
Submitted:
16 November 2023
Posted:
17 November 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Long Covid has become a blight, its economic consequences overwhelming. But Long Covid (LC) may represent persistent Covid-19 in those with preexisting Candida overgrowth (CO). The features shared by both are striking. LC may be a growing subset of CO. The altered tryptophan metabolism encountered in LC is driven by IFN-γ in reaction to CO. Females are robust producers of IFN-γ, yet estrogen promotes CO.The consequent decline in serotonin and melatonin triggers the mood swings and sleeplessness respectively in LC. The low serotonin, the primary messenger in the gut brain axis, creates autonomic dysfunction. The CO induced gut dysbiosis and leaky gut initiate an autoimmune path. TNFα seen in both LC and this opportunistic yeast is at the center of the cytokine triad that leads to neurodegeneration. The Western diet, high in carbohydrates, alcohol, and glutamate, is exploited by this commensal turned parasite. Residual SARS CoV2 and Candida generate abundant ROS, inducing mitochondrial dysfunction. Alcohol tastes different for many with LC, because acetaldehyde requires functioning hepatic mitochondria. This article delves into the complex physiology and biochemistry that drive the symptoms of this conspicuous connection. Recent research has revealed innovative approaches that might address this global scourge, including D-mannose for the immune dysfunction and butyrate for the gut dysbiosis created by and seen in both LC and intestinal Candidiasis.
Keywords:
D-mannose
; butyrate
; acetaldehyde
; serotonin
; autoimmunity
Introduction
An unholy combination of virus and fungus promoting gut dysbiosis is not new and has been previously reported for HIV [1,2]. Linkage between Candida induced gut dysbiosis and post viral fatigue syndrome has been suspected for almost 30 years [3]. Linkage between CO and not only the immune system [4] but also the gut brain axis is becoming more apparent. Patients with a depleted T-cell count are more prone to Candidiasis. HIV consumes CD4+ T cells and SARS CoV2 consumes CD8+ T cells. This supports reports of CO induced dysbiosis in CFS [3], AIDS, and LC.
The pre-existing Candida associated gut dysbiosis may be undiagnosed in an otherwise healthy individual. Symptoms are nonspecific and fall into a range depending on their severity. Nutritional deterioration in the Western diet has been insidious. Carbohydrates dominate the menu. Glutamates enhance taste. Alcohol eases the stress. SARS CoV2 has teamed with the commensal turned opportunist Candida albicans to mount an increasing challenge to the health of our gut microbiota. The sequestration of the pandemic has amplified the need for alcohol for those dependent. Many of these “light drinkers” and others were already vitamin and micronutrient deficient. The increase in many “modern” diseases is readily apparent - AD, CA [5] and autoimmune diseases [6]. Although the search for a therapeutic solution to LC has been almost frenetic and solutions at times have appeared tantalizingly close, ultimate success has been elusive.
Discussion

Table 1.
Features common to chronic intestinal candidiasis and LC. CO potentiates SARS CoV2 and SARS CoV2 energizes CO. Together they create LC.
Table 1.
Features common to chronic intestinal candidiasis and LC. CO potentiates SARS CoV2 and SARS CoV2 energizes CO. Together they create LC.
| 1. Altered tryptophan metabolism (7,8) 2. Increased indoleamine 2,3-dioxygenase activity (7,9,10) 3. Low serum serotonin (11,12,13,14,15,16) 4. Low butyrate produced by gut microbiota (17,18,19) 5. TNFα mediated (20, 5) 6. Oxidative stress associated neurodegeneration (3,21) 7. Elevated dementia triad cytokines - TNFα, IL-1β, IL-6 (22,23,24,25) 8. Low (exhausted) IFN-γ (26,27] 9. Low (exhausted) mannose binding lectins (MBLs) (28,29,30,31) 10. Complement mediated fog (29,32,33) 11. High oligomannose glycan shield (34,35,36) 12. Alcohol intolerance (37,38) 13. Dysbiosis, Leaky gut (39,40) 14. Increased autoimmunity (39,40,41) 15. Brain fog and fatigue (29,42) 16. Accelerated AD (43,44,45) 17. More common in females during their reproductive years (46,47) |
The overwhelming majority of those with LC may be those with pre-existing CO and some degree of gut dysbiosis that subsequently contracted Covid-19. CO might be at play in both LC and CFS [3], all mediated by oxidative stress [21].
1 Microbiome, Serotonin, Altered Tryptophan Metabolism
A fungal microbiome has already been established for severe Covid [48]. This gut microbiome may explain the symptomatic overlap between CO and LC. Faecal samples collected in early 2021 from 10 people with COVID-19 showed elevated levels of gut fungi, especially of Candida species, relative to 10 healthy [48]. SARS CoV2 induced gut dysbiosis can promote CO [49] and CO induced gut dysbiosis can promote LC [50].
The gut microbiome in those with LC is low in Bifidobacterium spp and Clostridium spp that produce butyrate [51]. The gut microbiome in those with CO is also low in butyrate producing bacterial species [52].
The vast majority of the body’s serotonin comes from intestinal enterochromaffin cells. The altered tryptophan metabolism seen with CO results in a reduction of serotonin (see Figure 1). In LC this reduction of serotonin may explain its anosmia and ageusia [53]. These symptoms may be related to the downstream effect of the serotonin induced autonomic imbalance on the gut brain axis. Loss of taste and smell have been well described in LC, but it has also been described post vaccination [54]. One study of 2289 elderly participants reported that those with poor olfaction had a 46% higher cumulative risk for death after 10 years v those with good olfaction [55]. The gut enterochromaffin cells produce 95% of the body’s serotonin [16], but rely on the essential amino acid tryptophan from either diet or intestinal bacteria to produce serotonin. Gut dysbiosis compromises the microbiota balance causing a decrease in critical micronutrients, e.g., butyrate, tryptophan, and dopamine [16]. CO may displace many of these good bacteria with consequent loss of vital micronutrients. The LC microbiome reflects this same loss of micronutrients, especially tryptophan [10]. The effect of decreased serotonin and dopamine on downstream monoaminergic and GABAergic neurotransmission is complex but clearly creates an autonomic imbalance.
T cells produce IFN-γ in response to CO [56], which shifts tryptophan metabolism from the serotonin pathway to the kynurenine pathway and NAD+ production (see Figure 1). This altered tryptophan metabolism is common to both CO and LC. Low serotonin [11,12,13] and melatonin ensue along with mood swings and insomnia respectively. Reports on IFN-γ levels in LC differ. Some report elevated IFN-γ [57], while others report marked decreases in IFN-γ [26] However, the latter is generally considered to be due to immune exhaustion [16]. Upregulation of indoleamine dioxygenase (IDO-1 and IDO-2), courtesy of IFN-γ, is directly related to Covid severity and CO [7,9] and may underlie potential for LC [10]. Females are robust producers of IFN-γ. IFN-γ in response to CO drives the pivot from the serotonin pathway to the kynurenine pathway [56]. This shift also occurs in LC [10]. Females are robust producers of IFN-γ and this prompts upregulation of the kynurenine pathway (see Figure 1).
Quinolinic acid and 3(OH)kynurenine are neurotoxic and upregulated when B6 is deficient [58]. The two NAD+ produced by the kynurenine pathway help address the shortfall in ATP due to the oxidative stress induced mitochondrial dysfunction. Candida requires much more thiamine in such an environment [59].
2 Alcohol and Mitochondria
Alcohol intolerance is a primary complaint in LC (alcohol tastes different) and CFS. Hepatic metabolism of alcohol by alcohol dehydrogenase yields acetaldehyde, but further degradation by aldehyde dehydrogenase occurs in mitochondria. To control CO the body oxidizes the yeast through granulocytes [60]. Even more ROS are released. When oxidatively stressed mitochondrial function is further compromised, acetaldehyde can cause headache, hangover, brain fog, fatigue [61].
These symptoms are seen in CO, which produces acetaldehyde from ethanol and/or glucose. The yeast induced acetaldehyde depletes thiamine [62], a deficiency linked to chronic alcoholism and Wernicke Korsakoff syndrome. This is exacerbated by glucose or ethanol loading [42] and leads to acetaldehyde induced brain fog. SARS CoV2 induced increases in acetaldehyde post Covid-19 may suggest the presence of early LC. Could a glass of wine diagnose LC? [37].
Although some reports suggest benefit from light alcohol intake against vascular dementia, there is no such efficacy against AD [63]. The brain fog of alcohol induced acetaldehyde (hangover) is equivalent to that induced by chronic intestinal candidiasis after glucose loading [48].
Thiamine deficiency, often encountered with excessive ethanol intake, is a required cofactor for pyruvate dehydrogenase. Without this enzyme pyruvate is shunted from producing acetyl CoA (requires vitamin B5 aka pantothenate) to lactate instead. Several other dehydrogenases in the Krebs Cycle also require thiamine pyrophosphate (TPP). These two thiamine-dependent pathways may mediate mitochondrial dysfunction and its associated fatigue. Thiamine deficiency rates range from 20% to over 90% [64].
CO is marked by increased production of ROS by the fungus [65] and by the host. SARS CoV2 can only aggravate this increase in ROS with concomitant loss of mitochondria and decrease in oxidative phosphorylation [66]. Susceptible individuals may already have an antioxidant shortfall
Persistent spike protein [67], demonstrated in those with LC, creates a nearly insurmountable challenge. Secondary neurodegenerative changes in an organ highly dependent on oxygen may accelerate [68], when combined with minimal alcohol abuse and thiamine deficiency [38]. Candida albicans is able to generate significant amounts of reactive oxygen species (ROS) [65]. In order to avoid lysis this forces cells to curtail their own ROS production from mitochondrial oxidative phosphorylation. The kynurenine pathway produces NAD+ via a non oxidative pathway, requiring B2 and B6, to create ATP (see Figure 2 and Figure 3). B1 is required for both oxidative phosphorylation and the pentose phosphate shunt, which bypasses the mitochondria to produce NADPH and ultimately ATP. This latter is especially important for erythrocytes, which lack mitochondria.
3 MBLs, Brain Fog, and Amyloid
MBLs are an integral part of innate immunity, the first line of defense, while antibodies comprise the bulk of adaptive immune defense. Candida [35], SARS CoV2, and many types of cancer cells are protected by high mannose glycan shields to avoid exposure to immunoglobulins. SARS CoV2, its vaccine, and 31 different types of cancer cells produce a CD147 epitope [69] that is adorned by a high oligomannose glycan shield. This oligomannose glycan shield on Candida albicans and SARS CoV2 attracts MBLs, which are vital to fungal elimination [31]. These oligomannosides not only serve to protect them but also to upregulate TNFα [20,34]. Candidiasis is linked to low MBL [28,29,30,31].
Recurrent Covid-19 is also linked to low MBL [32], generally felt due to immune exhaustion. The protective benefit of the oligomannose glycan shield in avoiding immunoglobulin detection is evident when MBLs are low [70]. Low MBLs may mark the onset of brain fog in LC [29]. Brain fog early in CO may be glucose or ethanol induced and acetaldehyde mediated, but the more serious cognitive decline that develops later may be MBL/lectin complement pathway [33] and cytokine mediated.
Covid-19 and its vaccines present the CD147 epitope and its oligomannose glycan shield. The MBL assault triggers the complement and clotting cascades. The ensuing MBL induced microthrombosis, closely linked to d-dimers, may be the prelude to neurodegenerative disease. The presence of amyloid beta like peptides in microglia associated with candidiasis (27,49), already demonstrated in AD, and the linkage of Covid-19 [44,45] and CO with neurodegenerative symptoms paints a worrisome long term picture for some long haulers. Brain amyloid β (Aβ)-like peptide aggregates characterize multiple Candida-associated neurodegenerative conditions, including Alzheimer's disease [43]. Amyloid precursor protein surrounds cleaved amyloid beta (Aβ) peptides around the CNS yeast cells. This activates NF-κB and induces production of IL-1β, IL-6, and TNFα [22]. This triad can activate NF-kB and NF-kB can activate the triad [23]. This triad has been detected in Alzheimer's disease, Parkinson's disease, amyotrophic lateral sclerosis, multiple sclerosis, and acquired immunodeficiency dementia [24].
6 Leaky Gut and Autoimmunity
The leaky gut syndrome induced by both Covid 19 [71] and CO [39] induces an environment that promotes autoimmune disease [40,41], e.g.,T1DM [72]. Post vax T1DM has also been reported [73,74]. CO can also induce T2DM [75,76], as can LC, but both types increase risk of CO. Both also increase risk for LC. There are numerous reports of T1DM appearing immediately post vax. Other reports claim that the vaccine protects against developing T2DM post Covid-19, but are silent regarding T1DM. Autoimmune T1DM post vax is well described(73,74,76). CO is linked to leaky gut and leaky gut is linked to autoimmunity. LC has now been classified as an autoimmune disease [6]. The balance between leaky gut and autoimmunity is precarious [77].
Prevention and Therapy
Many experienced clinicians advocate significant dietary changes to eliminate CO, i.e., the Candida cleanse detox. But changing one’s diet can be more difficult than changing one’s religion. Fortunately there may be other recently discovered approaches that cover LC as well, including D-mannose, butyrate, and tryptophan/serotonin/melatonin. Vitamin D, magnesium, and the active forms of the B vitamins, especially B1, B2, B3, B6 have proven efficacious. Prebiotics and probiotics in mild cases to fecal transplants in more severe cases have also been recommended.
1 Mannose
D-mannose, usually prescribed for UTI, suppresses TNFα [78], central to the dementia induced by the triad of TNFα, IL-6, IL-1β [24]. TNFα (inhibits mannosidase) is also central to the synthesis of the high oligomannose glycan shields on both Candida albicans and SARS CoV2, not to mention 31 different types of cancer cells [69]. TNFα upregulates IL-1β, linked to autoimmune disease. Mannose suppresses IL-1β [79]. Mannose may be especially helpful in preventing leaky gut syndrome [80,81], tightly linked to autoimmune disease. In fact, beneficial applications of mannose are rapidly expanding. D-Mannose offers newly recognized efficacy for diabetes, obesity, lung disease, autoimmune diseases and recently anti-tumor activity [82]. Once considered ill advised for diabetics, it has now been heralded as beneficial for diabetics.
2 Butyrate
Butyrate opposes growth of pathogenic yeast [18] and appears to increase serotonin, especially in the hippocampus [83]. Serotonin actively attenuates fungal virulence [84]. AD related amyloid beta bodies are well described in the hippocampus and amyloid beta bodies are linked to Candida [22,43]. SB exerts significant antifungal activity on pathogenic yeasts [17]. Short chain fatty acids, especially butyrate, suppress IDO and increase serotonin (see Figure 1) [85,86].
3 Serotonin and tryptophan
The altered tryptophan metabolism creates a significant serotonin shortfall. Serotonin supplementation can help fill the void. It attenuates fungal virulence [14,15] and may restore a proper immune response [87]. Less of the essential amino acid tryptophan is supplied by gut microbiota during LC and CO. Many clinicians encourage supplementing with tryptophan [88].
4 Vitamin D and magnesium
Men with the highest compared to lowest 1,25(OH)2D and activation ratios, i.e., active form/storage form (1,25(OH)2D/25(OH)D), are more likely to possess butyrate-producing bacteria associated with favorable gut microbial health [89]. Vitamin D, low in both LC and CO, suppresses TNFα [90,91]. Vitamin D levels are inversely correlated with TNFα. Vitamin D is also a fungicide effective against Candida albicans [92,93]. Vitamin D modulates the immune actions of IFN-γ [94].
Magnesium, a required cofactor for many enzymatic steps in the synthesis of vitamin D, also suppresses the neurodegeneration-inducing triad of TNFα, IL-1β, and IL-6 [95]. This cytokine triad drives LC and candidiasis [24,25,26,27] and neurodegenerative disease in general [25].
5 Covid boosters
Covid boosters are a personal choice, but the latest data are worrisome. Boost at your own risk. Each boost increases the likelihood of a recurrence and the associated risk of LC [96]. The CD147 epitope is present on the spike proteins S of the virus and its vaccines [97,98]. Post vax autoimmune complications include T1DM, CFS, multiple sclerosis, alopecia, Bell’s palsy, Guillain-Barré syndrome, immune thrombocytopenia, rheumatoid arthritis [99]. Post vax neurological complications were seen in 31.2% of 19,108 post Covid vaccinees [100]. Post vax microthrombosis is the most well known complication, especially amongst our athletes. MBL activity is strongly correlated with d-dimers [102]. D-dimers were reported in 62% of 900 Covid vaccinees [102].
6 Vitamins
Gut microbiota is responsible for the production of several water-soluble vitamins, including biotin, folate, niacin, ascorbate, riboflavin and thiamine. Gut dysbiosis results in a reduced amount of such vitamins [103]. Some vitamins modulate the gut microbiome by increasing the abundance of presumed commensals (vitamins A, B2, D, E, and beta-carotene), increasing or maintaining microbial diversity (vitamins A, B2, B3, C, K) and richness (vitamin D), increasing short chain fatty acid production (vitamin C), or increasing the abundance of short chain fatty acid producers (vitamins B2, E). Others, such as vitamins A and D, modulate the gut immune response or barrier function [104]. Thiamine (B1) deficiency can cause [105] or be caused by gut dysbiosis [69,106].
Conclusion
Any strategy that limits the production of TNFα has multiple benefits. This cytokine inhibits mannosidase and limits mannose trimming in the Golgi. Suppressing TNFα suppresses the “triad”. This triad is central not only to neurodegenerative changes but also to processing the high oligomannose glycan shield present on invading pathogens and many types of cancer cells. Improving the suboptimal gut microbiome should address CO, LC, and the leaky gut associated with autoimmune disease. This opens a new approach to treatment of autoimmune disease, one that bypasses specifically designed inhibitors e.g., Janus kinase inhibitors for vitiligo [108]. Vitiligo is characterized by a microbiome that lacks butyrate producing bacteria and that is less rich and less diverse than that of healthy controls [108]. The myriad benefits of a broad approach, one that may improve the gut microbiome and limit the “triad” without a major dietary makeover, are alluring.
References
- Silvere D Zaongo, Jing Ouyang, Stéphane Isnard, Xin Zhou, Vijay Harypursat, Hongjuan Cui, et al. (2023) Candida albicans can foster gut dysbiosis and systemic inflammation during HIV infection, Gut Microbes, 15:1 . [CrossRef]
- Park SY, Faraci G, Nanda S, Ter-Saakyan S, Love TMT, et al. Gut microbiome in people living with HIV is associated with impaired thiamine and folate syntheses. Microb Pathog. 2021 Nov;160:105209. [CrossRef]
- Cater RE 2nd. Chronic intestinal candidiasis as a possible etiological factor in the chronic fatigue syndrome. Med Hypotheses. 1995 Jun;44(6):507-15. [CrossRef]
- Valand N, Girija UV. Candida Pathogenicity and Interplay with the Immune System. Adv Exp Med Biol. 2021;1313:241-272. [CrossRef]
- Chambers P (2023) The CD147 Epitope on SARS CoV2 and the Spike in Cancer, Autoimmunity and Organ Fibrosis. Medical & Clinical Research, 8(11), 01-13 https://www.medclinrese.org/open-access/the-cd147-epitope-on-sars-cov2-and-the-spike-in-cancer-autoimmunity-and-organ-fibrosis.pdf].
- Autoimmune Registry COVID-19 Added to the list of Autoimmune Diseases https://www.autoimmuneregistry.org/long-covid-announcement.
- Silvia Bozza, Francesca Fallarino, Lucia Pitzurra, Teresa Zelante, Claudia Montagnoli, Silvia Bellocchio, et al. A Crucial Role for Tryptophan Catabolism at the Host/Candida albicans Interface. J Immunol 1 March 2005; 174 (5): 2910–2918. [CrossRef]
- Kaur G, Ji X, Rahman I. SARS-CoV2 Infection Alters Tryptophan Catabolism and Phospholipid Metabolism Metabolites. 2021 Sep 28;11(10):659. [CrossRef]
- Guo L, Schurink B, Roos E, Nossent EJ, Duitman JW, Vlaar AP, et al. Indoleamine 2,3-dioxygenase (IDO)-1 and IDO-2 activity and severe course of COVID-19. J Pathol. 2022 Mar;256(3):256-261. [CrossRef]
- Eroğlu İ, Eroğlu BÇ, Güven GS. Altered tryptophan absorption and metabolism could underlie long-term symptoms in survivors of coronavirus disease 2019 (COVID-19). Nutrition. 2021 Oct;90:111308. [CrossRef]
- Harris E. Long COVID Linked With Viral Persistence, Serotonin Decline. JAMA. Published online November 01, 2023. [CrossRef]
- Wong AC, Devason AS, Umana IC, Cox TO, Dohnalová L, Litichevskiy L, et al. Serotonin reduction in post-acute sequelae of viral infection. Cell. 2023 Oct 26;186(22):4851-4867.e20. [CrossRef]
- Bird, L. Low serotonin linked to long COVID. Nat Rev Immunol. Nat Rev Immunol (2023) . [CrossRef]
- Mayr A, Hinterberger G, Dierich MP, Lass-Flörl C. Interaction of serotonin with Candida albicans selectively attenuates fungal virulence in vitro. Int J Antimicrob Agents. 2005 Oct;26(4):335-7. [CrossRef]
- Lass-Flörl C, Fuchs D, Ledochowski M, Speth C, Dierich MP, Würzner R. Antifungal properties of 5-hydroxytryptamine (serotonin) against Candida species in vitro. J Med Microbiol. 2003 Feb;52(Pt 2):169-171. [CrossRef]
- Banskota S, Khan WI. Gut-derived serotonin and its emerging roles in immune function, inflammation, metabolism and the gut-brain axis. Curr Opin Endocrinol Diabetes Obes. 2022 Apr 1;29(2):177-182. [CrossRef]
- Noureldein MH, Bitar S, Youssef N, Azar S, Eid AA. Butyrate modulates diabetes-linked gut dysbiosis: epigenetic and mechanistic modifications. J Mol Endocrinol. 2020 Jan;64(1):29-42. [CrossRef]
- Nguyen LN, Lopes LC, Cordero RJ, Nosanchuk JD. Sodium butyrate inhibits pathogenic yeast growth and enhances the functions of macrophages. J Antimicrob Chemother. 2011 Nov;66(11):2573-80. [CrossRef]
- Wang, B., Zhang, L., Wang, Y. et al. Alterations in microbiota of patients with COVID-19: potential mechanisms and therapeutic interventions. Sig Transduct Target Ther 7, 143 (2022). [CrossRef]
- Jouault T, Lepage G, Bernigaud A, Trinel PA, Fradin C, Wieruszeski JM, Strecker G, Poulain D. Beta-1,2-linked oligomannosides from Candida albicans act as signals for tumor necrosis factor alpha production. Infect Immun. 1995 Jun;63(6):2378-81. [CrossRef]
- Al-Hakeim, H.K., Al-Rubaye, H.T., Al-Hadrawi, D.S. et al. Long-COVID post-viral chronic fatigue and affective symptoms are associated with oxidative damage, lowered antioxidant defenses and inflammation: a proof of concept and mechanism study. Mol Psychiatry 28, 564–578 (2023). [CrossRef]
- Wu, Y., Du, S., Johnson, J.L. et al. Microglia and amyloid precursor protein coordinate control of transient Candida cerebritis with memory deficits. Nat Commun 10, 58 (2019). [CrossRef]
- Zhou, Q., Zhang, L., Dong, Y. et al. The role of SARS-CoV-2-mediated NF-κB activation in COVID-19 patients. Hypertens Res (2023). [CrossRef]
- Ishijima T, Nakajima K. Inflammatory cytokines TNFα, IL-1β, and IL-6 are induced in endotoxin- stimulated microglia through different signaling cascades. Sci Prog. 2021 Oct;104(4):368504211054985. [CrossRef]
- Schultheiß, Christoph, Edith Willscher, Lisa Paschold, Cornelia Gottschick, et al. 2022. The IL-1β, IL-6, and TNF Cytokine Triad Is Associated with Post-Acute Sequelae of COVID-19 Cell Reports Medicine 3 (6): 100663. [CrossRef]
- Williams ES, Martins TB, Shah KS, Hill HR, Coiras M, Spivak AM, Planelles V. Cytokine Deficiencies in Patients with Long-COVID. J Clin Cell Immunol. 2022;13(6):672. https://pubmed.ncbi.nlm.nih.gov/36742994/].
- Szkaradkiewicz, E. Szponar, E. Krzemińska-Jaśkowiak, T. Tułecka, Serum interferon-gamma (IFN-γ) in chronic oral candidosis, Medical Mycology, Volume 36, Issue 5, January 1998, Pages 269–273, . [CrossRef]
- Hammad NM, El Badawy NE, Ghramh HA, Al Kady LM. Mannose-Binding Lectin: A Potential Therapeutic Candidate against Candida Infection. Biomed Res Int. 2018 May 2;2018:2813737. [CrossRef]
- Bulla R, Rossi L, Furlanis G, Agostinis C, Toffoli M, Balduit A, et al. A likely association between low mannan-binding lectin level and brain fog onset in long COVID patients. Front Immunol. 2023 Jun 16;14:1191083. [CrossRef]
- Damiens S, Poissy J, François N, Salleron J, Jawhara S, Jouault T, et al. Mannose-binding lectin levels and variation during invasive candidiasis. J Clin Immunol. 2012 Dec;32(6):1317-23. [CrossRef]
- Choteau, L., Parny, M., François, N. et al. Role of mannose-binding lectin in intestinal homeostasis and fungal elimination. Mucosal Immunol 9, 767–776 (2016). [CrossRef]
- Hurler L, Szilágyi Á, Mescia F, Bergamaschi L, Mező B, Sinkovits G, et al. Complement lectin pathway activation is associated with COVID-19 disease severity, independent of MBL2 genotype subgroups. Front Immunol. 2023 Mar 27;14:1162171. [CrossRef]
- Harpf V, Rambach G, Würzner R, Lass-Flörl C, Speth C. Candida and Complement: New Aspects in an Old Battle. Front Immunol. 2020 Jul 14;11:1471. [CrossRef]
- Lillegard JB, Sim RB, Thorkildson P, Gates MA, Kozel TR. Recognition of Candida albicans by mannan-binding lectin in vitro and in vivo. J Infect Dis. 2006 Jun 1;193(11):1589-97. [CrossRef]
- Sendid B, Lecointe K, Collot M, Danzé PM, Damiens S, Drucbert AS, et al. Dissection of the anti-Candida albicans mannan immune response using synthetic oligomannosides reveals unique properties of β-1,2 mannotriose protective epitopes. Sci Rep. 2021 May 24;11(1):10825. [CrossRef]
- Bai Y, Huang W, Ma LT, Jiang JL, Chen ZN. Importance of N-glycosylation on CD147 for its biological functions. Int J Mol Sci. 2014 Apr 15;15(4):6356-77. [CrossRef]
- Pho, K. Could a glass of wine diagnose long COVID? https://www.kevinmd.com/2021/03/could-a-glass-of-wine-diagnose-long-covid.html.
- Chen CH, Joshi AU, Mochly-Rosen D. The Role of Mitochondrial Aldehyde Dehydrogenase 2 (ALDH2) in Neuropathology and Neurodegeneration. Acta Neurol Taiwan. 2016 Dec 15;25(4)(4):111-123. http://www.ncbi.nlm.nih.gov/pmc/articles/pmc10618051/.
- Kinashi Y, Hase K. Partners in Leaky Gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Front Immunol. 2021 Apr 22;12:673708. [CrossRef]
- Fasano A. Leaky gut and autoimmune diseases. Clin Rev Allergy Immunol. 2012 Feb;42(1):71-8. [CrossRef]
- Mousa WK, Chehadeh F, Husband S. Microbial dysbiosis in the gut drives systemic autoimmune diseases. Front Immunol. 2022 Oct 20;13:906258. [CrossRef]
- Zimitat C, Nixon PF. Glucose loading precipitates acute encephalopathy in thiamine-deficient rats. Metab Brain Dis. 1999 Mar;14(1):1-20. [CrossRef]
- Wu Y, Du S, Bimler LH, Mauk KE, Lortal L, Kichik N, et al. Toll-like receptor 4 and CD11b expressed on microglia coordinate eradication of Candida albicans cerebral mycosis. Cell Rep. 2023 Oct 31;42(10):113240. [CrossRef]
- Dubey, S., Das, S., Ghosh, R., Dubey, M. J., Chakraborty, A. P., Roy, D., et al. (2023). The effects of SARS-CoV-2 infection on the cognitive functioning of patients with pre-existing dementia. Journal of Alzheimer’s Disease Reports, 7(1), 119–128. [CrossRef]
- Xia X, Wang Y, Zheng J. COVID-19 and Alzheimer's disease: how one crisis worsens the other. Transl Neurodegener. 2021 Apr 30;10(1):15. [CrossRef]
- Kumwenda P, Cottier F, Hendry AC, Kneafsey D, Keevan B, Gallagher H, et al. Estrogen promotes innate immune evasion of Candida albicans through inactivation of the alternative complement system. Cell Rep. 2022 Jan 4;38(1):110183. [CrossRef]
- Loster JE, Wieczorek A, Loster BW. Correlation between age and gender in Candida species infections of complete denture wearers: a retrospective analysis. Clin Interv Aging. 2016;11:1707-1714 . [CrossRef]
- Krämer K. Daily briefing: Severe COVID linked to bad fungal microbiome. Nature. 2023 Oct 24. [CrossRef]
- Jawhara S. How Gut Bacterial Dysbiosis Can Promote Candida albicans Overgrowth during Colonic Inflammation. Microorganisms. 2022 May 12;10(5):1014. [CrossRef]
- Lagree K, Chen P. Candida makes a lasting impression in COVID-19. Nat Immunol. 2023 Nov;24(11):1782-1784. [CrossRef]
- Zhu Z, Zhu L, Jiang L. Dynamic regulation of gut Clostridium-derived short-chain fatty acids. Trends Biotechnol. 2022 Mar;40(3):266-270. [CrossRef]
- Hu W, Xu D, Zhou Z, Zhu J, Wang D, Tang J. Alterations in the gut microbiota and metabolic profiles coincide with intestinal damage in mice with a bloodborne Candida albicans infection. Microb Pathog. 2021 May;154:104826. [CrossRef]
- Sen A. Does serotonin deficiency lead to anosmia, ageusia, dysfunctional chemesthesis and increased severity of illness in COVID-19? Med Hypotheses. 2021 Aug;153:110627. [CrossRef]
- Lechien JR, Diallo AO, Dachy B, Le Bon SD, Maniaci A, Vaira LA, Saussez S. COVID-19: Post-vaccine Smell and Taste Disorders: Report of 6 Cases. Ear Nose Throat J. 2021 Sep 1:1455613211033125. [CrossRef]
- Bojing Liu, PhD*, Zhehui Luo, PhD*, Jayant M. Pinto, MD, Eric J. Shiroma, ScD, Gregory J. Tranah, PhD, Karin Wirdefeldt, MD, PhD, Relationship Between Poor Olfaction and Mortality Among Community-Dwelling Older Adults Ann Int Med 170(10):673-681 . [CrossRef]
- Gozalbo D, Gil ML. IFN-gamma in Candida albicans infections. Front Biosci (Landmark Ed). 2009 Jan 1;14(5):1970-8. [CrossRef]
- Li, J., Zhou, Y., Ma, J. et al. The long-term health outcomes, pathophysiological mechanisms and multidisciplinary management of long COVID. Sig Transduct Target Ther 8, 416 (2023). [CrossRef]
- Müller N, Myint AM, Schwarz MJ. The impact of neuroimmune dysregulation on neuroprotection and neurotoxicity in psychiatric disorders--relation to drug treatment. Dialogues Clin Neurosci. 2009;11(3):319-32. [CrossRef]
- Wolak N, Tomasi M, Kozik A, Rapala-Kozik M. Characterization of thiamine uptake and utilization in Candida spp. subjected to oxidative stress. Acta Biochim Pol. 2015;62(3):445-55. [CrossRef]
- Stevenhagen A, van Furth R. Interferon-gamma activates the oxidative killing of Candida albicans by human granulocytes. Clin Exp Immunol. 1993 Jan;91(1):170-5. [CrossRef]
- Reddy MGS, Kakodkar P, Nayanar G. Capacity of Candida species to produce acetaldehyde at various concentrations of alcohol. J Oral Maxillofac Pathol. 2022 Apr-Jun;26(2):161-165. [CrossRef]
- Takabe M, Itokawa Y. Thiamine depletion after ethanol and acetaldehyde administration to rabbits. J Nutr Sci Vitaminol (Tokyo). 1983 Oct;29(5):509-14. [CrossRef]
- Wiegmann C, Mick I, Brandl EJ, Heinz A, Gutwinski S. Alcohol and Dementia - What is the Link? A Systematic Review. Neuropsychiatr Dis Treat. 2020 Jan 9;16:87-99. [CrossRef]
- Marrs C, Lonsdale D. Hiding in Plain Sight: Modern Thiamine Deficiency. Cells. 2021 Sep 29;10(10):2595. [CrossRef]
- Schröter C, Hipler UC, Wilmer A, Künkel W, Wollina U. Generation of reactive oxygen species by Candida albicans in relation to morphogenesis. Arch Dermatol Res. 2000 May;292(5):260-4. [CrossRef]
- Ballard JWO, Towarnicki SG. Mitochondria, the gut microbiome and ROS. Cell Signal. 2020 Nov;75:109737. [CrossRef]
- Swank, Z., Senussi, Y., Manickas-Hill, Z., Yu, X.G., Li, J.Z., et al. (2022) Persistent Circulating SARS-CoV-2 Spike Is Associated with Post-Acute COVID-19 Sequelae. Clinical Infectious Diseases, 2022, ciac722. [CrossRef]
- Shandilya S, Kumar S, Kumar Jha N, Kumar Kesari K, Ruokolainen J. Interplay of gut microbiota and oxidative stress: Perspective on neurodegeneration and neuroprotection. J Adv Res. 2021 Sep 17;38:223-244. [CrossRef]
- Zhang J, Wang Z, Zhang X, Dai Z, Zhi-Peng W, Yu J, et al. Large-Scale Single-Cell and Bulk Sequencing Analyses Reveal the Prognostic Value and Immune Aspects of CD147 in Pan-Cancer. Front Immunol. 2022 Apr 6;13:810471. [CrossRef]
- Hayes B, Stanley J, Peppers BP. COVID-19 Recurrence Without Seroconversion in a Patient With Mannose-Binding Lectin Deficiency. Allergy Rhinol (Providence). 2021 Jun 11;12:21526567211024140. [CrossRef]
- Kim HS. Do an Altered Gut Microbiota and an Associated Leaky Gut Affect COVID-19 Severity? mBio. 2021 Jan 12;12(1):e03022-20. [CrossRef]
- Rathmann W, Kuss O, Kostev K. Incidence of newly diagnosed diabetes after Covid-19. Diabetologia. 2022 Jun;65(6):949-954. [CrossRef]
- Sakurai K, Narita D, Saito N, Ueno T, Sato R, Niitsuma S, Takahashi K, Arihara Z. Type 1 diabetes mellitus following COVID-19 RNA-based vaccine. J Diabetes Investig. 2022 Jul;13(7):1290-1292. [CrossRef]
- Moon H, Suh S, Park MK. Adult-Onset Type 1 Diabetes Development Following COVID-19 mRNA Vaccination. J Korean Med Sci. 2023 Jan 9;38(2):e12. [CrossRef]
- Roe K. How major fungal infections can initiate severe autoimmune diseases. Microb Pathog. 2021 Dec;161(Pt A):105200. [CrossRef]
- Nikolic DM, Dimitrijevic-Sreckovic V, Ranin LT, Stojanovic MM, Ilic ID, Gostiljac DM, et al. Homeostatic microbiome disruption as a cause of insulin secretion disorders. Candida albicans, a new factor in pathogenesis of diabetes: A STROBE compliant cross-sectional study. Medicine (Baltimore). 2022 Nov 11;101(45):e31291. [CrossRef]
- Paray BA, Albeshr MF, Jan AT, Rather IA. Leaky Gut and Autoimmunity: An Intricate Balance in Individuals Health and the Diseased State. International Journal of Molecular Sciences. 2020; 21(24):9770. [CrossRef]
- Xiao P, Hu Z, Lang J, Pan T, Mertens RT, Zhang H, et al. Mannose metabolism normalizes gut homeostasis by blocking the TNF-α-mediated proinflammatory circuit. Cell Mol Immunol. 2023 Feb;20(2):119-130. [CrossRef]
- Torretta S, Scagliola A, Ricci L, Mainini F, Di Marco S, Cuccovillo I, Kajaste-Rudnitski A, Sumpton D, Ryan KM, Cardaci S. D-mannose suppresses macrophage IL-1β production. Nat Commun. 2020 Dec 11;11(1):6343. [CrossRef]
- Dong, L., Xie, J., Wang, Y. et al. Mannose ameliorates experimental colitis by protecting intestinal barrier integrity. Nat Commun 13, 4804 (2022). [CrossRef]
- Zhang W, Cheng H, Gui Y, Zhan Q, Li S, Qiao W, et al. Mannose Treatment: A Promising Novel Strategy to Suppress Inflammation. Front Immunol. 2021 Sep 27;12:756920. [CrossRef]
- Dhanalakshmi, M., Sruthi, D., Jinuraj, K.R. et al. Mannose: a potential saccharide candidate in disease management. Med Chem Res 32, 391–408 (2023). [CrossRef]
- Sun J, Wang F, Hong G, Pang M, Xu H, Li H, et al. Antidepressant-like effects of sodium butyrate and its possible mechanisms of action in mice exposed to chronic unpredictable mild stress. Neurosci Lett. 2016 Apr 8;618:159-166. [CrossRef]
- Mayr A, Hinterberger G, Dierich MP, Lass-Flörl C. Interaction of serotonin with Candida albicans selectively attenuates fungal virulence in vitro. Int J Antimicrob Agents. 2005 Oct;26(4):335-7. [CrossRef]
- Roth W, Zadeh K, Vekariya R, Ge Y, Mohamadzadeh M. Tryptophan Metabolism and Gut-Brain Homeostasis. Int J Mol Sci. 2021 Mar 15;22(6):2973. [CrossRef]
- Hodgkinson K, El Abbar F, Dobranowski P, Manoogian J, Butcher J, Figeys D, et al.. Butyrate's role in human health and the current progress towards its clinical application to treat gastrointestinal disease. Clin Nutr. 2023 Feb;42(2):61-75. [CrossRef]
- González Delgado S, Garza-Veloz I, Trejo-Vazquez F, Martinez-Fierro ML. Interplay between Serotonin, Immune Response, and Intestinal Dysbiosis in Inflammatory Bowel Disease. Int J Mol Sci. 2022 Dec 9;23(24):15632. [CrossRef]
- Yusufu I, Ding K, Smith K, Wankhade UD, Sahay B, Patterson GT, et al. A Tryptophan-Deficient Diet Induces Gut Microbiota Dysbiosis and Increases Systemic Inflammation in Aged Mice. Int J Mol Sci. 2021 May 8;22(9):5005. [CrossRef]
- Thomas, R.L., Jiang, L., Adams, J.S. et al. Vitamin D metabolites and the gut microbiome in older men. Nat Commun 11, 5997 (2020). [CrossRef]
- Książek A, Zagrodna A, Bohdanowicz-Pawlak A, Lwow F, Słowińska-Lisowska M. Relationships between Vitamin D and Selected Cytokines and Hemogram Parameters in Professional Football Players-Pilot Study. Int J Environ Res Public Health. 2021 Jul 2;18(13):7124. [CrossRef]
- Chen YC, Sung HC, Chuang TY, Lai TC, Lee TL, Lee CW, et al. Vitamin D3 decreases TNF-α-induced inflammation in lung epithelial cells through a reduction in mitochondrial fission and mitophagy. Cell Biol Toxicol. 2022 Jun;38(3):427-450. [CrossRef]
- Bouzid D, Merzouki S, Bachiri M, Ailane SE, Zerroug MM. Vitamin D3 a new drug against Candida albicans. J Mycol Med. 2017 Mar;27(1):79-82. [CrossRef]
- Lei J, Xiao W, Zhang J, Liu F, Xin C, Zhou B, et al. Antifungal activity of vitamin D3 against Candida albicans in vitro and in vivo. Microbiol Res. 2022 Dec;265:127200. [CrossRef]
- Ragab D, Soliman D, Samaha D, Yassin A. Vitamin D status and its modulatory effect on interferon gamma and interleukin-10 production by peripheral blood mononuclear cells in culture. Cytokine. 2016 Sep;85:5-10. [CrossRef]
- Nielsen FH. Magnesium deficiency and increased inflammation: current perspectives. J Inflamm Res. 2018 Jan 18;11:25-34. [CrossRef]
- Nabin K Shrestha, Patrick C Burke, Amy S Nowacki, James F Simon, Amanda Hagen, et al. Effectiveness of the Coronavirus Disease 2019 Bivalent Vaccine, Open Forum Infectious Diseases, Volume 10, Issue 6, June 2023, ofad209, . [CrossRef]
- Wang, K., Chen, W., Zhang, Z., Deng, Y., et al. (2020) CD147-Spike Protein Is a Novel Route for SARS-CoV-2 Infection to Host Cells. Signal Transduction and Targeted Therapy, 5, 283. [CrossRef]
- Chambers, P.W. (2021) Basigin Binds Spike S on SARS-CoV2. Open Access Library Journal, 8: e8064. [CrossRef]
- Ricke, D.O., Immediate onset signatures of autoimmune diseases after vaccination (2023) Global Translational Medicine 2023, 2(3), 1455 . [CrossRef]
- Salsone M, Signorelli C, Oldani A, Alberti VF, Castronovo V, Mazzitelli S, et al. NEURO-COVAX: An Italian Population-Based Study of Neurological Complications after COVID-19 Vaccinations. Vaccines. 2023; 11(10):1621. [CrossRef]
- Eriksson O, Hultström M, Persson B, Lipcsey M, Ekdahl KN, Nilsson B, Frithiof R. Mannose-Binding Lectin is Associated with Thrombosis and Coagulopathy in Critically Ill COVID-19 Patients. Thromb Haemost. 2020 Dec;120(12):1720-1724. [CrossRef]
- Hoffa, Charles, Doctor who vaccinated 900 calls blood clots at capillary level an ‘absolutely new phenomenon’ World Tribune 2021 https://www.worldtribune.com/doctor-who-vaccinated-900-calls-blood-clots-at-capillary-level-an-absolutely-new-phenomenon/].
- Luca M, Chattipakorn SC, Sriwichaiin S, Luca A. Cognitive-Behavioural Correlates of Dysbiosis: A Review. Int J Mol Sci. 2020 Jul 8;21(14):4834. [CrossRef]
- Pham VT, Dold S, Rehman A, Bird JK, Steinert RE. Vitamins, the gut microbiome and gastrointestinal health in humans. Nutr Res. 2021 Nov;95:35-53. [CrossRef]
- Lonsdale D, Marrs C. Thiamine Deficiency Disease, Dysautonomia, and High Calorie Malnutrition, 1st Edition, 2017, Academic Press.
- Tetsuka, S., Hashimoto, R. Alcohol-Related Central Nervous System Disorders Associated with Vitamin B Deficiency. SN Compr. Clin. Med. 3, 528–537 (2021). [CrossRef]
- Huang SC, Wei JC, Wu DJ, Huang YC (2010) Vitamin B(6) supplementation improves pro-inflammatory responses in patients with rheumatoid arthritis. Eur J Clin Nutr 64(9):1007- 1013. [CrossRef]
- Bzioueche H, Simonyté Sjödin K, West CE, Khemis A, Rocchi S, Passeron T, Tulic MK. Analysis of Matched Skin and Gut Microbiome of Patients with Vitiligo Reveals Deep Skin Dysbiosis: Link with Mitochondrial and Immune Changes. J Invest Dermatol. 2021 Sep;141(9):2280-2290. [CrossRef]
Figure 1.
The serotonin and kynurenine pathways.
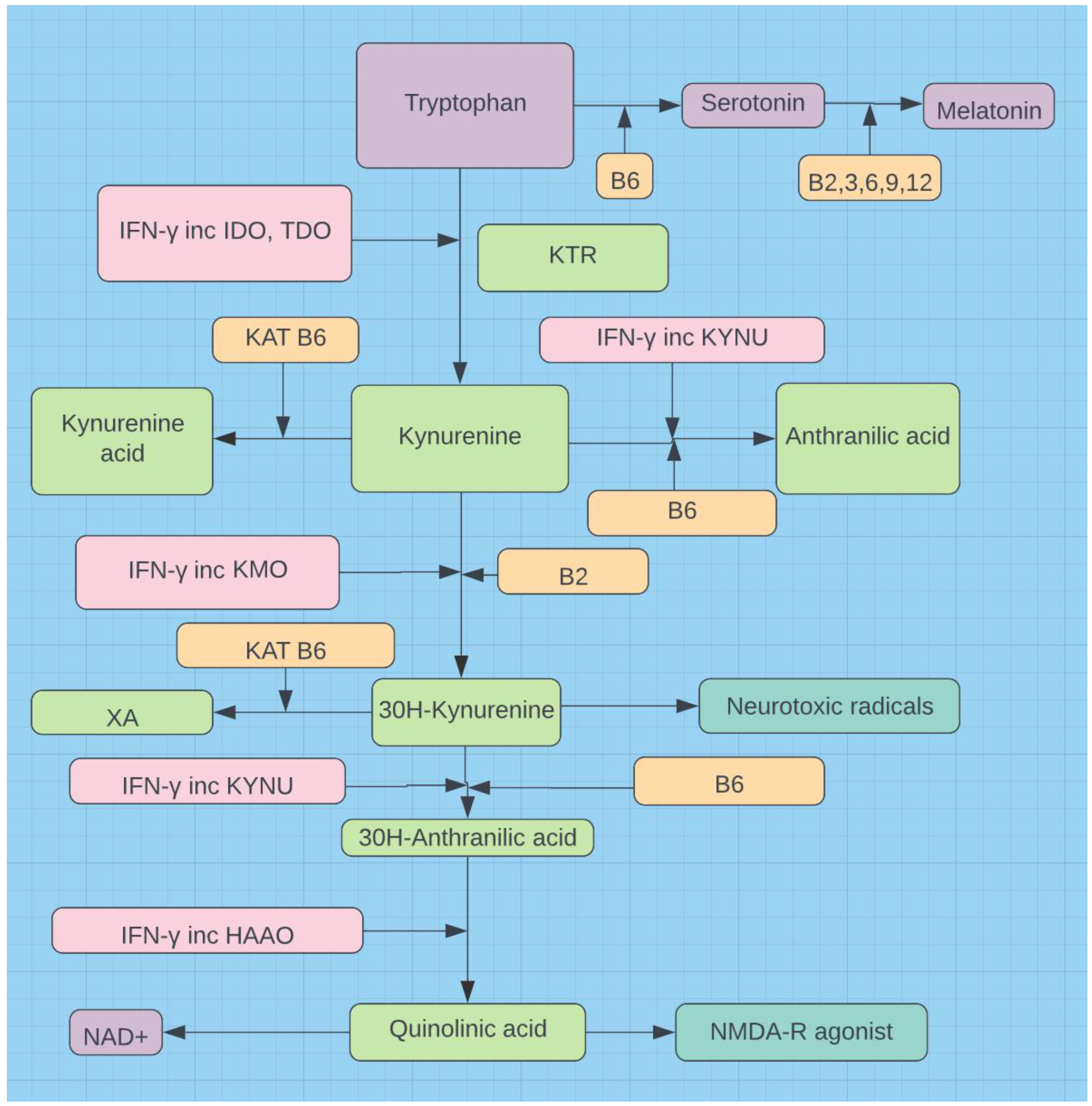
Figure 2.
ATP production per molecule of glucose drops from 34 to 2 in the environment of severe oxidative stress.
Figure 2.
ATP production per molecule of glucose drops from 34 to 2 in the environment of severe oxidative stress.
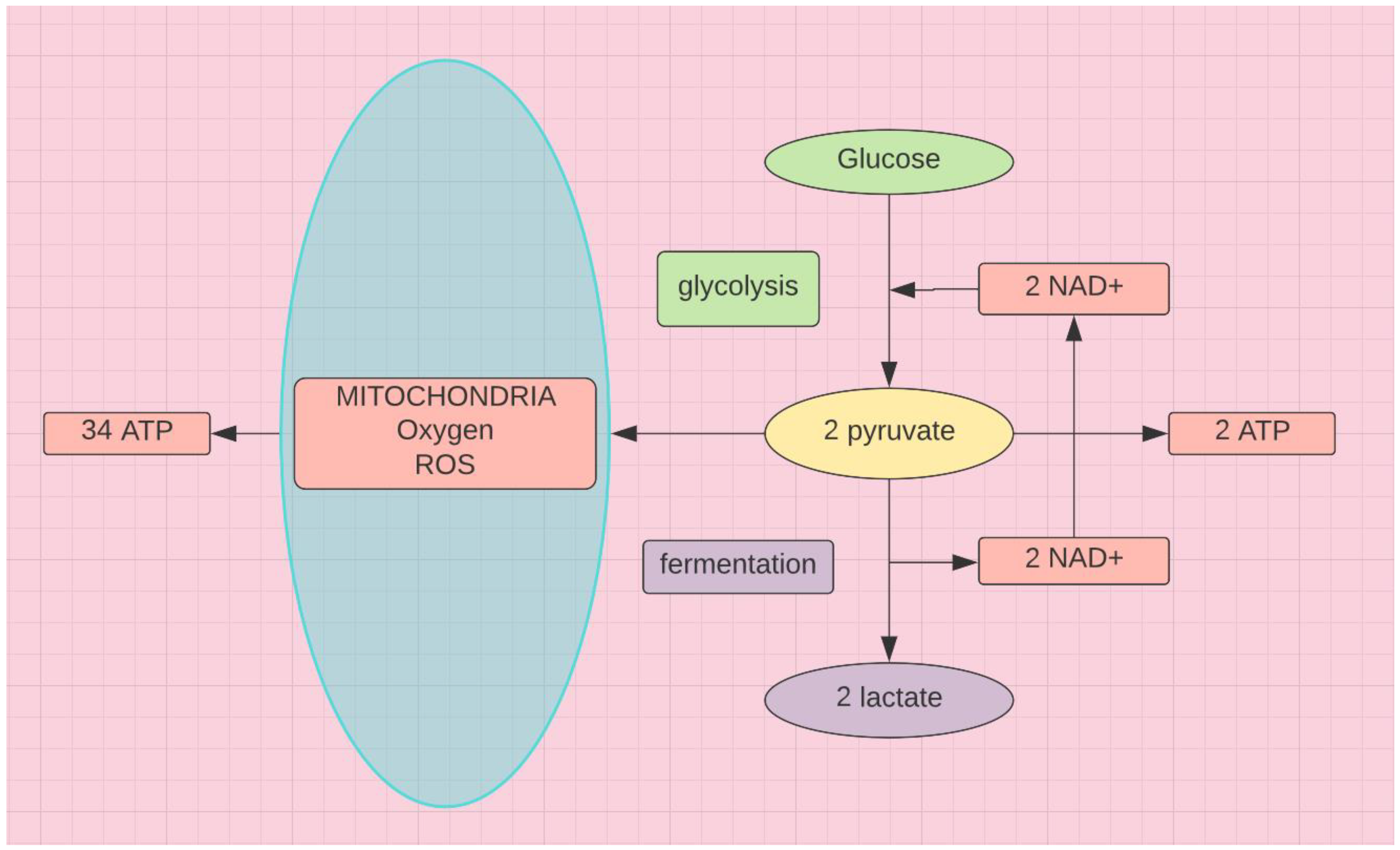
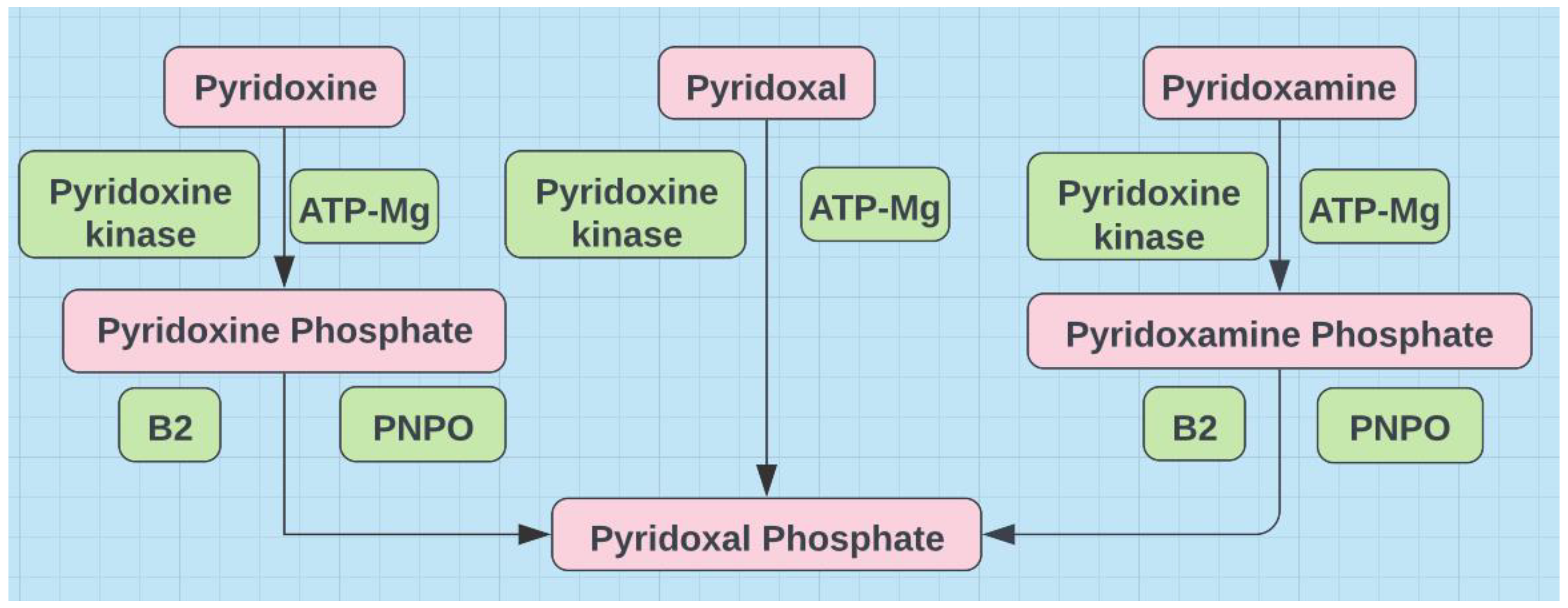
Figure 3.
Pyridoxal phosphate (P5P) is the active form and requires B2 and magnesium for synthesis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



