Submitted:
18 November 2023
Posted:
21 November 2023
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
When it is in the template RNA, the naturally occurring m1A epitranscriptomic RNA modification was recently reported to be able to stop the RNA polymerization reaction catalyzed by the RNA dependent RNA polymerase (RdRp) of SARS-CoV-2. In this report, we report that m1A via its triphosphate form (m1ATP) can be incorporated into RNA by the same RdRp. These two findings point a new direction for antiviral drug development based on m1A for combatting COVID-19. More broadly, it is possible that the large pool of epigenetic RNA as well as DNA modifications could serve as a treasury for drug discovery aimed at combating various infectious and other diseases.
Keywords:
SARS-CoV-2
; COVID-19
; RdRp
; m1A triphosphate
; Antiviral Drug
Introduction
Infectious diseases caused by RNA viruses such as HCoV 229E, HCoV OC43, HCV and HIV are notably annoying or difficult to treat. The recent COVID-19 pandemic caused by SARS-CoV-2 once again demonstrated this notion [1,2,3]. For all infectious diseases, one common feature is that different people show different symptoms, which range from asymptomatic to death. This observation has led to the invention of vaccines [4,5,6]. During the pandemic, driven by the curiosity to find additional causes for individualized symptoms of infectious diseases beyond those described in the literature such as induced immunity [7], we studied the effects of epitranscriptomic RNA modifications on the catalytic activity of SARS-CoV-2 RNA dependent RNA polymerase (RdRp) [8,9]. We reasoned that the epitranscriptomic systems of different individuals are not identical, and therefore they can modify the viral RNA genome differently. The difference in the modifications may affect viral life cycles by inhibiting or activating the RdRp, and as a result may contribute to the difference of clinical outcomes of COVID-19. Among other observations during the studies, we found that the m1A modification in an RNA template is capable to stop the RNA polymerization reaction catalyzed by SARS-CoV-2 RdRp [8]. With this finding, we were curious to know if m1A could be incorporated into RNA by the same RdRp via its triphosphate (m1ATP). If the answer were positive, prodrugs based on m1ATP and other modified nucleoside triphosphates could be developed to fight SARS-CoV-2 and other RNA viruses.
Results and Discussion
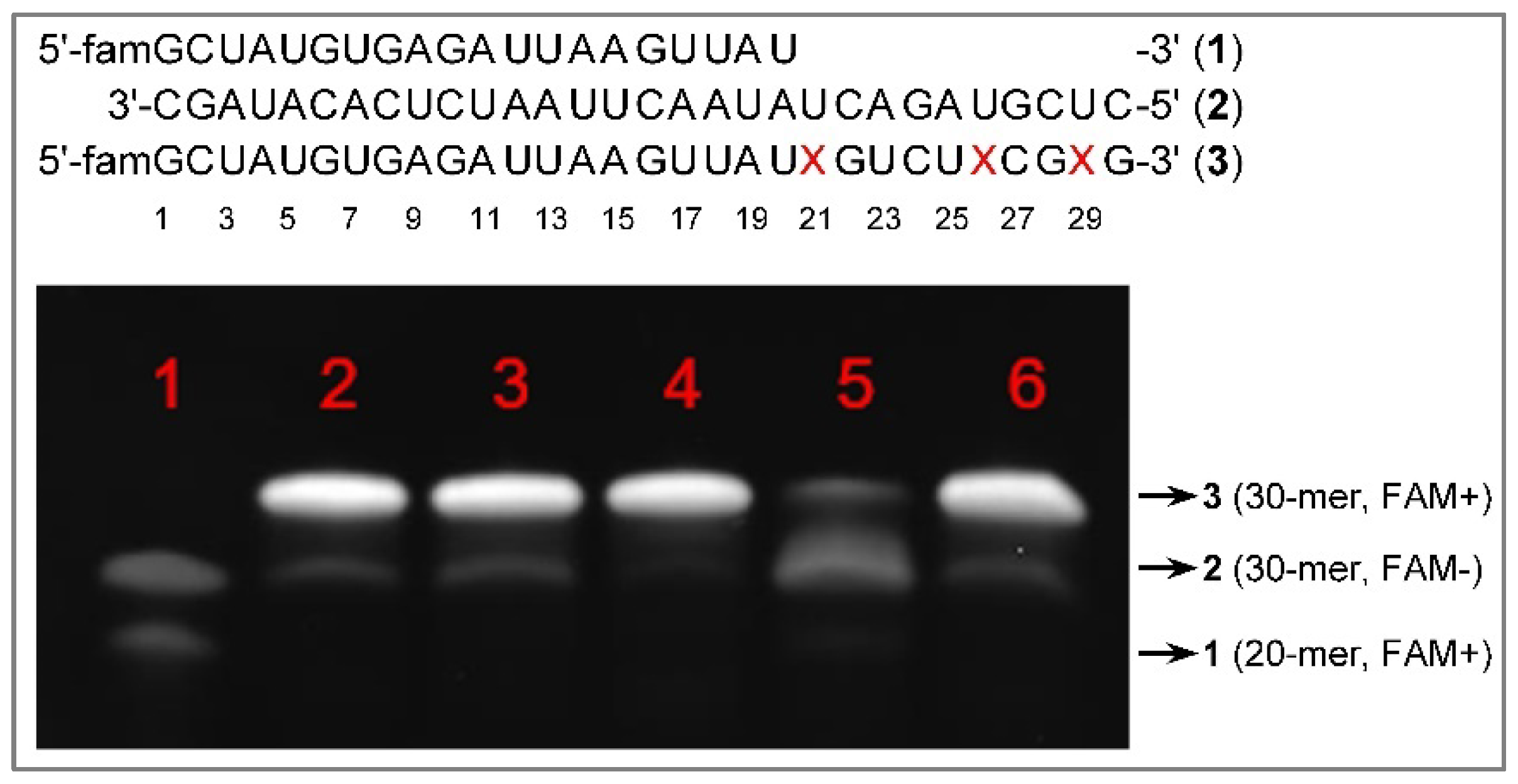
To test if m1A can be incorporated into RNA via m1ATP by SARS-CoV-2 RdRp, the 20-mer primer RNA (1), which has a FAM at its 5′-end, and the 30-mer template RNA (2) were synthesized. The preformed primer-template duplex (1-2) was subjected to various RNA polymerization conditions catalyzed by RdRp. The results were analyzed using denatured polyacrylamide gel electrophoresis (PAGE, Figure 1). Lane 1 is a control, and only the preformed primer-template duplex (1-2) was loaded. The two bands indicate that the duplex can be denatured and the gel can separate the primer and template. Lane 2 is also a control. The RNA polymerization reaction was carried out in the presence of all the four natural NTPs (ATP, CTP, GTP and UTP), and no m1ATP was added. As expected, the primer was all converted to the 30-mer RNA 3, where X is A. From this lane, we can also see that the 30-mer extended primer 3, which has a FAM, can be well separated from the 30-mer template 2, which does not have a FAM. Lane 3 is the result of m1A incorporation. As can be seen, when the RNA polymerization reaction was conducted in the presence of m1ATP, CTP, GTP and UTP, even though there was no ATP, the reaction proceeded smoothly. This indicates that m1A can be incorporated into RNA via m1ATP by SARS-CoV-2 RdRp.
To see if the RNA polymerization reaction could still proceed in the absence of both m1ATP and ATP, the reaction was conducted in the presence of only CTP, GTP and UTP (lane 5). As can be seen, the reaction did not go well. Only a small portion of the primer was extended to the 30-mer 3, which might also be the 28-mer RNA resulted from stopping the reaction at the last U (position 29) in 2 because there is a slight chance that the gel could not resolve 28-mer and 30-mer. The majority of the primer was only partially extended, and the reaction might have stopped at any of the three U positions in the single-stranded region of the primer-template duplex. Careful examination of the gel image can also reveal that a small portion of the primer remained unreacted, but this may not be easy to see in Figure 1 as the band is very light. The results in this lane (lane 5) indicate that either m1ATP or ATP is needed for the reaction to pass the positions occupied by U in the single stranded region of template 2 at the efficiency observed in experiments related to lanes 2 and 3. This further confirms that m1A can be incorporated into RNA via m1ATP by SARS-CoV-2 RdRp. The close to disappearance of primer and the appearance of smear in the region between 1 and 3 in lane 5 is consistent with the low fidelity of the RdRp as reported in the literature [10,11]. Experiments related to lanes 4 and 6 were designed and conducted before we knew that m1ATP could be an effective substrate of the RdRp. Indeed, we thought that the chance for the RdRp to incorporate m1A into RNA via m1ATP were low considering that m1A in template RNA inhibited the RdRp. In case that were the case, the experiments related to lanes 4 and 6 would determine if m1ATP could inhibit the incorporation of ATP by RdRp. Now that m1A can be incorporated, these experiments are irrelevant.
From the above experiments and the data we published earlier regarding inhibition of SARS-CoV-2 RdRp by m1A in RNA template [8], it is clear that m1ATP based prodrugs may be investigated for the treatment of COVID-19. The reason is that once m1ATP is formed from a prodrug in SARS-CoV-2 infected cells, the viral RdRp could incorporate m1A into its RNA genome. Once m1A is in the RNA genome, replication of genome by the viral RdRp would be stopped, and the viral life cycle would be interrupted. It is noted that for this drugging mechanism to function, there is no need for all A in the viral RNA genome to become m1A. Perhaps, only a few or even only one A in a key position is needed to be replaced by m1A to stop the functioning of the entire viral genome.
Besides m1A, there are over 300 epitranscriptomic RNA modifications [12,13,14]. Among them, we have found that m3C in RNA template does not inhibit SARS-CoV-2 RdRp [8], which was surprising considering that m1A inhibits the RdRp and both m1A and m3C disrupt canonical base pairing. However, even though m3C does not inhibit RdRp, it is unlikely that the nucleotide incorporated across it is precisely G due to G-C base pair disruption by the modification. Therefore, if m3CTP could be an effective substrate of RdRp, m3CTP based prodrugs, which would function through random mutagenesis like favipiravir [15,16], could also be studied. Besides m1A and m3C, many of the other epitranscriptomic modifications could be studied for similar purposes [17].
In addition to SARS-CoV-2, other RNA viruses such as HCoV 229E, HCoV OC43, HCV and HIV [18,19] could also be considered to be combatted using prodrugs based on m1A and other epitranscriptomic RNA modifications in a similar manner. Of course, the concept can also go beyond RNA viruses and extend to DNA viruses and even non-infectious diseases. Indeed, the large pool of epitranscriptomic RNA modifications and their variations could become a treasury for drug discovery. Importantly, drugs based on these modifications could likely have less adverse effect for reasons such as lower chance to generate toxic metabolites [20].
Conclusions
In conclusion, we found that m1A can be incorporated into RNA via m1ATP by SARS-CoV-2 RdRp. Earlier, we reported that m1A in RNA template could inhibit the same RdRp [8]. From these two discoveries, opportunities appear that m1A based prodrugs could be developed to combat SARS-CoV-2. Using a similar approach, the development of prodrugs based on other epitranscriptomic RNA modifications to fight SARS-CoV-2 and other viruses could be considered. Further investigation of the possibility of using m1A based prodrugs to combat COVID-19 under more biologically relevant conditions as well as using prodrugs based on other RNA modifications for treating COVID-19 and other infectious diseases are being pursued.
Supporting Information
Experimental details for SARS-CoV-2 RdRp catalyzed RNA polymerization reaction.
Acknowledgment
Financial support from NIH (GM109288), NSF (1954041), Robert and Kathleen Lane Endowed Fellowship (A.A.), and HRI Fellowship (A.A.); the assistance from D.W. Seppala (electronics), J.L. Lutz (NMR), and A. Galerneau (MS); and NSF equipment grants (2117318, NMR; 1048655 & 1531454, MS); are gratefully acknowledged.
Conflicts of Interest
Michigan Tech has interest in IP related to this report.
References
- Alsulimani, A.; Bhardwaj, T.; Janahi, E. M.; Almalki, A. H.; Tewari, B. N.; Wahid, M.; Alkhanani, M. F.; Somvanshi, P.; Haque, S. Systematic structure guided clustering of chemical lead compounds targeting RdRp of SARS-CoV-2. Minerva Biotechnology and Biomolecular Research 2022, 34, 114–121. [Google Scholar] [CrossRef]
- Wang, Y. Y.; Anirudhan, V.; Du, R. K.; Cui, Q. H.; Rong, L. J. RNA-dependent RNA polymerase of SARS-CoV-2 as a therapeutic target. Journal of Medical Virology 2021, 93, 300–310. [Google Scholar] [CrossRef]
- Min, J. S.; Kwon, S.; Jin, Y. H. SARS-CoV-2 RdRp inhibitors selected from a cell-based SARS-CoV-2 RdRp activity assay system. Biomedicines 2021, 9, 996. [Google Scholar] [CrossRef]
- Moradpour, J.; Chit, A.; Besada-Lombana, S.; Grootendorst, P. Overview of the global vaccine ecosystem. Expert Review of Vaccines 2023, 22, 749–763. [Google Scholar] [CrossRef]
- Fox, N.; Adams, P.; Grainger, D.; Herz, J.; Austin, C. The value of vaccines: A tale of two parts. Vaccines (Basel) 2022, 10, 2057. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, J. P. The value of vaccines. Current Opinion in Immunology 2022, 78, 102243. [Google Scholar] [CrossRef] [PubMed]
- McCoy, K.; Peterson, A.; Tian, Y.; Sang, Y. M. Immunogenetic association underlying severe COVID-19. Vaccines (Basel) 2020, 8, 700. [Google Scholar] [CrossRef] [PubMed]
- Apostle, A.; Yin, Y.; Chillar, K.; Eriyagama, A.; Arneson, R.; Burke, E.; Fang, S.; Yuan, Y. Effects of epitranscriptomic RNA modifications on the catalytic activity of the SARS-CoV-2 replication complex. ChemBioChem 2023, 24, e202300095. [Google Scholar] [CrossRef] [PubMed]
- Apostle, A.; Yin, Y.; Chillar, K.; Eriyagama, A.; Arneson, R.; Burke, E.; Fang, S. Effects of epitranscriptomic RNA modifications on the catalytic activity of SARS-CoV-2 replication complex. ChemRxiv [Preprint] 2023. [Google Scholar] [CrossRef] [PubMed]
- Yin, X. Y.; Popa, H.; Stapon, A.; Bouda, E.; Garcia-Diaz, M. Fidelity of ribonucleotide incorporation by the SARS-CoV-2 replication complex. Journal of Molecular Biology 2023, 435, 167973. [Google Scholar] [CrossRef]
- Moeller, N. H.; Shi, K.; Demir, Ö.; Belica, C.; Banerjee, S.; Yin, L. L.; Durfee, C.; Amaro, R. E.; Aihara, H. Structure and dynamics of SARS-CoV-2 proofreading exoribonuclease exon. Proceedings of the National Academy of Sciences of the United States of America 2022, 119, e2106379119. [Google Scholar] [CrossRef] [PubMed]
- Modomics - a database of RNA modifications. Available online: https://genesilico.pl/modomics/.
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nature Reviews Neuroscience 2020, 21, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Mersinoglu, B.; Cristinelli, S.; Ciuffi, A. The impact of epitranscriptomics on antiviral innate immunity. Viruses-Basel 2022, 14, 1666. [Google Scholar] [CrossRef]
- Shannon, A.; Selisko, B.; Le, N. T.; Huchting, J.; Touret, F.; Piorkowski, G.; Fattorini, V.; Ferron, F.; Decroly, E.; Meier, C.; Coutard, B.; Peersen, O.; Canard, B. Rapid incorporation of favipiravir by the fast and permissive viral RNA polymerase complex results in SARS-CoV-2 lethal mutagenesis. Nature Communications 2020, 11, 4682. [Google Scholar] [CrossRef] [PubMed]
- Hassanipour, S.; Arab-Zozani, M.; Amani, B.; Heidarzad, F.; Fathalipour, M.; Martinez-de-Hoyo, R. The efficacy and safety of favipiravir in treatment of COVID-19: A systematic review and meta-analysis of clinical trials. Scientific Reports 2021, 11, 11022. [Google Scholar] [CrossRef] [PubMed]
- Petushkov, I.; Esyunina, D.; Kulbachinskiy, A. Effects of natural RNA modifications on the activity of SARS-CoV-2 RNA-dependent RNA polymerase. FEBS Journal 2023, 290, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Blanco, M. A.; Ooi, E. E.; Sessions, O. M. RNA viruses, pandemics and anticipatory preparedness. Viruses-Basel 2022, 14, 2176. [Google Scholar] [CrossRef] [PubMed]
- Cross, S. T.; Michalski, D.; Miller, M. R.; Wilusz, J. RNA regulatory processes in RNA virus biology. Wiley Interdisciplinary Reviews-RNA 2019, 10, e1536. [Google Scholar] [CrossRef] [PubMed]
- Jockusch, S.; Tao, C. J.; Li, X. X.; Anderson, T. K.; Chien, M. C.; Kumar, S.; Russo, J. J.; Kirchdoerfer, R. N.; Ju, J. Y. A library of nucleotide analogues terminate RNA synthesis catalyzed by polymerases of coronaviruses that cause SARS and COVID-19. Antiviral Research 2020, 180, 104857. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Image of denatured PAGE for the analysis of SARS-CoV-2 RdRp catalyzed RNA polymerization reactions. X in 3 is the nucleoside A or m1A depending on the NTP used. The mixture of RdRp, primer-template duplex 1-2 and NTPs was incubated at 37 oC for 2 h. The reaction products were analyzed with PAGE. The image was taken after staining with GelRed. Lane 1: RNA duplex only. Lane 2: ATP, CTP, GTP and UTP, no m1ATP. Lane 3: m1ATP, CTP, GTP and UTP, no ATP. Result indicates that m1A can be incorporated into RNA via m1ATP. Lane 4: m1ATP only for 15 min, then ATP, CTP, GTP and UTP. Results indicate m1ATP does not inhibit RdRp. Lane 5: CTP, GTP, UTP, no ATP, no m1ATP. Results indicate that ATP and m1ATP in other lanes were needed.
Figure 1.
Image of denatured PAGE for the analysis of SARS-CoV-2 RdRp catalyzed RNA polymerization reactions. X in 3 is the nucleoside A or m1A depending on the NTP used. The mixture of RdRp, primer-template duplex 1-2 and NTPs was incubated at 37 oC for 2 h. The reaction products were analyzed with PAGE. The image was taken after staining with GelRed. Lane 1: RNA duplex only. Lane 2: ATP, CTP, GTP and UTP, no m1ATP. Lane 3: m1ATP, CTP, GTP and UTP, no ATP. Result indicates that m1A can be incorporated into RNA via m1ATP. Lane 4: m1ATP only for 15 min, then ATP, CTP, GTP and UTP. Results indicate m1ATP does not inhibit RdRp. Lane 5: CTP, GTP, UTP, no ATP, no m1ATP. Results indicate that ATP and m1ATP in other lanes were needed.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



