
Submitted:
09 October 2023
Posted:
17 November 2023
You are already at the latest version
Abstract
It is generally accepted that atherosclerosis is a chronic inflammatory disease. The link between atherosclerosis and other inflammatory diseases such as psoriasis, type 2 diabetes mellitus (T2DM), and rheumatoid arthritis (RA) via metabolic, inflammatory, and immunoregulatory pathways is well established. The aim of our review was to summarize the associations between selected microRNAs (miRs) and long non-coding RNAs (lncRNAs) and atherosclerosis, psoriasis, T2DM, and RA. MicroRNAs are short noncoding RNA molecules involved in cell signaling, intracellular communication, and gene expression. We reviewed the role of miR-146a, miR-210, miR-143, miR-223, miR-126, miR-21, miR-155, miR-145, miR-200, miR-133, miR-135, miR-221, miR-424, and let-7 in atherosclerosis, psoriasis, T2DM, and RA. LncRNAs are RNA transcripts longer than 200 nucleotides that are involved in cellular processes such as apoptosis, metabolism, inflammation, cell differentiation, and proliferation. We evaluated the role of lncRNA-H19, lncRNA-MEG3, lncRNA-UCA1, and lncRNA-XIST in atherosclerosis and psoriasis, T2DM, and RA. Extracellular vesicles (EVs) are a method of intracellular signal transduction. Their function depends on surface expression, cargo, and the cell from which they originate.
Keywords:
Atherosclerosis
; psoriasis
; microRNA
; extracellular vesicles
; long non-coding RNA
1. Introduction
It is now widely accepted that inflammation plays an important role in atherosclerosis and cardiovascular disease (CVD). It is also involved in all phases of the atherosclerotic process, from early endothelial dysfunction to advanced atherosclerotic plaques [1,2]. People with inflammatory diseases such as psoriasis, rheumatoid arthritis, chronic obstructive pulmonary disease, diabetes, and periodontitis may be at increased risk for CVD [1,2]. Elevated circulating markers such as C-reactive protein (CRP), interleukins (IL) (IL-1β, IL-6, IL-8, IL-12, IL-17, IL-32, IL-36), tumor necrosis factor α (TNFα), and vascular endothelial growth factor (VEGF) are commonly found in conditions of low-grade inflammation, such as CVD, psoriasis, RA, and DM [3,4,5]. Immune cells of innate or adaptive immunity (monocytes, neutrophils, mast cells, macrophages, and lymphocytes) are thought to play an important role in chronic inflammation [6]. Moreover, they were reported to be responsible for the production of inflammatory cytokines [3,4,5].
Psoriasis is a multifactorial disease with a strong genetic background [7,8,9]. Psoriasis affects approximately 2.5% of the population worldwide [7,10]. HLA-Cw6 is one of the most strongly associated psoriasis susceptibility alleles [7]. Psoriasis is considered to be primarily a T cell-mediated immune disease, and Th1 and Th17 lymphocytes are expected to be the main players via the release of inflammatory cytokines (i.e. IL-1α, IL-1β, IL-17, IL-22, TNF-α) leading to keratinocyte proliferation, migration of inflammatory cells and enhanced inflammatory response in the skin [11,12,13]. Patients with psoriasis can also be affected by psoriatic arthritis and are at greater risk for CVD and metabolic syndrome. TNF-α and IL-1 are the driving forces of chronic systemic inflammation that predispose patients with psoriasis to CVD and metabolic syndrome [11].
Type 2 diabetes mellitus (T2DM) is a chronic multisystemic disease that is characterized by hyperglycemia and insulin resistance. Hyperglycemia promotes the formation of reactive oxygen species (ROS) that are involved in endothelial dysfunction and inflammation. Metabolic syndrome (high blood pressure, hyperglycemia, abdominal obesity, high serum triglycerides and low serum high-density cholesterol (HDL)) is a major predisposing factor for both CVD and T2DM. CVD is a major cause of death for patients with T2DM [5]. The prevalence is increasing, 784 million people are expected to suffer from T2DM by 2045 [14].
Rheumatoid arthritis (RA) is a chronic inflammatory disease that mainly affects the joints, however, extraarticular involvement is not uncommon [15]. The global prevalence of RA in 2017 was 0.27%, however, there is a strong geographical and socioeconomic factor to the disease (the prevalence is higher in northern latitudes and higher-income countries) [16]. Dendritic cells play a crucial role in initiating and maintaining inflammation in RA. They modulate Th1, Th2, and Th17 lymphocyte activity by secreting inflammatory cytokines (IL-1 β, IL-6, IL-12, IL-18, IL-23, TGF- β). TNF- α, IL-1 β, and IL-6 play a key role in establishing and maintaining inflammation in the synovia [4].
The aim of our review was to summarize the associations between selected microRNAs (miRs) and long non-coding RNAs (lncRNAs) and atherosclerosis, psoriasis, T2DM, and RA.
2. METHODS
We reviewed articles that investigated the roles of genetic and epigenetic factors in the pathogenesis of atherosclerosis and psoriasis, T2DM, and RA. We used the PubMed database as our bibliographic source. The following search sequence was used to obtain articles: (atherosclerosis) AND ((psoriasis)) OR (T2DM) OR (diabetes) OR (type 2 diabetes mellitus) OR (rheumatoid arthritis) OR (RA)) AND ((microRNA) OR (miR) OR (long non-coding RNA) OR (lncRNA) OR (extracellular vesicles) OR (EVs) OR (exosomes) OR (microvesicles) OR (apoptotic bodies)).
Only articles written in English were included. Abstracts of all acquired articles were read. Articles, that had promising abstracts, were then read in their entirety. A targeted search of the PubMed database with specific search terms (e.g. (psoriasis) AND/OR (atherosclerosis) AND ((miR-200) OR (microRNA-200))) was then conducted to search for potential additional articles. We also checked publications that were cited in the articles that we found. The last search in the PubMed database was conducted in June 2023.
3. PATHOGENESIS OF ATHEROSCLEROSIS IN SELECTED INFLAMMATORY CONDITIONS
Several studies have shown that psoriasis and atherosclerosis are linked by dyslipidemia, increased levels of lipoprotein a and altered metabolic, inflammatory and immunoregulatory pathways [17,18,19]. In addition, the immune system is thought to play a crucial role in the pathogenesis of psoriasis and various innate and adaptive immune cells as well as proinflammatory mediators are also involved in the development of atherosclerosis [1,2,17].
One of the crucial cytokines involved in the pathogenesis of psoriasis and RA, IL-17, has been reported to play an important role in the production of reactive oxygen species (ROS) [10,20]. ROS are on the list of factors that can cause endothelial dysfunction [6]. IL-17 is one of the main factors for endothelial dysfunction in patients with RA [21]. Hyperglycemia causes endothelial dysfunction in T2DM through increased oxidative stress via ROS and reactive nitrogen species (RNS) [22]. Endothelial dysfunction causes changes in the production of endothelial cells (i.e. expression of adhesion molecules, ROMO1 expression and production of inflammatory cytokines…), and they lead to morphological changes in the vessel wall (inflammation, atherosclerotic changes) [1,2,6]. IL-1β, IL-6, IL-17, TNF-α and hs-CRP are involved in the pathogenesis of atherosclerosis, psoriasis and RA [23,24].
Endothelial dysfunction measured by functional ultrasound studies of the brachial artery, and carotid intima-media thickness (CIMT) were reported to be an important marker of subclinical atherosclerosis in psoriasis [25,26]. A meta-analysis involving psoriasis patients showed that psoriasis patients had significantly thicker CIMT (WMD 0.11 mm; 95% CI 0.08-0.15) and impaired flow-mediated dilation (WMD -2.79%; -4.14% to -1.43%) than the control group [26]. Psoriasis patients with a mean age > 45 years had a much thicker CIMT, while impaired FMD appeared to be more pronounced in psoriasis patients with a mean age of <45 years. The authors concluded in their meta-analysis that patients with psoriasis were associated with excessive risk of subclinical atherosclerosis [26].
Patients with T2DM have a significantly higher risk of cardiovascular events compared to people without T2DM [27]. A meta-analysis showed that the CIMT of patients with T2DM was thicker (0.13 (95% CI 0.12-0.14) mm thicker) compared to controls [28].
A meta-analysis of studies examining the CIMT in patients with RA showed that patients with RA have a significantly thicker CIMT compared to controls [29].
Dyslipidemia plays a key role in atherogenesis. Although the mechanisms by which LDL provokes lesion formation are not fully understood, it is speculated that oxidative modification of LDL particles leads to the accumulation of oxidized LDL in the vessel wall, the development and progression of plaques [1,2]. Oxidized LDL cholesterol has been reported to induce proinflammatory and proatherogenic effects via various mechanisms (e.g., via NF-κB, caspase-1 activation) [1,2,30,31]. Patients with psoriasis have an atherogenic lipid profile with elevated LDL cholesterol levels [32]. In addition, impaired HDL function in psoriasis patients also increases the production of oxidized LDL cholesterol [32].
Patients with T2DM have a characteristic proatherogenic lipid profile with low HDL levels and elevated LDL and triglyceride levels [33]. The so-called lipid paradox has been reported in patients with RA. In patients in the active stage of the disease, total cholesterol, LDL and HDL levels are lowered, while they increase with the administration of drugs targeting the inflammatory pathways in RA. The relationship between lipid profile and CVD risk is therefore U-shaped. The anti-inflammatory and antiatherogenic functions of HDL are impaired in patients with RA [34].
Macrophages filled with oxidized LDL cholesterol (i.e. foam cells) play a crucial role in the development of the atherosclerotic process [1,2]. Foam cells and other anti-gen-presenting cells (e.g. dendritic cells) can present oxidized LDL particles and heat shock proteins to the innate and adaptive immune system, leading to a chronic, low-grade inflammatory response [31].
4. GENE EXPRESSION IN PSORIASIS, T2DM, AND RA PROMOTING ATHEROSCLEROSIS DEVELOPMENT
The gene expression profiles of psoriasis (GSE30999) and atherosclerosis (GSE28829) were downloaded from the Gene Expression Omnibus (GEO) database and the common differentially expressed genes (DEGs) of psoriasis and atherosclerosis were identified. Functional analysis of DEGs emphasizes the important role of chemokines and cytokines in these two diseases. In addition, the lipopolysaccharide-mediated signaling pathway is closely related to both. Finally, 16 important hub genes were identified using cytoHubba, including LCP2, CD53, LYN, CSF2RB, C1QB, MMP9, PLEK, PTPRC, FYB, IL1RN, RAC2, CCL5, IRF8, BCL2A1, NCF2 and TLR2 [37]. Results of the study revealed some common pathogenetic mechanisms of psoriasis and atherosclerosis [37].
The IL-23R rs6682925T/C polymorphism and inheritance of the HLA, FUT2, UBE2L3, and SH2B3 gene variants increase the risk of major adverse cardiovascular events (MACE) in psoriasis [12,38]. The IL-23A polymorphism rs2066808 is associated with an increased risk of developing psoriasis and could also increase the genetic risk for premature CAD [39,40].
Eder and colleagues conducted a study on 411 patients with psoriasis and discovered that the HLA-C*06-02 and HLA-B*13-02 alleles are associated with a higher risk of atherosclerosis [41].
Mutations in CARD14, which is a NF-κB regulatory protein, have been described in psoriatic patients. CARD14 is also expressed in the cells of the aortic endothelium, which could result in atherosclerotic progression and increased systemic inflammation [42].
Gene expression profiles of atherosclerosis and T2DM were obtained from the GEO database to identify DEGs of atherosclerosis and T2DM. GSE28829 (atherosclerosis) and GSE20966 (T2DM) were used as test sets, the results were then validated with GSE43292 (atherosclerosis) and GSE25724 (T2DM). Genes related to immune activation and cytokines were found to have important roles in the pathogenesis of these diseases. Three important hub genes were identified using cytoHubba after validation with GSE43292 and GSE25724 (CD4, PLEK, and THY1). Further validation using clinical samples identified CD4 and PLEK as the key genes in atherosclerosis and T2DM [43].
Gene expression profiles of atherosclerosis (GSE14905) and RA (GSE55235 and GSE55457) were obtained from the GEO database and DEGs were identified. 12 important hub genes were identified using cytoHubba: CYBB, LAPTM5, CSF1R, HCK, ITGAM, CD86, C1QA, ITGB2, PTPRC, CTSS, LCP2 and CD53. Expression levels of the following genes were then verified: CSF1R, CD86, PTPRC and CD53. Genes related to phagocytosis, neutrophil activation and leukocyte migration were identified as being significantly enriched in the pathogenesis of RA and atherosclerosis [44].
5. THE ROLE OF MICRORNAS IN ATHEROSCLEROSIS, PSORIASIS, T2DM AND RA
MicroRNAs (miRs) are classified as short, non-coding RNA molecules that are involved in cell signaling, intracellular communication, and gene expression [45]. They regulate both physiological and pathological processes including autoimmune disorders, metabolism, inflammation, and cancerogenesis, just to name a few [46]. In the last couple of years, research into microRNAs has yielded several specific microRNAs that have been identified as being involved in atherosclerosis and other inflammatory diseases.
5.1. MiR-146a
MiR-146a has an anti-inflammatory function as it inhibits the NF-κB pathway and modulates the expression of cytokines (IL-6, IL-8, TNFα, and others) [47,48]. It also mediates the proliferation and migration of VSMCs [49,50]. Elevated levels of miR-146a have been found in atherosclerotic lesions and in the blood of patients with psoriasis. [47,51,52,53]. Leal and colleagues also reported a significant association between miR-146a levels and the PASI score as well as the body surface area index [47]. Ele-Refaei and El-Esawy described a decrease in miR-146a levels after 12 weeks of aggressive treatment [53].
The expression levels of miR-146a correlate with the expression levels of IL-6 and TNFα in patients with atherosclerosis. MiR-146a levels are higher in patients with ca-rotid atherosclerosis and correlate with the degree of stenosis and stability of the atherosclerotic plaque [48,54,55,56,57]. According to Zhelankin and colleagues, miR-146a-5p could be used as a biomarker for acute coronary syndrome (ACS) [55]. MiR-146a polymorphism rs2910164 has been extensively studied in the context of atherosclerosis and psoriasis. The G allele of rs2910164 is associated with an increased risk of psoriasis [58]. Two meta-analyses concluded that it may be associated with a lower risk of CHD, but further research is needed [59,60].
Alipoor et al. performed a meta-analysis and concluded that miR-146a expression is downregulated in whole blood and PBMCs from patients with T2DM compared to controls [61]. A more recent meta-analysis by Zhu and Leung also concluded that miR-146a is downregulated in PBMCs, but also that it is upregulated in adipose tissue [62].
However, a meta-analysis concluded that the rs2910164 polymorphism is not associated with susceptibility to T2DM [63]. Plasma miR-146a is a marker of subclinical atherosclerosis in patients with T2DM and correlates with CIMT [56]. According to Shen and colleagues, the CC genotype of the rs2910164 polymorphism was associated with an increased risk of plaque vulnerability in patients with T2DM, but not with carotid atherosclerosis [64].
In RA, circulating and PBMC levels of miR-146a are elevated and correlate with erythrocyte sedimentation rate and disease activity [65,66]. The CC genotype of the rs2910164 polymorphism is considered a protective factor for RA in the Egyptian-Caucasian population according to Liu et al [66]
MiR-146a is an important biomarker in the inflammatory diseases studied in this article. It is upregulated in patients with atherosclerosis, psoriasis and RA. It is downregulated in patients with T2DM, but is recognized as a marker for atherosclerosis in patients with T2DM as it correlates with CIMT. The available data suggest that it is a promising biomarker for atherosclerosis in a healthy population as well as in patients with psoriasis, T2DM or RA. New research is needed to investigate its potential as a biomarker for atherosclerosis in patients with psoriasis or RA.
5.2. MiR-210
MiR-210 is involved in VSMC migration and endothelial cell apoptosis [67]. Hypoxic conditions induce the expression of miR-210 in the endothelium, and its expression is also enhanced by TGF-β and IL-23 [68,71].
Increased expression of miR-210 has been found both in the serum of patients with atherosclerosis and in the plaques themselves [54,69]. Eken and colleagues reported decreased miR-210 expression in unstable atherosclerotic plaques and concluded that miR-210 stabilizes the fibrotic cap of advanced atherosclerotic lesions [70].
MiR-210 also has an immunomodulatory function, affecting the levels of IL-10 and IL-17 and the differentiation of Th1 and Th17 cells [71,72]. Zhao et al. and Wu et al. reported increased miR-210 expression in CD4+ cells as well as in psoriatic plaques in patients with psoriasis compared to healthy controls [71,72].
Plasma levels of miR-210 were elevated in patients with T2DM compared to healthy controls [73]. In another study, higher levels also correlated with higher BMI in T2DM patients [74]. Amr and colleagues reported increased plasma miR-210 levels in T2DM patients compared to healthy controls, as well as higher levels in T2DM patients with CHD compared to those without CHD [75]. Zhou and colleagues reported decreased expression of miR-210 in erythrocytes compared to healthy controls [76]. A small study investigated the expression of miR-210 in plasma-derived extracellular vesicles in T2DM patients with ischemic heart disease. They reported a downregulation of miR-210 compared to healthy controls and concluded that plasma-derived extracellular vesicles and their cargo could be a potential biomarker for ischemic heart disease in T2DM [77]. Zhu and Leung concluded that miR-221 is significantly upregulated in the serum of T2DM patients and is one of the most important biomarkers for T2DM [62].
In patients with RA, serum levels of miR-210 were lower compared to healthy controls and they were inversely correlated with TNF-α and IL-1β. Abdul-Maksoud et al. concluded that miR-210 could be used as a biomarker for RA [78]. However, Huang et al. did not report a statistically significant difference in PBMC miR-210 levels between patients with RA and healthy controls [79].
The importance of miR-210 as a biomarker for T2DM and atherosclerosis in patients with T2DM makes it an interesting target for future studies and a potential target for treatments. Larger studies on patients with atherosclerosis, psoriasis and RA are needed, as the sample sizes of the studies investigated were quite small (< 50 patients per group).
5.3. MiR-143
The expression of miR-143 in endothelial cells is upregulated by stable laminar blood flow [80]. It is then transferred to VSMCs via extracellular vesicles, where it has an antiatherogenic effect by regulating VSMC proliferation and differentiation [80,81]. MiR-143 has been shown to promote ROS production in mouse cardiomyocytes, but its role in oxidative stress in humans is not known [82].
In patients with ACS, plasma levels of miR-143 were significantly decreased compared to healthy controls and inversely correlated with the degree of carotid artery stenosis [83].
The role of miR-143 in psoriasis is still unclear. Løvendorf and colleagues reported significantly higher levels of miR-143 in PBMCs and a positive correlation with the PASI score [84]. A more recent study by Zheng and colleagues found decreased levels of miR-143 in patients with psoriasis vulgaris and a negative correlation with disease severity [85]. Their sample size was also larger; however, further studies are needed to evaluate the role of miR-143 in PBMC in psoriasis.
MiR-143-3p is significantly upregulated in VSMCs in patients with T2DM and is a promising biomarker for T2DM according to the results of a meta-analysis conducted by Zhu and Leung [62]. Its overexpression has been associated with insulin resistance and dysregulation of glucose metabolism in animal models.
It is also overexpressed in the plasma of patients with RA, where a positive correlation between its levels and CRP as well as the clinical swollen joint count and subjective pain score has been described [86].
MiR-143 is an interesting target for future studies of atherosclerosis in inflammatory diseases due to its effect on VSMCs.
5.4. MiR-223
MiR-223 is involved in the regulation of inflammation, cholesterol metabolism and VSMC migration, proliferation and apoptosis [87,88,89]. It is thought to have an anti-inflammatory function by suppressing the NLRP3 inflammasome, IL-1β and IL-10 [89,90].
Singh and colleagues reported upregulation of miR-223 in patients with unstable coronary artery disease [91]. They suggested that miR-223 may be a marker of plaque instability [91]. Guo and colleagues also found increased expression of miR-223 in patients with coronary atherosclerosis and a significant correlation between miR-223 levels and disease severity [92]. On the other hand, Zhu and colleagues reported decreased miR-223 levels in patients with carotid atherosclerosis and a correlation with plaque stability, although their sample size was smaller [93].
MiR-223 promotes proliferation and inhibits apoptosis in keratinocytes [94]. Løvendorf and colleagues found increased levels of miR-223 in patients with psoriasis and a positive correlation with the PASI score [84]. MiR-223 was significantly downregulated after 3-5 weeks of treatment with methotrexate [84]. Pivarci and colleagues found no significant difference in miR-223 levels between patients with psoriasis and healthy controls, but described a significant downregulation after treatment with etanercept [95]. However, Alatas and colleagues reported a significant downregulation of miR-223 in the blood of patients with psoriasis compared to healthy controls [96]. The correlation between miR-223 levels and psoriasis is unclear, as all three studies examined provided contradictory results.
In the serum of patients with T2DM, miR-223 was significantly downregulated in circulating microvesicles and could even be used as a biomarker for progression from prediabetes to diabetes [97]. Zhu and Leung also concluded that miR-223 is significantly downregulated in the plasma of patients with T2DM [62].
Plasma levels of miR-223 were increased 2.5-fold in patients with RA compared to controls, but there was no correlation with disease activity or Framingham risk scores [90].
5.5. MiR-155
The expression of miR-155 is stimulated by inflammation. TNF-α stimulates its transcription through NF-κB. Its transcription is also stimulated by ox-LDL. MiR-155 is thought to have an anti-inflammatory function as it suppresses TNF-α. In atherosclerosis, authors reported both upregulation and downregulation of miR-155, but they were consistent that it is significantly dysregulated [98,99,100].
Fichtlscherer et al. reported decreased levels of miR-155 in the blood of patients with stable CHD compared to healthy controls [98]. Wang and colleagues also reported reduced miR-155 levels in early coronary atherosclerotic plaques found in healthy heart donors [100]. However, a larger study by Li et al. found increased levels of miR-155 in plasma and atherosclerotic plaques [99]. Expression of miR-155 is stimulated by oxidized LDL and TNF-α [99]. Li and colleagues found that miR-155 has an anti-inflammatory function because it suppresses TNF-α expression [99].
MiR-155 is another microRNA whose levels correlate with the severity of psoriasis [101]. García-Rodríguez and colleagues described increased levels of miR-155 in PBMCs from patients with psoriasis compared to controls, which decreased with disease remission [101]. Alatas and colleagues reported significantly increased expression of miR-155 in patients with psoriasis compared to healthy controls [96]. MiR-155 plays a role in keratinocyte proliferation and inhibition of apoptosis [102].
The meta-analysis conducted by Zhu and Leung concluded that miR-155 is not a suitable biomarker for T2DM, as 7 studies reported upregulation and 11 studies reported downregulation [62].
In RA, miR-155 expression was increased in patients and correlated with serum and plasma levels of TNF-α and IL-1β. Abdul-Maksoud and colleagues reported increased serum levels of miR-155 correlated with TNF-α and IL-1β levels in patients with RA [78]. Elmesmari and colleagues also reported increased levels of miR-155 in mononuclear cells from patients with RA compared to healthy controls [103].
The increased expression of miR-155 in atherosclerosis, psoriasis and RA and its correlation with TNF-α and IL-1β make it an interesting target for future research and a potential therapeutic target.
5.6. MiR-145
MiR-145-5p is involved in a variety of human diseases, including cancers, asthma, rheumatoid arthritis and many others [104]. It is also involved in phenotype switching of VSMCs in atherosclerosis [105]. MiR-145 is involved in the development of psoriasis and RA via regulating the Wnt/β-catenin pathway [106]. Yuan and colleagues reported that it has an anti-inflammatory function by suppressing the production of IL-1β, TNF-α and IL-6 after ischemic injury [107].
Studies showed competing evidence for the expression profile of miR-145-5p in atherosclerosis. Minin and colleagues reported increased expression of miR-145-5p in the serum of hypertensive patients with carotid plaques compared to hypertensive patients without carotid plaques [108]. Li and colleagues reported a slight increase in plasma miR-145 levels in patients with atherosclerosis, but this was not statistically significant [99]. However, several studies reported lower miR-145 levels in patients with atherosclerosis. A study by Zhang and colleagues reported downregulation of miR-145-5p in patients with coronary stenosis [109]. Lv and colleagues also reported decreased plasma levels of miR-145 in patients with ACS and a strong inverse correlation of miR-145 levels with the degree of carotid artery stenosis [110]. Meng et al. reported decreased plasma levels of miR-145 in patients with ACS compared to controls and a negative correlation with coronary stenosis [83]. Two miR-145 polymorphisms (rs353291 and rs41291957) are also associated with atherosclerosis [83,111].
The expression of miR-145-5p in the serum of patients with psoriasis was lower compared to healthy controls [106]. Wang and Cao also claimed that upregulation of miR-145-5p would inhibit the progression of psoriasis [106].
In adipose tissue of patients with T2DM, miR-145-5p is significantly upregulated and was recognized as a potential biomarker for T2DM in the meta-analysis by Zhu and Leung [62].
In RA, miR-145-5p was overexpressed in fibroblast-like synoviocytes and in the plasma of patients with RA. [86,112]. Hong et al. reported increased expression of miR-145 in fibroblast-like synoviocytes (FLS) from patients with RA compared to patients with osteoarthritis [112]. Increased expression of miR-145 promotes the expression of matrix metalloproteinases in FLS from patients with RA [113].
It is difficult to assess the role of miR-145 in the studied diseases, therefore further studies are needed. The studies conducted in patients with psoriasis, T2DM and RA were relatively small. On the other hand, the studies in patients with atherosclerosis were larger, but their conclusions are different.
5.7. MiR-200
The miR-200 family (miR-200a, miR-200b, miR-200c, miR-141 and miR-429) is involved in apoptosis, senescence, inflammation and endothelial dysfunction associated with atherosclerosis [114]. The expression of all members of the miR-200 family is induced by ROS. Members of the MiR-200 family (but especially miR-200c) promote inflammation by inducing ROS, MMP-1, MMP-9, IL-6 and COX-2 and reduce the antioxidant capacity of the cell by decreasing the transcription of manganese superoxide dismutase [114,115].
Plasma levels of miR-200c were significantly increased in patients with carotid plaques and correlated with plaque instability and pro-inflammatory molecules such as MMP-1, MMP-9, IL-6 and COX-2 [115].
In another study, Magenta and colleagues found an upregulation of miR-200c in psoriatic skin lesions and a significant correlation between miR-200c and the severity of psoriasis (PASI score) [114]. In patients with psoriasis, miR-200c also correlates with diastolic dysfunction and cardiac hypertrophy [114]. Circulating levels of miR-200a correlate with arterial stiffness and cardiac hypertrophy in psoriasis patients [114]. Wang and colleagues found a correlation between miR-200a in CD4+ T cells and PASI score, IL-17 levels and IL-23 levels [116].
Kujawa and colleagues reported increased levels of miR-200a/b/c in aortic endothelial cells from patients with T2DM compared to healthy controls. They also reported increased endothelial permeability possibly due to increased levels of the miR-200 family [117]. Ofori and colleagues reported increased levels of miR-200c in pancreatic islets from patients with T2DM compared to controls [118]. The meta-analysis conducted by Zhu and Leung concluded that miR-200 is significantly downregulated in the plasma of patients with T2DM [62]. Lo and colleagues also reported decreased miR-200 family levels in aortic endothelial cells stimulated with high glucose levels [119].
No significant difference in plasma levels of miR-200b-5p and miR-200c-3p was found in patients with RA compared to healthy controls [120].
Further studies are needed to determine the role of miR-200 in atherosclerosis and RA, as only two studies with relatively small sample sizes were found at the time of writing this review. According to the reviewed studies, it is dysregulated in different directions in T2DM and psoriasis.
5.8. MiR-133
MiR-133 has 3 genomic sites in human DNA (on chromosomes 6, 18 and 20) and together with miR-1 plays a crucial role in heart development [121]. MiR-133a upregulates IL-1β and TNF-α and is associated with increased ROS production in mice [122].
Studies have shown that miR-133 has a crucial regulatory role in important atherosclerotic processes: VSMC differentiation, angiogenesis, inflammation and apoptosis [123,124,125,126].
It is upregulated after ischemic heart injury and in patients with hypertension [121,127]. The levels of miR-133a and miR-133b correlate negatively correlate with cardio-ankle vascular index in patients with metabolic syndrome [127]. Plasma miR-133a levels also correlate with the degree of coronary stenosis. Wang et al. concluded that this is a sensitive predictor for the diagnosis of coronary artery disease and AMI [127].
In psoriasis, miR-133 appears to be downregulated. Chicarro and colleagues reported lower miR-133a levels in psoriasis lesions compared to skin samples from healthy controls [128]. The levels of miR-133a-3p increased to those of healthy skin after 3 months of treatment [128].
De Gonzalo-Calvo et al. and Ghasemi et al. reported significantly increased serum levels of miR-133 in patients with T2DM compared to healthy controls [129,130]. Al-Muhtaresh reported increased miR-133 levels in whole blood of patients with T2DM and CAD compared to patients with T2DM without CAD [131]. On the other hand, the meta-analysis by Zhu and Leung concluded that miR-133a-3p is significantly downregulated in patients with T2DM [62].
No studies were found that investigated the role of miR-133 in human patients with RA.
The studies reviewed identify miR-133a as an important marker of coronary stenosis in otherwise healthy patients and patients with T2DM. Further studies with larger sample sizes are needed to validate these findings. The only study that investigated the association between miR-133 and psoriasis was related to psoriatic skin lesions. We found no studies examining circulating levels of miR-133 in psoriasis, making comparison with other conditions difficult.
5.9. MiR-135
MiR-135 is involved in the regulation of endothelial cell proliferation and apoptosis, inflammation and angiogenesis [132,133,134]. MiR-135b is upregulated by ox-LDL and is associated with increased levels of TNF-α, IL-1β, IL-6 and IL-8. It also regulates the expression of I-CAM, V-CAM and E-selectins [132]. Xu and colleagues detected increased expression of miR-135b-5p in the serum of patients with CAD compared to healthy controls, noting that miR-135b-5p may be involved in the migration and proliferation of VSMCs and endothelial cells [135]. However, Li and colleagues reported downregulation of miR-135a-5p in patients with atherosclerosis compared to healthy adults [136]. They claimed that it has an atheroprotective function by suppressing the migration and proliferation of VSMCs in atherosclerosis [136].
Joyce and colleagues reported an upregulation of miR-135b in psoriatic skin lesions compared to healthy skin samples [137]. Chicarro and colleagues confirmed these findings and also discovered that miR-135b levels decreased after treatment and correlated with the PASI score [128].
Sarookhani and colleagues reported increased expression of miR-135a in the plasma of patients with T2DM and prediabetic patients compared to healthy controls [138]. Monfared et al. reported similar results and also found a correlation between serum miR-135a levels and HbA1c levels in prediabetic patients [139]. In another study, Monfared and colleagues reported that miR-135a in saliva could be used as a biomarker for T2DM [140].
Liu and colleagues reported a significant downregulation of miR-135a-5p in synovial tissue from patients with RA compared to healthy controls [141].
To better understand the role of miR-135 in atherosclerosis, psoriasis, T2DM and RA, further studies with larger sample sizes are needed. All studies examined had relatively small samples and reported conflicting results.
5.10. MiR-221
TNF- α induces the expression of miR-221. The function of miR-221-3p in the intima of atherosclerotic vessels is the repression of peroxisome proliferator-activated receptor-γ coactivator 1 α (PGC-1 α) [142]. Post-transcriptional repression of PGC-1 α leads to accumulation of ROS in endothelial cells and induces apoptosis. MiR-221 is highly expressed in VSMCs and endothelial cells, where it regulates proliferation and apoptosis [143].
Under atherosclerotic conditions, it downregulates eNOS, inhibits vascular repair mechanisms and promotes VSMC calcification [139]. Minami and colleagues reported elevated levels of miR-221 in the serum of patients with atherosclerosis [144].
MiR-221 is also upregulated in psoriasis and is strongly associated with psoriatic arthritis [145,146,147]. Zibert and colleagues reported increased expression of miR-221 in psoriatic lesions compared to the skin of healthy controls [145]. Meng et al. and Wade et al. reported increased serum levels of miR-221 in patients with psoriasis and psoriatic arthritis compared to healthy controls [146,147].
MiR-221-3p was significantly upregulated in the serum of patients with T2DM and was reported by Zhu and Leung to be the second most important biomarker for T2DM [62].
The expression of miR-221 is increased in PBMCs and in the serum of patients with RA compared to healthy controls [148,149]. However, Cieśla and colleagues found no significant difference in plasma levels of miR-221 between patients with RA and healthy controls [150].
MiR-221 is upregulated in all diseases that were the subject of this study. MiR-221 plays an important role in maintaining inflammation. It is induced by one of the major inflammatory cytokines TNF- α and causes the accumulation of ROS in cells. Studies in mice have shown that suppression of miR-221 protects against the development of atherosclerosis [151]. MiR-221 has the potential to be a useful clinical biomarker in the future, but more research is needed to test whether it could also be a therapeutic target.
5.11. MiR-424
MiRin-424 appears to have an anti-inflammatory function in the human body [152,153]. Its main target is apolipoprotein C3. Suppression of APOC3 inhibits NF-κB and its signallg pathway. [154]. It regulates the function of VSMCs in atherosclerosis by regulating the NF-κB signalling pathway [152].
The levels of miR-424-5p are downregulated in the peripheral blood of patients with atherosclerosis [152]. Mir-424-5p is also involved in the accumulation of lipids in foam cells [154].
MiR-424 is involved in the regulation of keratinocyte proliferation [153]. Ichihara et al. reported reduced levels of miR-424 in skin samples and serum of patients with psoriasis compared to healthy controls [153]. Alatas et al. also reported reduced miR-424 levels in patients with psoriasis compared to healthy controls [96]. In the hair shafts of psoriasis patients, miR-424 levels were significantly higher compared to healthy controls [155].
At the time of writing, there were no published articles discussing miR-424 in patients with T2DM.
Wang and colleagues reported higher levels of miR-424 in synovial samples from patients with RA compared to patients with osteoarthritis [156].
Since miR-424 suppresses the NF-κB signalling pathway, it has the potential to be a therapeutic target in the future. Enhanced suppression of NF-κB by miR-424 could alleviate inflammation and be useful in the treatment of atherosclerosis, psoriasis, T2DM, RA and other diseases.
5.12. Let-7
Let-7 was the second microRNA to be discovered, hence the atypical name. The let-7 family has a tumor suppressive function and is highly expressed in endothelial cells and VSMCs [114]. They stimulate eNOS, prevent activation of the NF-κB pathway and inhibit apoptosis of endothelial cells [157,158].
In the study by Yu et al [159], let-7b-5p was downregulated in the serum of patients with atherosclerosis compared to healthy controls. Long and colleagues also report decreased levels of let-7b in patients with large-vessel atherosclerosis who have suffered an ischemic stroke compared to healthy controls, and increased levels in patients who have suffered a stroke of other etiology (small-vessel atherosclerosis, cardioembolism, or undetermined etiology) [160]. Huang and colleagues reported a correlation between let-7 serum levels and CIMT in patients with hypertension [161]. Increased expression of let-7c in hypertensive patients with atherosclerosis compared to patients without atherosclerosis was also reported by Minin et al [108].
Let-7a inhibits INF-γ secretion and proliferation of T cells in psoriasis [162]. Hu and colleagues discovered decreased expression of let-7a in patients with psoriasis compared to healthy controls [162]. Pasquali and colleagues also reported decreased expression of let-7b-5p in patients with psoriasis compared to healthy controls [163]. However, Alatas et al. reported upregulation of let-7c-5p and let-7d-5p in patients with psoriasis [96].
In patients with CAD and T2DM, plasma levels of let-7b were found to be associated with regression of T2DM after dietary intervention [164]. In human atherosclerotic plaque tissue, let-7b levels were significantly lower in tissue samples from diabetics compared to non-diabetics [165]. However, Aljaibeji and colleagues reported increased let-7b-5p levels in the serum of patients with T2DM compared to healthy controls [166]. Zhu and Leung found that let-7-f and let-7-i were significantly downregulated in the serum of patients with T2DM in whole blood and serum, respectively [62].
Interestingly, plasma levels of let-7a were significantly increased in patients with RA compared to healthy controls [167]. Ormseth and colleagues included let-7c-5p in a panel of plasma microRNAs that predicted coronary artery calcification in patients with RA. However, no significant association was found between let-7c-5p alone and coronary calcification [168].
We summarize the main findings of the studies that investigated the role of mi-croRNAs in atherosclerosis, psoriasis, T2DM and RA in Table 1.
MiR-146a, miR-200 and miR-223 were downregulated in patients with T2DM but upregulated in atherosclerosis, psoriasis and RA. One possible explanation is that they are downregulated by hyperglycemia. Plasma miR-200 levels are decreased by hyperglycemia, but there is no evidence of the effect of hyperglycemia on miR-146a and miR-223, suggesting that further research is needed to test this hypothesis [119].
In the studies examined, miR-143 was upregulated in psoriasis, T2DM and RA, but downregulated in patients with ACS and carotid stenosis. One possible explanation for these results is that impaired blood flow is a stronger mediator of miR-143 expression than inflammation, which occurs in T2DM, RA and psoriasis.
LONG NON-CODING RNA IN ATHEROSCLEROSIS, PSORIASIS, T2DM AND RA
Long non-coding RNA (lncRNA) are RNA transcripts that are longer than 200 nucleotides and do not encode proteins [169]. In recent years, they have been extensively studied and it has been discovered that they are involved in a variety of cellular processes such as apoptosis, metabolism, inflammation, cell differentiation and proliferation [169].
6.1. LncRNA-H19
One of the lncRNAs that has been associated with atherosclerosis is lncRNA-H19, which is highly evolutionarily conserved and regulates lipid metabolism, cell proliferation, apoptosis, inflammation and angiogenesis [170]. It has been shown that LncRNA-H19 promotes the expression of acid phosphatase 5 (ACP5) [171]. Ox-LDL promotes the expression of lncRNA-H19, which promotes endothelial inflammation induced by Ox-LDL [172]. LncRNA-H19 promotes the secretion of TNF-α, IL-1β, IL-6, increases ROS production and the expression of ICAM1, VCAM1 and selectins. LncRNA-H19 regulates the NF-κB and MAPK signaling pathways and acts as a molecular sponge for let7 [172].
In patients who had suffered an ischemic stroke and had large artery atherosclerosis, the levels of both lncRNA-H19 and ACP5 were significantly increased compared to patients who had suffered an ischemic stroke and had a different stroke etiology [171]. In another study, the lncRNA H19 polymorphism rs217727 was found to be associated with the risk of small vessel ischemic stroke in the Chinese Han population [173]. Bitarafan and colleagues also found elevated levels of lncRNA-H19 in patients with CAD compared to healthy controls, but the difference between the groups was not statistically significant [174]. Several other studies also reported elevated levels of lncRNA-H19 in the serum of patients with atherosclerosis [172,175,176,177].
Through its interaction with miR-130b-3p, lncRNA-H19 is also important for the differentiation of keratinocytes and inhibits apoptosis [178]. However, in psoriasis patients, studies have found that lncRNA-H19 is underexpressed in psoriasis lesions compared to healthy skin controls [178,179].
Fawzy and colleagues, and Cheng and colleagues reported elevated levels of lncRNA-H19 in the plasma of patients with T2DM compared to healthy controls [180,181]. Similar results were obtained by Tello-Flores and colleagues. They reported increased levels of lncRNA-H19 in the serum of patients with T2DM and poor glycemic control compared to healthy controls [182]. On the other hand, Alfaifi and colleagues reported decreased serum levels of lncRNA-H19 in patients with T2DM compared to healthy controls [183]. Similar results were reported by Alrefai and colleagues who detected decreased plasma levels of lncRNA-H19 in patients with T2DM [184].
Mahmoudi and colleagues reported significantly higher lncRNA-H19 levels in patients with RA compared to healthy controls [185]. They also reported a significant correlation between lncRNA-H19 and the severity of RA.
The present data show that lncRNA-H19 is significantly elevated in atherosclerosis, but its role in T2DM is inconclusive. Further studies with larger samples are needed to clarify its role in psoriasis, RA and T2D.
6.2. lncRNA-MEG3
LncRNA-MEG3 is upregulated by TNF-α in adipocytes where it promotes inflammation and downregulated in keratinocytes, where it has an anti-inflammatory function [186,187]. It is involved in endothelial dysfunction, where it enhances inflammation induced by the NLRP3 inflammasome [188]. LncRNA-MEG3 is another lncRNA that appears to be involved in both atherosclerosis and psoriasis as well as tumor suppression through accumulation of p53 [189,190,191]. It regulates proliferation and apoptosis of endothelial cells and vascular smooth muscle cells through miR-26a, miR-21 and miR-223 interactions [191,192]. LncRNA-MEG3 also regulates angiogenesis through modulation of the VEGF signaling pathway and miR-9 interactions [179]. Studies by Bai et al. and Wu et al. showed decreased expression of lncRNA-MEG3 in atherosclerotic arteries in patients with CAD [191,192].
The expression of lncRNA-MEG3 was downregulated in skin samples from psoriatic patients compared to healthy skin samples [190]. Jia and colleagues proposed that lncRNA-MEG3 has a direct binding site for miR-21 and thus regulates apoptosis and pro-liferation in psoriatic keratinocytes [190].
Chang and colleagues reported significant upregulation of lncRNA-MEG3 in PBMCs in patients with T2DM and vascular complications compared to patients with T2DM without vascular complications and controls [193].
In patients with RA, levels of lncRNA-MEG3 were elevated in plasma, PBMCs and synovial fluid compared to healthy controls [194]. However, Wahba and colleagues reported a down-regulation of lncRNA-MEG3 in the serum of patients with RA compared to healthy controls. They also reported that the rs941576 polymorphism of lncRNA-MEG3 is associated with increased severity of RA in Egyptian patients [195].
6.3. lncRNA-UCA1
LncRNA-UCA1 downregulates NF-κB [196]. The role of lncRNA-UCA1 in psoriasis is not clearly understood. Studies have reported both increased and decreased expression of lncRNA-UCA1 in psoriasis [196,197]. Tian and colleagues reported that lncRNA-UCA1 is upregulated in vascular smooth muscle cells (VSMCs) treated with oxidized LDL and directly affects VSMC proliferation in atherosclerosis by modulating miR-26a expression [198].
In T2DM patients, lncRNA-UCA1 was downregulated in serum, serum exosomes and VSMCs compared to healthy controls [199]. It has been reported to promote VSMC proliferation via miR-582-5p under hyperglycemic conditions.
LncRNA-UCA1 was underexpressed in fibroblast-like synoviocytes from patients with RA compared to healthy controls [200].
6.4. LncRNA-XIST
The LncRNA-X-inactive specific transcript (lncRNA-XIST) is important for the inactivation of the X chromosome in female mammals in the placenta [201]. It is also involved in inflammation and carcinogenesis in various types of cancer. Higher levels of lncRNA-XIST have been associated with poorer prognosis in solid tumors [202,203].
Wang and colleagues reported higher lncRNA-XIST levels in patients with psoria-sis compared to healthy controls and a positive correlation between lncRNA-XIST levels and the PASI score, TNF-α, IL-17 and IL-22 levels [203]. To date, only the role of lncRNA-XIST in the transformation of atherosclerotic VSMCs is known. It promotes their migration and proliferation and reduces apoptosis [204,205,206].
Patients with T2DM have been reported to have decreased expression of lncRNA-XIST compared to healthy controls [207,208]. Interestingly, lncRNA-XIST was significantly upregulated in patients with CAD who also had T2DM compared to patients with CAD but without T2DM [209].
Liu and colleagues reported increased expression of lncRNA-XIST in synovial tissues of patients with RA compared to healthy controls [210].
We summarize the main findings of the studies that have investigated the role of lncRNAs in atherosclerosis, psoriasis, T2DM and RA in Table 2. This area of research is underdeveloped, but we expect more studies to be published in the future. The main limitation of the studies examined is the small sample sizes. LncRNA-MEG3 is upregulated in T2DM but downregulated in atherosclerosis and psoriasis (the two studies examining lncRNA-MEG3 in RA reported conflicting results). This could be due to the fact that hyperglycemia upregulates lncRNA-MEG3, however, experimental studies are needed to evaluate this hypothesis.
7. THE ROLE OF EXTRACELLULAR VESICLES IN ATHEROSCLEROSIS, PSORIASIS, T2DM, AND RA
Extracellular vesicles (EV) are a method of intercellular signaling used by almost all cells in the human body. Extracellular vesicles consist of a bilipid layer membrane that encapsulates a cargo of proteins, lipids, mRNA and microRNA [211]. Their classification is still controversial, but they can be divided into three categories based on their size: small or exosomes (< 100nm in diameter), medium or microvesicles (MV) (100–1000nm in diameter) and large or apoptotic bodies (> 1000nm in diameter) [212]. The function of EVs is highly dependent on their surface expression as well as their content [213].
Microvesicles not only act as biomarkers for disease, but also contribute to inflammation and promote the pathogenesis of atherosclerosis [214]. In vitro studies have shown that MVs produced under pathological conditions promote endothelial dysfunction [215,216]. Patients with psoriasis have higher concentrations of circulating EVs of endothelial origin of all sizes and higher concentrations of platelet-derived MVs less than 500 nm in size in their blood compared to healthy controls [217]. Zhang and colleagues reported increased plasma concentrations of platelet-derived MVs in patients with T2DM compared to obese and healthy subjects [218].
Increased levels of platelet-derived CD41+ MVs were found in patients with peripheral artery disease compared to healthy individuals [219]. In a hospital-based cross-sectional study conducted in 40 psoriasis patients and 12 healthy participants, a significant correlation was found between platelet-derived MVs and IL-12 and IL-17. CD41+ MVs were also significantly more common in psoriasis patients [220]. Tamagawa-Mineoka and colleagues discovered that the amount of platelet-derived MVs strongly correlated with the severity of psoriasis, however, their study was only conducted in 21 psoriasis patients and 22 healthy controls [221]. Ho and colleagues found increased levels of CD41+ and CD31+ platelet MVs compared to healthy controls [222]. Platelets with such surface markers have been associated with unstable atherosclerotic plaques [212].
Endothelial and platelet MV levels are decreased in psoriasis patients treated with anti-TNF-α medications, which may reduce the risk of cardiovascular complications [223,224].
Tan and colleagues reported increased levels of platelet MVs in patients with T2DM and clinically apparent atherosclerosis compared to healthy controls and T2DM patients without clinically apparent atherosclerosis [225].
MVs derived from the platelet-poor plasma of patients with RA promoted the production of IL-6 and IL-8 and the expression of CD54+ in endothelial cells as well as the adhesion of monocytes to the endothelium [226]. Michael and colleagues reported significantly higher levels of platelet- and leukocyte-derived MVs in patients with RA compared to patients with osteoarthritis and healthy controls. They found no correlation between MV levels and disease activity [227].
The concentrations of endothelial cell-derived EVs are elevated in both psoriasis and atherosclerosis and correlate with disease severity [213,228]. EVs produced by endothelial cells can induce the expression of adhesion molecules and selectins involved in leukocyte diapedesis [229]. Endothelial apoptotic bodies promote atherosclerosis via their miR-126 cargo; however, they may also have an anti-inflammatory and cytoprotective function [212]. MiR-126 may inhibit inflammation by mediating the production of VCAM-1 and thus reducing inflammation.
The exact role of miR-126 in psoriasis is still debatable. Murzina and colleagues discovered that levels of miR-126 correlates with disease severity and response to treatment in children with psoriasis [230]. Feng and colleagues also discovered a positive correlation between miR-126 and the severity of psoriasis and inflammation [231]. However, Pelosi and her research team and Duan and colleagues reported a negative correlation between miR-126 levels and the risk of developing psoriasis and the severity of the disease [232,233].
Levels of CD105+ endothelial EVs are significantly increased in psoriasis patients compared to healthy controls [234]. Chironi and colleagues reported a correlation between CD105+ EVs and CIMT [235]. Kandiyil and colleagues discovered an association between CD105+ EVs and stroke severity and clinical outcome [236]. Several studies have also reported increased expression of CD105+ in unstable atherosclerotic plaques [237,238,239].
Marei and colleagues reported significantly increased levels of CD42- CD31+ endothelial EVs in patients with T2DM and ACS compared to healthy controls [240]. Rodríguez-Carrio and colleagues reported increased levels of total EVs in patients with RA compared to healthy controls and a correlation with cardiovascular risk factors [241].
Smooth muscle cells, endothelial cells and macrophages are all affected by senescence in the process of atherosclerosis. Even though these cells are in the last phase of their life cycle, they still secrete EVs as well as pro-inflammatory cytokines. EVs carrying miR-21 induce senescence in endothelial cells, implying that they may also be involved in the progression of atherosclerosis [212]. This is possibly another overlap between the pathological pathways of psoriasis and atherosclerosis, as there is evidence that miR-21 is upregulated in psoriasis [46]. In atherosclerotic conditions, platelet-derived EVs stimulate the activity of IL-1, IL-6 and IL-8, all of which are important mediators in psoriasis [242,243]. EVs secreted by macrophages, neutrophils, mesenchymal stem cells, keratinocytes and adipocytes have also been associated with inflammation [230].
Figure 1 schematically represents the role of extracellular vesicles in the pathological processes involved in atherosclerosis and psoriasis.
8. Conclusions
It has been well established that patients who suffer from inflammatory diseases, such as T2DM, RA, and psoriasis have an increased risk of developing atherosclerosis. Systemic chronic inflammation induces the formation of focal atherosclerotic plaques at predilection sites (i.e. bifurcations of the arteries). Inflammatory cytokines regulate miR and lncRNA expression and function through complex feedback loops. Targeting inflammatory cytokines to treat atherosclerotic disease has proven effective, however, high costs of treatment and adverse side effects are the main issues. Specific miRNA therapeutics are already in different stages of development for various diseases (hepatitis C, various cancers, Alport syndrome) [244]. As the technology develops and becomes more accessible, we can expect miRNA-based medication development for treatment of atherosclerosis and its complications. In order for them to be established, they would have to be safe, more efficient and cost-effective than already established treatments, so they will more likely be developed for acute complications (myocardial infarction and stroke) first.
In this paper we reviewed the studies that investigated the associations between selected miRs and lncRNAs in atherosclerosis, psoriasis, T2DM, and RA. The interest in researching microRNAs, long non-coding RNAs and extracellular vesicles has increased recently because of the advancements in technology and the potential for novel treatment options. Information about the molecular profiles of EVs and their specific function in pathological pathways is still sparse. Advancements in the isolation of EVs and the subsequent analysis are needed in order to move this field one step closer to clinical implementation. Further research with larger number of participants is needed since the majority of the studies reviewed had a relatively small sample size.
Funding
“This research received no external funding”.
Conflicts of Interest
“The authors declare no conflict of interest.”
References
- Libby, P. Inflammation during the life cycle of the atherosclerotic plaque. Cardiovasc Res. 2021;117(13):2525-2536. [CrossRef]
- Libby, P. The changing landscape of atherosclerosis. Nature. 2021;592(7855):524-533. [CrossRef]
- Henein MY, Vancheri S, Longo G, Vancheri F. The Role of Inflammation in Cardiovascular Disease. Int J Mol Sci. 2022;23(21):12906. Published 2022 Oct 26. [CrossRef]
- Lin YJ, Anzaghe M, Schülke S. Update on the Pathomechanism, Diagnosis, and Treatment Options for Rheumatoid Arthritis. Cells. 2020;9(4):880. Published 2020 Apr 3. [CrossRef]
- Luc K, Schramm-Luc A, Guzik TJ, Mikolajczyk TP. Oxidative stress and inflammatory markers in prediabetes and diabetes. J Physiol Pharmacol. 2019;70(6):10.26402/jpp.2019.6.01. [CrossRef]
- Tibaut M, Mankoč Ramuš S, Petrovič D. The C allele of the reactive oxygen species modulator 1 (ROMO1) polymorphism rs6060566 is a biomarker predicting coronary artery stenosis in Slovenian subjects with type 2 diabetes mellitus. BMC Med Genomics. 2020;13(1):184. Published 2020 Dec 10. [CrossRef]
- Borroni RG, Costanzo A. HLA-C*06 and psoriasis: susceptibility, phenotype, course and response to treatment. Br J Dermatol. 2018;178(4):825. [CrossRef]
- Chen L, Tsai TF. HLA-Cw6 and psoriasis. Br J Dermatol. 2018;178(4):854-862. [CrossRef]
- Huang YW, Tsai TF. HLA-Cw1 and Psoriasis. Am J Clin Dermatol. 2021;22(3):339-347. [CrossRef]
- Caiazzo G, Fabbrocini G, Di Caprio R, et al. Psoriasis, Cardiovascular Events, and Biologics: Lights and Shadows. Front Immunol. 2018;9:1668. Published 2018 Aug 13. [CrossRef]
- Furue M, Tsuji G, Chiba T, Kadono T. Cardiovascular and Metabolic Diseases Comorbid with Psoriasis: Beyond the Skin. Intern Med. 2017;56(13):1613-1619. [CrossRef]
- Choudhary S, Patel R, Pradhan D, et al. Psoriasis and cardiovascular disorders: association or epiphenomenon? Meta-analysis of observational studies. 3 Biotech. 2020;10(3):104. [CrossRef]
- Reich, K. The concept of psoriasis as a systemic inflammation: implications for disease management. J Eur Acad Dermatol Venereol. 2012;26 Suppl 2:3-11. [CrossRef]
- Ahmad E, Lim S, Lamptey R, Webb DR, Davies MJ. Type 2 diabetes. Lancet. 2022;400(10365):1803-1820. [CrossRef]
- Smolen JS, Aletaha D, McInnes IB. Rheumatoid arthritis [published correction appears in Lancet. 2016 Oct 22;388(10055):1984]. Lancet. 2016;388(10055):2023-2038. [CrossRef]
- Finckh A, Gilbert B, Hodkinson B, et al. Global epidemiology of rheumatoid arthritis. Nat Rev Rheumatol. 2022;18(10):591-602. [CrossRef]
- Praveenkumar U, Ganguly S, Ray L, Nanda SK, Kuruvila S. Prevalence of Metabolic Syndrome in Psoriasis Patients and its Relation to Disease Duration: A Hospital Based Case-Control Study. J Clin Diagn Res. 2016;10(2):WC01-WC5. [CrossRef]
- Shibata S, Tada Y, Hau CS, et al. Adiponectin regulates psoriasiform skin inflammation by suppressing IL-17 production from γδ-T cells. Nat Commun. 2015;6:7687. Published 2015 Jul 15. [CrossRef]
- Gonzalez-Cantero A, Gonzalez-Cantero J, Sanchez-Moya AI, et al. Subclinical atherosclerosis in psoriasis. Usefulness of femoral artery ultrasound for the diagnosis, and analysis of its relationship with insulin resistance. PLoS One. 2019;14(2):e0211808. Published 2019 Feb 8. [CrossRef]
- Ruiz de Morales JMG, Puig L, Daudén E, et al. Critical role of interleukin (IL)-17 in inflammatory and immune disorders: An updated review of the evidence focusing in controversies. Autoimmun Rev. 2020;19(1):102429. [CrossRef]
- Marder W, Khalatbari S, Myles JD, et al. Interleukin 17 as a novel predictor of vascular function in rheumatoid arthritis. Ann Rheum Dis. 2011;70(9):1550-1555. [CrossRef]
- Kaur R, Kaur M, Singh J. Endothelial dysfunction and platelet hyperactivity in type 2 diabetes mellitus: molecular insights and therapeutic strategies. Cardiovasc Diabetol. 2018;17(1):121. Published 2018 Aug 31. [CrossRef]
- Kothiwala SK, Khanna N, Tandon N, et al. Prevalence of metabolic syndrome and cardiovascular changes in patients with chronic plaque psoriasis and their correlation with disease severity: A hospital-based cross-sectional study. Indian J Dermatol Venereol Leprol. 2016;82(5):510-518. [CrossRef]
- Bordy R, Totoson P, Prati C, Marie C, Wendling D, Demougeot C. Microvascular endothelial dysfunction in rheumatoid arthritis. Nat Rev Rheumatol. 2018;14(7):404-420. [CrossRef]
- Ikeda N, Kogame N, Iijima R, Nakamura M, Sugi K. Carotid artery intima-media thickness and plaque score can predict the SYNTAX score. Eur Heart J. 2012;33(1):113-119. [CrossRef]
- Fang N, Jiang M, Fan Y. Association Between Psoriasis and Subclinical Atherosclerosis: A Meta-Analysis. Medicine (Baltimore). 2016;95(20):e3576. [CrossRef]
- Emerging Risk Factors Collaboration, Sarwar N, Gao P, et al. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: a collaborative meta-analysis of 102 prospective studies [published correction appears in Lancet. 2010 Sep 18;376(9745):958. Hillage, H L [corrected to Hillege, H L]]. Lancet. 2010;375(9733):2215-2222. [CrossRef]
- Brohall G, Odén A, Fagerberg B. Carotid artery intima-media thickness in patients with Type 2 diabetes mellitus and impaired glucose tolerance: a systematic review. Diabet Med. 2006;23(6):609-616. [CrossRef]
- Wang P, Guan SY, Xu SZ, et al. Increased carotid intima-media thickness in rheumatoid arthritis: an update meta-analysis. Clin Rheumatol. 2016;35(2):315-323. [CrossRef]
- Que X, Hung MY, Yeang C, et al. Oxidized phospholipids are proinflammatory and proatherogenic in hypercholesterolaemic mice [published correction appears in Nature. 2018 Sep;561(7724):E43]. Nature. 2018;558(7709):301-306. [CrossRef]
- Kim TW, Febbraio M, Robinet P, et al. The critical role of IL-1 receptor-associated kinase 4-mediated NF-κB activation in modified low-density lipoprotein-induced inflammatory gene expression and atherosclerosis. J Immunol. 2011;186(5):2871-2880. [CrossRef]
- Mehta NN, Li R, Krishnamoorthy P, et al. Abnormal lipoprotein particles and cholesterol efflux capacity in patients with psoriasis. Atherosclerosis. 2012;224(1):218-221. [CrossRef]
- Bahiru E, Hsiao R, Phillipson D, Watson KE. Mechanisms and Treatment of Dyslipidemia in Diabetes. Curr Cardiol Rep. 2021;23(4):26. Published 2021 Mar 2. [CrossRef]
- Venetsanopoulou AI, Pelechas E, Voulgari PV, Drosos AA. The lipid paradox in rheumatoid arthritis: the dark horse of the augmented cardiovascular risk. Rheumatol Int. 2020;40(8):1181-1191. [CrossRef]
- Chiu JJ, Chien S. Effects of disturbed flow on vascular endothelium: pathophysiological basis and clinical perspectives. Physiol Rev. 2011;91(1):327-387. [CrossRef]
- Popescu D, Rezus E, Badescu MC, et al. Cardiovascular Risk Assessment in Rheumatoid Arthritis: Accelerated Atherosclerosis, New Biomarkers, and the Effects of Biological Therapy. Life (Basel). 2023;13(2):319. Published 2023 Jan 23. [CrossRef]
- Su W, Zhao Y, Wei Y, Zhang X, Ji J, Yang S. Exploring the Pathogenesis of Psoriasis Complicated With Atherosclerosis via Microarray Data Analysis. Front Immunol. 2021;12:667690. Published 2021 May 27. [CrossRef]
- Zhang M, Cai ZR, Zhang B, et al. Functional polymorphisms in interleukin-23 receptor and susceptibility to coronary artery disease. DNA Cell Biol. 2014;33(12):891-897. [CrossRef]
- Eirís N, González-Lara L, Santos-Juanes J, Queiro R, Coto E, Coto-Segura P. Genetic variation at IL12B, IL23R and IL23A is associated with psoriasis severity, psoriatic arthritis and type 2 diabetes mellitus. J Dermatol Sci. 2014;75(3):167-172. [CrossRef]
- Vázquez-Vázquez C, Posadas-Sánchez R, Pérez-Hernández N, et al. The rs2066808 Polymorphism Located Near the IL-23A Gene Is Associated with Premature Coronary Artery Disease in Mexican Population (GEA Study). DNA Cell Biol. 2019;38(8):880-886. [CrossRef]
- Eder L, Abji F, Rosen CF, Chandran V, Cook RJ, Gladman DD. The Association of HLA-class I Genes and the Extent of Atherosclerotic Plaques in Patients with Psoriatic Disease. J Rheumatol. 2016;43(10):1844-1851. [CrossRef]
- Harden JL, Lewis SM, Pierson KC, et al. CARD14 expression in dermal endothelial cells in psoriasis. PLoS One. 2014;9(11):e111255. Published 2014 Nov 4. [CrossRef]
- Fu Y, Xu L, Zhang H, et al. Identification and Validation of Immune-Related Genes Diagnostic for Progression of Atherosclerosis and Diabetes. J Inflamm Res. 2023;16:505-521. Published 2023 Feb 10. [CrossRef]
- Xiao L, Yang Z, Lin S. Identification of hub genes and transcription factors in patients with rheumatoid arthritis complicated with atherosclerosis. Sci Rep. 2022;12(1):4677. Published 2022 Mar 18. [CrossRef]
- Tibaut M, Caprnda M, Kubatka P, et al. Markers of Atherosclerosis: Part 2 - Genetic and Imaging Markers. Heart Lung Circ. 2019;28(5):678-689. [CrossRef]
- Salvi V, Gianello V, Tiberio L, Sozzani S, Bosisio D. Cytokine Targeting by miRNAs in Autoimmune Diseases. Front Immunol. 2019;10:15. Published 2019 Jan 29. [CrossRef]
- Hermann H, Runnel T, Aab A, et al. miR-146b Probably Assists miRNA-146a in the Suppression of Keratinocyte Proliferation and Inflammatory Responses in Psoriasis. J Invest Dermatol. 2017;137(9):1945-1954. [CrossRef]
- Huang P, He XY, Xu M. The Role of miRNA-146a and Proinflammatory Cytokines in Carotid Atherosclerosis. Biomed Res Int. 2020;2020:6657734. Published 2020 Dec 9. [CrossRef]
- Xue L, Luo S, Ding H, et al. Upregulation of miR-146a-5p is associated with increased proliferation and migration of vascular smooth muscle cells in aortic dissection. J Clin Lab Anal. 2019;33(4):e22843. [CrossRef]
- Dong S, Xiong W, Yuan J, Li J, Liu J, Xu X. MiRNA-146a regulates the maturation and differentiation of vascular smooth muscle cells by targeting NF-κB expression. Mol Med Rep. 2013;8(2):407-412. [CrossRef]
- Vaher H, Kivihall A, Runnel T, et al. SERPINB2 and miR-146a/b are coordinately regulated and act in the suppression of psoriasis-associated inflammatory responses in keratinocytes. Exp Dermatol. 2020;29(1):51-60. [CrossRef]
- Leal B, Carvalho C, Ferreira AM, et al. Serum Levels of miR-146a in Patients with Psoriasis. Mol Diagn Ther. 2021;25(4):475-485. [CrossRef]
- Ele-Refaei AM, El-Esawy FM. Effect of Narrow-Band Ultraviolet B Phototherapy and Methotrexate on MicroRNA (146a) Levels in Blood of Psoriatic Patients [published correction appears in Dermatol Res Pract. 2016;2016:7168587]. Dermatol Res Pract. 2015;2015:145769. [CrossRef]
- Raitoharju E, Lyytikäinen LP, Levula M, et al. miR-21, miR-210, miR-34a, and miR-146a/b are up-regulated in human atherosclerotic plaques in the Tampere Vascular Study. Atherosclerosis. 2011;219(1):211-217. [CrossRef]
- Zhelankin AV, Stonogina DA, Vasiliev SV, et al. Circulating Extracellular miRNA Analysis in Patients with Stable CAD and Acute Coronary Syndromes. Biomolecules. 2021;11(7):962. Published 2021 Jun 29. [CrossRef]
- Guo W, Li XN, Li J, et al. Increased plasma miR-146a levels are associated with subclinical atherosclerosis in newly diagnosed type 2 diabetes mellitus. J Diabetes Complications. 2020;34(12):107725. [CrossRef]
- Gao W, Li R, Yu J, et al. LncRNA SCIRT is downregulated in atherosclerosis and suppresses the proliferation of human aortic smooth muscle cells (HAOSMCs) by sponging miR-146a in cytoplasm. J Cardiothorac Surg. 2021;16(1):324. Published 2021 Nov 6. [CrossRef]
- Gong HB, Zhang SL, Wu XJ, Pu XM, Kang XJ. Association of rs2910164 polymorphism in MiR-146a gene with psoriasis susceptibility: A meta-analysis. Medicine (Baltimore). 2019;98(6):e14401. [CrossRef]
- Bao Q, Li R, Wang C, et al. Association between microRNA-146a rs2910164 polymorphism and coronary heart disease: An updated meta-analysis. Medicine (Baltimore). 2022;101(46):e31860. [CrossRef]
- Bao MH, Xiao Y, Zhang QS, et al. Meta-Analysis of miR-146a Polymorphisms Association with Coronary Artery Diseases and Ischemic Stroke. Int J Mol Sci. 2015;16(7):14305-14317. Published 2015 Jun 24. [CrossRef]
- Alipoor B, Ghaedi H, Meshkani R, et al. Association of MiR-146a Expression and Type 2 Diabetes Mellitus: A Meta-Analysis. Int J Mol Cell Med. 2017;6(3):156-163. [CrossRef]
- Zhu H, Leung SW. MicroRNA biomarkers of type 2 diabetes: evidence synthesis from meta-analyses and pathway modelling. Diabetologia. 2023;66(2):288-299. [CrossRef]
- Cheng L, Zhou M, Zhang D, Chen B. Association of miR-146a polymorphism rs2910164 and type 2 diabetes risk: a meta-analysis. J Int Med Res. 2020;48(8):300060520931313. [CrossRef]
- Shen J, Zhang M, Sun M, Tang K, Zhou B. The relationship of miR-146a gene polymorphism with carotid atherosclerosis in Chinese patients with type 2 diabetes mellitus. Thromb Res. 2015;136(6):1149-1155. [CrossRef]
- Bae SC, Lee YH. MiR-146a levels in rheumatoid arthritis and their correlation with disease activity: a meta-analysis. Int J Rheum Dis. 2018;21(7):1335-1342. [CrossRef]
- Liu F, Liang Y, Zhao Y, Chen L, Wang X, Zhang C. Meta-analysis of association of microRNAs genetic variants with susceptibility to rheumatoid arthritis and systemic lupus erythematosus. Medicine (Baltimore). 2021;100(17):e25689. [CrossRef]
- Tao J, Xia L, Cai Z, et al. Interaction Between microRNA and DNA Methylation in Atherosclerosis. DNA Cell Biol. 2021;40(1):101-115. [CrossRef]
- Ivan M, Harris AL, Martelli F, Kulshreshtha R. Hypoxia response and microRNAs: no longer two separate worlds. J Cell Mol Med. 2008;12(5A):1426-1431. [CrossRef]
- Signorelli SS, Volsi GL, Pitruzzella A, et al. Circulating miR-130a, miR-27b, and miR-210 in Patients With Peripheral Artery Disease and Their Potential Relationship With Oxidative Stress. Angiology. 2016;67(10):945-950. [CrossRef]
- Eken SM, Jin H, Chernogubova E, et al. MicroRNA-210 Enhances Fibrous Cap Stability in Advanced Atherosclerotic Lesions. Circ Res. 2017;120(4):633-644. [CrossRef]
- Wu R, Zeng J, Yuan J, et al. MicroRNA-210 overexpression promotes psoriasis-like inflammation by inducing Th1 and Th17 cell differentiation. J Clin Invest. 2018;128(6):2551-2568. [CrossRef]
- Zhao M, Wang LT, Liang GP, et al. Up-regulation of microRNA-210 induces immune dysfunction via targeting FOXP3 in CD4(+) T cells of psoriasis vulgaris. Clin Immunol. 2014;150(1):22-30. [CrossRef]
- Li X, Jia Z, Zhao X, Xu M, Chen M. Expression of miR-210 in the peripheral blood of patients with newly diagnosed type 2 diabetes mellitus and its effect on the number and function of endothelial progenitor cells. Microvasc Res. 2020;131:104032. [CrossRef]
- Chen X, Tian F, Sun Z, Zeng G, Tang P. Elevation of Circulating miR-210 Participates in the Occurrence and Development of Type 2 Diabetes Mellitus and Its Complications. J Diabetes Res. 2022;2022:9611509. Published 2022 Nov 23. [CrossRef]
- Amr KS, Abdelmawgoud H, Ali ZY, Shehata S, Raslan HM. Potential value of circulating microRNA-126 and microRNA-210 as biomarkers for type 2 diabetes with coronary artery disease. Br J Biomed Sci. 2018;75(2):82-87. [CrossRef]
- Zhou Z, Collado A, Sun C, et al. Downregulation of Erythrocyte miR-210 Induces Endothelial Dysfunction in Type 2 Diabetes. Diabetes. 2022;71(2):285-297. [CrossRef]
- Zhang L, Zhang J, Qin Z, et al. Diagnostic and Predictive Values of Circulating Extracellular Vesicle-Carried microRNAs in Ischemic Heart Disease Patients With Type 2 Diabetes Mellitus. Front Cardiovasc Med. 2022;9:813310. Published 2022 Feb 28. [CrossRef]
- Abdul-Maksoud RS, Sediq AM, Kattaia A, et al. Serum miR-210 and miR-155 expression levels as novel biomarkers for rheumatoid arthritis diagnosis. Br J Biomed Sci. 2017;74(4):209-213. [CrossRef]
- Huang Q, Chen SS, Li J, et al. miR-210 expression in PBMCs from patients with systemic lupus erythematosus and rheumatoid arthritis. Ir J Med Sci. 2018;187(1):243-249. [CrossRef]
- Kumar S, Kim CW, Simmons RD, Jo H. Role of flow-sensitive microRNAs in endothelial dysfunction and atherosclerosis: mechanosensitive athero-miRs. Arterioscler Thromb Vasc Biol. 2014;34(10):2206-2216. [CrossRef]
- Vengrenyuk Y, Nishi H, Long X, et al. Cholesterol loading reprograms the microRNA-143/145-myocardin axis to convert aortic smooth muscle cells to a dysfunctional macrophage-like phenotype. Arterioscler Thromb Vasc Biol. 2015;35(3):535-546. [CrossRef]
- Zhou C, Yang Y, Hu L, et al. Effects of miR-143 regulation on cardiomyocytes apoptosis in doxorubicin cardiotoxicity based on integrated bioinformatics analysis. Toxicol In Vitro. 2023;93:105662. [CrossRef]
- Meng L, Yu X, Han H, et al. Circulating miR-143 and miR-145 as promising biomarkers for evaluating severity of coronary artery stenosis in patients with acute coronary syndrome. Clin Biochem. 2023;111:32-40. [CrossRef]
- Løvendorf MB, Zibert JR, Gyldenløve M, Røpke MA, Skov L. MicroRNA-223 and miR-143 are important systemic biomarkers for disease activity in psoriasis. J Dermatol Sci. 2014;75(2):133-139. [CrossRef]
- Zheng YZ, Chen CF, Jia LY, Yu TG, Sun J, Wang XY. Correlation between microRNA-143 in peripheral blood mononuclear cells and disease severity in patients with psoriasis vulgaris. Oncotarget. 2017;8(31):51288-51295. Published 2017 Apr 20. [CrossRef]
- Yue J, Lau TCK, Griffith JF, et al. Circulating miR-99b-5p as a novel predictor of erosion progression on high-resolution peripheral quantitative computed tomography in early rheumatoid arthritis: A prospective cohort study. Int J Rheum Dis. 2019;22(9):1724-1733. [CrossRef]
- Shan Z, Qin S, Li W, et al. An Endocrine Genetic Signal Between Blood Cells and Vascular Smooth Muscle Cells: Role of MicroRNA-223 in Smooth Muscle Function and Atherogenesis. J Am Coll Cardiol. 2015;65(23):2526-2537. [CrossRef]
- Vickers KC, Landstreet SR, Levin MG, et al. MicroRNA-223 coordinates cholesterol homeostasis. Proc Natl Acad Sci U S A. 2014;111(40):14518-14523. [CrossRef]
- Ann SJ, Bang H, Lee CJ, et al. LncRNA HSPA7 in human atherosclerotic plaques sponges miR-223 and promotes the proinflammatory vascular smooth muscle cell transition. Exp Mol Med. 2021;53(12):1842-1849. [CrossRef]
- Parrizas M, Mundet X, Castaño C, et al. miR-10b and miR-223-3p in serum microvesicles signal progression from prediabetes to type 2 diabetes. J Endocrinol Invest. 2020;43(4):451-459. [CrossRef]
- Singh S, de Ronde MWJ, Kok MGM, et al. MiR-223-3p and miR-122-5p as circulating biomarkers for plaque instability. Open Heart. 2020;7(1):e001223. [CrossRef]
- Guo JF, Zhang Y, Zheng QX, Zhang Y, Zhou HH, Cui LM. Association between elevated plasma microRNA-223 content and severity of coronary heart disease. Scand J Clin Lab Invest. 2018;78(5):373-378. [CrossRef]
- Zhu L, Wang Y, Qiao F. microRNA-223 and microRNA-126 are clinical indicators for predicting the plaque stability in carotid atherosclerosis patients [published online ahead of print, 2022 Oct 3]. J Hum Hypertens. 2022;10.1038/s41371-022-00760-3. [CrossRef]
- Wang R, Wang FF, Cao HW, Yang JY. MiR-223 regulates proliferation and apoptosis of IL-22-stimulated HaCat human keratinocyte cell lines via the PTEN/Akt pathway. Life Sci. 2019;230:28-34. [CrossRef]
- Pivarcsi A, Meisgen F, Xu N, Ståhle M, Sonkoly E. Changes in the level of serum microRNAs in patients with psoriasis after antitumour necrosis factor-α therapy. Br J Dermatol. 2013;169(3):563-570. [CrossRef]
- Alatas ET, Kara M, Dogan G, Akın Belli A. Blood microRNA expressions in patients with mild to moderate psoriasis and the relationship between microRNAs and psoriasis activity. An Bras Dermatol. 2020;95(6):702-707. [CrossRef]
- Ormseth MJ, Solus JF, Vickers KC, Oeser AM, Raggi P, Stein CM. Utility of Select Plasma MicroRNA for Disease and Cardiovascular Risk Assessment in Patients with Rheumatoid Arthritis. J Rheumatol. 2015;42(10):1746-1751. [CrossRef]
- Fichtlscherer S, De Rosa S, Fox H, et al. Circulating microRNAs in patients with coronary artery disease. Circ Res. 2010;107(5):677-684. [CrossRef]
- Li X, Kong D, Chen H, et al. miR-155 acts as an anti-inflammatory factor in atherosclerosis-associated foam cell formation by repressing calcium-regulated heat stable protein 1. Sci Rep. 2016;6:21789. Published 2016 Feb 22. [CrossRef]
- Wang R, Dong LD, Meng XB, Shi Q, Sun WY. Unique MicroRNA signatures associated with early coronary atherosclerotic plaques. Biochem Biophys Res Commun. 2015;464(2):574-579. [CrossRef]
- García-Rodríguez S, Arias-Santiago S, Blasco-Morente G, et al. Increased expression of microRNA-155 in peripheral blood mononuclear cells from psoriasis patients is related to disease activity. J Eur Acad Dermatol Venereol. 2017;31(2):312-322. [CrossRef]
- Xu L, Leng H, Shi X, Ji J, Fu J, Leng H. MiR-155 promotes cell proliferation and inhibits apoptosis by PTEN signaling pathway in the psoriasis. Biomed Pharmacother. 2017;90:524-530. [CrossRef]
- Elmesmari A, Fraser AR, Wood C, et al. MicroRNA-155 regulates monocyte chemokine and chemokine receptor expression in Rheumatoid Arthritis. Rheumatology (Oxford). 2016;55(11):2056-2065. [CrossRef]
- Kadkhoda S, Ghafouri-Fard S. Function of miRNA-145-5p in the pathogenesis of human disorders. Pathol Res Pract. 2022;231:153780. [CrossRef]
- Chin DD, Poon C, Wang J, et al. miR-145 micelles mitigate atherosclerosis by modulating vascular smooth muscle cell phenotype. Biomaterials. 2021;273:120810. [CrossRef]
- Wang Y, Cao Y. miR-145-5p inhibits psoriasis progression by regulating the Wnt/β-catenin pathway. Am J Transl Res. 2021;13(9):10439-10448. Published 2021 Sep 15.
- Yuan M, Zhang L, You F, et al. MiR-145-5p regulates hypoxia-induced inflammatory response and apoptosis in cardiomyocytes by targeting CD40. Mol Cell Biochem. 2017;431(1-2):123-131. [CrossRef]
- Minin EOZ, Paim LR, Lopes ECP, et al. Association of Circulating miR-145-5p and miR-let7c and Atherosclerotic Plaques in Hypertensive Patients. Biomolecules. 2021;11(12):1840. Published 2021 Dec 7. [CrossRef]
- Zhang X, Zai L, Tao Z, Wu D, Lin M, Wan J. miR-145-5p affects autophagy by targeting CaMKIIδ in atherosclerosis. Int J Cardiol. 2022;360:68-75. [CrossRef]
- Lv Y, Yi Y, Jia S, Peng X, Yang H, Guo R. The miR-145 rs353291 C allele increases susceptibility to atherosclerosis. Front Biosci (Landmark Ed). 2020;25(3):577-592. Published 2020 Jan 1. [CrossRef]
- Hall IF, Climent M, Viviani Anselmi C, et al. rs41291957 controls miR-143 and miR-145 expression and impacts coronary artery disease risk. EMBO Mol Med. 2021;13(10):e14060. [CrossRef]
- Hong BK, You S, Yoo SA, et al. MicroRNA-143 and -145 modulate the phenotype of synovial fibroblasts in rheumatoid arthritis. Exp Mol Med. 2017;49(8):e363. Published 2017 Aug 4. [CrossRef]
- Wang X, Tang K, Wang Y, et al. Elevated microRNA 145 5p increases matrix metalloproteinase 9 by activating the nuclear factor κB pathway in rheumatoid arthritis. Mol Med Rep. 2019;20(3):2703-2711. [CrossRef]
- Magenta A, D'Agostino M, Sileno S, et al. The Oxidative Stress-Induced miR-200c Is Upregulated in Psoriasis and Correlates with Disease Severity and Determinants of Cardiovascular Risk. Oxid Med Cell Longev. 2019;2019:8061901. Published 2019 Dec 19. [CrossRef]
- Magenta A, Sileno S, D'Agostino M, et al. Atherosclerotic plaque instability in carotid arteries: miR-200c as a promising biomarker. Clin Sci (Lond). 2018;132(22):2423-2436. Published 2018 Nov 21. [CrossRef]
- Wang XY, Chen XY, Li J, Zhang HY, Liu J, Sun LD. MiR-200a expression in CD4+ T cells correlates with the expression of Th17/Treg cells and relevant cytokines in psoriasis vulgaris: A case control study. Biomed Pharmacother. 2017;93:1158-1164. [CrossRef]
- Kujawa M, O'Meara M, Li H, et al. MicroRNA-466 and microRNA-200 increase endothelial permeability in hyperglycemia by targeting Claudin-5. Mol Ther Nucleic Acids. 2022;29:259-271. Published 2022 Jul 6. [CrossRef]
- Ofori JK, Karagiannopoulos A, Nagao M, et al. Human Islet MicroRNA-200c Is Elevated in Type 2 Diabetes and Targets the Transcription Factor ETV5 to Reduce Insulin Secretion. Diabetes. 2022;71(2):275-284. [CrossRef]
- Lo WY, Yang WK, Peng CT, Pai WY, Wang HJ. MicroRNA-200a/200b Modulate High Glucose-Induced Endothelial Inflammation by Targeting O-linked N-Acetylglucosamine Transferase Expression [published correction appears in Front Physiol. 2018 Jun 19;9:786]. Front Physiol. 2018;9:355. Published 2018 Apr 18. [CrossRef]
- Balzano F, Deiana M, Dei Giudici S, et al. MicroRNA Expression Analysis of Centenarians and Rheumatoid Arthritis Patients Reveals a Common Expression Pattern. Int J Med Sci. 2017;14(7):622-628. Published 2017 Jun 14. [CrossRef]
- Gangwar RS, Rajagopalan S, Natarajan R, Deiuliis JA. Noncoding RNAs in Cardiovascular Disease: Pathological Relevance and Emerging Role as Biomarkers and Therapeutics. Am J Hypertens. 2018;31(2):150-165. [CrossRef]
- Jin X, Chen D, Zheng RH, Zhang H, Chen YP, Xiang Z. miRNA-133a-UCP2 pathway regulates inflammatory bowel disease progress by influencing inflammation, oxidative stress and energy metabolism. World J Gastroenterol. 2017;23(1):76-86. [CrossRef]
- Torella D, Iaconetti C, Catalucci D, et al. MicroRNA-133 controls vascular smooth muscle cell phenotypic switch in vitro and vascular remodeling in vivo. Circ Res. 2011;109(8):880-893. [CrossRef]
- Liao XB, Zhang ZY, Yuan K, et al. MiR-133a modulates osteogenic differentiation of vascular smooth muscle cells. Endocrinology. 2013;154(9):3344-3352. [CrossRef]
- Navickas R, Gal D, Laucevičius A, Taparauskaitė A, Zdanytė M, Holvoet P. Identifying circulating microRNAs as biomarkers of cardiovascular disease: a systematic review. Cardiovasc Res. 2016;111(4):322-337. [CrossRef]
- Šatrauskienė A, Navickas R, Laucevičius A, et al. Mir-1, miR-122, miR-132, and miR-133 Are Related to Subclinical Aortic Atherosclerosis Associated with Metabolic Syndrome. Int J Environ Res Public Health. 2021;18(4):1483. Published 2021 Feb 4. [CrossRef]
- Wang F, Long G, Zhao C, et al. Plasma microRNA-133a is a new marker for both acute myocardial infarction and underlying coronary artery stenosis. J Transl Med. 2013;11:222. Published 2013 Sep 23. [CrossRef]
- Chicharro P, Rodríguez-Jiménez P, Llamas-Velasco M, et al. Expression of miR-135b in Psoriatic Skin and Its Association with Disease Improvement. Cells. 2020;9(7):1603. Published 2020 Jul 2. [CrossRef]
- de Gonzalo-Calvo D, van der Meer RW, Rijzewijk LJ, et al. Serum microRNA-1 and microRNA-133a levels reflect myocardial steatosis in uncomplicated type 2 diabetes. Sci Rep. 2017;7(1):47. Published 2017 Mar 3. [CrossRef]
- Ghasemi H, Karimi J, Khodadadi I, Tavilani H. Correlation between miR-103 and miR-133a Expression and the Circulating ANGPTL8 in Type 2 Diabetic Patients and Healthy Control Subjects. Clin Lab. 2019;65(11):10.7754/Clin.Lab.2019.190436. [CrossRef]
- Al-Muhtaresh HA, Salem AH, Al-Kafaji G. Upregulation of Circulating Cardiomyocyte-Enriched miR-1 and miR-133 Associate with the Risk of Coronary Artery Disease in Type 2 Diabetes Patients and Serve as Potential Biomarkers. J Cardiovasc Transl Res. 2019;12(4):347-357. [CrossRef]
- Zhang X, Lu J, Zhang Q, Luo Q, Liu B. CircRNA RSF1 regulated ox-LDL induced vascular endothelial cells proliferation, apoptosis and inflammation through modulating miR-135b-5p/HDAC1 axis in atherosclerosis. Biol Res. 2021;54(1):11. Published 2021 Mar 23. [CrossRef]
- Kadkhoda S, Eslami S, Mahmud Hussen B, Ghafouri-Fard S. A review on the importance of miRNA-135 in human diseases. Front Genet. 2022;13:973585. Published 2022 Sep 6. [CrossRef]
- Chicharro P, Rodríguez-Jiménez P, Llamas-Velasco M, et al. Expression of miR-135b in Psoriatic Skin and Its Association with Disease Improvement. Cells. 2020;9(7):1603. Published 2020 Jul 2. [CrossRef]
- Xu Z, Han Y, Liu J, et al. MiR-135b-5p and MiR-499a-3p Promote Cell Proliferation and Migration in Atherosclerosis by Directly Targeting MEF2C. Sci Rep. 2015;5:12276. Published 2015 Jul 17. [CrossRef]
- Li D, An Y. MiR-135a-5p inhibits vascular smooth muscle cells proliferation and migration by inactivating FOXO1 and JAK2 signaling pathway. Pathol Res Pract. 2021;224:153091. [CrossRef]
- Joyce CE, Zhou X, Xia J, et al. Deep sequencing of small RNAs from human skin reveals major alterations in the psoriasis miRNAome. Hum Mol Genet. 2011;20(20):4025-4040. [CrossRef]
- Sarookhani MR, Honardoost M, Foroughi F. Plasma miR-135a; a potential biomarker for diagnosis of new type 2 diabetes (T2DM). Bali Medical Journal. 2018; 7(2):296-301. [CrossRef]
- Monfared YK, Honardoost M, Sarookhani MR, Farzam SA. Circulating miR-135 May Serve as a Novel Co-biomarker of HbA1c in Type 2 Diabetes. Appl Biochem Biotechnol. 2020;191(2):623-630. [CrossRef]
- Monfared YK, Mirzaii-Dizgah MR, Khodabandehloo E, Sarookhani MR, Hashemipour S, Mirzaii-Dizgah I. Salivary microRNA-126 and 135a: a potentially non-invasive diagnostic biomarkers of type- 2 diabetes. J Diabetes Metab Disord. 2021;20(2):1631-1638. Published 2021 Oct 28. [CrossRef]
- Liu J, Xue J, Xu B, et al. miR-135a-5p mediated down-regulation of STAT6 inhibits proliferation and induces apoptosis of fibroblast-like synoviocytes in rheumatoid arthritis. Am J Transl Res. 2022;14(5):3092-3103. Published 2022 May 15.
- Xue Y, Wei Z, Ding H, et al. MicroRNA-19b/221/222 induces endothelial cell dysfunction via suppression of PGC-1α in the progression of atherosclerosis. Atherosclerosis. 2015;241(2):671-681. [CrossRef]
- Chistiakov DA, Sobenin IA, Orekhov AN, Bobryshev YV. Human miR-221/222 in Physiological and Atherosclerotic Vascular Remodeling. Biomed Res Int. 2015;2015:354517. [CrossRef]
- Minami Y, Satoh M, Maesawa C, et al. Effect of atorvastatin on microRNA 221 / 222 expression in endothelial progenitor cells obtained from patients with coronary artery disease. Eur J Clin Invest. 2009;39(5):359-367. [CrossRef]
- Zibert JR, Løvendorf MB, Litman T, Olsen J, Kaczkowski B, Skov L. MicroRNAs and potential target interactions in psoriasis. J Dermatol Sci. 2010;58(3):177-185. [CrossRef]
- Meng Z, Qiu J, Zhang H. MiR-221-3p as a Potential Biomarker for Patients with Psoriasis and Its Role in Inflammatory Responses in Keratinocytes. Skin Pharmacol Physiol. 2021;34(5):300-306. [CrossRef]
- Wade SM, McGarry T, Wade SC, Fearon U, Veale DJ. Serum MicroRNA Signature as a Diagnostic and Therapeutic Marker in Patients with Psoriatic Arthritis. J Rheumatol. 2020;47(12):1760-1767. [CrossRef]
- Abo ElAtta AS, Ali YBM, Bassyouni IH, Talaat RM. Upregulation of miR-221/222 expression in rheumatoid arthritis (RA) patients: correlation with disease activity. Clin Exp Med. 2019;19(1):47-53. [CrossRef]
- Cunningham CC, Wade S, Floudas A, et al. Serum miRNA Signature in Rheumatoid Arthritis and "At-Risk Individuals". Front Immunol. 2021;12:633201. Published 2021 Mar 3. [CrossRef]
- Cieśla M, Kolarz B, Majdan M, Darmochwał-Kolarz D. Plasma micro-RNA-22 is associated with disease activity in well-established rheumatoid arthritis. Clin Exp Rheumatol. 2022;40(5):945-951. [CrossRef]
- Wang H, He F, Liang B, et al. LincRNA-p21 alleviates atherosclerosis progression through regulating the miR-221/SIRT1/Pcsk9 axis. J Cell Mol Med. 2021;25(19):9141-9153. [CrossRef]
- Li C, Zhang M, Dai Y, Xu Z. MicroRNA-424-5p regulates aortic smooth muscle cell function in atherosclerosis by blocking APOC3-mediated nuclear factor-κB signalling pathway. Exp Physiol. 2020;105(6):1035-1049. [CrossRef]
- Ichihara A, Jinnin M, Yamane K, et al. microRNA-mediated keratinocyte hyperproliferation in psoriasis vulgaris. Br J Dermatol. 2011;165(5):1003-1010. [CrossRef]
- Zhang ZZ, Chen JJ, Deng WY, Yu XH, Tan WH. CTRP1 decreases ABCA1 expression and promotes lipid accumulation through the miR-424-5p/FoxO1 pathway in THP-1 macrophage-derived foam cells. Cell Biol Int. 2021;45(11):2226-2237. [CrossRef]
- Tsuru Y, Jinnin M, Ichihara A, et al. miR-424 levels in hair shaft are increased in psoriatic patients. J Dermatol. 2014;41(5):382-385. [CrossRef]
- Wang S, Xu J, Guo Y, et al. MicroRNA-497 Reduction and Increase of Its Family Member MicroRNA-424 Lead to Dysregulation of Multiple Inflammation Related Genes in Synovial Fibroblasts With Rheumatoid Arthritis. Front Immunol. 2021;12:619392. Published 2021 Mar 26. [CrossRef]
- Bao MH, Feng X, Zhang YW, Lou XY, Cheng Y, Zhou HH. Let-7 in cardiovascular diseases, heart development and cardiovascular differentiation from stem cells. Int J Mol Sci. 2013;14(11):23086-23102. Published 2013 Nov 21. [CrossRef]
- Zhu L, Li Q, Qi D, et al. Atherosclerosis-associated endothelial cell apoptosis by miRNA let7-b-mediated downregulation of HAS-2 [published online ahead of print, 2019 Nov 17]. J Cell Biochem. 2019;10.1002/jcb.29537. [CrossRef]
- Yu C, Wu B, Jiang J, Yang G, Weng C, Cai F. Overexpressed lncRNA ROR Promotes the Biological Characteristics of ox-LDL-Induced HUVECs via the let-7b-5p/HOXA1 Axis in Atherosclerosis. Front Cardiovasc Med. 2021;8:659769. Published 2021 Sep 13. [CrossRef]
- Long G, Wang F, Li H, et al. Circulating miR-30a, miR-126 and let-7b as biomarker for ischemic stroke in humans. BMC Neurol. 2013;13:178. Published 2013 Nov 16. [CrossRef]
- Huang YQ, Huang C, Chen JY, Li J, Feng YQ. Plasma expression level of miRNA let-7 is positively correlated with carotid intima-media thickness in patients with essential hypertension. J Hum Hypertens. 2017;31(12):843-847. [CrossRef]
- Hu XP, Xie Q, Chen CF, Zhang W, Yu B. Let-7a Inhibits T-Cell Proliferation and IFN-γ Secretion by Down-Regulating STAT3 Expression in Patients with Psoriasis. Cell Physiol Biochem. 2017;42(1):115-125. [CrossRef]
- Pasquali L, Svedbom A, Srivastava A, et al. Circulating microRNAs in extracellular vesicles as potential biomarkers for psoriatic arthritis in patients with psoriasis. J Eur Acad Dermatol Venereol. 2020;34(6):1248-1256. [CrossRef]
- Rangel-Zuñiga OA, Vals-Delgado C, Alcala-Diaz JF, et al. A set of miRNAs predicts T2DM remission in patients with coronary heart disease: from the CORDIOPREV study. Mol Ther Nucleic Acids. 2020;23:255-263. Published 2020 Nov 11. [CrossRef]
- Brennan E, Wang B, McClelland A, et al. Protective Effect of let-7 miRNA Family in Regulating Inflammation in Diabetes-Associated Atherosclerosis. Diabetes. 2017;66(8):2266-2277. [CrossRef]
- Aljaibeji H, Elemam NM, Mohammed AK, et al. Let7b-5p is Upregulated in the Serum of Emirati Patients with Type 2 Diabetes and Regulates Insulin Secretion in INS-1 Cells. Exp Clin Endocrinol Diabetes. 2022;130(1):22-29. [CrossRef]
- Tang J, Lin J, Yu Z, et al. Identification of circulating miR-22-3p and let-7a-5p as novel diagnostic biomarkers for rheumatoid arthritis. Clin Exp Rheumatol. 2022;40(1):69-77. [CrossRef]
- Ormseth MJ, Solus JF, Sheng Q, et al. Plasma miRNAs improve the prediction of coronary atherosclerosis in patients with rheumatoid arthritis. Clin Rheumatol. 2021;40(6):2211-2219. [CrossRef]
- Schmitz SU, Grote P, Herrmann BG. Mechanisms of long noncoding RNA function in development and disease. Cell Mol Life Sci. 2016;73(13):2491-2509. [CrossRef]
- Hurst LD, Smith NG. Molecular evolutionary evidence that H19 mRNA is functional. Trends Genet. 1999;15(4):134-135. [CrossRef]
- Huang Y, Wang L, Mao Y, Nan G. Long Noncoding RNA-H19 Contributes to Atherosclerosis and Induces Ischemic Stroke via the Upregulation of Acid Phosphatase 5. Front Neurol. 2019;10:32. Published 2019 Feb 4. [CrossRef]
- Cao L, Zhang Z, Li Y, Zhao P, Chen Y. LncRNA H19/miR-let-7 axis participates in the regulation of ox-LDL-induced endothelial cell injury via targeting periostin. Int Immunopharmacol. 2019;72:496-503. [CrossRef]
- Zhu R, Liu X, He Z. Long non-coding RNA H19 and MALAT1 gene variants in patients with ischemic stroke in a northern Chinese Han population. Mol Brain. 2018;11(1):58. Published 2018 Oct 10. [CrossRef]
- Bitarafan S, Yari M, Broumand MA, et al. Association of Increased Levels of lncRNA H19 in PBMCs with Risk of Coronary Artery Disease. Cell J. 2019;20(4):564-568. [CrossRef]
- Zhang L, Cheng H, Yue Y, Li S, Zhang D, He R. H19 knockdown suppresses proliferation and induces apoptosis by regulating miR-148b/WNT/β-catenin in ox-LDL -stimulated vascular smooth muscle cells. J Biomed Sci. 2018;25(1):11. Published 2018 Feb 7. [CrossRef]
- Pan, JX. LncRNA H19 promotes atherosclerosis by regulating MAPK and NF-kB signaling pathway. Eur Rev Med Pharmacol Sci. 2017;21(2):322-328.
- Han Y, Ma J, Wang J, Wang L. Silencing of H19 inhibits the adipogenesis and inflammation response in ox-LDL-treated Raw264.7 cells by up-regulating miR-130b. Mol Immunol. 2018;93:107-114. [CrossRef]
- He Y, Yin X, Yan J, Li X, Sun Q. The lncRNA H19/miR-766-3p/S1PR3 Axis Contributes to the Hyperproliferation of Keratinocytes and Skin Inflammation in Psoriasis via the AKT/mTOR Pathway. Mediators Inflamm. 2021;2021:9991175. Published 2021 Dec 28. [CrossRef]
- Gupta R, Ahn R, Lai K, et al. Landscape of Long Noncoding RNAs in Psoriatic and Healthy Skin. J Invest Dermatol. 2016;136(3):603-609. [CrossRef]
- Fawzy MS, Abdelghany AA, Toraih EA, Mohamed AM. Circulating long noncoding RNAs H19 and GAS5 are associated with type 2 diabetes but not with diabetic retinopathy: A preliminary study. Bosn J Basic Med Sci. 2020;20(3):365-371. Published 2020 Aug 3. [CrossRef]
- Cheng XW, Chen ZF, Wan YF, Zhou Q, Wang H, Zhu HQ. Long Non-coding RNA H19 Suppression Protects the Endothelium Against Hyperglycemic-Induced Inflammation via Inhibiting Expression of miR-29b Target Gene Vascular Endothelial Growth Factor a Through Activation of the Protein Kinase B/Endothelial Nitric Oxide Synthase Pathway. Front Cell Dev Biol. 2019;7:263. Published 2019 Nov 1. [CrossRef]
- Tello-Flores VA, Valladares-Salgado A, Ramírez-Vargas MA, et al. Altered levels of MALAT1 and H19 derived from serum or serum exosomes associated with type-2 diabetes. Noncoding RNA Res. 2020;5(2):71-76. Published 2020 Apr 11. [CrossRef]
- Alfaifi M, Verma AK, Alshahrani MY, et al. Assessment of Cell-Free Long Non-Coding RNA-H19 and miRNA-29a, miRNA-29b Expression and Severity of Diabetes. Diabetes Metab Syndr Obes. 2020;13:3727-3737. Published 2020 Oct 14. [CrossRef]
- Alrefai AA, Khader HF, Elbasuony HA, Elzorkany KM, Saleh AA. Evaluation of the expression levels of lncRNAs H19 and MEG3 in patients with type 2 diabetes mellitus [published online ahead of print, 2023 Jun 9]. Mol Biol Rep. 2023;10.1007/s11033-023-08569-0. [CrossRef]
- Mahmoudi Z, Karamali N, Roghani SA, et al. Efficacy of DMARDs and methylprednisolone treatment on the gene expression levels of HSPA5, MMD, and non-coding RNAs MALAT1, H19, miR-199a-5p, and miR-1-3p, in patients with rheumatoid arthritis. Int Immunopharmacol. 2022;108:108878. [CrossRef]
- Lu Y, Qie D, Yang F, Wu J. LncRNA MEG3 aggravates adipocyte inflammation and insulin resistance by targeting IGF2BP2 to activate TLR4/NF-κB signaling pathway. Int Immunopharmacol. 2023;121:110467. doi:10.1016/j.intimp.2023.110467. [CrossRef]
- Tang ZL, Zhang K, Lv SC, Xu GW, Zhang JF, Jia HY. LncRNA MEG3 suppresses PI3K/AKT/mTOR signalling pathway to enhance autophagy and inhibit inflammation in TNF-α-treated keratinocytes and psoriatic mice [published correction appears in Cytokine. 2022 May;153:155853]. Cytokine. 2021;148:155657. [CrossRef]
- Zhang Y, Liu X, Bai X, et al. Melatonin prevents endothelial cell pyroptosis via regulation of long noncoding RNA MEG3/miR-223/NLRP3 axis. J Pineal Res. 2018;64(2):10.1111/jpi.12449. [CrossRef]
- Yu B, Wang S. Angio-LncRs: LncRNAs that regulate angiogenesis and vascular disease. Theranostics. 2018;8(13):3654-3675. Published 2018 Jun 8. [CrossRef]
- Jia HY, Zhang K, Lu WJ, Xu GW, Zhang JF, Tang ZL. LncRNA MEG3 influences the proliferation and apoptosis of psoriasis epidermal cells by targeting miR-21/caspase-8. BMC Mol Cell Biol. 2019;20(1):46. Published 2019 Oct 28. [CrossRef]
- Bai Y, Zhang Q, Su Y, Pu Z, Li K. Modulation of the Proliferation/Apoptosis Balance of Vascular Smooth Muscle Cells in Atherosclerosis by lncRNA-MEG3 via Regulation of miR-26a/Smad1 Axis. Int Heart J. 2019;60(2):444-450. [CrossRef]
- Wu Z, He Y, Li D, et al. Long noncoding RNA MEG3 suppressed endothelial cell proliferation and migration through regulating miR-21. Am J Transl Res. 2017;9(7):3326-3335. Published 2017 Jul 15.
- Chang WW, Zhang L, Yao XM, et al. Upregulation of long non-coding RNA MEG3 in type 2 diabetes mellitus complicated with vascular disease: a case-control study. Mol Cell Biochem. 2020;473(1-2):93-99. [CrossRef]
- Chatterjee S, Bhattcharjee D, Misra S, Saha A, Bhattacharyya NP, Ghosh A. Increase in MEG3, MALAT1, NEAT1 significantly predicts the clinical parameters in patients with rheumatoid arthritis. Per Med. 2020;17(6):445-457. [CrossRef]
- Wahba AS, Ibrahim ME, Mesbah NM, Saleh SM, Abo-Elmatty DM, Mehanna ET. Long non-coding RNA MEG3 and its genetic variant rs941576 are associated with rheumatoid arthritis pathogenesis in Egyptian patients. Arch Physiol Biochem. 2022;128(6):1571-1578. [CrossRef]
- Shefler A, Patrick MT, Wasikowski R, et al. Skin-Expressing lncRNAs in Inflammatory Responses. Front Genet. 2022;13:835740. Published 2022 Apr 26. [CrossRef]
- Ma XL, Wen GD, Yu C, Zhao Z, Gao N, Liu ZY. LncRNA UCA1 negatively regulates NF-kB activity in psoriatic keratinocytes through the miR125a-A20 axis. Kaohsiung J Med Sci. 2021;37(3):172-180. [CrossRef]
- Tian S, Yuan Y, Li Z, Gao M, Lu Y, Gao H. LncRNA UCA1 sponges miR-26a to regulate the migration and proliferation of vascular smooth muscle cells. Gene. 2018;673:159-166. [CrossRef]
- Yang JL, Han NH. LncRNA UCA1 stimulates the repair of hyperglycemic vascular smooth muscle cells through targeting miR-582-5p. Eur Rev Med Pharmacol Sci. 2020;24(24):12859-12866. [CrossRef]
- Yan ZF, Zhao XY, Liu W, Liu XP. UCA1 impacts progress of rheumatoid arthritis by inducing the apoptosis of fibroblast-like synoviocyte. Eur Rev Med Pharmacol Sci. 2018;22(4):914-920. [CrossRef]
- Raposo AC, Casanova M, Gendrel AV, da Rocha ST. The tandem repeat modules of Xist lncRNA: a swiss army knife for the control of X-chromosome inactivation. Biochem Soc Trans. 2021;49(6):2549-2560. [CrossRef]
- Yang J, Qi M, Fei X, Wang X, Wang K. Long non-coding RNA XIST: a novel oncogene in multiple cancers. Mol Med. 2021;27(1):159. Published 2021 Dec 20. [CrossRef]
- Wang Y, Jiang F, Chen F, Zhang D, Wang J. LncRNA XIST Engages in Psoriasis via Sponging miR-338-5p to Regulate Keratinocyte Proliferation and Inflammation. Skin Pharmacol Physiol. 2022;35(4):196-205. [CrossRef]
- Zhang Y, Tang Y, Yan J. LncRNA-XIST Promotes Proliferation and Migration in ox-LDL Stimulated Vascular Smooth Muscle Cells through miR-539-5p/SPP1 Axis. Oxid Med Cell Longev. 2022;2022:9911982. Published 2022 Jan 4. [CrossRef]
- Yang K, Xue Y, Gao X. LncRNA XIST Promotes Atherosclerosis by Regulating miR-599/TLR4 Axis. Inflammation. 2021;44(3):965-973. [CrossRef]
- Mo L, Jiang HB, Tian GR, Lu GJ. The proliferation and migration of atherosclerosis-related HVSMCs were inhibited by downregulation of lncRNA XIST via regulation of the miR-761/BMP9 axis. Kaohsiung J Med Sci. 2022;38(1):18-29. [CrossRef]
- Wang M, Hou Z, Li X, et al. Relationship of serum lncRNA XIST and miR-30d-5p levels with diabetic peripheral neuropathy in type 2 diabetes. Am J Transl Res. 2022;14(12):9001-9006. Published 2022 Dec 15.
- Liu BY, Li L, Bai LW, Xu CS. Long Non-coding RNA XIST Attenuates Diabetic Peripheral Neuropathy by Inducing Autophagy Through MicroRNA-30d-5p/sirtuin1 Axis. Front Mol Biosci. 2021;8:655157. Published 2021 Apr 28. [CrossRef]
- Sohrabifar N, Ghaderian SMH, Alipour Parsa S, Ghaedi H, Jafari H. Variation in the expression level of MALAT1, MIAT and XIST lncRNAs in coronary artery disease patients with and without type 2 diabetes mellitus. Arch Physiol Biochem. 2022;128(5):1308-1315. [CrossRef]
- Liu W, Song J, Feng X, Yang H, Zhong W. LncRNA XIST is involved in rheumatoid arthritis fibroblast-like synoviocytes by sponging miR-126-3p via the NF-κB pathway. Autoimmunity. 2021;54(6):326-335. [CrossRef]
- Ferdin Jana and Lenassi Metka. Extracellular Vesicles and their Clinical Potential. Medicinski razgledi. 2016; 55 (1): 63–82.
- Konkoth A, Saraswat R, Dubrou C, et al. Multifaceted role of extracellular vesicles in atherosclerosis. Atherosclerosis. 2021;319:121-131. [CrossRef]
- Thulin Å, Christersson C, Alfredsson J, Siegbahn A. Circulating cell-derived microparticles as biomarkers in cardiovascular disease. Biomark Med. 2016;10(9):1009-1022. [CrossRef]
- Amabile N, Rautou PE, Tedgui A, Boulanger CM. Microparticles: key protagonists in cardiovascular disorders. Semin Thromb Hemost. 2010;36(8):907-916. [CrossRef]
- Han WQ, Chang FJ, Wang QR, Pan JQ. Microparticles from Patients with the Acute Coronary Syndrome Impair Vasodilatation by Inhibiting the Akt/eNOS-Hsp90 Signaling Pathway. Cardiology. 2015;132(4):252-260. [CrossRef]
- Zu L, Ren C, Pan B, et al. Endothelial microparticles after antihypertensive and lipid-lowering therapy inhibit the adhesion of monocytes to endothelial cells. Int J Cardiol. 2016;202:756-759. [CrossRef]
- Pelletier F, Garnache-Ottou F, Angelot F, et al. Increased Levels of Circulating Endothelial-Derived Microparticles and Small-Size Platelet-Derived Microparticles in Psoriasis. Journal of Investigative Dermatology, Volume 131, Issue 7, 2011, Pages 1573-1576, ISSN 0022-202X. [CrossRef]
- Zhang X, McGeoch SC, Johnstone AM, et al. Platelet-derived microparticle count and surface molecule expression differ between subjects with and without type 2 diabetes, independently of obesity status. J Thromb Thrombolysis. 2014;37(4):455-463. [CrossRef]
- Zeiger F, Stephan S, Hoheisel G, Pfeiffer D, Ruehlmann C, Koksch M. P-Selectin expression, platelet aggregates, and platelet-derived microparticle formation are increased in peripheral arterial disease. Blood Coagul Fibrinolysis. 2000;11(8):723-728. [CrossRef]
- Papadavid E, Diamanti K, Spathis A, et al. Increased levels of circulating platelet-derived microparticles in psoriasis: Possible implications for the associated cardiovascular risk. World J Cardiol. 2016;8(11):667-675. [CrossRef]
- Tamagawa-Mineoka R, Katoh N, Kishimoto S. Platelet activation in patients with psoriasis: increased plasma levels of platelet-derived microparticles and soluble P-selectin. J Am Acad Dermatol. 2010;62(4):621-626. [CrossRef]
- Ho JC, Lee CH, Lin SH. No Significant Reduction of Circulating Endothelial-Derived and Platelet-Derived Microparticles in Patients with Psoriasis Successfully Treated with Anti-IL12/23. Biomed Res Int. 2016;2016:3242143. [CrossRef]
- Pelletier F, Garnache-Ottou F, Biichlé S, et al. Effects of anti-TNF-α agents on circulating endothelial-derived and platelet-derived microparticles in psoriasis. Exp Dermatol. 2014;23(12):924-925. [CrossRef]
- Hu SC, Lan CE. Psoriasis and Cardiovascular Comorbidities: Focusing on Severe Vascular Events, Cardiovascular Risk Factors and Implications for Treatment. Int J Mol Sci. 2017;18(10):2211. Published 2017 Oct 21. [CrossRef]
- Tan KT, Tayebjee MH, Lim HS, Lip GY. Clinically apparent atherosclerotic disease in diabetes is associated with an increase in platelet microparticle levels. Diabet Med. 2005;22(12):1657-1662. [CrossRef]
- Atehortúa L, Rojas M, Vásquez G, et al. Endothelial activation and injury by microparticles in patients with systemic lupus erythematosus and rheumatoid arthritis. Arthritis Res Ther. 2019;21(1):34. Published 2019 Jan 23. [CrossRef]
- Michael BNR, Kommoju V, Kavadichanda Ganapathy C, Negi VS. Characterization of cell-derived microparticles in synovial fluid and plasma of patients with rheumatoid arthritis. Rheumatol Int. 2019;39(8):1377-1387. [CrossRef]
- Shao S, Fang H, Li Q, Wang G. Extracellular vesicles in Inflammatory Skin Disorders: from Pathophysiology to Treatment. Theranostics. 2020;10(22):9937-9955. Published 2020 Aug 7. [CrossRef]
- Hoyer FF, Giesen MK, Nunes França C, Lütjohann D, Nickenig G, Werner N. Monocytic microparticles promote atherogenesis by modulating inflammatory cells in mice. J Cell Mol Med. 2012;16(11):2777-2788. [CrossRef]
- Murzina E, Dosenko V, Drevytska T, Litus O, Bardova K, Vozianova S. Relationship between mir-126 expression in children with psoriasis, disease progression and therapeutic response. J Med Life. 2021;14(5):667-675. [CrossRef]
- Feng S, Wang L, Liu W, Zhong Y, Xu S. MiR-126 correlates with increased disease severity and promotes keratinocytes proliferation and inflammation while suppresses cells' apoptosis in psoriasis. J Clin Lab Anal. 2018;32(9):e22588. [CrossRef]
- Pelosi A, Lunardi C, Fiore PF, et al. MicroRNA Expression Profiling in Psoriatic Arthritis. Biomed Res Int. 2018;2018:7305380. Published 2018 Apr 23. [CrossRef]
- Duan Y, Zou J, Mao J, et al. Plasma miR-126 expression correlates with risk and severity of psoriasis and its high level at baseline predicts worse response to Tripterygium wilfordii Hook F in combination with acitretin. Biomed Pharmacother. 2019;115:108761. [CrossRef]
- Takeshita J, Mohler ER, Krishnamoorthy P, et al. Endothelial cell-, platelet-, and monocyte/macrophage-derived microparticles are elevated in psoriasis beyond cardiometabolic risk factors. J Am Heart Assoc. 2014;3(1):e000507. Published 2014 Feb 28. [CrossRef]
- Chironi GN, Simon A, Boulanger CM, et al. Circulating microparticles may influence early carotid artery remodeling. J Hypertens. 2010;28(4):789-796. [CrossRef]
- Kandiyil N, MacSweeney ST, Heptinstall S, May J, Fox SC, Auer DP. Circulating Microparticles in Patients with Symptomatic Carotid Disease Are Related to Embolic Plaque Activity and Recent Cerebral Ischaemia. Cerebrovasc Dis Extra. 2019;9(1):9-18. [CrossRef]
- Ren S, Fan X, Peng L, et al. Expression of NF-κB, CD68 and CD105 in carotid atherosclerotic plaque. J Thorac Dis. 2013;5(6):771-776. [CrossRef]
- Li X, van der Meer JJ, van der Loos CM, et al. Microvascular endoglin (CD105) expression correlates with tissue markers for atherosclerotic plaque vulnerability in an ageing population with multivessel coronary artery disease. Histopathology. 2012;61(1):88-97. [CrossRef]
- Novikova OA, Nazarkina ZK, Cherepanova AV, et al. Isolation, culturing and gene expression profiling of inner mass cells from stable and vulnerable carotid atherosclerotic plaques. PLoS One. 2019;14(6):e0218892. Published 2019 Jun 26. [CrossRef]
- Marei I, Chidiac O, Thomas B, et al. Angiogenic content of microparticles in patients with diabetes and coronary artery disease predicts networks of endothelial dysfunction. Cardiovasc Diabetol. 2022;21(1):17. Published 2022 Feb 2. [CrossRef]
- Rodríguez-Carrio J, Alperi-López M, López P, et al. Altered profile of circulating microparticles in rheumatoid arthritis patients. Clin Sci (Lond). 2015;128(7):437-448. [CrossRef]
- Georgescu A, Simionescu M. Extracellular Vesicles: Versatile Nanomediators, Potential Biomarkers and Therapeutic Agents in Atherosclerosis and COVID-19-Related Thrombosis. Int J Mol Sci. 2021;22(11):5967. Published 2021 May 31. [CrossRef]
- Mavropoulos A, Rigopoulou EI, Liaskos C, Bogdanos DP, Sakkas LI. The role of p38 MAPK in the aetiopathogenesis of psoriasis and psoriatic arthritis. Clin Dev Immunol. 2013;2013:569751. [CrossRef]
- Diener C, Keller A, Meese E. Emerging concepts of miRNA therapeutics: from cells to clinic. Trends Genet. 2022;38(6):613-626. [CrossRef]
Figure 1.
Schematic representation of the role of extracellular vesicles in the pathogenesis of atherosclerosis and psoriasis. Legend: SMC: Smooth muscle cell, M: Macrophages, N: Neutrophils, EVs: extracellular vesicles, P: platelet, EC: Endothelial cell. Created with BioRender.com.
Figure 1.
Schematic representation of the role of extracellular vesicles in the pathogenesis of atherosclerosis and psoriasis. Legend: SMC: Smooth muscle cell, M: Macrophages, N: Neutrophils, EVs: extracellular vesicles, P: platelet, EC: Endothelial cell. Created with BioRender.com.
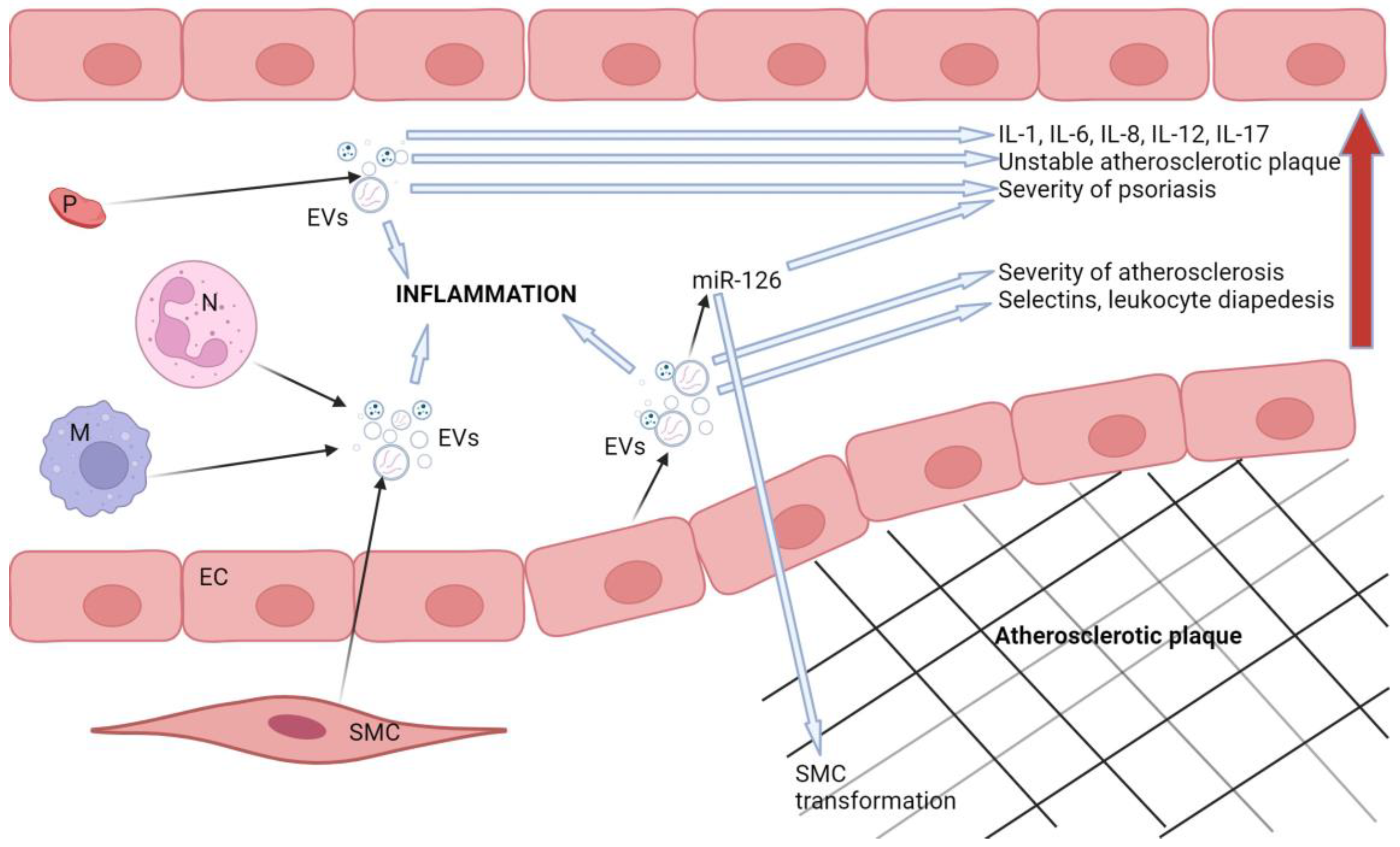

Table 1.
Summary of the clinical studies that investigated the roles of microRNAs and were reviewed in this article.
Table 1.
Summary of the clinical studies that investigated the roles of microRNAs and were reviewed in this article.
| Type ofmicroRNA | Atherosclerosis | Psoriasis | Type 2 diabetes mellitus | Rheumatoid arthritis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Authors of the study | No. of patients/samples (Cases/controls) | Main findings | Authors of the study | No. of patient (Cases/controls) | Main findings | Authors of the study | No. of patients (Cases/controls) | Main findings | Authors of the study | No. of patients (Cases/controls) | Main findings | |
| miR-146a | Huang et al. | 180/90 | ↑ | Leal et al. | 99/78 | ↑ | Alipoor et al. | 344/316 | ↓ | Bae et al. | 683/477 | ↑ |
| Raitoharju et al. | 30/20 | ↑ | Ele-Refaei et al. | 40/20 | ↑ | Zhu and Leung | Adipose 24 | ↑ | ||||
| Zhelankin et al. | 50/30 | ↑ | Vaher et al. | 26/26 | ↑ | Zhu and Leung | PBMC 140 | ↓ | ||||
| Guo et al. | 42/58 | ↑ | ||||||||||
| Gao et al. | 56/56 | ↑ | ||||||||||
| miR-210 | Signorelli et al. | 27/27 | ↑ | Zhao et al. | 18/18 | ↑ | Li et al. | 32/32 | ↑ | Abdul-Maksoud et al. | 100/100 | ↓ |
| Raitoharju et al. | 30/20 | ↑ | Wu et al. | 30/30 | ↑ | Amr et al. | 100/20 | ↑ | Huang et al. | 38/45 | X | |
| Zhou et al. | 10/10 | ↓ | ||||||||||
| Zhang et al | 32/20 | ↓ | ||||||||||
| Zhu and Leung | 540 | ↑ | ||||||||||
| miR-143 | Meng et al. | 279/65 | ↓ | Løvendorf et al. | 55/33 | ↑ | Zhu and Leung | 112 | ↑ | Yue et al. | 117/6 | ↑ |
| Zheng et al. | 194/175 | ↓ | ||||||||||
| miR-223 | Singh et al. | 250/250 | ↑ | Løvendorf et al. | 55/33 | ↑ | Parrizas et al. | 1184/838 | ↓ | Ormseth et al. | 168/91 | ↑ |
| Guo et al. | 300/100 | ↑ | Pivarcsi et al. | 43/22 | x | Zhu and Leung | Plasma 309 | ↓ | ||||
| Zhu et al. | 52/25 | ↓ | Alatas et al. | 52/54 | ↓ | |||||||
| miR-155 | Fichtlscherer et al. | 31/14 | ↓ | García-Rodríguez et al. | 11/11 | ↑ | Zhu and Leung | Whole blood 120 | ↑ | Abdul-Maksoud et al. | 100/100 | ↑ |
| Li et al. | 70/55 | ↑ | Alatas et al. | 52/54 | ↑ | Zhu and Leung | PBMC 80 | ↓ | Elmesmari et al. | 24/22 | ↑ | |
| Wang et al. | 3/x | |||||||||||
| miR-145 | Meng et al. | 279/65 | ↓ | Wang et al. | 45/40 | ↓ | Zhu and Leung | 24 | ↑ | Yue et al. | 117/6 | ↑ |
| Minin et al. | 105/72 | ↑ | Hong et al. | 5/5 | ↑ | |||||||
| Lv et al. | 328/374 | ↓ | ||||||||||
| Li et al. | 70/55 | x | ||||||||||
| miR-200 | Magenta et al. | 24/19 | ↑ | Magenta et al. | 29/29 | ↑ | Kujawa et al. | 5/5 | ↑ | Balzano et al. | 28/20 | x |
| Wang et al. | 189/109 | ↑ | Ofori et al. | 9/27 | ↑ | |||||||
| Zhu and Leung | Plasma 242 | ↓ | ||||||||||
| miR-133 | Wang et al. | 154/92 | ↑ | Chicharro et al. | 44/5 | ↓ | De Gonzalo-Calvo et al. | 72/x | ↑ | - | - | - |
| Al-Muhtaresh et al. | 30/30 | ↑ | Ghasemi et al | 35/35 | ↑ | |||||||
| Al-Muhtaresh et al. | 30/30 | ↑ | ||||||||||
| Zhu and Leung | 169 | ↓ | ||||||||||
| miR-135 | Xu et al. | 77/45 | ↑ | Chicharro et al. | 44/5 | ↓ | Sarookhani et. al | 30/30 | ↑ | Liu et al. | 3/x | ↓ |
| Li et al. | 47/47 | ↓ | Joyce et al. | 26/26 | ↑ | Monfared et al. | 80/40 | ↑ | ||||
| miR-221 | Minami et al. | 44/22 | ↑ | Zibert et al. | 13/13 | ↑ | Zhu and Leung | Serum 793 | ↑ | Abo ElAtta et al. | 30/20 | ↑ |
| Meng et al. | 46/42 | ↑ | Cunningham et al. | 50/20 | ↑ | |||||||
| Wade et al. | 31/20 | ↑ | Ciesla et al. | 50/24 | x | |||||||
| Mir-424 | Li et al. | 75/60 | ↓ | Ichihara et al. | 15/15 | ↓ | - | - | - | - | - | - |
| Alatas et al. | 52/54 | ↓ | ||||||||||
| Let-7 | Long et al. | 179/50 | ↑/↓ | Alatas et al. | 52/54 | ↑ | Aljaibeji et al. | 29/25 | ↑ | Cunningham et al. | 50/20 | ↑ |
| Huang et al. | 60/60 | ↑ | Hu et al. | 40/38 | ↓ | Zhu and Leung | Let-7-f Whole blood 89 | ↓ | Tang et al. | 76/36 | ↑ | |
| Minin et al. | 105/72 | ↑ | Zhu and Leung | Let-7-iserum 54 | ↓ | |||||||
| Yu et al. | 30/22 | ↓ | ||||||||||
Legend: ↑: increased expression/upregulation, ↓: decreased expression/downregulation, x: there was no statistically significant difference.
Table 2.
Summary of the clinical studies that investigated the roles of microRNAs and were reviewed in this article.
Table 2.
Summary of the clinical studies that investigated the roles of microRNAs and were reviewed in this article.
| Type of lncRNA | Atherosclerosis | Psoriasis | Type 2 diabetes mellitus | Rheumatoid arthritis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Authors of the study | No. of patients/samples (Cases/controls) | Main findings | Authors of the study | No. of patient (Cases/controls) | Main findings | Authors of the study | No. of patients (Cases/controls) | Main findings | Authors of the study | No. of patients (Cases/controls) | Main findings | |
| H19 | Cao et al. | 27/20 | ↑ | Gupta et al. | 18/16 | ↓ | Fawzy et al. | 119/110 | ↑ | Mahmoudi et al. | 25/25 | ↑ |
| Bitarafan et al. | 50/50 | ↑ | He et al. | 6/6 | ↓ | Cheng et al. | 30/30 | ↓ | ||||
| Zhang et al. | 40/40 | ↑ | Tello-Flores et al. | 60/60 | ↑ | |||||||
| Huang et al. | 80/85 | ↑ | Alfaifi et al. | 200/200 | ↓ | |||||||
| Pan et al. | 42/37 | ↑ | Alrefai et al. | 65/65 | ↓ | |||||||
| Han et al. | 30/30 | ↑ | ||||||||||
| MEG3 | Bai et al. | 40/35 | ↓ | Jia et al. | 19/19 | ↓ | Alrefai et al. | 65/65 | ↑ | Chatterjee et al. | 82/15 | ↑ |
| Wu et al. | 15/15 | ↓ | Chang et al. | 53/62 | ↑ | Wahba et al. | 100/100 | ↓ | ||||
| UCA1 | - | - | - | Ma et al. | 20/x | ↓ | Yang et al. | 40/40 | ↓ | - | - | - |
| XIST | Sohrabifar et al. | 25/25 | x | - | - | - | Wang et al. | 76/76 | ↓ | Liu et al. | 20/7 | ↑ |
| Sohrabifar et al. | 25/25 | ↑ | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



