
Submitted:
20 November 2023
Posted:
21 November 2023
You are already at the latest version
Abstract
Celtis sinensis is an adaptable species that is widely grown in southern China. In June 2022, a leaf blotch disease of C. sinensis was observed in Nanjing, Jiangsu, China. Based on morphological characterization, three isolates were identified as Alternaria species. Phylogenetic analysis of combined ITS, GAPDH, TEF1-α, RPB2 and Alt a 1 sequences identified the three isolates we obtained as Alternaria alternata and Alternaria koreana. Koch’s postulates were fulfilled in the greenhouse, and the pathogenicity of A. alternata and A. koreana was determined by leaf inoculation tests on C. sinensis seedlings. The symptoms of indoor inoculation were consistent with those in the field. A. alternata and A. koreana can grow at 15-35 °C, with an optimal growth temperature of 25 °C. The results of fungicide sensitivity experiments indicated that A. alternata and A. koreana were the most sensitive to prochloraz, which may be a useful strategy for the future prevention and control management of A. alternata and A. koreana. This study provides the first step for further research on A. alternata and A. koreana as pathogens of C. sinensis and provides a theoretical basis for future control strategies.
Keywords:
Celtis sinensis
; Alternaria alternata
; Alternaria koreana
; fungi
; identification
; fungicides
1. Introduction
Celtis sinensis is an important member of the Ulmaceae family [1] that is native to China, Japan, and Korea [2]. Mainly distributed in the central and eastern regions of China, with straight trunks and beautiful tree shapes, it is a common street and landscaping tree species and a valuable resistant tree species that can adsorb toxic gases such as sulfur dioxide and chlorine and has a certain degree of adsorption capacity for dust [3]. C. sinensis demonstrates remarkable resilience, as it is able to withstand both dry conditions and moist, infertile soils. These trees possess a strong adaptability to diverse environmental factors. Moreover, the branches, leaves, roots, and bark of C. sinensis hold important medicinal value and can be utilized in the treatment of burns and urticaria [4]. In addition, it has a wide range of industrial uses. The roots and branches can be used to make artificial cotton and furniture, and the fruits can also be used as raw materials for lubricant production [5]. However, in Xuanwu District, Nanjing, Jiangsu Province, a large number of leaf spot and leaf curling occurred in C. sinensis, which seriously affected the ornamental value of C. sinensis and damaged the ecological environment.
Alternaria is a widely distributed group of fungi in the natural environment and is also economically important as a dematiaceous fungus [6]. More than 95% of the species are plant parthenogenetic parasites. They can grow and multiply in low-temperature and humid environments, causing a variety of plant diseases, as well as fruit rot, which severely jeopardizes the growth of crops and plants and causes enormous economic losses while destroying natural ecosystems globally [7]. In addition, the spores of Alternaria can also release allergens [8], triggering diseases such as asthma [9] and posing a major threat to human health. The pathogenic fungi of Alternaria establish their population by colonizing the surface of seeds with mycelium or conidia [10] on diseased plant residues and in the soil. These serve as primary infection sources for the following year. The conidia of the fungi are spread and dispersed by air currents, leading to secondary infections, thus perpetuating the cycle of transmission and spread of disease.
At present, the most direct and effective method for the control of diseases caused by Alternaria is chemical control [11]. Fungicides such as dithiocarbamates, triazoles, strobilurins, iprodione, or copper fungicides are used in most areas for disease control [12]. Copper-based fungicides are widely used due to their long residual effectiveness and resistance to washing [13]. Methoxyacrylate, dimethomorph, and mancozeb fungicides have also been registered for the control of brown spot disease in countries such as the United States, Israel, and Spain [14]. Due to the difficulty of mixing copper-based formulations with other pesticides and the single-site action of methoxyacrylate and dimethomorph fungicides, resistant populations can quickly emerge in the field, leading to decreased or ineffective efficacy of these fungicides. Recently, nicotinamide fungicides such as cyazofamid have been registered in the United States for the control of this disease [15]. These fungicides can be used for preseed disinfection, as well as direct application to the soil, effectively providing preventive protection [16].
This study reveals the characteristics of the pathogen related to leaf spot in C. sinensis in Xuanwu District, Nanjing, Jiangsu Province, China. This disease has caused significant damage to the local ecosystem. This study was conducted through the following methodologies: to determine the causative agent of brown spot disease in C. sinensis using Koch's postulates; to identify the pathogen through molecular biological and morphological identification methods; and to screen fungicides with significant inhibitory effects on the mycelial growth of the pathogen through culture-based plate phenotype experiments.
2. Materials and Methods
2.1. Sampling and fungal isolations
In June 2022, the disease was observed on the leaves of C. sinensis on the campus of Nanjing Forestry University, Nanjing City, Jiangsu Province (119°46'43"E, 32°02'38"N), and 20 diseased leaves were collected as samples from across 0.75-1.5 meters of C. sinensis. The samples were collected and returned to the laboratory, and the diseased leaves were wiped with absorbent paper with sterile water to remove surface dust and impurities. One hundred small tissues (3×3 mm2) were cut from lesion margins and surface-sterilized in 75% ethanol for 30 s, followed by 5% NaClO for 90 s, rinsed in sterile water 3 times, dried on sterilized filter paper, plated onto potato dextrose agar (PDA) for 4 days at 25 °C [17]. On the third day, fungal hyphae emerging from leaf tissue were shifted to fresh PDA [18]. The isolated strains were preliminarily classified according to morphological characteristics and ITS sequence alignment for subsequent experiments.
2.2. Pathogenicity tests
Healthy C. sinensis and seedlings were obtained from the laboratory of Nanjing Forestry University, and the seedlings had heights of approximately 80 cm. Before the pathogenicity experiment, the leaf surface was wiped with 75% alcohol twice, and sprayed with sterile water three times. The water stains on the surface of the leaves were wiped dry with absorbent paper. The colony morphology of 100 fungal samples was assessed, and they were divided into different groups. Three isolates (11, 12 and 13) from the group with the highest frequency of occurrence were selected for pathogenicity testing and inoculation of leaf blades and seedlings, respectively [19]. Five isolated leaves were inoculated with each isolate, wounds were created on both sides of the leaf veins with a sterile needle, and a 6 mm mycelium block was placed face down on each puncture wound and removed after 24 hours [20]. In addition, five leaves were inoculated with sterilized PDA agar blocks as a control. The samples were cultured for four days under environmental conditions of 25 °C temperature, 70%-80% humidity, and 24 hours of light exposure per day. Nine healthy C. sinensis seedlings were selected on each plant, 3 leaves were inoculated with spores of one of the 3 isolates by puncturing the leaf with a sterile needle as a wound suspension (106 conidia/mL), and the other three plantlets were inoculated with sterile water as a control group. The plantlets were covered with plastic bags and sprayed with water to maintain a high level of humidity.
2.3. Morphological identification and biological characteristics
Three isolates (11, 12 and 13) were cultured on PDA medium at 25 °C for 7 days in a constant temperature incubator. Morphological identification relied on colony appearance and conidia characteristics. Morphology of conidia using Zeiss Axio Imager A2m microscope (Carl Zeiss, Germany) (n=30).
Three isolates (11, 12, and 13) were chosen for the purpose of conducting morphological observations and biological characterization. Isolates were cultured on potato dextrose agar medium (PDA), Czapek dox agar medium (CDA), Richard medium (Richard), corn meal agar medium (CMA), oatmeal agar medium (OA), and potato saccharose agar medium (PSA) for 7 days at 25 °C, and the appearance and coloration of the colonies were documented.
In order to ascertain the isolates' optimal growth temperature, mycelial plugs with a diameter of 6 mm were positioned on freshly prepared PDA medium, which had a diameter of 90 mm, and incubated at 5 °C, 15 °C, 20 °C, 25 °C, 30 °C, and 35 °C. The colony growth diameter was measured and recorded daily.
2.4. DNA extraction, PCR amplification, and sequencing
The isolated fungi were cultured on PDA medium for 7 days, and mature mycelium was scraped off using a sterilized surgical blade. DNA extraction was carried out utilizing the CTAB method [21]. All DNA extracts were preserved at a temperature of -20 °C for future utilization.
Polymerase chain reaction was performed on ITS [22], TEF1-α [23], GAPDH [24], RPB2 [25], and Alt a 1 [26] five genes, which were sequenced using primers ITS1/4, EF1-728F/EF1-986R, GPD1/GPD2, RPB2-5F2/RPB2-7C and Alt-for/Alt-rev, respectively. PCR was carried out in a 25 µL reaction mixture containing 21 μL of Tmix, 1 μL of DNA template, 1 μL of Taq DNA polymerase, 1 μL of forward primer and 1 μL of reverse primer (Table 1). The PCR products were sequenced at Sangon Biotech Co. Ltd. (Nanjing, China).
2.5. Phylogenetic analyses
The phylogenetic tree was constructed using sequences from the fungal cultures in this study and sequences related to Alternaria sp. in GenBank (Table 2); Alternaria alternantherae was used as the outgroup. Sequence alignment analysis using MAFFT version 7 (https://mafft.cbrc.jp/alignment/software/) [27], to trim sequences to ensure a high degree of sequence alignment [28]. Phylogenetic analysis through concatenation of five loci (ITS, TEF1-α, GAPDH, RPB2, and Alt a 1). Phylogenetic trees based on combined genes were constructed using two independent optimality search criteria: maximum likelihood (ML) and Bayesian inference (BI) criteria. The ML analysis used the GTR + F + I + G4 model, and branch stability was determined by 1,000 bootstrap replicates. BI analysis used the GTR + I + G + F model, including two parallel runs of 2,000,000 generations. The resulting trees were plotted using FigTree v.1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/).
2.6. Evaluation of fungicides against A. alternata and A. koreana
Fungicides were kindly provided by Mrs. Tingting Dai of Nanjing Forestry University as 96% prochloraz [29], 97% myclobutanil [30], 97% tebuconazole [31], and 98% pyraclostrobin [32]. Each fungicide was prepared as a stock solution, which was then diluted to different concentrations. The solutions were added to sterilized PDA medium at a rate of 2% to create agar plates containing the respective concentrations of the fungicide. PDA plates without the use of any fungicide were used as a control. A fungal plug was extracted from the colony's edge using a 6mm punch and relocated to the center of the medium. After incubation at 25 °C for 7 days, the diameter of the fungal mycelium was measured. The EC50 value was calculated using GraphPad Prism 8.
3. Results
3.1. Field observations and fungal isolation
The survey results indicated that nearly 50% of the C. sinensis in Xuanwu District, Nanjing, have shown signs of disease. Most of the spots were produced on the apex part of the leaves (Figure 1A), and the spots were brown with darker margins and round to irregular in shape (Figure 1B), on which dark brown chains of conidia were visible. Over time, the spots gradually expand, and the leaves become dark brown, dry, curled, and necrotic and eventually fall off. In total, 90 fungal colonies developed from the tissue pieces and were grouped into three types according to their colony characteristics, with frequencies of 84.4% (76 of 90), 10% (9 of 90), and 5.5% (5 of 90). Three isolates from the highest frequency were selected for purification. According to ITS sequences, 3 fungi belonged to Alternaria and were used for further research.
3.2. Pathogenicity test
Three representative isolates (11, 12 and 13) were inoculated on healthy leaves, and black spots were observed on the leaves after 4 days (Figure 2). The induced symptoms matched those observed in the wild. Conversely, the control maintained healthy with no symptoms of disease. Observing the size of the lesions, those caused by isolates 11 and 13 had smaller affected areas. In contrast, the lesions caused by isolate 12 exhibited a larger affected area. The isolates obtained from the inoculated diseased leaves displayed similar morphological characteristics and shape to the originally isolated strains. DNA was extracted and subjected to PCR using the primer ITS1/4, and the obtained sequences were similar to the original ones. The symptoms observed on seedlings inoculated in a controlled environment closely resembled those seen in the natural field conditions, thus meeting the criteria of Koch’s postulates. Therefore, A. alternata and A. koreana are the pathogens of leaf spot disease of C. sinensis.
3.3. Morphological characteristics
Three representative isolates were grown on PDA medium and hatched for 7 days at 25 °C, and morphological characteristics were recorded based on visual observations (Figure 3). The colonies of isolates 11 and 13 on PDA were initially gray and became grayish brown over time. Conidiophores produced numerous conidia in long chains, mostly unbranched. Conidia were obclavate, dark brown, with two to five transverse and zero to two longitudinal or oblique septa, and measured 10.1 to 35.9 × 7.6 to 24.3 μm (n=30). The colony of isolate 12 on PDA was grayish-green. Hyaline (young) hyphae or brown (old) hyphae, with septa and smooth walls, were slightly constricted at the septa and branched. Conidiophores produced numerous conidia in long chains. These conidia were globose to ellipsoidal and 13.5 to 34.5 × 7.0 to 14.1 μm (n = 30). The morphological characteristics of the 3 isolates matched those of Alternaria spp. The three isolates showed different morphological characteristics on different media.
3.4. Multigene phylogenetic analysis
BLAST results showed that the ITS, GAPDH, TEF1-α, RPB2 and Alt a 1 sequences of isolate 12 were highly similar (≥98%) to those of A. koreana (culture ex-type SPL2-1), while sequences of isolates 11 and 13 were highly similar (≥99%) to those of A. alternata (culture representative CBS 918.96). The sequences of genes/region ITS, GAPDH, TEF1-α, RPB2 and Alt a 1 from the three isolates (11, 12 and 13) were deposited in GenBank, and the accession numbers are shown in Table 2. Phylogenetic analyses using concatenated sequences placed isolate 12 in the clade of A. koreana and isolates 11 and 13 in the clade of A. alternata. Based on the phylogeny and morphology, isolate 12 was identified as A. koreana, and isolates 11 and 13 were identified as A. alternata (Figure 4).
3.5. Biological characteristics
The colonies of isolates 11 and 13 on CDA appeared as yellowish colonies with flattened colonies (Figure 5B, N). On Richard's medium, the colony appeared grayish-white, with very few hyphae and weak growth (Figure 5C, O). The colony of isolate 11 was dark green with dense mycelium on CMA and OA media, and on PSA media, the colony was dark gray in the middle with a brown ring around it with abundant mycelium (Figure 5D, E). The colony of isolate 13 was light green and radial on CMA, OA and PSA media (Figure 5P, Q, R). The colony of isolate 12 on CDA was almost identical to the colony morphology on PDA and PSA, with grayish-white colonies with dense hyphae (Figure 5G, H, L), showing a brownish color on Richard, with reticulated colonies and fewer hyphae (Figure 4I), a whitish-green color on CMA, with loosely packed colonies (Figure 5J), and a dark green color on PSA medium, with fuzzy colonies (Figure 5K).
Isolates 11, 12, and 13 were cultured between 5°C-35°C for 7 days, and the colony diameter reached a maximum and grew most vigorously at 25°C, but at 5°C, the mycelium hardly grew, which shows that 25°C is the optimum culture temperature for isolates 11, 12, and 13 (Figure 6).
3.6. Susceptibility of Alternaria isolates to fungicides
The three isolates showed similar biological responses to the four fungicides (Figure 7). All four fungicides showed significant growth inhibition of the isolates on PDA media. Prochloraz had a lower EC50 on mycelial growth than the other three fungicides and showed the best inhibitory effect on isolates 11, 12, and 13 (Table 3). Pyraclostrobin had the highest EC50 on mycelial growth of 13 isolates and the weakest inhibitory effect but still had a strong inhibitory effect on 11 and 12 isolates. Based on the EC50 values, myclobutanil inhibited isolates 11 and 13 almost equally but more strongly against isolate 12. Isolate 12 was the most sensitive to tebuconazole compared to 11 and 13, and isolate 13 was the least sensitive. The results indicated that prochloraz was the most effective fungicide against Alternaria spp. in this study.
4. Discussion
Celtis sinensis is a common deciduous tree that not only has some value in food and medicine but also plays an important role in urban greening and the economy. However, leaf spot disease deteriorates the appearance of leaves and severely affects the ornamental value of C. sinensis. This disease also leads to leaf damage and interferes with photosynthesis, which prevents C. sinensis from fully utilizing sunlight for nutrient synthesis, resulting in slow growth of the plant and weakening of the leaves, which in turn affects the health and growth of the whole tree and reduces the quality of the timber [33], resulting in an economic loss to the timber industry, as well as affecting the ecological function of C. sinensis. Timely identification of the pathogen responsible for C. sinensis leaf spot is essential. By combining morphological identification, molecular analysis, and phylogenetic research [34], A. alternata and A. koreana were identified as the causal agents of C. sinensis leaf spot in China. To our knowledge, this is the first report of A. alternata and A. koreana causing leaf spot in C. sinensis.
The majority of pathogenic species within the Alternaria genus are the primary cause of the most detrimental plant diseases, commonly referred to as “leaf spot” or “leaf blight”. These diseases are characterized by the development of circular or irregular necrotic lesions on the surface of the leaf, accompanied by a distinct concentric ring-like pattern at the center of the lesions. It is difficult to classify species of Alternaria based on spore morphology characteristics. The challenge arises because a considerable number of small-spore Alternaria species, commonly referred to as A. alternata, exhibit closely similar morphological characteristics [35]. Currently, there is very little research available on A. koreana species. The spore size observed in this study closely resembles the findings reported by Romain et al. [36]. However, there is actually a large amount of research available on Alternaria spp., and there is considerable variation in spore sizes reported. The spore sizes observed in this study are notably different from those described by Ramirez et al. [37], but they are similar to the description by Sun et al. [38]. Due to its morphological characteristics being susceptible to environmental conditions and the presence of many uncontrollable factors, it is highly prone to variations [39].
Currently, the classification of Alternaria species primarily relies on the utilization of morphological characteristics and multigene phylogenetic analysis methods. Lawrence et al [40], Woudenberg et al [41], Grum-Grzhimaylo et al [42] and Ghafri et al [43] used multigene analyses to reconstruct phylogenetic relationships within the genus Alternaria, resulting in the division of the genus into 28 clades, each represented by a type specimen. Phylogenetic analysis of Alternaria using multiple nucleotide sequences, such as ITS, mtLSU, endoPG, TUB, mtSSU, ATP, EF-1α, gpd, Alt a 1, CAL, CHS, ACT, OPA2-1, IGS, HIS, TMA22, PGS1, and REV3, is often employed because these genes play a role in the identification of the genus Alternaria as well as similar interspecifics. Alternaria sect. Alternaria contains most of the small-spored Alternaria species with concatenated conidia, including important plant, human and postharvest pathogens [44]. Alternaria sect. Alternaria consists of only 11 phylogenetic species and one species complex [40].
Temperature is typically regarded as the primary environmental factor influencing the prevalence of plant diseases [45]. In this study, the optimal growth temperature for the representative isolates was 25 °C, mycelial growth was stagnant at 5 °C, and growth was weaker at 15 °C, 20 °C, 30 °C, and 35 °C than at 25 °C. This aligns with the findings of prior studies regarding the ideal temperature for the growth of A. alternata [46]. Therefore, it is advisable to implement timely pathogen control measures before the optimal growth temperature is reached. Leaf spot of C. sinensis tend to occur in March and April, with outbreaks peaking in May and June and continuing through October. During this period, the pathogen thrives in suitable environments and produces highly infectious conidia that infect the plant host.
Evaluating the biological characteristics of pathogens is of utmost significance in preventing and controlling plant disease outbreaks, as it can lay the groundwork for scientifically informed disease prevention and control measures. It is important to detect the presence of pathogens as early as possible to effectively prevent and control plant diseases [47]. Leaf spot of C. sinensis caused by Alternaria not only affect the health and appearance of plants but also have an impact on their economic value. Fungal leaf spot disease usually spreads under wet conditions and can spread to other healthy C. sinensis or plants by wind, raindrops, insects or artificial means, thus spreading the disease further and causing the occurrence of leaf spot disease on large areas of C. sinensis as well as the epidemic occurrence of other plant diseases, which disrupts the ecological balance. Therefore, the negative impacts of C. sinensis leaf spot on agriculture, forestry, urbanization, ecosystems and the economy are manifold, and measures need to be taken to prevent and manage this plant disease. It is necessary to inhibit the growth of fungal mycelium, thus further controlling the spread and transmission of the disease to prevent greater losses. The temperature experiment conducted in this study conclusively demonstrates the importance of determining the timing for fungicide application before the pathogen reaches its optimal growth temperature.
Different fungicides exert their fungicidal effects through various mechanisms based on their type, chemical structure, etc. [48]. Prochloraz inhibits sterol synthesis in fungal cell membranes, leading to cell wall rupture and cell death. It also interferes with fungal DNA synthesis and protein synthesis, resulting in fungicidal effects [49]. In 2009, researchers conducted in vitro susceptibility tests using six fungicides against the pathogen Alternaria solani. The results showed that prochloraz, with EC50 values ranging from 0.03 to 0.11 µg/ml, was effective in inhibiting the growth of the pathogen [50]. This result is consistent with the results of the present study that prochloraz has the best inhibitory effect on A. alternata and A. koreana, causing leaf spot in C. sinensis. However, by comparing the EC50 values, the sensitivity of prochloraz to A. alternata and A. koreana are much less than that to A. solani, and the inhibitory effect is still unsatisfactory. In 2023, the EC50 value of fludioxonil was only 0.089 ± 0.020 in Wang et al.'s study [51], whereas the EC50 value of prochloraz in this study was 2.92, which clearly shows that the inhibitory effect of fludioxonil on A. alternata was stronger. Fewer fungicides were selected in this study, and the selection of efficient, environmentally friendly, and economical fungicides needs to be further explored.
5. Conclusions
In this study, we performed the isolation of fungal pathogens and carried out pathogenicity tests. We identified three isolates (11-13) responsible for causing leaf spot in C. sinensis through multilocus phylogenetic analyses involving ITS, GAPDH, TEF1-α, RPB2, Alt a 1 locis, as well as physical attributes. Three isolates were ascertained that A. alternata and A. koreana. The pathogen's susceptibility to the four fungicides was ascertained through phenotypic experiments conducted on culture medium plates. These sensitivity data can serve as a point of reference for assessing potential changes in the sensitivity of Alternaria spp. to various fungicides in the future. To our knowledge, this is the inaugural report that A. alternata and A. koreana causing leaf spot in C. sinensis in China and worldwide. This finding will yield valuable insights for future investigations focused on the prevention and treatment strategies for this recently emerged disease.
Author Contributions
Q.-Q.W., X.-Y.Z., Y.W., and Y.-J.Z. designed the study and were involved in writing the paper; Q.-Q.W. and Y.-J.Z. were responsible for sample collections; Q.-Q.W. was responsible for pathogenicity tests; Q.-Q.W., X.-Y.Z., and Y.W. were involved in morphological identification; Q.-Q.W., X.-Y.Z. and Y.W. were involved in phylogenetic analyses; Q.-Q.W., X.-Y.Z., Y.W., and Y.-J.Z. contributed to planning and editing of the paper. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
All data generated or analyzed during this study are included in this article.
Acknowledgments
The authors would like to thank those who provided assistance and advice for this study.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fan, J.; Chen, Y.; Luo, M.; Liang, Z.; Nong, X. The chloroplast genome characteristics, comparative genomics and gene resource mining of Celtis sinensis (Persoon, 1805). MITOCHONDRIAL DNA B 2022, 7, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.B.; Lee, K.H.; Nguyen, T.T.T. First Report of Powdery Mildew Caused by Erysiphe michikoae on Celtis sinensis in Korea. Plant Dis. 2020, 104, 984–985. [Google Scholar] [CrossRef]
- Guozheng, W.; Yunyan, Z.; Jiehan, Z.; Mengyuan, Z.; Yimin, H.; Yongjing, T.; Jingbo, Z.; Zhongsheng, W. Characterization of the first complete chloroplast genome sequence of Celtis sinensis (Cannabaceae) and its phylogenetic implications. MITOCHONDRIAL DNA B 2019, 4. [Google Scholar] [CrossRef]
- Huizheng, W.; Shilong, W.; Yubin, L. First Report of Botryosphaeria dothidea Causing Leaf Spot and Wilt on Celtis sinensis in China. Plant Dis. 2020. [Google Scholar] [CrossRef]
- Liang, L.; Li, H.; Zhou, L.F.; Chen, F.M. Lasiodiplodia pseudotheobromae causes stem canker of Chinese hackberry in China. J. For. Res. 2020, 31, 2571–2580. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. STUDIES IN MYCOLOGY 2013, 171–212. [Google Scholar] [CrossRef]
- Qin, Q.M.; Fan, Y.Y.; Jia, Q.L.; Duan, S.S.; Liu, F.J.; Jia, B.X.; Wang, G.Q.; Guo, W.H.; Wang, C. The Potential of Alternaria Toxins Production by A. alternata in Processing Tomatoes. TOXINS. 2022, 14. [Google Scholar] [CrossRef]
- Mitakakis, T.Z.; Barnes, C.; Tovey, E.R. Spore germination increases allergen release from Alternaria. J. Allergy Clin. Immunol. 2001, 107, 388–390. [Google Scholar] [CrossRef]
- Bush, R.K.; Prochnau, J.J. Alternaria-induced asthma. J. Allergy Clin. Immunol. 2004, 113, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Pochon, S.; Terrasson, E.; Guillemette, T.; Iacomi-Vasilescu, B.; Georgeault, S.; Juchaux, M.; Berruyer, R.; Debeaujon, I.; Simoneau, P.; Campion, C. The Arabidopsis thaliana-Alternaria brassicicola pathosystem: A model interaction for investigating seed transmission of necrotrophic fungi. Plant Methods 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Baez-Flores, M.E.; Troncoso-Rojas, R.; Osuna, M.A.I.; Dominguez, M.R.; Pryor, B.; Tiznado-Hernandez, M.E. Differentially expressed cDNAs in Alternaria alternata treated with 2-propenyl isothiocyanate. Microbiol. Res. 2011, 166, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Timmer, L.W.; Peever, T.L.; Solel, Z.; Akimitsu, K. Alternaria diseases of citrus - Novel pathosystems. Phytopathol. Mediterr. 2003, 42, 1000–1014. [Google Scholar]
- Vicent, A.; Badal, J.; Asensi, M.J.; Sanz, N.; Armengol, J.; Garcia-Jimenez, J. Laboratory evaluation of citrus cultivars susceptibility and influence of fruit size on Fortune mandarin to infection by Alternaria alternata pv. citri. Eur. J. Plant Pathol. 2004, 110, 245–251. [Google Scholar] [CrossRef]
- Vega, B.; Dewdney, M.M. Distribution of Qol Resistance in Populations of Tangerine-Infecting Alternaria alternata in Florida. Plant Dis. 2014, 98, 67–76. [Google Scholar] [CrossRef]
- Vega, B.; Dewdney, M.M. Sensitivity of Alternaria alternata from Citrus to Boscalid and Polymorphism in Iron-Sulfur and in Anchored Membrane Subunits of Succinate Dehydrogenase. Plant Dis. 2015, 99, 231–239. [Google Scholar] [CrossRef]
- Song, J.H.; Wang, Y.F.; Yin, W.X.; Huang, J.B.; Luo, C.X. Effect of Chemical Seed Treatment on Rice False Smut Control in Field. Plant Dis. 2021, 105, 3218–3223. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Si, Y.Z.; Li, D.W.; Xu, W.; Sun, J.W.; Zhu, L.H. First Report of Leaf Blotch of Salix babylonica Caused by Botryosphaeria dothidea in China. Plant Dis. 2021, 105, 224–224. [Google Scholar] [CrossRef]
- Zhu, L.H.; Xu, W.; Huang, L.; Ye, J.R.; Li, D.W. Pathogenicity and Biological Characteristics of Septotinia populiperda Causing Leaf Blotch of Willow. Plant Dis. 2022, 106, 1262–1270. [Google Scholar] [CrossRef]
- Liao, Y.C.Z.; Wan, Y.; Si, Y.Z.; Geng, J.X.; Li, D.W.; Zhu, L.H. Leaf spot of Ligustrum japonicum caused by Diaporthe eres newly reported in China. Crop Prot. 2023, 164. [Google Scholar] [CrossRef]
- Si, Y.Z.; Sun, J.W.; Wan, Y.; Chen, Y.N.; He, J.; Li, W.Z.; Li, D.W.; Zhu, L.H. Neofusicoccum cryptomeriae sp. nov. and N. parvum Cause Stem Basal Canker of Cryptomeria japonica in China. J. Fungi 2023, 9. [Google Scholar] [CrossRef]
- Jiao, H.; MeiLing, S.; DeWei, L.; LiHua, Z.; JianRen, Y.; Lin, H. A real-time PCR for detection of pathogens of anthracnose on Chinese fir using TaqMan probe targeting ApMat gene. Pest Manage. Sci. 2022, 79. [Google Scholar] [CrossRef]
- White TJ, B.T., Lee SB, Taylor JW. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protocols: A guide to Methods and Applications 1990, 18, 315–322.
- Carbone I, K.L. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Berbee ML, P.M., Hubbard S Cochliobolus phylogenetics and the origin of known, highly virulent pathogens, inferred from ITS and glyceraldehydes-3-phosphate dehydrogenase gene sequences. Mycologia 1999, 91, 964–977. [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 allergen homologs from Alternaria and related taxa: analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlic, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, B.; Luo, C.; Fu, Y.; Zhu, F. Fungicidal Actions and Resistance Mechanisms of Prochloraz to Penicillium digitatum. Plant Dis. 2021, 105, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Ypema, H.L.; Ypema, M.; Gubler, W.D. Sensitivity of Uncinula necator to Benomyl, Triadimefon, Myclobutanil, and Fenarimol in California. Plant Dis. 1997, 81, 293–297. [Google Scholar] [CrossRef]
- Gama, A.B.; Baggio, J.S.; Rebello, C.S.; Lourenco, S.d.A.; Gasparoto, M.C.d.G.; da Silva Junior, G.J.; Peres, N.A.; Amoriml, L. Sensitivity of Colletotrichum acutatum Isolates from Citrus to Carbendazim, Difenoconazole, Tebuconazole, and Trifloxystrobin. Plant Dis. 2020, 104, 1621–1628. [Google Scholar] [CrossRef]
- Han, S.; Lu, J.; Gao, J.; Cheng, J.; Xu, W.; Tao, L.; Zhang, Y. Pyraclostrobin induced AMPK/mTOR pathways mediated autophagy in RAW264.7 macrophages. J. Environ. Sci. Health Part B. 2021, 56, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.J.; Zheng, X.R.; Li, H.; Chen, F.M. Alternaria alternata, the Causal Agent of a New Needle Blight Disease on Pinus bungeana. J. Fungi 2023, 9. [Google Scholar] [CrossRef]
- Meng, J.X.; Zhang, X.Y.; Han, X.S.; Fan, B. Application and Development of Biocontrol Agents in China. PATHOGENS 2022, 11. [Google Scholar] [CrossRef]
- Zhu, X.Q.; Xiao, C.L. Phylogenetic, Morphological, and Pathogenic Characterization of Alternaria Species Associated with Fruit Rot of Blueberry in California. Phytopathology 2015, 105, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Romain, B.B.N.D.; Hassan, O.; Kim, J.S.; Chang, T. Alternaria koreana sp. nov., a new pathogen isolated from leaf spot of ovate-leaf Atractylodes in South Korea. Mol. Biol. Rep. 2022, 49, 413–420. [Google Scholar] [CrossRef]
- Ramirez-Villacis, D.; Barriga-Medina, N.; Llerena-Llerena, S.; Pazmino-Guevara, C.; Leon-Reyes, A. First Report of Alternaria alternata Causing Leaf Spot on Broccoli in Ecuador. Plant Dis. 2023. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.F.; Jiang, X.; Li, Z.; Wang, H.; Yan, Y.; Yang, H. First report of Alternaria alternata causing leaf blight on Trollius chinensis in China. Plant Dis. 2023. [Google Scholar] [CrossRef]
- Pryor, B.M.; Michailides, T.J. Morphological, pathogenic, and molecular characterization of Alternaria isolates associated with Alternaria late blight of pistachio. Phytopathology. 2002, 92, 406–416. [Google Scholar] [CrossRef]
- Lawrence, D.P.; Gannibal, P.B.; Peever, T.L.; Pryor, B.M. The sections of Alternaria: formalizing species-group concepts. Mycologia 2013, 105, 530–546. [Google Scholar] [CrossRef] [PubMed]
- Woudenberg, J.H.C.; Truter, M.; Groenewald, J.Z.; Crous, P.W. Large-spored Alternaria pathogens in section Porri disentangled. Stud. Mycol. 2014, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Bondarenko, S.A.; Debets, A.J.M.; Bilanenko, E.N. On the diversity of fungi from soda soils. Fungal Diversity. 2016, 76, 27–74. [Google Scholar] [CrossRef]
- Ghafri, A.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Al-Saady, N.; Al-Sadi, A.M. A new section and a new species of Alternaria encountered from Oman. PHYTOTAXA 2019, 405, 279–289. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 1–21. [Google Scholar] [CrossRef]
- Giner, M.M.; Garcia, J.S.C.; Camacho, C.N. Airborne Alternaria spores in SE Spain (1993-98) - Occurrence patterns, relationship with weather variables and prediction models. GRANA 2001, 40, 111–118. [Google Scholar] [CrossRef]
- Cabral, L.D.; Rodriguez, A.; Delgado, J.; Patriarca, A. Understanding the effect of postharvest tomato temperatures on two toxigenic Alternaria spp. strains: growth, mycotoxins and cell-wall integrity-related gene expression. J. Sci. Food Agric. 2019, 99, 6689–6695. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.M.; Li, Y.L.; Fan, J.L.; Zhao, Y.J.; Borriss, R.; Fan, B. Two Lysine Sites That Can Be Malonylated Are Important for LuxS Regulatory Roles in Bacillus velezensis. MICROORGANISMS 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Gisi, U.; Sierotzki, H. Fungicide modes of action and resistance in downy mildews. Eur. J. Plant Pathol. 2008, 122, 157–167. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Zhang, B.; Luo, C.X.; Fu, Y.P.; Zhu, F.X. Fungicidal Actions and Resistance Mechanisms of Prochloraz to Penicillium digitatum. Plant Dis. 2021, 105, 408–415. [Google Scholar] [CrossRef]
- Stepanovic, M.; Vuksa, P.; Potocnik, I.; Milijasevic, S.; Rekanovic, E.; Todorovic, B. Integrated Management of Viral Sensitivity of Alternaria solani Isolates to Some Fungicides. In Proceedings of the II INTERNATIONAL SYMPOSIUM ON TOMATO DISEASES; 2009; pp. 377–380. [Google Scholar] [CrossRef]
- Wang, F.; Saito, S.; Xiao, C.L. Fungicide Resistance of Alternaria alternata and A. arborescens Isolates from Mandarin Fruit and Its Influence on Control of Postharvest Alternaria Rot. Plant Dis. 2023, 107, 1538–1543. [Google Scholar] [CrossRef]
Figure 1.
Symptoms of infected leaves in C. sinensis in the wild. A. Diseased leaves naturally. B. Close-up view of diseased leaves.
Figure 1.
Symptoms of infected leaves in C. sinensis in the wild. A. Diseased leaves naturally. B. Close-up view of diseased leaves.

Figure 2.
Pathogenicity of fungal isolates. A. Symptomless leaves of controls treated with PDA after 4 days. B-D. Symptoms on leaves inoculated with mycelium blocks of isolates 11, 12 and 13 after 4 days. E. Symptomless leaves of controls treated with sterile water after 5 days. F-H. Symptomless leaves of controls treated with sterile water after 5 days. Symptoms on leaves inoculated with 10 µl of conidial suspension (106 conidia/mL) of isolates 11, 12 and 13 after 5 days.
Figure 2.
Pathogenicity of fungal isolates. A. Symptomless leaves of controls treated with PDA after 4 days. B-D. Symptoms on leaves inoculated with mycelium blocks of isolates 11, 12 and 13 after 4 days. E. Symptomless leaves of controls treated with sterile water after 5 days. F-H. Symptomless leaves of controls treated with sterile water after 5 days. Symptoms on leaves inoculated with 10 µl of conidial suspension (106 conidia/mL) of isolates 11, 12 and 13 after 5 days.

Figure 3.
Morphological characteristics of fungal isolates. A. 7-day-old front and back view colony of 11 on PDA. B-C. Conidial chain and conidia of isolate 11, Bars: E = 100 μm, F =10 μm. D. 7-day-old front and back view colony of 12 on PDA. E-F. Conidial chain and conidia of isolate 12, Bars: I = 100 μm, J =10 μm. G. 7-day-old front and back view colony of 13 on PDA. H-I. Conidial chain and conidia of isolate 13, Bars: M = 100 μm, N =10 μm.
Figure 3.
Morphological characteristics of fungal isolates. A. 7-day-old front and back view colony of 11 on PDA. B-C. Conidial chain and conidia of isolate 11, Bars: E = 100 μm, F =10 μm. D. 7-day-old front and back view colony of 12 on PDA. E-F. Conidial chain and conidia of isolate 12, Bars: I = 100 μm, J =10 μm. G. 7-day-old front and back view colony of 13 on PDA. H-I. Conidial chain and conidia of isolate 13, Bars: M = 100 μm, N =10 μm.
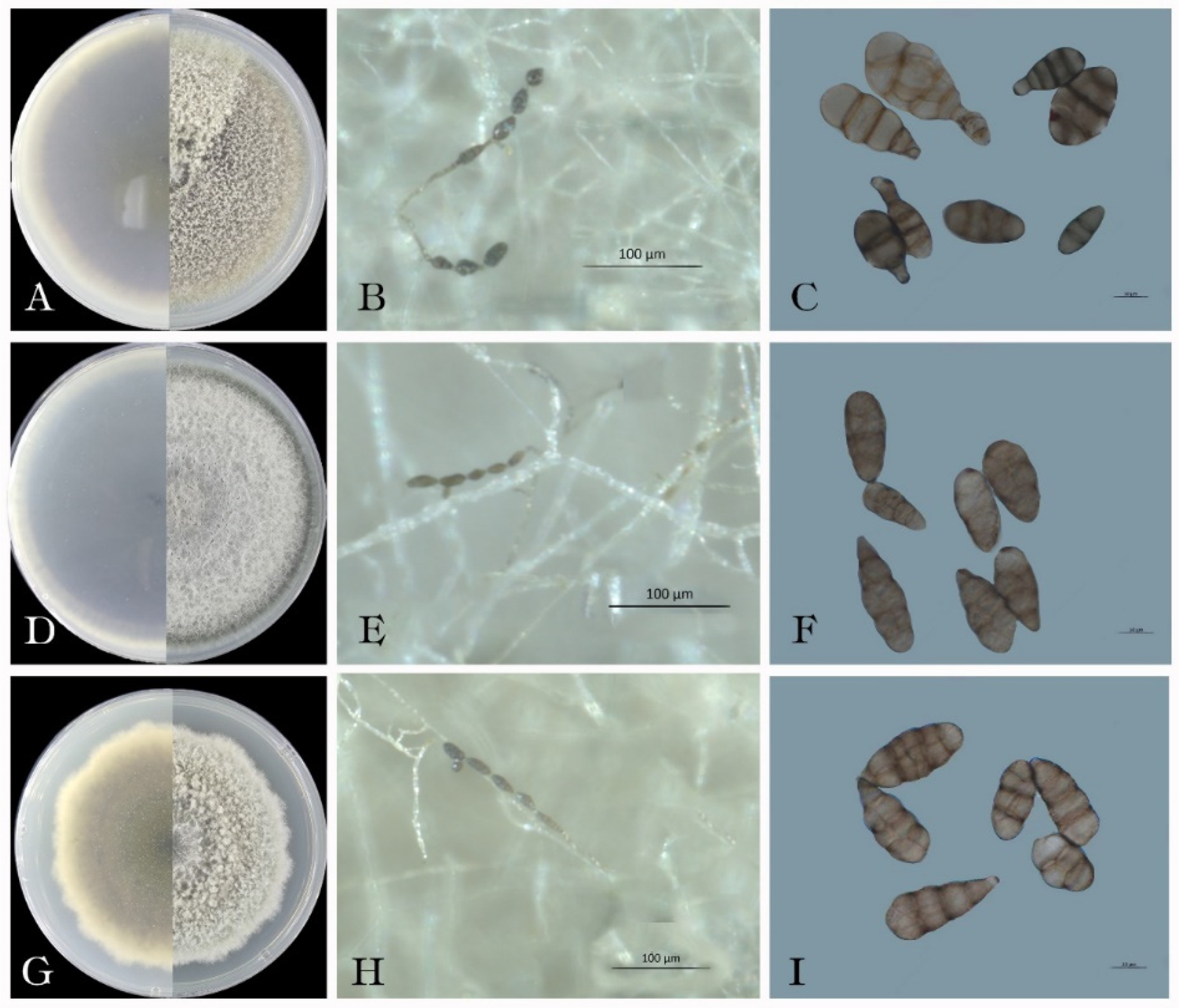
Figure 4.
Phylogenetic relationship of 11, 12 and 13 and related taxa derived from concatenated sequences of the ITS, GAPDH, TEF1-α, RPB2 and Alt a 1 genes using the maximum likelihood algorithm and Bayesian analysis. Bootstrap values >75% (1,000 replications) and Bayesian posterior probability (PP ≥ 0.90) are shown at the nodes (ML/PP). Alternaria alternantherae (CBS 124392) was used as an outgroup. Bar = 0.002 substitutions per nucleotide position. T indicates ex-types.
Figure 4.
Phylogenetic relationship of 11, 12 and 13 and related taxa derived from concatenated sequences of the ITS, GAPDH, TEF1-α, RPB2 and Alt a 1 genes using the maximum likelihood algorithm and Bayesian analysis. Bootstrap values >75% (1,000 replications) and Bayesian posterior probability (PP ≥ 0.90) are shown at the nodes (ML/PP). Alternaria alternantherae (CBS 124392) was used as an outgroup. Bar = 0.002 substitutions per nucleotide position. T indicates ex-types.
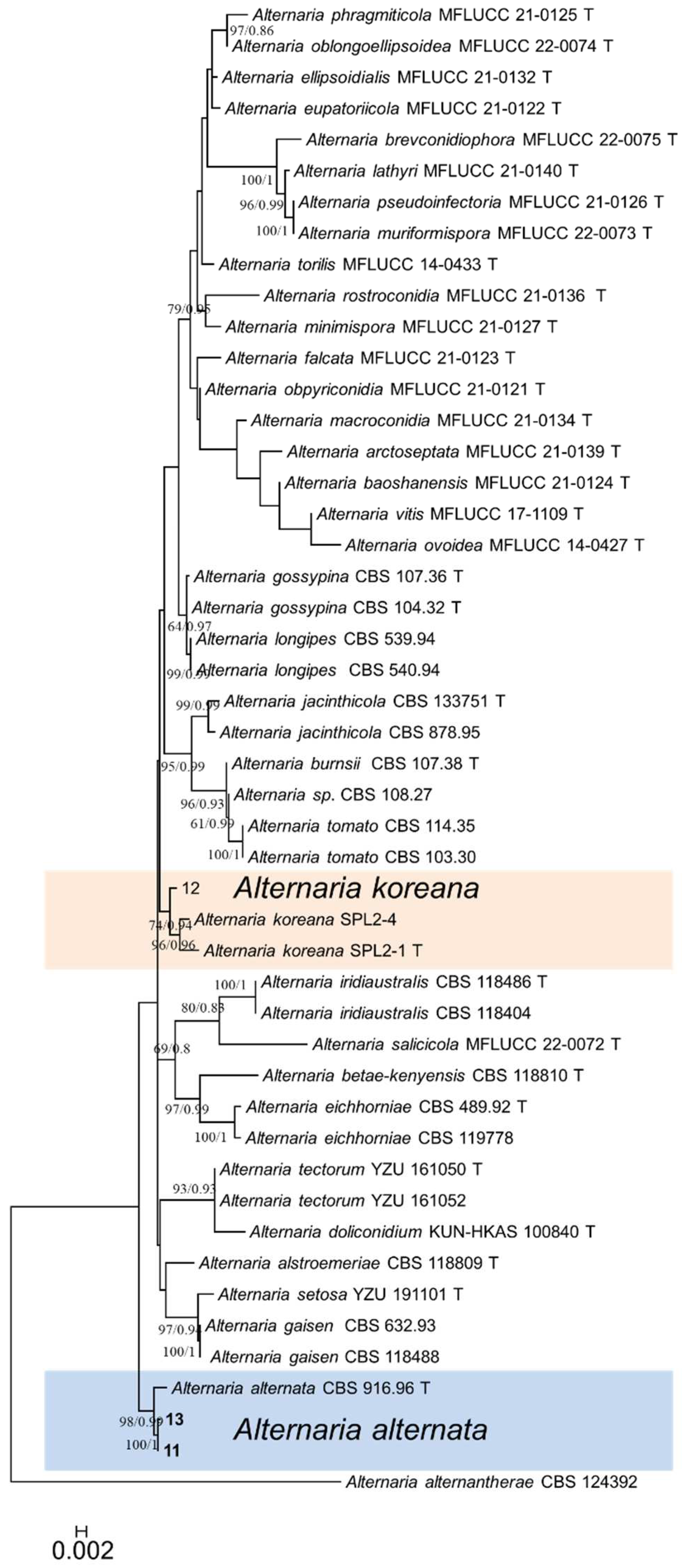
Figure 5.
Colony formation of isolates 11, 12, and 13 isolated from Celtis sinensis hatched for 7 days on different media at 25°C. A-F. Colony morphology of isolate 11 cultured on PDA, CDA, Richard, CMA, OA, and PSA media for 7 days. G-L. Colony morphology of isolate 12 cultured on PDA, CDA, Richard, CMA, OA, and PSA media for 7 days. M-R. Colony morphology of isolate 13 cultured on PDA, CDA, Richard, CMA, OA, and PSA media for 7 days.
Figure 5.
Colony formation of isolates 11, 12, and 13 isolated from Celtis sinensis hatched for 7 days on different media at 25°C. A-F. Colony morphology of isolate 11 cultured on PDA, CDA, Richard, CMA, OA, and PSA media for 7 days. G-L. Colony morphology of isolate 12 cultured on PDA, CDA, Richard, CMA, OA, and PSA media for 7 days. M-R. Colony morphology of isolate 13 cultured on PDA, CDA, Richard, CMA, OA, and PSA media for 7 days.
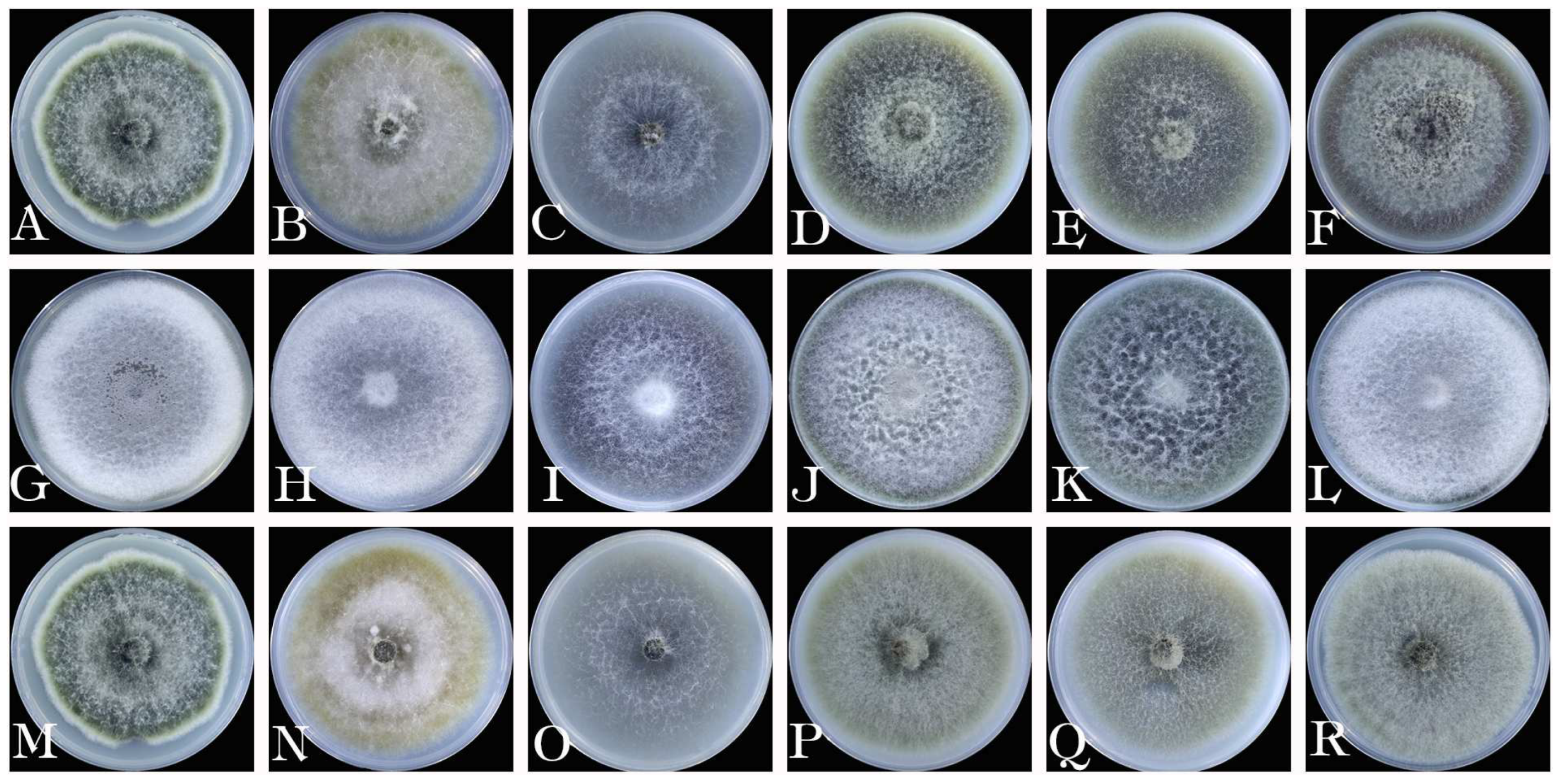
Figure 6.
The impact of temperature on the growth of colony diameters in isolates 11, 12, and 13 after 7 days of culture on PDA media.
Figure 6.
The impact of temperature on the growth of colony diameters in isolates 11, 12, and 13 after 7 days of culture on PDA media.
Figure 7.
The suppression effect on isolates 11, 12 and 13 on plates by corresponding doses of four different fungicides (prochloraz, myclobutanil, tebuconazole, and pyraclostrobin) based on fresh PDA for 7 days.
Figure 7.
The suppression effect on isolates 11, 12 and 13 on plates by corresponding doses of four different fungicides (prochloraz, myclobutanil, tebuconazole, and pyraclostrobin) based on fresh PDA for 7 days.


Table 1.
Primers used for PCR amplification in the molecular identification of three isolates (11–13).
Table 1.
Primers used for PCR amplification in the molecular identification of three isolates (11–13).
| Gene | Primer | Sequence (5’-3’) | PCR amplification cycle parameters: | |
|---|---|---|---|---|
| Internal transcribed spacer (ITS) |
ITS1 ITS4 |
TCCGTAGGTGAACCTGCGG TCCTCCGCTTATTGATATGC |
95 °C: 3 min, (95 °C: 30 s, 55 °C: 30 s,72 °C: 45 s) × 30 cycles, 72 °C: 10 min | |
| Elongation factor 1-alpha (TEF1-α) |
EF1-728F EF1-986R |
CATCGAGAAGTTCGAGAAGG TACTTGAAGGAACCCTTACC |
95 °C: 3 min, (95 °C: 30 s, 59 °C: 30 s, 72 °C: 20 s) × 30 cycles, 72 °C: 10 min | |
| Glycerol-3-phosphate dehydrogenase (GAPDH) |
GPD1 GPD2 |
CAACGGCTTCGGTCGCATTG GCCAAGCAGTTG GTTGT |
95 °C: 3 min, (95 °C: 30 s, 59.5 °C: 30 s, 72 °C: 45 s) × 30 cycles, 72 °C: 10 min | |
| RNA polymerase second largest subunit (RPB2) |
RPB2-5F2 RPB2-7cR |
GGGGWGAYCAGAAGA AGGC CCCATRGCTTGTYYRCCCAT |
95 °C: 3 min, (95 °C: 30 s, 53 °C: 30 s, 72 °C: 50 s) × 30 cycles, 72 °C: 10 min | |
| Alternaria major allergen gene (Alt a 1) |
Alt-al-for Alt-al-rev |
ATGCAGTTCACCACCATCGC ACGAGGGTGAYGTAGGCGTC |
95 °C: 3 min, (95 °C: 30 s, 60.5 °C: 30 s, 72 °C: 30 s) × 30 cycles, 72 °C: 10 min | |
Table 2.
The isolates of Alternaria species used in this study for phylogenetic analysis.
| Species | Isolate | Accession Numbers | |||||||
|---|---|---|---|---|---|---|---|---|---|
| ITS | GAPDH | TEF1-α | RPB2 | Alt a 1 | |||||
| Alternaria alstroemeriae | CBS 118809T | KP124297 | KP124154 | KP125072 | KP124765 | KP123845 | |||
| Alternaria sp. | CBS 108.27 | KC584236 | KC584162 | KC584727 | KC584468 | - | |||
| A. alternantherae | CBS 124392 | KC584179 | KC584096 | KC584633 | KC584374 | KP123846 | |||
| A. alternata | CBS 916.96T | AF347031 | AY278808 | KC584634 | KC584375 | AY563301 | |||
| A. alternata | 11 | OP476716 | OP609771 | OP609768 | OP604538 | OP609775 | |||
| A. alternata | 13 | OP476718 | OP609773 | OP609770 | OP604540 | OP609776 | |||
| A. arctoseptata | MFLUCC 21-0139T |
- | OK236702 | 0K236608 | OK236655 | OK236755 | |||
| A. baoshanensis | MFLUCC 21-0124T |
MZ622003 | OK236706 | OK236613 | OK236659 | OK236760 | |||
| A. betae-kenyensis | CBS 118810T | KP124419 | KP124270 | KP125197 | KP124888 | KP123966 | |||
| A. breviconidiophora | MFLUCC 22-0075T |
MZ621997 | OK236698 | OK236604 | OK236651 | - | |||
| A. burnsii | CBS 107.38T | KP124420 | JQ646305 | KP125198 | KP124889 | KP123967 | |||
| A. doliconidium | KUN-HKAS 100840T |
NR158361 | - | - | - | - | |||
| A. ellipsoidialis | MFLUCC 21-0132T |
MZ621989 | OK236690 | OK236596 | OK236643 | OK236743 | |||
| A. eupatoriicola | MFLUCC 21-0122T |
MZ621982 | OK236683 | OK236589 | OK236636 | OK236736 | |||
| A. eichhorniae | CBS 489.92T | KC146356 | KP124276 | KP125204 | KP124895 | KP123973 | |||
| A. eichhorniae | CBS 119778 | KP124426 | KP124277 | KP125205 | KP124896 | KP123973 | |||
| A. falcata | MFLUCC 21-0123T |
MZ621992 | OK236693 | OK236599 | OK236646 | OK236746 | |||
| A. gaisen | CBS 632.93 | KC584197 | KC584116 | KC584658 | KC584399 | KP123974 | |||
| A. gaisen | CBS 118488 | KP124427 | KP124278 | KP125206 | KP124897 | KP123975 | |||
| A. gossypina | CBS 104.32T | KP124430 | JQ646312 | KP125209 | KP124900 | JQ646395 | |||
| A. gossypina | CBS 107.36T | KP124431 | JQ646310 | KP125210 | KP124901 | JQ646393 | |||
| A. iridiaustralis | CBS 118404 | KP124434 | KP124283 | KP125213 | KP124904 | KP123980 | |||
| A. iridiaustralis | CBS 118486T | KP124435 | KP124284 | KP125214 | KP124905 | KP123981 | |||
| A. jacinthicola | CBS 878.95 | KP124437 | KP124286 | KP125216 | KP124907 | KP123983 | |||
| A. jacinthicola | CBS 133751T | KP124438 | KP124287 | KP125217 | KP124908 | KP123984 | |||
| A. koreana | SPL2-1 (KACC49833)T |
LC621613 | LC621647 | LC621715 | LC621681 | LC631831 | |||
| A. koreana | SPL2-4 | LC621615 | LC621649 | LC621717 | LC621683 | LC631832 | |||
| A. koreana | 12 | OP476717 | OP609772 | OP609769 | OP604539 | OP609774 | |||
| A. lathyri | MFLUCC 21-0140T |
MZ621974 | OK236675 | OK236581 | OK236628 | OK236728 | |||
| A. longipes | CBS 539.94 | KP124441 | KP124290 | KP125220 | KP124911 | KP123987 | |||
| A. longipes | CBS 540.94 | AY278835 | AY278811 | KC584667 | KC584409 | AY563304 | |||
| A. macroconidia | MFLUCC 21-0134T |
MZ622001 | OK236704 | OK236610 | OK236657 | OK236757 | |||
| A. minimispora | MFLUCC 21-0127T |
MZ621980 | OK236681 | OK236587 | OK236634 | OK236734 | |||
| A. muriformispora | MFLUCC 22-0073T |
MZ621976 | OK236677 | OK236583 | OK236630 | OK236730 | |||
| A. ovoidea | MFLUCC 14-0427T |
MZ622005 | OK236708 | OK236614 | OK236661 | OK236761 | |||
| A. phragmiticola | MFLUCC 21-0125T |
MZ621994 | OK236696 | OK236602 | OK236649 | OK236749 | |||
| A. pseudoinfectoria | MFLUCC 21-0126T |
MZ621984 | OK236685 | OK236591 | OK236638 | OK236738 | |||
| A. rostroconidia | MFLUCC 21-0136T |
MZ621969 | OK236670 | OK236576 | OK236623 | OK236723 | |||
| A. salicicola | MFLUCC 22-0072T |
MZ621999 | OK236700 | OK236606 | OK236653 | OK236753 | |||
| A. setosa | YZU 191101T | OP341770 | OP352306 | OP374459 | OP352294 | OP293717 | |||
| A. tectorum | YZU 161050T | OP341728 | OP352303 | OP374456 | OP352291 | OP293714 | |||
| A. tectorum | YZU 161052 | OP341817 | OP352304 | OP374457 | OP352292 | OP293715 | |||
| A. tomato | CBS 103.30 | KP124445 | KP124294 | KP125224 | KP124915 | KP123991 | |||
| A. tomato | CBS 114.35 | KP124446 | KP124295 | KP125225 | KP124916 | KP123992 | |||
| A. torilis | MFLUCC 14-0433T |
MZ621988 | OK236688 | OK236594 | OK236641 | OK236741 | |||
| A. vitis | MFLUCC 17-1109T |
MG764007 | - | - | - | - | |||
T ex-type isolate; bolded are the isolates for this study.
Table 3.
Concentration at 50% of Maximum Effect (EC50 values) of isolates.
| Fungicide | EC50 Values (µg/mL) | ||
| 11 | 12 | 13 | |
| Prochloraz | 2.92 | 3.89 | 3.98 |
| Myclobutanil | 84.54 | 54.86 | 17.23 |
| Tebuconazole | 34.1 | 17.23 | 141.9 |
| Pyraclostrobin | 50.21 | 32.27 | 421.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



