, ASP114, PHE389, VAL91, LEU94, TYR408, and TRP413 for DOM. Multiple studies have consistently provided evidence indicating that the activation of D2 receptors triggers the vomiting center located in the CTZ [84,85,86]. QUA effectively inhibits the response of the D2 receptor, thereby impeding dopamine activity. Therefore, it can be confidently concluded that QUA exhibits strong inhibitory potency for the D2 receptor compared to other receptors responsible for emesis. This conclusion is supported by the higher docking scores of QUA with D2 receptors compared to other receptors. Additionally, in vivo, combined therapy with DOM+QUA-50 had shown higher efficacy than other combinations.
Drug discovery and development is a protracted, financially burdensome, and precarious undertaking, spanning 10 to 15 years on average [87]. The approval of a novel pharmaceutical for clinical application entails an average expenditure exceeding $1 to $2 billion [88]. The drug candidate undergoes rigorous optimization during the preclinical stage before progressing to the phase I clinical trial [89]. Approximately 90% of drug candidates fail during the progression of clinical studies, specifically during phase I, II, and III clinical trials, primarily due to issues related to toxicity and a lack of clinical efficacy [90,91,92]. In silico toxicological investigations play a crucial and significant role in safe and cost-effective drug development [93]. In our present study, QUA demonstrated no toxic effects on hepatotoxicity, immunotoxicity, and cytotoxicity. However, it did exhibit toxicity effects in terms of carcinogenicity and mutagenicity.
The study indicates that QUA exhibits significant antiemetic effects in response to CuSO4.5H2O-induced emesis, potentially attributed to its ability to antagonize D2 receptors. The in vivo findings also suggest that the antiemetic efficacy of QUA remains consistent and dependable when administered at a reduced dosage.
Studies using certain laboratory animals provide necessary and important data regarding the beneficial and harmful effects of new drug candidates and their possible biopharmaceutical considerations [94]. Thus, each pre-clinical study helps medicinal scientists evaluate biologically active molecules potential for clinical studies. These studies allow for the determination of test dose and dosage frequency, the right administration routes, the drug metabolism profile, and the development of error correction machines during clinical trials. In this study, we have seen that all standard anti-emetic drugs clearly inhibit the emesis tendency in animals. The test sample's QUA also exerted dose-dependent anti-emetic effects in animals. In comparison to the control and standard groups, QUA exerted strong anti-emetic effects in animals. Although this well known drug candidate is yet to undergo extensive toxicological studies in animal models, in this study QUA treatment did not show any toxicological phenomena, nor did it cause the death of any chicks, which demonstrates its safety in this animal model. Moreover, our in-vivo findings also corroborate the outcomes of in-silico studies, demonstrating the possibility of considering this bioactive in the emesis of laboratory animals.
The major limitation of this study is that the findings may be influenced by various factors, such as the differences in physiology and geometry of the animal’s stomach and the digestive capability of foods taken before the test started, which also impact emesis induction. Another possible limitation is the placement of the gavage tube at the time of inducer (copper sulfate) delivery in the gastric compartment; therefore, it is difficult to determine the specific chemical placement and timing of movement of the emetic agent along the GI tract. In addition, this is a physiologic behavioral experiment; in this case, all animals would not respond similarly due to the variance of the environment, such as laboratory light, noise during the experiment, and test time differences, though we followed the optimum laboratory protocols mentioned in the study design section.
5. Conclusions
The findings of this study provide evidence that QUA exhibits notable antiemetic properties and effectively mitigates CuSO4.5H2O-induced retching in chicks, potentially through its peripheral mechanism of action. The results of the molecular docking study confirm that QUA has a higher affinity for dopamine receptors, particularly D2 receptors, compared to other receptors known to induce emesis. Our test compound, QUA, also shows synergistic effects when administered in conjunction with established antiemetic drugs. So, the compound QUA might be a good option in the treatment of emesis. Similarly, the computational toxicity (A Random Forest (RF) algorithm method) analysis reveals that QUA demonstrates toxic properties in relation to carcinogenicity and mutagenicity. Further animals and more desirable study (human organ on chips) are required to check validity of the toxicity of QUA. The implications of these findings extend to future non-clinical, preclinical, and clinical investigations, necessitating further in vivo studies to validate the observed outcomes. Researchers in the field of medicinal chemistry must be encouraged to conduct thorough investigations on this highly promising natural lead compound and its various derivatives.
Author Contributions
Conceptualization, R.C., M.S.B., and M.T.I.; data curation, M.S.B. and R.H; Methodology A.I.R, R.H., H.D.M.C.; formal analysis; M.S.B., A.I.R., H.D.M.C. and I.M.A.; investigation, M.S.B., R.H., and R.C., resources, A.I.R. and MT.I., software, H.D.M.C. and I.R.A.d.M; validation, I.M.A. and I.R.A.M.; visualization, A.I.R., M.S.B., H.D.M.C., and R.C., writing-original draft, R.C., and M.S.B., writing review & editing, R.C., H.D.M.C., I.R.A.d.M., A.I.M. and M.T.I., supervision, M.T.I., project administration, I.R.A.d.M. and M.T.I.
Funding
This study was approved and funded by the BSMRSTU Research Center (BSMRSTU-RC) (Approval No. 2023-33).
Institutional Review Board Statement
This study was approved and funded by the Bangabandhu Sheikh Mujibur Rahman Science and Technology University Research Center (BSMRSTU-RC) (Approval No. 2023-33).
Informed Consent Statement
Not applicable.
Acknowledgments
Researchers Supporting Project number (RSPD2023R744), King Saud University, Riyadh, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhong, W.; Shahbaz, O.; Teskey, G.; Beever, A.; Kachour, N.; Venketaraman, V.; Darmani, N.A. Mechanisms of Nausea and Vomiting: Current Knowledge and Recent Advances in Intracellular Emetic Signaling Systems. Int. J. Mol. Sci. 2021, 22, 5797. [Google Scholar] [CrossRef]
- de la Puente-Redondo, V.; TINGLEY III, F.D.; Schneider, R.P.; Hickman, M.A. The Neurokinin-1 Antagonist Activity of Maropitant, an Antiemetic Drug for Dogs, in a Gerbil Model. J. Vet. Pharmacol. Ther. 2007, 30, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Balaban, C.D.; Yates, B.J. What Is Nausea? A Historical Analysis of Changing Views. Auton. Neurosci. 2017, 202, 5–17. [Google Scholar] [CrossRef] [PubMed]
- MacDougall, M.R.; Sharma, S. Physiology, Chemoreceptor Trigger Zone. In StatPearls [Internet]; StatPearls Publishing, 2022. [Google Scholar]
- Iqbal, I.M.; Spencer, R. Postoperative Nausea and Vomiting. Anaesth. Intensive Care Med. 2012, 13, 613–616. [Google Scholar] [CrossRef]
- Kobrinsky, N.L. Regulation of Nausea and Vomiting in Cancer Chemotherapy. A Review with Emphasis on Opiate Mediators. Am. J. Pediatr. Hematol. Oncol. 1988, 10, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Dodds, L.J. The Control of Cancer Chemotherapy-induced Nausea and Vomiting. J. Clin. Pharm. Ther. 1985, 10, 143–166. [Google Scholar] [CrossRef]
- Salunkhe, S.S.; Bhatia, N.M.; Kawade, V.S.; Bhatia, M.S. Development of Lipid Based Nanoparticulate Drug Delivery Systems and Drug Carrier Complexes for Delivery to Brain. J. Appl. Pharm. Sci. 2015, 5, 110–129. [Google Scholar] [CrossRef]
- Becker, D.E. Nausea, Vomiting, and Hiccups: A Review of Mechanisms and Treatment. Anesth. Prog. 2010, 57, 150–157. [Google Scholar] [CrossRef]
- Bhuia, M.S.; Kamli, H.; Islam, T.; Sonia, F.A.; Kazi, M.A.; Siam, M.S.H.; Rahman, N.; Bappi, M.H.; Mia, M.N.; Hossen, M.M. Antiemetic Activity of Trans-Ferulic Acid Possibly through Muscarinic Receptors Interaction Pathway: In Vivo and in Silico Study. Results Chem. 2023, 101014. [Google Scholar] [CrossRef]
- Gan, T.J. Mechanisms Underlying Postoperative Nausea and Vomiting and Neurotransmitter Receptor Antagonist-Based Pharmacotherapy. CNS Drugs 2007, 21, 813–833. [Google Scholar] [CrossRef]
- Shames, B. Anatomy and Physiology of the Duodenum. In Shackelford’s Surgery of the Alimentary Tract, 2 Volume Set; Elsevier, 2019; pp. 786–803. [Google Scholar] [CrossRef]
- 1Khan, I.A.; Aziz, A.; Sarwar, H.S.; Munawar, S.H.; Manzoor, Z.; Anwar, H. Evaluation of Antiemetic Potential of Aqueous Bark Extract of Cinnamon Loureiroi. Can. J. App. Sci 2014, 1, 26–32. [Google Scholar]
- Chang, T. Nausea and Vomiting. In Supportive Care in Pediatric Oncology: A Practical Evidence-Based Approach; Springer, 2014; pp. 159–175. [Google Scholar]
- Bhargava, K.P.; Dixit, K.S.; Gupta, Y.K. Enkephalin Receptors in the Emetic Chemoreceptor Trigger Zone of the Dog. Br. J. Pharmacol. 1981, 72, 471–475. [Google Scholar] [CrossRef]
- Andrews, P.L.R.; Sanger, G.J. Nausea and the Quest for the Perfect Anti-Emetic. Eur. J. Pharmacol. 2014, 722, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Hasan, M.M.; Ahmed, S.W. Natural Antiemetics: An Overview. Pak. J. Pharm. Sci 2014, 27, 1583–1598. [Google Scholar]
- Denholm, L.; Gallagher, G. Physiology and Pharmacology of Nausea and Vomiting. Anaesth. Intensive Care Med. 2018, 19, 513–516. [Google Scholar] [CrossRef]
- McKarns, S.C. A Review of Neuroreceptors for Clinical and Experimental Neuropharmacology in Central Nervous System Disorders. Curr. Rev. Clin. Exp. Pharmacol. Former. Curr. Clin. Pharmacol. 2023, 18, 192–241. [Google Scholar] [CrossRef]
- Navari, R.M. Pharmacological Management of Chemotherapy-Induced Nausea and Vomiting: Focus on Recent Developments. Drugs 2009, 69, 515–533. [Google Scholar] [CrossRef] [PubMed]
- Naylor, R.J.; Inall, F.C. The Physiology and Pharmacology of Postoperative Nausea and Vomiting. Anaesthesia 1994, 49, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.D.; Leslie, R.A. The Area Postrema and Vomiting. Front. Neuroendocrinol. 1994, 15, 301–320. [Google Scholar] [CrossRef]
- Hornby, P.J. Central Neurocircuitry Associated with Emesis. Am. J. Med. 2001, 111, 106–112. [Google Scholar] [CrossRef]
- Holmes, A.M.; Rudd, J.A.; Tattersall, F.D.; Aziz, Q.; Andrews, P.L.R. Opportunities for the Replacement of Animals in the Study of Nausea and Vomiting. Br. J. Pharmacol. 2009, 157, 865–880. [Google Scholar] [CrossRef] [PubMed]
- Percie du Sert, N.; Andrews, P.L.R. The Ferret in Nausea and Vomiting Research: Lessons in Translation of Basic Science to the Clinic. Biol. Dis. Ferret 2014, 735–778. [Google Scholar] [CrossRef]
- Xu, P.; Huang, S.; Krumm, B. E.; Zhuang, Y.; Mao, C.; Zhang, Y.; Wang, Y.; Huang, X. P.; Liu, Y. F.; He, X.; Li, H.; Yin, W.; Jiang, Y.; Zhang, Y.; Roth, B. L.; Xu, H. E. Structural genomics of the human dopamine receptor system. Cell res. 2023, 33, 604–616. [Google Scholar] [CrossRef]
- Haubrich, J.; Hagena, H.; Tsanov, M.; Manahan-Vaughan, D. Editorial: Dopaminergic control of experience encoding, memory and cognition. Front. Behave. Neurosci. 2023, 17, 1230576. [Google Scholar] [CrossRef] [PubMed]
- Bhuia, M. S.; Islam, T.; Rokonuzzman, M.; Shamsh Prottay, A. A.; Akter, F.; Hossain, M. I.; Chowdhury, R.; Kazi, M. A.; Khalipha, A. B. R.; Coutinho, H. D. M.; Islam, M. T. Modulatory effects of phytol on the antiemetic property of domperidone, possibly through the D2 receptor interaction pathway: in vivo and in silico studies. 3 Biotech 2023, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, L.; Darmani, N. A. Dopamine receptors in emesis: Molecular mechanisms and potential therapeutic function. Pharmacol. res. 2020, 161, 105124. [Google Scholar] [CrossRef] [PubMed]
- Mannoury la Cour, C.; Salles, M. J.; Pasteau, V.; Millan, M. J. Signaling pathways leading to phosphorylation of Akt and GSK-3β by activation of cloned human and rat cerebral D₂and D₃ receptors. Molecu. Pharmacol. 2011, 79, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Hasan, M.M.; Ahmed, S.W.; Mahmood, Z.A.; Azhar, I.; Habtemariam, S. Anti-Emetic Effects of Bioactive Natural Products. Phytopharmacology 2013, 4, 390–433. [Google Scholar]
- Horn, C.C. The Physiology of Vomiting. Nausea vomiting diagnosis Treat. 2017, 15–25. [Google Scholar] [CrossRef]
- McNicol, E.; Horowicz-Mehler, N.; Fisk, R.A.; Bennett, K.; Gialeli-Goudas, M.; Chew, P.W.; Lau, J.; Carr, D. Management of Opioid Side Effects in Cancer-Related and Chronic Noncancer Pain: A Systematic Review. J. Pain 2003, 4, 231–256. [Google Scholar] [CrossRef]
- Athavale, A.; Athavale, T.; Roberts, D.M. Antiemetic Drugs: What to Prescribe and When. Aust. Prescr. 2020, 43, 49. [Google Scholar] [CrossRef] [PubMed]
- Dzobo, K. The Role of Natural Products as Sources of Therapeutic Agents for Innovative Drug Discovery. Compr. Pharmacol. 2022, 408. [Google Scholar] [CrossRef]
- Muschietti, L. V; Ulloa, J.L.; Redko, F.D.C. The Role of Flavonoids as Modulators of Inflammation and on Cell Signaling Pathways. Nat. Prod. as Source Mol. with Ther. Potential Res. Dev. Challenges Perspect. 2018, 159–208. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef] [PubMed]
- Bhimanwar, R.; Kothapalli, L.; Khawshi, A. Quercetin as Natural Bioavailability Modulator: An Overview. Res. J. Pharm. Technol. 2020, 13, 2045–2052. [Google Scholar] [CrossRef]
- Lakhanpal, P.; Rai, D.K. Quercetin: A Versatile Flavonoid. Internet J. Med. Updat. 2007, 2, 22–37. [Google Scholar] [CrossRef]
- Vafadar, A.; Shabaninejad, Z.; Movahedpour, A.; Fallahi, F.; Taghavipour, M.; Ghasemi, Y.; Akbari, M.; Shafiee, A.; Hajighadimi, S.; Moradizarmehri, S. Quercetin and Cancer: New Insights into Its Therapeutic Effects on Ovarian Cancer Cells. Cell Biosci. 2020, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Hu, M.J.; Wang, Y.Q.; Cui, Y.L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef]
- Ahn, J.; Lee, H.; Kim, S.; Park, J.; Ha, T. The Anti-Obesity Effect of Quercetin Is Mediated by the AMPK and MAPK Signaling Pathways. Biochem. Biophys. Res. Commun. 2008, 373, 545–549. [Google Scholar] [CrossRef]
- Hu, D.; Gu, X.; Wang, G.; Zhou, Z.; Sun, L.; Pei, J. Performance and Mechanism of Lignin and Quercetin as Bio-Based Anti-Aging Agents for Asphalt Binder: A Combined Experimental and Ab Initio Study. J. Mol. Liq. 2022, 359, 119310. [Google Scholar] [CrossRef]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Sreedhar, R.; Giridharan, V. V; Watanabe, K. Molecular Targets of Quercetin with Anti-Inflammatory Properties in Atopic Dermatitis. Drug Discov. Today 2016, 21, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Vessal, M.; Hemmati, M.; Vasei, M. Antidiabetic Effects of Quercetin in Streptozocin-Induced Diabetic Rats. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2003, 135, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef] [PubMed]
- Ferenczyova, K.; Kalocayova, B.; Bartekova, M. Potential Implications of Quercetin and Its Derivatives in Cardioprotection. Int. J. Mol. Sci. 2020, 21, 1585. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Garrick, J.M.; Roquè, P.J.; Pellacani, C. Mechanisms of Neuroprotection by Quercetin: Counteracting Oxidative Stress and More. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Azeem, M.; Hanif, M.; Mahmood, K.; Ameer, N.; Chughtai, F.R.S.; Abid, U. An Insight into Anticancer, Antioxidant, Antimicrobial, Antidiabetic and Anti-Inflammatory Effects of Quercetin: A Review. Polym. Bull. 2023, 80, 241–262. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-H.; Jung, S.-M.; ngo Lee, J.-H.; Kim, J.-H.; Yoon, I.-S.; Lee, J.-H.; Choi, S.-H.; Lee, S.-M.; Chang, C.-G.; Kim, H.-C. Quercetin Inhibits the 5-Hydroxytryptamine Type 3 Receptormediated Ion Current by Interacting with Pre-Transmembrane Domain I. Mol. Cells (Springer Sci. Bus. Media BV) 2005, 20. [Google Scholar] [CrossRef]
- Sliwoski, G.; Kothiwale, S.; Meiler, J.; Lowe, E.W. Computational Methods in Drug Discovery. Pharmacol. Rev. 2014, 66, 334–395. [Google Scholar] [CrossRef]
- Singh, S.S. Preclinical Pharmacokinetics: An Approach towards Safer and Efficacious Drugs. Curr. Drug Metab. 2006, 7, 165–182. [Google Scholar] [CrossRef]
- Akita, Y.; Yang, Y.; Kawai, T.; Kinoshita, K.; Koyama, K.; Takahashi, K.; Watanabe, K. New Assay Method for Surveying Anti-Emetic Compounds from Natural Sources. Nat. Prod. Sci. 1998, 4, 72–77. [Google Scholar]
- El-Mageed, H.R.A.; Abdelrheem, D.A.; Ahmed, S.A.; Rahman, A.A.; Elsayed, K.N.M.; Ahmed, S.A.; El-Bassuony, A.A.; Mohamed, H.S. Combination and Tricombination Therapy to Destabilize the Structural Integrity of COVID-19 by Some Bioactive Compounds with Antiviral Drugs: Insights from Molecular Docking Study. Struct. Chem. 2021, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.U.; Zoete, V.; Michielin, O.; Guex, N. Defining and Searching for Structural Motifs Using DeepView/Swiss-PdbViewer. BMC Bioinformatics 2012, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Ali, M.T.; Shawan, M.M.A.K.; Sarwar, M.G.; Khan, M.A.K.; Halim, M.A. Halogen-Directed Drug Design for Alzheimer’s Disease: A Combined Density Functional and Molecular Docking Study. Springerplus 2016, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hossain, R.; Sarkar, C.; Hassan, S.M.H.; Khan, R.A.; Arman, M.; Ray, P.; Islam, M.T.; Daştan, S.D.; Sharifi-Rad, J.; Almarhoon, Z.M. In Silico Screening of Natural Products as Potential Inhibitors of SARS-CoV-2 Using Molecular Docking Simulation. Chin. J. Integr. Med. 2022, 28, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.A.; Abdelrahman, A.H.M.; Badr, E.A.A.; Almansour, N.M.; Alzahrani, O.R.; Ahmed, M.N.; Soliman, M.E.S.; Naeem, M.A.; Shawky, A.M.; Sidhom, P.A. Naturally Occurring Plant-Based Anticancerous Candidates as Prospective ABCG2 Inhibitors: An in Silico Drug Discovery Study. Mol. Divers. 2022, 26, 3255–3277. [Google Scholar] [CrossRef] [PubMed]
- AVS, S.K.; Sinha, S.; Donakonda, S. Virus-Host Interaction Analysis in Colorectal Cancer Identifies Core Virus Network Signature and Small Molecules. Comput. Struct. Biotechnol. J. 2022, 20, 4025–4039. [Google Scholar] [CrossRef]
- Dey, D.; Hossain, R.; Biswas, P.; Paul, P.; Islam, M.A.; Ema, T.I.; Gain, B.K.; Hasan, M.M.; Bibi, S.; Islam, M.T. Amentoflavone Derivatives Significantly Act towards the Main Protease (3CLPRO/MPRO) of SARS-CoV-2: In Silico Admet Profiling, Molecular Docking, Molecular Dynamics Simulation, Network Pharmacology. Mol. Divers. 2023, 27, 857–871. [Google Scholar] [CrossRef]
- Hossain, R.; Al-Khafaji, K.; Khan, R.A.; Sarkar, C.; Islam, M.S.; Dey, D.; Jain, D.; Faria, F.; Akbor, R.; Atolani, O. Quercetin and/or Ascorbic Acid Modulatory Effect on Phenobarbital-Induced Sleeping Mice Possibly through Gabaa and Gabab Receptor Interaction Pathway. Pharmaceuticals 2021, 14, 721. [Google Scholar] [CrossRef]
- Banerjee, P.; Eckert, A.O.; Schrey, A.K.; Preissner, R. ProTox-II: A Webserver for the Prediction of Toxicity of Chemicals. Nucleic Acids Res. 2018, 46, W257–W263. [Google Scholar] [CrossRef]
- Adane, F.; Assefa, W.; Alem, M.B.; Dessalegn, M. Sub-Chronic Toxicity of the Aqueous Leaf Extract of Ocimum Lamiifolium Hochst. Ex Benth on Biochemical Parameters and Histopathology of Liver and Kidney in Rats: In Vivo and in-Silico Toxicity Studies. BMC Complement. Med. Ther. 2023, 23, 30. [Google Scholar] [CrossRef] [PubMed]
- Brianna; Lee, S. H. Chemotherapy: How to Reduce Its Adverse Effects While Maintaining the Potency? Med. Oncol. 2023, 40, 88. [Google Scholar] [CrossRef]
- Gelberg, H. Pathophysiological Mechanisms of Gastrointestinal Toxicity. Compr. Toxicol. 2018, 139. [Google Scholar] [CrossRef]
- Wallig, M.A. Digestive System. In Fundamentals of Toxicologic Pathology; Elsevier, 2018; pp. 395–442. [Google Scholar]
- Horn, C.C.; Meyers, K.; Lim, A.; Dye, M.; Pak, D.; Rinaman, L.; Yates, B.J. Delineation of Vagal Emetic Pathways: Intragastric Copper Sulfate-Induced Emesis and Viral Tract Tracing in Musk Shrews. Am. J. Physiol. Integr. Comp. Physiol. 2014, 306, R341–R351. [Google Scholar] [CrossRef] [PubMed]
- Boeckxstaens, G.; Camilleri, M.; Sifrim, D.; Houghton, L.A.; Elsenbruch, S.; Lindberg, G.; Azpiroz, F.; Parkman, H.P. Fundamentals of Neurogastroenterology: Physiology/Motility–Sensation. Gastroenterology 2016, 150, 1292–1304. [Google Scholar] [CrossRef] [PubMed]
- Coluzzi, F.; Rocco, A.; Mandatori, I.; Mattia, C. Non-Analgesic Effects of Opioids: Opioid-Induced Nausea and Vomiting: Mechanisms and Strategies for Their Limitation. Curr. Pharm. Des. 2012, 18, 6043–6052. [Google Scholar] [CrossRef]
- Babic, T.; Browning, K.N. The Role of Vagal Neurocircuits in the Regulation of Nausea and Vomiting. Eur. J. Pharmacol. 2014, 722, 38–47. [Google Scholar] [CrossRef]
- Sanger, G.J.; Andrews, P.L.R. An Analysis of the Pharmacological Rationale for Selecting Drugs to Inhibit Vomiting or Increase Gastric Emptying during Treatment of Gastroparesis. Aliment. Pharmacol. Ther. 2023, 57, 962–978. [Google Scholar] [CrossRef]
- Khani, A.; Oskuyi, A.E.; Asghari, R.; Khalkhli, H.R.; Sharifi, H. Olanzapine Enhances the Effect of Conventional Drugs in Chemotherapy Inducing Nausea and Vomiting: A Randomized Clinical Trial. Casp. J. Intern. Med. 2022, 13, 356. [Google Scholar] [CrossRef]
- Mitchelson, F. Pharmacological Agents Affecting Emesis: A Review (Part I). Drugs 1992, 43, 295–315. [Google Scholar] [CrossRef]
- Jacoby, H.I. Safety Pharmacology and the GI Tract. Toxicol. Gastrointest. Tract 2018, 53–81. [Google Scholar]
- Kumar, D.; Jain, V.; Rai, B. Capturing the Synergistic Effects between Corrosion Inhibitor Molecules Using Density Functional Theory and ReaxFF Simulations-A Case for Benzyl Azide and Butyn-1-Ol on Cu Surface. Corros. Sci. 2022, 195, 109960. [Google Scholar] [CrossRef]
- Perwitasari, D.A.; Gelderblom, H.; Atthobari, J.; Mustofa, M.; Dwiprahasto, I.; Nortier, J.W.R.; Guchelaar, H.-J. Anti-Emetic Drugs in Oncology: Pharmacology and Individualization by Pharmacogenetics. Int. J. Clin. Pharm. 2011, 33, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.C.; Borison, H.L. Copper Sulphate Emesis: A Study of Afferent Pathways from the Gastrointestinal Tract. Am. J. Physiol. Content 1951, 164, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Niijima, A.; Jiang, Z.-Y.; Daunton, N.G.; Fox, R.A. Effect of Copper Sulphate on the Rate of Afferent Discharge in the Gastric Branch of the Vagus Nerve in the Rat. Neurosci. Lett. 1987, 80, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Krovat, E.M.; Steindl, T.; Langer, T. Recent Advances in Docking and Scoring. Curr. Comput. Aided. Drug Des. 2005, 1, 93–102. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular Docking and Structure-Based Drug Design Strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed]
- Bhuia, M.S.; Rokonuzzman, M.; Hossain, M.I.; Ansari, S.A.; Ansari, I.A.; Islam, T.; Al Hasan, M.S.; Mubarak, M.S.; Islam, M.T. Anxiolytic-like Effects by trans-Ferulic Acid Possibly Occur through GABAergic Interaction Pathways. Pharmaceuticals 2023, 16, 1271. [Google Scholar] [CrossRef] [PubMed]
- Plewczynski, D.; Łaźniewski, M.; Augustyniak, R.; Ginalski, K. Can We Trust Docking Results? Evaluation of Seven Commonly Used Programs on PDBbind Database. J. Comput. Chem. 2011, 32, 742–755. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, D.O.; Briggs, D.B.; Strominger, N.L. Mechanisms of Radiation-Induced Emesis in the Dog. Pharmacol. Ther. 1988, 39, 367–371. [Google Scholar] [CrossRef]
- Belkacemi, L.; Darmani, N.A. Dopamine Receptors in Emesis: Molecular Mechanisms and Potential Therapeutic Function. Pharmacol. Res. 2020, 161, 105124. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Morita, M.; Horii, A.; Nishiike, S.; Kitahara, T.; Uno, A. Neural Mechanisms of Motion Sickness. J. Med. Investig. 2001, 48, 44–59. [Google Scholar]
- Berdigaliyev, N.; Aljofan, M. An Overview of Drug Discovery and Development. Future Med. Chem. 2020, 12, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Blackstone, E.A.; Joseph, P.F. The Economics of Biosimilars. Am. Heal. drug benefits 2013, 6, 469. [Google Scholar]
- Ruggeri, B.A.; Camp, F.; Miknyoczki, S. Animal Models of Disease: Pre-Clinical Animal Models of Cancer and Their Applications and Utility in Drug Discovery. Biochem. Pharmacol. 2014, 87, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Schuster, D.; Laggner, C.; Langer, T. Why Drugs Fail-a Study on Side Effects in New Chemical Entities. Curr. Pharm. Des. 2005, 11, 3545–3559. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Gao, W.; Hu, H.; Zhou, S. Why 90% of Clinical Drug Development Fails and How to Improve It? Acta Pharm. Sin. B 2022, 12, 3049–3062. [Google Scholar] [CrossRef] [PubMed]
- Bhuia, M.S.; Aktar, M.A.; Chowdhury, R.; Ferdous, J.; Rahman, M.A.; Hasan, M.S.A.; Islam, M.T. Therapeutic potentials of ononin with mechanistic insights: A comprehensive review. Fo. Biosci. 2023, 2023, 103302. [Google Scholar] [CrossRef]
- Brogi, S.; Ramalho, T.C.; Medina-Franco, J.L.; Kuca, K.; Valko, M. IN SILICO METHODS FOR DRUG DESIGN AND DISCOVERY. [CrossRef]
- White, H.S. Clinical Significance of Animal Seizure Models and Mechanism of Action Studies of Potential Antiepileptic Drugs. Epilepsia 1997, 38, S9–S17. [Google Scholar] [CrossRef]
Figure 1.
Structures of quercetin and selected reference drugs screened against the emesis inducing receptor.
Figure 1.
Structures of quercetin and selected reference drugs screened against the emesis inducing receptor.
Figure 2.
Latency (sec) of retches observed in test sample, controls and combinations. [Values are mean ± S.E.M. (n = 5). acompared to the NC (vehicle), bcompared to the DOM (PC); ccompared to the OND; dcompared to the HYS; ecompared to the QUA-25; fcompared to the QUA-50; gcompared to the DOM+QUA-50; hcompared to the OND+QUA-50; p<0.05 (OND+QUA-50 vs HYS+QUA-50); p<0.0001(NC vs DOM, NC vs QUA-25, NC vs QUA-50, NC vs DOM+QUA-50, NC vs OND+QUA-50, NC vs HYS+QUA-50, DOM vs OND, DOM vs HYS, DOM vs QUA-50, DOM vs DOM+QUA-50, DOM vs OND+QUA-50, DOM vs HYS+QUA-50, OND vs QUA-25, OND vs QUA-50, OND vs DOM+QUA-50, OND vs OND+QUA-50, OND vs HYS+QUA-50, HYS vs QUA-25, HYS vs QUA-50, HYS vs DOM+QUA-50, HYS vs OND+QUA-50, HYS vs HYS+QUA-50, QUA-25 vs QUA-50, QUA-25 vs DOM+QUA-50, QUA-25 vs OND+QUA-50, QUA-25 vs HYS+QUA-50, QUA-50 vs DOM+QUA-50, QUA-50 vs OND+QUA-50, QUA-50 vs HYS+QUA-50, DOM+QUA-50 vs HYS+QUA-50).
Figure 2.
Latency (sec) of retches observed in test sample, controls and combinations. [Values are mean ± S.E.M. (n = 5). acompared to the NC (vehicle), bcompared to the DOM (PC); ccompared to the OND; dcompared to the HYS; ecompared to the QUA-25; fcompared to the QUA-50; gcompared to the DOM+QUA-50; hcompared to the OND+QUA-50; p<0.05 (OND+QUA-50 vs HYS+QUA-50); p<0.0001(NC vs DOM, NC vs QUA-25, NC vs QUA-50, NC vs DOM+QUA-50, NC vs OND+QUA-50, NC vs HYS+QUA-50, DOM vs OND, DOM vs HYS, DOM vs QUA-50, DOM vs DOM+QUA-50, DOM vs OND+QUA-50, DOM vs HYS+QUA-50, OND vs QUA-25, OND vs QUA-50, OND vs DOM+QUA-50, OND vs OND+QUA-50, OND vs HYS+QUA-50, HYS vs QUA-25, HYS vs QUA-50, HYS vs DOM+QUA-50, HYS vs OND+QUA-50, HYS vs HYS+QUA-50, QUA-25 vs QUA-50, QUA-25 vs DOM+QUA-50, QUA-25 vs OND+QUA-50, QUA-25 vs HYS+QUA-50, QUA-50 vs DOM+QUA-50, QUA-50 vs OND+QUA-50, QUA-50 vs HYS+QUA-50, DOM+QUA-50 vs HYS+QUA-50).
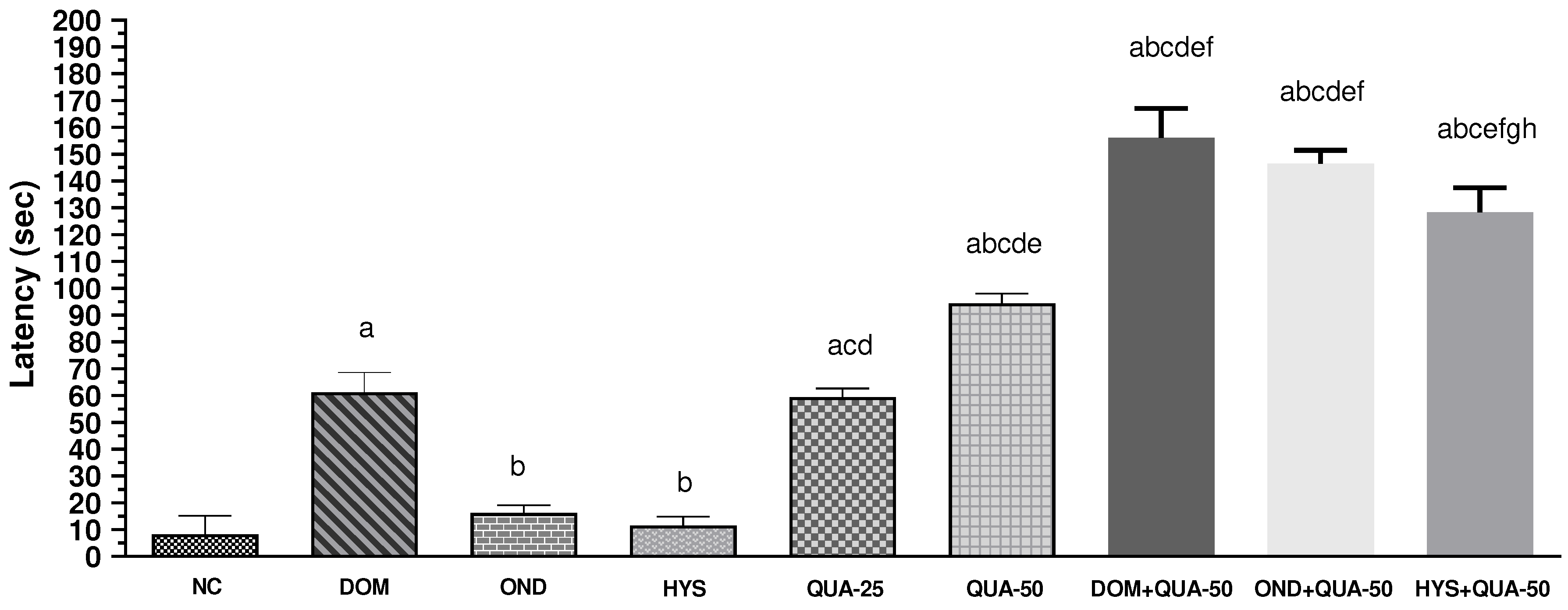
Figure 3.
Number of retches observed in test sample, controls and combinations. [Values are mean ± S.E.M. (n = 5). acompared to the NC (vehicle), bcompared to the DOM (positive control); ccompared to the OND; dcompared to the HYS; p<0.05 (NC vs HYS, OND vs QUA-25); p<0.01 (DOM vs OND, OND vs HYS, OND vs HYS+QUA-50); p<0.001 (OND vs QUA-50, OND vs OND+QUA-50); p<0.0001 (NC vs DOM, NC vs OND, NC vs QUA-25, NC vs QUA-50, NC vs DOM+QUA-50, NC vs OND+QUA-50, NC vs HYS+QUA-50, DOM vs HYS, OND vs DOM+QUA-50, HYS vs QUA-25, HYS vs QUA-50, HYS vs DOM+QUA-50, HYS vs OND+QUA-50, HYS vs HYS+QUA-50).
Figure 3.
Number of retches observed in test sample, controls and combinations. [Values are mean ± S.E.M. (n = 5). acompared to the NC (vehicle), bcompared to the DOM (positive control); ccompared to the OND; dcompared to the HYS; p<0.05 (NC vs HYS, OND vs QUA-25); p<0.01 (DOM vs OND, OND vs HYS, OND vs HYS+QUA-50); p<0.001 (OND vs QUA-50, OND vs OND+QUA-50); p<0.0001 (NC vs DOM, NC vs OND, NC vs QUA-25, NC vs QUA-50, NC vs DOM+QUA-50, NC vs OND+QUA-50, NC vs HYS+QUA-50, DOM vs HYS, OND vs DOM+QUA-50, HYS vs QUA-25, HYS vs QUA-50, HYS vs DOM+QUA-50, HYS vs OND+QUA-50, HYS vs HYS+QUA-50).
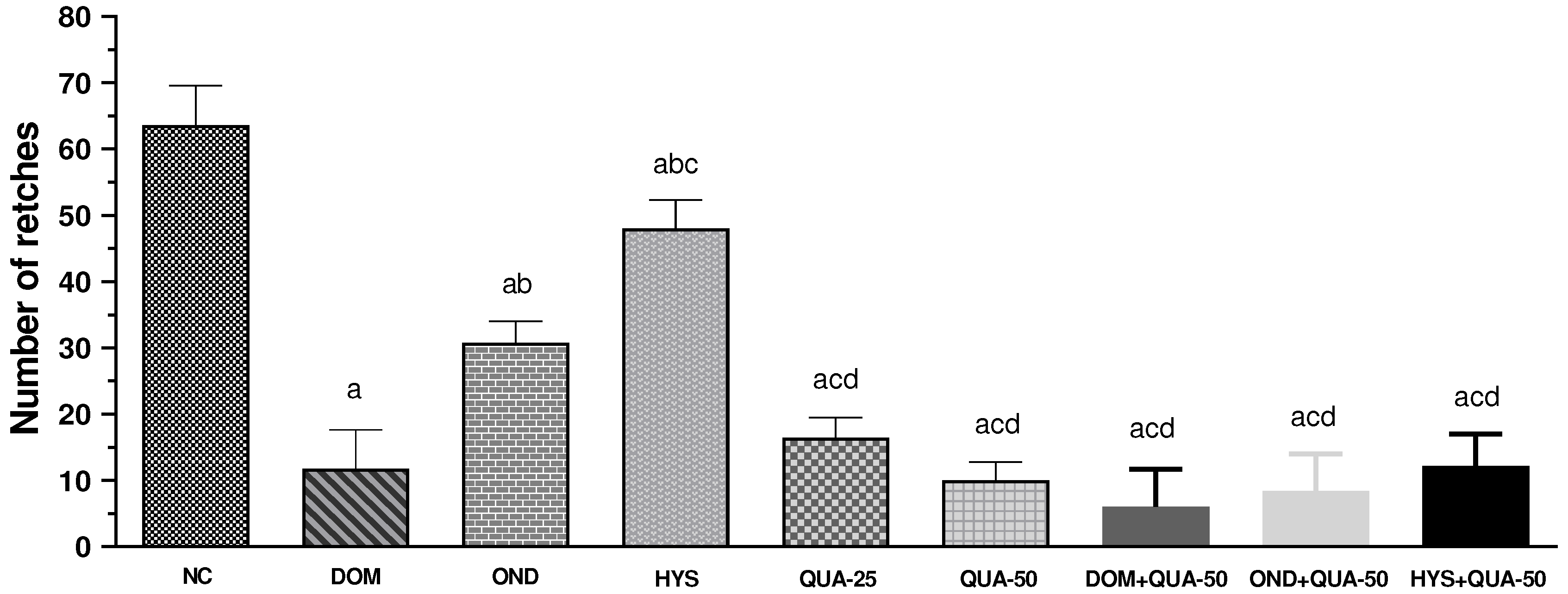
Figure 4.
The best Molecular docking interaction of the D2 receptor with (A) QUA or (B) DOM compound.
Figure 4.
The best Molecular docking interaction of the D2 receptor with (A) QUA or (B) DOM compound.
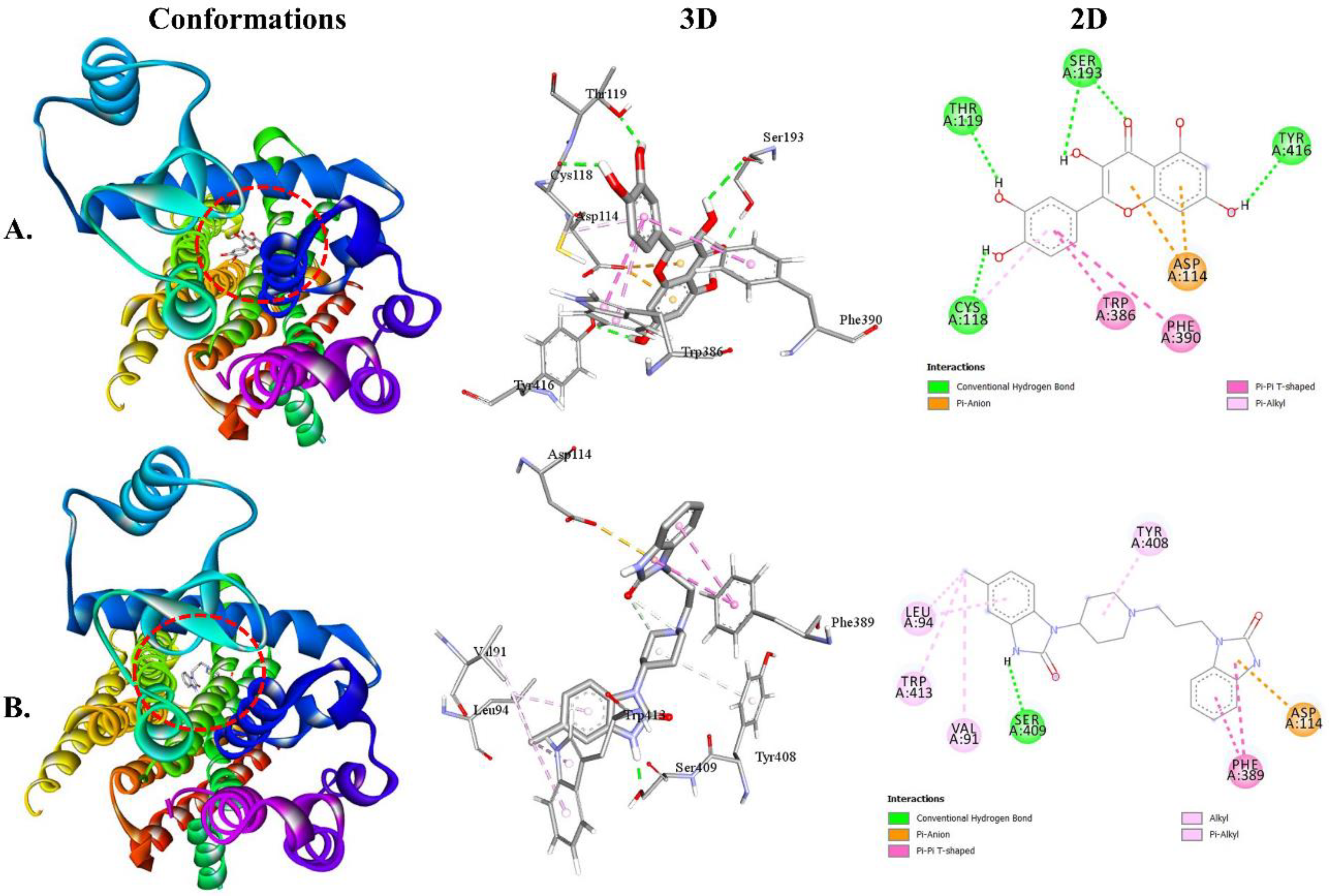
Figure 5.
The best Molecular docking interaction of the 5HT3 receptor with (A) QUA or (B) OND compound.
Figure 5.
The best Molecular docking interaction of the 5HT3 receptor with (A) QUA or (B) OND compound.
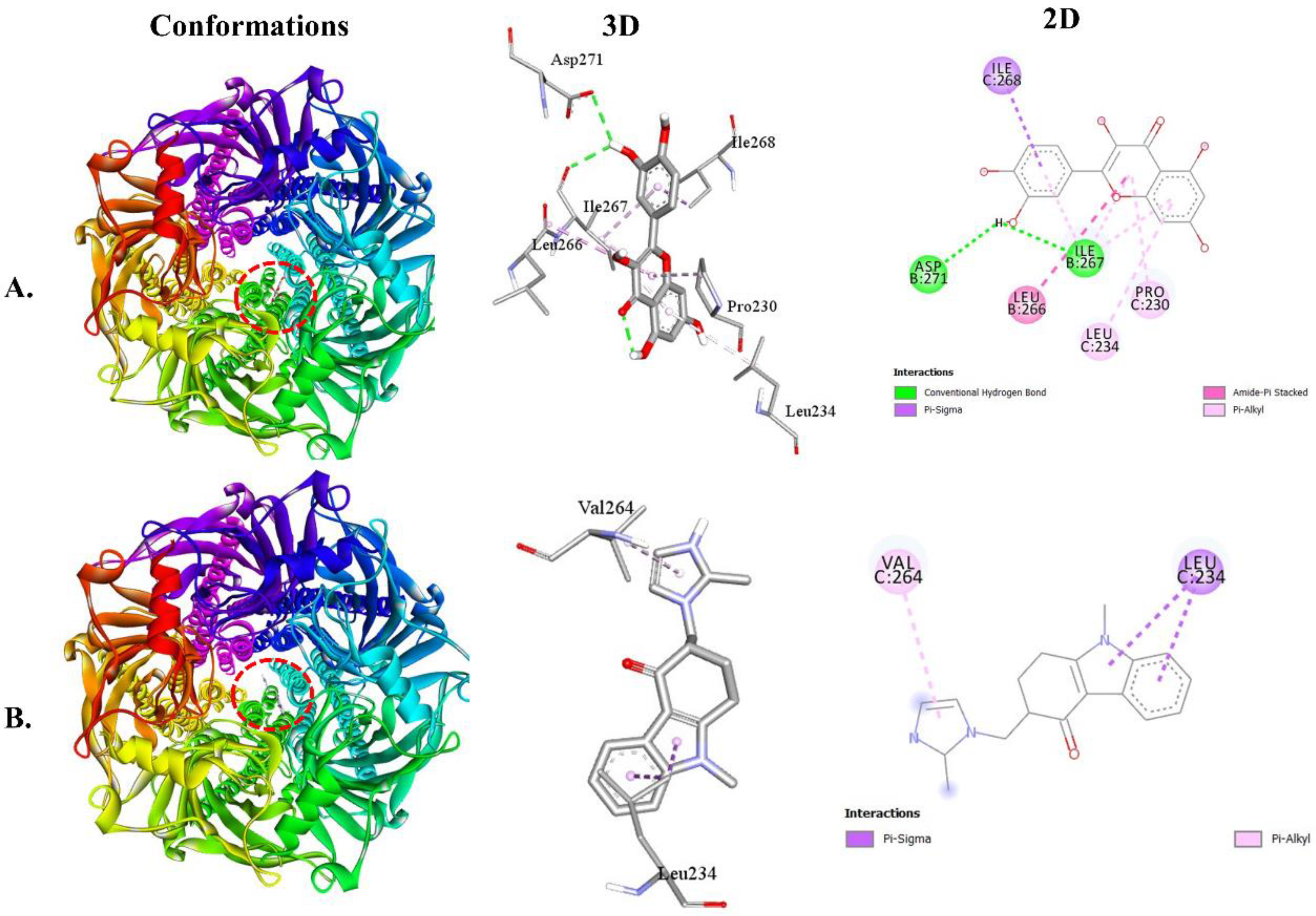
Figure 6.
The best molecular docking interaction of the M4 receptor with (A) QUA or (B) HYS compound.
Figure 6.
The best molecular docking interaction of the M4 receptor with (A) QUA or (B) HYS compound.
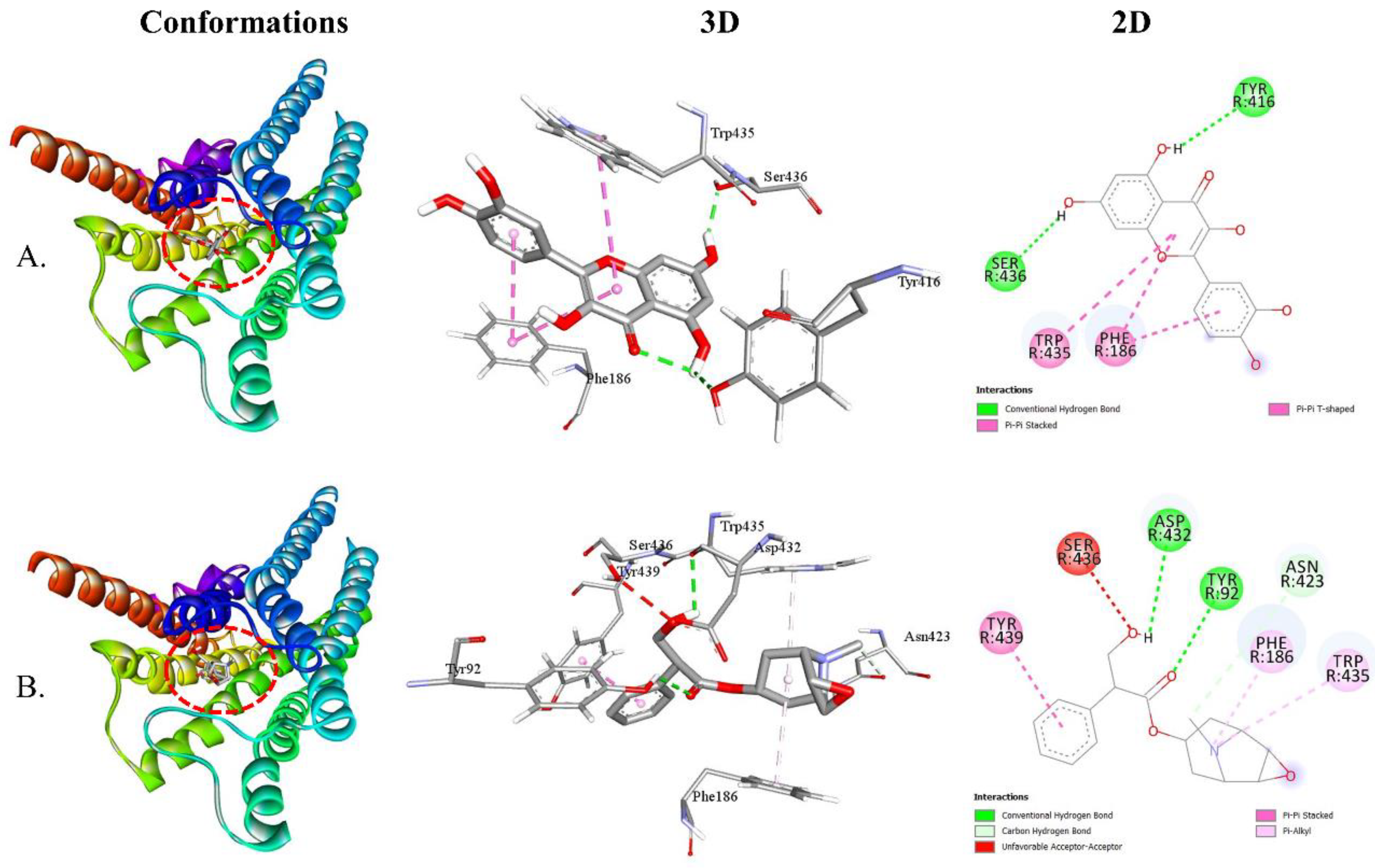
Figure 7.
The proposed mechanism of action for the antiemetic effects of QUA, OND, DOM, and HYS is based on their binding affinity with the D2, 5HT3, and M4 receptors [Briefly, QUA is evident to bind with 5HT3 (Lee et al., 2005). Our in silico studies suggest that it can also bind with D2 and M4 receptors. Upon going through the literature report and our present study, we suppose QUA may inhibit these three receptors. On the other hand, both DOM and OND display inhibitory effects on D2 and 5HT3 receptors, respectively, while HYS specifically blocks M4 receptor. By antagonizing these stomach receptors, these drugs prevent the stimulation of the vomiting center located in the medulla oblongata in animals. As a result, there is a lack of gastrointestinal tract contraction, muscle contraction, and initiation of efferent signals to organs, ultimately, inhibiting emesis].
Figure 7.
The proposed mechanism of action for the antiemetic effects of QUA, OND, DOM, and HYS is based on their binding affinity with the D2, 5HT3, and M4 receptors [Briefly, QUA is evident to bind with 5HT3 (Lee et al., 2005). Our in silico studies suggest that it can also bind with D2 and M4 receptors. Upon going through the literature report and our present study, we suppose QUA may inhibit these three receptors. On the other hand, both DOM and OND display inhibitory effects on D2 and 5HT3 receptors, respectively, while HYS specifically blocks M4 receptor. By antagonizing these stomach receptors, these drugs prevent the stimulation of the vomiting center located in the medulla oblongata in animals. As a result, there is a lack of gastrointestinal tract contraction, muscle contraction, and initiation of efferent signals to organs, ultimately, inhibiting emesis].
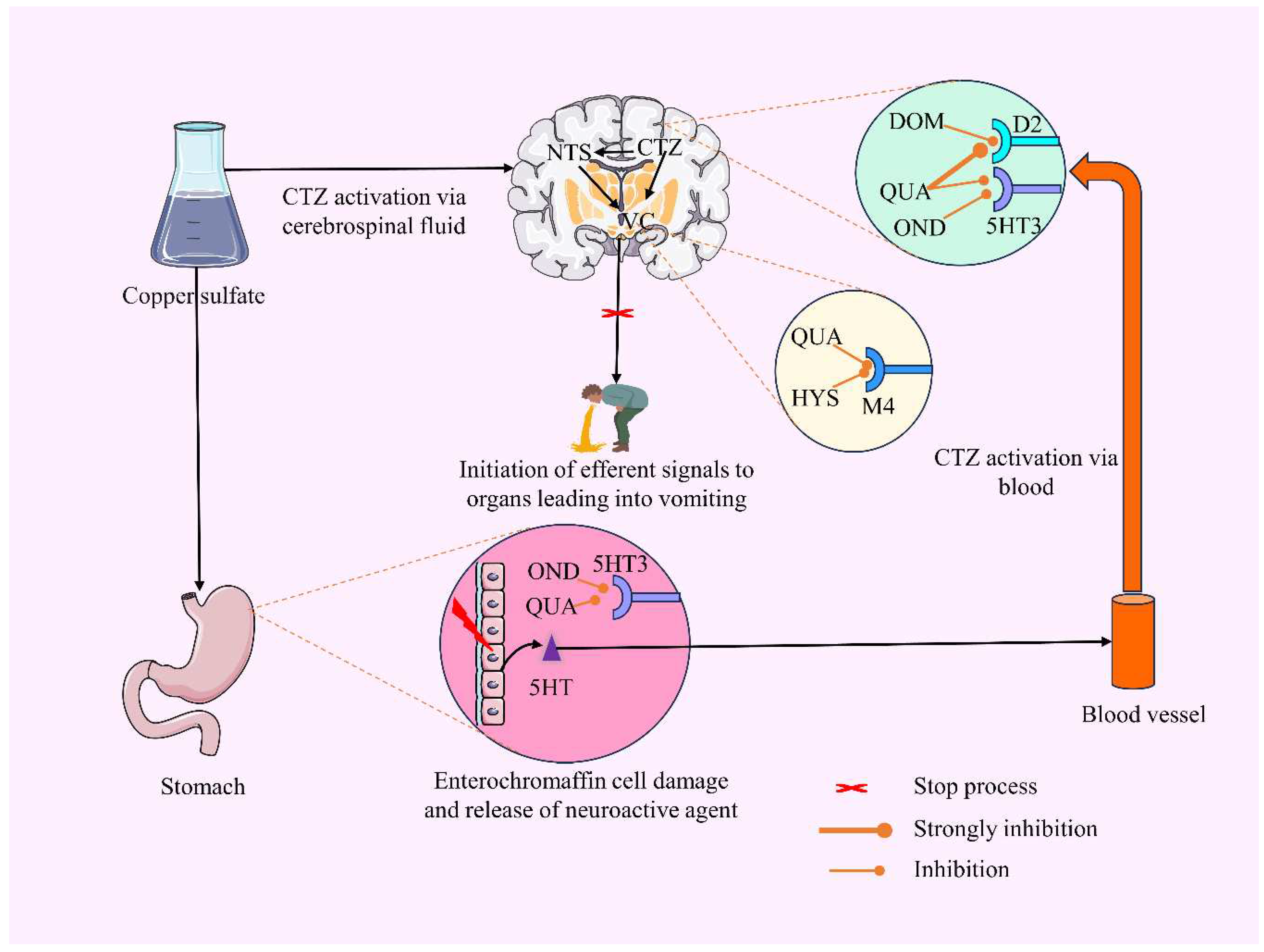
Table 1.
Percentage increase in latency and decrease in retches in treatment groups retches observed in test and referral groups.
Table 1.
Percentage increase in latency and decrease in retches in treatment groups retches observed in test and referral groups.
| Name of group | %Increase in latency | %Decrease in retches |
| NC (vehicle) | - | - |
| DOM | 86.48 | 81.63 |
| OND | 48.98 | 52.06 |
| HYS | 28.26 | 24.00 |
| QUA-50 | 91.25 | 84.43 |
| QUA-25 | 86.08 | 74.32 |
| DOM+QUA-50 | 94.73 | 90.66 |
| OND+QUA-50 | 94.36 | 87.04 |
| HYS+QUA-50 | 93.56 | 81.01 |
NC: Distilled water (Dose: 150 mg/kg); DOM: Domperidone (Dose: 6 mg/kg); OND: Ondansetron (Dose: 5 mg/kg); HYS: Hyoscine (Dose: 21 mg/kg); QUA-50: Quercetin (Dose: 50 mg/kg); QUA-25: Quercetin (Dose: 25 mg/kg); DOM+QUA-50: Domperidone+Quercetin (Dose: 6+50 mg/kg); ODN+QUA-50: Ondansetron+Quercetin (Dose: 5+50 mg/kg); HYS+QUA-50: Hyoscine+Quercetin (21 +50 mg/kg).
Table 2.
The best results of a molecular docking study of QUA and DOM with two (D2 and D3) receptors.
Table 2.
The best results of a molecular docking study of QUA and DOM with two (D2 and D3) receptors.
| Receptors | Binding Affinity | No. of HB | HB residues | HB length (Å) | Other bond residues |
| D2-QUA | -9.7 | 4 | SER193 TYR416 THR119 CYS118 |
1.85 2.20 2.14 2.18 |
ASP114 TRP386 PHE390 |
| D3-QUA | -8.5 | 3 | VAL189 TYR365 VAL111 |
2.36 2.09 3.56 |
ASP110 ILE183 PHE345 HIS349 VAL107 ILE183 |
| D2-DOM | -9.8 | 1 | SER409 | 1.89 |
ASP114 PHE389 VAL91 LEU94 TYR408 TRP413 |
| D3-DOM | -9.4 | 3 | GLY1107 VAL1103 PHE1104 |
2.83 1.77 3.53 |
GLU1011 ASP1020 LEU1032 ALA1074 |
Table 3.
The best results of a molecular docking study of QUA and OND with 5HT3 receptors.
| Receptors | Binding Affinity | No. of HB | HB residues | HB length (Å) | Other bond residues |
|---|---|---|---|---|---|
| 5HT3-QUA | -7.9 | 2 | ILE267 ASP271 |
2.51 2.95 |
ILE268 LEU266 ILE267 LEU234 PRO230 |
| 5HT3-OND | -8.1 |
- | - | - | LEU234 VAL264 |
Table 4.
The best results of a molecular docking study of QUA and HYS with five (M1, M2, M3, M4, and M5) receptors.
Table 4.
The best results of a molecular docking study of QUA and HYS with five (M1, M2, M3, M4, and M5) receptors.
| Receptors | Binding Affinity | No. of HB | HB residues | HB length (Å) | Other bond residues |
|---|---|---|---|---|---|
| M1-QUA | -7.8 |
2 | TYR106 ILE180 |
2.03 2.77 |
TYR404 |
| M2-QUA | -8.2 |
2 | PHE181 TYR104 |
2.43 1.96 |
TRP422 |
| M3-QUA | -9 |
2 | TRP206 ARG183 |
2.08 3.22 |
ARG171 MET187 VAL210 TYR175 |
| M4-QUA | -9.2 |
2 | TYR416 SER436 |
2.54 2.06 |
PHE186 TRP435 |
| M5-QUA | -8 |
2 | ILE185 HIS478 |
2.76 2.19 |
TYR481 TRP477 |
| M1-HYS | -7.1 | 4 | THR189 CYS178 TYR404 LEU183 |
2.01 2.33 3.62 3.41 |
TYR179 |
| M2-HYS | -7.5 | - | - | - | TYR426 |
| M3-HYS | -8.6 |
2 | ASN507 TYR529 |
2.26 3.47 |
CYS532 TYR148 TRP503 TYR506 ALA235 |
| M4-HYS | -8.9 |
3 | TYR92 ASP432 ASN423 |
2.81 2.71 3.70 |
TYR439 PHE186 TRP435 |
| M5-HYS | -7.7 |
ILE185 HIS478 SER189 |
1.93 2.62 2.95 |
TRP477 TYR481 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



