
Submitted:
21 November 2023
Posted:
22 November 2023
You are already at the latest version
Abstract
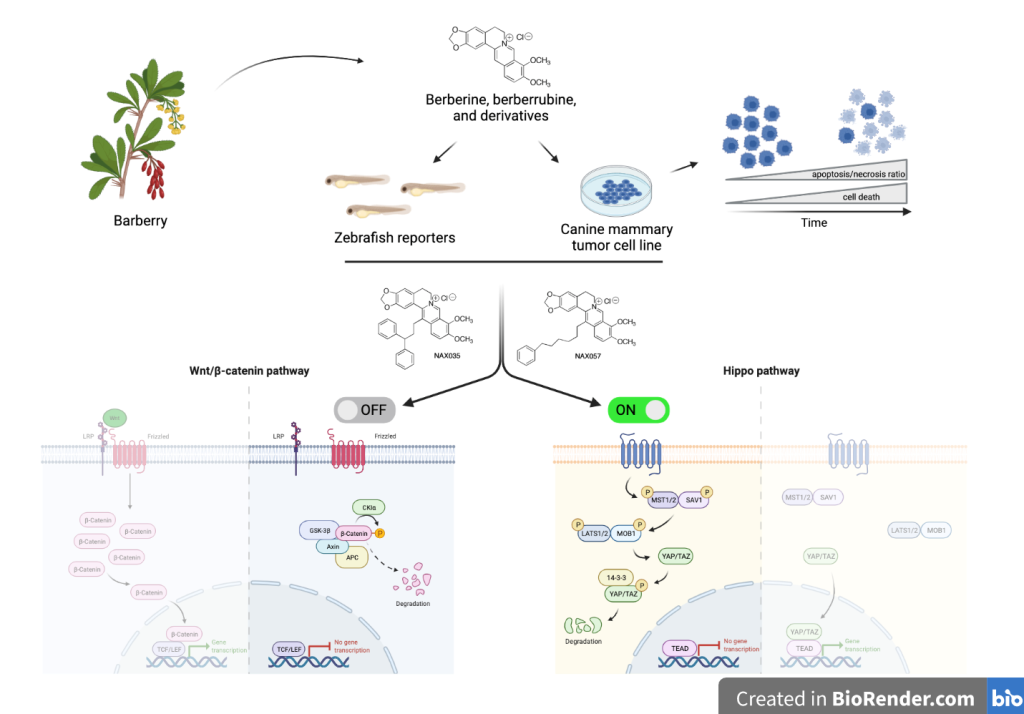
The heterogeneous nature of human breast cancer (HBC) can still lead to therapy inefficacy and high le-thality and new therapeutics are needed as well as new spontaneous animal models to benefit translational HBC research. Dogs are primarily investigated since they spontaneously develop tumors which share many features with human cancers. In recent years, different natural phytochemicals including berberine, a plant alkaloid, have been reported to have antiproliferative activity in vitro in human cancers and in rodent animal models. In this study, we report the antiproliferative activity and mechanism of action of berberine, of its active metabolite berberrubine, and of eight analogues, on a canine mammary carcinoma cell line and in transgenic zebrafish models. We demonstrate both in vitro and in vivo the significant effects of specific analogues on cell viability, via induction of apoptosis, also identifying their role in inhibiting the Wnt/β-catenin pathway and activating the Hippo signals with a downstream reduction of CTGF expres-sion. Particularly the berberine analogues NAX035 and NAX057 show the highest therapeutic efficacy, de-serving further analyses to elucidate their mechanism of action more in detail, and in vivo studies on spon-taneous neoplastic diseases aiming at improving veterinary treatments of cancer as well as translational cancer research.
Keywords:
berberine derivates
; dog
; mammary tumors
; zebrafish
; Hippo
; Wnt
; β-catenin
1. Introduction
Human breast cancer (HBC) is the most common type of tumor in women accounting for 30% of all diagnosed cancers [1]. Despite recent advancements in detection, diagnosis and treatment of HBC, the heterogeneous nature of this disease leads, in some cases, to therapy inefficacy [2]. For this reason, spontaneous animal models of HBC are highly important to study the molecular mechanisms underlying the onset and progression of these tumors. They can serve as a useful intermediary between traditional preclinical models and human clinical trials and therefore for the identification of new therapies [3,4].
Canine mammary tumors (CMTs) are a highly heterogeneous group of tumors representing the most common type of tumors in female dogs. Approximately 50% of diagnosed CMTs are malignant [5] but recently, an increase in malignant vs. benign tumors was observed with a similar trend that has been also described in human oncologic patients [6]. CMTs are spontaneous in dogs as well as in women, and many anatomic, clinical, and histological features of CMTs are similar to those reported in HBC [7,8,9]. As companion animals, dogs share environment and lifestyle with humans and become consequently exposed to many of the same carcinogens. Better than genetically modified mice, dogs have higher physiological similarities with humans, such as comparable telomerase activities [10] and a high incidence of spontaneous cancers [11], making them a valuable cancer model for HBC research [12]. This spontaneous animal model could be highly beneficial to translational HBC research, particularly for the identification of new therapeutic targets and the collection of highly predictive data that may accelerate human cancer clinical research.
In the last decade, the Wnt/β-catenin pathway, involving β-catenin and the adenomatous polyposis coli (APC) proteins, has emerged as an important player in many human tumor types, including HBC [13,14,15]. The reduced membrane expression of E-cadherin and β-catenin has been associated with a poor prognosis in feline and canine MTs as well as in HBC [16,17]. E-cadherin in association with other adhesion molecules of the β-catenin class binds to the actin cytoskeleton. This association is important for the epithelial cell function and for tissue integrity [18]. Moreover, β-catenin has a double function as an adhesion molecule and as an activator of the Wnt/β-catenin pathway. Specifically, in normal cells, free cytoplasmic β-catenin is entrapped by a complex containing axin, glycogen synthase kinase 3 (GSK3), and APC that facilitates the phosphorylation of β-catenin which consequently can be degraded. In tumor cells, this complex is not able to phosphorylate β-catenin that, as a result, cannot be degraded [19,20]. To the best of our knowledge, only little information is known about the role played by the Wnt/β-catenin signaling pathway in CMTs [21,22,23].
Additionally, the Hippo pathway has been identified as highly important in HBC onset and progression. Cordenonsi and co-authors demonstrated that the activity of the transducer of the Hippo pathway TAZ is required to sustain tumor-initiation capacities and self-renewal of cancer stem cells (CSCs) in the breast. TAZ protein level is elevated in CSCs and in poorly differentiated HBC with a poor prognosis [24,25]. To date, the role of Hippo pathway in CMTs has not been deeply investigated and not fully clarified [23,26,27,28].
Berberine (BBR) is a bitter-tasting isoquinoline quaternary alkaloid isolated from many kinds of plants such as Hydrastis canadensis, Berberis vulgaris and other plant species. A mainstay of Traditional Chinese Medicine, BBR is also in use in Ayurvedic and Native American herbal medicines for its anti-microbial and anti-inflammatory properties [29,30]. BBR has multiple pharmacological properties including antibacterial, anti-inflammatory, antidiabetic, hepatoprotective, and neuroprotective. It also controls the expansion of blood vessels and the inhibition of platelet aggregation and, therefore, has a wide spectrum of medical applications against, inter alia, gastroenteritis, hyperlipidemia, non-alcoholic fatty liver disease, coronary artery disease, hypertension, diabetes, and Alzheimer’s disease [31,32,33,34]. There is an increasing interest in the clinical efficacy of BBR as manifested by almost 90 completed and ongoing clinical studies, including five for cancer indications.
In vitro studies using HBC cell lines demonstrated that BBR inhibits cancer cell migration and proliferation and affects cell viability, inducing apoptosis [15,35,36,37]. More recently, it has been shown that BBR could be a promising drug able to suppress cell growth and cell invasiveness of triple-negative HBC (TNBC) cells through IL-8-related mechanisms [38,39]. BBR was also demonstrated to be a promising drug in prevention and treatment of colon cancer through the inhibition of Wnt/β-catenin signaling [40] and in exerting anticancer activity delaying the development of MTs in transgenic mice for the HER-2/neu oncogene [41,42,43].
Berberrubine (BRR) is the first-pass metabolism product of BBR [44], and little is known about its antitumor properties [45,46]. BRR has been reported to induce topoisomerase II-mediated DNA cleavage [47], and to bind G-quadruplex nucleic acid structures [48].
The aim of our study was to evaluate the potential of BBR, BRR, and eight structurally related analogues as anticancer agents. We conducted in vitro experiments to examine the biological effects of these compounds on a malignant CMT cell line in order to identify candidates that might become potential anticancer agents. The analogues under investigation possess aromatic groups attached to the 13-position of the parent alkaloid skeleton through a hydrocarbon linker [49]. Their effects on Wnt/β-catenin signaling [50] and on some relevant human tumor types have been reported, both in vitro [36,51,52,53,54,55] and in vivo [41,56].
To elucidate the mechanism involved in the antitumor effect of BBR, of BRR, and of selected analogues, we investigated the association with the Wnt/β-catenin signaling and Hippo pathways. More specifically, we used Wnt/β-catenin and Hippo signaling pathway reporter zebrafish lines to confirm the in vivo effect of these BBR and BRR analogues on these pathways. We expected to identify at least one analogue able to kill cancer cells mainly by apoptosis and better clarify how the Wnt/β-catenin and Hippo signaling pathways are involved.
2. Materials and Methods
2.1. Chemicals
Berberine (BBR - chloride form, Trust & We, Shanghai, China), berberrubine (BRR), and analogues NAX012, NAX014, NAX035, NAX053, NAX057, NAX060, NAX085, NAX118 were provided by Naxospharma (Figure 1).
2.2. Cell Culture and Drug Treatment
The canine mammary carcinoma cell line CF33 (kindly provided by Dr. R. De Maria, University of Turin, Italy) was maintained in Dulbecco Modified Eagle Medium (DMEM) supplemented with 10% Fetal Bovine Serum (FBS), 100 units/ml penicillin, and 100μg/ml streptomycin under a humidified atmosphere of 5% CO2 at 37°C. BBR, BRR, and analogues were dissolved in dimethyl sulfoxide (DMSO), aliquoted, and stored at −20 °C with a stock concentration of 40mM. The final concentration of DMSO did not exceed 0.1%.
For the in vitro experiments, CF33 cells were plated and treated separately with BBR, BRR, NAX012, NAX014, NAX035, NAX053, NAX057, NAX060, NAX085 and NAX118. Two different experiments were performed. In the first experiment, we used different drug concentrations, from 2 μM to 40 μM, for a steady incubation time of 24 hours. In the second experiment, we used the same drug concentration, 10 μM, for different incubation times (from 3 to 24 hours). Control cells were incubated with a respective maximal percentage of DMSO alone (<0.1%). The two experiments were performed in triplicate.
2.3. Cell Viability Assay
CF33 cells were plated in a 96-well plate at a density of 1x104 cells/well in 100 μl of complete medium. After 24 hours, a medium containing the specific drug was added. To measure cell viability, cells were incubated with the different drugs and the in vitro toxicology assay kit TOX8 (Sigma-Aldrich) was used, following manufacturer’s instructions. The experiment was performed in triplicate. Experimental results were expressed as percentage of cell viability in comparison with control DMSO-treated cells.
2.4. Quantification of Necrosis/Apoptosis by Flow Cytometry
In order to assess the rate of necrosis and apoptosis induced by BBR, BRR, and analogues at the same concentration (10 µM) and at the different incubation times (6, 12, 18, and 24 hours of incubation), two biological replicates (both performed in technical triplicates) of CF33 were analyzed by flow cytometry using the Annexin V-Fluorescein isothiocyanate (FITC) Apoptosis Detection Kit (eBioscience, Thermo Scientific) following the manufacturer’s instruction. Briefly, cells were trypsinized and centrifuged at 1100 rpm for 10 min at 4°C. Cells were resuspended in 200 µL of binding buffer with 5 µL of annexin V–FITC and incubated for 10 min in the dark at room temperature. After the incubation, 200 µL of binding buffer were added to the cells that were subsequently centrifuged at 1100 rpm for 10 min at 4°C, and the supernatant was discarded. Then, 10 µL of Propidium Iodide (PI) and 900 µL of binding buffer were added. After 5 minutes of incubation in the dark at room temperature, the cells were acquired using the flow cytometer CyFlow Space (Partec-System, Sysmex Europe GmbH, Norderstedt-Amburgo, Germany) and the data were analyzed with the open-source software FCSalyzer (version 0.9.16-alpha). For each tube, 20,000 events were analyzed. Necrotic cells were positive for both Annexin-V FITC and PI while apoptotic cells were positive for Annexin-V FITC only.
2.5. Protein Extraction and Western Blot Analysis on CF33 Cells
At the end of incubation time (24 hours), CF33 cells treated with BBR, BRR or analogues at a final concentration of 2 μM, were washed twice with ice-cold phosphate buffered saline (PBS) and total lysate was obtained by solubilization in 5% sodium deoxycholate supplemented with protease inhibitors (Sigma-Aldrich). Protein concentration was determined by the bicinchoninic acid- protein assay (Pierce), using bovine serum albumin as standard. Protein fractions were stored at −80 °C. Proteins were resolved by SDS-PAGE using 7.5% polyacrylamide gels [57] and the same amount of protein (20g) was loaded for all the samples. After electrophoresis, proteins were transferred onto nitrocellulose which was stained with Ponceau S (Sigma-Aldrich). The proteins were transferred to the membrane and blocked in 5% skim milk before being probed overnight at 4°C with the primary antibodies. The blots were probed with the following antibodies: rabbit polyclonal antibodies specific for WW domain-containing transcription regulator protein 1 (WWTR1) (1:1000, Sigma-Aldrich #HPA007415), recognizing both YAP and TAZ human proteins; rabbit monoclonal antibodies to non-phosphorylated (active) β-Catenin (1:1000, Cell Signaling #8814); mouse monoclonal antibodies to β-catenin (1:1000, BD Biosciences #610154). After washing the membrane, an HRP-conjugate secondary antibody was added and incubated with the membrane for 1 hour at room temperature. The membrane was washed and analyzed by a chemiluminescence analyzer using the chemiluminescence ECL reagent.
2.6. RT-PCR and Semi-Quantitative PCR
Total RNA was isolated from control CF33 cells and CF33 cells 24 hours after treatment with 2 μM of drug using RNeasy Micro Kit (Qiagen) following the supplier’s protocol. The obtained RNA was then quantified using Qubit Fluorometric Quantitation (LifeTechnologies) and 500 ng were reverse transcribed using RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific), according to manufacturer’s instructions. Five μl of cDNA products were amplified with 1 unit of GoTaq (Promega) in the buffer provided by the manufacturer that contains MgCl2, with dNTPs and in the presence of the specific primers for β-catenin, YAP, TAZ, CTGF. Actin-beta was used as a housekeeping gene. Primers and PCR Conditions are summarized in Table 1. A first cycle of 2 minutes at 95°C was followed by 30 seconds at 95°C, 30 seconds at 58°C and 30 seconds at 72 °C for 27 cycles. The used number of cycles was chosen so that none of the samples reached a plateau at the end of the amplification protocol; therefore, all the samples were in the exponential phase of amplification. Each set of reactions always included a negative control, where the sample was replaced by nuclease-free water and was performed in triplicate.
2.7. Gel Electrophoresis, Acquisition of Gel Images and Quantitative Analysis
The PCR products were loaded onto a 2% agarose gel stained with gel-red (Promega, Italy). A 100 bp DNA ladder molecular weight marker (Life Technologies, Rockville, MD) was run on each gel to confirm the expected molecular weight of the amplification product. Images of the RT-PCR agarose gels were acquired with BioRad gel imaging systems and band quantification was performed using ImageJ software. The ratio between the gene of interest and actin-beta was calculated to normalize for initial variations in sample concentration and as a control for reaction efficiency. Mean and standard deviation of all experiments performed were calculated after normalization to actin-beta.
2.8. Ethics Statements
Zebrafish embryos and adults were raised, staged, and maintained at the Zebrafish Facility of the University of Padua, under standard conditions [58]. All husbandry and experimental procedures complied with European Legislation for the Protection of Animals used for Scientific Purposes (Directive 2010/63/EU) and with Italian law on animal experimentation (D.L. 4 March 2014, n.26). All the procedures were carried out under authorization n. 407/2015-PR from the Italian Ministry of Health. The project was also examined and approved by the Ethics Committee of the University of Padua with protocol number 18746.
2.9. Zebrafish Housing and Maintenance
Zebrafish used for all the experiments were taken from the breeding stocks of the Zebrafish Facility of the University of Padua and fed four times a day on a variable diet of dried food and live Artemia. Fish were kept in a 14:10 light:dark cycle in a 10L multi tank constant flow system. Water temperature was held at a constant 28.5 ̊C and replaced at a rate of 10% per day. Eggs were collected between 9:00–11:00 am and transferred in petri dishes. The following day, dead eggs were removed, and dishes were cleaned. Zebrafish embryos were incubated at 28.5 ̊C in system water mixed with methylene blue (2 ml 0.1% methylene blue per 1 L). Humane endpoints were not used considering that all the experimental procedures were performed in wild type and transgenic larvae under 5 days post fertilization. For anesthesia or euthanasia of zebrafish embryos and larvae, Tricaine was added to the fish water at 0.16 or 0.3 mg/mL, respectively. The study was carried out in compliance with the ARRIVE guidelines as reported by Percie du Sert and colleagues [59]. All the experimental procedures have been performed by specifically trained personnel by the veterinary staff working in the Zebrafish Facility of the University of Padua.
2.10. Zebrafish Tg(7xTCF-Xla.-Siam:mCherry) and Tg(Hsa.CTGF:mCherry) Transgenic Lines
In order to confirm the in vivo effect of BBR and NAX035, NAX053 and NAX057 on Wnt/β-catenin and Hippo pathways, we used two zebrafish transgenic lines named Tg(7xTCF-Xla.-Siam:mCherry) [60] and Tg(Hsa.CTGF:mCherry), respectively [61]. Fish of all strains, maintained in Padua Zebrafish facility, were monitored daily for the presence of signs of sickness, pain, distress, suffering, or moribund conditions; all treated zebrafish larvae were euthanized before the phenotypic analysis.
2.11. LC50
The median lethal concentration (LC50) of BBR and analogues identified as the drug dosage able to kill 50% of 24 hours post fertilization (hpf) zebrafish larvae incubated with the drugs for 24 hours, was calculated with the Quest Graph™ LD50 Calculator, (AAT Bioquest, Inc., Sunnyvale, CA, United States). Different dosages of drugs were directly added to the fish water in 96-well plates. For each treatment we used at least 96 embryos, and the experiment was performed in triplicate.
2.12. In Vivo Drug Treatments
Zebrafish embryos were incubated with 100 μM BBR (BBR LC50) and with 10 μM of NAX035, NAX053 and NAX057 (LC50) at 24 hpf for 24 or 48 hours. Drugs were added directly to the fish water in 6-well plates. For each treatment performed in triplicates, at least 10 embryos were used.
2.13. Microscopy and Image Acquisition
The mCherry-expressing embryos belonging to the transgenic lines named Tg(7xTCF-Xla.-Siam:mCherry) and Tg(Hsa.CTGF:mCherry) were analyzed using a Leica M165FC epifluorescent microscope. All pictures were acquired with a Leica DC 500 digital camera and contrast and brightness elaborated with Adobe Photoshop 6.0 software. In each transgenic embryo, to quantify the level of fluorescence (that is strictly associated with the activity of Wnt/β-catenin of Hippo pathways) the integrity density was calculated using ImageJ software. With ImageJ we were able to analyze the embryos’ fluorescence in the head, cardiac region, and spinal cord. The yolk sac was not included since it usually shows autofluorescence after drug treatment.
2.14. Statistical Analysis
Statistical analysis was performed using GraphPad Prism 9 software. To verify mean differences among groups, the one-way ANOVA with Tukey’s multiple comparison test was used when data were normally distributed. The Kruskal-Wallis test with Dunn’s multiple comparison test was used when data were not normally distributed. Shapiro-Wilk test was used to check normality. Level of significance was set at p < 0.05.
3. Results
3.1. BBR, BRR, and Analogues Induce a Dose-Dependent Inhibition of Tumor Cell Viability
To evaluate the in vitro anti-proliferative effect of BBR, BRR, and analogues (Figure 1) on CF33 cells, we measured cell viability after treatment. In the first experiment, cells were treated with BBR and analogues for 24 hours using different dosages (2 μM, 5 μM, 10 μM, 20 μM, and 40μM) (Figure 2A). The half maximal inhibitory concentration (IC50) was analyzed for each compound and reported in Table S1. Consequently, we performed a second cell viability assay at different time points, using a concentration of 10 μM for all the compounds (Figure 2B). In this experiment, cells were treated with each drug at the dosage of 10 μM and cell viability was measured at different times (3, 6, 12, 15, 18, and 24 hours) (Figure 2B). Treatments with BBR, BRR, and analogues exerted a dose- and time-dependent inhibition of cell viability (Figure 2).
NAX012, NAX035, NAX053, NAX057, and NAX060 were identified as promising drug candidates able to kill more than 50% of tumor cells after 3h. NAX035, NAX053, NAX057, and NAX060 were able to kill more than 85% of tumor cells after 24h (Figure 2B) and were selected for further analyses. BRR was also included as a comparison being an apparently less efficient compound.
3.2. Induction of Necrosis/Apoptosis in Canine Mammary Tumor Cells by BBR Analogues
Next, we investigated whether the inhibitory effects of BBR and analogues were due to apoptotic or necrotic cell death. CF33 cells were exposed to 10 μM of each drug and early apoptosis and late apoptosis/necrosis were measured after 6, 12, 18, and 24 hours of treatment by flow cytometry (Figure 3). Over time, an increased rate of apoptosis versus necrosis was observed for most drugs, from 6 to 18 hours (Figure 3A–C).
3.3. NAX035, NAX053, NAX057, and NAX060 Induce a Downregulation of Wnt/β-Catenin and an Activation of the Hippo Signaling Pathways in CF33 Cells
To evaluate the effect of BBR and the most efficacious analogues on Wnt/β-catenin and Hippo pathways, we analyzed the protein expression of total β-catenin, active β-catenin, YAP, and TAZ after treatment.
Western blot analysis revealed a decreased amount of total β-catenin in CF33 cells treated with NAX035, NAX053, NAX057, and NAX060. Interestingly, cells treated with the same compounds showed a notable lower expression of active β-catenin when compared to the controls and to other analogues, suggesting a down-regulation of the Wnt/β-catenin pathway (Figure 4).
Considering the Hippo pathway, western blot analysis also revealed a down-regulation of YAP and TAZ proteins in cells treated with NAX035, NAX053, NAX057, and NAX060 when compared to the control, which was more evident for TAZ (Figure 4). These results suggest a down-regulation of the Wnt/β-catenin and an activation of the Hippo pathway, respectively, when cells are treated with NAX035, NAX053, NAX057, and NAX060, when compared to the control cells.
We focused on genes specifically involved in the Wnt/β-catenin and Hippo pathways. β-catenin mRNA expression was not significantly altered after treatment with NAX035, NAX053, NAX057 and BRR in comparison with cells treated with BBR and control. β-catenin mRNA expression was significantly higher (p < 0.01) in cells treated with NAX060 when compared to the control (Figure 5A).
Interestingly, YAP mRNA expression was significantly higher in cells treated with BBR, NAX053, and NAX060, when compared to the control (p < 0.05) (Figure 5B), whereas TAZ mRNA expression was not altered after treatment (Figure 5C). We also looked at the mRNA expression of CTGF, which is regulated by the Hippo pathway. Notably, CTGF mRNA expression was significantly lower in cells treated with NAX035 (p < 0.01) and NAX057 (p < 0.001) when compared to the control (Figure 5D), indicating an activation of the tumor-suppressor Hippo pathway after treatment with these BBR analogues.
3.4. BBR and NAX057 Reduce the Activity of Wnt/β-Catenin and Activate Hippo Signaling Pathways in Zebrafish Embryos
Next, we used zebrafish embryos to study the toxicity response to BBR, BRR and analogues by assessing Median Lethal Concentration (LC50) at 24 and 48 hours of drug treatment. The LC50 of BBR, BRR, NAX085, and NAX118 was defined at 200 μM. NAX012 and NAX014 had a LC50 of 100 μM. NAX035 and NAX060 had a LC50 of 20 μM. Finally, NAX053 and NAX057 had a LC50 of 10 μM (Table S2). For all tested compounds, zebrafish embryo LC50 values were not dependent on the duration of exposure, such that longer exposures (48h) were not associated with lower LC50 values (data not shown).
We tested BBR, NAX035, NAX053, and NAX057 on Zebrafish Tg(7xTCF.Xlasiamois:nlsmCherry) and Tg(Hsa.CTGF:mCherry) transgenic lines, in which the expression of the reporter gene mCherry is regulated by specific cell signaling pathways-responsive elements (52,53). By fluorescent microscopy imaging, we analyzed the amount of fluorescence indicating the expression of specific target genes in treated fish compared with controls. The analysis of these responsive transgenic lines, in which the expression of the reporter protein mCherry is directly associated with the activity of canonical Wnt/β-catenin and Hippo signaling pathways, is simple and relatively immediate. In these in vivo experiments, we decided to use the drug concentrations of 100 μM for BBR, 10 μM for NAX035, and 5 μM for NAX053 and NAX057 because these concentrations were tested on zebrafish and did not induce alteration in fish morphology and development. Indeed, fluorescence analysis and quantification need to be performed on animals that are correctly formed and developed. Tg(7xTCF.Xlasiamois:nlsmCherry) reporter animals treated with BBR, NAX035, NAX053, and NAX057 showed a significant lower fluorescence when compared to the control (p < 0.0001) (Figure 6A), suggesting that these compounds down-regulated the canonical Wnt/β-catenin pathway in vivo.
Interestingly, Hippo pathway reporter animals showed a lower fluorescence when treated with BBR (p < 0.001), NAX035, and NAX057 (p < 0.05), when compared to the control (Figure 6B), while transgenic embryos treated with NAX053 showed an increased fluorescence compared to untreated siblings (p < 0.0001) (Figure 6B).
4. Discussion
HBC is the most common invasive cancer in women. Treatment of HBC depends on the subtype of breast cancer, the stage of the disease, sensitivity to hormones, patient's age, and overall health [1]. The main treatment options include radiation therapy, chemotherapy, surgery, and hormone therapy. With treatment, a woman who receives a diagnosis of HBC has a percentage chance of surviving for at least 5 years ranging from almost 100 to 22 percent depending on the stage of the disease at diagnosis and the tumor subtype [62]. For this reason, new treatment strategies are needed.
In recent years, phytochemicals, considered as bioactive ingredients present in plant products, became interesting drugs to be studied for their anti-tumor activity. This effect was demonstrated both in vitro and in vivo [63]. BBR is a phytochemical which has been reported to inhibit carcinogenesis in rats and mice [64] and in certain types of human cancers as revealed by both in vitro and in vivo studies [38,41,65,66] and in one clinical administration to human patients [40].
Regarding breast cancer, most in vitro studies on HBC cell lines have demonstrated BBR antitumoral effects with several involved molecules and pathways mainly regarding cell proliferation and cell cycle, apoptosis, autophagy, and metastasis [65,67]. Only one study investigated BBR effects in vitro on a canine mammary tumor cell line (CF41.Mg) showing a decrease of cell viability after 24h of BBR treatment (100 μM) [67]. In vivo mouse models also showed reduction of tumor volume, tumor weight or vessel density after BBR exposure [41,66,68,69,70,71,72,73]. In a recent study, Pierpaoli and collaborators explored the efficacy of the oral administration of a BBR derivative (NAX014) in a mouse model of HER2-overexpressing breast cancer. The authors showed no signs of toxicity after oral administration of a high dose of the compound, suggesting its safety [56].
A major challenge today is to design novel drugs that target tumor cells specifically, with minimal cytotoxic effects on normal cells. Marverti and collaborators showed a prominent decrease of cell viability on two ovarian cancer cell lines, but a minimal effect on normal cells [74], when treated with BBR. This might indicate that BBR specifically targets tumor cells.
In this study, we chose eight semi-synthetic BBR analogues to investigate their effects on the CMT CF33 cell line for the first time, in comparison with BBR. In order to study the in vivo effect of these compounds, we used a transgenic zebrafish model. Despite this preliminary study being performed with simple methodology, this type of comparison of the two models has never been performed before for CMTs. Our data demonstrated that the BBR analogues NAX035, NAX053, NAX057, and NAX060 affected cell viability, exerting a higher cytotoxic effect on cancer cells acting at a lower concentration in comparison with BBR. Generally, most of the tested drugs increased cell death by inducing cell apoptosis, which is a desirable effect for novel anticancer drugs. Apoptosis is a programmed cell death often overcome by tumors, whereas necrosis is often present in tumors and can be associated with increased metastases [75]. In line with our results, the BBR capacity to induce apoptosis is well known [65]. Wang and co-authors demonstrated that BBR inhibits proliferation and induces apoptosis of human cervical cancer (HeLa229) cells in a dose and time-dependent manner, by the up-regulation of p53 and the downregulation of Bcl2 and Ptgs2 mRNA expression levels [76]. Additionally, the pro-apoptotic activity of BBR is well documented in various breast cancer cell lines and many other cell cancer types [35].
In recent years, several data have emerged regarding the involvement of Wnt/β-catenin signaling in tumors. In particular, the increase of Wnt/β-catenin signaling seems to be important for human cancer onset and progression, including tumor initiation, tumor growth, cell death, cell differentiation and metastasis onset [77]. In this study, we showed that BBR and other BBR analogues downregulated Wnt/β-catenin signaling. Our data were confirmed both at the protein level in vitro, by a decreased expression of active β-catenin in the tested canine cell line, and in vivo on zebrafish. In other studies, BBR has been found to deregulate the Wnt/β-catenin signaling, inhibiting proliferation, migration, and invasion of HBC cells in vitro [78], and human colorectal cancer growth both in vitro and in vivo in a mouse model [79,80]. Similarly, it was found by clinical administration that BBR potently attenuated intestinal polyps in familial adenomatous polyposis human patients via inhibition of Wnt signaling [40]. In our study, we showed that at the gene expression level β-catenin mRNA, instead, did not show a significant decrease after drug treatment, exception made for cells treated with NAX060 which curiously presented a significant increase in comparison with control cells. We can speculate that the downregulation of active β-catenin could be due to post-transcriptional or post-translational mechanisms. To date, it is well known that the mechanism whereby nuclear β-catenin drives or inhibits expression of Wnt target genes is more diverse and less characterized [81]. Additional studies will be necessary to understand better the mechanistic underpinnings of the active β-catenin-observed plasticity. A deeper and more comprehensive characterization of the protein networks which regulate β-catenin transcription using zebrafish as a model to dissect molecular mechanisms involved in tumorigenesis could possibly drive us to confirm the identification of attractive new therapeutic targets.
The Hippo pathway is a well-conserved signaling pathway able to regulate organ size and tissue homeostasis [82]. This pathway can be downregulated leading to oncogenesis through a variety of mechanisms [83]. These mechanisms include the induction of hyperproliferation, cellular invasion, metastasis, and might play a role in cancer cell maintenance and chemotherapy resistance [84]. Our in vitro study on zebrafish for Hippo pathway indicated an effect for BBR and for other analogues on this signaling pathway. Particularly, there was a decrease of YAP and, mainly, TAZ proteins in cells treated with NAX035, NAX053, NAX057, and NAX060. Notably, the mRNA expression of the downstream gene CTGF was lower in cells treated with NAX035 and NAX057, when compared to the control. The Hippo pathway has already been found to be implicated in canine mammary carcinogenesis and the described post-transcriptional/post-translational modifications of YAP and TAZ proteins justify the changes at the protein but not at the mRNA level that were found in our and other studies [23,85]. Mechanistically, YAP and TAZ accumulate within the cytoplasm to translocate into the nucleus and activate downstream tumor-promoting genes when the Hippo pathway is switched off, whereas decreased cytoplasmic YAP/TAZ levels, due to their phosphorylation and degradation, is seen when the tumor-suppressing Hippo pathway is activated [86]. More recently, a Hippo-independent regulation of YAP/TAZ has been described, that could possibly explain non-decreased YAP levels in our study [87]. Several anti-cancer mechanisms of BBR-related compounds have been extensively studied [88], but only one study detected cell cycle arrest and apoptosis induced by a novel synthetic cyclizing-berberine on human cancer cell lines by activating YAP phosphorylation [89]. No data are published on this Hippo-related anticancer effect of BBR analogues.
The analysis of the reporter gene expression in zebrafish treated for 48 hours particularly with BBR and NAX057 (NAX035 with no statistical significance) confirmed a downregulation of CTGF. These data suggest an activation of the Hippo pathway, in comparison with controls, as also supported by a significant decrease of CTGF mRNA expression when cells were treated with NAX035 and NAX057. Instead, treatment with NAX053 indicated an in vivo up-regulation of CTGF. On one side, human studies have shown that CTGF can have pro-tumorigenic or anti-tumorigenic effects in different situations [90,91]. On the other hand, CTGF regulation is not controlled exclusively by the Hippo pathway [92]. Additional analysis to better characterize the response to these drugs in the in vivo zebrafish model should be performed, measuring the expression of other genes and proteins.
Further studies will be necessary to clarify the interesting interaction between Wnt/β-catenin and Hippo pathways in response to BBR and other analogues. We have only touched the tip of the iceberg in understanding the intricacies and interconnections of Wnt/β-catenin and Hippo pathways responsible for the onset and progression of cancer proliferation particularly in canine mammary tumors.
5. Conclusions
In conclusion, our results demonstrated that NAX035 and NAX057 could be considered promising anti-tumoral drugs. In short, (i) they kill approximately 50% of tumor cells at low concentration, (ii) they induce cell death mainly by apoptosis, (iii) they downregulate the Wnt/β-catenin pathway and, (iv) interestingly, they also seem to act on the activation of the well-known tumor-suppressor Hippo pathway. Our results, obtained on a canine mammary tumor cell line, are in line with similar results demonstrated only for BBR in in vitro and in vivo human cancer studies, but with less toxicity also proved in our in vivo zebrafish model. Considering many similarities between canine mammary tumor and human breast cancer, these BBR analogues might represent relevant candidates to be tested in further in vivo animal models.
Supplementary Materials
The following supporting information are available Figure S1: Protein expression of YAP, TAZ, β-catenin, and active β-catenin after treatment with berberine, berberrubine, NAX035, NAX053, NAX057, and NAX060 of a canine mammary cancer cell line. Table S1: Half maximal inhibitory concentration and standard deviation of different compounds after 24 hours of treatment of CF33 cells. Table S2: Different drugs toxicity effects and standard deviation on zebrafish larvae treated for 24 hours.
Author Contributions
Conceptualization, G.B., V.Z.; methodology, A.S., G.B., V.Z.; validation, G.B., A.V., R.S., F.B., G.S.; formal analysis, A.S., G.B.; investigation, G.B., A.S., A.V., R.S., F.B., G.S.; resources, M.E.G., L.C., S.F., C.G., P.L.; data curation, A.S.; writing—original draft preparation, G.B., A.S., V.Z.; writing—review and editing, A.S., R.S., A.V., F.B., P.L.; visualization, A.S.; supervision, V.Z.; project administration, V.Z.; funding acquisition, M.E.G., L.C., S.F., V.Z. All authors have read and agreed to the published version of the manuscript.
Funding
G.B. is supported by BIRD200003, University of Padua.
Institutional Review Board Statement
Zebrafish embryos and adults were raised, staged, and maintained at the Zebrafish Facility of the University of Padua, under standard conditions (50). All husbandry and experimental procedures complied with European Legislation for the Protection of Animals used for Scientific Purposes (Directive 2010/63/EU) and with Italian law on animal experimentation (D.L. 4 March 2014, n.26). All the procedures were carried out under authorization n. 407/2015-PR from the Italian Ministry of Health. The project was also examined and approved by the Ethics Committee of the University of Padua with protocol number 18746.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siegel, R.; Miller, K.D.; Ahmedin, J. Cáncer Statistics. Ca Cáncer Journal 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Saeg, F.; Anbalagan, M. Breast Cancer Stem Cells and the Challenges of Eradication: A Review of Novel Therapies. Stem Cell Investig 2018, 5, 39. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; O’Hare, M.J.; Stein, R. Models of Breast Cancer: Is Merging Human and Animal Models the Future? Breast Cancer Res 2004, 6, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Mondal, P.; Bailey, K.L.; Cartwright, S.B.; Band, V.; Carlson, M.A. Large Animal Models of Breast Cancer. Frontiers in Oncology 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Rao, N. a. S.; van Wolferen, M.E.; van den Ham, R.; van Leenen, D.; Groot Koerkamp, M.J.A.; Holstege, F.C.P.; Mol, J.A. CDNA Microarray Profiles of Canine Mammary Tumour Cell Lines Reveal Deregulated Pathways Pertaining to Their Phenotype. Anim Genet 2008, 39, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, G.; Alonso-Diez, Á.; Pérez-Alenza, D.; Peña, L. From Conventional to Precision Therapy in Canine Mammary Cancer: A Comprehensive Review. Frontiers in Veterinary Science 2021, 8, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Abadie, J.; Nguyen, F.; Loussouarn, D.; Peña, L.; Gama, A.; Rieder, N.; Belousov, A.; Bemelmans, I.; Jaillardon, L.; Ibisch, C.; et al. Canine Invasive Mammary Carcinomas as Models of Human Breast Cancer. Part 2: Immunophenotypes and Prognostic Significance. Breast Cancer Research and Treatment 2018, 167, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-H.; Park, H.-M.; Son, K.-H.; Shin, T.-J.; Cho, J.-Y. Transcriptome Signatures of Canine Mammary Gland Tumors and Its Comparison to Human Breast Cancers. Cancers (Basel) 2018, 10, 317. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Lee, K.-H.; Nam, A.-R.; Cho, J.-Y. Genome-Wide Methylation Profiling in Canine Mammary Tumor Reveals MiRNA Candidates Associated with Human Breast Cancer. Cancers (Basel) 2019, 11, 1466. [Google Scholar] [CrossRef]
- Nasir, L.; Devlin, P.; Mckevitt, T.; Rutteman, G.; Argyle, D.J. Telomere Lengths and Telomerase Activity in Dog Tissues: A Potential Model System to Study Human Telomere and Telomerase Biology. Neoplasia 2001, 3, 351–359. [Google Scholar] [CrossRef]
- Meuten J Donald Tumors in Domestic Animals, 5th ed.; Ames: John Wiley and Sons lnc, 2017.
- Jaillardon, L.; Abadie, J.; Godard, T.; Campone, M.; Loussouarn, D.; Siliart, B.; Nguyen, F. The Dog as a Naturally-Occurring Model for Insulin-like Growth Factor Type 1 Receptor-Overexpressing Breast Cancer: An Observational Cohort Study. BMC Cancer 2015, 15, 664. [Google Scholar] [CrossRef] [PubMed]
- Restucci, B.; Maiolino, P.; Martano, M.; Esposito, G.; Filippis, D.D.E.; Borzacchiello, G.; Muzio, L.L.O. Expression of B-Catenin, E-Cadherin and APC in Canine Mammary Tumors. Anticancer Research 2007, 27, 3083–3090. [Google Scholar] [PubMed]
- Yu, F.; Yu, C.; Li, F.; Zuo, Y.; Wang, Y.; Yao, L.; Wu, C.; Wang, C.; Ye, L. Wnt/β-Catenin Signaling in Cancers and Targeted Therapies. Signal Transduct Target Ther 2021, 6, 307. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-L.; Lin, S.-G.; Mao, Y.-W.; Wu, J.-X.; Hu, C.-D.; Lv, R.; Zeng, H.-D.; Zhang, M.-H.; Lin, L.-Z.; Ouyang, S.-S.; et al. Wnt/β-Catenin Signalling Pathway in Breast Cancer Cells and Its Effect on Reversing Tumour Drug Resistance by Alkaloids Extracted from Traditional Chinese Medicine. Expert Rev Mol Med 2023, 25, e21. [Google Scholar] [CrossRef] [PubMed]
- Turashvili, G.; Bouchal, J.; Burkadze, G.; Kolar, Z. Wnt Signaling Pathway in Mammary Gland Development and Carcinogenesis. Pathobiology 2006, 73, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Zappulli, V.; De Cecco, S.; Trez, D.; Caliari, D.; Aresu, L.; Castagnaro, M. Immunohistochemical Expression of E-Cadherin and β-Catenin in Feline Mammary Tumours. Journal of Comparative Pathology 2012, 147, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Aplin, A.E.; Howe, A.; Alahari, S.K.; Juliano, R.L. Signal Transduction and Signal Modulation by Cell Adhesion Receptors: The Role of Integrins, Cadherins, Immunoglobulin-Cell Adhesion Molecules, and Selectins. Pharmacol Rev 1998, 50, 197–263. [Google Scholar] [PubMed]
- Geyer, F.C.; Lacroix-Triki, M.; Savage, K.; Arnedos, M.; Lambros, M.B.; MacKay, A.; Natrajan, R.; Reis-Filho, J.S. Β-Catenin Pathway Activation in Breast Cancer Is Associated with Triple-Negative Phenotype but Not with CTNNB1 Mutation. Modern Pathology 2011, 24, 209–231. [Google Scholar] [CrossRef]
- Abreu de Oliveira, W.A.; El Laithy, Y.; Bruna, A.; Annibali, D.; Lluis, F. Wnt Signaling in the Breast: From Development to Disease. Frontiers in Cell and Developmental Biology 2022, 10. [Google Scholar] [CrossRef]
- Hussain, S.; Saxena, S.; Shrivastava, S.; Mohanty, A.K.; Kumar, S.; Singh, R.J.; Kumar, A.; Wani, S.A.; Gandham, R.K.; Kumar, N.; et al. Gene Expression Profiling of Spontaneously Occurring Canine Mammary Tumours: Insight into Gene Networks and Pathways Linked to Cancer Pathogenesis. PLoS One 2018, 13, e0208656. [Google Scholar] [CrossRef]
- Yu, F.; Rasotto, R.; Zhang, H.; Pei, S.; Zhou, B.; Yang, X.; Jin, Y.; Zhang, D.; Lin, D. Evaluation of Expression of the Wnt Signaling Components. Journal of Veterinary Science 2017, 18, 359–367. [Google Scholar] [CrossRef]
- Sammarco, A.; Gomiero, C.; Sacchetto, R.; Beffagna, G.; Michieletto, S.; Orvieto, E.; Cavicchioli, L.; Gelain, M.E.; Ferro, S.; Patruno, M.; et al. Wnt/β-Catenin and Hippo Pathway Deregulation in Mammary Tumors of Humans, Dogs, and Cats. Vet Pathol 2020, 57, 774–790. [Google Scholar] [CrossRef] [PubMed]
- Cordenonsi, M.; Zanconato, F.; Azzolin, L.; Forcato, M.; Rosato, A.; Frasson, C.; Inui, M.; Montagner, M.; Parenti, A.R.; Poletti, A.; et al. The Hippo Transducer TAZ Confers Cancer Stem Cell-Related Traits on Breast Cancer Cells. Cell 2011, 147, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, X.; Song, G. The Hippo Pathway: A Master Regulatory Network Important in Cancer. Cells 2021, 10, 1416. [Google Scholar] [CrossRef] [PubMed]
- Beffagna, G.; Sacchetto, R.; Cavicchioli, L.; Sammarco, A.; Mainenti, M.; Ferro, S.; Trez, D.; Zulpo, M.; Michieletto, S.; Cecchinato, A.; et al. A Preliminary Investigation of the Role of the Transcription Co-Activators YAP/TAZ of the Hippo Signalling Pathway in Canine and Feline Mammary Tumours. Veterinary Journal 2016, 207, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Guillemette, S.; Rico, C.; Godin, P.; Boerboom, D.; Paquet, M. In Vitro Validation of the Hippo Pathway as a Pharmacological Target for Canine Mammary Gland Tumors. Journal of Mammary Gland Biology and Neoplasia 2017, 22, 203–214. [Google Scholar] [CrossRef]
- Rico, C.; Boerboom, D.; Paquet, M. Expression of the Hippo Signalling Effectors YAP and TAZ in Canine Mammary Gland Hyperplasia and Malignant Transformation of Mammary Tumours. Veterinary and Comparative Oncology 2018, 1–6. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, Y.-F. Natural Compounds as Anticancer Agents: Experimental Evidence. World J Exp Med 2012, 2, 45–57. [Google Scholar] [CrossRef]
- Chen, X.-W.; Di, Y.M.; Zhang, J.; Zhou, Z.-W.; Li, C.G.; Zhou, S.-F. Interaction of Herbal Compounds with Biological Targets: A Case Study with Berberine. ScientificWorldJournal 2012, 2012, 708292. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, C.; Yang, W. Role of Berberine in Alzheimer’s Disease. Neuropsychiatr Dis Treat 2016, 12, 2509–2520. [Google Scholar] [CrossRef]
- Chang, W. Non-Coding RNAs and Berberine: A New Mechanism of Its Anti-Diabetic Activities. Eur J Pharmacol 2017, 795, 8–12. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Baggioni, A. Berberine and Its Role in Chronic Disease. Adv Exp Med Biol 2016, 928, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Berberine and Inflammatory Bowel Disease: A Concise Review. Pharmacol Res 2016, 113, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Tillhon, M.; Guamán Ortiz, L.M.; Lombardi, P.; Scovassi, A.I. Berberine: New Perspectives for Old Remedies. Biochem Pharmacol 2012, 84, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Pierpaoli, E.; Arcamone, A.G.; Buzzetti, F.; Lombardi, P.; Salvatore, C.; Provinciali, M. Antitumor Effect of Novel Berberine Derivatives in Breast Cancer Cells. Biofactors 2013, 39, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Guamán Ortiz, L.M.; Lombardi, P.; Tillhon, M.; Scovassi, A.I. Berberine, an Epiphany Against Cancer. Molecules 2014, 19, 12349–12367. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; You, D.; Jeong, Y.; Yu, J.; Kim, S.W.; Nam, S.J.; Lee, J.E. Berberine Down-Regulates IL-8 Expression through Inhibition of the EGFR/MEK/ERK Pathway in Triple-Negative Breast Cancer Cells. Phytomedicine 2018, 50, 43–49. [Google Scholar] [CrossRef]
- Rauf, A.; Abu-Izneid, T.; Khalil, A.A.; Imran, M.; Shah, Z.A.; Emran, T.B.; Mitra, S.; Khan, Z.; Alhumaydhi, F.A.; Aljohani, A.S.M.; et al. Berberine as a Potential Anticancer Agent: A Comprehensive Review. Molecules 2021, 26, 7368. [Google Scholar] [CrossRef]
- Zhang, J.; Cao, H.; Zhang, B.; Cao, H.; Xu, X.; Ruan, H.; Yi, T.; Tan, L.; Qu, R.; Song, G.; et al. Berberine Potently Attenuates Intestinal Polyps Growth in ApcMin Mice and Familial Adenomatous Polyposis Patients through Inhibition of Wnt Signalling. J Cell Mol Med 2013, 17, 1484–1493. [Google Scholar] [CrossRef]
- Pierpaoli, E.; Damiani, E.; Orlando, F.; Lucarini, G.; Bartozzi, B.; Lombardi, P.; Salvatore, C.; Geroni, C.; Donati, A.; Provinciali, M. Antiangiogenic and Antitumor Activities of Berberine Derivative NAX014 Compound in a Transgenic Murine Model of HER2/Neu-Positive Mammary Carcinoma. Carcinogenesis 2015, 36, 1169–1179. [Google Scholar] [CrossRef]
- Ponnusamy, L.; Kothandan, G.; Manoharan, R. Berberine and Emodin Abrogates Breast Cancer Growth and Facilitates Apoptosis through Inactivation of SIK3-Induced MTOR and Akt Signaling Pathway. Biochim Biophys Acta Mol Basis Dis 2020, 1866, 165897. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M.; Kitaguchi, D.; Morinami, S.; Kurashiki, Y.; Hashida, H.; Miyata, S.; Yamaguchi, M.; Sakai, M.; Murata, N.; Tanaka, S. Berberine-Induced Nucleolar Stress Response in a Human Breast Cancer Cell Line. Biochem Biophys Res Commun 2020, 528, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Spinozzi, S.; Colliva, C.; Camborata, C.; Roberti, M.; Ianni, C.; Neri, F.; Calvarese, C.; Lisotti, A.; Mazzella, G.; Roda, A. Berberine and Its Metabolites: Relationship between Physicochemical Properties and Plasma Levels after Administration to Human Subjects. J Nat Prod 2014, 77, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, A.; Ikekawa, T.; Ikeda, Y.; Shirakawa, S.; Iigo, M. Antitumor Activity of Berberrubine Derivatives. Gan 1976, 67, 321–325. [Google Scholar] [PubMed]
- Ikekawa, T.; Ikeda, Y. Antitumor Activity of 13-Methyl-Berberrubine Derivatives. J Pharmacobiodyn 1982, 5, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Kwon, Y.; Kim, J.H.; Muller, M.T.; Chung, I.K. Induction of Topoisomerase II-Mediated DNA Cleavage by a Protoberberine Alkaloid, Berberrubine. Biochemistry 1998, 37, 16316–16324. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Y.; Saha, U.; Fiorillo, G.; Lombardi, P.; Kumar, G.S. Calorimetric Insights into the Interaction of Novel Berberrubine Derivatives with Human Telomeric G-Quadruplex DNA Sequence. Journal of Thermal Analysis and Calorimetry 2018, 132, 623–631. [Google Scholar] [CrossRef]
- Lombardi, P.; Buzzetti, F.; Arcamone, A.G. Benzoquinolizinium Salt Derivatives as Anticancer Agents 2011.
- Albring, K.F.; Weidemüller, J.; Mittag, S.; Weiske, J.; Friedrich, K.; Geroni, M.C.; Lombardi, P.; Huber, O. Berberine Acts as a Natural Inhibitor of Wnt/β-Catenin Signaling--Identification of More Active 13-Arylalkyl Derivatives. Biofactors 2013, 39, 652–662. [Google Scholar] [CrossRef]
- Guamán Ortiz, L.M.; Tillhon, M.; Parks, M.; Dutto, I.; Prosperi, E.; Savio, M.; Arcamone, A.G.; Buzzetti, F.; Lombardi, P.; Scovassi, A.I. Multiple Effects of Berberine Derivatives on Colon Cancer Cells. Biomed Res Int 2014, 2014, 924585. [Google Scholar] [CrossRef]
- Guamán Ortiz, L.M.; Croce, A.L.; Aredia, F.; Sapienza, S.; Fiorillo, G.; Syeda, T.M.; Buzzetti, F.; Lombardi, P.; Scovassi, A.I. Effect of New Berberine Derivatives on Colon Cancer Cells. Acta Biochim Biophys Sin (Shanghai) 2015, 47, 824–833. [Google Scholar] [CrossRef]
- Pierpaoli, E.; Fiorillo, G.; Lombardi, P.; Salvatore, C.; Geroni, C.; Piacenza, F.; Provinciali, M. Antitumor Activity of NAX060: A Novel Semisynthetic Berberine Derivative in Breast Cancer Cells. BioFactors 2018, 44, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Abrams, S.L.; Follo, M.Y.; Steelman, L.S.; Lertpiriyapong, K.; Cocco, L.; Ratti, S.; Martelli, A.M.; Candido, S.; Libra, M.; Murata, R.M.; et al. Abilities of Berberine and Chemically Modified Berberines to Inhibit Proliferation of Pancreatic Cancer Cells. Adv Biol Regul 2019, 71, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, K.; Ke, R.; Saini, K.S.; Viswakarma, N.; Nair, R.S.; Das, S.; Chen, Z.; Rana, A.; Rana, B. Berberine Represses β-Catenin Translation Involving 4E-BPs in Hepatocellular Carcinoma Cells. Mol Pharmacol 2021, 99, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pierpaoli, E.; Piacenza, F.; Fiorillo, G.; Lombardi, P.; Orlando, F.; Salvatore, C.; Geroni, C.; Provinciali, M. Antimetastatic and Antitumor Activities of Orally Administered NAX014 Compound in a Murine Model of HER2-Positive Breast Cancer. Int J Mol Sci 2021, 22, 2653. [Google Scholar] [CrossRef]
- Sacchetto, R.; Testoni, S.; Gentile, A.; Damiani, E.; Rossi, M.; Liguori, R.; Drögemüller, C.; Mascarello, F. A Defective SERCA1 Protein Is Responsible for Congenital Pseudomyotonia in Chianina Cattle. Am J Pathol 2009, 174, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev Dyn 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Sert, N.P. du; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE Guidelines 2.0: Updated Guidelines for Reporting Animal Research. PLOS Biology 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Moro, E.; Ozhan-Kizil, G.; Mongera, A.; Beis, D.; Wierzbicki, C.; Young, R.M.; Bournele, D.; Domenichini, A.; Valdivia, L.E.; Lum, L.; et al. In Vivo Wnt Signaling Tracing through a Transgenic Biosensor Fish Reveals Novel Activity Domains. Dev Biol 2012, 366, 327–340. [Google Scholar] [CrossRef]
- Astone, M.; Lai, J.K.H.; Dupont, S.; Stainier, D.Y.R.; Argenton, F.; Vettori, A. Zebrafish Mutants and TEAD Reporters Reveal Essential Functions for Yap and Taz in Posterior Cardinal Vein Development. Sci Rep 2018, 8, 10189. [Google Scholar] [CrossRef]
- Kuzmickiene, I.; Atkocius, V.; Aleknavicius, E.; Ostapenko, V. Impact of Season of Diagnosis on Mortality among Breast Cancer Survivors. J Cancer Res Ther 2018, 14, S1091–S1097. [Google Scholar] [CrossRef]
- Mouhid, L.; Corzo-Martínez, M.; Torres, C.; Vázquez, L.; Reglero, G.; Fornari, T.; Ramírez de Molina, A. Improving In Vivo Efficacy of Bioactive Molecules: An Overview of Potentially Antitumor Phytochemicals and Currently Available Lipid-Based Delivery Systems. J Oncol 2017, 2017, 7351976. [Google Scholar] [CrossRef] [PubMed]
- Anis, K.V.; Rajeshkumar, N.V.; Kuttan, R. Inhibition of Chemical Carcinogenesis by Berberine in Rats and Mice. J Pharm Pharmacol 2001, 53, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Xiong, R.-G.; Huang, S.-Y.; Wu, S.-X.; Zhou, D.-D.; Yang, Z.-J.; Saimaiti, A.; Zhao, C.-N.; Shang, A.; Zhang, Y.-J.; Gan, R.-Y.; et al. Anticancer Effects and Mechanisms of Berberine from Medicinal Herbs: An Update Review. Molecules 2022, 27, 4523. [Google Scholar] [CrossRef]
- Xu, J.; Long, Y.; Ni, L.; Yuan, X.; Yu, N.; Wu, R.; Tao, J.; Zhang, Y. Anticancer Effect of Berberine Based on Experimental Animal Models of Various Cancers: A Systematic Review and Meta-Analysis. BMC Cancer 2019, 19, 589. [Google Scholar] [CrossRef]
- Sefidabi, R.; Mortazavi, P.; Hosseini, S. Antiproliferative Effect of Berberine on Canine Mammary Gland Cancer Cell Culture. Biomedical Reports 2017, 6, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jing, Z.; Lv, J.; Zhang, Z.; Lin, J.; Cao, X.; Zhao, Z.; Liu, P.; Mao, W. Berberine Activates Caspase-9/Cytochrome c-Mediated Apoptosis to Suppress Triple-Negative Breast Cancer Cells in Vitro and in Vivo. Biomedicine & Pharmacotherapy 2017, 95, 18–24. [Google Scholar] [CrossRef]
- Refaat, A.; Abdelhamed, S.; Yagita, H.; Inoue, H.; Yokoyama, S.; Hayakawa, Y.; Saiki, I. Berberine Enhances Tumor Necrosis Factor-related Apoptosis-inducing Ligand-mediated Apoptosis in Breast Cancer. Oncology Letters 2013, 6, 840–844. [Google Scholar] [CrossRef]
- Kim, S.; Lee, J.; You, D.; Jeong, Y.; Jeon, M.; Yu, J.; Kim, S.W.; Nam, S.J.; Lee, J.E. Berberine Suppresses Cell Motility Through Downregulation of TGF-Β1 in Triple Negative Breast Cancer Cells. Cellular Physiology and Biochemistry 2018, 45, 795–807. [Google Scholar] [CrossRef]
- Karnam, K.C.; Ellutla, M.; Bodduluru, L.N.; Kasala, E.R.; Uppulapu, S.K.; Kalyankumarraju, M.; Lahkar, M. Preventive Effect of Berberine against DMBA-Induced Breast Cancer in Female Sprague Dawley Rats. Biomedicine & Pharmacotherapy 2017, 92, 207–214. [Google Scholar] [CrossRef]
- Damiani, E.; Pierpaoli, E.; Orlando, F.; Donati, A.; Provinciali, M. Sidestream Dark Field Videomicroscopy for in Vivo Evaluation of Vascularization and Perfusion of Mammary Tumours in HER2/Neu Transgenic Mice. Clinical and Experimental Pharmacology and Physiology 2015, 42, 225–229. [Google Scholar] [CrossRef]
- Su, K.; Hu, P.; Wang, X.; Kuang, C.; Xiang, Q.; Yang, F.; Xiang, J.; Zhu, S.; Wei, L.; Zhang, J. Tumor Suppressor Berberine Binds VASP to Inhibit Cell Migration in Basal-like Breast Cancer. Oncotarget 2016, 7, 45849–45862. [Google Scholar] [CrossRef]
- Marverti, G.; Ligabue, A.; Lombardi, P.; Ferrari, S.; Monti, M.G.; Frassineti, C.; Costi, M.P. Modulation of the Expression of Folate Cycle Enzymes and Polyamine Metabolism by Berberine in Cisplatin-Sensitive and -Resistant Human Ovarian Cancer Cells. International Journal of Oncology 2013, 43, 1269–1280. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. Apoptosis, Autophagy, Necroptosis, and Cancer Metastasis. Mol Cancer 2015, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-Y.; Yu, H.-Z.; Huang, S.-M.; Zheng, Y.-L. P53, Bcl-2 and Cox-2 Are Involved in Berberine Hydrochloride-Induced Apoptosis of HeLa229 Cells. Mol Med Rep 2016, 14, 3855–3861. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Li, L.; Zou, X.; Xu, L.; Yi, P. Berberine Attenuated Proliferation, Invasion and Migration by Targeting the AMPK/HNF4α/WNT5A Pathway in Gastric Carcinoma. Frontiers in Pharmacology 2018, 9. [Google Scholar]
- Dian, L.; Xu, Z.; Sun, Y.; Li, J.; Lu, H.; Zheng, M.; Wang, J.; Drobot, L.; Horak, I. Berberine Alkaloids Inhibit the Proliferation and Metastasis of Breast Carcinoma Cells Involving Wnt/β-Catenin Signaling and EMT. Phytochemistry 2022, 200, 113217. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Y.; Shi, C.-J.; Fu, W.-M.; Zhang, J.-F. Berberine Inhibits Tumour Growth in Vivo and in Vitro through Suppressing the LincROR-Wnt/β-Catenin Regulatory Axis in Colorectal Cancer. J Pharm Pharmacol 2023, 75, 129–138. [Google Scholar] [CrossRef]
- Nie, Q.; Peng, W.W.; Wang, Y.; Zhong, L.; Zhang, X.; Zeng, L. β-Catenin Correlates with the Progression of Colon Cancers and Berberine Inhibits the Proliferation of Colon Cancer Cells by Regulating the β-Catenin Signaling Pathway. Gene 2022, 818, 146207. [Google Scholar] [CrossRef]
- Bian, J.; Dannappel, M.; Wan, C.; Firestein, R. Transcriptional Regulation of Wnt/β-Catenin Pathway in Colorectal Cancer. Cells 2020, 9, 2125. [Google Scholar] [CrossRef]
- Wu, Z.; Guan, K.-L. Hippo Signaling in Embryogenesis and Development. Trends Biochem Sci 2021, 46, 51–63. [Google Scholar] [CrossRef]
- Calses, P.C.; Crawford, J.J.; Lill, J.R.; Dey, A. Hippo Pathway in Cancer: Aberrant Regulation and Therapeutic Opportunities. Trends Cancer 2019, 5, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337. [Google Scholar] [CrossRef]
- Zhu, C.; Li, L.; Zhao, B. The Regulation and Function of YAP Transcription Co-Activator. Acta Biochimica et Biophysica Sinica 2014, 47, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-L.; Li, Q.-Y.; Jin, M.-J.; Lu, C.-F.; Mu, Z.-Y.; Xu, W.-Y.; Song, J.; Zhang, Y.; Zhang, S.-Y. A Review: Hippo Signaling Pathway Promotes Tumor Invasion and Metastasis by Regulating Target Gene Expression. J Cancer Res Clin Oncol 2021, 147, 1569–1585. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.S.; Jiang, J. Hippo-Independent Regulation of Yki/Yap/Taz: A Non-Canonical View. Frontiers in Cell and Developmental Biology 2021, 9. [Google Scholar]
- Wang, Y.; Liu, Y.; Du, X.; Ma, H.; Yao, J. The Anti-Cancer Mechanisms of Berberine: A Review. Cancer Manag Res 2020, 12, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Liu, H.; Wang, J.; Wang, M.; Shao, R. Cyclizing-Berberine A35 Induces G2/M Arrest and Apoptosis by Activating YAP Phosphorylation (Ser127). Journal of Experimental & Clinical Cancer Research 2018, 37, 98. [Google Scholar] [CrossRef]
- Kang, Y.; Siegel, P.M.; Shu, W.; Drobnjak, M.; Kakonen, S.M.; Cordon-Cardo, C.; Guise, T.A.; Massague, J. A Multigenic Program Mediating Breast Cancer Metastasis to Bone. Cancer Cell 2003, 3, 537–549. [Google Scholar]
- Chang, C.-C.; Shih, J.-Y.; Jeng, Y.-M.; Su, J.-L.; Lin, B.-Z.; Chen, S.-T.; Chau, Y.-P.; Yang, P.-C.; Kuo, M.-L. Connective Tissue Growth Factor and Its Role in Lung Adenocarcinoma Invasion and Metastasis. J Natl Cancer Inst 2004, 96, 364–375. [Google Scholar] [CrossRef]
- Rebolledo, D.L.; Acuña, M.J.; Brandan, E. Role of Matricellular CCN Proteins in Skeletal Muscle: Focus on CCN2/CTGF and Its Regulation by Vasoactive Peptides. International Journal of Molecular Sciences 2021, 22, 5234. [Google Scholar] [CrossRef]
Figure 1.
Biochemical structure of berberine (BBR), berberrubine (BRR), and analogues.
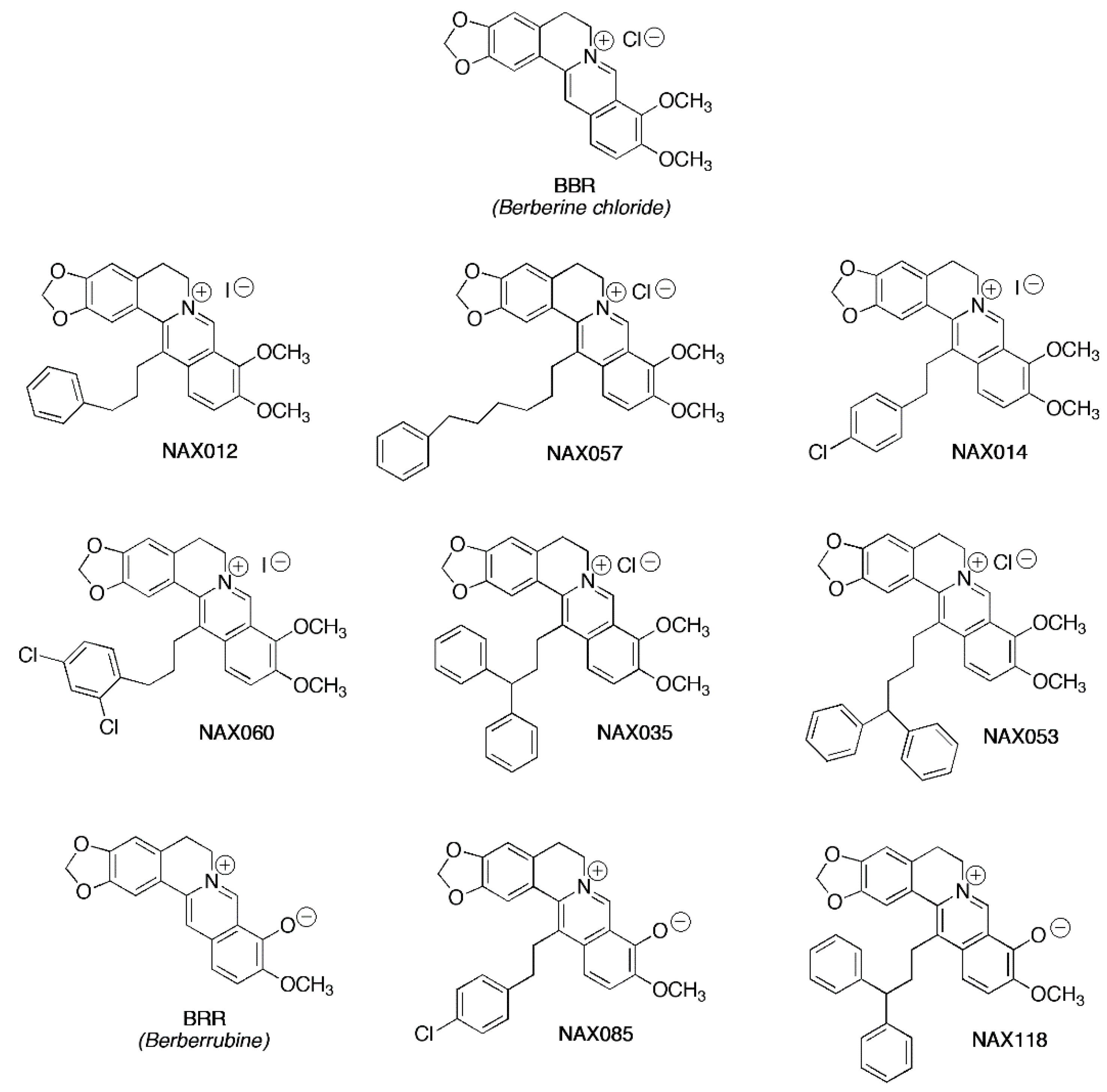
Figure 2.
Effect of berberine (BBR), berberrubine (BRR) and other analogues on CF33 cell viability. A) Cells were treated with 2 μM, 5 μM, 10 μM, 20 μM, and 40 μM of the different compounds for 24 hours. B) Cells were treated with 10 μM of the different compounds and cell viability was measured at different time points (3, 6, 12, 15, 18, and 24 hours). BBR, BRR and analogues exerted a dose- and time-dependent inhibition of cell viability.
Figure 2.
Effect of berberine (BBR), berberrubine (BRR) and other analogues on CF33 cell viability. A) Cells were treated with 2 μM, 5 μM, 10 μM, 20 μM, and 40 μM of the different compounds for 24 hours. B) Cells were treated with 10 μM of the different compounds and cell viability was measured at different time points (3, 6, 12, 15, 18, and 24 hours). BBR, BRR and analogues exerted a dose- and time-dependent inhibition of cell viability.
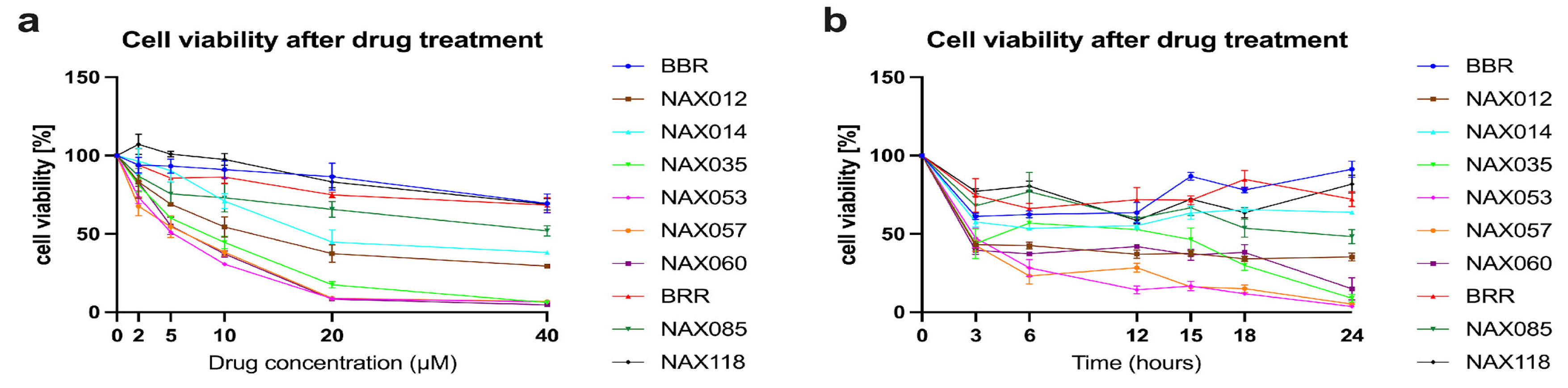
Figure 3.
Cell death after drug treatment. Early apoptosis and late apoptosis/necrosis rates of CF33 cells treated with 10 μM of the different compounds at A) 6h, B) 12h, C) 18h, and D) 24h, measured using flow cytometry with double staining for annexin V and propidium iodide.
Figure 3.
Cell death after drug treatment. Early apoptosis and late apoptosis/necrosis rates of CF33 cells treated with 10 μM of the different compounds at A) 6h, B) 12h, C) 18h, and D) 24h, measured using flow cytometry with double staining for annexin V and propidium iodide.
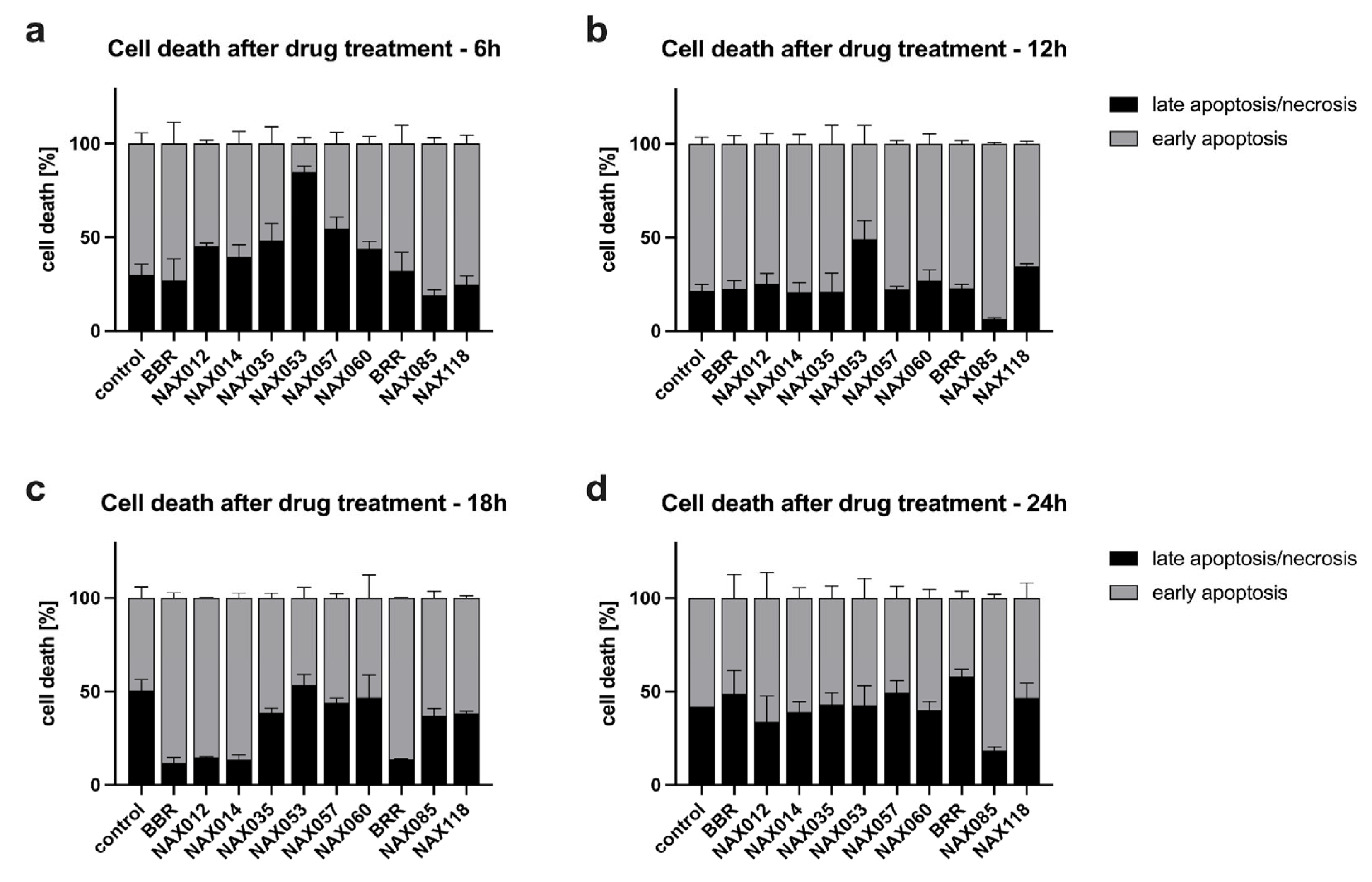
Figure 4.
Protein expression after drug treatment. Representative cropped western blots of YAP/TAZ, β-catenin and active β-catenin in protein lysates obtained from CF33 cells treated with berberine (BBR), NAX035, NAX053, NAX057, NAX060 and berberrubine (BRR) for 24 hours. Bands of the expected sizes for YAP (~70 kDa), TAZ (~55 kDa), β-catenin (~92 kDa) and active β-catenin (~92 kDa) are present in all samples. The experiment was performed in quadruplicate. Each lane corresponds to a replicate. Ponceau red was used as a loading control.
Figure 4.
Protein expression after drug treatment. Representative cropped western blots of YAP/TAZ, β-catenin and active β-catenin in protein lysates obtained from CF33 cells treated with berberine (BBR), NAX035, NAX053, NAX057, NAX060 and berberrubine (BRR) for 24 hours. Bands of the expected sizes for YAP (~70 kDa), TAZ (~55 kDa), β-catenin (~92 kDa) and active β-catenin (~92 kDa) are present in all samples. The experiment was performed in quadruplicate. Each lane corresponds to a replicate. Ponceau red was used as a loading control.
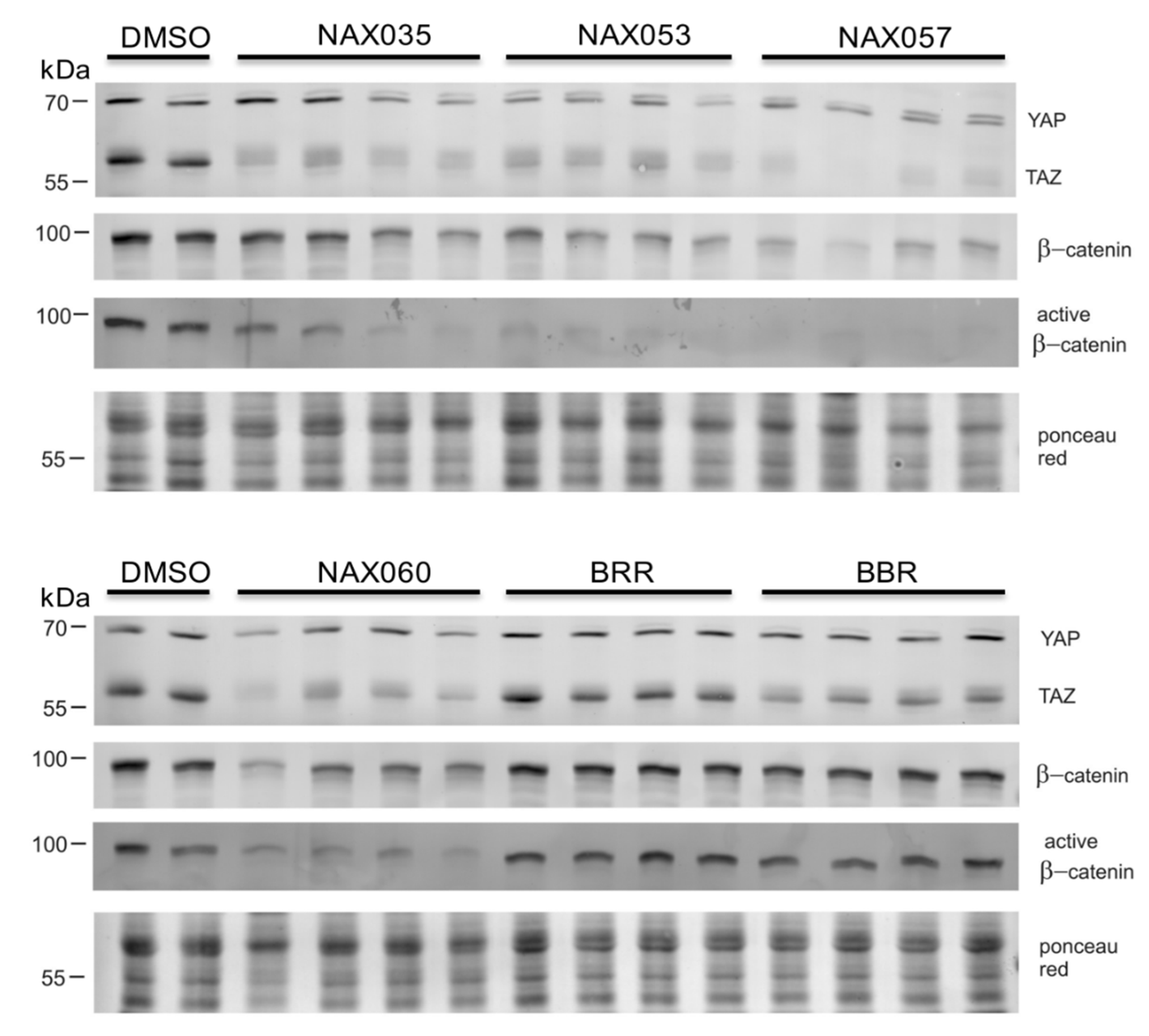
Figure 5.
mRNA expression after drug treatment. Gene expression analysis by semi-quantitative PCR of genes directly or indirectly involved in Wnt/β-catenin and Hippo pathways in CF33 cells treated with berberine (BBR), NAX035, NAX053, NAX057, NAX060 and berberrubine (BRR). A) β-catenin, B) YAP, C) TAZ, and D) CTGF mRNA expression levels in CF33 cells. Relative gene expression levels are shown following normalization with beta-actin. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Figure 5.
mRNA expression after drug treatment. Gene expression analysis by semi-quantitative PCR of genes directly or indirectly involved in Wnt/β-catenin and Hippo pathways in CF33 cells treated with berberine (BBR), NAX035, NAX053, NAX057, NAX060 and berberrubine (BRR). A) β-catenin, B) YAP, C) TAZ, and D) CTGF mRNA expression levels in CF33 cells. Relative gene expression levels are shown following normalization with beta-actin. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
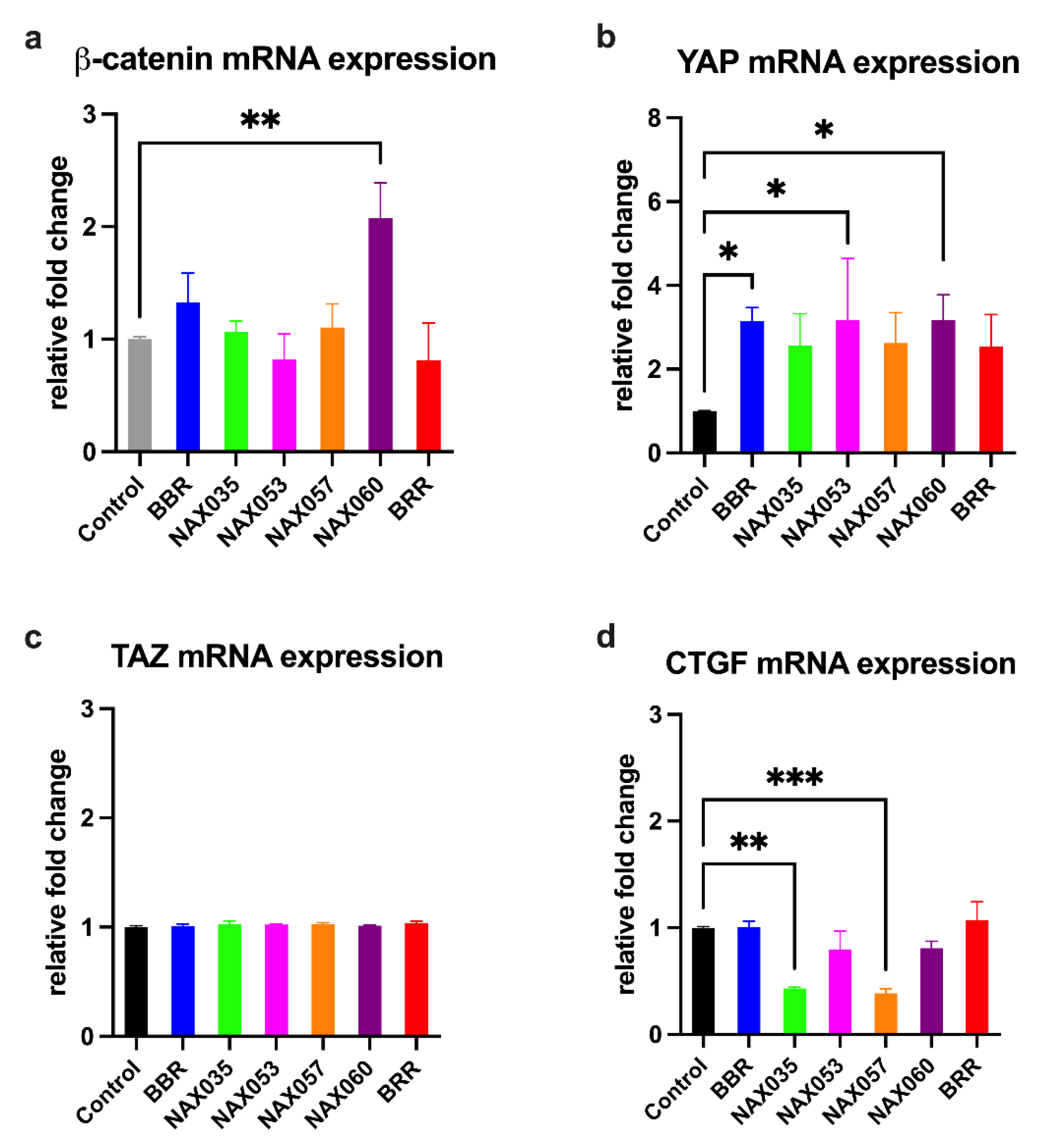
Figure 6.
Wnt/β-catenin and Hippo pathways zebrafish reporters. Tissue specificity expression of mCherry under control of A) TCF/LEF and of B) CTGF genes. Quantification of mCherry fluorescence in control transgenic zebrafish in comparison with transgenic zebrafish treated with berberine (BBR) and other analogues (NAX035, NAX053, NAX057). *, p < 0.05; ***, p < 0.001; ****, p < 0.0001.
Figure 6.
Wnt/β-catenin and Hippo pathways zebrafish reporters. Tissue specificity expression of mCherry under control of A) TCF/LEF and of B) CTGF genes. Quantification of mCherry fluorescence in control transgenic zebrafish in comparison with transgenic zebrafish treated with berberine (BBR) and other analogues (NAX035, NAX053, NAX057). *, p < 0.05; ***, p < 0.001; ****, p < 0.0001.
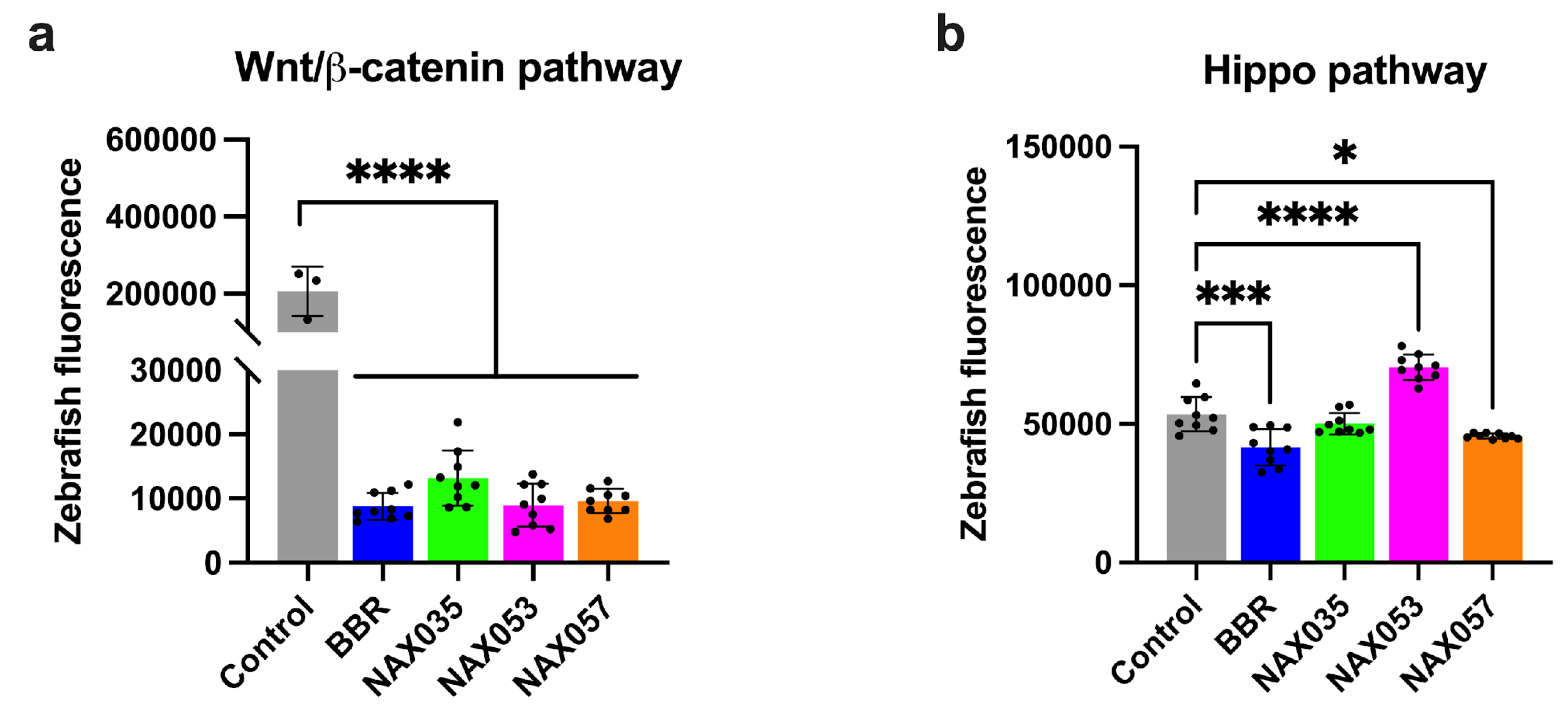

Table 1.
Primers and condition used for the semi-quantitative PCR analysis.
| Primer F (5’-3’) | Primer R (5’-3’) | Amplicon length (pb) | Number of cycles | |
|---|---|---|---|---|
| ACTB | TGGCACCACACCTTCTACAA | CCAGAGGCGTACAGGGATAG | 182 | 25 |
| β-catenin | ACACGTGCAATCCCTGAACT | CACCATCTGAGGAGAACGCA | 138 | 26 |
| TAZ | TCCAATCACCAGTCCTGCAT | AGCTCCTTGGTGAAGCAGAT | 125 | 28 |
| YAP | CCCAGACTACCTTGAAGCCA | CTTCCTGCAGACTTGGCATC | 107 | 28 |
| CTGF | CGACTGGAAGACACGTTTGG | AGGAGGCGTTGTCATTGGTA | 136 | 27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



