
Submitted:
23 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
Juvenile hormones (JH) play crucial roles in insect development, reproduction, and other physi-ological functions. Because JH is specific to arthropods, including insects, it has long been stud-ied for use in pest control. Accordingly, this study aimed to identify the substances that act as JH disruptors from edible plants. Curcuminoids generally exhibit a wide range of biological ac-tivities, such as antioxidant, anti-inflammatory, antibacterial, and insecticidal properties, and they exhibit insect growth inhibitory effects. Demethoxycurcumin and bisdemethoxycurcumin, two curcuminoids from the turmeric plant Curcuma longa L. inhibit the formation of a metho-prene-tolerant (Met)–Taiman (Tai) heterodimer complex as shown through in vitro yeast two-hybrid assays. An artificial diet containing 1% (w/v) demethoxycurcumin or bisdemethox-ycurcumin reduced the number of larvae, leading to no pupal development. Building on our re-sults regarding curcuminoids, researchers can use our study as a reference to develop eco-friendly pesticides.
Keywords:
Juvenile hormone
; Methoprene-tolerant
; diterpene
; Juvenile hormone disruptor
; Curcuminoids
; Demethoxycurcumin
; Bisdemethoxycurcumin
1. Introduction
While chemical insecticides with diverse insecticidal effects have long been utilized to control major pests, issues including ecosystem degradation, environmental contamination, and the emergence of resistance in target pests due to the improper use and overuse of these insecticides have arisen [1]. Therefore, eco-friendly insecticides that exhibit effective insecticidal effects without considerably affecting the environment have been actively researched, and several have been characterized from different types of plant extracts [2]. Plants use secondary metabolites as natural defenses against pests such as insects, including nicotine, rotenone, lianas, sabadala, pyrethrum, neem, and turmeric [3]. Turmeric is derived from the root of Curcuma longa L., a perennial herb belonging to the Zingiberaceae family [4,5], and contains approximately 60–70% carbohydrates, 6–8% protein, 5–10% fat, 3–7% minerals, and 6–13% moisture [5,6,7,8]. Within turmeric, there are more than 50 structurally related compounds known as curcuminoids, which make up approximately 3–5% of the total composition. The main curcuminoids include three commercially available compounds, curcumin, demethoxycurcumin (DMC), and bisdemethoxycurcumin (BDMC) [6,9]. Curcuminoids are well-known for their various pharmacological properties, including antidepressant, antioxidant, anti-inflammatory, hepatoprotective, antidiabetic, anticancer, and antibacterial effects [7,10,11,12,13,14]. However, research on insecticidal properties related to curcuminoids has been limited. Some research on insecticidal properties, studies have explored the efficacy of curcuminoids, including ovicidal activity on Helicoverpa armigera eggs [15] In particular, curcuminoids I, II, III inhibited the P-glycoprotein ATPase in H. armigera [16] and insecticidal activity against Aedes aegypti [17], and other curcuminoids showed high efficacy against Aedes albopictus and Culex pipiens larvae [18]. In particular, curcumin, demethoxycurcumin, curcumin-BF2 complex, and a monocarbonyl tetramethoxy-curcumin derivative showed remarkable larvicidal activity, indicating their potential as alternative agents for mosquito control strategies [18]. Also found that in C. pipiens, curcumin increased mortality in the early larval stages through acetylcholine esterase 1 (AchE1) inhibition [19]. Additionally, research has explored the anti-insect effects of curcuma essential oil, such as feeding inhibition, oviposition deterrence, and reproductive inhibition [20]. However, to our knowledge, studies on the juvenile hormones (JHs) of insects related to curcuminoids have not been conducted.
The intricate interplay between juvenile hormone (JH) and ecdysone in insect development is a finely tuned process. JH, synthesized in the corpus allatum interacts with ecdysone to maintain larval development during molts [21]. Beyond its role in molting, JH plays a multifaceted role in crucial physiological functions, encompassing reproduction, developmental regulation, pheromone production, and caste differentiation, particularly in social insects [22]. Methoprene-tolerant (Met) was identified as a JH receptor in the mutant studies of D. melanogaster, accelerating research on JH signaling in D. melanogaster [23]. Derived from the germ cell-expressed gene, Met is a member of the basic helix-loop-helix (bHLH) Per-Arnt-Sim (PAS) family [24,25,26]. Like other members of the bHLH-PAS transcription factor family, Met requires the formation of heterodimeric partner with other bHLH-PAS proteins for its activation [27]. JHs act through a receptor complex consisting of Met and steroid receptor coactivator (SRC; Taiman in D. melanogaster or βFtz-F1 Interacting Steroid Receptor Coactivator FISC in A. aegypti) to induce the transcription of specific genes [28]. Mets bind to JH with a high affinity, activating transcription [24,25,26]. Subsequently, Met heterodimerizes with an SRC [23]. To probe the impact of these interactions on the juvenile hormone signaling pathway, in vitro yeast two-hybrid assay systems have been employed, utilizing the JH-dependent heterodimer-binding properties of Met and SRC. These experimental approaches have facilitated the exploration of Juvenile Hormone Disruptor (JHD) activities in various plant extracts, studied not only in A. aegypti [29] but also in the Indian meal moth Plodia interpunctella [30]. Furthermore, an in vitro assay system capable of quantifying the disruption activity of plant extracts and diterpenes on JH-mediated Met/Taiman (Tai) heterodimer formation in D. melanogaster was developed [31]. The outcomes of these studies collectively underscore the prevalence of JHD diterpenes in the plant kingdom, emphasizing their propensity to interfere with JH-mediated endocrine regulation in insects [32].
In this study, we carefully validated the JHD activity of 30 herbal medicines and edible plant extracts, building on previous findings that highlighted their potent JHD effects [31]. Both the plant extracts from C. longa L and two curcuminoids, DMC and BDMC, showed strong JHD activity in the in vitro assay system of D. melanogaster Met-Tai. DMC and BDMC also blocked larval/pupal development, preventing the formation of pupae and emergence of adults.
2. Materials and Methods
2.1. Chemicals and Insects[29]
JH III and methoprene were purchased from Sigma-Aldrich (St. Louis, MO, USA). Three curcuminoids, curcumin, DMC, and BDMC, were purchased from Aladdin (Pudong, Shanghai, China). The plant diterpene methyl lucidone (LE3G) was isolated from Lindera erythrocarpa, as described in a previous study [29]. Plant extracts (i.e., methanol extracts of 30 edible plant species) were obtained from the Korean Plant Extracts Bank (Daejeon, South Korea). Each reagent was prepared as a stock solution in either dimethyl sulfoxide (DMSO) or methanol. D. melanogaster was grown in an instant fruit fly medium (Hansol Tech, Seoul, South Korea).
2.2. Yeast β-galactosidase Assay
The yeast two-hybrid binding assay, utilizing a quantitative β-galactosidase assay, was conducted with the Y187 strain transformed with the D. melanogaster juvenile hormone (JH) receptor and its interacting partner, Met-Tai, following the established protocol outlined in a prior study [31]. The Y187 strain, which was modified according to the protocol specified in the previous study [31], was grown in culture For the screening of JHD activity, 100 µL of cultured yeast cells with an optical density at 600 nm (OD600) ranging from 0.2 to 0.3 were exposed to a combination of 0.1 ppm JH III and 10 ppm individual plant extracts. This exposure was performed in 96-well plates, with each plate including a positive control comprising 0.1 ppm JH III with 10 ppm methyl lucidone and a negative control involving 0.1 ppm JH III with the control solvent (DMSO). Following a 3-hour incubation period, the cells were subjected to an assay for OD420, providing a quantitative measure of β-galactosidase activity. The OD420 values were then normalized to an arbitrary unit of JHD. Methyl lucidone served as a positive control due to its strong interference with JH III-mediated Met-Tai binding in the D. melanogaster β-galactosidase assay systems tested [31]. A single arbitrary unit of JHD activity was determined based on the binding interference observed with 10 ppm methyl lucidone. To assess the specific JHD activity of each plant extract, the average of duplicate experiments was calculated in arbitrary units using the following formula:
· A: The activity of the JHD, adjusted to A = 0 if A < 0. This adjustment ensures that negative values do not contribute to the specific JHD activity calculation.
· OD420 Control: The absorbance of yeast cells treated with JH III.
· OD420 PE: The absorbance of yeast cells treated with 0.1 ppm of JH III and 10 ppm of each plant extract.
· OD420 ML10: The absorbance of cells treated with 0.1 ppm of JH III and 10 ppm of methyl lucidone.
The formula essentially compares the observed effects of the plant extract with the positive and negative controls, providing a standardized measure of the specific JHD activity that ensures accurate and meaningful comparisons across different experimental conditions.
2.3. Bioassay
The experimental design commenced by introducing twenty male and twenty female flies separately into individual vials. Each vial was then subjected to a specific dietary regimen. The treatments included 1% (w/v) concentrations of JHD, namely LE3G, DMC, and BDMC, 0.05% (w/v) of the Juvenile Hormone Analog (JHA) methoprene, and a control group treated with 1% (w/v) ethanol. All these compounds were thoroughly incorporated into 3 g of artificial feed. To ensure the proper solidification of the diet and the subsequent evaporation of ethanol, the mixed diets were allowed to stand undisturbed at room temperature overnight. Subsequently, the vials containing the experimental diets were maintained at a controlled breeding temperature of 25 °C. Following a one-day period of oviposition, during which each vial accumulated approximately 150–250 eggs, the adult flies were removed from the vials. After an additional 3–4 days, second-instar larvae were collected from each vial, forming the basis for subsequent analyses.
2.4. RNA Extraction, Primers, and Quantitative Real Time Polymerase Chain Reaction (qPCR) Analysis
Total RNA was isolated from second instar larvae using the RNeasy isolation kit (Qiagen, Hilden, Germany). Subsequently, cDNA was synthesized for quantitative polymerase chain reaction (qPCR) with the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA) using 1 μg of RNA, quantified with a NanoDrop ND-1000 Spectrophotometer (Thermo Scientific, Waltham, MA, USA). The qRT-PCR primers were designed through NCBI Primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/), and the primer sequences can be found in Table 1. For qRT-PCR, the SsoAdvanced Universal SYBR Green Supermix (Bio-Rad) was employed in 96-well plates on a CFX Connect Real-Time Polymerase Chain Reaction System (Bio-Rad). The thermal cycling program consisted of an initial denaturation at 95 °C for 30 s, followed by 40 cycles of 95 °C for 10 s and 60 °C for 30 s. A final melting curve analysis spanned 95 °C for 5 s and 65 °C for 0.5 s. Validation of amplification efficiency and specificity was performed using CFX Maestro Software (Bio-Rad). To normalize the signal intensity of Kr-h1, it was compared to that of rp49. All RNA extractions and qRT-PCR analyses were conducted in triplicate, and the means of each sample were compared.
2.5. Data Analysis
The significance of treatment and control mean differences was assessed using the Student’s t-test. Statistical analyses and figure preparations were conducted using the GraphPad Prism 7 software.
3. Results
3.1. JHD Activity of Plant Extracts
In order to evaluate the JHD activities of different edible plant species on the formation of Met-Tai heterodimers in Drosophila melanogaster, 30 plant extracts, previously identified in a comprehensive study and mainly used as herbal medicines and edible plant extracts, were selected for investigation [32]. The selected plant extracts were tested for JHD activity in D. melanogaster using yeast-two hybrid β-galactosidase assays (Table 2). Among the extracts tested, 14 showed substantial JHD activity, with the highest activity observed in C. longa L. Notably, the curcuminoids present in Curcuma longa L. are linear diarylheptanoids that share similar size and structural characteristics with plant diterpenes and are recognized as well-known secondary metabolites of Curcuma plants. Motivated by these findings, we further investigated the JHD activities of three commercially available curcuminoids-curcumin, demethoxycurcumin (DMC), and bisdemethoxycurcumin (BDMC)-that are constituents of Curcuma longa L. (Figure 1). Interestingly, two of these curcuminoids, DMC and BDMC, exhibited robust interference with the heterodimer binding of MET-Tai while showing no anti-yeast activity. These results highlight the potential of curcuminoids, particularly DMC and BDMC, as potent modulators of JHD activity and shed light on their intricate role in interfering with juvenile hormone signaling pathways. Further investigations into the molecular mechanisms underlying this interference and the specific effects on insect physiology are warranted to fully understand the implications for pest control strategies.
3.2. Changes in the Emergence Rates of D. melanogaster Larvae According to Feeding
LE3G, an isolated diterpene from the extract of L. erythrocarpa, has previously been reported to interfere with larval development in mosquitoes, fruit flies, and moths [29,31,32], and this substance was used as a positive control in our study. Building on this, we explored the impact of two commercially available curcuminoids, demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC), on larval development in D. mlanogaster. Similar to the larvicidal effects observed with LE3G, the introduction of DMC or BDMC into the diets of D. melanogaster larvae resulted in a concentration-dependent inhibition of larval development. Notably, diets containing 1% (w/v) DMC or BDMC extract exhibited a pronounced disruption in larval development, preventing the progression to pupal stages (Figure 2). Furthermore, in diets supplemented with the JHD (LE3G, DMC, and BDMC at 1%, w/v), no adult flies were observed (Figure 3). This outcome suggests a significant impediment to the normal developmental transitions, emphasizing the potency of these compounds in disrupting the life cycle of D. melanogaster. These findings not only validate the larvicidal activity of LE3G but also extend it to the curcuminoids DMC and BDMC.
3.3. DMC and BDMC Effects on JH-dependent gene expression
Krüppel homolog 1 (Kr-h1), identified as an early juvenile hormone (JH) inducible gene, assumes a pivotal role in insect metamorphosis by orchestrating the activation of the JH/Met/Tai complex. In this study, we employed kr-h1 as a marker gene to elucidate its expression pattern in second-instar D. melanogaster larvae under the influence of various treatments. Upon treating D. melanogaster larvae with LE3G, demethoxycurcumin (DMC), and bisdemethoxycurcumin (BDMC), a consistent trend emerged, revealing a decrease in Kr-h1 expression compared to the control group (Figure 4). This downregulation suggests a potential inhibitory effect of these compounds on the JH/Met/Tai complex, aligning with their observed interference with larval development. In contrast, methoprene-treated flies exhibited an increase in Kr-h1 expression (Figure 4). Methoprene, a JH analog, is known to mimic the effects of JH in insects, promoting a response akin to natural JH induction. This elevation in Kr-h1 expression in methoprene-treated flies supports the expected activation of the JH signaling pathway. These findings underscore the modulatory impact of LE3G, DMC, and BDMC on Kr-h1 expression, indicative of their influence on the JH-regulated processes crucial for insect metamorphosis.
4. Discussion
Secondary metabolites in plants are produced as defense mechanisms against insects and other herbivores and have been used in this way throughout their evolutionary history [33,34]. Such secondary metabolites have become a research focus regarding the development of eco-friendly pesticides [35,36]. Plant-derived pesticides are considered ideal defense technologies because of their low toxicity, effectiveness against specific pests, and minimal environmental damage [37,38]. C. longa L, possessing insecticidal activity, has been utilized as an insecticide due to its diverse bioactive constituents that disrupt insect behavior and growth, making it possibly applicable for pest control in agriculture [39]. Curcuminoids, major components of C. longa rhizome powder, are known to show high insecticidal activity when used against various pests, such as Culex pipiens larvae [40]. Additionally, 45–60% growth inhibition and 10–15% mortality were confirmed as a result of insect growth inhibition assay in Schistocerca gregaria (Forsskål, 1775) and Dysdercus koenigii (Fabricius, 1775) by injecting the nymphs with curcuminoids [41]. In our study, D. melanogaster larvae fed with 1% curcuminoids failed to develop normally, suggesting that this substance inhibits larval/pupal development in relation to JH (Figure 3).
Curcuminoids, the yellow polyphenols responsible for the biological activity of turmeric oleoresin [42], are linear diarylheptanoids. Chemically, they are composed of two benzene rings linked by unsaturated chains, and they are a mixture of curcumin [1,7-bis(4-hydroxy-3-methoxy-phenyl)-hepta-1,6-diene-3,5-dione] and two derivatives, DMC and BDMC [43]. Curcumin contains methoxy groups on both ends of the benzene rings, whereas DMC lacks one of these methoxy groups. Conversely, BDMC has no methoxy groups and possesses a simpler structure than curcumin and DMC. Considering this structural difference, it is inferred that DMC and BDMC show higher Met-Tai binding inhibitory activity compared to curcumin, which is likely due to the variation in the number of methoxy groups on both ends of the benzene rings (Figure 5). Furthermore, methoprene and pyriproxyfen have similar ring-shaped backbone structures (Figure 5) similar to JH [44]. The cyclopentenedione rings of curcuminoids and LE3G consist of two α,β-unsaturated ketone functional groups [45]. These compounds may interfere with or disrupt the binding of the JH/Met/Tai complex by interacting with the methoxy and hydroxyl groups on both ends. However, explaining the chemical binding with JH as a singular determinant is challenging, as the interaction with JH receptors is regulated by various structural features and chemical properties of different compounds. Considering that curcuminoids have phenolic rings, α- and β-unsaturated carbonyl groups, and multiple substituents, these structural features may influence its interaction with JH receptors.
In a previous study, LE3G, an extract of L. erythrocarpa, showed the opposite pattern of Kr-h1 expression than that of JHD and methoprene [31]. JHD hinders larval development by interrupting heterodimer binding between Met and Tai through JH-mediating adjustments, whereas methoprene promotes heterodimer binding between Met and Tai by activating JH-mediating adjustments [46]. DMC and BDMC interfered with the development of Drosophila larvae (Figure 2), confirming that these substances had opposite effects compared with methoprene on Kr-h1 expression (Figure 4). Additionally, it is believed that when insects ingest a diet containing a mixture of DMC or BDMC, it disrupts the regulation of JH-mediated genes while also interfering with normal JH-dependent development. Therefore, the use of insecticidal methods using DMC or BDMC holds great promise for the formulation of novel pest control strategies. The appeal lies in their safety profile, as these substances biodegrade to non-toxic products. In addition, their potential suitability for integration into comprehensive pest management programs enhances their attractiveness as environmentally friendly alternatives. This is expected to serve as a model for the development of new synthetic analogs with favorable biological properties. Furthermore, as both curcuminoids are components of the tumeric, they would be classified as substances harmless to humans and animals. As a result, they can enhance environmental protection and the value of agricultural products. By maintaining the sustainability of insecticide production and preserving a healthy environment, these plant-derived insecticides are considered potential candidates for new insecticidal substances.
5. Conclusions
This study highlights the insecticidal effects of curcuminoids, which are known to be the major constituents of Curcuma longa L, particularly against Drosophila melanogaster larvae. Components of the curcuminoids, demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC), were shown to affect D. melanogaster larval and pupal development, suggesting that they interfere with juvenile hormone (JH)-related processes. Structural differences among curcuminoids, particularly in the number of methoxy groups, likely contribute to variations in their binding affinity with JH/Met/Tai complex. DMC and BDMC, whose effects on Kr-h1 expression were found to be opposite to those of methoprene, disrupt the heterodimeric bond between Met-taiman, which is important for JH-mediated regulation. More importantly, the use of DMC or BDMC as an eco-friendly pesticide suggests that these substances can be presented as a safe alternative to chemical pesticides with less environmental damage. In addition, the fact that these substances are classified as harmless to humans and animals and are environmentally friendly can contribute to environmental protection and enhancement of agricultural value. The potential applications of the plant-derived pesticides presented in this study are highlighted as sustainable and environmentally friendly alternatives in agriculture, and these substances are considered as potential candidates for new pesticide materials.
Author Contributions
Conceptualization, J.J., S.S., and H.O.; formal analysis, J.J., and J.S.; investigation, J.J., J.S., S.S., D.P., P.H., and O.H; writing—original draft preparation, J.J., S.S. and H.O.; funding acquisition, P.H., D.P and H.O.
Funding
This research was funded by the Rural Development Administration Republic of Korea grant number PJ0148452023 and Brain Pool Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science and ICT (2022H1D3A2A01053247)
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Lacey, L.A.; Lacey, C.M. The medical importance of riceland mosquitoes and their control using alternatives to chemical insecticides. Journal of the American Mosquito Control Association. Supplement 1990, 2, 1–93. [Google Scholar] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Rattan, R.S. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop protection 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Amalraj, A.; Kuttappan, S.; Matharu, A. Herbs, Spices and Their Roles in Nutraceuticals and Functional Foods; Elsevier, 2022. [Google Scholar]
- Zhang, H.A.; Kitts, D.D. Turmeric and its bioactive constituents trigger cell signaling mechanisms that protect against diabetes and cardiovascular diseases. Molecular and cellular biochemistry 2021, 476, 3785–3814. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: biological, pharmaceutical, nutraceutical, and analytical aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The essential medicinal chemistry of curcumin: miniperspective. Journal of medicinal chemistry 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Gupta, S.C.; Tyagi, A.K.; Aggarwal, B.B. Curcumin, a component of golden spice: from bedside to bench and back. Biotechnology advances 2014, 32, 1053–1064. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Huebbe, P.; Ernst, I.M.; Chin, D.; Wagner, A.E.; Rimbach, G. Curcumin—from molecule to biological function. Angewandte Chemie International Edition 2012, 51, 5308–5332. [Google Scholar] [CrossRef]
- Motterlini, R.; Foresti, R.; Bassi, R.; Green, C.J. Curcumin, an antioxidant and anti-inflammatory agent, induces heme oxygenase-1 and protects endothelial cells against oxidative stress. Free Radical Biology and Medicine 2000, 28, 1303–1312. [Google Scholar] [CrossRef]
- Labban, L. Medicinal and pharmacological properties of Turmeric (Curcuma longa): A review. Int J Pharm Biomed Sci 2014, 5, 17–23. [Google Scholar]
- Durgaprasad, S.; Pai, C.G.; Alvres, J.F. A pilot study of the antioxidant effect of curcumin in tropical pancreatitis. Indian Journal of Medical Research 2005, 122, 315. [Google Scholar] [PubMed]
- Ravindran, P.; Babu, K.N.; Sivaraman, K. Turmeric: the genus Curcuma; CRC press, 2007. [Google Scholar] [CrossRef]
- Basak, S.; Sarma, G.C.; Rangan, L. Ethnomedical uses of Zingiberaceous plants of Northeast India. Journal of ethnopharmacology 2010, 132, 286–296. [Google Scholar] [CrossRef]
- Solsoloy, A.; Cacayorin, N.; Cano, L. Insecticidal and fungicidal action of some indigenous plant extracts against cotton pests. Cotton Research Journal (Philippines) 1991. [Google Scholar]
- Aurade, R.M.; Jayalakshmi, S.K.; Sreeramulu, K. Modulatory effects of natural curcuminoids on P-glycoprotein ATPase of insecticide-resistant pest Helicoverpa armigera (Lepidopetera: Noctüidae). The Journal of Membrane Biology 2010, 236, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.N.; Chandra, A.; Nair, M.G. Novel bioactivities of Curcuma longa constituents. Journal of natural products 1998, 61, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Matiadis, D.; Liggri, P.G.; Kritsi, E.; Tzioumaki, N.; Zoumpoulakis, P.; Papachristos, D.P.; Balatsos, G.; Sagnou, M.; Michaelakis, A. Curcumin derivatives as potential mosquito larvicidal agents against two mosquito vectors, Culex pipiens and Aedes albopictus. International Journal of Molecular Sciences 2021, 22, 8915. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.; Goswami, D.; Rawal, R.M. Revealing the molecular interplay of curcumin as Culex pipiens Acetylcholine esterase 1 (AChE1) inhibitor. Scientific Reports 2021, 11, 17474. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Silva, A.S.; Varshney, R.; Chávez-González, M.L.; Singh, P. Curcuma-based botanicals as crop protectors: From knowledge to application in food crops. Current Research in Biotechnology 2021, 3, 235–248. [Google Scholar] [CrossRef]
- Truman, J.W.; Riddiford, L.M. The morphostatic actions of juvenile hormone. Insect biochemistry and molecular biology 2007, 37, 761–770. [Google Scholar] [CrossRef]
- Jindra, M.; Palli, S.R.; Riddiford, L.M. The juvenile hormone signaling pathway in insect development. Annu Rev Entomol 2013, 58, 181–204. [Google Scholar] [CrossRef]
- Li, M.; Mead, E.A.; Zhu, J. Heterodimer of two bHLH-PAS proteins mediates juvenile hormone-induced gene expression. Proc Natl Acad Sci U S A 2011, 108, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Oda, M.; Makita, S.; Chinzei, Y. Characterization of the Drosophila Methoprene -tolerant gene product. Juvenile hormone binding and ligand-dependent gene regulation. FEBS J 2005, 272, 1169–1178. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.P.; Iwema, T.; Epa, V.C.; Takaki, K.; Rynes, J.; Jindra, M. Ligand-binding properties of a juvenile hormone receptor, Methoprene-tolerant. Proc Natl Acad Sci U S A 2011, 108, 21128–21133. [Google Scholar] [CrossRef] [PubMed]
- Kayukawa, T.; Minakuchi, C.; Namiki, T.; Togawa, T.; Yoshiyama, M.; Kamimura, M.; Mita, K.; Imanishi, S.; Kiuchi, M.; Ishikawa, Y.; et al. Transcriptional regulation of juvenile hormone-mediated induction of Kruppel homolog 1, a repressor of insect metamorphosis. Proc Natl Acad Sci U S A 2012, 109, 11729–11734. [Google Scholar] [CrossRef]
- Kewley, R.J.; Whitelaw, M.L.; Chapman-Smith, A. The mammalian basic helix–loop–helix/PAS family of transcriptional regulators. The international journal of biochemistry & cell biology 2004, 36, 189–204. [Google Scholar] [CrossRef]
- Tumova, S.; Milacek, M.; Šnajdr, I.; Muthu, M.; Tuma, R.; Reha, D.; Jedlicka, P.; Bittova, L.; Novotna, A.; Majer, P. Unique peptidic agonists of a juvenile hormone receptor with species-specific effects on insect development and reproduction. Proceedings of the National Academy of Sciences 2022, 119, e2215541119. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Oh, H.W.; Fang, Y.; An, S.B.; Park, D.S.; Song, H.H.; Oh, S.R.; Kim, S.Y.; Kim, S.; Kim, N.; et al. Identification of plant compounds that disrupt the insect juvenile hormone receptor complex. Proc Natl Acad Sci U S A 2015, 112, 1733–1738. [Google Scholar] [CrossRef]
- Oh, H.W.; Yun, C.S.; Jeon, J.H.; Kim, J.A.; Park, D.S.; Ryu, H.W.; Oh, S.R.; Song, H.H.; Shin, Y.; Jung, C.S.; et al. Conifer Diterpene Resin Acids Disrupt Juvenile Hormone-Mediated Endocrine Regulation in the Indian Meal Moth Plodia interpunctella. J Chem Ecol 2017, 43, 703–711. [Google Scholar] [CrossRef]
- Shin, S.W.; Jeon, J.H.; Jeong, S.A.; Kim, J.A.; Park, D.S.; Shin, Y.; Oh, H.W. A plant diterpene counteracts juvenile hormone-mediated gene regulation during Drosophila melanogaster larval development. PLoS One 2018, 13, e0200706. [Google Scholar] [CrossRef]
- Shin, S.W.; Jeon, J.H.; Yun, C.S.; Jeong, S.A.; Kim, J.A.; Park, D.S.; Shin, Y.; Oh, H.W. Species-Specific Interactions between Plant Metabolites and Insect Juvenile Hormone Receptors. J Chem Ecol 2018, 44, 1022–1029. [Google Scholar] [CrossRef]
- Wink, M. Plant Secondary Metabolites Modulate Insect Behavior-Steps Toward Addiction? Front Physiol 2018, 9, 364. [Google Scholar] [CrossRef]
- Khare, S.; Singh, N.; Singh, A.; Hussain, I.; Niharika, K.; Yadav, V.; Bano, C.; Yadav, R.K.; Amist, N. Plant secondary metabolites synthesis and their regulations under biotic and abiotic constraints. Journal of Plant Biology 2020, 63, 203–216. [Google Scholar] [CrossRef]
- Mrid, R.B.; Benmrid, B.; Hafsa, J.; Boukcim, H.; Sobeh, M.; Yasri, A. Secondary metabolites as biostimulant and bioprotectant agents: A review. Science of the Total Environment 2021, 777, 146204. [Google Scholar] [CrossRef]
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K. Plant secondary metabolites as defense tools against herbivores for sustainable crop protection. International journal of molecular sciences 2022, 23, 2690. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Dubey, N. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of fruit and vegetables. Postharvest biology and Technology 2004, 32, 235–245. [Google Scholar] [CrossRef]
- Rajendran, S.; Sriranjini, V. Plant products as fumigants for stored-product insect control. Journal of stored products Research 2008, 44, 126–135. [Google Scholar] [CrossRef]
- Damalas, C.A. Potential uses of turmeric (‘Curcuma longa’) products as alternative means of pest management in crop production. Plant omics 2011, 4, 136–141. [Google Scholar]
- Sagnou, M.; Mitsopoulou, K.; Koliopoulos, G.; Pelecanou, M.; Couladouros, E.; Michaelakis, A. Evaluation of naturally occurring curcuminoids and related compounds against mosquito larvae. Acta tropica 2012, 123, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, H.; Walia, S.; Saxena, V.S. Isolation, characterization and insect growth inhibitory activity of major turmeric constituents and their derivatives against Schistocerca gregaria (Forsk) and Dysdercus koenigii (Walk). Pest Management Science: formerly Pesticide Science 2000, 56, 1086–1092. [Google Scholar] [CrossRef]
- Sahne, F.; Mohammadi, M.; Najafpour, G.D.; Moghadamnia, A.A. Enzyme-assisted ionic liquid extraction of bioactive compound from turmeric (Curcuma longa L.): Isolation, purification and analysis of curcumin. Industrial crops and products 2017, 95, 686–694. [Google Scholar] [CrossRef]
- Siviero, A.; Gallo, E.; Maggini, V.; Gori, L.; Mugelli, A.; Firenzuoli, F.; Vannacci, A. Curcumin, a golden spice with a low bioavailability. Journal of Herbal Medicine 2015, 5, 57–70. [Google Scholar] [CrossRef]
- Sláma, K. The history and current status of juvenoids. In Proceedings of the Proceedings of 3rd International Conference on Urban Pests, 1999; pp. 9–25.
- Senthil Kumar, K.; Wang, S.-Y. Pharmacological Applications of Lucidone: A Naturally Occurring Cyclopentenedione. Medicinal Plants-Recent Advances in Research and Development 2016, 273–295. [Google Scholar] [CrossRef]
- Shin, S.-W.; Jeon, J.-H.; Kim, J.-A.; Park, D.-S.; Shin, Y.-J.; Oh, H.-W. Inducible Expression of Several Drosophila melanogaster Genes Encoding Juvenile Hormone Binding Proteins by a Plant Diterpene Secondary Metabolite, Methyl Lucidone. Insects 2022, 13, 420. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
JHD activities of extracts from Curcuma longa L and two curcuminoids, demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC). (A) Met-Tai binding triggered by the addition of 0.1 ppm of JH III and the inhibition of the Met-Tai binding activity by treatment with 10 ppm of methyl lucidone (LE3G) was used as a positive control. The plant extract and compounds were added to the yeast culture at each shown concentration. (B) Anti-yeast activity tests of C. longa L with JHD activity. Corresponding compounds were tested for their anti-yeast activity to investigate whether the reduced β-galactosidase activity resulted from JHD activity or anti-yeast toxicity. Mean values with associated error bars are presented as means ± standard deviation (n = 3).
Figure 1.
JHD activities of extracts from Curcuma longa L and two curcuminoids, demethoxycurcumin (DMC) and bisdemethoxycurcumin (BDMC). (A) Met-Tai binding triggered by the addition of 0.1 ppm of JH III and the inhibition of the Met-Tai binding activity by treatment with 10 ppm of methyl lucidone (LE3G) was used as a positive control. The plant extract and compounds were added to the yeast culture at each shown concentration. (B) Anti-yeast activity tests of C. longa L with JHD activity. Corresponding compounds were tested for their anti-yeast activity to investigate whether the reduced β-galactosidase activity resulted from JHD activity or anti-yeast toxicity. Mean values with associated error bars are presented as means ± standard deviation (n = 3).
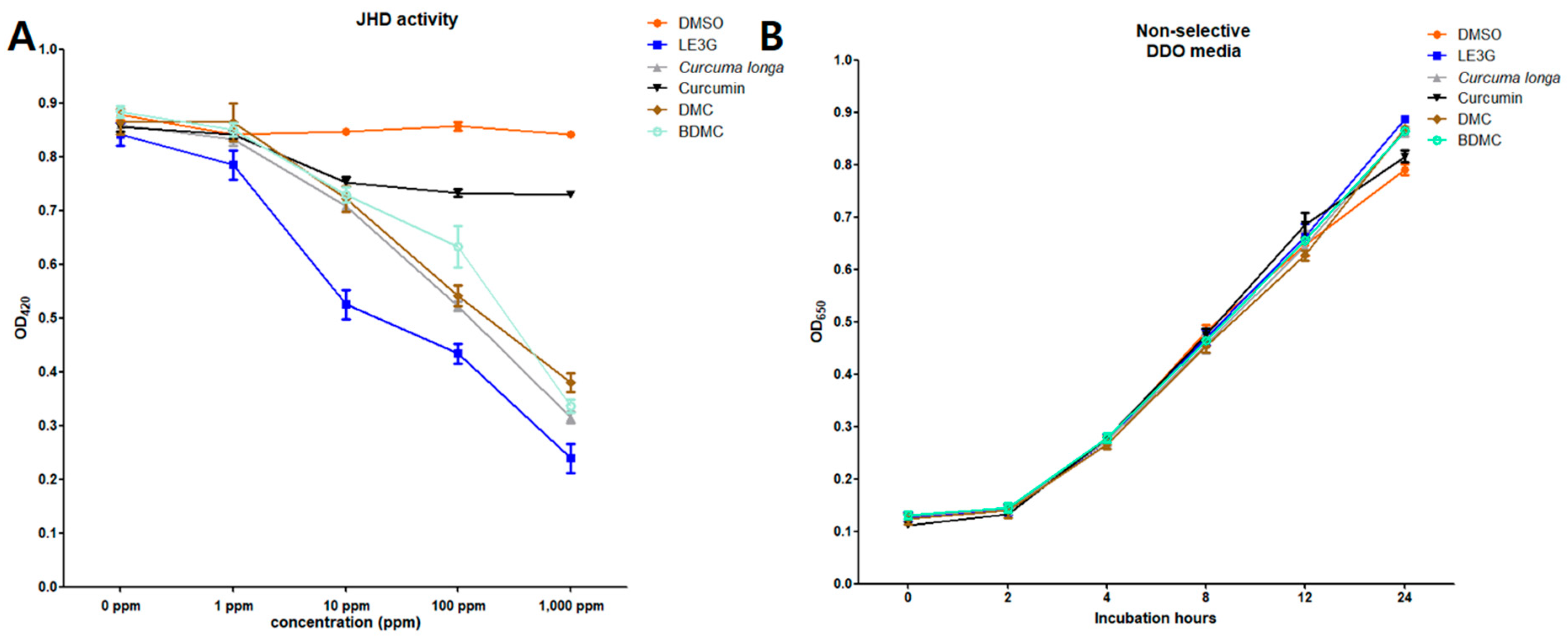
Figure 2.
Effect of JHA (methoprene) or JHD (LE3G, DMC, and BDMC) compounds on Drosophila melanogaster larval development.
Figure 2.
Effect of JHA (methoprene) or JHD (LE3G, DMC, and BDMC) compounds on Drosophila melanogaster larval development.

Figure 3.
Effect of DMC and BDMC on the growth of Drosophila melanogaster larvae. Artificial diets were supplemented with either 1% (w/v) ethanol (control) or JHD compounds (LE3G, DMC, and BDMC], or 0.05% (w/v) JHA (Methoprene). Mean values with associated error bars are presented as means ± standard deviation (n = 3).. * p < 0.01 and ** p < 0.001 (t-test).
Figure 3.
Effect of DMC and BDMC on the growth of Drosophila melanogaster larvae. Artificial diets were supplemented with either 1% (w/v) ethanol (control) or JHD compounds (LE3G, DMC, and BDMC], or 0.05% (w/v) JHA (Methoprene). Mean values with associated error bars are presented as means ± standard deviation (n = 3).. * p < 0.01 and ** p < 0.001 (t-test).
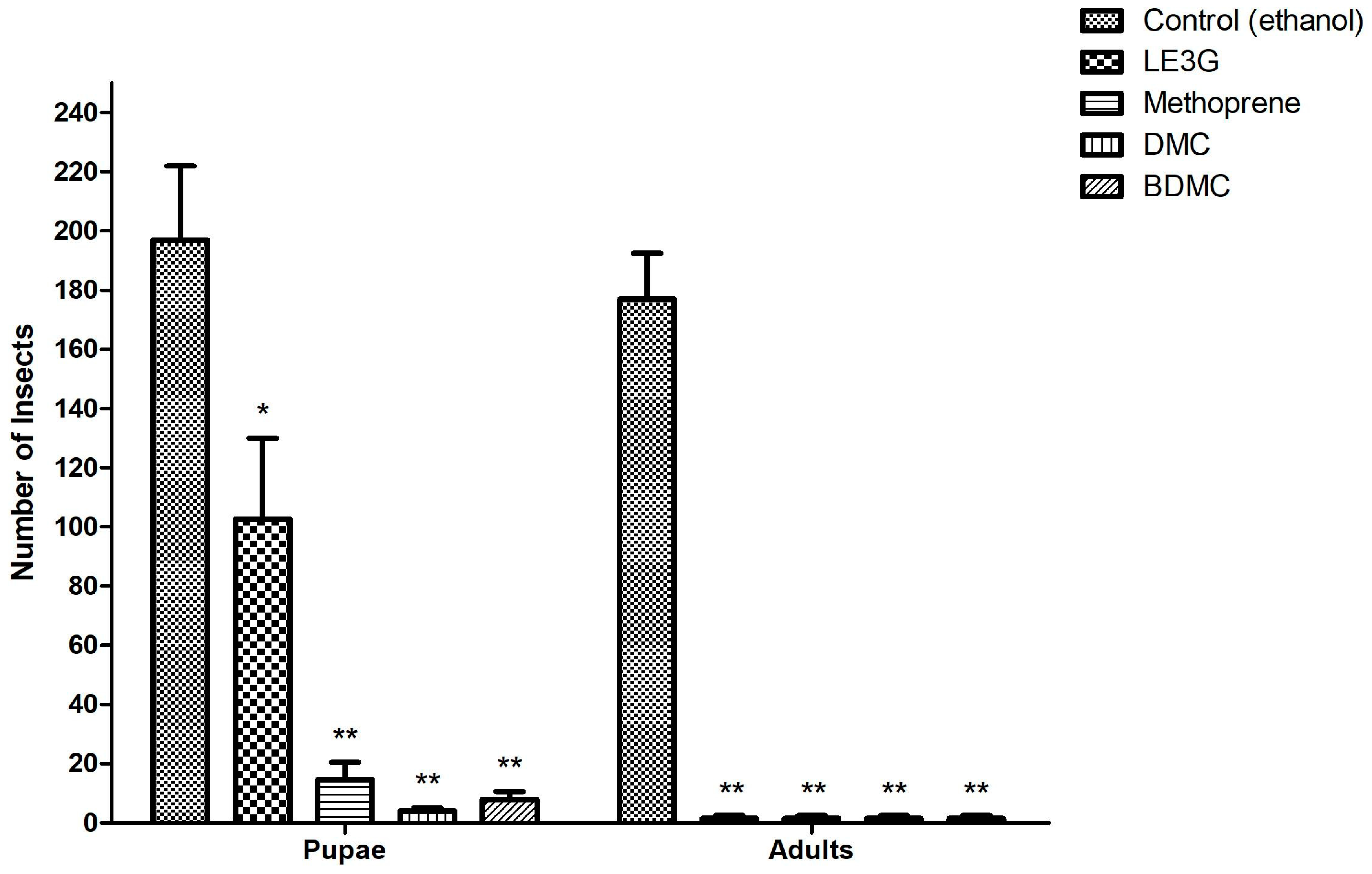
Figure 4.
Krüppel homolog 1 (Kr-h1) expression after DMC or BDMC treatments. Total RNA was extracted from second instar larvae that were fed with a diet containing each component and analyzed using qRT-PCR. Values and error bars indicate means ± SD (n = 3). *, p < 0.05, **, p < 0.01 (t-test).
Figure 4.
Krüppel homolog 1 (Kr-h1) expression after DMC or BDMC treatments. Total RNA was extracted from second instar larvae that were fed with a diet containing each component and analyzed using qRT-PCR. Values and error bars indicate means ± SD (n = 3). *, p < 0.05, **, p < 0.01 (t-test).
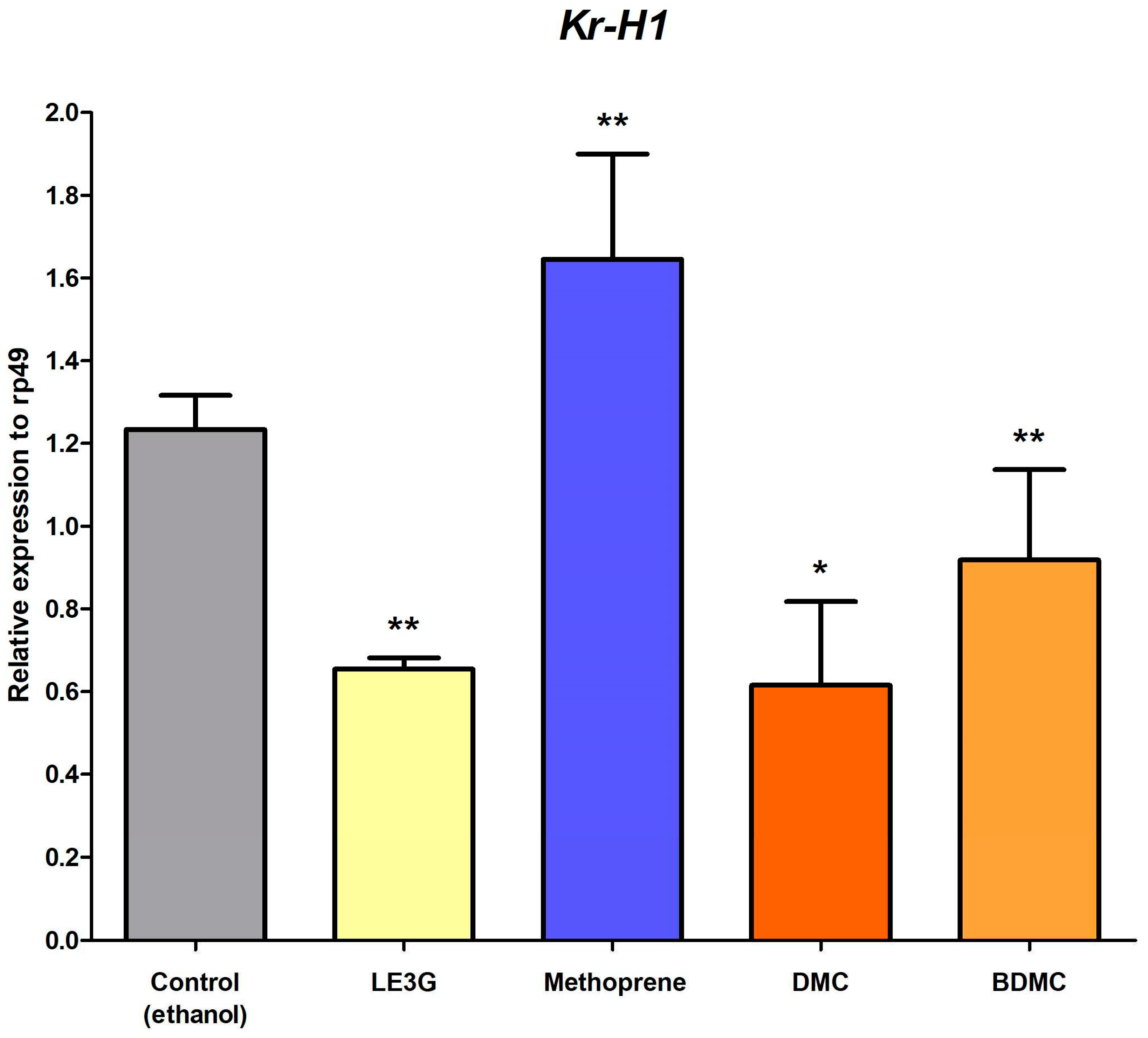
Figure 5.
Chemical structures of JHD and JH analog (JHa). JHD: (A) Curcumin, (B) DMC, (C) BDMC, (D) LE3G, JHa: (E) Methoprene, and (F) Pyriproxyfen. Red and blue shapes indicate α,β-unsaturated ketone moiety.
Figure 5.
Chemical structures of JHD and JH analog (JHa). JHD: (A) Curcumin, (B) DMC, (C) BDMC, (D) LE3G, JHa: (E) Methoprene, and (F) Pyriproxyfen. Red and blue shapes indicate α,β-unsaturated ketone moiety.
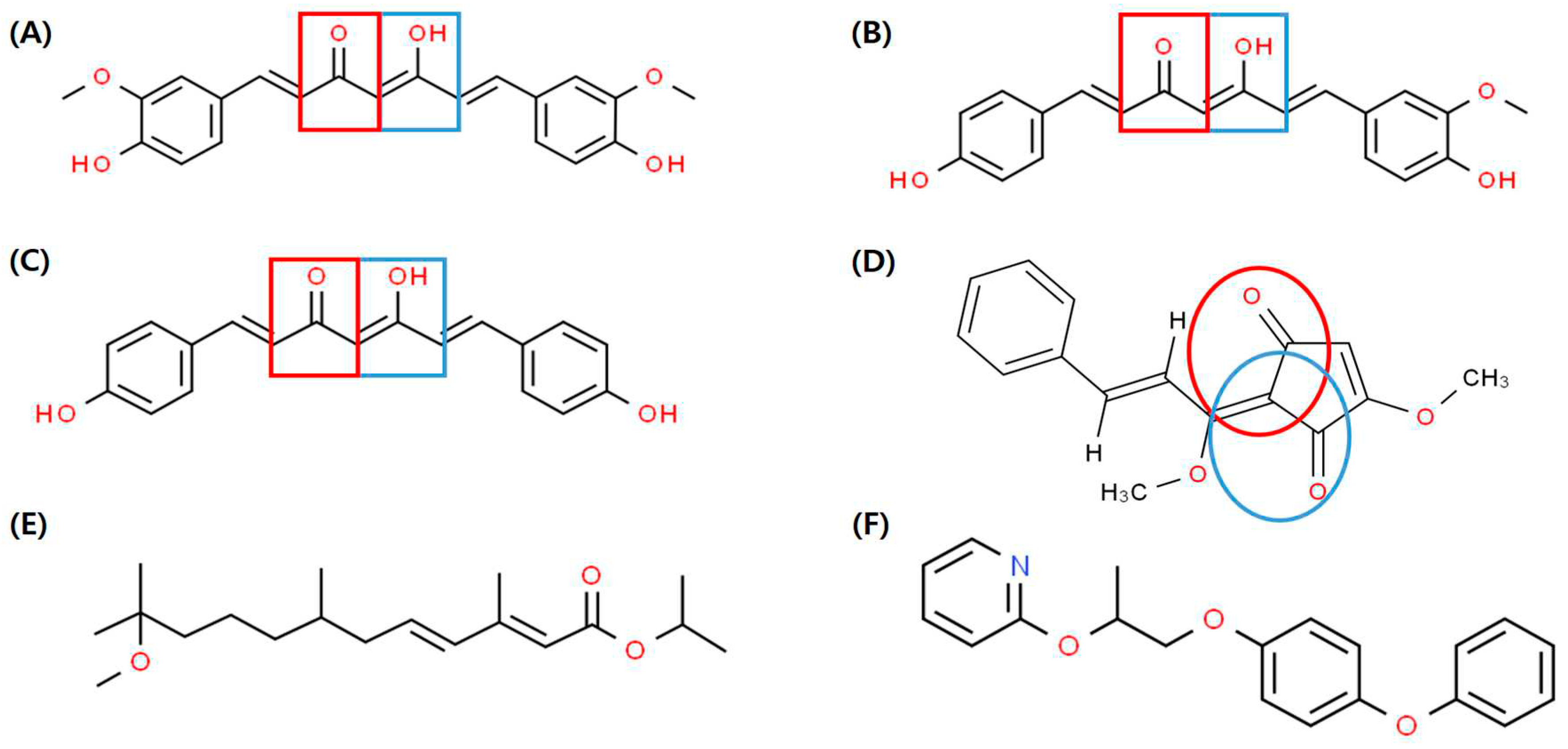

Table 1.
Primer sequences used for qRT-PCR.
| Gene | Primer |
|---|---|
| Krüppel homolog 1 | Forward 5′-TCACACATCAAGAAGCCAACT-3′ Reverse 5′-GCTGGTTGGCGGAATAGTAA-3′ |
| Rp49 | Forward 5′-ATGCTAAGCTGTCGCACAAATG-3′ Reverse 5′-GTTCGATCCGTAACCGATGT-3′ |
Table 2.
Juvenile hormone disrupter (JHD) activity of plant extracts.
| Species | Plant part | Family | JHD activity |
|---|---|---|---|
| Curcuma longa L. | Root | Zingiberaceae | 1.217175 |
| Pulsatilla koreana Nakai | Root | Ranunculaceae | 1.101688 |
| Machilus thunbergii Siebold & Zucc. | Trunk-bark | Lauraceae | 1.056931 |
| Echinosophora koreensis Nakai | Root | Fabaceae | 0.960778 |
| Pinus densiflora Siebold & Zucc. | Trunk-bark | Pinaceae | 0.936298 |
| Alpinia officinarum | Root | Zingiberaceae | 0.904591 |
| Smilax sieboldii Miq. | Leaf | Liliaceae | 0.886733 |
| Scutellaria baicalensis | Flower | Labiatae | 0.825496 |
| Portulaca oleracea L. | Whole | Portulacaceae | 0.806894 |
| Magnolia kobus DC | Leaf | Magnoliaceae | 0.785417 |
| Broussonetia papyrifera (L.) L’Hér. ex Vent. | Leaf | Moraceae | 0.768187 |
| Syringa patula (Palib.) Nakai | Leaf | Oleaceae | 0.759335 |
| Agrimonia pilosa Ledeb. | Whole | Rosaceae | 0.750968 |
| Cudrania tricuspidata (Carr.) Bureau ex Lavallée | Fruit | Moraceae | 0.745534 |
| Psoralea corylifolia (Babchi) | Seed | Fabaceae | 0.658419 |
| Phlomis umbrosa Turcz. | Whole | Labiatae | 0.65772 |
| Cudrania tricuspidata (Carr.) Bureau ex Lavallée | Trunk | Moraceae | 0.613586 |
| Zingiber officinale | Root | Zingiberaceae | 0.546682 |
| Myristica fragrans | Seed | Myristicaceae | 0.535624 |
| Saururus chinensis (Lour.) Baill. | Whole | Saururaceae | 0.493735 |
| Glycyrrhiza uralensis | Root | Fabaceae | 0.469728 |
| Picrasma quassioides (D. Don) Benn. | Trunk | Simaroubaceae | 0.433324 |
| Sophora flavescens | Root | Fabaceae | 0.391224 |
| Morus alba L. | Root | Moraceae | 0.390836 |
| Actinostemma lobatum Maxim. | Whole | Cucurbitaceae | 0.381283 |
| Salvia miltiorrhiza Bunge | Whole | Labiatae | 0.363577 |
| Cudrania tricuspidata (Carr.) Bureau ex Lavallée | Root | Moraceae | 0.257315 |
| Eclipta prostrata | Whole | Compositae | 0.237714 |
| Magnolia obovata Thunb | Trunk-bark | Lauraceae | 0.176234 |
| Angelica keiskei | Leaf | Apiaceae | 0.134399 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



