
Submitted:
17 November 2023
Posted:
17 November 2023
You are already at the latest version
Abstract
Sulforaphane, the native but unstable form of SFX-01, is an antioxidant that activates NRF2 and inhibits NF-KB pathways to achieve its actions. Resolving the mechanism(s) by which SFX-01 serves to control the various osteoclastogenic stages may expose pathways that could be explored for therapeutic use. Here we seek to identify the stage of osteoclastogenesis targeted by SFX-01 and explore whether, like SFN, it exerts its actions via the NRF2 and NF-KB pathways. Osteoclasts generated from the bone marrow (BM) of mice were cultured with SFX-01 at different timepoints to examine each phase of osteoclastogenesis separately. This showed that SFX-01 exerted actions throughout the process of osteoclastogenesis, but had its largest effects in the early osteoclast precursor differentiation stage. Thus, treatment with SFX-01 for the duration of culture, for the initial 3 days differentiation or for as little as the first 24 hours was sufficient for effective inhibition. This aligned with data suggesting that SFX-01 reduced DC-STAMP levels, osteoclast nuclear number and modified cytoskeletal architecture. Pharmacological regulation of NRF2 pathways, via selective inhibitors/activators, supported anti-osteoclastogenic roles for an SFX-01-mediated by NRF2 activation, as well as a need for tight NF-KB pathway regulation in osteoclast formation/function.
Keywords:
sulforaphane
; osteoclasts
; osteoclastogenesis
; NRF2 and NF-KB
1. Introduction
Several clinical conditions including osteoporosis, Paget’s disease and other cancer-associated rarer bone diseases are characterised by bone loss and increased osteoclast activity [1,2] . Targeting of osteoclasts in skeletal pathobiology has thus been explored and has generated pharmacological agents capable of modulating osteoclastogenesis, resorption and ensuing bone loss. These agents, including bisphosphonates, selective estrogen receptor modulators and anti-RANKL antibodies, are effective at restricting osteoclastic bone resorption but nonetheless exert undesirable side effects such as impaired bone remodelling [2]. For this reason, there remains demand for alternative approaches to control osteoclast formation and bone resorptive activity; antioxidant agents have been attributed with such anti-resorptive therapeutic potential [3,4].
Sulforaphane (4-methylsulfinylbutyl isothiocyanate, SFN) is a phytochemical produced from a biologically inactive precursor, glucoraphanin, during biting and chewing of certain plant sources [5,6,7,8,9]; this precursor is found in cruciferous vegetables and many nutraceuticals [10]. SFN is usually well tolerated and is known to exert potent antioxidant effects by modifying cytoplasmic KEAP1 conformation, releasing NRF2 for nuclear translocation and transcriptional activation of genes with antioxidant response elements (ARE) in their promoters [11,12,13,14,15]. While cellular SFN uptake leads to an initial burst in ROS production, its rapid activation of the KEAP1-NRF2-ARE system induces an overall antioxidative or anti-inflammatory profile of cellular behaviours [16,17,18]. SFN is, however, highly unstable, rapidly degraded by heat and light, and highly sensitive to oxygen and pH [19]. SFN pharmacokinetics are therefore highly variable and inconsistent, making it difficult to ensure effective levels for sufficient duration for an attractive therapeutic, and its clinical advances have been restricted.
Technology for synthesising and concurrently stabilising SFN has therefore been developed to unlock its pharmaceutical potential. This has yielded a stable alpha-cyclodextrin: SFN complex, SulforadexTM (SFX-01), that is safe, well tolerated in humans, but rarely compared to SFN. SFX-01 is, nonetheless, known to recapitulate the vital targeting of NRF2 and STAT3 [20] to inhibit lung metastasis by endocrine resistant stem-like cells in estrogen receptor (ER)-positive breast cancer [20]. This clinical candidature of SFX-01 is reinforced by its efficacy in a mouse model of relapsing experimental autoimmune encephalomyelitis in which it reduces residual disability, decreases maximum severity of relapses and improves recovery [21]. Beneficial effects of SFX-01 have also been observed in a preclinical human glioblastoma model [22]. This has led to the examination of SFX-01 as a therapeutic in Phase 2 trials in ER-positive metastatic breast cancer and spontaneous subarachnoid haemorrhage patients [23].
Few studies have, however, explored the effects of SFX-01 in skeletal pathophysiology. SFX-01 administration has been found to reduce levels of gait asymmetry that emerge spontaneously in a natural osteoarthritis model in STR/Ort mice [24], in which such asymmetries are otherwise predictive of disease progression [25]. This was accompanied by increases in trabecular bone mass and indices of bone strength, and higher procollagen type-I N-propeptide (PINP, bone formation) and, notably, lower type-I collagen cross-linked C-telopeptide (CTX-1, bone resorption) serum markers. It has also been shown that divergent basal bone mass in different mouse strains is linked both to in vitro osteoclastogenic potential and that this aligns with sensitivity to SFX-01 [26]. These data suggest that SFX-01 represents an alternative approach to control osteoclast formation and resorptive activity.
There are extensive studies using SFN that support this notion. SFN inhibits RANKL-induced NF-KB activation in RAW 264.7 cells [27] and restricts expression of the cell fusion molecules, DC-STAMP and OC-STAMP, by inducing STAT1 phosphorylation [27]. This aligns with microarrays in RAW 264.7 cells and primary osteoclasts in which SFN lowered OSCAR, NFATc1 and TRAP, DC-STAMP and OC-STAMP mRNA levels whilst STAT1, a macrophage and osteoclast fusion regulator, was instead increased [28]. Hyeon et al. (2013) explored links between SFN and NRF2, to find both more osteoclast multinucleation and resorption, with raised oxidative stress, in cells from NRF2 knockout mice, and SFN-related inhibition of osteoclast differentiation in cells from wildtype mice [29], suggesting that SFN likely acts through NRF2 to suppress osteoclastogenesis. In vivo data showing elevated metaphyseal cancellous bone volume in both normal and ovariectomised mice treated with SFN indicate that SFN can indeed reduce the bone loss due to estrogen-deficiency [30].
These data indicate that SFN and SFX-01 may have therapeutic value in skeletal diseases where bone resorption or accelerated remodelling is causal. It is therefore necessary to define whether SFX-01 also exerts antiresorptive actions and, if so, which osteoclastogenic stage(s) it targets and its mechanism(s) of action. This will verify its therapeutic potential and likely identify alternative antiresorptive candidates.
It is appropriate to also reflect on the methods used to study osteoclastogenesis. Initial advances involved cell isolation from fragmented neonatal bones and their deposition onto bone/dentine to allow for pit excavation [31]. Long-term in vitro osteoclast formation now uses haematopoietic precursors from marrow, spleen or peripheral blood [32,33]; supplementation with, for example, PTH, M-CSF and RANKL with similar deposition onto bone/dentine. This has the advantage of allowing separate examination of key osteoclast formation, activation and resorption phases, impossible on plastic. Almost all studies reporting inhibitory SFN effects osteoclastogenesis have relied, however, on RAW 264.7 cell lines or primary cells seeded onto plastic, showing direct effects on differentiation, not resorption [27,28,34]. One study elegantly extended this to study cells seeded on dentine, reporting that SFN reduces resorption pit number, not area of dentine resorbed per osteoclast, concluding that SFN strongly inhibited osteoclast formation at an early precursor differentiation stage [27]. Here, using primary osteoclasts on dentine, we examine how SFX-01 influences each of the key osteoclast formation, activation and resorption phases, and explore whether, like SFN, it exerts its actions via the NRF2 and NF-KB pathways.
2. Materials and Methods
2.1. Animals
All the procedures conducted in the facility were in accordance with the Animals Act (Scientific Procedures) 1986 and approved by the Royal Veterinary College (RVC) Research Ethics Committee. Bone marrow (BM) cells were isolated from C57BL/6, CBA, and STR/Ort strains. Mice were housed at 21 ± 2°C with 12-h light/dark cycles with free access to food and water.
2.2. Osteoclast cell culture
Osteoclast precursors were isolated from the bone marrow of mouse long bones (6-8 weeks old) and cultured as described previously [26] [33]. Briefly, the resultant BM suspension was then centrifuged at 1500 rpm and resuspended in MEM supplemented with 10% foetal calf serum (FCS), 2 mM l-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin and 0.25 μg/ml amphotericin, 100 nM prostaglandin E2 (PGE2) and 50 ng/ml M-CSF. The cell suspension was cultured for 24 h in 75 cm² flasks at 5% CO2/95% atmospheric air. The next day (Day 1), the non-adherent cell suspension was removed, centrifuged, and resuspended in MEM containing 10% FCS, 2 mM l-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin and 0.25 μg/ml amphotericin,100 nM PGE2 and 200 ng/ml M-CSF as well as 5 ng/ml RANKL (R&D Systems Europe Ltd.)—referred to as S2MEM, see below. Cells were plated onto 5 mm diameter dentine discs (106 cells/disc) in 96-well plates and incubated overnight at 37°C/5% CO2 to allow for attachment of osteoclast precursors. After 24 h (Day 2), the dentine discs were transferred to six-well plates (3–4 discs/well in 3 ml S2MEM) and maintained for 3 days. S2MEM was acidified to pH 6.9 (Day 5) by addition of HCl to activate osteoclast resorption for 48 h (Day 7).
To establish stages at which SFX-01 exerts actions on osteoclast formation, differentiation and resorption, cells were cultured in osteoclastogenic medium, with or without 2.5μΜ SFX-01 during very specifically defined and different time points; 2.5 μΜ was used as it has been previously showed to be the optimal concentration to inhibit osteoclastogenesis [26]. Cells were exposed to SFX-01 (or vehicle) either: i) for the duration of post-plating culture from day 1-7 (during culture; DC; D1-D7); ii) at only the early stage (ES; D1-D2) for 24 hours after plating; iii) during only the differentiation stage and excluding the resorption phase (DS, between D2 and D5); iv) during only the later resorption phase (LS; D5-D7) (Figure 1). All cultures were replenished with fresh control medium during the periods not supplemented with SFX-01 and in all cases, experimental protocols were terminated on day 7 of culture.
For some experiments, pharmacological inhibitors/activators of NRF2 and NF-KB were used initially alone for the duration of the culture (DC), at one of two different concentrations (1μM and 10μM). For NRF2 activation and inhibition, RA839 and Trigonelline (Trig) were used respectively; while for NF-KB activation and inhibition, Betulinic acid (BA) and SC-514 were used (all at 1μM or 10μM). Each compound was also added in conjugation with SFX-01 (2.5 μM) during the early stage (ES; D1-D2) of culture, for 24 hours after plating (at which time SFX-01 was found to be most effective).
All experiments used bone marrow cells derived from STR/Ort mice, with the sole exception of the experiment seeking to identify dose-dependent effects of pharmacological activators/ inhibitors of the NRF2 and NF-KB pathways in which CBA and C57BL/6 mice were used instead.
2.3. Analysis of osteoclast formation and bone resorption
Discs containing adherent cells/osteoclasts were fixed in 2% glutaraldehyde and stained to demonstrate tartrate-resistant acid phosphatase (TRAP) activity. Osteoclasts were defined as TRAP-positive cells containing two or more nuclei and/or clear evidence of resorption pit formation. Osteoclast number and the area resorbed on each disc were assessed ‘blind’ by transmitted light microscopy and reflective light microscopy and dot-counting morphometry using Image J, respectively [33]
2.4. Immunocytochemistry
Osteoclasts grown in tissue culture 96-well plates for 6 days were treated with SFX01 as described above at specific, defined timepoints (ES, DS or DC), and then fixed with 4% paraformaldehyde in PBS for 20 minutes. Cells were washed twice with 0.05% PBS Tween 20 (PBS-T) and then exposed to 1% BSA in PBS-T for 30 minutes to block non-specific binding sites, prior to incubation for 2 hours with an anti-DCSTAMP antibody (clone 1A2 primary antibody, 1:500). Fluorescently tagged goat anti-mouse IgG was used as secondary antibody (1:1000) and added to for 1 hour at room temperature in the dark. For F-actin ring staining, phalloidiniFluor488 reagent was added for 30 min at room temperature. After washing with PBS, DAPI was used to stain nuclei. Cells were imaged using a DMIRB microscope to allow for simultaneous imaging of three fluorescent labels.
2.5. ELISA for 4-HNE
The levels of lipid peroxidation as a stable marker for oxidative stress were measured using the 4-hydroxynonenal (4-HNE) ELISA kit (ab238538, Abcam, USA) according to manufacturer’s instructions. BM cells extracted from STR/Ort mice were cultured for 6 days on plastic discs as described above and SFX-01 was added only during the early (ES) and/or differentiation stage (DS) stages along with appropriate untreated control group. The cultures were replenished with fresh control medium during the periods not supplemented with SFX-01. The absorbance was read immediately on the microplate reader at 450nm. The concentrations of the 4-HNE adduct were quantified by comparison to absorbance of a known 4-HNE-BSA standard curve.
2.4. Statistics
All statistical analysis was performed using GraphPad Prism 8. To show the difference between two groups, a two-tailed paired t-test was performed. Between three or more groups, a repeated measures one-way analysis of variance or a repeated measures mixed-effects model was performed with Fisher's LSD post hoc analysis. Each individual reported set of data represents n = 4–5 biological ‘experimental’ replicates using cells derived from different animals. Individual experiments comprised 6–8 technical replicates (number of discs) per group.
3. Results
3.1. SFX-01 preferentially targets early stages of osteoclastogenesis but exerts lasting restriction upon osteoclast resorptive function
Initial studies explored the effects of SFX-01 supplementation either throughout the culture (DC; D1-D7) or only during the late (LS; D5-D7) or differentiation stages (DS; D2-D5; Figure 1). These studies indicate that SFX-01 achieved identical levels of reduction in osteoclast number and total area resorbed irrespective of treatment duration (Figure 2). These data also show that osteoclast number and total area resorbed, but not area resorbed/osteoclast, were significantly lower following SFX-01 treatment for the duration of culture from day 1-7 (DC) or D2-D5 (DS) compared with treatment during only the late stage (D5-D7). It was nonetheless evident that treatment with SFX-01 solely at the late stage (LS) did achieve significant, albeit less marked, reduction in both the number of osteoclasts and total area resorbed (vs. control). SFX-01 failed to exert significant modification in area resorbed/osteoclast under any of the dosing regimens (Figure 2).
3.2. SFX-01 exposure solely at a really early stage (D1-2) is sufficient for lasting inhibition of osteoclastogenesis/ resorption
The data above support a predominant effect of SFX-01 on osteoclast differentiation, which commences at the point when mononuclear osteoclast precursors are initially exposed to M-CSF and RANKL in the first 24 hours of culture. To explore how early and persistent these effects of SFX-01 are on osteoclast differentiation, transient SFX01 exposure was administered at only the very earliest stage (ES; D1-D2) of M-CSF and RANKL exposure during the initial 24 hours after plating. The results show that SFX-01 for the initial 24 hours was sufficient to inhibit osteoclastogenesis to a similar extent to that seen with SFX-01 throughout the culture (DC; D1-D7) or during later differentiation stages (DS; D2-D5). There were no significant differences found between DS, DC or ES (p>0.05), suggesting that this short and early exposure is sufficient for lasting inhibition, reducing osteoclast formation by around 50% in all cases compared with control (Figure 3A).
As early osteoclast differentiation accommodates vital osteoclast precursor multinucleation, the effects of early exposure to SFX-01 were additionally explored with DAPI staining to assess levels of multinucleation. These studies indicated that the addition of SFX-01 at ES generated cells with smaller overall size containing fewer nuclei compared to n the controls (treated with M-CSF and RANKL; Figure 3B).
This multinucleation, cell-cell fusion also involves morphological changes in cell architecture. To assess whether SFX-01 modifies these morphological changes involved in multinucleation, cells were cultured on plastic and labelled for polymerised cytoskeletal actin using phalloidin and for nuclear DNA using DAPI, with or without early prior SFX-01 exposure during the early stage. Visualisation of the control cultures revealed overt cytoskeletal spreading and the presence of cellular phalloidin-positive polymerized actin in osteoclasts (Figure 4A), but a complete absence of actin polymerisation in SFX-01 treated cultures (Figure 4B). The effects of SFX-01 on cell-cell fusion at ES were further assessed by triple-labelling for polymerised actin, nuclear DNA and for DC-STAMP using selective antibody labelling. Visualisation of labelling indicated a diffuse extra-nuclear distribution of DC-STAMP that co-localised with polymerised actin in control cells where multinucleation was supported by clusters of DAPI-stained nuclei (Figure 5A-D). In contrast, cells exposed to SFX-01 during DC, ES or DS sages exhibited only nuclear labelling for DCSTAMP, no co-localisation with polymerised actin (that was absent in these cells) and a failure in multinucleation (Figure 5E-P). The similarity in the outcomes of SFX-01 treatment in cells exposed during DC, ES and DS, confirms that the inhibitory actions of SFX-01 start during the early phase (Figure 5M-P).
3.3. NRF2 and NF-KB activators inhibit osteoclastogenesis
This section seeks to identify whether pharmacological activators/inhibitors of the NRF2 and NF-KB pathways independently affect osteoclastogenesis. Effects on osteoclastogenesis and resorption of the alkaloid trigonelline (Trig), a recognized inhibitor, and RA839 an activator of NRF2 have not previously been investigated. These studies were performed in osteoclasts from C57BL/6 and CBA mice, which we have previously found have greater sensitivity to exogenous anti-osteoclastogenic factors, and all activators/inhibitors were each added at 1 and 10μΜ. Trends were identical in cells from both mouse strains and these data are combined for statistical comparison. These studies showed that the NRF2 inhibitor, Trig did not effect on osteoclast number, total area resorbed or area resorbed/osteoclast at either 1μΜ or 10μΜ (Figure 6). In contrast, RA839 (NRF2 activator) exerted inhibition of osteoclast number, total area resorbed and area resorbed/osteoclast more than 50% reaching levels of statistical significance at 10μM.
Similar studies performed with NF-KB modulators show that the activator, betulinic acid (BA) significantly and concentration-dependently limited osteoclastogenesis and resorption (Figure 6). This suggests that NF-KB activation restricts osteoclast formation and resorption and, thus, it may be predicted that the NF-KB inhibitor, SC-514 may have opposing effects. Differentiation of osteoclasts in medium supplemented with SC-514 did not however conform to this simple expectation; SC-514 instead exerted mild, yet significant inhibition of osteoclast number, but no effects on total or per osteoclast resorption.
3.4. Co-administration of NRF2 activator/inhibitor does not further augment the effects of SFX-01
To further explore relationships between the actions of SFX-01 (2.5μΜ) and RA839 (10μΜ; NRF2 activator), the independent effects of each was compared to their combined action. Treatment during only the early phase (D1-D2, ES) in cells derived from STR/Ort mice showed that RA839 reduces the number of osteoclasts, less so than SFX-01 (Figure 7A), and that there is no additional inhibition by RA839 in the presence of SFX-01. In contrast with previous studies, early stage SFX-01 exposure also significantly reduced area resorbed/osteoclast. Nonetheless, there was a lack of any additional inhibition exerted by RA839 on either total area resorbed or area resorbed /osteoclast in the presence of SFX-01.
This conclusion was not, however, consistent with the actions of the NRF2 inhibitor Trig which would therefore have been expected to reverse or limit the inhibitory effects of SFX-01 on osteoclastogenesis if its actions were achieved solely by NRF2 activation. Trig significantly reduced osteoclast number and area resorbed/osteoclast (Figure 7B). When Trig was combined with SFX-01, there were only borderline differences compared to SFX-01 or Trig alone, and there was clearly a lack of any additional inhibition exerted by Trig on any of the measured parameters, suggesting that the mechanism of SFX-01 action upon osteoclastogenesis and resorption may involve modulation of the NRF2 pathway.
To explore whether SFX-01 modulates oxidative stress, the 4-HNE adduct was measured using ELISA in medium collected from osteoclast cultures treated at different stages with SFX-01. These data show that the collection on day 7 of medium conditioned by osteoclast cultures treated with SFX-01 during differentiation stages (DS, D2-D5) expressed lower 4-HNE levels, compared to control vehicle-treated cells, while those treated only during ES show less marked changes that did not reach levels of significance (Figure 8).
3.5. Co-administration of an NF-KB activator but not inhibitor augments SFX-01 effects of SFX-01
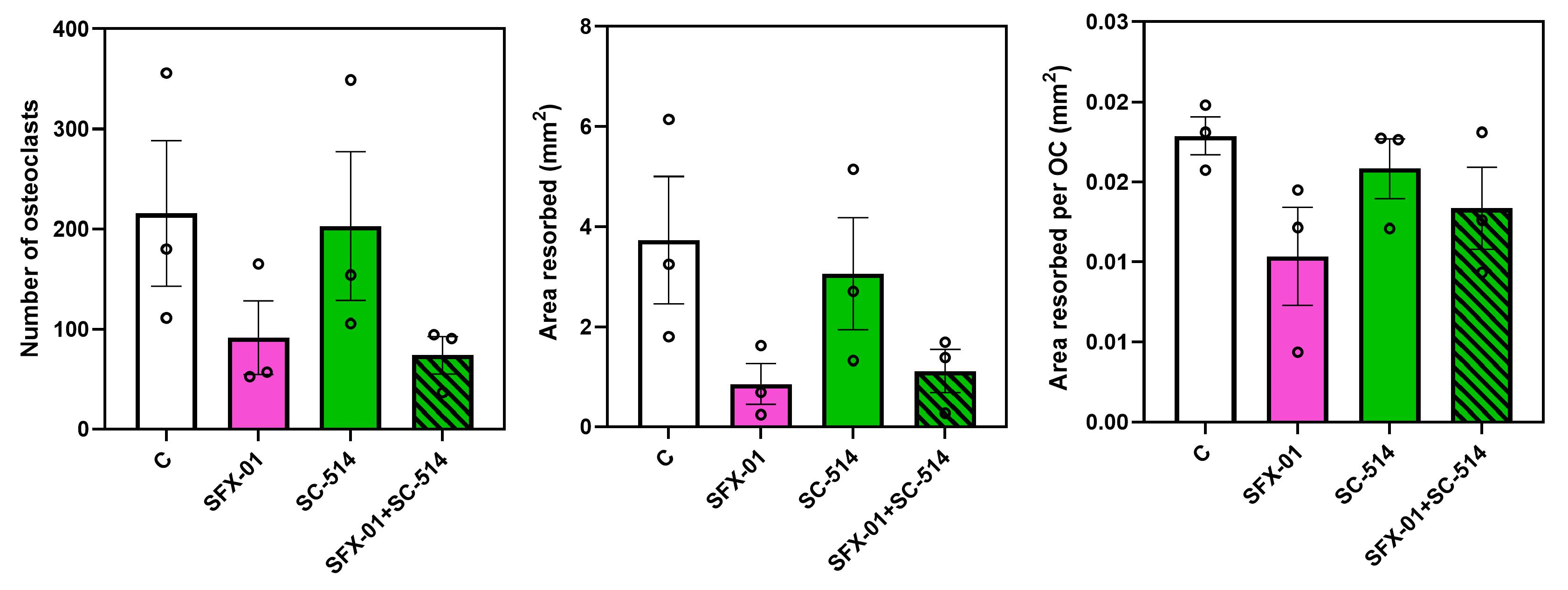
Treatment with 1μΜ BA, the NF-KB activator, during only early (D1-D2, ES) stages restricted osteoclastogenesis and resorption (Figure 9A), with lower osteoclast number, total resorbed area and area resorbed/ osteoclast by more than 50% (vs. controls). Intriguingly, combined BA/SFX-01 treatment generated significantly lower area resorbed/osteoclast but only modestly lower levels of total osteoclast numbers and area resorbed (non-significant) than with SFX01 alone. SC-514 (10μM) did not show any effect on osteoclast formation or resorption (Figure 9). Likewise, combined SC-514 and SFX-01 treatment failed to modify levels of osteoclast formation and resorption seen in cultures treated with SFX-01 alone, indicating that SC-514 did not have any effect either in the presence or absence of SFX-01.
4. Discussion
These data show that SFX-01 preferentially, rapidly and irreversibly targets early osteoclastogenic stages by inhibiting the cell-cell fusion and cell-spreading required for multinucleation. Based on the use of pharmacological agents, it appears that SFX-01 appears to achieve these effects, akin to SFN, via the NRF2 pathway. These are the first data exploring the mechanism by which SFX-01 influences osteoclastogenesis.
Herein, SFX-01 can target all stages of osteoclastogenesis but exerts the most profound actions on early osteoclast precursor differentiation to exert lasting restriction. This aligns with previous findings with SFN showing that it similarly targets osteoclastogenesis and specifically decreases osteoclast formation rate, with less TRAP+ cells [27,28,35] and expression of osteoclast differentiation-associated genes such as NFATc1, NF-KB, c-Fos, DC-STAMP [28]. These conclusions regarding SFN are based on the seeding of cells (RAW 264.7 or BMCs) onto plastic plates and thereby any possibility that SFN might directly influence resorption was negated. Only one study by Kim et al. (2005) describes assays using dentine to explore resorption, but no data for these assays are shown. To determine which osteoclastogenic stage was targeted by SFN, lymphocyte-free precursors were seeded on plastic plates and SFN was added at different timepoints to reveal that a minimum 3 days of exposure was required for inhibition [27]. This aligns somewhat with the data herein, in which 7 days, 3 days or as little as 24 hour (1 day) of SFX-01 treatment was sufficient for effective inhibition.
Mononuclear osteoclasts promote cell–cell fusion by increasing the expression levels of the fusogens DC-STAMP or OC-STAMP, which ultimately leads to osteoclast formation and bone resorption [36]. Additionally, it has been shown that Atp6v0d2-deficient mice exhibit impaired multinucleation emphasising its importance in cell fusion [37]. SFN decreases osteoclast cell fusion genes such as DC-STAMP, OC-STAMP, and Atp6v0d2 in sRANKL-induced multinucleated osteoclasts [28,35] and osteoclasts appear to be smaller in size with fewer nuclei. Also, the immunohistochemical data presented herein offer support for the notion that SFX-01 affects fusion/multinucleation, with some evidence indicating a decrease in the cellular levels of DC-STAMP and reduced labelling for polymerised actin.
There are many studies indicating that NRF2 plays an important role in bone cell regulation and homeostasis. Global NRF2 deletion increases RANKL expression and thereby increases number of osteoclasts and resorption [29,38,39,40]. Within this context, it is established that SFN activates NRF2, which via binding to the antioxidant response element in cognate genes serves to upregulate protective enzymes with known anti-inflammatory roles [17]. There are several methods available for measuring ROS. Herein, we quantified oxidative stress by assessment of the lipid peroxidation marker, 4-HNE. The results indicated that exposure of cells to SFX-01 also decreased levels of oxidative stress in addition to lowering levels of osteoclastogenesis. This indicates an SFX-01-induced modulation of oxidation status and suggests that this occurs through NRF2 activation.
As a prelude to examining whether SFX-01 also achieves its effects on osteoclasts via the modulation of NFR2, our studies were initially focused on whether pharmacological NRF2 modulators, namely Trig and RA839, exerted direct effects on osteoclast formation and function. The alkaloid, Trig, has been used in many studies as an inhibitor of NRF2 in other cell types [41,42,43]. On the other hand, RA839 has only recently been characterised as a non-covalent KEAP1 binding partner and selective activator of NRF2 [44]; neither have been used in assays of osteoclastogenesis previously. Initially we used each at both 1 and 10μM and found that Trig (NRF2 inhibitor), did not modify osteoclastogenesis or resorption in BM precursors, but intriguingly that RA839 (NRF2 activator) significantly inhibited osteoclast formation and osteoclast resorption/cell when administered at 10μM. These data imply that direct NRF2 activation with RA839 abrogates all osteoclast maturation and resorptive phases extending to the activity of individual osteoclasts, but that inhibition of NRF2 (Trig) is insufficient to stimulate osteoclastogenesis or resorption, under these assay conditions. These data align to some extent with the findings that SFX-01 achieves its inhibition via activation of this NRF2 pathway.
To further explore interaction between SFX-01 and direct NRF2 pathway modulation, the effects of RA839 (10μΜ) were examined in the presence and absence of known inhibitory concentrations of SFX-01 (2.5μΜ). As previous data had defined early stage SFX-01 supplementation sufficient for inhibition, all compounds were added only for the initial 24 hours (ES). This confirmed that RA839 administration, in the absence of SFX-01, reduced osteoclast number and area resorbed/ osteoclast in BM precursors. Combined addition of SFX-01/RA839 to BM cultures revealed similar inhibition levels in osteoclastogenesis and resorption achieved by administration of SFX-01 alone, implying that the pharmacological NRF2 activator was rendered ineffective by the presence of SFX-01. These results suggest that SFX-01 likely exerts its action via activation of NRF2 to inhibit osteoclastogenesis and total resorptive function.
The results with Trig (NRF2 inhibitor) are more difficult to interpret. Whilst without any measurable effect even at 10μΜ on cells derived from C57BL/6 and CBA mice, Trig instead significantly inhibited both osteoclast formation and resorption/OC in BM cells derived from STR/Ort mice. These differences may be explained by different mouse strain-related sensitivities based upon bone mass [26]. Nonetheless, there were no adjunct, additional, activities exerted by Trig over those exerted by SFX-01 alone in these studies, suggesting that Trig was ineffective when combined with SFX-01. The possibility that the NRF2-inhibitory functions of Trig may remain cryptic, unless combined with an activation of NRF2, in this case by SFX-01, would however require much more detailed pharmacological profiling. Likewise, it remains possible that Trig may not be very effective as an inhibitor of NRF2 under the conditions of these assays.
Treatment of BM cells with BA (NF-KB activator) revealed a concentration-dependent inhibition of osteoclast formation and resorption when added for the duration of the culture (DC). This aligns with prior studies that have found that BA suppresses osteoclast formation and acts as an inhibitor of bone resorption [45,46]. It nonetheless conflicts somewhat with the notion that NF-KB activation is required for OC formation; further studies into the actions of BA are required to resolve these apparent conflicts. Similar studies performed using SC-514, NF-KB inhibitor showed that it also restricted osteoclastogenesis without modifying resorption, even though one might predict based upon data from BA, that this NF-KB inhibitor would have opposing effects [47] .
To explore interaction between the effects of SFX-01 and direct NF-KB pathway modulation, the effects of BA (10μΜ) were examined with and without SFX-01 (2.5μΜ) supplementation only for these initial 24 hours (ES). Treatment of mouse cells consistently indicated that BA more efficiently targets area resorbed/osteoclast than it does osteoclastogenesis compared with SFX-01. Treatment with SC-514 failed to exert statistical differences in osteoclast behaviour; albeit using a relatively small sample size.
It is important to emphasise that these studies explore the actions of stable SFX-01, with a strong focus upon the NRF2 and NF-KB pathways that are most strongly implicated in mediating the effects of its unstable counterpart SFN. It is important to stress that the NRF2 and NF-KB pathways are not the only potential targets for SFN in osteoclasts. Recent studies indicate that SFN may also inhibit osteoclastogenesis by suppressing the autophagic pathway, with fewer autophagosomes in RAW 264.7 cells and attenuation in autophagy markers [34] . Simultaneous addition of rapamycin, an mTOR inhibitor that activates autophagy, was capable of reversing the inhibition of osteoclastogenesis induced by SFN [34]. The possibility that SFX-01 may also achieve its effects via these mechanisms has not however been explored. It is also feasible that the chemical stabilisation of SFN, SFX01, may allow alternative mechanisms to be targeted. These would perhaps be best identified by some non-targeted screening; the studies described here serve to both verify a cellular, osteoclast precursor target for SFX-01 actions and also define timeframes over which exposure to SFX-01 could be explored. In conclusion, these studies show that SFX-01 preferentially and more effectively targets early stages of osteoclastogenesis rather than resorption and that it does so relatively rapidly and irreversibly. They also identify prominent roles for NRF2 activation over NF-KB pathway involvement in SFX-01-related inhibition of osteoclastogenesis.
5. Conclusions
The studies show that SFX-01 preferentially and more effectively targets the early stages of osteoclastogenesis rather than resorption and that it does so relatively rapidly and irreversibly. The data also imply support roles for NRF2 activation much more strongly than they do roles for NF-KB involvement in SFX-01-related inhibition of osteoclastogenesis.
Author Contributions
Conceptualization, P.L., I.O., and A.A.P; methodology, P.L., I.O., and A.A.P; software, P.L.; validation, P.L.; formal analysis, P.L.; investigation, P.L.; resources, A.A.P X.X.; data curation, P.L., and A.A.P; writing—original draft preparation, P.L. and A.A.P ; writing—review and editing, P.L., I.O., and A.A.P; visualization, P.L.; supervision, I.O., and A.A.P; project administration, A.A.P.; funding acquisition, A.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported predominantly by iCASE funding from the BBSRC and Evgen Pharma PLC and from an EPSRC, Osteoarthritis Technology Network Plus grant sub-project award entitled: a multidisciplinary approach to the prevention and treatment of OA (EP/N027264/1).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
No humans involved.
Data Availability Statement
Data available on request from the authors.
Acknowledgments
We are grateful to Evgen Pharma for providing us with SFX-01.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Helfrich, M.H. Osteoclast diseases. Microsc Res Tech 2003, 61, 514–532. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yamauchi, K.; Mitsunaga, T. A review on osteoclast diseases and osteoclastogenesis inhibitors recently developed from natural resources. Fitoterapia 2020, 142, 104482. [Google Scholar] [CrossRef] [PubMed]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin Cases Miner Bone Metab 2017, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Xue, P.; Hu, X.; Powers, J.; Nay, N.; Chang, E.; Kwon, J.; Wong, S.W.; Han, L.; Wu, T.H.; Lee, D.J.; et al. CDDO-Me, Sulforaphane and tBHQ attenuate the RANKL-induced osteoclast differentiation via activating the NRF2-mediated antioxidant response. Biochem Biophys Res Commun 2019, 511, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Zalcmann At Fau - Talalay, P.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. 2001. (0031-9422 (Print)). [CrossRef]
- Schroeter, H. Phytochemicals: Mechanisms of Action: Edited by Mark S Meskin, Wayne R Bidlack, Audra J Davies, Douglas S Lewis, and R Keith Randolph, 203 pages, hardcover, $119.95. CRC Press, Boca Raton, FL. The American Journal of Clinical Nutrition 2004, 80, 1670–1670. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol Med 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Bello, C.; Maldini, M.; Baima, S.; Scaccini, C.; Natella, F. Glucoraphanin and sulforaphane evolution during juice preparation from broccoli sprouts. Food Chem 2018, 268, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Vanduchova, A.; Anzenbacher, P.; Anzenbacherova, E. Isothiocyanate from Broccoli, Sulforaphane, and Its Properties. J Med Food 2019, 22, 121–126. [Google Scholar] [CrossRef]
- Mahn, A.; Saavedra, A.; Paz Rubio, M. Kinetic study of sulforaphane stability in blanched and un-blanched broccoli (Brassica oleracea var. italica) florets during storage at low temperatures. J Food Sci Technol 2018, 55, 4687–4693. [Google Scholar] [CrossRef]
- Kensler, T.W.; Egner, P.A.; Agyeman, A.S.; Visvanathan, K.; Groopman, J.D.; Chen, J.G.; Chen, T.Y.; Fahey, J.W.; Talalay, P. Keap1-nrf2 signaling: A target for cancer prevention by sulforaphane. Top Curr Chem 2013, 329, 163–177. [Google Scholar] [CrossRef]
- Alfieri, A.; Srivastava, S.; Siow, R.C.M.; Cash, D.; Modo, M.; Duchen, M.R.; Fraser, P.A.; Williams, S.C.R.; Mann, G.E. Sulforaphane preconditioning of the Nrf2/HO-1 defense pathway protects the cerebral vasculature against blood-brain barrier disruption and neurological deficits in stroke. Free Radic Biol Med 2013, 65, 1012–1022. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wang, X.; Zhao, S.; Ma, C.; Cui, J.; Zheng, Y. Sulforaphane Protects against Cardiovascular Disease via Nrf2 Activation. Oxid Med Cell Longev 2015, 2015, 407580. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Zhang, M.Q.; Chen, Z.Y.; Wu, X.B.; Xia, Z.B.; Chai, J.Y.; Yin, X.P. Sulforaphane prevents PC12 cells from oxidative damage via the Nrf2 pathway. Mol Med Rep 2019, 19, 4890–4896. [Google Scholar] [CrossRef] [PubMed]
- Krajka-Kuźniak, V.A.-O.; Baer-Dubowska, W.A.-O. Modulation of Nrf2 and NF-κB Signaling Pathways by Naturally Occurring Compounds in Relation to Cancer Prevention and Therapy. Are Combinations Better Than Single Compounds? LID - 10.3390/ijms22158223 [doi] LID - 8223. Int J Mol Sci 2021, 22, 8223. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, L.O.; Kipp, M.; Lucius, R.; Pufe, T.; Wruck, C.J. Sulforaphane suppresses LPS-induced inflammation in primary rat microglia. Inflamm Res 2010, 59, 443–450. [Google Scholar] [CrossRef]
- Zakkar, M.; Van der Heiden, K.; Luong le, A.; Chaudhury, H.; Cuhlmann, S.; Hamdulay, S.S.; Krams, R.; Edirisinghe, I.; Rahman, I.; Carlsen, H.; et al. Activation of Nrf2 in endothelial cells protects arteries from exhibiting a proinflammatory state. Arterioscler Thromb Vasc Biol 2009, 29, 1851–1857. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, B.R.; Hansen, J.M. Extracellular redox status regulates Nrf2 activation through mitochondrial reactive oxygen species. The Biochemical Journal 2009, 424, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Jin, Z.; Xu, X.; Zhuang, H.; Shen, W. Preparation and stability of the inclusion complex of astaxanthin with hydroxypropyl-beta-cyclodextrin. Food Chem 2008, 109, 264–268. [Google Scholar] [CrossRef]
- Simoes, B.M.; Santiago-Gomez, A.; Chiodo, C.; Moreira, T.; Conole, D.; Lovell, S.; Alferez, D.; Eyre, R.; Spence, K.; Sarmiento-Castro, A.; et al. Targeting STAT3 signaling using stabilised sulforaphane (SFX-01) inhibits endocrine resistant stem-like cells in ER-positive breast cancer. Oncogene 2020, 39, 4896–4908. [Google Scholar] [CrossRef]
- Galea, I.; Copple, I.M.; Howat, D.W.; Franklin, S. SFX-01 reduces residual disability after experimental autoimmune encephalomyelitis. Mult Scler Relat Disord 2019, 30, 257–261. [Google Scholar] [CrossRef]
- Colapietro, A.; Rossetti, A.; Mancini, A.; Martellucci, S.; Ocone, G.; Pulcini, F.; Biordi, L.; Cristiano, L.; Mattei, V.; Delle Monache, S.; et al. Multiple Antitumor Molecular Mechanisms Are Activated by a Fully Synthetic and Stabilized Pharmaceutical Product Delivering the Active Compound Sulforaphane (SFX-01) in Preclinical Model of Human Glioblastoma. Pharmaceuticals (Basel) 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Zolnourian, A.H.; Franklin, S.; Galea, I.; Bulters, D.O. Study protocol for SFX-01 after subarachnoid haemorrhage (SAS): A multicentre randomised double-blinded, placebo controlled trial. BMJ Open 2020, 10, e028514. [Google Scholar] [CrossRef]
- Javaheri, B.; Poulet, B.; Aljazzar, A.; de Souza, R.; Piles, M.; Hopkinson, M.; Shervill, E.; Pollard, A.; Chan, B.; Chang, Y.; et al. Stable sulforaphane protects against gait anomalies and modifies bone microarchitecture in the spontaneous STR/Ort model of osteoarthritis. Bone 2017, 103, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Poulet, B.; de Souza, R.; Knights, C.B.; Gentry, C.; Wilson, A.M.; Bevan, S.; Chang, Y.M.; Pitsillides, A.A. Modifications of gait as predictors of natural osteoarthritis progression in STR/Ort mice. Arthritis Rheumatol 2014, 66, 1832–1842. [Google Scholar] [CrossRef] [PubMed]
- Louka, P.; Orriss, I.R.; Pitsillides, A.A. High bone mass in mice can be linked to lower osteoclast formation, resorptive capacity, and restricted in vitro sensitivity to inhibition by stable sulforaphane. Cell Biochem Funct 2022, 40, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kang, S.Y.; Shin, H.H.; Choi, H.S. Sulforaphane inhibits osteoclastogenesis by inhibiting nuclear factor-kappaB. Mol Cells 2005, 20, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Inoue, H.; Takahashi, N.; Katsumata-Tsuboi, R.; Uehara, M. Sulforaphane inhibits osteoclast differentiation by suppressing the cell-cell fusion molecules DC-STAMP and OC-STAMP. Biochem Biophys Res Commun 2017, 483, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Hyeon, S.; Lee, H.; Yang, Y.; Jeong, W. Nrf2 deficiency induces oxidative stress and promotes RANKL-induced osteoclast differentiation. Free Radic Biol Med 2013, 65, 789–799. [Google Scholar] [CrossRef]
- Thaler, R.; Maurizi, A.; Roschger, P.; Sturmlechner, I.; Khani, F.; Spitzer, S.; Rumpler, M.; Zwerina, J.; Karlic, H.; Dudakovic, A.; et al. Anabolic and Antiresorptive Modulation of Bone Homeostasis by the Epigenetic Modulator Sulforaphane, a Naturally Occurring Isothiocyanate. J Biol Chem 2016, 291, 6754–6771. [Google Scholar] [CrossRef]
- Chambers, T.J.; Revell, P.A.; Fuller, K.; Athanasou, N.A. Resorption of bone by isolated rabbit osteoclasts. J Cell Sci 1984, 66, 383–399. [Google Scholar] [CrossRef]
- Matsuzaki, K.; Udagawa, N.; Takahashi, N.; Yamaguchi, K.; Yasuda, H.; Shima, N.; Morinaga, T.; Toyama, Y.; Yabe, Y.; Higashio, K.; et al. Osteoclast differentiation factor (ODF) induces osteoclast-like cell formation in human peripheral blood mononuclear cell cultures. Biochem Biophys Res Commun 1998, 246, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Orriss, I.R.; Arnett, T.R. Rodent osteoclast cultures. Methods Mol Biol 2012, 816, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Fu, X.; Liu, Y.; Ji, Y.; Shang, Z. Sulforaphane Inhibits Osteoclastogenesis via Suppression of the Autophagic Pathway. Molecules 2021, 26. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Inoue, H.; Takahashi, N.; Katsumata-Tsuboi, R.; Uehara, M. Sulforaphene attenuates multinucleation of pre-osteoclasts by suppressing expression of cell-cell fusion-associated genes DC-STAMP, OC-STAMP, and Atp6v0d2. Biosci Biotechnol Biochem 2017, 81, 1220–1223. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, H.; Suzuki, T.; Miyauchi, Y.; Iwasaki, R.; Kobayashi, T.; Sato, Y.; Miyamoto, K.; Hoshi, H.; Hashimoto, K.; Yoshida, S.; et al. Osteoclast stimulatory transmembrane protein and dendritic cell-specific transmembrane protein cooperatively modulate cell-cell fusion to form osteoclasts and foreign body giant cells. J Bone Miner Res 2012, 27, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Rho, J.; Jeong, D.; Sul, J.Y.; Kim, T.; Kim, N.; Kang, J.S.; Miyamoto, T.; Suda, T.; Lee, S.K.; et al. v-ATPase V0 subunit d2-deficient mice exhibit impaired osteoclast fusion and increased bone formation. Nat Med 2006, 12, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, H.; Shinohara, F.; Kajiya, M.; Kodama, T. The Keap1/Nrf2 protein axis plays a role in osteoclast differentiation by regulating intracellular reactive oxygen species signaling. J Biol Chem 2013, 288, 23009–23020. [Google Scholar] [CrossRef]
- Park, C.K.; Lee, Y.; Kim, K.H.; Lee, Z.H.; Joo, M.; Kim, H.H. Nrf2 is a novel regulator of bone acquisition. (1873-2763 (Electronic)). [CrossRef]
- Sun, Y.X.; Xu, A.H.; Yang, Y.; Li, J. Role of Nrf2 in bone metabolism. J Biomed Sci 2015, 22, 101. [Google Scholar] [CrossRef]
- Arlt, A.; Sebens, S.; Krebs, S.; Geismann, C.; Grossmann, M.; Kruse, M.L.; Schreiber, S.; Schafer, H. Inhibition of the Nrf2 transcription factor by the alkaloid trigonelline renders pancreatic cancer cells more susceptible to apoptosis through decreased proteasomal gene expression and proteasome activity. Oncogene 2013, 32, 4825–4835. [Google Scholar] [CrossRef]
- Roh, J.-L.; Kim, E.H.; Jang, H.; Shin, D. Nrf2 inhibition reverses the resistance of cisplatin-resistant head and neck cancer cells to artesunate-induced ferroptosis. Redox Biology 2017, 11, 254–262. [Google Scholar] [CrossRef]
- Qin, W.; Guan, D.; Ma, R.; Yang, R.; Xing, G.; Shi, H.; Tang, G.; Li, J.; Lv, H.; Jiang, Y. Effects of trigonelline inhibition of the Nrf2 transcription factor in vitro on Echinococcus granulosus. Acta Biochim Biophys Sin (Shanghai) 2017, 49, 696–705. [Google Scholar] [CrossRef]
- Winkel, A.F.; Engel, C.K.; Margerie, D.; Kannt, A.; Szillat, H.; Glombik, H.; Kallus, C.; Ruf, S.; Gussregen, S.; Riedel, J.; et al. Characterization of RA839, a Noncovalent Small Molecule Binder to Keap1 and Selective Activator of Nrf2 Signaling. J Biol Chem 2015, 290, 28446–28455. [Google Scholar] [CrossRef]
- Kim, K.J.; Lee, Y.; Hwang, H.G.; Sung, S.H.; Lee, M.; Son, Y.J. Betulin Suppresses Osteoclast Formation via Down-Regulating NFATc1. J Clin Med 2018, 7. [Google Scholar] [CrossRef]
- Jeong, D.H.; Kwak, S.C.; Lee, M.S.; Yoon, K.H.; Kim, J.Y.; Lee, C.H. Betulinic Acid Inhibits RANKL-Induced Osteoclastogenesis via Attenuating Akt, NF-kappaB, and PLCgamma2-Ca(2+) Signaling and Prevents Inflammatory Bone Loss. J Nat Prod 2020, 83, 1174–1182. [Google Scholar] [CrossRef]
- Liu, H.; Talalay, P. Relevance of anti-inflammatory and antioxidant activities of exemestane and synergism with sulforaphane for disease prevention. Proc Natl Acad Sci U S A 2013, 110, 19065–19070. [Google Scholar] [CrossRef]
Figure 1.
Schematic illustration of dosing regimen to study the effects of SFX-01 on osteoclast differentiation and maturation. DC: treatment throughout (D1-D7). ES: treatment for first 24 hours (D1-D2). DS: treatment for differentiation stage (D2-D5). LS: SFX-01 for the resorption phase (D5-D7).
Figure 1.
Schematic illustration of dosing regimen to study the effects of SFX-01 on osteoclast differentiation and maturation. DC: treatment throughout (D1-D7). ES: treatment for first 24 hours (D1-D2). DS: treatment for differentiation stage (D2-D5). LS: SFX-01 for the resorption phase (D5-D7).
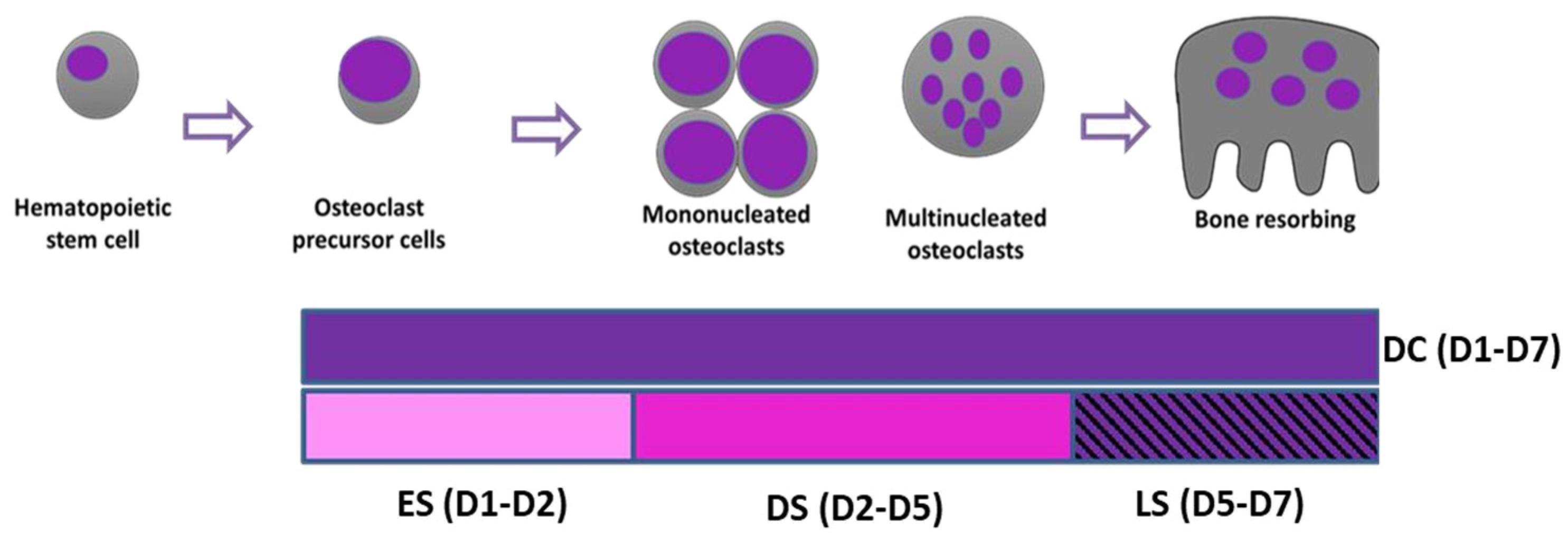
Figure 2.
SFX-01 targets osteoclast formation. A) 2.5μΜ SFX-01 was added at different time points during osteoclast culture, either throughout the culture (DC; D1-D7) or at later resorption stage (LS; D5-D7) or (differentiation stages (DS; D2-D5): A) TRAP+ osteoclast numbers (top left), area resorbed (middle), area resorbed/OC (top right, all mean ±SE) from STR/Ort mice. B Representative reflective light images of osteoclasts at different stages after SFX-01addition. $ and $$ denote p ≤ 0.05 and p ≤ 0.01 difference from control, respectively. *, ** and *** denote p ≤ 0.05, p ≤0.01 and p ≤0.001 differences within groups DC, LS, DS. C, respectively. Combined data from n=6 separate experiments, 6-8 replicates/condition/experiment.
Figure 2.
SFX-01 targets osteoclast formation. A) 2.5μΜ SFX-01 was added at different time points during osteoclast culture, either throughout the culture (DC; D1-D7) or at later resorption stage (LS; D5-D7) or (differentiation stages (DS; D2-D5): A) TRAP+ osteoclast numbers (top left), area resorbed (middle), area resorbed/OC (top right, all mean ±SE) from STR/Ort mice. B Representative reflective light images of osteoclasts at different stages after SFX-01addition. $ and $$ denote p ≤ 0.05 and p ≤ 0.01 difference from control, respectively. *, ** and *** denote p ≤ 0.05, p ≤0.01 and p ≤0.001 differences within groups DC, LS, DS. C, respectively. Combined data from n=6 separate experiments, 6-8 replicates/condition/experiment.
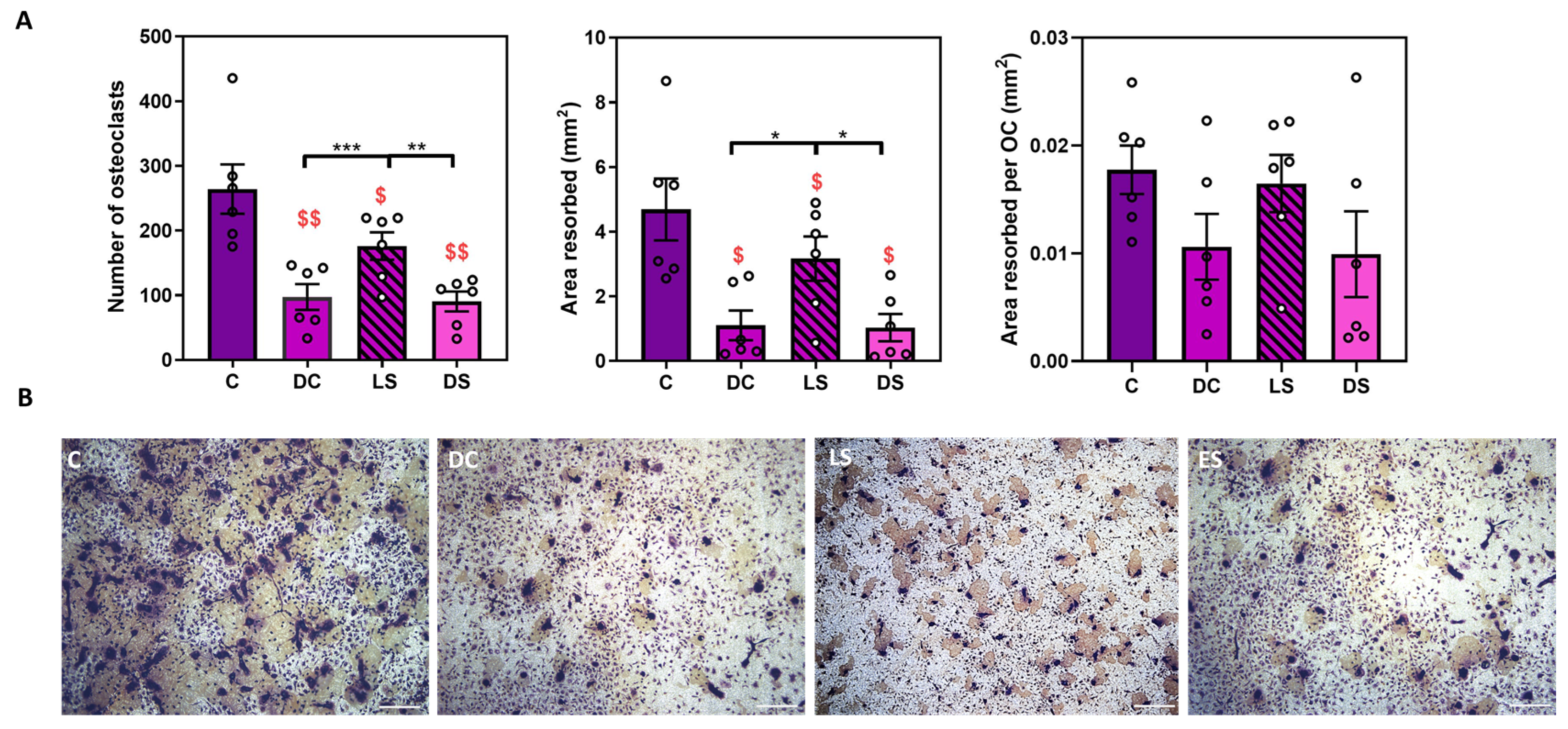
Figure 3.
24 hours exposure to SFX-01 is sufficient to inhibit osteoclastogenesis. (A) 2.5μΜ SFX-01 was added at defined timepoints during osteoclast culture either throughout the culture (DC; D1-D7) or at differentiation (DS; D2-D5), or only at early stage (ES; D1-D2) for 24 hours after plating on dentine and imaged and quantified conventionally: TRAP+ osteoclast numbers (top left), area resorbed (middle), area resorbed/OC (top right, all mean ±SE) from STR/Ort mice. $ and $$ denote p ≤ 0.05 and p ≤ 0.01 difference from control, respectively. Combined data from n=6 experiments, 6-8 replicates/condition. (B) 2.5μΜ SFX-01 was added during the early stage (ES) for 24 hours during osteoclast culture on plastic and imaged after DAPI nuclear staining to assess levels of multinucleation. SFX-01 reduces cell size and the number of nuclei comprising a single osteoclast.
Figure 3.
24 hours exposure to SFX-01 is sufficient to inhibit osteoclastogenesis. (A) 2.5μΜ SFX-01 was added at defined timepoints during osteoclast culture either throughout the culture (DC; D1-D7) or at differentiation (DS; D2-D5), or only at early stage (ES; D1-D2) for 24 hours after plating on dentine and imaged and quantified conventionally: TRAP+ osteoclast numbers (top left), area resorbed (middle), area resorbed/OC (top right, all mean ±SE) from STR/Ort mice. $ and $$ denote p ≤ 0.05 and p ≤ 0.01 difference from control, respectively. Combined data from n=6 experiments, 6-8 replicates/condition. (B) 2.5μΜ SFX-01 was added during the early stage (ES) for 24 hours during osteoclast culture on plastic and imaged after DAPI nuclear staining to assess levels of multinucleation. SFX-01 reduces cell size and the number of nuclei comprising a single osteoclast.
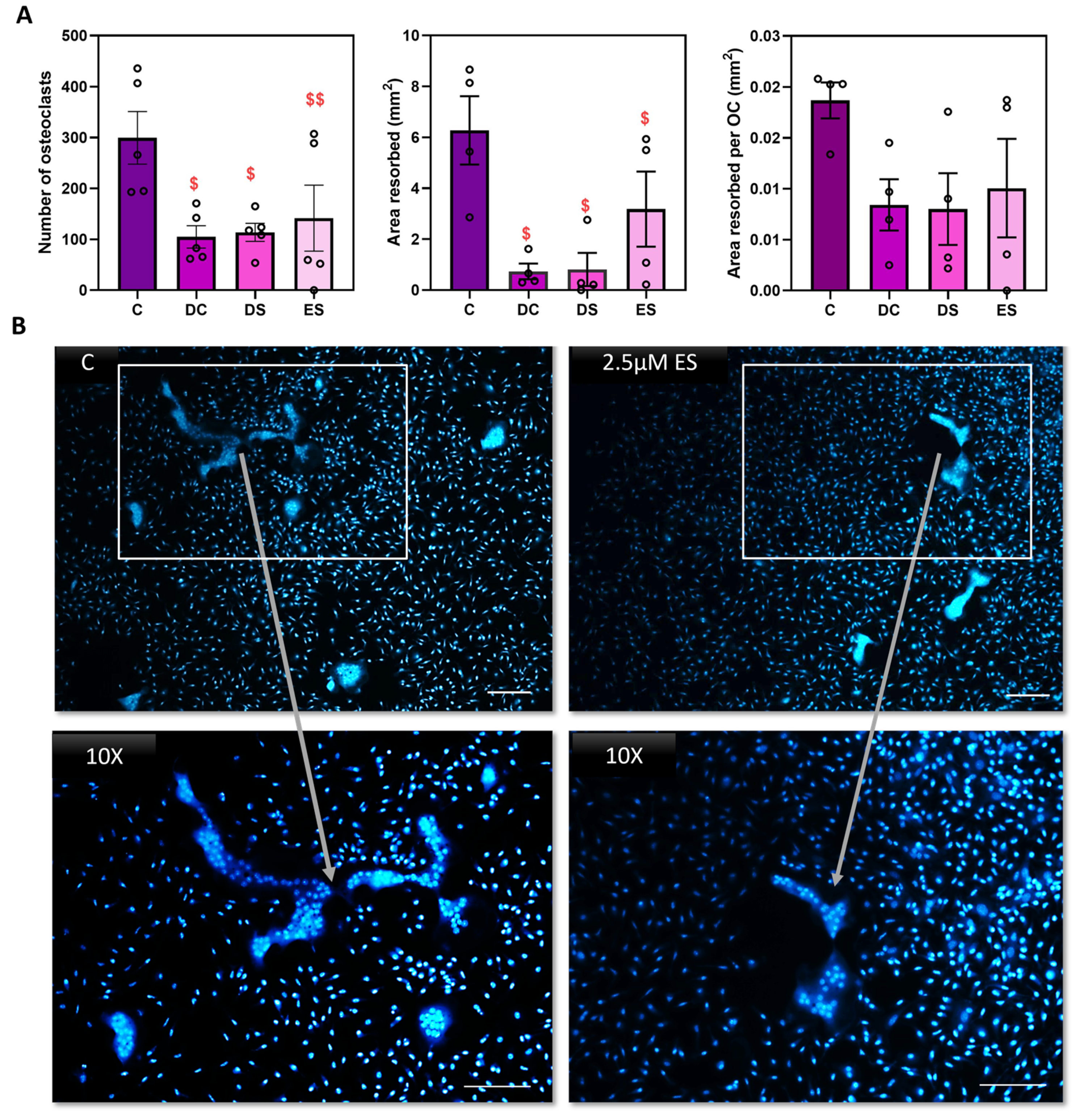
Figure 4.
Immunofluorescent labelling of mature OCs treated with SFX-01 at ES reveals modified polymerised actin organisation. Osteoclasts cultured for 6 days and then immunocytochemically labelled with phalloidin (for polymerised actin, green) and DAPI (for DNA, blue) and imaged by confocal microscopy indicating the difference in cytoskeletal architecture in untreated cells (A) and cells treated with SFX-01 (B).
Figure 4.
Immunofluorescent labelling of mature OCs treated with SFX-01 at ES reveals modified polymerised actin organisation. Osteoclasts cultured for 6 days and then immunocytochemically labelled with phalloidin (for polymerised actin, green) and DAPI (for DNA, blue) and imaged by confocal microscopy indicating the difference in cytoskeletal architecture in untreated cells (A) and cells treated with SFX-01 (B).
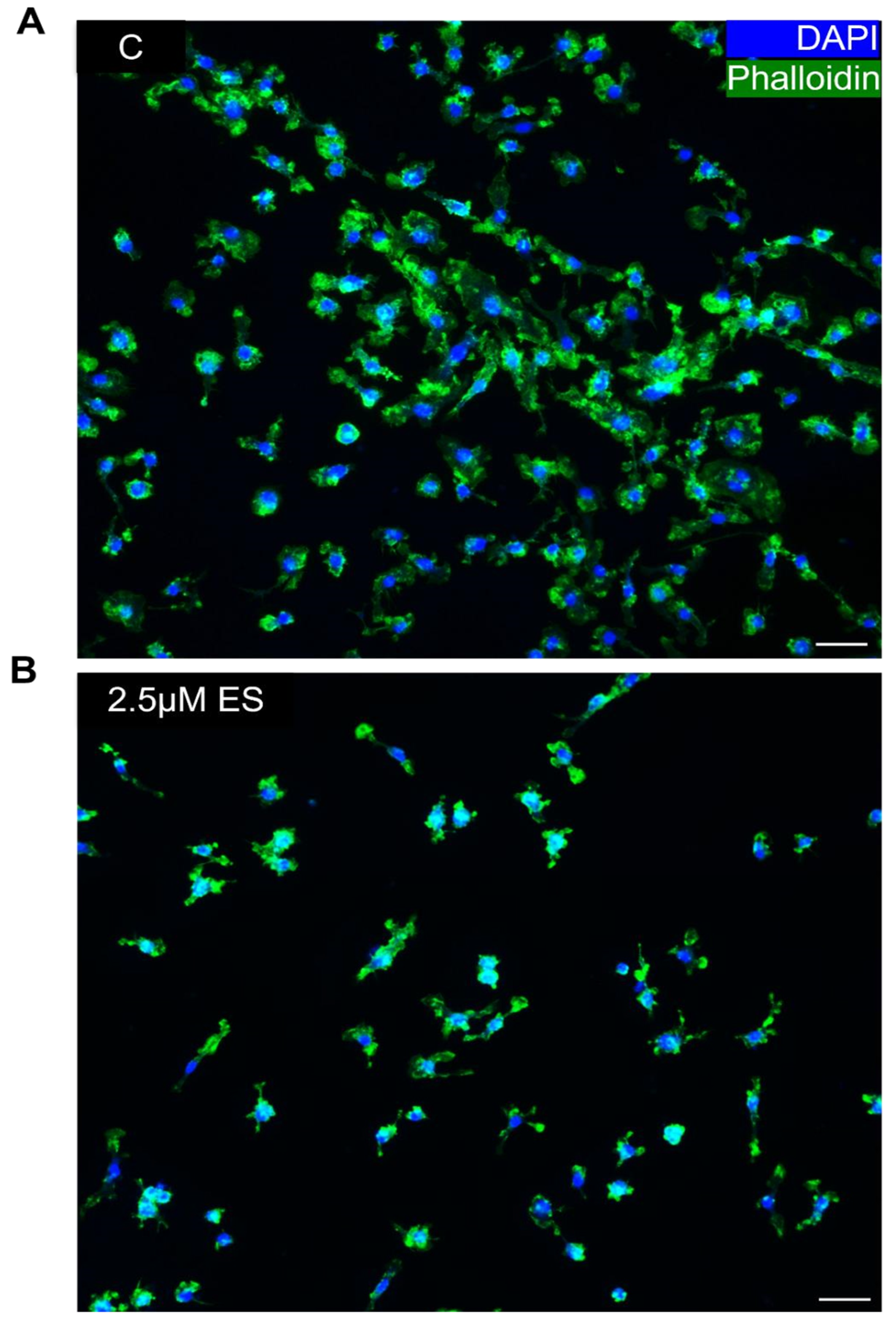
Figure 5.
Incubation with SFX-01 indicates inhibition of cell-cell fusion/multinucleation of osteoclasts grown on plastic. Osteoclasts cultured for 6 days and then immunocytochemically labelled for polymerized actin (phalloidin, green), DC-STAMP (red) and nuclear DNA (DAPI, blue). (AD) Control cultures showed positive labelling for cytoskeletal actin, cell spreading, cell fusion and multinucleation. These features were inhibited by the addition of SFX-01 at all timepoints, but particularly prominently at ES and DC (E-H, M-P). DS treatment likewise inhibited cell fusion (I-L) but less markedly.
Figure 5.
Incubation with SFX-01 indicates inhibition of cell-cell fusion/multinucleation of osteoclasts grown on plastic. Osteoclasts cultured for 6 days and then immunocytochemically labelled for polymerized actin (phalloidin, green), DC-STAMP (red) and nuclear DNA (DAPI, blue). (AD) Control cultures showed positive labelling for cytoskeletal actin, cell spreading, cell fusion and multinucleation. These features were inhibited by the addition of SFX-01 at all timepoints, but particularly prominently at ES and DC (E-H, M-P). DS treatment likewise inhibited cell fusion (I-L) but less markedly.
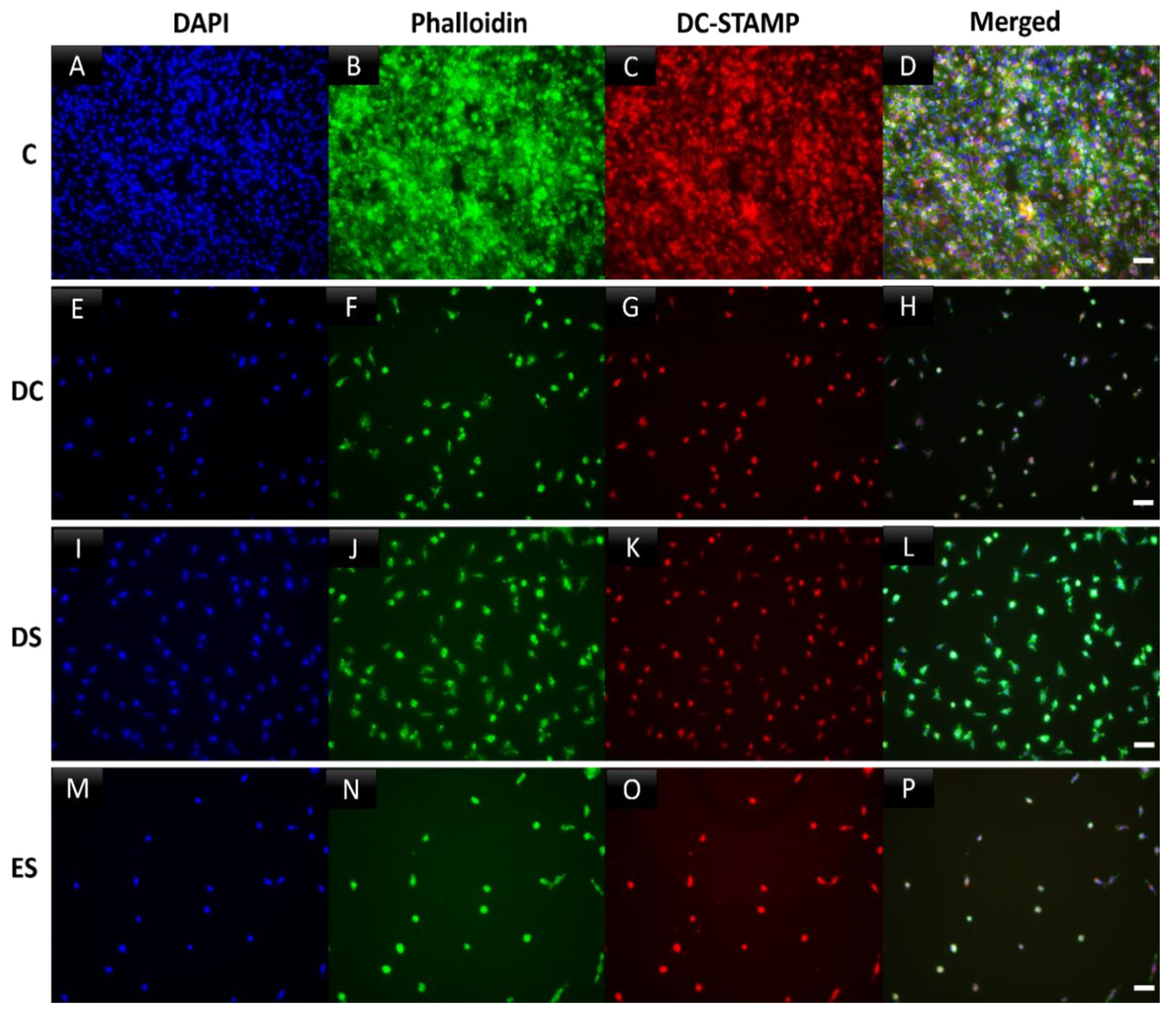
Figure 6.
Regulation of osteoclast behaviour by NRF2 and NF-KB modulators. RA839 (NRF2 activator) and BA (NF-KB activator) exert most marked inhibitory effects on osteoclast formation/resorption when added for duration of culture (7 days). TRAP+ osteoclast numbers, area resorbed, area resorbed/OC (mean± SEM). $ and $$ denote p ≤ 0.05 and p ≤ 0.01 difference from control. *denotes p ≤ 0.05 differences between 1μM and 10μΜ for each compound. Combined data from n=5 experiments, 6-8 replicates/condition. Open /closed circles and crosses denote BM cells from C57/Bl6 and squares CBA, respectively.
Figure 6.
Regulation of osteoclast behaviour by NRF2 and NF-KB modulators. RA839 (NRF2 activator) and BA (NF-KB activator) exert most marked inhibitory effects on osteoclast formation/resorption when added for duration of culture (7 days). TRAP+ osteoclast numbers, area resorbed, area resorbed/OC (mean± SEM). $ and $$ denote p ≤ 0.05 and p ≤ 0.01 difference from control. *denotes p ≤ 0.05 differences between 1μM and 10μΜ for each compound. Combined data from n=5 experiments, 6-8 replicates/condition. Open /closed circles and crosses denote BM cells from C57/Bl6 and squares CBA, respectively.
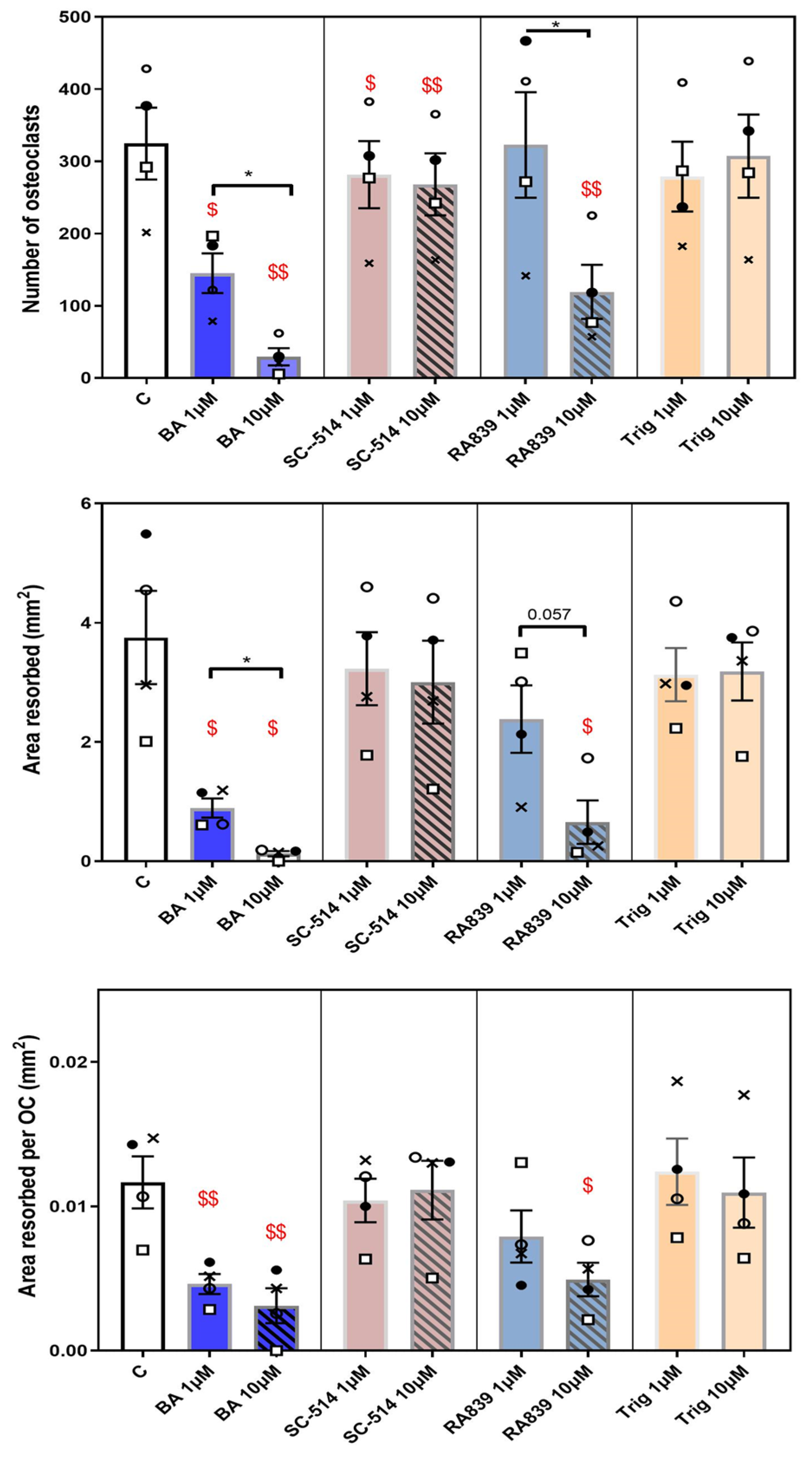
Figure 7.
Regulation of osteoclast behaviour by NRF2 and SFX-01. A) RA839 (NRF2 activator) inhibits osteoclast formation/resorption at early stage (ES; D1-D2) but there is no additional inhibition exerted when SFX-01 is co-administered; indicating that inhibition by SFX-01 is stronger than RA839. B) Trig (NRF2 inhibitor) also inhibits osteoclast formation/resorption at early stage (ES; D1-D2); no additional inhibition with SFX-01. TRAP+ osteoclast numbers (top), area resorbed (centre) and area resorbed/OC (bottom, mean± SE) from STR/Ort mice. $, $$ and $$$ denote p ≤ 0.05, p ≤ 0.01 and p≤ 0.001 from the control; * p ≤ 0.05, ***p<0.001 denote statistical difference between the different groups. Combined data from n=4 experiments, 6-8 replicates/condition.
Figure 7.
Regulation of osteoclast behaviour by NRF2 and SFX-01. A) RA839 (NRF2 activator) inhibits osteoclast formation/resorption at early stage (ES; D1-D2) but there is no additional inhibition exerted when SFX-01 is co-administered; indicating that inhibition by SFX-01 is stronger than RA839. B) Trig (NRF2 inhibitor) also inhibits osteoclast formation/resorption at early stage (ES; D1-D2); no additional inhibition with SFX-01. TRAP+ osteoclast numbers (top), area resorbed (centre) and area resorbed/OC (bottom, mean± SE) from STR/Ort mice. $, $$ and $$$ denote p ≤ 0.05, p ≤ 0.01 and p≤ 0.001 from the control; * p ≤ 0.05, ***p<0.001 denote statistical difference between the different groups. Combined data from n=4 experiments, 6-8 replicates/condition.
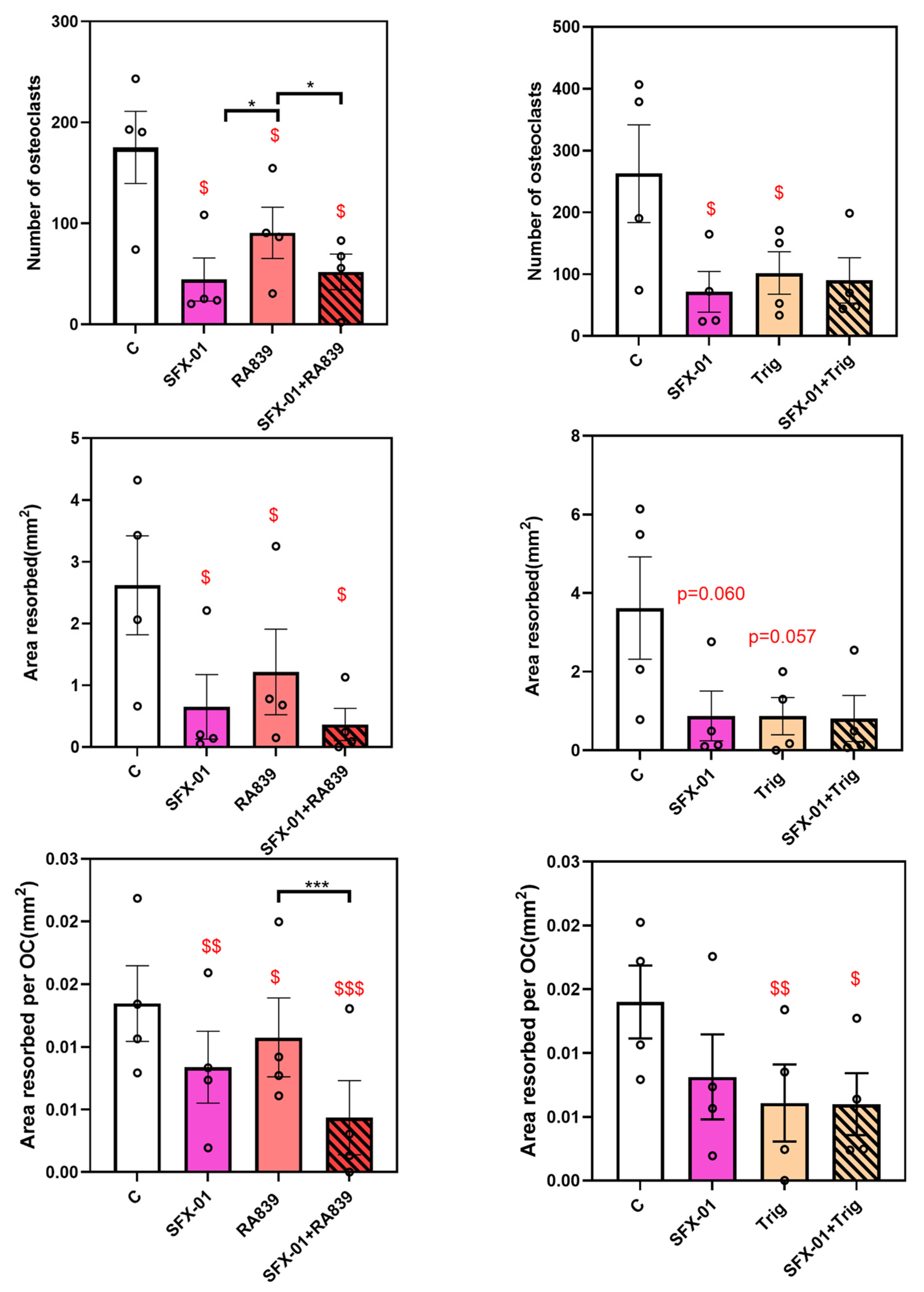
Figure 8.
Assay for the 4-HNE adduct indicates that cultures treated with SFX-01 (DS; D2-D5) expressed lower oxidative stress levels than the untreated cells.
Figure 8.
Assay for the 4-HNE adduct indicates that cultures treated with SFX-01 (DS; D2-D5) expressed lower oxidative stress levels than the untreated cells.
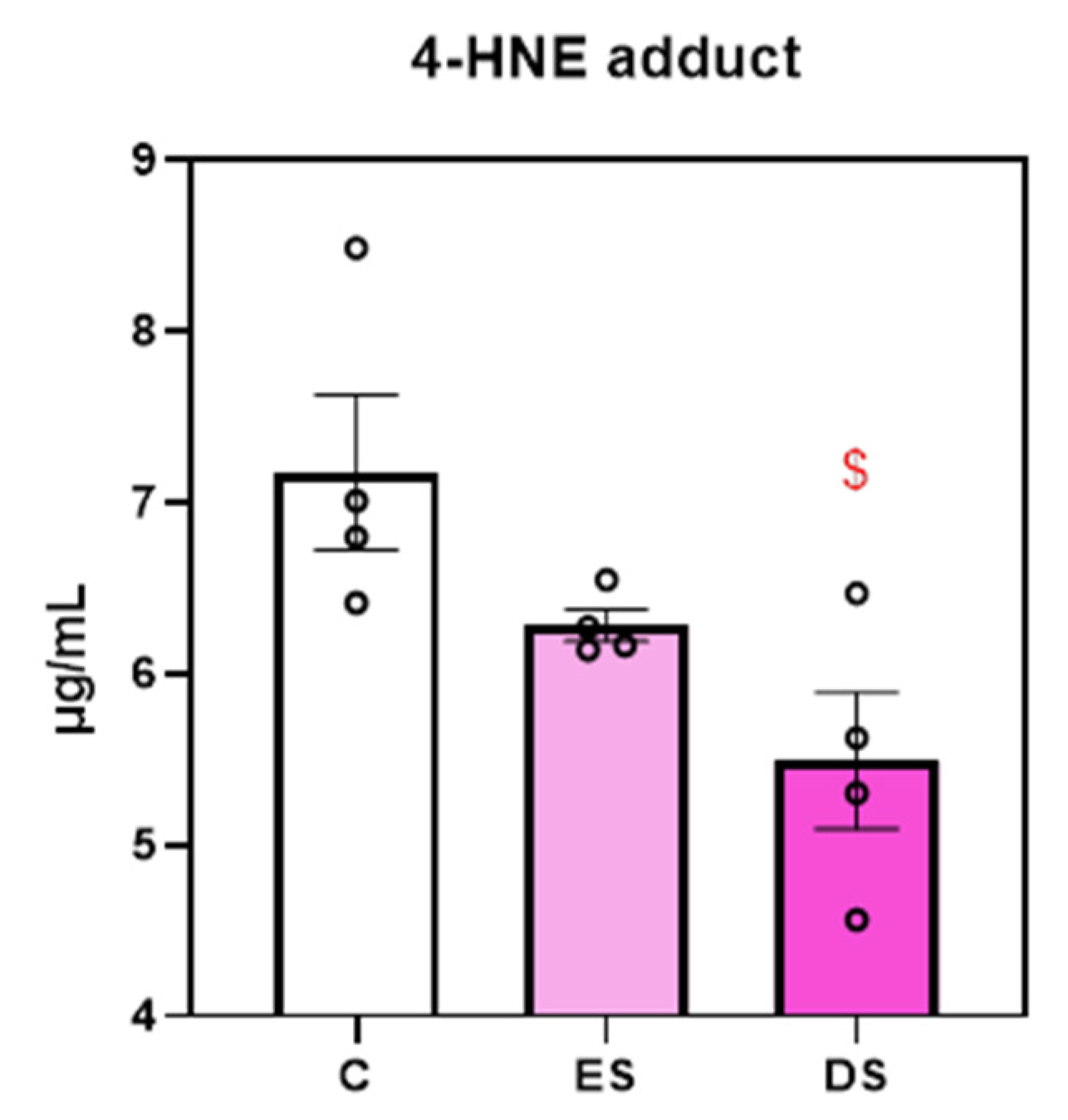
Figure 9.
Regulation of osteoclast behaviour by NF-KB and SFX-01. BA (NF-KB activator) inhibits osteoclast formation/resorption at early stage (ES; D1-D2); when supplemented with SFX-01 there was no additional inhibition greater than either with BA or SFX-01 alone. B) SC-514 (NF-KB inhibitor) did not modify osteoclast formation/resorption at early stage (ES; D1-D2) alone with SFX-01. TRAP+ osteoclast numbers (left), area resorbed (centre) and area resorbed/OC (right, mean± SE) from STR/Ort mice. $, $$ and $$$ denote p ≤ 0.05, p ≤ 0.01 and p≤ 0.001 from the control. * p ≤ 0.05, ***p<0.001 denote statistical difference between different groups.
Figure 9.
Regulation of osteoclast behaviour by NF-KB and SFX-01. BA (NF-KB activator) inhibits osteoclast formation/resorption at early stage (ES; D1-D2); when supplemented with SFX-01 there was no additional inhibition greater than either with BA or SFX-01 alone. B) SC-514 (NF-KB inhibitor) did not modify osteoclast formation/resorption at early stage (ES; D1-D2) alone with SFX-01. TRAP+ osteoclast numbers (left), area resorbed (centre) and area resorbed/OC (right, mean± SE) from STR/Ort mice. $, $$ and $$$ denote p ≤ 0.05, p ≤ 0.01 and p≤ 0.001 from the control. * p ≤ 0.05, ***p<0.001 denote statistical difference between different groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



