
Submitted:
22 November 2023
Posted:
22 November 2023
You are already at the latest version
Abstract
Phosphate (Pi) starvation is a critical factor limiting crop growth, development, and productivity. Rice (Oryza sativa) R2R3-MYB transcription factors function in the transcriptional regulation of plant responses to various abiotic stresses and micronutrient deprivation, but little is known about their roles in Pi starvation signaling and Pi homeostasis. Here we identified the R2R3-MYB transcription factor gene OsMYB58, which shares high sequence similarity with AtMYB58. OsMYB58 expression was induced more strongly by Pi starvation than by other micronutrient deficiencies. Overexpressing OsMYB58 in Arabidopsis thaliana and rice inhibited plant growth and development under Pi-deficiency conditions. In addition, overexpression of OsMYB58 in plants exposed to Pi deficiency strongly affected root development, including seminal root, lateral root, and root hair formation. Overexpressing OsMYB58 strongly decreased the expression of the rice microRNAs, OsmiR399a and OsmiR399j. By contrast, overexpressing OsMYB58 strongly increased the expression of rice PHOSPHATE 2 (OsPHO2), whose expression is repressed by miR399 during Pi starvation signaling. OsMYB58 functions as a transcriptional repressor of the expression of its target genes, as determined by a transcriptional activity assay. These results demonstrate that OsMYB58 negatively regulates OsmiR399-dependent Pi starvation signaling by enhancing OsmiR399s expression.
Keywords:
MYB transcription factor
; phosphate starvation signaling
; OsMYB58
; OsmiR399
; PHR-miR399-PHO2
1. Introduction
MYB transcription factors, which are widespread in crops, play important roles in regulating plant signaling pathways in response to various stresses [1,2]. MYB domain repeats (R) in transcription factors are divided into the R1, R2, and R3 types based on amino acid sequence similarity [3]. The R region comprises 51-52 conserved amino acid residues arranged in a helix-turn-helix (HTH) structure that includes 3 tryptophans or other hydrophobic residues, spaced 18 amino acids apart [4]. MYB transcription factor classes are divided into the 1R-MYB, 2R-MYB (R2R3-MYB), 3R-MYB, and 4R-MYB classes [5]. Functional studies have revealed that R2R3-MYB transcription factors function in a wide variety of plant responses, including stress tolerance, plant hormonal regulation, metabolism, development, and cell differentiation [1,3].
R2R3-MYB transcription factors in plants function in transcriptional regulation in response to diverse abiotic stresses, such as salt stress, drought, cold stress, abscisic acid (ABA) treatment, oxidative damage, and phosphate (inorganic phosphate; Pi) deficiency [3,6,7,8,9,10]. Overexpressing OsMYB2 in rice (Oryza sativa L.) enhanced plant tolerance to salt, cold, and drought stress [6]. TaMYB31-B in hexaploid wheat (Triticum aestivum L.) positively regulates plant responses to drought by regulating the expression of wax biosynthesis genes and drought-responsive genes [7]. TcMYB29a in Taxus chinensis is a transcriptional activator that functions in ABA-mediated signaling by binding to MYB-recognizing element (MRE) motifs in the promoters of taxol-biosynthesis-related genes [8]. AtMYB30 in Arabidopsis thaliana (Arabidopsis) is involved in systemic reactive oxygen species (ROS) signaling in response to high-light stress [9]. The transcription factor TaMYB4-7D in T. aestivum mediates Pi uptake and translocation under Pi-deficiency conditions by binding to four MBS motifs in the promoter region of TaPHT1;9-4B [10]. The interaction of PuMYB40 and PuWRKY75 in Populus ussuriensis positively regulates adventitious root formation under low-Pi conditions [11].
Rice contains 99 R2R3-MYB transcription factor genes, accounting for 52.1% of the 190 MYB genes [12]. A few R2R3-MYB transcription factors in rice were recently shown to be associated with Pi-starvation responses. For instance, OsMYB2P-1 functions as a key Pi-dependent regulator of Pi-starvation signaling by controlling the expression of Pi transporter (PT) genes, such as OsPT2, OsPT6, OsPT8, and OsPT10 [13]. OsMYB4P acts as a transcriptional activator of Pi-homeostasis-related genes to increase Pi acquisition in rice [14]. Overexpressing OsMYB4P strongly induced the expression of PT genes, including OsPT1, OsPT2, OsPT4, OsPT7, and OsPT8, in shoots under Pi-deficiency conditions [14]. OsMYB1 mediates root elongation in the Pi starvation responses and acts as an important regulator of both Pi-starvation signaling and gibberellic acid (GA) biosynthesis [15]. Overexpressing OsMYB5P enhanced plant tolerance of Pi deficiency by controlling the transcription of PT genes [16].
Pi is an important macronutrient for plant growth and development and a component of various biological molecules such as nucleic acids, membrane lipids, and ATP [17,18]. These molecules function as essential regulators of Pi-starvation signaling and cellular mechanisms to systemically enhance Pi uptake, transport, and utilization [19]. Pi-starvation signaling is mediated by the PHR-miR399-PHO2 molecular network [17,20,21]. The rice miRNA399 (OsmiR399) family comprises 11 members (OsmiR399a to k) that are important regulators of Pi-starvation signaling [17]. During exposure to Pi-deficiency stress, the expression of OsmiR399s is widely induced, whereas the expression of their target gene OsPHO2 ultimately decreases [17,21]. OsPHR1 and OsPHR2, the homologs of Arabidopsis PHR1, encode transcription factors that positively regulate the expression of OsIPS1 [20,22]. OsIPS1 represses the activity of OsmiR399s via a target mimicry mechanism [20,22]. In Pi-starvation signaling, the PHR-miR399-PHO2 systemic regulatory network influences the activity of two important Pi transporters, PHO1 and PT2, to help maintain Pi homeostasis [23,24]. In addition, the OsPHR2-OsmiR827-OsSPX regulatory network is associated with Pi-starvation signaling in rice [20,22]. OsmiR827 is highly expressed under Pi deficiency, which induces OsmiR827 expression, thereby decreasing the expression of OsSPX genes [22].
Pi deficiency leads to changes in plant morphology, physiology, and biochemistry by decreasing the acquisition and utilization efficiency of Pi from the soil [25,26]. Although the functions of many R2R3-MYB transcription factors in plant responses to abiotic stress have been extensively investigated, more research is needed on their roles in Pi-deficiency responses. In this study, we characterized the functions of rice OsMYB58 in PHR-miR399-PHO2-dependent Pi starvation signaling. Through physiological and biological analyses, we demonstrated that OsMYB58, which functions as a transcriptional repressor essential for OsmiR399s expression, plays a key role in maintaining Pi homeostasis. Understanding the role of OsMYB58 in PHR-miR399-PHO2-dependent mechanisms during Pi-starvation signaling will help enhance crop yields and growth in nutrient-poor soils.
2. Results
2.1. OsMYB58 expression is enhanced under phosphate-deficiency conditions.
We previously isolated and characterized several R2R3-type MYB transcription factors that function in phosphate starvation signaling [14,16]. In the current study, to identify Pi-deficiency-related R2R3-type MYB transcription factors in rice, we investigated the expression levels of several R2R3-type OsMYB transcription factor genes in wild-type rice plants under various nutrient deficiency conditions. Among the many R2R3-type MYB genes with high expression under nutrient-deficiency conditions, OsMYB58 was highly expressed in rice under Pi-deficiency conditions (Figure 1a). The full protein sequence of OsMYB58 (LOC_Os02g46780) shared high levels of sequence similarity with two Arabidopsis two genes, AtMYB58 (AT1G16490; 60.5% similarity) and AtMYB63 (AT1G79180; 59.1% similarity) (Supplementary Figure S1a) [27]. The R2R3 MYB transcription factor OsMYB58 was classified in the same clade as OsMYB63 (LOC_Os04g50770) by phylogenetic analysis (Supplementary Figure S1b). OsMYB58 is homologous to OsMYB63, with 72% sequence similarity, as revealed using the Phytozome database (https://phytozome-next.jgi.doe.gov/) [28]. To investigate the changes in OsMYB58 expression in rice shoots and roots, we extracted total RNA from the shoots and roots of plants under high-Pi and low-Pi conditions. The expression levels of OsMYB58 were similar in shoots and roots under high-Pi conditions, whereas it was expressed at higher levels in roots than shoots under low-Pi conditions (Figure 1b). In the shoot, OsMYB58 expression showed a clear increase, followed by a decrease, in the course of 7 days of exposure to low Pi (Figure 1c). On the other hand, OsMYB58 expression consistently increased in roots under low-Pi conditions (Figure 1d). These results suggest that OsMYB58 is associated with Pi-starvation signaling.
2.2. Heterologously overexpressing OsMYB58 in Arabidopsis disrupts Pi homeostasis in response to Pi deficiency.
To explore the roles of OsMYB58 in the Pi-starvation response, we heterologously overexpressed OsMYB58 in Arabidopsis Col-0 plants (Supplementary Figure S2a). Using RT-PCR analysis, we selected two lines of OsMYB58-overexpressing Arabidopsis plants (OsMYB58-AraOX) with different OsMYB58 expression levels for further analysis (Supplementary Figure S2b). To investigate the phenotypic alterations of wild-type Col-0 and OsMYB58-AraOX plants in response to Pi deficiency, we subjected 4-day-old seedlings to Pi deficiency for 7 days. The shoots and roots of OsMYB58-AraOX plants grew more slowly than those of Col-0 plants under both high-Pi and low-Pi conditions (Figure 2a). In addition, the shoots and roots of OsMYB58-AraOX plants weighed less than those of Col-0 plants under both high- and low-Pi conditions (Figure 2c and 2d). The root architecture of OsMYB58-AraOX was significantly altered at both Pi levels (Figure 2b), with reductions in the lengths of primary and lateral roots, as well as the number of lateral roots (Figure 2b, 2e, and 2f). Pi concentrations in both shoots and roots were significantly lower in OsMYB58-AraOX plants compared to Col-0 (Figure 2g and 2h). These results indicate that the phenotypic alterations in Arabidopsis plants under both low-Pi and high-Pi conditions in response to OsMYB58 overexpression were due to disturbed Pi homeostasis.
2.3. OsMYB58 modulates plant growth during the Pi-deficiency response.
To characterize the role of OsMYB58 in the Pi-starvation response, we generated two types of transgenic rice plants, OsMYB58 overexpression plants (OsMYB58-OX; Supplementary Figure S3a) and T-DNA tagged knock-out mutants (OsMYB58-KO; Supplementary Figure S4a), via Agrobacterium-mediated transformation (Supplementary Figure S3b). The expression levels of OsMYB58 were similar in most OsMYB58-OX lines, as revealed by RT-PCR analysis; we randomly selected line #5 for further analysis (Supplementary Figure S3c). We selected three OsMYB58-KO T1 lines based on their growth on medium containing hygromycin due to the presence of the hygromycin phosphotransferase (HPH) gene. We performed genotyping PCR (Supplementary Figure S4b) and RT-PCR (Supplementary Figure S4c) analysis of these three selected OsMYB58-KO lines to confirm the presence of the T-DNA insertion and used line #6 in all our experiments.
To investigate the phenotypic alterations due to OsMYB58 overexpression or knockout, we exposed 7-day-old wild-type (WT), OsMYB58-OX, and OsMYB58-KO plants to either high-Pi or low-Pi conditions for 7 days. Shoot and root growth were strongly inhibited in OsMYB58-OX plants compared to WT and OsMYB58-KO plants under both high-Pi and low-Pi conditions (Figure 3a). The fresh weights of shoots and roots were significantly reduced in OsMYB58-OX plants compared to WT under both conditions (Figure 3b and 3c). By contrast, the fresh weights of shoots or roots were slightly reduced in OsMYB58-KO plants compared to WT regardless of Pi concentration (Figure 3b and 3c). The shoots and primary roots were significantly shorter in OsMYB58-OX than in WT under both high- and low-Pi conditions (Figure 3d and 3e). The shoot length was reduced in OsMYB58-KO compared to the WT, while the primary root length was similar to the WT, under both sets of conditions (Figure 3d and 3e). These results indicate that OsMYB58 affects plant growth and developmental processes regardless of the Pi concentrations in plants.
2.4. OsMYB58 represses root development in rice.
Pi deficiency significantly suppresses primary root growth by disrupting the root meristem, with effects such as reducing cell elongation and interrupting cell proliferation and cell differentiation [29]. To investigate the changes in root architecture in response to OsMYB58 overexpression or knockout, we transferred 3-day-old WT, OsMYB58-OX, and OsMYB58-KO seedlings to growth medium containing high or low Pi concentrations and allowed them to grow for 7 days. The root architecture of OsMYB58-KO was similar to that of the WT, whereas root growth was dramatically reduced in OsMYB58-OX under both high-Pi and low-Pi conditions (Figure 4a). An overall comparison of roots, including seminal roots, lateral roots, and root hairs, indicated that the lengths and numbers of these structures were lower in OsMYB58-OX than in the WT, while those in OsMYB58-KO were similar to the WT, under both high- and low-Pi conditions (Figure 4b-g).
To investigate the changes in Pi concentrations in plants in response to OsMYB58 overexpression or knockout, we treated 7-day-old WT, OsMYB58-OX, and OsMYB58-KO seedlings with high Pi and low Pi. The Pi levels in shoots and roots were lower in OsMYB58-OX than in the WT under both high-Pi and low-Pi conditions (Figure 5a and 5b). However, the Pi concentrations in shoots and roots of OsMYB58-KO plants were similar to those of WT plants under both conditions (Figure 5a and 5b). These results suggest that OsMYB58 modulates Pi levels by reducing Pi uptake in plants through alteration of root architecture during Pi-deficiency responses.
2.5. OsMYB58 is associated with Pi-responsive gene expression.
Pi-responsive genes and Pi transporters (PTs) improve plant development and growth to help maintain Pi homeostasis via Pi-starvation signaling [30]. To investigate whether Pi-responsive and PT genes are regulated by OsMYB58, we treated 7-day-old WT, OsMYB58-OX, and OsMYB58-KO seedlings with high Pi and low Pi and subjected samples from the seedlings to quantitative RT-PCR (qRT-PCR). OsmiR399a, OsmiR399j, OsIPS1, OsPT2, and OsPT4 transcript levels were significantly reduced in the shoots and roots of OsMYB58-OX plants compared to the WT (Figure 6). In OsMYB58-KO plants, OsmiR399a, OsmiR399j, and OsIPS1 were more highly expressed in shoots and roots compared to the WT (Figure 6a-6c). The transcript levels of OsPHO2, the target genes of OsmiR399, significantly increased in shoots and roots of OsMYB58-OX but slightly decreased in OsMYB58-KO compared to the WT (Figure 6d). The expression levels of OsPT2 and OsPT4 in shoots and roots of OsMYB58-OX plants were similar to those of the WT (Figure 6e and 6f). Overall, the effects of OsMYB58 on Pi-responsive and PT gene expression were more varied under low-Pi conditions than under high-Pi conditions (Figure 6). These results suggest that OsMYB58 functions in Pi starvation signaling by regulating the transcription of Pi-responsive genes, especially OsmiR399, an important component of PHR-miR399-PHO2-mediated mechanisms.
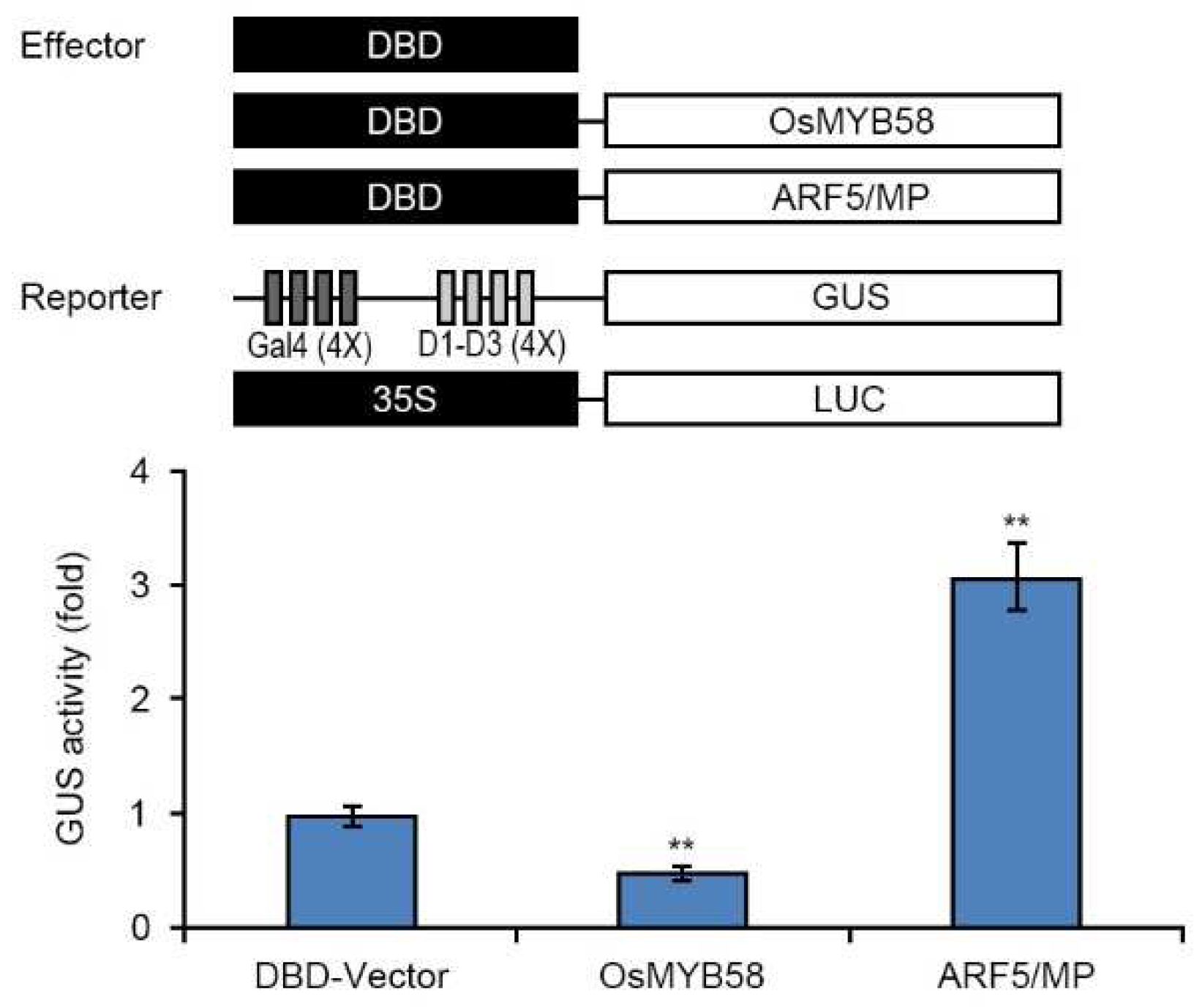
Finally, to test the transcriptional activity of OsMYB58, we performed a transcriptional activity assay via transient expression using Arabidopsis protoplasts. We generated an effector construct harboring OsMYB58 with the yeast GAL4 DNA binding domain (DBD) and a reporter construct harboring constitutively expressed GUS with four upstream GAL4 DNA binding sites, Gal4 (4X)-D1-D3 (4X); the LUC (luciferase) gene driven by the constitutive 35S promoter was used as an internal control (Figure 7). We co-transfected Arabidopsis protoplasts with various combinations of effector and reporter constructs and measured GUS activity. The GUS activity was significantly lower in protoplasts harboring the effector and reporter constructs compared to protoplasts harboring empty vector (DBD-vector) alone (Figure 7). These results indicate that OsMYB58 functions as a transcriptional repressor in Pi starvation signaling.
3. Discussion
Pi deficiency in soil has adverse effects on plant growth and development, such as damaging root architecture, inhibiting leaf development, and impairing fruit production via Pi starvation signaling [29,31]. Although numerous transcription factors are reported to function in Pi starvation signaling, few functional studies have focused on the roles of rice MYB transcription factors in this process. To date, only five MYB transcription factors have been shown to function in Pi starvation signaling in rice: OsPHR2, OsMYB1, OsMYB2P-1, OsMYB4P, and OsMYB5P [15,16,32]. Here we characterized the role of OsMYB58 in Pi starvation signaling, demonstrating that it acts as a transcriptional repressor that regulates the expression of Pi-responsive genes. OsMYB58 expression strongly increased in response to Pi deficiency. Overexpressing OsMYB58 inhibited plant growth and root development by impairing Pi homeostasis in shoots and roots of Arabidopsis and rice. OsmiR399a and OsmiR399j levels strongly decreased in response to OsMYB58 overexpression in roots under Pi-deficiency conditions. Our results demonstrate that OsMYB58 plays an important role in the PHR-miR399-PHO mechanism for root-to-shoot translocation of Pi.
3.1. OsMYB58 is associated with plant response to Pi deficiency.
Plant MYB transcription factors have important roles in regulating many processes in plants, including growth and development, secondary metabolism, phytohormone signaling, pathogen resistance, and abiotic stress tolerance [33]. Many R2R3 MYB transcription factors are associated with transcriptional regulation in response to various environmental stresses, such as salt, drought, cold, and nutrient deficiency [1,2,3]. Five R2R3 MYB transcription factors have been shown to regulate the Pi-deficiency response in rice [13,14,15,16,23,32]. However, the roles of R2R3 MYB transcription factors in Pi starvation signaling to maintain Pi homeostasis in rice are poorly understood. Here we demonstrated that OsMYB58 is strongly expressed in shoots and roots under Pi deficiency compared to other nutrient deficiencies (Figure 1). The OsMYB58 protein sequence, which contains an R2R3-MYB domain at the N-terminal region, is similar to that of OsMYB63 (LOC_Os04g50770) in rice and AtMYB58 (AT1G16490)/AtMYB63 (AT1G79180) in Arabidopsis (Supplementary Figure 1). OsMYB58 was recently shown to regulate the expression of the cellulose synthase gene OsCesA7 [28]. The correlation network between Pi starvation signaling and lignin biosynthesis by OsMYB58 has not yet been studied, and it is imperative to continue the functional study of OsMYB58 using that and other approaches.
3.2. OsMYB58 inhibits plant growth and development by disrupting Pi homeostasis.
Overexpressing MYB transcription factor genes in rice reduced the damage caused by Pi deficiency to plant growth and shoot and root development by enhancing Pi uptake [13,14,16,23]. Plants overexpressing OsPHR2, OsMYB2P-1, OsMYB4P, and OsMYB5P showed common phenotypes, including increased shoot, primary root, seminal root, and lateral root growth, due to increased Pi levels under Pi-deficiency conditions [13,14,16,23]. By contrast, in the current study, OsMYB58-overexpressing Arabidopsis and rice plants were hypersensitive, as compared to the WT, to both low and high Pi levels in terms of shoot growth and root architecture (Figure 2, Figure 3 and Figure 4). Overexpressing OsMYB58 disrupted Pi homeostasis in rice by inhibiting Pi uptake in shoots and roots (Figure 5). These results contrast with those from other studies of Pi-responsive MYB transcription factors. OsSPX1 and OsSPX2, which are not MYB transcription factors, negatively regulate Pi levels, as their overexpression reduced Pi concentrations in shoots [34]. Our results demonstrate that OsMYB58 acts as a negative regulator of plant growth and development based on cellular Pi concentration.
3.3. OsMYB58 is a negative regulator of Pi starvation signaling.
During Pi starvation signaling in rice, the pathway regulating Pi homeostasis is mediated by two major pathways: PHR2-miR399-PHO2 and PHR2-miR827-SPX/MSF [22,35,36]. The PHR2-miR399-PHO2 pathway modulates Pi-regulated plant phenotypes and Pi accumulation, while the PHR2-miR827-SPX/MSF pathway only regulates Pi accumulation, without inducing phenotypic alterations in response to Pi deficiency [22]. OsmiR399-dependent OsPHO2 expression influences Pi uptake and remobilization to help maintain Pi homeostasis [37,38]. OsPHO2 acts as a positive regulator of OsPT2 gene expression in Pi starvation signaling [37,38]. We demonstrated that the expression of OsmiR399a and OsmiR399j significantly decreased in OsMYB58-OX plants, whereas the expression of OsPHO2 strongly increased, under both high-Pi and low-Pi conditions (Figure 6). In addition, OsMYB58 acted as a transcriptional repressor in a transcriptional activity assay (Figure 7). The decline in Pi levels in OsMYB58-OX plants was likely caused by the reduced expression of OsPT2 and OsPT4 (Figure 5 and Figure 6). These findings demonstrate that OsMYB58 modulates cellular Pi concentrations in shoots and roots through the PHR2-miR399-PHO pathway during Pi starvation signaling. Thus, OsMYB58 functions in OsmiR399-dependent Pi-starvation signaling by negatively regulating Pi homeostasis. Our findings demonstrate that OsMYB58 functions as a negative regulator of OsmiR399-dependent Pi-starvation signaling.
4. Materials and Methods
4.1. In silico analysis
The sequences of OsMYB58 (Os02g46780) homologs were obtained from NCBI (http://blast.ncbi.nlm.gov/Blast,cgi). Multiple protein sequence alignment was performed with the Clustal Omega program (https://www.ebi.ac.uk/Tools/msa/clustalo/) and MEGA X (https://www.megasoftware.net/).
4.2. Generation of MYB58 transgenic plants
The OsMYB58:pH7WG2D.1 (OsMYB58-OX) constructs were introduced into Agrobacterium tumefaciens (EHA105 and GV2260) by electroporation. Transformation of rice and Arabidopsis was performed as previously described [14]. Dr. Ki Hong Jung (Kyung Hee University) provided OsMYB58 T-DNA tagged knock-out mutant (OsMYB58-KO) rice plants.
4.3. Plant materials and growth conditions
Physiological experiments and the generation of transgenic plants were performed using Oryza sativa L. ‘Dongjin’ and Arabidopsis thaliana Col-0 plants. Hydroponic experiments were performed in either a high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) medium as described previously [14]. Rice or Arabidopsis plants were cultivated in growth chambers at 32˚C or 22˚C, respectively, under a 16 h light/8 h dark cycle.
4.4. Northern bolt and gene expression analysis
Total RNA was isolated from 100 mg of rice tissue using TRIzol reagent (Sigma-Aldrich) according to the manufacturer’s instructions for northern blotting and quantitative reverse-transcription PCR (qRT-PCR). The 20 μM sample of total RNA was separated on a 1.2% (w/v) denaturing formaldehyde agarose gel via gel electrophoresis, transferred to a Hybrid-N+ membrane (GE Healthcare), and cross-linked using a commercial UV-light crosslinking instrument (UVP). The membrane was hybridized overnight with a [32P]-dCTP-labeled probe (Stratagene) in a solution containing 20% (w/v) SDS, 20X SSPE, 100 g/L PEG (8,000 mwt), 250 mg/L heparin, and 10 ml/L herring sperm DNA (10 mg/ml) at 65°C. Probes for the OsMYB58 gene were generated using a primer set designed for its open reading frame. The filters were washed twice in 2X SSC and 0.2% (w/v) SDS at 65°C for 10 min, twice in 1X SSC and 0.2% (w/v) SDS at 65°C for 10 min, and once in 0.1X SSC and 0.2%(w/v) SDS at 65°C for 20 min. The dried membrane was placed on X-ray film at -72°C for 1 day and then developed and exposed.
For qRT-PCR, cDNA was synthesized using 0.5 μg of total RNA with a 1st Strand cDNA synthesis Kit (Takara, Dalian, CA, Japan) and subjected to RT-PCR analysis to measure gene expression using qRT-PCR Detection Systems (Bio-Rad, Hercules, CA, USA) following a standard protocol. The primer sequences were designed using NCBI Primer-BLAST and Primer3 (https://bioinfo.ut.ee/primer3/) and are provided in Table S1. PCR (10 µl reactions) was performed using TB Green Premix Ex Taq™ II (Takara, Dalian, CA, Japan) as follows: 95°C for 5 min, and 40 cycles of 95°C for 15 sec, 60℃ for 15 sec, and 72°C for 15 sec. The relative expression levels in all the samples were automatically calculated and analyzed three times using a CFX384 Real-time PCR Detection System and CFX Manager software (Bio-Rad, Hercules, CA, USA) following a standard protocol.
4.5. Measurement of inorganic Pi content in plants
After measurement of fresh weight, samples were frozen and dried at 80˚C for 3 days. Inorganic Pi contents were measured as previously described [14].
4.6. Transcriptional activity assay via transient expression of OsMYB58 in Arabidopsis protoplasts
To investigate the transcriptional activity of OsMYB58, we performed a transcriptional activity assay by introducing constructs harboring reporter genes driven by the OsMYB58 promoter into Arabidopsis protoplasts prepared from leaf tissues by PEG-mediated transformation and measuring reporter activity, as described previously [14,16].
5. Conclusions
In summary, we characterized the role of the transcription factor OsMYB58 in Pi-starvation signaling. OsMYB58 was dramatically upregulated under Pi deficiency compared to other nutrient-deficiency conditions. Overexpressing OsMYB58 in Arabidopsis and rice strongly inhibited plant growth and root development, whereas OsMYB58-KO plants showed enhanced growth and development under both high- and low-Pi conditions. In addition, OsMYB58-overexpressing transgenic Arabidopsis and rice showed greatly reduced Pi concentrations in both shoots and roots. The transcript levels of OsmiR399a and OsmiR399j strongly increased in shoots and roots, whereas OsPHO2 transcript levels decreased, in plants overexpressing OsMYB58. OsMYB58 is a transcriptional repressor, as determined by a transcriptional activity assay. Our results suggest that OsMYB58 negatively regulates Pi-starvation signaling in rice in an OsmiR399-dependent manner.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figure S1. Sequence alignment and phylogenetic tree analysis of R2R3-type MYB transcription factors in Arabidopsis and rice. Supplementary Figure S2. Diagram of DNA construct and RT-PCR analysis in OsMYB58 overexpressing Arabidopsis plants. Supplementary Figure S3. Generation of OsMYB58 overexpressing rice plants. Supplementary Figure S4. Generation of OsMYB58 T-DNA tagging knock-out mutant rice plants. Supplementary Table S1. Lists of primers in this study.
Author Contributions
Conceptualization, D.H.K. and D.B.; methodology, W.T.Y.; formal analysis, D.B., W.T.Y., S.H., H.J.K., S.M.; investigation, D.B., W.T.Y., S.H.; resources, K.H.J.; data curation, D.B., W.T.Y. S.H.; writing—original draft preparation, D.B. and W.T.Y.; writing—review and editing, D.H.K.; supervision, D.H.K..; project administration, D.H.K.; funding acquisition, D.H.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (grant number 2020R1A6A1A03047729) and the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (No.2021R1F1A1056993).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, J; Han, G; Sun, C; Sui, N. Research advances of MYB transcription factors in plant stress resistance and breeding. Plant Signal Behav. 2019, 14, 1613131. [Google Scholar] [CrossRef] [PubMed]
- Ambawat, S.; Sharma, P.; Yadav, R. N.; Yadav, C. R. MYB transcription factor genes as regulators for plant responses: an overview. Physiol Mol Biol Plants. 2013, 19, 307–321. [Google Scholar] [CrossRef]
- Wu, Y.; Wen, J.; Xia, Y.; Zhang, L.; Du, H. Evolution and functional diversification of R2R3-MYB transcription factors in plants. Hortic Res. 2022, 9, uhac058. [Google Scholar] [CrossRef]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr Opin Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Dai, X.; Zhang, W.H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J Exp Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cheng, X.; Liu, X.; Wu, H.; Bi, H.; Xu, H. The Wheat MYB Transcription Factor TaMYB31 is involved in drought stress responses in Arabidopsis. Front Plant Sci. 2018, 9, 1426. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Xu, L.; Li, L.; Wan, W.; Jiang, J. TcMYB29a, an ABA-responsive R2R3-MYB transcriptional factor, upregulates taxol biosynthesis in Taxus chinensis. Front Plant Sci. 2022, 13, 804593. [Google Scholar] [CrossRef] [PubMed]
- Fichman, Y.; Zandalinas, S.I.; Sengupta, S.; Burks, D.; Myers, R.J. Jr.; Azad, R.K.; Mittler, R. MYB30 orchestrates systemic reactive oxygen signaling and plant acclimation. Plant Physiol. 2020, 184, 666–675. [Google Scholar] [CrossRef]
- Wang, P.; Li, G.; Li, G.; Yuan, S.; Wang, C.; Xie, Y.; Guo, T.; Kang, G.; Wang, D. TaPHT1;9-4B and its transcriptional regulator TaMYB4-7D contribute to phosphate uptake and plant growth in bread wheat. New Phytol. 2021, 231, 1968–1983. [Google Scholar] [CrossRef]
- Wang, H.; Pak, S.; Yang, J.; Wu, Y.; Li, W.; Feng, H.; Yang, J.; Wei, H.; Li, C. Two high hierarchical regulators, PuMYB40 and PuWRKY75, control the low phosphorus driven adventitious root formation in Populus ussuriensis. Plant Biotechnol J. 2022, 20, 1561–1577. [Google Scholar] [CrossRef]
- Kang, L.; Teng, Y.; Cen, Q.; Fang, Y.; Tian, Q.; Zhang, X.; Wang, H.; Zhang, X.; Xue, D. Genome-wide identification of R2R3-MYB transcription factor and expression analysis under abiotic stress in rice. Plants (Basel). 2022, 11, 1928. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Wang, Y.; Yang, A.; Zhang, W.H. OsMYB2P-1, an R2R3 MYB transcription factor, is involved in the regulation of phosphate-starvation responses and root architecture in rice. Plant Physiol. 2012, 159, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.T.; Baek, D.; Yun, D.J.; Hwang, W.H.; Park, D.S.; Nam, M.H.; Chung, E.S.; Chung, Y.S.; Yi, Y.B.; Kim, D.H. Overexpression of OsMYB4P, an R2R3-type MYB transcriptional activator, increases phosphate acquisition in rice. Plant Physiol Biochem. 2014, 80, 259–267. [Google Scholar] [CrossRef]
- Gu, M.; Zhang, J.; Li, H.; Meng, D.; Li, R.; Dai, X.; Wang, S.; Liu, W.; Qu, H.; Xu, G. Maintenance of phosphate homeostasis and root development are coordinately regulated by MYB1, an R2R3-type MYB transcription factor in rice. J Exp Bot. 2017, 68, 3603–3615. [Google Scholar] [CrossRef]
- Yang, W.T.; Baek, D.; Yun, D.J.; Lee, K.S.; Hong, S.Y.; Bae, K.D.; Chung, Y.S.; Kwon, Y.S.; Kim, D.H.; Jung, K.H.; Kim, D.H. Rice OsMYB5P improves plant phosphate acquisition by regulation of phosphate transporter. PLoS One. 2018, 13, e0194628. [Google Scholar] [CrossRef]
- Hu, B.; Wang, W.; Deng, K.; Li, H.; Zhang, Z.; Zhang, L.; Chu, C. MicroRNA399 is involved in multiple nutrient starvation responses in rice. Front Plant Sci. 2015, 6, 188. [Google Scholar] [CrossRef]
- Cuyas, L.; David, P.; de Craieye, D.; Ng, S.; Arkoun, M.; Plassard, C.; Faharidine, M.; Hourcade, D.; Degan, F.; Pluchon, S.; Nussaume, L. Identification and interest of molecular markers to monitor plant Pi status. BMC Plant Biol. 2023, 23, 401. [Google Scholar] [CrossRef]
- Wang, F.; Deng, M.; Xu, J.; Zhu, X.; Mao, C. Molecular mechanisms of phosphate transport and signaling in higher plants. Semin Cell Dev Biol. 2018, 74, 114–122. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, H.; Huang, H.; Duan, K.; Wu, Z.; Wu, P. Regulation of OsSPX1 and OsSPX3 on expression of OsSPX domain genes and Pi-starvation signaling in rice. J Integr Plant Biol. 2009, 51, 663–674. [Google Scholar] [CrossRef]
- Bari, R.; Datt Pant, B.; Stitt, M.; Scheible, W.R. PHO2, microRNA399, and PHR1 define a phosphate-signaling pathway in plants. Plant Physiol. 2006, 141, 988–999. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, S.; Sun, C.; Xu, Y.; Chen, Y.; Yu, C.; Qian, Q.; Jiang, D.A.; Qi, Y. Auxin response factor (OsARF12), a novel regulator for phosphate homeostasis in rice (Oryza sativa). New Phytol. 2014, 201, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Jiao, F.; Wu, Z.; Li, Y.; Wang, X.; He, X.; Zhong, W.; Wu, P. OsPHR2 is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants. Plant Physiol. 2008, 146, 1673–1686. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zhu, C.; Li, F.; Tang, J.; Wang, Y.; Lin, A.; Liu, L.; Che, R.; Chu, C. LEAF TIP NECROSIS1 plays a pivotal role in the regulation of multiple phosphate starvation responses in rice. Plant Physiol. 2011, 156, 1101–1115. [Google Scholar] [CrossRef]
- Raghothama, K.G. Phosphate acquisition. Annu Rev Plant Physiol Plant Mol Biol. 1999, 50, 665–693. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, B.; Zheng, S.; Zhang, X.; Wang, X.; Dong, W.; Xie, Q.; Wang, G.; Xiao, Y.; Chen, F.; Yu, N.; Wang, E. A phosphate starvation response-centered network regulates mycorrhizal symbiosis. Cell. 2021, 184, 5527–5540.e18. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Kondo, M.; Aya, K.; Miyao, A.; Sato, Y.; Antonio, B.A.; Namiki, N.; Nagamura, Y.; Matsuoka, M. Identification of transcription factors involved in rice secondary cell wall formation. Plant Cell Physiol. 2013, 54, 1791–1802. [Google Scholar] [CrossRef]
- Noda, S.; Koshiba, T.; Hattori, T.; Yamaguchi, M.; Suzuki, S.; Umezawa, T. The expression of a rice secondary wall-specific cellulose synthase gene, OsCesA7, is directly regulated by a rice transcription factor, OsMYB58/63. Planta. 2015, 242, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Péret, B.; Desnos, T.; Jost, R.; Kanno, S.; Berkowitz, O.; Nussaume, L. Root architecture responses: in search of phosphate. Plant Physiol. 2014, 166, 1713–1723. [Google Scholar] [CrossRef]
- Péret, B.; Clément, M.; Nussaume, L.; Desnos, T. Root developmental adaptation to phosphate starvation: better safe than sorry. Trends Plant Sci. 2011, 16, 442–450. [Google Scholar] [CrossRef]
- Rouached, H.; Arpat, A.B.; Poirier, Y. Regulation of phosphate starvation responses in plants: signaling players and cross-talks. Mol Plant. 2010, 3, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Chen, A.; Sun, S.; Xu, G. Complex Regulation of Plant Phosphate Transporters and the Gap between Molecular Mechanisms and Practical Application: What Is Missing? Mol Plant. 2016, 9, 396–416. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, A.; Smita, S.; Lenka, S.K.; Rajwanshi, R.; Chinnusamy, V.; Bansal, K.C. Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis. BMC Genomics. 2012, 13, 544. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ruan, W.; Shi, J.; Zhang, L.; Xiang, D.; Yang, C.; Li, C.; Wu, Z.; Liu, Y.; Yu, Y.; Shou, H.; Mo, X.; Mao, C.; Wu, P. Rice SPX1 and SPX2 inhibit phosphate starvation responses through interacting with PHR2 in a phosphate-dependent manner. Proc Natl Acad Sci U S A. 2014, 111, 14953–14958. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ying, S.; Huang, H.; Li, K.; Wu, P.; Shou, H. Involvement of OsSPX1 in phosphate homeostasis in rice. Plant J. 2009, 57, 895–904. [Google Scholar] [CrossRef]
- Wang, C.; Huang, W.; Ying, Y.; Li, S.; Secco, D.; Tyerman, S.; Whelan, J.; Shou, H. Functional characterization of the rice SPX-MFS family reveals a key role of OsSPX-MFS1 in controlling phosphate homeostasis in leaves. New Phytol. 2012, 196, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ai, P.; Sun, S.; Zhao, J.; Fan, X.; Xin, W.; Guo, Q.; Yu, L.; Shen, Q.; Wu, P.; Miller, A.J.; Xu, G. Two rice phosphate transporters, OsPht1;2 and OsPht1;6, have different functions and kinetic properties in uptake and translocation. Plant J. 2009, 57, 798–809. [Google Scholar] [CrossRef]
- Liu, F.; Wang, Z.; Ren, H.; Shen, C.; Li, Y.; Ling, H.Q.; Wu, C.; Lian, X.; Wu, P. OsSPX1 suppresses the function of OsPHR2 in the regulation of expression of OsPT2 and phosphate homeostasis in shoots of rice. Plant J. 2010, 62, 508–517. [Google Scholar] [CrossRef]
Figure 1.
Transcriptional expression of OsMYB58 in rice under nutrient deficiency conditions. The upper panels of each figure indicated that transcript levels of OsMYB58 were analyzed using the northern blot. The bottom graphs of each figure indicated that the relative values of band intensity were calculated by rRNA intensity. (a) Total RNA extracted from rice wild-type plants (Oryza sativa L. ‘Dongjin’) growing in various nutrient deficiency conditions. Rice were transferred to nitrogen (N-; 0.25 mM), phosphate (P-; 0.0125 mM), potassium (K-; 0.01 mM), or iron (Fe-; 0.01 mM)-deficient media and grown for 6 hours. (b) Total RNA extracted from shoots and roots of rice plants after 3 days of treatment to high Pi (P+; 0.25 mM KH2PO4) or low Pi (P-; 0.0125 mM KH2PO4). (c and d) Rice samples were treated to low Pi at different time points. Total RNA extracted from separate parts of shoot (c) and root (d) parts of harvested samples. The rRNA was a loading control.
Figure 1.
Transcriptional expression of OsMYB58 in rice under nutrient deficiency conditions. The upper panels of each figure indicated that transcript levels of OsMYB58 were analyzed using the northern blot. The bottom graphs of each figure indicated that the relative values of band intensity were calculated by rRNA intensity. (a) Total RNA extracted from rice wild-type plants (Oryza sativa L. ‘Dongjin’) growing in various nutrient deficiency conditions. Rice were transferred to nitrogen (N-; 0.25 mM), phosphate (P-; 0.0125 mM), potassium (K-; 0.01 mM), or iron (Fe-; 0.01 mM)-deficient media and grown for 6 hours. (b) Total RNA extracted from shoots and roots of rice plants after 3 days of treatment to high Pi (P+; 0.25 mM KH2PO4) or low Pi (P-; 0.0125 mM KH2PO4). (c and d) Rice samples were treated to low Pi at different time points. Total RNA extracted from separate parts of shoot (c) and root (d) parts of harvested samples. The rRNA was a loading control.
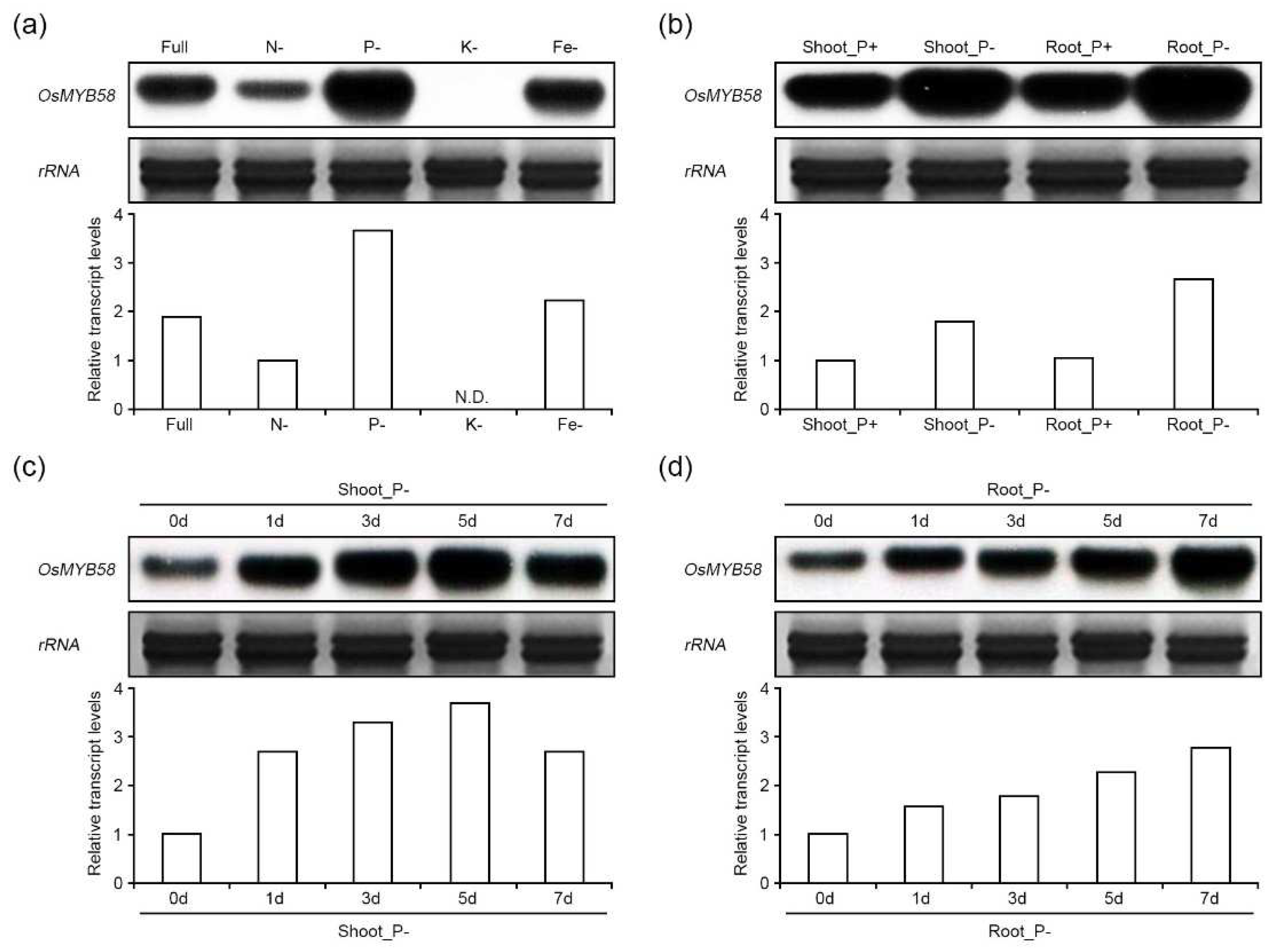
Figure 2.
Physiological analysis of OsMYB58 overexpressing Arabidopsis plants under low Pi conditions. (a) 4-day-old seedlings of Arabidopsis wild-type (Col-0) and OsMYB58 overexpressing plants (OsMYB58-AraOX) were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days and then the photos were taken. Scale bar indicated the 1.8 cm. (b) Comparison of root architectures between Col-0 and OsMYB58-AraOX seedlings depicted in (a). Scale bar indicated the 1.8 cm. (c-f) After 7 days to high Pi or low Pi, physiological changes in shoot and root were analyzed by various methodological measurements, including shoot fresh weight (c), root fresh weight (d), primary root length (e), and the number of lateral roots (f). (g and h) Pi concentrations were measured in the shoot (g) and root (h) of Col-0 and OsMYB58-AraOX after treatment to high Pi or low Pi for 7 days. Error bars represent the mean ± standard deviation (SD) of three biological replicates with 10 seedlings for each experiment. Asterisks represent significant differences from the Col-0 (*; 0.01 < p-value ≤ 0.05, **; p-value ≤ 0.01, Student’s t-test).
Figure 2.
Physiological analysis of OsMYB58 overexpressing Arabidopsis plants under low Pi conditions. (a) 4-day-old seedlings of Arabidopsis wild-type (Col-0) and OsMYB58 overexpressing plants (OsMYB58-AraOX) were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days and then the photos were taken. Scale bar indicated the 1.8 cm. (b) Comparison of root architectures between Col-0 and OsMYB58-AraOX seedlings depicted in (a). Scale bar indicated the 1.8 cm. (c-f) After 7 days to high Pi or low Pi, physiological changes in shoot and root were analyzed by various methodological measurements, including shoot fresh weight (c), root fresh weight (d), primary root length (e), and the number of lateral roots (f). (g and h) Pi concentrations were measured in the shoot (g) and root (h) of Col-0 and OsMYB58-AraOX after treatment to high Pi or low Pi for 7 days. Error bars represent the mean ± standard deviation (SD) of three biological replicates with 10 seedlings for each experiment. Asterisks represent significant differences from the Col-0 (*; 0.01 < p-value ≤ 0.05, **; p-value ≤ 0.01, Student’s t-test).
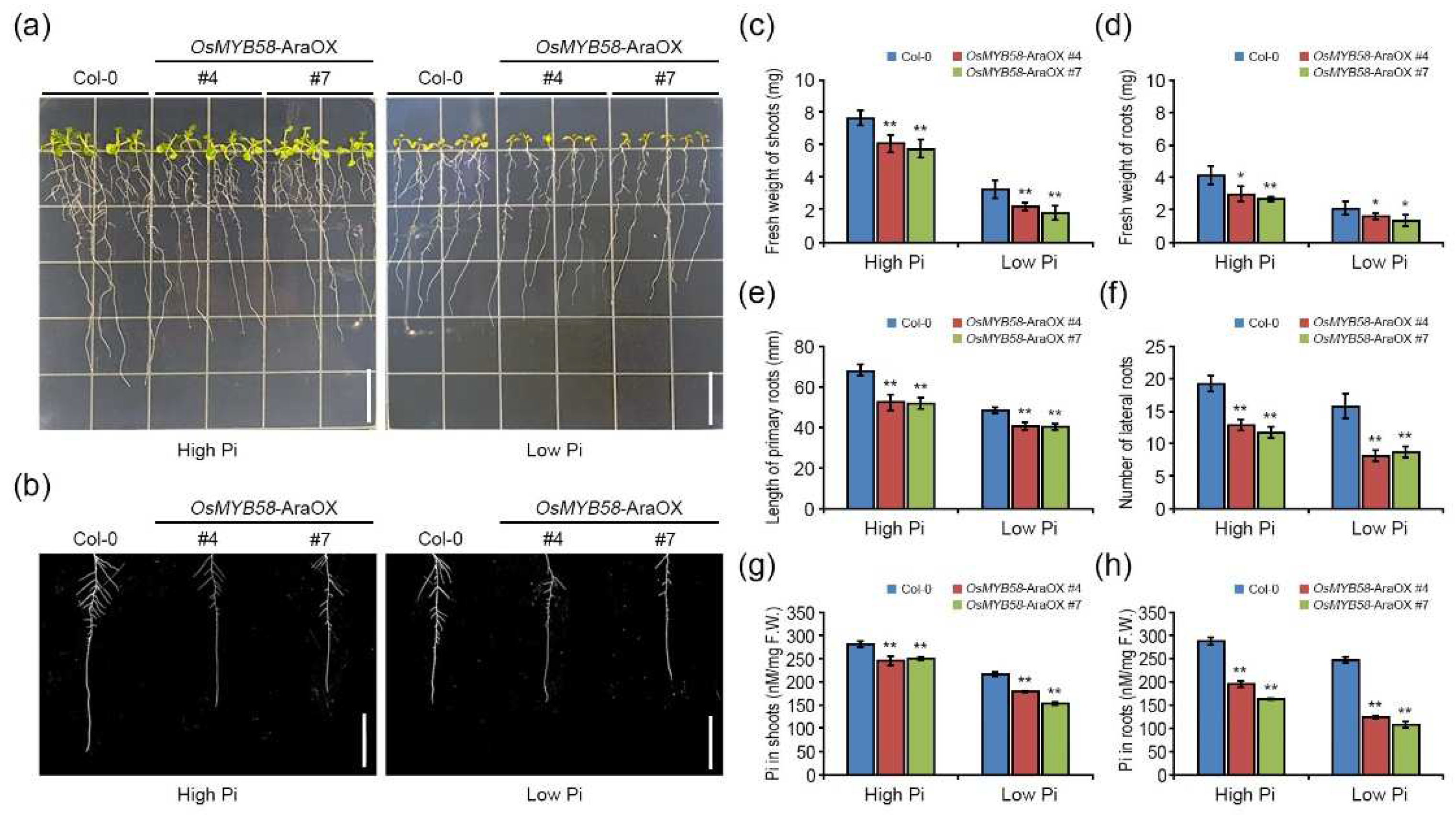
Figure 3.
Physiological analysis of OsMYB58 overexpressing and knock-out mutant rice plants. (a) 7-day-old seedlings of rice wild-type (WT), OsMYB58 overexpressing plants (OsMYB58-OX), and OsMYB58 T-DNA tagging knock-out mutant (OsMYB58-KO) were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days and then the photos were taken. Scale bar indicated the 5 cm. (b-e) After 7 days to high Pi or low Pi, physiological changes in shoot and root were analyzed by various methodological measurements, including fresh weight of shoots (b), fresh weight of roots (c), length of shoots (d), and length of primary roots (e). Error bars represent the mean ± standard deviation (SD) of three biological replicates with five seedlings for each experiment. Asterisks represent significant differences from the WT (*; 0.01 < p-value ≤ 0.05, **; p-value ≤ 0.01, Student’s t-test).
Figure 3.
Physiological analysis of OsMYB58 overexpressing and knock-out mutant rice plants. (a) 7-day-old seedlings of rice wild-type (WT), OsMYB58 overexpressing plants (OsMYB58-OX), and OsMYB58 T-DNA tagging knock-out mutant (OsMYB58-KO) were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days and then the photos were taken. Scale bar indicated the 5 cm. (b-e) After 7 days to high Pi or low Pi, physiological changes in shoot and root were analyzed by various methodological measurements, including fresh weight of shoots (b), fresh weight of roots (c), length of shoots (d), and length of primary roots (e). Error bars represent the mean ± standard deviation (SD) of three biological replicates with five seedlings for each experiment. Asterisks represent significant differences from the WT (*; 0.01 < p-value ≤ 0.05, **; p-value ≤ 0.01, Student’s t-test).
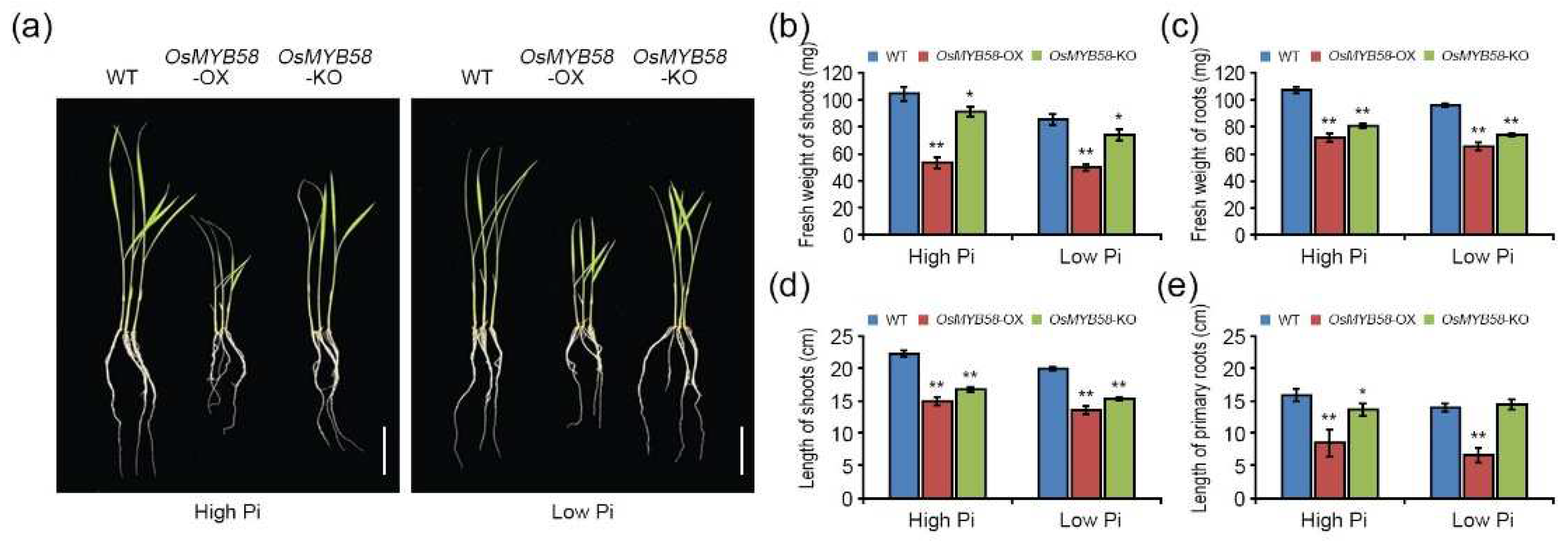
Figure 4.
Physiological alteration in root architecture of OsMYB58-OX and OsMYB58-KO plants. (a) 3-day-old seedlings of rice WT, OsMYB58-OX, and OsMYB58-KO were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days and then the photos were taken in root architecture. The white arrow indicated the primary roots and the yellow arrowhead indicated the seminal roots. Scale bar in upper or middle panels indicated the 1 cm. Scale bar in bottom panels indicated the 0.5 mm. (b-g) After 7 days to high Pi or low Pi, physiological alteration in root architecture was analyzed by various methodological measurements, including length of seminal roots (b), number of seminal roots (c), length of lateral roots (d), number of lateral roots (e), length of root hair (f), and number of lateral roots per 1cm primary roots (g). Error bars represent the mean ± standard deviation (SD) of three biological replicates with five seedlings for each experiment. Asterisks represent significant differences from the WT (*; 0.01 < p-value ≤ 0.05, **; p-value ≤ 0.01, Student’s t-test).
Figure 4.
Physiological alteration in root architecture of OsMYB58-OX and OsMYB58-KO plants. (a) 3-day-old seedlings of rice WT, OsMYB58-OX, and OsMYB58-KO were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days and then the photos were taken in root architecture. The white arrow indicated the primary roots and the yellow arrowhead indicated the seminal roots. Scale bar in upper or middle panels indicated the 1 cm. Scale bar in bottom panels indicated the 0.5 mm. (b-g) After 7 days to high Pi or low Pi, physiological alteration in root architecture was analyzed by various methodological measurements, including length of seminal roots (b), number of seminal roots (c), length of lateral roots (d), number of lateral roots (e), length of root hair (f), and number of lateral roots per 1cm primary roots (g). Error bars represent the mean ± standard deviation (SD) of three biological replicates with five seedlings for each experiment. Asterisks represent significant differences from the WT (*; 0.01 < p-value ≤ 0.05, **; p-value ≤ 0.01, Student’s t-test).
Figure 5.
Pi accumulation in OsMYB58-OX and OsMYB58-KO plants. Pi concentrations were measured in the shoots (a) and roots (b) of rice WT, OsMYB58-OX, and OsMYB58-KO plants under high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) conditions. Error bars represent the mean ± standard deviation (SD) of three biological replicates with five seedlings for each experiment. Asterisks represent significant differences from the WT (**; p-value ≤ 0.01, Student’s t-test).
Figure 5.
Pi accumulation in OsMYB58-OX and OsMYB58-KO plants. Pi concentrations were measured in the shoots (a) and roots (b) of rice WT, OsMYB58-OX, and OsMYB58-KO plants under high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) conditions. Error bars represent the mean ± standard deviation (SD) of three biological replicates with five seedlings for each experiment. Asterisks represent significant differences from the WT (**; p-value ≤ 0.01, Student’s t-test).
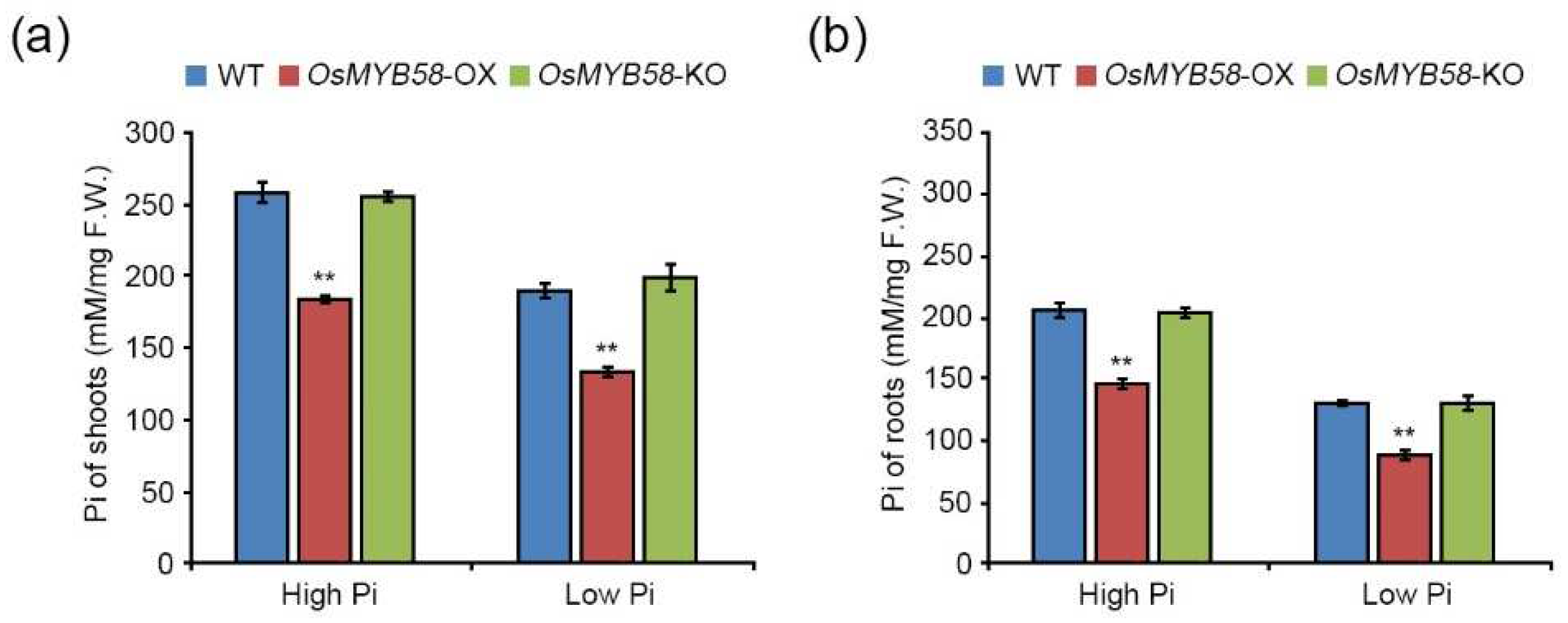
Figure 6.
Transcripts comparison of Pi-responsive genes and Pi transporters in OsMYB58-OX and OsMYB58-KO plants. 7-day-old seedlings of rice WT, OsMYB58-OX, and OsMYB58-KO were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days. For qRT-PCR analysis, total RNA was extracted from shoots and roots of high Pi- or low Pi-treated plants. The qRT-PCR analyzed the transcript levels of Pi-responsive genes, such as OsmiR399a (a), OsmiR399j (b), OsIPS1 (c), OsPHO2 (d), OsPT2 (e), and OsPT4 (f) using specific primers in Supplementary Table S1. Expression of OsACTIN1 was used for the normalization. Error bars represent the mean ± standard deviation (SD) of three biological replicates. Asterisks represent significant differences from the WT (*; 0.01 < p-value ≤ 0.05,**; p-value ≤ 0.01, Student’s t-test).
Figure 6.
Transcripts comparison of Pi-responsive genes and Pi transporters in OsMYB58-OX and OsMYB58-KO plants. 7-day-old seedlings of rice WT, OsMYB58-OX, and OsMYB58-KO were transferred to medium including high Pi (0.25 mM KH2PO4) or low Pi (0.0125 mM KH2PO4) for 7 days. For qRT-PCR analysis, total RNA was extracted from shoots and roots of high Pi- or low Pi-treated plants. The qRT-PCR analyzed the transcript levels of Pi-responsive genes, such as OsmiR399a (a), OsmiR399j (b), OsIPS1 (c), OsPHO2 (d), OsPT2 (e), and OsPT4 (f) using specific primers in Supplementary Table S1. Expression of OsACTIN1 was used for the normalization. Error bars represent the mean ± standard deviation (SD) of three biological replicates. Asterisks represent significant differences from the WT (*; 0.01 < p-value ≤ 0.05,**; p-value ≤ 0.01, Student’s t-test).
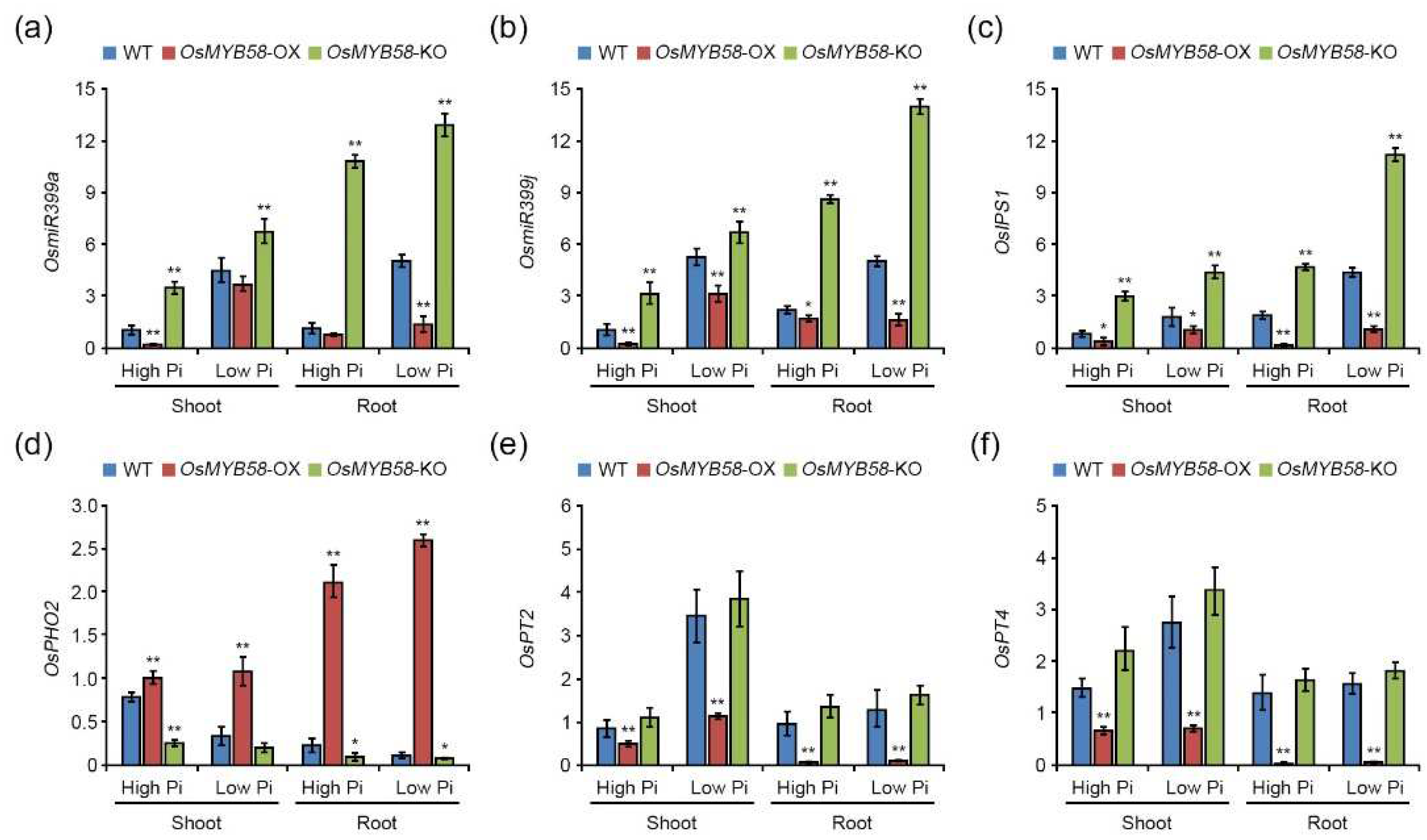
Figure 7.
Transcriptional activity of OsMYB58 in Arabidopsis protoplast transient system. A schematic diagram showed the effector and reporter plasmid DNA used in the transient expression assay. Combinations with each effector along with two reporters were co-transfected into protoplasts from 2-week-old Arabidopsis leaves. ARF5/MP was used as an experimental positive control, and 35S:LUC plasmid DNA was used as an internal control. After normalization by LUC activity, GUS activity in each transfected protoplast sample was calculated. Error bars represent the mean ± standard deviation (SD) of three biological replicates. Asterisks represent significant differences from the BD-vector (**; p-value ≤ 0.01, Student’s t-test).
Figure 7.
Transcriptional activity of OsMYB58 in Arabidopsis protoplast transient system. A schematic diagram showed the effector and reporter plasmid DNA used in the transient expression assay. Combinations with each effector along with two reporters were co-transfected into protoplasts from 2-week-old Arabidopsis leaves. ARF5/MP was used as an experimental positive control, and 35S:LUC plasmid DNA was used as an internal control. After normalization by LUC activity, GUS activity in each transfected protoplast sample was calculated. Error bars represent the mean ± standard deviation (SD) of three biological replicates. Asterisks represent significant differences from the BD-vector (**; p-value ≤ 0.01, Student’s t-test).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



