
Submitted:
20 November 2023
Posted:
22 November 2023
You are already at the latest version
Abstract
Obesity affects nearly 660 million adults worldwide and is known for its many comorbidities. Although the phenomenon of obesity is not fully understood, science regularly reveals new determinants of this pathology. Among them, the persistent organic pollutants (POPs) have been recently highlighted. Mainly lipophilic, POPs are normally stored in adipose tissue and can lead to adverse metabolic effects when released into the bloodstream. The main objective of this narrative review is to discuss the different pathways by which physical activity may counteract POPs adverse effects. The research that we carried out seems to indicate that physical activity could positively influence several pathways negatively influenced by POPs, such as insulin resistance, inflammation, lipid accumulation, adipogenesis and gut microbiota dysbiosis that are associated with the development of obesity. This review also indicates how, through the controlled mobilization of POPs, physical activity could be a valuable approach to reduce the concentration of POPs in the bloodstream. These findings suggest that physical activity should be used to counteract the adverse effects of POPs. However, future studies should accurately assess its impact in specific situations such as bariatric surgery where weight loss promotes POPs blood release.
Keywords:
Endocrine disruptors
; Inflammation
; Insulin resistance
; Lipid
; Adipogenesis
; Microbiota
; Bariatric surgery
; Adipose tissue
1. Introduction
Obesity is an excessive accumulation of fat mass within the body contributing to ectopic lipid deposits. Obesity is clinically characterized by a BMI (i.e., Body Mass Index; Supplementary Materials) greater than 30kg/m2 and is linked to metabolic disorders, psychosocial consequences and impaired quality of life. These deposits depend on the cellular modification of the adipose tissue alienating to morbi-mortalities. Globally, obesity is increasing and is considered as a true pandemic. Its prevalence tripled between 1975 and 2016. Worldwide, more than 660 million adults are concerned. Moreover, this increase also affects children and is observed in all socio-economic groups. The latest WHO report specific to the European Region, counts 59% of adults and nearly 1 child out of 3 (29% of boys and 27% of girls) as being overweight or living with obesity. Android obesity, which is specifically characterized by visceral fat accumulation, may be accompanied by comorbidities such as cardiovascular disease [1], hormone-dependent cancers (National Cancer Institute, Obesity and Cancer 2017), type 2 diabetes (T2DM) [2], and metabolic dysfunction-associated fatty liver disease (MAFLD) [3]. Owing to these comorbidities, obesity is estimated to cause millions of deaths [4] each year worldwide (WHO, 2021). It is therefore crucial to address the causes contributing to the increasing prevalence of obesity. In most cases, no single determinant is exclusively responsible for the development of obesity. In this narrative review, we focus on one of the factors contributing to this multifactorial pathology. Specifically, we highlight the role of exposure to endocrine disruptors, in particular persistent organic pollutants (POPs), in the development of obesity and the potentially protective effects of physical activity as a non-medicinal intervention to counteract POPs effects.
1.1. What Are Persistent Organic Pollutants (POPs)?
POPs are organic compounds coming mainly from pesticides and industrial chemicals. These molecules have four specific properties. Firstly, these molecules have the capacity to be transported over very long distances (e.g., between two countries, traveling through air, water, soil and the food chain). Secondly, they are bioaccumulable, implying that these molecules enter the organism, have the capacity to accumulate in the organism and, even more seriously, can be transmitted from generation to generation. Thirdly, they are toxic, which means that they are a significant danger to the health of human beings, and to the wildlife. Fourthly, they are persistent, meaning that it is difficult to degrade these molecules and they can stay in the environment during many years (Stockholm Convention, UN environment program).
In many cases, POPs do not follow a linear dose-response relationship [5]. In other words, POPs can be more dangerous at low doses than at moderate or high doses. Some POPs have been reported to have an inverted U-shaped dose-response relationship. While many studies evaluate the individual effects of several POPs, the human body is affected by mixtures of POPs [5]. The assessment of mixtures is essential, as POPs can have synergistic effects on each other. When a POP is added to a mixture, it can have an additional, multiplying, or antagonistic effect. Adding a new POP to a mixture may also have no effect if its concentration doesn’t allow it.
Many chemical molecules are no longer authorized on the market because of their POPs related properties. However, problems remain, due to the POPs relatively long-life expectancy, their half-life in organic tissues and their unconscious industrial production around the world. Another problem consists of the presence of POPs used before their embargo and therefore remaining in living environment, for example in building materials. Currently, molecules used before their ban are regularly found in our environment. Furthermore, POPs banned in many countries are still observed and/or produced in specific locations/occasions [6,7]. In addition, some POPs are still used in the context of Integrated Vector Management (e.g., dichlorodiphenyltrichloroethane [DDT]). Eradication of POPs remains difficult because of their characteristics and because many molecular alternatives (e.g., industrial, pesticides) still need to be found, knowing that these alternatives must meet multiple criteria to be considered viable (e.g., yield, price, efficacy, safety). POPs ability to travel great distances represents a threat for populations. It should be mentioned that new molecules are regularly added to the official comprehensive list of POPs (established under the Stockholm ConventionListing of POPs in the Stockholm Convention; Stockholm Convention Secretariat, http://www.pops.int/TheConvention/ThePOPs/AllPOPs/tabid/2509/Default.aspx, 12/03/2023). If humans are concerned by these molecules, it is due to their regular exposure to food and water pollution [8]. Following this exposure, and owing to their lipophilic properties, POPs will be stored in adipose tissue and will be able, under certain circumstances, to be released into the bloodstream [9]. The presence of POPs in the human body may cause many disruptions by affecting physiological mechanisms. In this regard, in vitro, in vivo models and epidemiological research have highlighted that POPs could be obesogens [10].
1.2. POPs as Obesogens
At the beginning of the 21st century, links between POPs and adipose tissue were highlighted in the scientific literature. Based on several publications over the last 20 years, the idea emerged that POPs may be determinants of obesity. Indeed, some POPs would have an obesogenic effect [10,11]. Obesogenic molecules are “chemicals that alter homeostatic metabolic setpoints, disrupt appetite controls, perturb lipid homeostasis to promote adipocyte hypertrophy, stimulate adipogenic pathways that enhance adipocyte hyperplasia, or otherwise alter adipocyte differentiation during development” [12]. More precisely, “obesogens are chemicals that elicit increased white adipose tissue mass (WAT) after exposure in vivo […]. Potential obesogens are chemicals that can induce differentiation of adipocytes in vitro but have not yet been demonstrated to increase WAT accumulation in vivo” as defined by Heindel and collaborators [10]. Several different categories of POPs are currently directly associated with obesity such as organochlorines pesticides [OCPs] (e.g., found in insecticides designed to fight against malaria), polychlorinated biphenyls [PCBs] (e.g., found in plastics, paints, and electrical components), per- and polyfluoroalkyl substances [PFASs] (e.g., found in cosmetics, in the textile sector and in phytosanitary products) and polybromodiphenylethers [PBDEs] (e.g., found in the textile and plastic sectors). Obesogenic effects have also been observed for other categories, like dioxins [13]. It should nevertheless be noted that the presence of multiple cofactors makes it difficult to accurately determine the impact of POPs on human obesity.
Taken together, this reflects the harmfulness of POPs to the body and to the consequences of obesity on individuals and society. There is a need to find solutions to effectively address this problem. Many countries are banning the use of these chemicals, but as explained above, some remain in our environment. Several articles have looked at the impact of different types of diet on exposure to POPs [14,15]. To our knowledge, only a few studies have evaluated the effect of physical activity on POPs concentration and/or adverse effects. Recent reviews [10,16] have examined the molecular pathways underlying the ability of POPs to promote the development of obesity. From these reviews, several crucial insights emerge such as adipogenesis, lipid accumulation, insulin resistance/alteration of insulin sensitivity, inflammatory function, and gut microbiota dysbiosis. The practice of physical activity could positively counteract these effects. It may also promote the elimination of POPs. Following a short explanation of the importance of physical activity in the context of obesity, the interest of physical activity for each insight will be discussed.
2. Physical Activity and POPs in the Context of Obesity
2.1. Physical Activity and Obesity
Physical activity (PA) is defined as "any bodily movement produced by skeletal muscle contraction resulting in an increase in energy expenditure over resting energy expenditure" [17]. Adapted physical activity is a form of PA with a preventive and curative aim considering the clinical and motivational state of the person in order to promote bio-psycho-social well-being [18]. The proven effects of PA on the reduction/prevention of diseases and associated disorders, particularly obesity-related comorbidities, and more globally on the improvement of quality of life, have led to the inclusion of the "physical activity prescription" in the Public Health Code of many countries.
According to the American College of Sports and Medicine (ACSM, 2013) and EASO consensus statement [19], PA recommendations to prevent obesity, are similar to those for the general population (i.e., at least 150 min/week of moderate-intensity PA or at least 75min/week of sustained-intensity physical activity combined with resistance training at least 2 times/week). However, to gain more benefits and allow for maintenance of the positive adaptations obtained after weight loss, more than 300min/week of moderate to sustained intensity PA is recommended (ACSM, 2011 and 2013). Despite this, it is nowadays considered that any PA, even less than the recommendations previously mentioned, is better than no activity.
Nevertheless, specific PA programs have proved more effective that the general recommendations. To be more precise, a meta-analysis highlighted a significantly greater decrease in waist circumference with combined- or aerobic-training alone than resistance-training alone [20]. Moderate- to high-intensity aerobic physical training is the most effective type of PA to significantly reduce visceral adipose tissue (VAT) in obesity [21]. Furthermore, an intervention on the mobilization of WAT, in particular reducing VAT, plays a key role in treating obesity-related inflammation. To conclude the meta-analysis observations, PA promotes the mobilization of free fatty acids (FFAs) from their storage sites by enhancing the sensitivity of adipocytes to the lipolytic influence of catecholamines and their muscular utilization through activation of the AMPK/PGC1α signaling pathway [22].
2.2. The Link between POPs, PA, Adipogenesis and Lipid Accumulation
Adipogenesis is the differentiation process whereby preadipocytes become adipocytes. Adipogenesis favorize lipid accumulation and the development of adipose tissues. Adipogenesis and lipid accumulation can be increased through several physiological pathways and transcription factors. Among these PPARγ, STATs, C/EBPα, C/EBPβ, C/EBPδ and SREBP-1 are central [23]. PPARγ is a nuclear receptor and a transcription factor regulating adipocyte differentiation and gene expression. STATs are proteins able to increase adipocytes differentiation in case of ectopic expression. C/EBPs is a set of proteins in which some have an active role at different phases of adipogenesis. SREBP-1 is another transcription factor and protein implicated in lipogenesis. Interestingly, many of this transcription factors interact with each other during adipogenesis. For example, ectopic level of C/EBPβ can be related to PPARγ expression [24] and it can also be true for ectopic expression of STAT5 [25].
It is established that some obesogenic POPs have the ability to increase adipogenesis [26]. POPs can influence the activity and/or expression of the main molecular pathways involved in lipid metabolism through transcription factors (i.e.,PPARγ, C/EBPα, C/EBPβ and SREBP-1), as observed under in vitro conditions [27,28,29]. This would lead to an increased differentiation of adipocytes. C/EBPδ may also be an interesting factor to consider when evaluating POPs effects but there is a lack of studies on this subject. Most studies investigated POPs with antiadipogenic properties. One study did not find effects of PDBE 99 exposure on C/EBPδ [28]. In addition, an increased expression of adipogenesis-specific gene markers such as LPL were observed in in vitro conditions [27].
To our knowledge, there is a lack of evidence about the direct or indirect effects of PA in relation with adipogenesis and lipid accumulation in the presence of POPs. Nevertheless, it is known that PA can be involved in the activity and/or expression level of PPARγ [30,31], C/EBPα, C/EBPβ, and C/EBPδ [32,33] as well as for SREBP-1 [34] as observed in animal models. Thus, better control of adipogenesis through regular PA could promote a reduced risk of obesity development. It is possible to expect that PA, by regulating the expression and activation of the previously cited adipogenesis factors affected by POPs, may prevent the development of obesity. Finally, it is important to mention that, to our knowledge, no study investigated the direct relationship between PA, previously cited transcription factors and POPs.
Glucocorticoid receptor (GR) is a key factor in adipogenesis. A study indicates the inhibition of adipogenesis with GR antagonists [35], and on the contrary, another study observes an increase of adipose tissue development factors (e.g., cell proliferation and triglyceride accumulation) with GR agonist [36]. In an interesting manner, some POPs may disrupt GR expression [37]. Independently of the effect of POPs, the protective effect of PA on GR has already been established [38]. Although studies seem to indicate that GR and POPs can interact with each other [39], the ability of PA to counteract the adverse effects of POPs via an effect on GR remains to be established. The understanding of this relation would allow a better comprehension of how PA may contribute to reduce POPs adverse effects in the context of obesity.
Other molecular mechanisms are involved in the development of adipose tissues and lipid accumulation. Notch pathway, TYK-2/STAT-3 pathway, FABP, FAS, AhR and hormonal actions are also central. Notch pathway is a signal transduction factor participating in lipid metabolism. The inhibition of the Notch pathway can reduce obesity development during high fat diet [40]. TYK-2 is part of the JAK family and refers to an enzyme as well as a gene encoding the enzyme. It is known that the alteration of the TYK-2/STAT-3 pathway can increase obesity development [41]. TYK-2 and STAT-3 are part of the JAK/STAT signaling pathway. FABP is a fatty acid/ lipophilic substances transport protein and FAS is an enzyme, participating to fatty acids biosynthesis. AhR (Aryl hydrocarbon Receptor) is a ligand-activated transcription factor implicated in adipocyte differentiation and related to PPARγ activity [42]. Its inhibition can prevent the development of obesity [43]. Even more interesting, inhibition of AhR has the potential to reverse obesity [44]. Key hormonal factors implicated in adipogenesis and lipid accumulation include epinephrine, norepinephrine, estrogens, and androgens. Epinephrine and norepinephrine are hormones highly implicated during lipolysis. Androgens have antiadipogenic effects and estrogens have proadipogenic effects, following the results observed on in vitro rat preadipocytes [45]. However, the pro and antiadipogenic effects of androgens and estrogens can be debated [23].
POPs have the capacity to negatively alter β-oxidation, to promote lipotoxicity, to alter lipid export and to promote triglyceride synthesis, contributing to enhanced lipid accumulation in the body [46]. Results reported differences between low fat diet and high fat diet. POPs may also be linked to the development of obesity by increasing activation of the Notch pathway, by causing alteration of the TYK-2/STAT-3 pathway and by increasing FABP expression and FAS upregulation, observed under in vitro conditions [29,47,48]. The ability to limit lipid accumulation would also be impaired by POPs. Indeed, POPs are associated with an inhibition of adrenergic, epinephrine and norepinephrine-induced lipolysis pathways [49]. Other studies reported the alteration of mitochondrial function in vivo, but with high POPs concentrations [50]. The ability to prevent the thermogenic response of adipocytes with an AhR agonist (i.e., PCB 126), were also observed under in vitro conditions [51]. For these reasons, and because of its importance in energy expenditure, the thermogenic response principle is central in the context of obesity. A recent in vitro study evaluated the effect of a mixture including 29 POPs [52]. Although 4 of these POPs were AhR agonists, the global mixture was assessed to antagonize AhR activity. However, and according to the results of the same study, some POPs antagonizing AhR activity can have non-monotone and non-linear dose response relationship. The obesogenic effect of POPs known to be AhR agonists has been demonstrated in other articles [13]. The alteration of AhR activity promoting the development of obesity could depend on POPs mixture composition and individual POPs concentration.
In the context of factors related to lipid accumulation, PA can positively influence some molecular pathways such as the Notch pathway and studies show that PA can modulate the TYK-2/STAT-3 pathway [53,54]. The levels of proteins involved in fatty acids transport and related to FABP would also be modified. For example a study observed a decrease of FAB4 plasma levels following PA [55]. In addition, a study reported the capacity of chronic PA to reduce FAS activity on obese rats but not for lean rats [56]. Moreover, it has been shown that PA improves mitochondrial respiration and, more globally, mitochondrial function including protein content [57,58]. Furthermore, PA is known to influence the production of hormones such as adrenaline [59], which could counteract the inhibition of adrenergic, epinephrine and norepinephrine induced lipolysis caused by POPs. It is important to mention that PA is recognized as a central approach in increasing energy expenditure [60], given that studies demonstrate its impact on the thermogenic function of adipose tissue and on reducing AhR, cytoplasmic levels notably in humans [61,62]. The positive impact of PA on β-oxidation, lipid export and triglyceride synthesis is also well known. New research should focus on establishing if these results are still observable in rodents’ models exposed to different POPs. To our knowledge, no study assessed the ability of PA to directly counteract the adverse effects of POPs on Notch pathway, on TYK-2/STAT-3 pathway on FABP, on FAS and on AhR. One study observed that an intervention including diet and PA may attenuate the obesogenic effect of PFASs [63].
A study indicated that POPs could disrupt the translocation and transactivation of androgen receptors. [64]. This result should be taken with caution because it could change depending on the POPs concentrations and combinations. This study also observed different results between different POPs mixtures or compounds, emphasizing the importance of the cocktail effect. Another study reported a low but significant agonist effect of DDT on estrogenic activity [65]. Several studies assessed the effect of PA alone with these hormones. An increase of dihydrotestosterone (i.e., androgen metabolite) is observed following PA and more precisely resistance training [66]. For estrogens, a meta-analysis concluded that PA reduces estradiol body concentrations but also positively influenced SHBG (Sex Hormone Binding Globulin), a hormonal regulator who reduces hormones activity [67]. To our knowledge no study directly compared PA and POPs on androgens and estrogens.
2.3. The Link between POPs, PA and Insulin Resistance/Insulin Sensitivity
Reduction of insulin sensitivity is one of the determinants of obesity and relations between insulin resistance and obesity are well known [68,69]. Lack of insulin sensitivity and insulin resistance are partially caused by the alteration of various physiological factors of insulin regulation. Examples of these factors include JNK, IRS, PTEN, PI3K-Akt pathway and GLUT4. JNK is a signal transducer implicated in cellular anabolism and catabolism related to insulin sensitivity, obesity and insulin resistance [70]. JNK also have a key role on macrophage activity. IRS are proteins whose role mostly consist in transmitting intracellular signals coming from insulin receptors. They participate to glucose metabolism. PTEN is a tumor suppressor known for its ability to inhibit the PI3K-Akt pathway [71] and so for GLUT4 translocation. Dysregulation of the PI3K-Akt pathway can lead to insulin resistance. Alteration of GLUT4 activity can limit glucose transport. Additionally, inflammatory factors such as TNFα can promote insulin resistance. Finally, it is interesting to note that thyroid function is a master regulator of lipid homeostasis and glycaemia homeostasis. Its dysregulation may participate to insulin resistance development [72].
As explained by a recent review [16] POPs can influence the previously cited mechanisms of insulin sensitivity. Thus, some POPs can increase TNFα expression and influence the JNK molecular pathway [73,74]. In addition, POPs can influence thyroid dysfunction in which ROS and JNK pathway may contribute [74]. Other insulin resistance factors such as dysfunction of insulin signaling, negative impact on the Akt pathway (i.e., reduction of phosphorylated Akt), on GLUT4 expression [75] and on insulin receptor/IRS were evidenced [76]. Moreover, POPs are known to increase PTEN expression [75] which inhibits the PI3K enzyme and Akt pathway [77]. This insulin resistance is even more problematic as it contributes to ROS production through its link with hyperglycemia and ROS promotes lipid peroxidation. Lipid peroxidation is part of a vicious circle because it promotes in turn insulin resistance. A recent review summarizes the major interactions between oxidative stress, inflammation, hyperglycemia and insulin resistance [78]. In addition, some researchers proposed that insulin resistance would cause an uncontrolled release of POPs into the bloodstream [79]. POPs are determinants of obesity, which itself is a determinant of the development of insulin resistance.
It is well known that PA is an effective strategy to prevent and reduce insulin resistance. Studies showed an improvement of insulin sensitivity through PI3K/Akt pathway activity [80], glucose transport via GLUT4 [81] and insulin receptor tyrosine phosphorylation as well as IRS phosphorylation [82,83]. A study reported many combined potential beneficial effects of PA on specific molecular pathways that impact insulin transduction [84], such as GLUT4 expression, insulin receptor expression, IRS2 protein expression, insulin stimulated receptor tyrosine phosphorylation, insulin stimulated tyrosine phosphorylation of IRS1, PI3K activity and insulin stimulated Akt phosphorylation. Positive effects on the activity of PTEN have also been observed following PA and these effects could participate to promote insulin sensitivity [85]. Comparing in a direct manner POPs exposure and PA on insulin resistance is then essential.
Few studies investigated the beneficial effect of PA as a countermeasure to the adverse effects of POPs on insulin resistance. A study compared the effect of tetrachlorodibenzo-p-dioxin (i.e., a POP), and PA on glucose metabolism and IRS/PI3K/Akt pathway [86]. Results showed that while PA had positive effects on some parameters (e.g., IRS2), the ability of PA during POP exposure to counteract altered insulin sensitivity appears to be limited. A second study indicated that children with maternal exposure to PFAS and reporting high PA scores displayed null HOMA-IR indexes on the contrary of children with lower PA scores [87]. An old study revealed an interaction between DDT and PA during the assessment of blood glucose levels on rats [88]. This study also showed that, when exposed to DDT, PA increased insulin levels of exercised rats in comparison with sedentary rats. A recent study, investigating different PFASs, seems to demonstrate that an intervention including diet and PA could protect the individual from diabetogenic effects [89]. Our knowledge about the interaction of POPs, PA and its effect on insulin sensitivity remains limited. Then, it would be interesting to determine the cumulative effect of PA and POPs on insulin related factors like JNK, PTEN and GLUT4.
2.4. The Link between POPs, PA and Inflammatory Function
POPs may influence certain factors related to inflammation, which is associated with insulin resistance and obesity [90]. The easiest way to observe the inflammation caused by chemicals substances is to measure proinflammatory cytokines (e.g., IL-1β and TNFα), anti-inflammatory cytokines (e.g., IL-10 and IL-4), and inflammatory markers (e.g., C-reactive Protein). It is also possible to measure hormones concentration more or less directly related to inflammation. Thus, leptin and adiponectin should be considered. As a reminder, leptin is recognized for its influence on satiety as well as inflammation. Another molecule, adiponectin, is recognized for its positive influence on inflammation (i.e., anti-inflammatory capacities), insulin sensitivity and lipid/glucose metabolism.
An intestinal exposure to POPs can promote NF-κB protein activation via the ATM/NEMO pathway, leading to an increase in IL-6 and TNFα, as observed in a rodent model [91]. NF-κB is a protein implicated in cytokine production but also in insulin resistance. POPs are also thought to be linked to inflammation via the AhR protein and expression of the inflammatory cytokine IL-1β, as found in an in vitro experimentation [92]. In addition, this study demonstrated an increase of macrophage polarization and a significant increase of CCL2, CCL3 and CCL4, cytokines involved in the inflammatory function. Other inflammation factors appear to be impacted by POPs exposure. A human study showed that these molecules contribute to altered levels of IFNγ, IL1-β, IL-2, IL-5, IL-8, IL-12p70, IL-17A TNFα and TNFβ [93]. Interestingly, another study indicates that POPs could be the cause of a chronic pro-inflammatory state [94]. Inflammatory markers such as C-reactive Protein seem to increase in presence of some POPs while others don’t show any effect or are inversely associated with it [95]. The impact of POPs on inflammation-related factors may also involves alteration of leptin signaling, leptin gene expression and leptin receptor expression [96]. Adiponectin can be negatively associated with POPs [97] as observed in a human study. The link between POPs and adiponectin could be explained indirectly by the hypoxia phenomenon caused by adipose tissue expansion. However, this may not be applicable to all POPs [98]. Other factors promoting inflammation that are impacted by POPs are to be considered such as ROS [99,100], adipose tissue dysfunction, altered lipid metabolism as well as macrophage infiltration [101]. According to the result of a study [102], intestinal inflammation can also be caused by POPs in a AhR-dependent manner.
Regarding factors related to inflammation, PA would limit the activation of the NF-κB protein, in particular by promoting the increase of its inhibitor IκB [103]. Another study shows that PA would promote the release of anti-inflammatory cytokines such as IL-1ra and IL-10. This increased concentration of anti-inflammatory cytokines would result from the production of IL-6 following PA [104]. Together with this IL-6 production, PA may inhibit other inflammatory factors like IL-1β and TNFα and results can change following sex, PA intensity and PA duration [104,105]. A meta-analysis focusing on overweight and obese adults indicated that PA in association with caloric restriction can be more efficient than caloric restriction alone to reduce TNFα levels. Results were dependent on lifestyle behaviors (i.e., sedentary levels) [106]. In parallel, we have already mentioned the influence of PA on the AhR protein, whose inflammatory role was discussed earlier. Although it is recognized that immediate assessment of inflammation following PA reveal elevation of inflammatory markers, chronic PA can reduce CRP concentrations [107]. A meta-analysis [108] highlights that PA was associated with a decrease in leptin and an increase in adiponectin in children. Interestingly, both hormones are strongly linked to the insulin resistance phenomenon and to the inflammatory function [109]. PA is also essential for reducing oxidative stress, and different training modalities may have different effects and targets [110]. Furthermore, oxidative stress is known to be linked to insulin resistance, which was previously described as a major determinant of obesity and POPs release into the bloodstream.
All the previously cited articles only assessed the effect of PA on inflammation without POPs exposure. Evidence for comparable effects in organisms contaminated by POPs remains scarce. PA was recognized as key approach to limit oxidative stress caused by some POPs, by increasing the activity of antioxidant enzymes such as SOD, CAT, GSH-Px, and MDA scavenging [111]. A study reported a decrease of IL-6, CCL2, and macrophages in the PA group exposed to one PCB in comparison with a sedentary group. Results also showed reduced oxidative stress and increases of various antioxidant enzymes (e.g., GPx) [112]. Regarding hormones, a study assessed the effect of PA on leptin/adiponectin ratio following maternal exposure to specific PFASs [87]. Interestingly, this ratio was used to assess adipose tissue dysfunction. Another study reported links between PFASs, leptin and adiponectin but PA did not appear to prevent this alterations [113]. A study also demonstrated the interaction between PA and POPs on inflammatory factors but in the context of wound healing [114]. Nevertheless, this study observed that IL-1β levels, TNFα levels, CCL2 levels and IL-6 levels following PA changed in function of POPs exposure. Knowledge linking PA, inflammation and POPs being limited, futures study should explore this research area.
2.5. The Link between POPs, PA and Gut Microbiota
Exposure to specific POPs can cause dysbiosis of the gut microbiome, as observed in a mouse model [115]. A recent review highlights how dysbiosis can contribute to the development of obesity [116]. Additionally, it is important to mention that the adverse effects of obesogenic chemicals may be facilitated by the alteration of gut microbiota [117]. Another study reported alteration of gut microbiota, metabolism and inflammatory function when exposed to POPs [102]. This study also reported an important relationship between POPs, gut microbiota and AhR. Our narrative review already exposed the effect of AhR during obesity development.
As described in a recent review, several research already focused on how different PA could modulate gut microbiota, for example by affecting the Bacteroides/Firmicutes ratio or the gut microbiota diversity [118]. Interestingly, PA in obese children can reduce the abundance of proteobacteria phylum and proteobacteria are known to be associated with the gut microbiota profile of adult people living with obesity [119,120]. It is possible to suggest that PA may protect individuals from microbiota dysbiosis in the early stages of their life when exposure to POPs is critical.
Following these insights, it was necessary to assess if there was a clear interaction between PA and POPs exposure. One study found that when mice were exposed to PCBs, the abundance of proteobacteria was decreased but PA attenuated this decrease [115]. A second study presented more mitigated results when observing the effect of PA on gut microbiota of mice following maternal exposure to PCBs [121].
2.6. Effects of PA on the Mobilization of POPs
Some of the above-mentioned studies suggest that PA could be a valuable strategy to counteract POPs adverse effects. However, one may also argue that PA could also promote the release of POPs into the bloodstream. This may be associated with adverse effects. Few articles focused on the effects of PA on POPs blood concentration levels. It is therefore essential to assess if PA can increase POPs excretion, elimination or on the contrary increase their interaction with organs.
Following their entrance in the body, most POPs can be found in adipose tissues where they are stored, but also in the blood flow. As suggested in a study [122] POPs moving in the bloodstream may be eliminated or transformed by several mechanisms. These include biotransformation (i.e., chemical reaction altering a substance) and biliary clearance/excretion [123]. Both mechanisms can be influenced by PA [124,125,126]. In fact, a study highlighted the ability of aerobic exercise to facilitate DDT degradation [111]. Interestingly, authors explain that anaerobic and aerobic conditions have different biotransformation rate. A more recent study seems to confirm the ability of PA to eliminate POPs from human body [127]. Results demonstrate that PA can reduce benzo(a)pyrene urine levels, with sex-dependent effect. In fact, the elimination potential was more important for female than male. Secondary results show better elimination of benzo(a)pyrene for people with low BMI.
Excretion of POPs via urine or sweat can also contribute to the reduction of POPs concentrations [128,129,130]. Nevertheless, the clearance through perspiration does not impact all POPs similarly [128]. The urinary system is known to be influenced by PA with an increase of diuresis [131]. Nonetheless, one study directly compared the effect of some POPs and an intervention including PA with the kidney function, which is part of the urinary system. Results showed alteration of the kidney function in relation with PFASs but PA did not prevent this association [132]. The sweating phenomenon is increased with PA [133]. It is interesting to note that the quantities of POPs excreted may depend on the type of activity involved in sweat production [130]. More participants should be included in similar studies to confirm these results and extend the identification of the associated mechanisms. Other authors [134] mitigate the importance of POPs excretion by sweat. However, it possible that by including all ways of excretion the results on health may be higher. Moreover, various POPs remain to be studied when assessing sweat and urine rate of excretion.
Although POPs can be eliminated, transformed, or excreted, PA can also increase the concentration of POPs into the bloodstream. Indeed, exercise-induced lipolysis may have the potential to promote the release of POPs into the bloodstream [135] since POPs are stored in adipose tissue. The results of this study nevertheless reveal different rates of release between different PCBs. Increasing the release of POPs into the bloodstream from adipose tissues may facilitate the global reduction of POPs accumulated through life if combined with elimination, transformation, and excretion process. To our knowledge no study has addressed this issue by considering the release/elimination ratio. Yet, this is a crucial information to assess the ability of PA to control the adverse effects of POPs in the context of obesity.
Furthermore, few articles focused on POPs blood concentrations during PA. Recent observations seem to indicate that the effect of PA on POPs blood concentration can vary as a function of individual characteristics (e.g., sex, age, country, exposure rate to POPs and body composition). In addition, these recent observations also reveal that each POPs category (e.g., PCBs, PBDEs, OCPs) can react differently to PA [122,136,137]. For example, in a study, OCPs blood concentrations were shown to be reduced following PA and PCBs blood concentrations did not change significantly [122]. This aspect is still debated in the current literature [138,139]. Another study observed that obese individuals had significantly more plasma concentrations of various POPs than lean individuals and athletes [140]. Only one POP showed significant lower plasma concentration in athletes than in lean individuals.
A schematic representation of the relations between POPs and PA in the context of obesity is presented in Figure 1.
2.7. Is PA an Accurate Solution to Prevent POPs Adverse Effects?
The present review emphasizes the interest of performing PA in relation to the adverse effects of POPs. However, PA could be potentially harmful in some situations if not used with caution. Four main difficulties need to be considered. The first one is the increased exposition to POPs during PA caused by environmental pollution. The second one is the increased lipolysis during PA which facilitate POPs liberation into the bloodstream. The third one is the presence of POPs in physical activity equipment and associated stuff.
Firstly, a study reports an increased risk (x 3) of exposure to high level POPs during PA [137]. A multifactorial approach including various respiratory parameters and mucociliary clearance (i.e., reduced during PA) can explain these results [141]. Then, practicing PA during high pollution levels periods and/or close to POPs sources would increase the inhalation of pollutants and their transport in the ventilatory tract. Another study observes the presence of a large variety of PFASs in the water of swimming pools and suggest that the presence of PFASs may be related to sunscreen, conditioners, and disinfectants [142]. Thereby, it is possible to suppose that PA should be practiced in non-polluted environment if possible. In addition, a study investigated the link between dietary intakes, some POPs and sport [143]. Results showed different dietary habits in comparison with the general population. This could increase the risk of exposure to POPs.
Secondly, PA increase lipolysis within the adipose tissue and reduces triglycerides [144,145]. POPs being lipophilic, lipolysis within the adipose tissue degradation would inevitably cause POPs release into the bloodstream. Although increasing the release of POPs to the bloodstream from adipose tissues may facilitate the global reduction of POPs accumulated through life as previously explained, it may also cause adverse effects by facilitating the interaction of POPs with vital organs (e.g., brain, kidney, liver). This has yet to be proven. Interestingly, PFASs can have hepatotoxic effects but PA could be an efficient strategy to prevent these effects [146]. Another study reported that weight loss can increase POPs concentrations in human milk [147].
Thirdly, many PA related equipment’s are fabricated with POPs and human are regularly in contact with them. For example, a review indicated that, tennis rackets, bicycle, fishing lines, climbing ropes, ski wax and boat equipment were fabricated using PFAS [148]. Textile products are also a source of exposure [149]. Priority should be given to clothing that presents a low risk of exposure to POPs during PA. Not only individuals can be exposed to POPs by skin contact, but it is also very likely for these chemicals to affect the environment [150,151,152,153], resulting in an increased risk of exposure for humans.
During specific situations such as bariatric surgery, both the surgical procedure and the recommended PA programs may increase POPs blood concentrations via environmental exposure and lipolysis.
3. When Physical Activity Is Highly Recommended but Potentially Harmful: The Case of Bariatric Surgery
3.1. Bariatric Surgery Is Associated with an Important Increase of POPs Blood Concentrations.
In some cases of morbid obesity, bariatric surgery may be required. However, the massive weight loss associated with this surgical procedure may promote the release of POPs, with harmful consequences. Indeed, a recent review emphasizes that an individual’s weight loss is accompanied by an increase in POPs concentration of 2–4% per kilogram-weight loss [9]. A recent study also evidences that blood concentration of POPs may double in the year following bariatric surgery [154]. However, the magnitude of the concentration increase may vary depending on the molecule considered. It is nevertheless important to note that in some cases the increases could exceed the current recommendations [155]. Interestingly, it is also observed that certain characteristics, such as the type of POPs released, may vary among populations following bariatric surgery. This makes each person a unique case to be treated. Furthermore, the increase of POPs concentration following bariatric surgery has been shown to be potentially associated with hormonal changes in humans [156]. Finally, the ability of POPs to spread to other organs (e.g., like the brain) via the bloodstream should be considered [157].
3.2. How to Implement PA Programs to Protect against POPs Release?
PA is highly recommended to improve patients’ health, wellbeing, and quality of life after bariatric surgery. PA also favors weight loss, preservation of muscle quality and improves cardiorespiratory capacities [158]. However, after bariatric surgery, it remains to be determined if PA would have beneficial or adverse effects on POPs release. This reasoning is based on the rationale linking PA, lipolysis, and POPs release into the bloodstream. As already mentioned, it is suggested that by promoting lipolysis, PA could increase POPs plasmatic concentrations beyond current sanitary regulation levels. Alternatively, one may suggest that PA could promote POPs elimination. These opposite effects may be modulated by the type of PA involved in the rehabilitation process. After bariatric surgery, aerobic training and resistance training can be combined. Resistance training can be implemented for its beneficial effects on muscle mass and strength and aerobic training for its beneficial effects on cardiovascular, visceral adiposity and respiratory capacities [159,160,161,162]. Each type of PA seems to have specific benefits to offer, but aerobic training promoting fat loss, it could induce a greater release of POPs into the bloodstream (Figure 2). Alternatively, resistance training would not expose to these adverse effects.
In that context, it could be suggested to periodize the different types of PA (i.e., aerobic training and resistance training) during the readaptation period to protect the individual against massive release of POPs into the bloodstream. For instance, given that POPs levels drastically increase during the first months following bariatric surgery, resistance training should be planned during this initial period to limit POPs blood release through reduced lipolysis. Aerobic training should be periodized after this initial phase when POPs release is reduced or after the spike. Alternatively, programming aerobic exercise shortly after surgery may contribute to exceed sanitary regulation levels. It may also cause a harmful release of obesogenic POPs mixtures into the bloodstream (Figure 3).
Consequently, aerobic training should be used later during patient monitoring (i.e., when POPs blood concentration naturally drops), but this remains to be proven. Early periodization of resistance training could prevent overcoming POPs sanitary regulation levels. Resistance training in combination with bariatric surgery may produce less POPs release into the bloodstream than aerobic training after bariatric surgery. When POPs blood concentrations start to drop, aerobic training could be implemented (Figure 4). However, POPs dangerousness not respecting a dose-response linear model, increased POPs blood concentration may not be the ideal biomarker to determine which type of PA should be used and when. Therefore, future studies should assess the relationship between PA modality, POPs mobilization rate, lipolysis-induced POPs blood release rate and POPs concentrations dangerousness.
4. Conclusion
POPs are increasingly known for their ability to promote the development of obesity. Among the main reasons, it is possible to mention the detrimental effects of POPs on adipogenesis, lipid accumulation, insulin resistance, inflammation, and gut dysbiosis. All these factors can be influenced by PA and our analysis of the literature suggests that PA could potentially be an effective intervention to counteract most of the adverse effects of identified POPs. However, only a few studies directly assessed the effects of PA on these adverse effects in the context of exposure to POPs. PA also appears to be associated to the direct mobilization of POPs, but the ins and outs of these mobilizations are less known. For example, it is unclear whether the excretion of POPs via sweat during PA is sufficiently important to limit their accumulation into the bloodstream consecutive to lipolysis. Future studies should address this question.
It is possible to assume that the “POPs adipose tissue profile” including diversity and quantity is the key factor to understand the development of obesity in some individuals. To give an example, it is logical to assume that a person with an ”obesogenic POPs profile“ (i.e., quantity and variety favoring the development of obesity) is more at risk to develop obesity than a person with a ”non-obesogenic POPs profile“ (i.e., quantity and variety less at risk for obesity but which may present other health risks). To our knowledge, no study has established a global profile for a person living with obesity. Although the cost may be very expensive, future studies should aim to assess these profiles, and to establish the effect of PA on all these profiles.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Methods used to select the articles included in the review.
Author Contributions
Q.A.S., and S.L.G., designed the narrative review, researched the articles included in the review and wrote the first draft of the manuscript. V.M., S.S.C., and N.C., edited, critically revised the manuscript. All authors contributed to and approved the final version of the manuscript.
Funding
Q.A.S. obtained a Ph.D grant from La Région SUD Provence-Alpes Côte d’Azur (https://www.maregionsud.fr/). This work has been partly supported by French government, through the UCAJEDI Investments in the Future project managed by the National Research Agency (ANR) with the reference number ANR-15-IDEX-01.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This article is a narrative review. No data were generated.
Acknowledgments
VM is supported by a junior Fellowship from the Institut Universitaire de France.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ACSM | American College of Sports Medicine |
| AhR | Aryl hydrocarbon Receptor |
| Akt | or Protein Kinase B (PKB) |
| AMPK | AMP-Activated Protein Kinase |
| APA | Adapted Physical Activity |
| ATM/NEMO pathway | Ataxia Telangiectasia Mutated/NF-κB essential modifier pathway |
| BaP | Benzo(a)Pyrene |
| C/EBPα | CCAAT Enhancer-Binding Protein alpha |
| C/EBPβ | CCAAT Enhancer-Binding Protein beta |
| C/EBPδ | CCAAT Enhancer-Binding Protein delta |
| CAT | Chloramphénicol AcetylTransferase |
| CCAAT | cytosin-cytosin-adenosin-adenosin-thymidin |
| CCL2 | Macrophage Chemoattractant Protein-1 (MCP-1) |
| CCL3 | Macrophage Inflammatory Protein-1 alpha (MIP-1α) |
| CCL4 | Macrophage Inflammatory Protein-1 beta (MIP-1β) |
| CRP | C Reactive Protein |
| DDT | DichloroDiphenylTrichloroethane |
| EASO | European Asylum Support Office |
| eNOS | endothelial Nitric Oxide Synthase |
| FABP | Fatty Acid-Binding Protein |
| FAS | Fatty Acid Synthase |
| FFAs | Free Fatty Acids |
| GLUT4 | Glucose Transporter type 4 |
| GR | Glucocorticoid Receptor |
| GSH-Px | Glutathion Peroxydase |
| IFNγ | Interferon-gamma |
| IκB | Inhibitor κB |
| IL-12p70 | Interleukin 12p70 |
| IL-17A | Interleukin 17A |
| IL-1Ra | Interleukin-1Ra |
| IL-1β | Interleukin 1beta |
| IL-2 | Interleukin 2 |
| IL-5 | Interleukin 5 |
| IL-6 | Interleukin 6 |
| IL-8: Interleukin 8 | |
| INSERM | Institut National de la Santé et de la Recherche Médicale |
| IRS | Insulin Receptor Substrate |
| IVM | Integrated Vector Management |
| JNK | Jun N-terminal Kinase |
| LPL | Lipoprotein Lipase |
| MDA | Malondialdehyde |
| NADPH oxidase | Nicotinamide Adenine Dinucleotide Phosphate oxidase |
| NF-κB | Nuclear Factor-Kappa B |
| OCPs | OrganoChlorines Pesticides |
| PA | Physical Activity |
| PBDEs | PolyBromoDiphénylEthers |
| PCBs | PolyChlorinated Biphenyls |
| PGC1α | Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-alpha |
| PI3K | Phosphoinositide 3-Kinase |
| POPs | Persistent Organic Pollutants |
| PPARγ | Peroxisome Proliferator-Activated Receptor gamma |
| PTP1B | Protein Tyrosine Phosphatase 1B |
| PTEN | Phosphatase and TENsin homolog |
| ROS | Reactive Oxygen Species |
| SHBG | Sex Hormone Binding Globulin |
| SOCS3 | Supressor Of Cytokine Signaling 3 |
| SOD | SuperOxyde Dismutase |
| SREBP-1 | Sterol Regulatory Element-Binding Protein-1 |
| TNFα | Tumor Necrosis Factor alpha |
| TYK-2/STAT-3 | Tyrosine Kinase-2/Signal Transducer and Activator of Transcription 3 |
| VAT | Visceral Adipose Tissue |
References
- Cercato, C.; Fonseca, F.A. Cardiovascular Risk and Obesity. Diabetol Metab Syndr 2019, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Ciangura, C.; Touizer, E.; Basdevant, A. Who Is Considered Obese? Why? Clinical and Therapeutic Implications. Journal of Visceral Surgery 2010, 147, e5–e9. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Tacke, F.; Arrese, M.; Chander Sharma, B.; Mostafa, I.; Bugianesi, E.; Wai-Sun Wong, V.; Yilmaz, Y.; George, J.; Fan, J.; et al. Global Perspectives on Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis: Hepatology. Hepatology 2019, 69, 2672–2682. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.L.; Vos, T.; Lozano, R.; Naghavi, M.; Flaxman, A.D.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; et al. Disability-Adjusted Life Years (DALYs) for 291 Diseases and Injuries in 21 Regions, 1990–2010: A Systematic Analysis for the Global Burden of Disease Study 2010. The Lancet 2012, 380, 2197–2223. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.-H.; Shioda, T.; Soto, A.M.; Vom Saal, F.S.; Welshons, W.V.; et al. Hormones and Endocrine-Disrupting Chemicals: Low-Dose Effects and Nonmonotonic Dose Responses. Endocrine Reviews 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Bartrons, M.; Catalan, J.; Penuelas, J. Spatial And Temporal Trends Of Organic Pollutants In Vegetation From Remote And Rural Areas. Sci Rep 2016, 6, 25446. [Google Scholar] [CrossRef] [PubMed]
- White, K.B.; Kalina, J.; Scheringer, M.; Přibylová, P.; Kukučka, P.; Kohoutek, J.; Prokeš, R.; Klánová, J. Temporal Trends of Persistent Organic Pollutants across Africa after a Decade of MONET Passive Air Sampling. Environ. Sci. Technol. 2021, 55, 9413–9424. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Pan, B.; Sakkiah, S.; Yavas, G.; Ge, W.; Zou, W.; Tong, W.; Hong, H. Persistent Organic Pollutants in Food: Contamination Sources, Health Effects and Detection Methods. IJERPH 2019, 16, 4361. [Google Scholar] [CrossRef]
- Jansen, A.; Lyche, J.L.; Polder, A.; Aaseth, J.; Skaug, M.A. Increased Blood Levels of Persistent Organic Pollutants (POP) in Obese Individuals after Weight Loss—A Review. Journal of Toxicology and Environmental Health, Part B 2017, 20, 22–37. [Google Scholar] [CrossRef]
- Heindel, J.J.; Howard, S.; Agay-Shay, K.; Arrebola, J.P.; Audouze, K.; Babin, P.J.; Barouki, R.; Bansal, A.; Blanc, E.; Cave, M.C.; et al. Obesity II: Establishing Causal Links between Chemical Exposures and Obesity. Biochemical Pharmacology 2022, 199, 115015. [Google Scholar] [CrossRef]
- Lee, D.-H.; Steffes, M.W.; Sjödin, A.; Jones, R.S.; Needham, L.L.; Jacobs, D.R. Low Dose Organochlorine Pesticides and Polychlorinated Biphenyls Predict Obesity, Dyslipidemia, and Insulin Resistance among People Free of Diabetes. PLoS ONE 2011, 6, e15977. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Porta, M.; Jacobs, D.R.; Vandenberg, L.N. Chlorinated Persistent Organic Pollutants, Obesity, and Type 2 Diabetes. Endocrine Reviews 2014, 35, 557–601. [Google Scholar] [CrossRef] [PubMed]
- Brulport, A.; Le Corre, L.; Chagnon, M.-C. Chronic Exposure of 2,3,7,8-Tetrachlorodibenzo-p-Dioxin (TCDD) Induces an Obesogenic Effect in C57BL/6J Mice Fed a High Fat Diet. Toxicology 2017, 390, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Fleury, S.; Rivière, G.; Allès, B.; Kesse-Guyot, E.; Méjean, C.; Hercberg, S.; Touvier, M.; Bemrah, N. Exposure to Contaminants and Nutritional Intakes in a French Vegetarian Population. Food and Chemical Toxicology 2017, 109, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Kahleova, H.; Tonstad, S.; Rosmus, J.; Fisar, P.; Mari, A.; Hill, M.; Pelikanova, T. The Effect of a Vegetarian versus Conventional Hypocaloric Diet on Serum Concentrations of Persistent Organic Pollutants in Patients with Type 2 Diabetes. Nutrition, Metabolism and Cardiovascular Diseases 2016, 26, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Aaseth, J.; Javorac, D.; Djordjevic, A.; Bulat, Z.; Skalny, A.; Zaitseva, I.; Aschner, M.; Tinkov, A. The Role of Persistent Organic Pollutants in Obesity: A Review of Laboratory and Epidemiological Studies. Toxics 2022, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Caspersen, C.J.; Powell, K.E.; Christenson, G.M. Physical Activity, Exercise, and Physical Fitness: Definitions and Distinctions for Health-Related Research. 6.
- Le Garf, S.; Anty, R. Place de l’Activité Physique Adaptée dans le parcours de soins : cas du patient présentant une stéatose hépatique non-alcoolique (NAFLD). Nutrition Clinique et Métabolisme 2022, 36, 247–255. [Google Scholar] [CrossRef]
- Müller, M.J. Reports of the EASO Physical Activity Working Group: Diverse Insights, Evidence-based Recommendations, and Future Perspectives. Obesity Reviews 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Dias, S.; Strasser, B.; Hoffmann, G. Impact of Different Training Modalities on Anthropometric and Metabolic Characteristics in Overweight/Obese Subjects: A Systematic Review and Network Meta-Analysis. PLoS ONE 2013, 8, e82853. [Google Scholar] [CrossRef]
- Vissers, D.; Hens, W.; Taeymans, J.; Baeyens, J.-P.; Poortmans, J.; Van Gaal, L. The Effect of Exercise on Visceral Adipose Tissue in Overweight Adults: A Systematic Review and Meta-Analysis. PLoS ONE 2013, 8, e56415. [Google Scholar] [CrossRef]
- Gleeson, M.; Bishop, N.C.; Stensel, D.J.; Lindley, M.R.; Mastana, S.S.; Nimmo, M.A. The Anti-Inflammatory Effects of Exercise: Mechanisms and Implications for the Prevention and Treatment of Disease. Nature Reviews Immunology 2011, 11, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harbor Perspectives in Biology 2012, 4, a008417. [Google Scholar] [CrossRef]
- Wu, Z.; Bucher, N.L.R.; Farmer, S.R. Induction of Peroxisome Proliferator-Activated Receptor ␥ during the Conversion of 3T3 Fibroblasts into Adipocytes Is Mediated by C/EBP, C/EBP␦, and Glucocorticoids. MOL. CELL. BIOL. 1996, 16. [Google Scholar] [CrossRef] [PubMed]
- Stewart, W.C.; Pearcy, L.A.; Floyd, Z.E.; Stephens, J.M. STAT5A Expression in Swiss 3T3 Cells Promotes Adipogenesis In Vivo in an Athymic Mice Model System. Obesity 2011, 19, 1731–1734. [Google Scholar] [CrossRef] [PubMed]
- González-Casanova, J.E.; Pertuz-Cruz, S.L.; Caicedo-Ortega, N.H.; Rojas-Gomez, D.M. Adipogenesis Regulation and Endocrine Disruptors: Emerging Insights in Obesity. BioMed Research International 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Liu, W.; Qin, H.; Pan, Y.; Luo, F.; Zhang, Z. Low Concentrations of Perfluorooctane Sulfonate Repress Osteogenic and Enhance Adipogenic Differentiation of Human Mesenchymal Stem Cells. Toxicology and Applied Pharmacology 2019, 367, 82–91. [Google Scholar] [CrossRef]
- Wen, Q.; Xie, X.; Zhao, C.; Ren, Q.; Zhang, X.; Wei, D.; Emanuelli, B.; Du, Y. The Brominated Flame Retardant PBDE 99 Promotes Adipogenesis via Regulating Mitotic Clonal Expansion and PPARγ Expression. Science of The Total Environment 2019, 670, 67–77. [Google Scholar] [CrossRef]
- Mangum, L.H.; Howell, G.E.; Chambers, J.E. Exposure to p,P′-DDE Enhances Differentiation of 3T3-L1 Preadipocytes in a Model of Sub-Optimal Differentiation. Toxicology Letters 2015, 238, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Cai, Y. Concurrent Exercise Improves Insulin Resistance and Nonalcoholic Fatty Liver Disease by Upregulating PPAR-γ and Genes Involved in the Beta-Oxidation of Fatty Acids in ApoE-KO Mice Fed a High-Fat Diet. Lipids Health Dis 2019, 18, 6. [Google Scholar] [CrossRef]
- Gu, X.; Ma, X.; Mo, L.; Wang, Q. The Role of Exercise Intensity on Fatty Liver in Rats. Chin J Physiol 2022, 65, 301. [Google Scholar] [CrossRef]
- Khalafi, M.; Mohebbi, H.; Symonds, M.E.; Karimi, P.; Akbari, A.; Tabari, E.; Faridnia, M.; Moghaddami, K. The Impact of Moderate-Intensity Continuous or High-Intensity Interval Training on Adipogenesis and Browning of Subcutaneous Adipose Tissue in Obese Male Rats. Nutrients 2020, 12, 925. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhou, H.; Jin, W.; Lee, H.J. Acute Exercise Regulates Adipogenic Gene Expression in White Adipose Tissue. Biol Sport 2016, 33, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Liu, Y.; Ma, Y.; Wen, D. High-Intensity Interval versus Moderate-Intensity Continuous Training: Superior Metabolic Benefits in Diet-Induced Obesity Mice. Life Sciences 2017, 191, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Asada, M.; Rauch, A.; Shimizu, H.; Maruyama, H.; Miyaki, S.; Shibamori, M.; Kawasome, H.; Ishiyama, H.; Tuckermann, J.; Asahara, H. DNA Binding-Dependent Glucocorticoid Receptor Activity Promotes Adipogenesis via Krüppel-like Factor 15 Gene Expression. Laboratory Investigation 2011, 91, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Kassotis, C.D.; Masse, L.; Kim, S.; Schlezinger, J.J.; Webster, T.F.; Stapleton, H.M. Characterization of Adipogenic Chemicals in Three Different Cell Culture Systems: Implications for Reproducibility Based on Cell Source and Handling. Sci Rep 2017, 7, 42104. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.; Berntsen, H.F.; Zimmer, K.E.; Verhaegen, S.; Frizzell, C.; Ropstad, E.; Connolly, L. Do Persistent Organic Pollutants Interact with the Stress Response? Individual Compounds, and Their Mixtures, Interaction with the Glucocorticoid Receptor. Toxicology Letters 2016, 241, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Dassonvalle, J.; Díaz-Castro, F.; Donoso-Barraza, C.; Sepúlveda, C.; Pino-de La Fuente, F.; Pino, P.; Espinosa, A.; Chiong, M.; Llanos, M.; Troncoso, R. Moderate Aerobic Exercise Training Prevents the Augmented Hepatic Glucocorticoid Response Induced by High-Fat Diet in Mice. IJMS 2020, 21, 7582. [Google Scholar] [CrossRef] [PubMed]
- Sargis, R.M.; Johnson, D.N.; Choudhury, R.A.; Brady, M.J. Environmental Endocrine Disruptors Promote Adipogenesis in the 3T3-L1 Cell Line through Glucocorticoid Receptor Activation. Obesity 2010, 18, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Bi, P.; Shan, T.; Liu, W.; Yue, F.; Yang, X.; Liang, X.-R.; Wang, J.; Li, J.; Carlesso, N.; Liu, X.; et al. Inhibition of Notch Signaling Promotes Browning of White Adipose Tissue and Ameliorates Obesity. Nat Med 2014, 20, 911–918. [Google Scholar] [CrossRef]
- Derecka, M.; Gornicka, A.; Koralov, S.B.; Szczepanek, K.; Morgan, M.; Raje, V.; Sisler, J.; Zhang, Q.; Otero, D.; Cichy, J.; et al. Tyk2 and Stat3 Regulate Brown Adipose Tissue Differentiation and Obesity. Cell Metabolism 2012, 16, 814–824. [Google Scholar] [CrossRef]
- Dou, H.; Duan, Y.; Zhang, X.; Yu, Q.; Di, Q.; Song, Y.; Li, P.; Gong, Y. Aryl Hydrocarbon Receptor (AhR) Regulates Adipocyte Differentiation by Assembling CRL4B Ubiquitin Ligase to Target PPARγ for Proteasomal Degradation. Journal of Biological Chemistry 2019, 294, 18504–18515. [Google Scholar] [CrossRef]
- Moyer, B.J.; Rojas, I.Y.; Kerley-Hamilton, J.S.; Nemani, K.V.; Trask, H.W.; Ringelberg, C.S.; Gimi, B.; Demidenko, E.; Tomlinson, C.R. Obesity and Fatty Liver Are Prevented by Inhibition of the Aryl Hydrocarbon Receptor in Both Female and Male Mice. Nutrition Research 2017, 44, 38–50. [Google Scholar] [CrossRef]
- Rojas, I.Y.; Moyer, B.J.; Ringelberg, C.S.; Tomlinson, C.R. Reversal of Obesity and Liver Steatosis in Mice via Inhibition of Aryl Hydrocarbon Receptor and Altered Gene Expression of CYP1B1, PPARα, SCD1, and Osteopontin. Int J Obes 2020, 44, 948–963. [Google Scholar] [CrossRef]
- Dieudonne, M.N.; Pecquery, R.; Leneveu, M.C.; Giudicelli, Y. Opposite Effects of Androgens and Estrogens on Adipogenesis in Rat Preadipocytes: Evidence for Sex and Site-Related Specificities and Possible Involvement of Insulin-Like Growth Factor 1 Receptor and Peroxisome Proliferator-Activated Receptorγ 21. Endocrinology 2000, 141, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhu, L.; Kang, Q.; Lee, H.K.; Li, D.; Chung, A.C.K.; Cai, Z. Chronic Exposure to Tetrabromodiphenyl Ether (BDE-47) Aggravates Hepatic Steatosis and Liver Fibrosis in Diet-Induced Obese Mice. Journal of Hazardous Materials 2019, 378, 120766. [Google Scholar] [CrossRef]
- Qi, W.; Zhou, L.; Zhao, T.; Ding, S.; Xu, Q.; Han, X.; Zhao, Y.; Song, X.; Zhao, T.; Zhang, X.; et al. Effect of the TYK-2/STAT-3 Pathway on Lipid Accumulation Induced by Mono-2-Ethylhexyl Phthalate. Molecular and Cellular Endocrinology 2019, 484, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Xu, Q.; Xu, Y.; Wang, Z.; Yang, L.; Guo, S.; Shi, Y.; Zhao, T.; Zhou, L.; Ye, L. Effect of Notch Pathway on Lipid Accumulation Induced by Mono-2-Ethylhexyl Phthalate on 3T3-L1 Cells. Ecotoxicology and Environmental Safety 2021, 208, 111472. [Google Scholar] [CrossRef] [PubMed]
- Irigaray, P.; Ogier, V.; Jacquenet, S.; Notet, V.; Sibille, P.; Mejean, L.; Bihain, B.E.; Yen, F.T. Benzo[a]Pyrene Impairs Beta-Adrenergic Stimulation of Adipose Tissue Lipolysis and Causes Weight Gain in Mice. A Novel Molecular Mechanism of Toxicity for a Common Food Pollutant. FEBS Journal 2006, 273, 1362–1372. [Google Scholar] [CrossRef]
- Chehade, L.; Khouri, H.; Malatier--Ségard, J.; Caron, A.; Mauger, J.-F.; Chapados, N.A.; Aguer, C. Acute Exposure to Environmentally Relevant Levels of DDT Alters Muscle Mitochondrial Function in Vivo in Rats but Not in Vitro in L6 Myotubes: A Pilot Study. Toxicology Reports 2022, 9, 487–498. [Google Scholar] [CrossRef]
- Gourronc, F.A.; Perdew, G.H.; Robertson, L.W.; Klingelhutz, A.J. PCB126 Blocks the Thermogenic Beiging Response of Adipocytes. Environ Sci Pollut Res 2020, 27, 8897–8904. [Google Scholar] [CrossRef]
- Doan, T.Q.; Berntsen, H.F.; Verhaegen, S.; Ropstad, E.; Connolly, L.; Igout, A.; Muller, M.; Scippo, M.L. A Mixture of Persistent Organic Pollutants Relevant for Human Exposure Inhibits the Transactivation Activity of the Aryl Hydrocarbon Receptor in Vitro. Environmental Pollution 2019, 254, 113098. [Google Scholar] [CrossRef] [PubMed]
- Shanaki, M.; Khosravi, M.; Khoshdooni-Farahani, A.; Dadashi, A.; Heydari, M.F.; Delfan, M.; Jafary, H.; Gorgani-Firuzjaee, S. High-Intensity Interval Training Reversed High-Fat Diet–Induced M1-Macrophage Polarization in Rat Adipose Tissue via Inhibition of NOTCH Signaling. JIR 2020, Volume 13, 165–174. [Google Scholar] [CrossRef]
- Trenerry, M.K.; Carey, K.A.; Ward, A.C.; Farnfield, M.M.; Cameron-Smith, D. Exercise-Induced Activation of STAT3 Signaling Is Increased with Age. Rejuvenation Research 2008, 11, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, I.; Ferré, R.; Plana, N.; Aragonès, G.; Girona, J.; Merino, J.; Heras, M.; Cabré, A.; Masana, L. Cambios de estilo de vida disminuyen las concentraciones plasmáticas de FABP4 en pacientes con riesgo cardiovascular. Revista Española de Cardiología 2012, 65, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, R.G.; Hollander, J.M.; Ney, D.; Boileau, R.; Jeffery, E.; Li Ji, L.M. Down-Regulates Fatty Acid Synthase and Body Fat in Obese Zucker Rats. Medicine & Science in Sports & Exercise 2002, 34, 1106–1114. [Google Scholar] [CrossRef]
- Mendham, A.E.; Goedecke, J.H.; Zeng, Y.; Larsen, S.; George, C.; Hauksson, J.; Fortuin-de Smidt, M.C.; Chibalin, A.V.; Olsson, T.; Chorell, E. Exercise Training Improves Mitochondrial Respiration and Is Associated with an Altered Intramuscular Phospholipid Signature in Women with Obesity. Diabetologia 2021, 64, 1642–1659. [Google Scholar] [CrossRef] [PubMed]
- Hey-Mogensen, M.; Højlund, K.; Vind, B.F.; Wang, L.; Dela, F.; Beck-Nielsen, H.; Fernström, M.; Sahlin, K. Effect of Physical Training on Mitochondrial Respiration and Reactive Oxygen Species Release in Skeletal Muscle in Patients with Obesity and Type 2 Diabetes. Diabetologia 2010, 53, 1976–1985. [Google Scholar] [CrossRef] [PubMed]
- Gashi, A.; Gontarev, S.; Zivkovic, V.; Gjorgovski, I.; Azemi, A. The Effect of Aerobic Physical Activity in Adrenaline Level in White Laboratory Rats. Med Arch 2020, 74, 84. [Google Scholar] [CrossRef] [PubMed]
- Westerterp, K.R. Control of Energy Expenditure in Humans. Eur J Clin Nutr 2017, 71, 340–344. [Google Scholar] [CrossRef]
- Bonfante, I.L.P.; Duft, R.G.; Mateus, K.C. da S.; Trombeta, J.C. dos S.; Finardi, E.A.R.; Ramkrapes, A.P.B.; Brunelli, D.T.; Mori, M.A. da S.; Chacon-Mikahil, M.P.T.; Velloso, L.A.; et al. Acute/Chronic Responses of Combined Training on Serum Pro-Thermogenic/Anti-Inflammatory Inducers and Its Relation With Fed and Fasting State in Overweight Type 2 Diabetic Individuals. Front. Physiol. 2022, 12, 736244. [Google Scholar] [CrossRef]
- Schenk, A.; Joisten, N.; Walzik, D.; Koliamitra, C.; Schoser, D.; Bloch, W.; Zimmer, P. Acute Exercise Impacts AhR and PD-1 Levels of CD8+ T-Cells—Exploratory Results from a Randomized Cross-over Trial Comparing Endurance versus Resistance Exercise. Eur J Appl Physiol 2021, 121, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, A.; Hauser, R.; Gold, D.R.; Kleinman, K.P.; Hivert, M.-F.; Fleisch, A.F.; Lin, P.-I.D.; Calafat, A.M.; Webster, T.F.; Horton, E.S.; et al. Association of Perfluoroalkyl and Polyfluoroalkyl Substances With Adiposity. JAMA Netw Open 2018, 1, e181493. [Google Scholar] [CrossRef] [PubMed]
- McComb, J.; Mills, I.G.; Muller, M.; Berntsen, H.F.; Zimmer, K.E.; Ropstad, E.; Verhaegen, S.; Connolly, L. Human Blood-Based Exposure Levels of Persistent Organic Pollutant (POP) Mixtures Antagonise Androgen Receptor Transactivation and Translocation. Environment International 2019, 132, 105083. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-S.; Kim, C.-Y.; Lee, H.-K.; Kang, I.-H.; Kim, M.-G.; Jung, K.-K.; Kwon, Y.-K.; Nam, H.-S.; Hong, S.-K.; Kim, H.-S.; et al. Estrogenic Activity of Persistent Organic Pollutants and Parabens Based on the Stably Transfected Human Estrogen Receptor-α Transcriptional Activation Assay (OECD TG 455). Toxicological Research 2011, 27, 181–184. [Google Scholar] [CrossRef] [PubMed]
- Horii, N.; Sato, K.; Mesaki, N.; Iemitsu, M. Increased Muscular 5α-Dihydrotestosterone in Response to Resistance Training Relates to Skeletal Muscle Mass and Glucose Metabolism in Type 2 Diabetic Rats. PLoS ONE 2016, 11, e0165689. [Google Scholar] [CrossRef] [PubMed]
- Ennour-Idrissi, K.; Maunsell, E.; Diorio, C. Effect of Physical Activity on Sex Hormones in Women: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Breast Cancer Res 2015, 17, 139. [Google Scholar] [CrossRef] [PubMed]
- Malone, J.I.; Hansen, B.C. Does Obesity Cause Type 2 Diabetes Mellitus (T2DM)? Or Is It the Opposite? Pediatr Diabetes 2019, 20, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose Tissue and Insulin Resistance in Obese. Biomedicine & Pharmacotherapy 2021, 137, 111315. [Google Scholar] [CrossRef]
- Solinas, G.; Becattini, B. JNK at the Crossroad of Obesity, Insulin Resistance, and Cell Stress Response. Molecular Metabolism 2017, 6, 174–184. [Google Scholar] [CrossRef]
- Verma, K.; Jaiswal, R.; Paliwal, S.; Dwivedi, J.; Sharma, S. An Insight into PI3k/Akt Pathway and Associated Protein–Protein Interactions in Metabolic Syndrome: A Recent Update. J of Cellular Biochemistry 2023, 124, 923–942. [Google Scholar] [CrossRef]
- Choi, Y.M.; Kim, M.K.; Kwak, M.K.; Kim, D.; Hong, E.-G. Association between Thyroid Hormones and Insulin Resistance Indices Based on the Korean National Health and Nutrition Examination Survey. Sci Rep 2021, 11, 21738. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Lee, E.; Moon, T.C.; Jung, H.; Lin, C.X.; Nam, K.-S.; Baek, S.H.; Min, H.-K.; Chang, H.W. Expression of Cyclooxygenase-2 and Pro-Inflammatory Cytokines Induced by 2,2,4,4,5,5-Hexachlorobiphenyl (PCB 153) in Human Mast Cells Requires NF-kB Activation. 2002, 25. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhu, X.; Wang, L.; Ding, G.; Wang, X.; Sheng, Y.; Lv, S.; Yu, J.; Liu, J.; Duan, Y. 2,3’,4,4’,5-Pentachlorobiphenyl Induced Thyroid Dysfunction by Increasing Mitochondrial Oxidative Stress. J. Toxicol. Sci. 2022, 47, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Sun, J.; Zhang, Y. Perfluorooctanoic Acid Impaired Glucose Homeostasis through Affecting Adipose AKT Pathway. Cytotechnology 2018, 70, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Rajesh, P.; Sathish, S.; Srinivasan, C.; Selvaraj, J.; Balasubramanian, K. Diethyl Hexyl Phthalate (DEHP) Is Associated with Insulin Resistance in Adipose Tissue of Male Rat: Protective Role of Antioxidant Vitamins (C & E). J of Cellular Biochemistry 2013, 114, 558–569. [Google Scholar] [CrossRef]
- Boosani, C.S.; Agrawal, D.K. PTEN Modulators: A Patent Review. Expert Opinion on Therapeutic Patents 2013, 23, 569–580. [Google Scholar] [CrossRef] [PubMed]
- et al. Oxidative Stress and Inflammatory Markers in Prediabetes and Diabetes. Journal of Physiology and Pharmacology 2019. [Google Scholar] [CrossRef]
- Lee, Y.-M.; Kim, K.-S.; Jacobs, D.R.; Lee, D.-H. Persistent Organic Pollutants in Adipose Tissue Should Be Considered in Obesity Research: Obesity and Persistent Organic Pollutants. Obesity Reviews 2017, 18, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Marinho, R.; Moura, L.P. de; Rodrigues, B. de A.; Pauli, L.S.S.; Silva, A.S.R. da; Ropelle, E.C.C.; Souza, C.T. de; Cintra, D.E.C.; Ropelle, E.R.; Pauli, J.R. Effects of Different Intensities of Physical Exercise on Insulin Sensitivity and Protein Kinase B/Akt Activity in Skeletal Muscle of Obese Mice. Einstein (São Paulo) 2014, 12, 82–89. [Google Scholar] [CrossRef]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and Skeletal Muscle Glucose Uptake. Physiological Reviews 2013, 93, 993–1017. [Google Scholar] [CrossRef]
- Heled, Y.; Shapiro, Y.; Shani, Y.; Moran, D.S.; Langzam, L.; Braiman, L.; Sampson, S.R.; Meyerovitch, J. Physical Exercise Enhances Protein Kinase C δ Activity and Insulin Receptor Tyrosine Phosphorylation in Diabetes-Prone Psammomys Obesus. Metabolism 2003, 52, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Luciano, E.; Carneiro, E.; Carvalho, C.; Carvalheira, J.; Peres, S.; Reis, M.; Saad, M.; Boschero, A.; Velloso, L. Endurance Training Improves Responsiveness to Insulin and Modulates Insulin Signal Transduction through the Phosphatidylinositol 3-Kinase/Akt-1 Pathway. European Journal of Endocrinology 2002, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Chibalin, A.V.; Yu, M.; Ryder, J.W.; Song, X.M.; Galuska, D.; Krook, A.; Wallberg-Henriksson, H.; Zierath, J.R. Exercise-Induced Changes in Expression and Activity of Proteins Involved in Insulin Signal Transduction in Skeletal Muscle: Differential Effects on Insulin-Receptor Substrates 1 and 2. Proc. Natl. Acad. Sci. U.S.A. 2000, 97, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, V.R.; Gaspar, R.C.; Severino, M.B.; Macêdo, A.P.A.; Simabuco, F.M.; Ropelle, E.R.; Cintra, D.E.; da Silva, A.S.R.; Kim, Y.-B.; Pauli, J.R. Exercise Counterbalances Rho/ROCK2 Signaling Impairment in the Skeletal Muscle and Ameliorates Insulin Sensitivity in Obese Mice. Front. Immunol. 2021, 12, 702025. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, J.; Zhu, Y.; Yan, H.; Lu, Y. Effects of Different Intensity Exercise on Glucose Metabolism and Hepatic IRS/PI3K/AKT Pathway in SD Rats Exposed with TCDD. IJERPH 2021, 18, 13141. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.M.; Papandonatos, G.D.; Li, N.; Sears, C.G.; Buckley, J.P.; Cecil, K.M.; Chen, A.; Eaton, C.B.; Kalkwarf, H.J.; Kelsey, K.T.; et al. Physical Activity Modifies the Relation between Gestational Perfluorooctanoic Acid Exposure and Adolescent Cardiometabolic Risk. Environmental Research 2022, 214, 114021. [Google Scholar] [CrossRef] [PubMed]
- Berdanier, C.D.; De Dennis, S.K. Effect of Exercise on the Responses of Rats to DDT. Journal of Toxicology and Environmental Health 1977, 2, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, A.; Hivert, M.-F.; Gold, D.R.; Hauser, R.; Kleinman, K.P.; Lin, P.-I.D.; Fleisch, A.F.; Calafat, A.M.; Ye, X.; Webster, T.F.; et al. Associations of Perfluoroalkyl and Polyfluoroalkyl Substances With Incident Diabetes and Microvascular Disease. Diabetes Care 2019, 42, 1824–1832. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E. Inflammation and Insulin Resistance. Journal of Clinical Investigation 2006, 116, 1793–1801. [Google Scholar] [CrossRef]
- Phillips, M.C.; Dheer, R.; Santaolalla, R.; Davies, J.M.; Burgueño, J.; Lang, J.K.; Toborek, M.; Abreu, M.T. Intestinal Exposure to PCB 153 Induces Inflammation via the ATM/NEMO Pathway. Toxicology and Applied Pharmacology 2018, 339, 24–33. [Google Scholar] [CrossRef]
- Wang, C.; Petriello, M.C.; Zhu, B.; Hennig, B. PCB 126 Induces Monocyte/Macrophage Polarization and Inflammation through AhR and NF-κB Pathways. Toxicology and Applied Pharmacology 2019, 367, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Imbeault, P.; Findlay, C.S.; Robidoux, M.A.; Haman, F.; Blais, J.M.; Tremblay, A.; Springthorpe, S.; Pal, S.; Seabert, T.; Krümmel, E.M.; et al. Dysregulation of Cytokine Response in Canadian First Nations Communities: Is There an Association with Persistent Organic Pollutant Levels? PLoS ONE 2012, 7, e39931. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Liu, Y.; Wang, J.; Zhao, Y.; Li, K.; Jing, Y.; Zhang, X.; Liu, Q.; Geng, X.; Li, G.; et al. Long-Term Persistent Organic Pollutants Exposure Induced Telomere Dysfunction and Senescence-Associated Secretary Phenotype. The Journals of Gerontology: Series A 2018, 73, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-S.; Hong, N.-S.; Jacobs, D.R.; Lee, D.-H. Interaction Between Persistent Organic Pollutants and C-Reactive Protein in Estimating Insulin Resistance Among Non-Diabetic Adults. J Prev Med Public Health 2012, 45, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, M.C.; Amero, P.; Santoro, A.; Monnolo, A.; Simeoli, R.; Di Guida, F.; Mattace Raso, G.; Meli, R. Polychlorinated Biphenyls (PCB 101, PCB 153 and PCB 180) Alter Leptin Signaling and Lipid Metabolism in Differentiated 3T3-L1 Adipocytes. Toxicology and Applied Pharmacology 2014, 279, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Jee, S.H. Association between Serum Levels of Adiponectin and Polychlorinated Biphenyls in Korean Men and Women. Endocrine 2015, 48, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Amine, Z.E.; Mauger, J.-F.; Imbeault, P. CYP1A1, VEGFA and Adipokine Responses of Human Adipocytes Co-Exposed to PCB126 and Hypoxia. Cells 2022, 11, 2282. [Google Scholar] [CrossRef] [PubMed]
- Berntsen, H.F.; Fonnum, F.; Walaas, S.I.; Bogen, I.L. Low-Chlorinated Non-Dioxin-like Polychlorinated Biphenyls Present in Blood and Breast Milk Induce Higher Levels of Reactive Oxygen Species in Neutrophil Granulocytes than High-Chlorinated Congeners. Basic Clin Pharmacol Toxicol 2016, 119, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive Oxygen Species (ROS) in Macrophage Activation and Function in Diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, H.; Xu, Q.; Han, X.; Zhao, Y.; Song, X.; Zhao, T.; Ye, L. The Effect of Di-2-Ethylhexyl Phthalate on Inflammation and Lipid Metabolic Disorder in Rats. Ecotoxicology and Environmental Safety 2019, 170, 391–398. [Google Scholar] [CrossRef]
- Zhang, L.; Nichols, R.G.; Correll, J.; Murray, I.A.; Tanaka, N.; Smith, P.B.; Hubbard, T.D.; Sebastian, A.; Albert, I.; Hatzakis, E.; et al. Persistent Organic Pollutants Modify Gut Microbiota–Host Metabolic Homeostasis in Mice Through Aryl Hydrocarbon Receptor Activation. Environmental Health Perspectives 2015, 123, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Sriwijitkamol, A.; Christ-Roberts, C.; Berria, R.; Eagan, P.; Pratipanawatr, T.; DeFronzo, R.A.; Mandarino, L.J.; Musi, N. Reversal by Exercise Training. 2006, 55. [Google Scholar]
- Petersen, A.M.W.; Pedersen, B.K. The Anti-Inflammatory Effect of Exercise. Journal of Applied Physiology 2005, 98, 1154–1162. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, X. Anti-Inflammatory Effect of Exercise Training through Reducing Inflammasome Activation-Related Inflammatory Cytokine Levels in Overweight/Obese Populations: A Systematic Review and Meta-Analysis. Complementary Therapies in Clinical Practice 2022, 49, 101656. [Google Scholar] [CrossRef]
- Liu, Y.; Hong, F.; Lebaka, V.R.; Mohammed, A.; Ji, L.; Zhang, Y.; Korivi, M. Calorie Restriction With Exercise Intervention Improves Inflammatory Response in Overweight and Obese Adults: A Systematic Review and Meta-Analysis. Front. Physiol. 2021, 12, 754731. [Google Scholar] [CrossRef]
- Kasapis, C.; Thompson, P.D. The Effects of Physical Activity on Serum C-Reactive Protein and Inflammatory Markers. Journal of the American College of Cardiology 2005, 45, 1563–1569. [Google Scholar] [CrossRef] [PubMed]
- Sirico, F.; Bianco, A.; D’Alicandro, G.; Castaldo, C.; Montagnani, S.; Spera, R.; Di Meglio, F.; Nurzynska, D. Effects of Physical Exercise on Adiponectin, Leptin, and Inflammatory Markers in Childhood Obesity: Systematic Review and Meta-Analysis. Childhood Obesity 2018, 14, 207–217. [Google Scholar] [CrossRef]
- Yadav, A.; Kataria, M.A.; Saini, V.; Yadav, A. Role of Leptin and Adiponectin in Insulin Resistance. Clinica Chimica Acta 2013, 417, 80–84. [Google Scholar] [CrossRef]
- Groussard, C.; Maillard, F.; Vazeille, E.; Barnich, N.; Sirvent, P.; Otero, Y.F.; Combaret, L.; Madeuf, E.; Sourdrille, A.; Delcros, G.; et al. Tissue-Specific Oxidative Stress Modulation by Exercise: A Comparison between MICT and HIIT in an Obese Rat Model. Oxidative Medicine and Cellular Longevity 2019, 2019, 1–11. [Google Scholar] [CrossRef]
- Li, K.; Zhu, X.; Wang, Y.; Zheng, S.; Dong, G. Effect of Aerobic Exercise Intervention on DDT Degradation and Oxidative Stress in Rats. Saudi Journal of Biological Sciences 2017, 24, 664–671. [Google Scholar] [CrossRef]
- Murphy, M.O.; Petriello, M.C.; Han, S.G.; Sunkara, M.; Morris, A.J.; Esser, K.; Hennig, B. Exercise Protects against PCB-Induced Inflammation and Associated Cardiovascular Risk Factors. Environ Sci Pollut Res 2016, 23, 2201–2211. [Google Scholar] [CrossRef] [PubMed]
- Domazet, S.L.; Jensen, T.K.; Wedderkopp, N.; Nielsen, F.; Andersen, L.B.; Grøntved, A. Exposure to Perfluoroalkylated Substances (PFAS) in Relation to Fitness, Physical Activity, and Adipokine Levels in Childhood: The European Youth Heart Study. Environmental Research 2020, 191, 110110. [Google Scholar] [CrossRef] [PubMed]
- Pillai, M.R.; Keylock, K.T.; Cromwell, H.C.; Meserve, L.A. Exercise Influences the Impact of Polychlorinated Biphenyl Exposure on Immune Function. PLoS ONE 2020, 15, e0237705. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.J.; Eum, S.Y.; Rampersaud, E.; Daunert, S.; Abreu, M.T.; Toborek, M. Exercise Attenuates PCB-Induced Changes in the Mouse Gut Microbiome. Environmental Health Perspectives 2013, 121, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-N.; Liu, X.-T.; Liang, Z.-H.; Wang, J.-H. Gut Microbiota in Obesity. WJG 2021, 27, 3837–3850. [Google Scholar] [CrossRef] [PubMed]
- Snedeker, S.M.; Hay, A.G. Do Interactions Between Gut Ecology and Environmental Chemicals Contribute to Obesity and Diabetes? Environmental Health Perspectives 2012, 120, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Campaniello, D.; Corbo, M.R.; Sinigaglia, M.; Speranza, B.; Racioppo, A.; Altieri, C.; Bevilacqua, A. How Diet and Physical Activity Modulate Gut Microbiota: Evidence, and Perspectives. Nutrients 2022, 14, 2456. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, R.; Nistal, E.; Estébanez, B.; Porras, D.; Juárez-Fernández, M.; Martínez-Flórez, S.; García-Mediavilla, M.V.; de Paz, J.A.; González-Gallego, J.; Sánchez-Campos, S.; et al. Exercise Training Modulates the Gut Microbiota Profile and Impairs Inflammatory Signaling Pathways in Obese Children. Exp Mol Med 2020, 52, 1048–1061. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the Gut Microbiota of Adults with Obesity: A Systematic Review. Eur J Clin Nutr 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Agarwal, M.; Hoffman, J.; Ngo Tenlep, S.Y.; Santarossa, S.; Pearson, K.J.; Sitarik, A.R.; Cassidy-Bushrow, A.E.; Petriello, M.C. Maternal Polychlorinated Biphenyl 126 (PCB 126) Exposure Modulates Offspring Gut Microbiota Irrespective of Diet and Exercise. Reproductive Toxicology 2023, 118, 108384. [Google Scholar] [CrossRef]
- Lee, Y.-M.; Shin, J.-Y.; Kim, S.-A.; Jacobs, D.R.; Lee, D.-H. Can Habitual Exercise Help Reduce Serum Concentrations of Lipophilic Chemical Mixtures? Association between Physical Activity and Persistent Organic Pollutants. Diabetes Metab J 2020, 44, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Minh, T.B.; Watanabe, M.; Tanabe, S.; Yamada, T.; Hata, J.; Watanabe, S. Specific Accumulation and Elimination Kinetics of Tris(4-Chlorophenyl)Methane, Tris(4-Chlorophenyl)Methanol, and Other Persistent Organochlorines in Humans from Japan. Environ Health Perspect 2001, 109, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Yiamouyiannis, C.A.; Sanders, R.A.; Watkins, J.B.; Martin, B.J. Chronic Physical Activity: Hepatic Hypertrophy and Increased Total Biotransformation Enzyme Activity. Biochemical Pharmacology 1992, 44, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Yiamouyiannis, C.A.; Martin, B.J.; Watkins, J.B. Chronic Physical Activity Alters Hepatobiliary Excretory Function in Rats. J Pharmacol Exp Ther 1993, 265, 321–327. [Google Scholar] [PubMed]
- Watkins, J.B.; Crawford, S.T.; Sanders, R.A. Chronic Voluntary Exercise May Alter Hepatobiliary Clearance of Endogenous and Exogenous Chemicals in Rats. Drug Metab Dispos 1994, 22, 537–543. [Google Scholar] [PubMed]
- Xu, Y.; Gao, H.; Du, Z.; Liu, H.; Cheng, Q.; Zhang, F.; Ye, J.; Wang, A.; Dou, Y.; Ma, B.; et al. A New Approach for Reducing Pollutants Level: A Longitudinal Cohort Study of Physical Exercises in Young People. BMC Public Health 2022, 22, 223. [Google Scholar] [CrossRef]
- Genuis, S.J.; Beesoon, S.; Birkholz, D. Biomonitoring and Elimination of Perfluorinated Compounds and Polychlorinated Biphenyls through Perspiration: Blood, Urine, and Sweat Study. ISRN Toxicology 2013, 2013, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Genuis, S.J.; Lane, K.; Birkholz, D. Human Elimination of Organochlorine Pesticides: Blood, Urine, and Sweat Study. BioMed Research International 2016, 2016, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Genuis, S.K.; Birkholz, D.; Genuis, S.J. Human Excretion of Polybrominated Diphenyl Ether Flame Retardants: Blood, Urine, and Sweat Study. BioMed Research International 2017, 2017, 1–14. [Google Scholar] [CrossRef]
- Deger, M.; Kapila, V.; Denys, M.A.; Aridogan, I.A.; Everaert, K.; Herve, F. The Impact of Movement, Physical Activity and Position on Urine Production: A Pilot Study. Int J Clin Pract 2021, 75. [Google Scholar] [CrossRef]
- Lin, P.-I.D.; Cardenas, A.; Hauser, R.; Gold, D.R.; Kleinman, K.P.; Hivert, M.-F.; Calafat, A.M.; Webster, T.F.; Horton, E.S.; Oken, E. Per- and Polyfluoroalkyl Substances and Kidney Function: Follow-up Results from the Diabetes Prevention Program Trial. Environment International 2021, 148, 106375. [Google Scholar] [CrossRef]
- Evans, G.H.; James, L.J.; Shirreffs, S.M.; Maughan, R.J. Optimizing the Restoration and Maintenance of Fluid Balance after Exercise-Induced Dehydration. Journal of Applied Physiology 2017, 122, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Imbeault, P.; Ravanelli, N.; Chevrier, J. Can POPs Be Substantially Popped out through Sweat? Environment International 2018, 111, 131–132. [Google Scholar] [CrossRef] [PubMed]
- Louis, C.; Tinant, G.; Mignolet, E.; Thomé, J.-P.; Debier, C. PCB-153 Shows Different Dynamics of Mobilisation from Differentiated Rat Adipocytes during Lipolysis in Comparison with PCB-28 and PCB-118. PLoS ONE 2014, 9, e106495. [Google Scholar] [CrossRef] [PubMed]
- Domazet, S.L.; Grøntved, A.; Jensen, T.K.; Wedderkopp, N.; Andersen, L.B. Higher Circulating Plasma Polychlorinated Biphenyls (PCBs) in Fit and Lean Children: The European Youth Heart Study. Environment International 2020, 136, 105481. [Google Scholar] [CrossRef]
- Harmouche-Karaki, M.; Mahfouz, Y.; Salameh, P.; Matta, J.; Helou, K.; Narbonne, J.-F. Patterns of PCBs and OCPs Exposure in a Sample of Lebanese Adults: The Role of Diet and Physical Activity. Environmental Research 2019, 179, 108789. [Google Scholar] [CrossRef]
- Domazet, S.L.; Jensen, T.K.; Grøntved, A. Response to Correspondence ENVINT_2020_552 “Can Habitual Exercise Really Increase Serum Concentrations of Persistent Organic Pollutants? ” Environment International 2020, 140, 105616. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-M.; Lee, D.-H. Can Habitual Exercise Really Increase Serum Concentrations of Persistent Organic Pollutants? Environment International 2020, 140, 105615. [Google Scholar] [CrossRef] [PubMed]
- Pelletier, C.; Després, J.-P.; Tremblay, A. Plasma Organochlorine Concentrations in Endurance Athletes and Obese Individuals. Med Sci Sports Exerc 2002, 34, 1971–1975. [Google Scholar] [CrossRef]
- Carlisle, A.J.; Sharp, N.C.C. Exercise and Outdoor Ambient Air Pollution. Br J Sports Med 2001, 35, 214–222. [Google Scholar] [CrossRef]
- Martinez, B.; Robey, N.M.; Da Silva, B.F.; Ditz, H.; Sobczak, W.J.; Deliz Quiñones, K.Y.; Bowden, J.A. Swimming with PFAS in Public and Private Pools. Chemosphere 2023, 310, 136765. [Google Scholar] [CrossRef] [PubMed]
- Falcó, G.; Bocio, A.; Llobet, J.M.; Domingo, J.L. Health Risks of Dietary Intake of Environmental Pollutants by Elite Sportsmen and Sportswomen. Food and Chemical Toxicology 2005, 43, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Kelley, G.A.; Kelley, K.S.; Vu Tran, Z. Aerobic Exercise, Lipids and Lipoproteins in Overweight and Obese Adults: A Meta-Analysis of Randomized Controlled Trials. Int J Obes 2005, 29, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Muscella, A.; Stefàno, E.; Lunetti, P.; Capobianco, L.; Marsigliante, S. The Regulation of Fat Metabolism during Aerobic Exercise. Biomolecules 2020, 10, 1699. [Google Scholar] [CrossRef] [PubMed]
- Borghese, M.M.; Liang, C.L.; Owen, J.; Fisher, M. Individual and Mixture Associations of Perfluoroalkyl Substances on Liver Function Biomarkers in the Canadian Health Measures Survey. Environ Health 2022, 21, 85. [Google Scholar] [CrossRef]
- Lignell, S.; Winkvist, A.; Bertz, F.; Rasmussen, K.M.; Glynn, A.; Aune, M.; Brekke, H.K. Environmental Organic Pollutants in Human Milk before and after Weight Loss. Chemosphere 2016, 159, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Glüge, J.; Scheringer, M.; Cousins, I.T.; DeWitt, J.C.; Goldenman, G.; Herzke, D.; Lohmann, R.; Ng, C.A.; Trier, X.; Wang, Z. An Overview of the Uses of Per- and Polyfluoroalkyl Substances (PFAS). Environ. Sci.: Processes Impacts 2020, 22, 2345–2373. [Google Scholar] [CrossRef]
- Xia, C.; Diamond, M.L.; Peaslee, G.F.; Peng, H.; Blum, A.; Wang, Z.; Shalin, A.; Whitehead, H.D.; Green, M.; Schwartz-Narbonne, H.; et al. Per- and Polyfluoroalkyl Substances in North American School Uniforms. Environ. Sci. Technol. 2022, 56, 13845–13857. [Google Scholar] [CrossRef] [PubMed]
- Carlson, G.L.; Tupper, S. Ski Wax Use Contributes to Environmental Contamination by Per- and Polyfluoroalkyl Substances. Chemosphere 2020, 261, 128078. [Google Scholar] [CrossRef]
- Fang, S.; Plassmann, M.M.; Cousins, I.T. Levels of Per- and Polyfluoroalkyl Substances (PFAS) in Ski Wax Products on the Market in 2019 Indicate No Changes in Formulation. Environ Sci Process Impacts 2020, 22, 2142–2146. [Google Scholar] [CrossRef]
- Grønnestad, R.; Vázquez, B.P.; Arukwe, A.; Jaspers, V.L.B.; Jenssen, B.M.; Karimi, M.; Lyche, J.L.; Krøkje, Å. Levels, Patterns, and Biomagnification Potential of Perfluoroalkyl Substances in a Terrestrial Food Chain in a Nordic Skiing Area. Environ. Sci. Technol. 2019, 53, 13390–13397. [Google Scholar] [CrossRef] [PubMed]
- Crawford, K.A.; Doherty, B.T.; Gilbert-Diamond, D.; Romano, M.E.; Claus Henn, B. Waxing Activity as a Potential Source of Exposure to Per- and Polyfluoroalkyl Substances (PFAS) and Other Environmental Contaminants among the US Ski and Snowboard Community. Environmental Research 2022, 215, 114335. [Google Scholar] [CrossRef]
- Fénichel, P.; Coquillard, P.; Brucker-Davis, F.; Marchand, P.; Cano-Sancho, G.; Boda, M.; Antignac, J.-P.; Iannelli, A.; Gugenheim, J.; Le Bizec, B.; et al. Sustained Bloodstream Release of Persistent Organic Pollutants Induced by Extensive Weight Loss after Bariatric Surgery: Implications for Women of Childbearing Age. Environment International 2021, 151, 106400. [Google Scholar] [CrossRef] [PubMed]
- Jansen, A.; Polder, A.; Müller, M.H.B.; Skjerve, E.; Aaseth, J.; Lyche, J.L. Increased Levels of Persistent Organic Pollutants in Serum One Year after a Great Weight Loss in Humans: Are the Levels Exceeding Health Based Guideline Values? Science of The Total Environment 2018, 622–623, 1317–1326. [Google Scholar] [CrossRef]
- Jansen, A.; Aaseth, J.O.; Lyche, J.L.; Berg, J.P.; Müller, M.H.B.; Lydersen, S.; Farup, P.G. Do Changes in Persistent Organic Pollutants after Bariatric Surgery Cause Endocrine Disruption? Chemosphere 2023, 313, 137461. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.H.; Ng, D.K.; Steele, K.; Schweitzer, M.; Groopman, J.D. Mobilization of Environmental Toxicants Following Bariatric Surgery. Obesity 2019, 27, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Bellicha, A.; Baak, M.A.; Battista, F.; Beaulieu, K.; Blundell, J.E.; Busetto, L.; Carraça, E.V.; Dicker, D.; Encantado, J.; Ermolao, A.; et al. Effect of Exercise Training before and after Bariatric Surgery: A Systematic Review and Meta-analysis. Obesity Reviews 2021, 22. [Google Scholar] [CrossRef]
- Schoenfeld, B.J.; Grgic, J.; Ogborn, D.; Krieger, J.W. Strength and Hypertrophy Adaptations Between Low- vs. High-Load Resistance Training: A Systematic Review and Meta-Analysis. Journal of Strength and Conditioning Research 2017, 31, 3508–3523. [Google Scholar] [CrossRef]
- Su, L.; Fu, J.; Sun, S.; Zhao, G.; Cheng, W.; Dou, C.; Quan, M. Effects of HIIT and MICT on Cardiovascular Risk Factors in Adults with Overweight and/or Obesity: A Meta-Analysis. PLoS ONE 2019, 14, e0210644. [Google Scholar] [CrossRef]
- Ismail, I.; Keating, S.E.; Baker, M.K.; Johnson, N.A. A Systematic Review and Meta-Analysis of the Effect of Aerobic vs. Resistance Exercise Training on Visceral Fat: Exercise for Visceral Fat. Obesity Reviews 2012, 13, 68–91. [Google Scholar] [CrossRef]
- In, G.; Taskin, H.E.; Al, M.; Alptekin, H.K.; Zengin, K.; Yumuk, V.; Ikitimur, B. Comparison of 12-Week Fitness Protocols Following Bariatric Surgery: Aerobic Exercise Versus Aerobic Exercise and Progressive Resistance. OBES SURG 2021, 31, 1475–1484. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Potential mechanisms showing how physical activity (PA) may counteract the adverse effects of persistent organic pollutants (POPs) in the context of obesity. These different factors may interact with each other by direct or indirect mechanisms. With its positive impact on insulin function, lipid accumulation, adipogenesis, inflammation and gut microbiota, PA can counteract a large variety of alteration caused by POPs. By increasing or improving physiological mechanisms such as sweat, urine, biotransformation and biliary clearance, PA may reduce POPs blood concentration. Red box [POPs potential negative influence], Green box [PA potential positive influence]. AhR [Aryl hydrocarbon Receptor], AI cytokines [Anti-inflammatory cytokines], Akt [or Protein Kinase B (PKB)], AMPK [AMP-Activated Protein Kinase], Ao-Enzymes [Antioxidant Enzymes], C/EBPα [CCAAT Enhancer-Binding Protein alpha], C/EBPβ [CCAAT Enhancer-Binding Protein beta], C/EBPδ [CCAAT Enhancer-Binding Protein delta], CRP [C-Reactive Protein], FABP [Fatty Acid-Binding Protein], FAS [Fatty Acid Synthase], GLUT4 [Glucose Transporter type 4], IL-1β [Interleukin 1beta], IR [Insulin Receptor], IRS [Insulin Receptor Substrate], JNK [Jun N-terminal Kinase], Lipid management [include alterations of lipolysis, thermogenic function, triglyceride synthesis, β-oxidation, and lipid export], M Respiration [Mitochondrial Respiration], NF-κB [Nuclear Factor-Kappa B], PA [Physical Activity], PI3K [Phosphoinositide 3-Kinase], POPs [Persistent Organic Pollutants], PPARγ [Peroxisome Proliferator-Activated Receptor gamma], PI cytokines [Proinflammatory cytokines], PTEN [Phosphatase and TENsin homolog], ROS [Reactive Oxygen Species], SREBP-1 [Sterol Regulatory Element-Binding Protein-1], TNFα [Tumor Necrosis Factor], TYK-2/STAT-3 pathway [Tyrosine Kinase 2/Signaling Transducer and Activator of Transcription 3 pathway].
Figure 1.
Potential mechanisms showing how physical activity (PA) may counteract the adverse effects of persistent organic pollutants (POPs) in the context of obesity. These different factors may interact with each other by direct or indirect mechanisms. With its positive impact on insulin function, lipid accumulation, adipogenesis, inflammation and gut microbiota, PA can counteract a large variety of alteration caused by POPs. By increasing or improving physiological mechanisms such as sweat, urine, biotransformation and biliary clearance, PA may reduce POPs blood concentration. Red box [POPs potential negative influence], Green box [PA potential positive influence]. AhR [Aryl hydrocarbon Receptor], AI cytokines [Anti-inflammatory cytokines], Akt [or Protein Kinase B (PKB)], AMPK [AMP-Activated Protein Kinase], Ao-Enzymes [Antioxidant Enzymes], C/EBPα [CCAAT Enhancer-Binding Protein alpha], C/EBPβ [CCAAT Enhancer-Binding Protein beta], C/EBPδ [CCAAT Enhancer-Binding Protein delta], CRP [C-Reactive Protein], FABP [Fatty Acid-Binding Protein], FAS [Fatty Acid Synthase], GLUT4 [Glucose Transporter type 4], IL-1β [Interleukin 1beta], IR [Insulin Receptor], IRS [Insulin Receptor Substrate], JNK [Jun N-terminal Kinase], Lipid management [include alterations of lipolysis, thermogenic function, triglyceride synthesis, β-oxidation, and lipid export], M Respiration [Mitochondrial Respiration], NF-κB [Nuclear Factor-Kappa B], PA [Physical Activity], PI3K [Phosphoinositide 3-Kinase], POPs [Persistent Organic Pollutants], PPARγ [Peroxisome Proliferator-Activated Receptor gamma], PI cytokines [Proinflammatory cytokines], PTEN [Phosphatase and TENsin homolog], ROS [Reactive Oxygen Species], SREBP-1 [Sterol Regulatory Element-Binding Protein-1], TNFα [Tumor Necrosis Factor], TYK-2/STAT-3 pathway [Tyrosine Kinase 2/Signaling Transducer and Activator of Transcription 3 pathway].
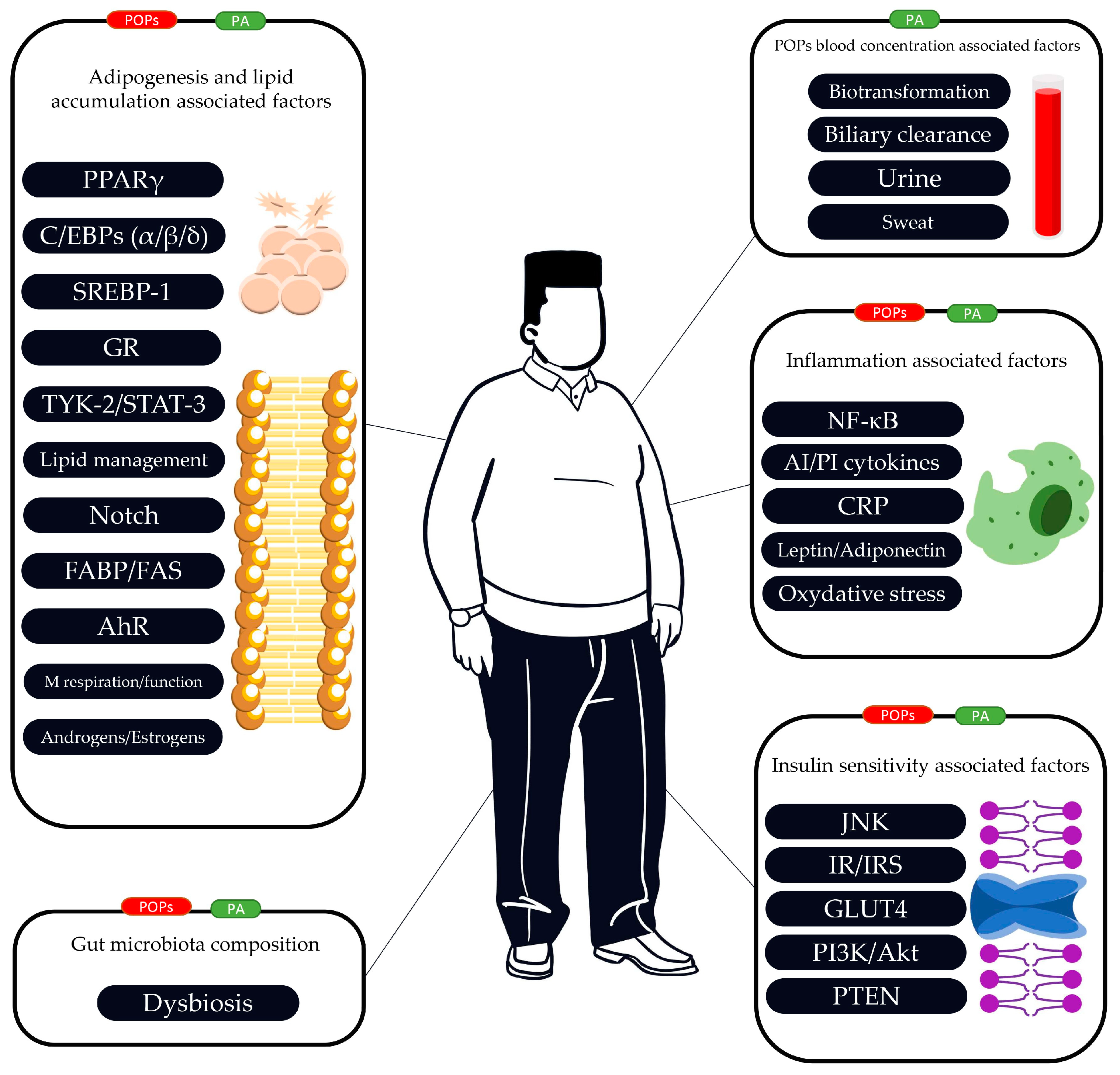
Figure 2.
The possible relationships between bariatric surgery, persistent organic pollutants (POPs) and physical activity (PA). Physical activity as well as bariatric surgery can promote POPs adipose tissue degradation, resulting in an increase of several determinants of POPs hazard. The balance between POPs release following adipose tissue degradation and POPs excretion/transformation may depend on the physical activity modality (aerobic vs. resistance training).
Figure 2.
The possible relationships between bariatric surgery, persistent organic pollutants (POPs) and physical activity (PA). Physical activity as well as bariatric surgery can promote POPs adipose tissue degradation, resulting in an increase of several determinants of POPs hazard. The balance between POPs release following adipose tissue degradation and POPs excretion/transformation may depend on the physical activity modality (aerobic vs. resistance training).
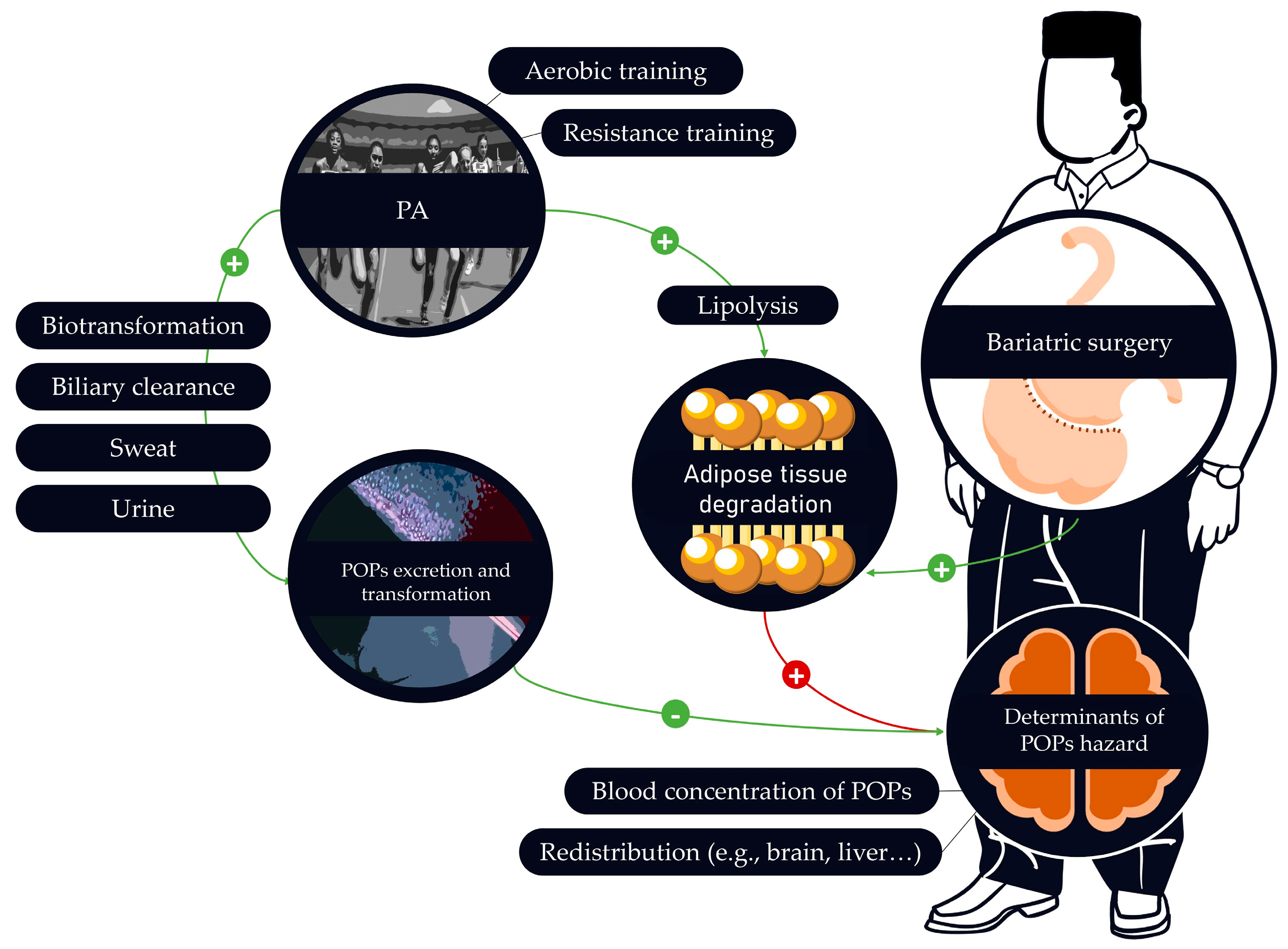
Figure 3.
The hypothetical adverse consequences of using aerobic training shortly after bariatric surgery. Aerobic training used shortly after bariatric surgery may increase lipolysis and POPs release into the blood.
Figure 3.
The hypothetical adverse consequences of using aerobic training shortly after bariatric surgery. Aerobic training used shortly after bariatric surgery may increase lipolysis and POPs release into the blood.
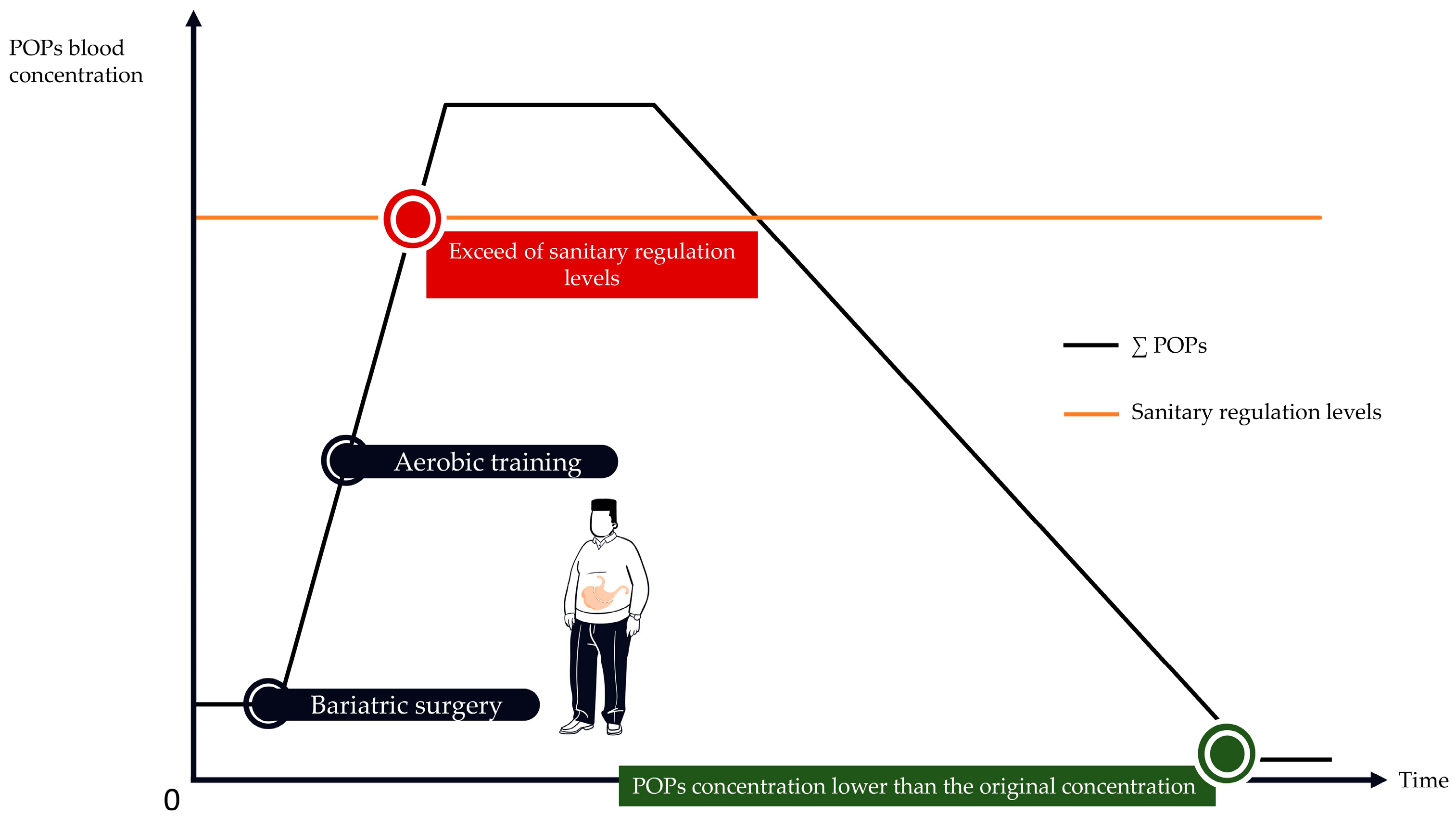
Figure 4.
Hypothetical utilization of resistance training and aerobic exercise following bariatric surgery to protect the individual from POPs blood release.
Figure 4.
Hypothetical utilization of resistance training and aerobic exercise following bariatric surgery to protect the individual from POPs blood release.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



