
Submitted:
22 November 2023
Posted:
23 November 2023
You are already at the latest version
Abstract
The diagnosis and the pathology of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS) remain under debate. However, there is a growing body of evidence for an autoimmune component in ME/CFS caused by the Epstein-Barr virus (EBV) and other viral infections. In this work, we took advantage of a large public dataset on the IgG antibodies to 3,054 EBV peptides to understand whether these immune responses could be used as putative biomarkers for disease diagnosis and triggers of pathological autoimmunity in ME/CFS patients using healthy controls (HCs) as a comparator cohort. We then aimed at predicting disease status of study participants using a Super Learner algorithm targeting an accuracy of 85% when splitting data into train and test datasets. When we compared data of all ME/CFS patients or data of a subgroup of these patients with non-infectious or unknown disease trigger to the dataset of HC, we could not find an antibody-based classifier that would meet the desired accuracy in the test dataset. In contrast, we could identify a 26-antibody classifier that could distinguish ME/CFS patients with an infectious disease trigger from HCs with 100% and 90% accuracies on the train and test sets, respectively. We finally performed a bioinformatic analysis of the EBV peptides associated with these 26 antibodies. We found no correlation between the importance metric of the selected antibodies in the classifier and the maximal sequence homology between human proteins and each EBV peptide recognized by these antibodies. In conclusion, these 26 antibodies against EBV have an effective potential for disease diagnosis of a subset of patients, but they are less likely to trigger pathological autoimmune responses that could explain the pathogenesis of ME/CFS.
Keywords:
Biomarker discovery
; disease pathogenesis
; autoimmunity
; antigenic mimicry
; machine learning
1. Introduction
The clinical manifestation of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS) is typically physical or mental post-exertional malaise, a persistent fatigue that is not alleviated by rest, together with other symptoms related to neurologic, autonomic, immunologic systems [1]. Several pathological mechanisms have been proposed to explain the origin of the disease and its progression over time [1,2,3,4,5]. Among these mechanisms, deleterious autoimmunity mostly driven by viruses is gaining traction in the literature [6,7,8]. SARS-CoV-2 is the newest causative agent of ME/CFS given that long-COVID patients might also receive a diagnosis of this disease [9,10,11,12]. However, the Epstein-Barr virus (EBV) among other herpesviruses remain the usual culprits of ME/CFS and now of long-COVID [13,14,15,16]. EBV is a particular interesting virus given its strong potential for antigenic mimicry with human proteins, especially, the EBNA1 protein that contains highly repetitive glycine-alanine motifs [17,18]. This potential of eliciting autoimmunity motivated serological investigations in patients with ME/CFS to identify key pathological EBV antigens and peptides [19,20,21,22]. However, these efforts did not lead to the identification of specific anti-EBV antibodies with a high accuracy of distinguishing patients with ME/CFS from heathy controls (HCs).
These disappointing findings could be simply explained by disease heterogeneity, selection bias, and misdiagnosis [23,24]. An alternative explanation is an inadequate choice of antibodies under analysis [25]. To overcome this limitation, a recent study performed a large screening of IgG antibody responses to more 3,000 EBV peptides in patients with ME/CFS and HCs [26]. In a subsequent study on the same data, antibody responses to two peptides were identified as candidate biomarkers for the subgroup of ME/CFS patients whose disease started with an infection. Notwithstanding providing the basis for a good disease diagnostic tool, these antibody responses were included in simple statistical models based on linear functions between covariates and disease status. Therefore, previous study was likely to fail detecting alternative antibody responses with more complex statistical relationships with disease status. Also, the same study did not evaluate eventual problems related to data overfitting.
The present paper aimed at re-analyzing the same dataset with the objective of using a machine learning approach where the identified analytical limitations could be tackled. We also took the chance of re-evaluating the role of molecular mimicry between EBV and human antigens on ME/CFS.
2. Materials and Methods
2.1. Study participants
Given that this study is based on a data re-analysis, the reader is recommended to consult the description of the original study in the respective reference [26]. Briefly, a total of 92 patients with ME/CFS patients were recruited from the Charité outpatient clinic for immunodeficiencies at the Institute of Medical Immunology in the Charité Universitatsmedizin Berlin, Germany. Fifty-four of these patients reported an acute infection at the beginning of their disease symptoms. The remaining patients (n=38) reported not knowing their disease trigger or reported a disease trigger other than an infection. A total of 50 self-reported HCs were recruited from the staff of the same clinic.
Age and gender distributions of the ME/CFS cohort as a whole or divided in its two subgroups were matched with the ones of HC. See the corresponding analysis in ref. [27].
2.2. Serological Data
The serological dataset under analysis is publicly available (see Supplementary File S1 of ref. [26]). Briefly, the serological dataset was generated by a seroarray that measured the signal intensities from IgG antibody responses to 3,054 EBV peptides. These peptides were derived from 14 EBV proteins: BALF2, BALF5, BFRF3, BLLF1, BLLF3, BLRF2, BMRF1, BZLF1, EBNA1, EBNA3, EBNA4, EBNA6, LMP1 and LMP2. The peptides had a length of 15 amino acids (15-mer) and they could overlap within the same protein.
To denote each peptide, we used the protein name and its starting position within the corresponding protein using the reference strain AG876. When the peptide name includes *, it referred to the starting position of the reference strain B95-8.
2.2. Data Splitting
Before conducting any analyses, the original dataset was divided into a train and test subsets using a 9:1 ratio while maintaining the proportions of ME/CFS patients and its subgroups, and HCs. This splitting ratio was approximately the optimal splitting ratio when applying a linear regression model that explains the data well at the cost of 81 covariates [28].
2.3. Machine Learning Approach
We performed an initial step where we ranked the antibodies according to their importance in discriminating ME/CFS patients from HCs (Supplementary Figure 1). In this step, we estimated 2500 Random Forests (RFs) using different hyperparameters: the number of trees (100, 500, 750, 1000, 2000), the number of features that could be used to split each node (fifty randomly generated values between 1 and 100), and the minimal node size (1,2,3,…,10). These hyperparameters were modified in each run through a grid approach. In each run, the mean decrease in the Gini index was used to determine the importance of each antibody in predicting the disease status of the study participants. The importance values for each antibody were averaged across the 2500 runs and then sorted from the most to the least important antibody for disease prediction.
We then created classifiers of disease status including an increasing number of the most important antibodies: Elastic-Net Logistic Regression (GLMNet), Random Forest (RF), Support Vector Machine (SVM), Linear Discriminant Analysis (LDA) and Extreme Gradient Boosting (XGBoost). For a given number of antibodies, the results from these classifiers were combined by a Super Learner (SL) algorithm that assigns different weights to each classifier estimated to the same data [29]. The performance of the resulting classifier was evaluated by accuracy (the proportion of individuals correctly classified) using the ROC01 criterion [30].
We started our SL-based analysis with the two most important antibodies as the respective features. Every time data from a new antibody response was added to the SL-based classifier (and its subclassifiers), we calculated the Spearman’s correlation coefficient R and remove highly correlated antibody responses (R > |0.8|), as done elsewhere [31]. We kept adding new antibody responses until we reached a maximum number of 100 antibody responses. The best classifier was the SL-based classifier with the lowest antibodies reaching a target accuracy of at least 85% in both the train and test sets; this accuracy is argued to be optimal for classification problems [32].
The above analysis was done to compare the cohorts of all ME/CFS patients, ME/CFS patients with reported infectious disease trigger, ME/CFS patients with non-infectious or unknown disease trigger against HCs.
2.4. Bioinformatic analysis
When we could find a SL-based classifier with the target accuracy in both train and test datasets, we then performed protein-protein alignments between the EBV peptide associated with each selected antibody and human proteins using the RefSeq reference protein database [33], as available in the National Centre for Biotechnology Information (https://blast.ncbi.nlm.nih.gov/). The degree of alignment was based on the E-score statistic [34]. In our analysis, we focused on the maximal E-score between each EBV peptide under analysis and human proteins. We then calculated the Spearman’s correlation coefficient R and the respective 95% confidence interval between the importance of each selected antibody for disease prediction and the respective maximal E-score of the peptide associated with that antibody. We finally performed a hypothesis test for the significance of this correlation coefficient using a significance level of 5%. The same analysis was repeated using the RefSeq non-redundant (nr) protein database.
2.5. Statistical Software
The statistical analyses were performed in the R software version 4.3.0 using the following packages: “caret” for multicollinearity analysis [35], “OptimalCutpoints” to obtain the accuracy based on the ROC01 criterion of each predictive model [30], “pROC” for the AUC estimation [36], “ranger” to perform Random-Forest [37], “SuperLearner” for the SL-based analysis [38].
3. Results
3.1. Analysis of all ME/CFS patients against HCs
The comparison between all the 92 patients with ME/CFS and 50 HCs was carried out to develop a classifier that could predict the disease status of these study participants. The train dataset was composed of forty-five HCs and 83 ME/CFS patients, while the test dataset comprised 5 HCs and 9 ME/CFS patients.
The overall average antibody importance distribution is presented in Figure 2A as a density plot. Our results showed that the overall average antibody importance was around 0.018 with the antibody against EBNA6_0066 being the most important one (importance = 0.36) (Figure 1A). Furthermore, 7 out of the 10 topmost antibodies are associated with peptides belonging to the family of Epstein-Bar nuclear antigen (EBNA) proteins (Figure 1A).
It is worth noting that antibodies against EBNA1_430 that contains a peptide candidate for a molecular mimicry with the human Anoctamin-2 protein [39,40] were similar in both patients with ME/CFS and HCs (Supplementary Figure 2). As a consequence, they only had an average importance of 0.010 (ranked in the 1866th place of the most important antibodies). Therefore, this finding suggested a negligible role of these antibodies in predicting disease status.
In the train dataset, the target accuracy of 85% was already achieved by an SL classifier including only two antibodies (Figure 1B). As a consequence, the underlying sensitivity and specificity were close or equal to 1 (data not shown). This result was explained by the high accuracy of the RF as a function of the number of antibodies included in the assembled models (Figure 1C).
In the test dataset, the accuracy estimates fluctuated around 0.5 and they were at best 64% using an SL classifier including 36 antibodies as features. This poor performance was largely explained by a low sensitivity of the classifiers (Figure 1D). Hence, the target accuracy of 85% was not achieved for the overall dataset, largely, due to a poor performance in predicting ME/CFS patients in the test dataset.
3.2. Analysis of ME/CFS patients with non-infectious or unknown disease trigger against HCs
We then compared the 38 ME/CFS patients with non-infected or with unknown disease trigger to the 50 HCs. This time the overall average antibody importance was around 0.012. The most important antibodies were EBNA1_0595, EBNA1_0530 and EBNA6_0400 with mean importance of 0.13, 0.12 and 0.11, respectively (Figure 2A). Once more, 7 out of the 10 topmost important antibodies recognized antigens from the EBNA protein group. In line with the analysis based on the whole cohort of ME/CFS, the antibodies against EBNA1_430 had an average importance of 0.005 that translated to a poor importance ranking (2384th place) among all the antibodies.
In the train dataset, an SL classifier based on the top 3 antibodies (EBNA1_0595 EBNA1_0530, and EBNA6_0400) reached an accuracy of 94%, a value higher than the target accuracy of 85% (Figure 3B). In contrast, the same classifier reached only 78% in the test dataset (Figure 2B). This value was the best accuracy that could be derived by not only this classifier, but also other SL classifiers including a higher number of antibodies.
In the test dataset, the target accuracy of the top 3 antibodies SL classifiers was not achieved by a relatively modest sensitivity (Figure 2C). Hence, this analysis suggested that these EBV antibodies were unable to discriminate this subset of ME/CFS patients from HC with high sensitivity and high specificity.
3.3. Analysis of ME/CFS patients with a putative infectious disease trigger against HCs
We conducted a final comparison between the 54 ME/CFS patients who reported an infection at their disease onset and the 50 HCs. This time the antibodies recognizing EBNA4_0529, EBNA3_0139, and EBNA3_0577 antigens had the highest importance values in the RF (0.26, 0.25, and 0.24, respectively; Figure 3A). Eight of the 10 topmost important antibodies belonged to the group of EBNA proteins. Once again, the antibodies against EBNA1_430 had a low average importance (0.009) and a poor ranking (1447th place) in terms of predictive importance.
In this analysis, we found an SL classifier including 26 antibodies that could reach an accuracy above the target value of 85% in both train and test datasets (99% and 90%, respectively; Figure 3B). These antibodies were associated with antigens derived from 9 different EBV proteins: BALF2 (n=2), BALF5 (n=3), BLLF1 (n=1), BMRF1 (n=1), EBNA1 (n=2), EBNA3 (n=4), EBNA4 (n=4), EBNA6 (n=7) and LMP2 (n=2). Twenty-two out of the 26 selected antibodies had increased levels in this subset of ME/CFS patients compared to HCs (Figure 3C). The estimated classifier had a sensitivity of 100% and a specificity of 80% (Figure 3D). The corresponding AUC for both the train and test datasets were 1.00 and 0.88, respectively (Figure 3E).
Note that there was an SL classifier including 42 antibodies that predicted disease status almost perfectly in both train and test datasets (Figure 4B). However, this perfect classification was achieved at the cost of ~2.5 antibodies per study participant.
3.4. Bioinformatic analysis
The final analysis aimed at testing the hypothesis whether the EBV peptides associated with the above 26 antibodies selected in the SL classifier could explain ME/CFS pathology via a mechanism of molecular mimicry. Under this hypothesis, we expected a positive correlation between the importance of each antibody selected against the best alignment score between the associated peptides and human proteins.
The best alignment score per peptide varied from 22.7 (BALF5_0025) to 32.9 (EBNA6_0070) when conducting the protein alignments in the human RefSeq reference protein dataset. In the case of EBNA6_0070, the highest alignment scores were associated with alignments with Homeobox-9A (HOXA-91) and Adrenergic Receptor Alpha (ADRA1B) proteins (Figure 4A). The peptide with the second best alignment was EBNA6_0488. In this case, this peptide had an extensive sequence homology with the human proteins CCCTC-binding factor (CTCF) and Adipocyte enhancer-binding proteins (AEBP1) (Figure 4B).
When we analyzed the average importance of these 26 antibodies against the best alignment scores of the respective peptides, we found a slight negative association between these two quantities, but this association was no statistically significant (R =-0.161, 95% CI=(-0.473;0.185); Figure 4C). When we repeated the same analysis using the non-redundant protein data, we obtained the same lack of correlation (R =-0.018, 95% CI=(-0.355;0.330); Supplementary Figure 3). Hence, these data did not support the hypothesis that antibodies recognizing important EBV peptides for disease predictive had a high potential for cross-reactivity with human proteins.
4. Discussion
This paper demonstrated the difficulty of finding anti-EBV antibodies that could be used as general markers of ME/CFS. Such a difficulty was clear when analyzing the whole cohort of ME/CFS patients due to the subset of patients who did not know their disease trigger or report a non-infectious disease trigger. Similar difficulty was obtained in studies on IgG antibodies against common pathogens [41] or multiple peptides derived from different herpesviruses [19].
In the present work, the best-case scenario was obtained for the patients with an infectious disease trigger where a set of 26 EBV-related antibodies led to a good accuracy in both train and test datasets. This finding on this subgroup of ME/CFS patients is not surprising given that our previous study also led to a similar conclusion using a different statistical methodology [27]. Overall, the present study reinforces the idea that patients with ME/CFS are very heterogenous as a clinical group and, as such, the quest of discovering a generic disease marker applicable to all patients is likely to fail. However, this problem can be surpassed by using an appropriate stratification (e.g., based on the hypothetical disease trigger) where specific disease biomarkers can emerge, as illustrated in this work.
Six of the selected 26 antibodies were associated with EBNA1_0005, EBNA3_0577, EBNA4_0566, EBNA6_0025, EBNA6_0488 EBNA6_0752 peptides. Antibodies against these antigens were found elevated in patients with multiple sclerosis (MS) [42]. Such a result is another piece of evidence that some patients with ME/CFS and MS share not only symptoms [43,44], but also serological alterations possibly linked to EBV [45]. In this line of thought, one could argue that rituximab that depletes the B-cell population and used in MS patients could be beneficial for this specific cohort of ME/CFS patients [2]. Unfortunately, the only large-scale clinical trial on the use of this drug in ME/CFS patients led to disappointed results [46]. A likely explanation for this finding is not targeting the specific cohort of ME/CFS patients with an infectious disease trigger .
Among the peptides recognized by the selected antibodies, EBNA6_0070 had the highest sequence homology with a human protein. This antigen was already discovered and amply discussed in our previous study [27]. The replication of this finding using a different statistical approach provided further support for the hypothetical role of this peptide in ME/CFS. However, in contrast with our previous work, antibodies against this candidate antigen were not on the top most important antibodies for disease prediction. This result suggests that the potential pathological effect of this EBV antigen via molecular mimicry might not be as straightforward as our initial study suggested. To resolve this question, one could perform functional studies where purified antibodies against this EBV peptide, when transferred to a recipient animal model, could or not elicit ME/CFS-like disease.
The EBNA6_0488 peptide had the second highest sequence homology with human proteins. This homology was related to two possible human 10-mer peptides belonging to CTCF and AEBP1. The former protein is a master transcription factor due to its more than 50,000 possible binding sites and its role as a chromatin barrier element [47,48]. Besides that, the level of this transcription factor is inversely correlated with the levels of DNA methylation [48]. CTCF in partnership with cohesion molecules is also important in many immunological pathways, such as interferon gamma production in Th1 cells and establishment and maintenance of regulatory T cells in visceral adipose tissue and skeletal muscle [49,50]. In this scenario, we speculate that the increased quantity of antibodies against EBNA6_0488 results in a cross-reactive antibody response to the CTCF peptide, thus, reducing the abundance of this transcription factor. This putative reduction could lead to altered gene expression and DNA methylation patterns, and abnormal immunological processes, including the maintenance of deleterious autoimmunity in check. This speculation is in line with findings from altered gene expression and DNA methylation profiles in ME/CFS patients [51,52,53,54]. As an extreme case, one study identified more than 12,000 CpG sites with altered DNA methylation levels in patients with ME/CFS compared to HCs [55]. Immunological abnormalities are also reported by many studies in ME/CFS patients (reviewed in ref. [7,56]). An alternative interpretation is that the increased levels of antibodies against EBNA6_0488 resulted from a putative CTCF overexpression during the disease progression in the cohort of ME/CFS patients with an infectious disease trigger. An overexpression of this transcription factor could be the result of a stress-induced response to restore homeostatic equilibrium within cells. However, altered gene expression was not reported for CTCF by any gene expression studies published so far. This negative reporting could be explained by not performing any patient’s stratification when analyzing data from these studies.
With respect to AEBP1, this protein is a ubiquitous transcriptional repressor involved in the regulation of adipogenesis, mammary gland development, inflammation, macrophage cholesterol homeostasis, and atherogenesis [57]. Interestingly, mutations on AEBP1-encoding gene were implicated with the onset of Ehlers-Danlos syndrome (EDS) [58,59]. Patients with EDS hypermobility-type can also receive a diagnosis of ME/CFS [60]. On the other hand, patients with a diagnosis of ME/CFS also show EDS as a co-morbidity [61]. In fact, the presence of EDS in a suspected case of ME/CFS has not been considered as an exclusionary condition for the respective disease diagnosis [62]. However, genome-wide association studies of ME/CFS did not report any genetic markers located on the AEBP1 gene [63,64,65,66,67]. In this scenario, antibody responses to EBNA6_0488 with the potential of being cross-reactive with AEBP1 should alter the regulation of biological processes where this protein is involved. In particular, the deficient regulation of inflammatory processes is particularly relevant for ME/CFS given the general idea that established ME/CFS translates into a persistent low-grade inflammatory process in patients [5]. Given that endothelial disfunction is also observed in patients with ME/CFS [68,69,70], such a disfunction could result from damaged endothelial cells via persistent low-grade inflammation in response to EBNA6_0488 mimicking an AEBP1 peptide. Hence, the identification of this molecular mimicry brings an unexpected link between EBV and AEBP1. As alluded above for CTCF, current gene expression studies did not highlight AEBP1 at the top of the proteins with the most significant differential abundance between patients with ME/CFS and HCs. The lack of patient’s stratification is once again a possible reason to not detect an altered abundance of AEBP1 in ME/CFS patients when compared to HCs.
Interestingly, the maximum sequence homology of the peptides recognized by the 26 selected antibodies and human proteins was not associated with the importance of the same antibodies in disease prediction. Moreover, antibodies against EBNA1_430 that contains a peptide mimicking a peptide from the human Anoctamin-2 protein [39,40] had a low importance in predicting ME/CFS patients. These results suggest that eventual molecular mimicries due to antibody reactivity between EBV and human antigens has a minor role in the underlying pathological mechanism in such a subset of patients. However, we cannot rule out the possibility of an eventual mimicry based on three-dimensional molecular structure of the respective peptides, but not at the level of their amino-acid sequence. We cannot also rule out that molecular mimicries based on sequence homology might be elicited by peptides from EBV proteins other than the ones evaluated in this study. This might be the case of two EBV peptides from BPLF1 and BHRF1 proteins that were able to elicit an immune response by self-reactive T cell-clones derived from patients with MS [71].
An interesting perspective of the above results can be given by the so-called danger theory [72]. The theory is based on the premise that the immune system is activated by danger or damage signals sent by infected (or stressed) cells to the immune system. These danger signals are independent of the intrinsic nature of antigens (self or non-self) seen by the immune system. As a corollary, autoimmune responses and autoimmune diseases are then understood as unintended consequences of persistent danger signals that ultimately include chronic presentation of multiple self and nonself antigens. This explains why chronic and low-grade infections by herpesviruses are among the most documented triggers of autoimmune diseases. In this scenario, the theory predicts exactly the lack of correlation between importance of the selected antibodies in predicting ME/CFS and the degree of molecular mimicry between the EBV peptides and human proteins.
The basic question of applying the danger theory to ME/CFS pathogenesis lies upon understanding which danger signals are at the core of the disease. According to the original proponents of the danger theory, general danger signals are the heat shock proteins (HSP), the vasoactive intestinal polypeptide (VIP), the cytokines TNFα and IL1β, among others [73]. A brief discussion about some of these danger signals in the context of ME/CFS is given below; a more comprehensive discussion of this topic will be done in a near future.
HSPs are highly conservative proteins in nature and they are produced in response to many different cellular stresses. In theory, antigens derived from these proteins were thought to belong to the so-called immunological homunculus, a limited set of dominant self antigens that allow the immune system to have picture of the self [74]. However, there is no consensus whether HSPs are indeed signaling danger or simply key regulatory and resolution elements of a stress or immune response [72,75]. This alternative interpretation of the functional role of HSPs might explain the lack of consistency in HSP-related responses across studies where patients with ME/CFS and HCs were challenged with physical exercise [76,77,78]. In addition, antibodies against endogenous and microbial HSP65 peptides were at the same level in patients with ME/CFS and HCs with the exception of a higher seroprevalence to a HSP65 peptide derived from Chlamydia pneumoniae in the former [79].
VIP is a neuromodulator present in the gut and the anterior chamber of the eye [73]. On the one hand, it has the capacity of activating dendritic cells [80] (thus, its suggestion as a possible danger signal). On the other hand, the binding of VIP to its receptor in immune cells also leads to anti-inflammatory actions [81]. In this line of thought, a loss of tolerance to VIP, other vasoactive neuropeptides or their receptors were hypothesized to be at the genesis of ME/CFS [82]. However, a follow-up study showed an elevated expression of VPACR2 – the VIP receptor – in immune cells and an increased frequency of the regulatory Foxp3+CD4+ T cells in ME/CFS patients in comparison to HCs [83]. Given the capacity of VIP inducing the generation of these regulatory cells [84], this finding is more in line with VIP as a regulation mediator in the context of ME/CFS.
TNFα and IL1β are the two classical pro-inflammatory cytokines. According to a systematic review [85], 20%-25% of the studies reported elevated levels of these cytokines in patients with ME/CFS when compared to HCs. However, the same systematic review did not perform a meta-analysis of the published data. Therefore, it is unclear whether the lack of significant findings related to these two cytokines is a consequence of insufficient statistical power due to reduced sample sizes used in the respective studies. It is worth noting that patients with ME/CFS from Italy had a higher frequency of an allele variant associated with elevated levels of TNFα (rs1800629:G>A) [86]. However, this finding was not replicated by another study with German patients [87].
In summary, this study provided a list of possible EBV peptides whose associated IgG antibody responses could be used in the diagnosis of suspected ME/CFS cases who reported an infection at their symptom’s onset. Two of these peptides had high sequence homologies with human proteins, but the corresponding antibody responses were not the most important ones for disease prediction. This finding suggested that the role of EBV on eventual ME/CFS-related autoimmunity should be reconsidered under alternative theories about the self-nonself discrimination problem.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
NS designed this research; AF conceptualized and performed all the computational implementation of the methodology; MS and TL performed preliminary analyses concerning the molecular mimicry between EBNA1 and Anoctamin-2; AF conducted the analysis; AF, CC, and NS interpreted the data; AF, CC and NS have participated in the original draft preparation, writing, reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
André Fonseca received funding from FCT – Fundação para a Ciência e Tecnologia, Portugal (refs. SFRH/BD/147629/2019 and UIDB/00006/2020). Clara Cordeiro and Nuno Sepúlveda received partial funding from FCT – Fundação para a Ciência e Tecnologia, Portugal (ref. UIDB/00006/2020).
Data Availability Statement
The dataset is freely available as Supplementary Material of Loebel et al [26]. The R scripts are available without restriction in the following address: https://github.com/Publications/Fonseca_etal.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rivera MC, Mastronardi C, Silva-Aldana CT, Arcos-Burgos M, Lidbury BA. Myalgic encephalomyelitis/chronic fatigue syndrome: A comprehensive review. Diagnostics 2019;9:91. [CrossRef]
- Fluge Ø, Tronstad KJ, Mella O. Pathomechanisms and possible interventions in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS). J Clin Invest 2021;131:e150377. [CrossRef]
- Stanculescu D, Larsson L, Bergquist J. Hypothesis: Mechanisms That Prevent Recovery in Prolonged ICU Patients Also Underlie Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Front Med 2021;8:628029. [CrossRef]
- Stanculescu D, Sepúlveda N, Lim CL, Bergquist J. Lessons From Heat Stroke for Understanding Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front Neurol 2021;12. [CrossRef]
- Nacul L, O’Boyle S, Palla L, Nacul FE, Mudie K, Kingdon CC, et al. How Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS) Progresses: The Natural History of ME/CFS. Front Neurol 2020;11:826. [CrossRef]
- Blomberg J, Gottfries CG, Elfaitouri A, Rizwan M, Rosén A. Infection elicited autoimmunity and Myalgic encephalomyelitis/chronic fatigue syndrome: An explanatory model. Front Immunol 2018;9:229. [CrossRef]
- Sotzny F, Blanco J, Capelli E, Castro-Marrero J, Steiner S, Murovska M, et al. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome – Evidence for an autoimmune disease. Autoimmun Rev 2018;17:601–9. [CrossRef]
- Morris G, Berk M, Galecki P, Maes M. The emerging role of autoimmunity in myalgic encephalomyelitis/chronic fatigue syndrome (ME/cfs). Mol Neurobiol 2014;49:741–56. [CrossRef]
- Petracek LS, Suskauer SJ, Vickers RF, Patel NR, Violand RL, Swope RL, et al. Adolescent and Young Adult ME/CFS After Confirmed or Probable COVID-19. Front Med 2021;8:668944. [CrossRef]
- Davis HE, Assaf GS, McCorkell L, Wei H, Low RJ, Re’em Y, et al. Characterizing long COVID in an international cohort: 7 months of symptoms and their impact. EClinicalMedicine 2021;38. [CrossRef]
- Jason LA, Dorri JA. ME/CFS and Post-Exertional Malaise among Patients with Long COVID. Neurol Int 2022;15:1–11. [CrossRef]
- Vélez-Santamaría R, Fernández-Solana J, Méndez-López F, Domínguez-García M, González-Bernal JJ, Magallón-Botaya R, et al. Functionality, physical activity, fatigue and quality of life in patients with acute COVID-19 and Long COVID infection. Sci Reports 2023 131 2023;13:1–9. [CrossRef]
- Ruiz-Pablos M, Paiva B, Montero-Mateo R, Garcia N, Zabaleta A. Epstein-Barr Virus and the Origin of Myalgic Encephalomyelitis or Chronic Fatigue Syndrome. Front Immunol 2021;12:656797. [CrossRef]
- Ariza, ME. Myalgic encephalomyelitis/chronic fatigue syndrome: The human herpesviruses are back! Biomolecules 2021;11:1–17. [CrossRef]
- Sepúlveda N, Carneiro J, Lacerda E, Nacul L. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome as a Hyper-Regulated Immune System Driven by an Interplay Between Regulatory T Cells and Chronic Human Herpesvirus Infections. Front Immunol 2019;10:2684. [CrossRef]
- Ruiz-Pablos M, Paiva B, Zabaleta A. Epstein–Barr virus-acquired immunodeficiency in myalgic encephalomyelitis—Is it present in long COVID? J Transl Med 2023;21:633. [CrossRef]
- Capone G, Calabrò M, Lucchese G, Fasano C, Girardi B, Polimeno L, et al. Peptide matching between Epstein-Barr virus and human proteins. Pathog Dis 2013;69:205–12. [CrossRef]
- Baboonian C, Venables PJW, Williams DG, Williams RO, Maini RN. Cross reaction of antibodies to a glycine/alanine repeat sequence of Epstein-Barr virus nuclear antigen-i with collagen, cytokeratin, and actin. Ann Rheum Dis 1991;50:772–5. [CrossRef]
- Blomberg J, Rizwan M, Böhlin-Wiener A, Elfaitouri A, Julin P, Zachrisson O, et al. Antibodies to human herpesviruses in myalgic encephalomyelitis/chronic fatigue syndrome patients. Front Immunol 2019;10:1946. [CrossRef]
- Lerner AM, Ariza ME, Williams M, Jason L, Beqaj S, Fitzgerald JT, et al. Antibody to Epstein-Barr virus deoxyuridine triphosphate nucleotidohydrolase and deoxyribonucleotide polymerase in a chronic fatigue syndrome subset. PLoS One 2012;7:e47891. [CrossRef]
- Kerr, JR. Epstein-Barr Virus Induced Gene-2 Upregulation Identifies a Particular Subtype of Chronic Fatigue Syndrome/Myalgic Encephalomyelitis. Front Pediatr 2019;7:59. [CrossRef]
- Domingues TD, Grabowska AD, Lee JS, Ameijeiras-Alonso J, Westermeier F, Scheibenbogen C, et al. Herpesviruses Serology Distinguishes Different Subgroups of Patients From the United Kingdom Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Biobank. Front Med 2021;8:686736. [CrossRef]
- Malato J, Graça L, Sepúlveda N. Impact of Misdiagnosis in Case-Control Studies of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Diagnostics (Basel, Switzerland) 2023;13:531. [CrossRef]
- Nacul L, Lacerda EM, Kingdon CC, Curran H, Bowman EW. How have selection bias and disease misclassification undermined the validity of myalgic encephalomyelitis/chronic fatigue syndrome studies? J Health Psychol 2019;24:1765–9. [CrossRef]
- Ariza, ME. Commentary: Antibodies to Human Herpesviruses in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients. Front Immunol 2020;11:1945. [CrossRef]
- Loebel M, Eckey M, Sotzny F, Hahn E, Bauer S, Grabowski P, et al. Serological profiling of the EBV immune response in Chronic Fatigue Syndrome using a peptide microarray. PLoS One 2017;12:e0179124. [CrossRef]
- Sepúlveda N, Malato J, Sotzny F, Grabowska AD, Fonseca A, Cordeiro C, et al. Revisiting IgG Antibody Reactivity to Epstein-Barr Virus in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome and Its Potential Application to Disease Diagnosis. Front Med 2022;9:921101. [CrossRef]
- Roshan V, Stewart JHM, Joseph R, Stewart HM. Optimal ratio for data splitting. Stat Anal Data Min ASA Data Sci J 2022;15:531–8. [CrossRef]
- Van Der Laan MJ, Polley EC, Hubbard AE. Super learner. Stat Appl Genet Mol Biol 2007;6:25. [CrossRef]
- López-Ratón M, Rodríguez-Álvarez MX, Cadarso-Suárez C, Gude-Sampedro F. Optimalcutpoints: An R package for selecting optimal cutpoints in diagnostic tests. J Stat Softw 2014;61:1–36. [CrossRef]
- Valletta JJ, Recker M. Identification of immune signatures predictive of clinical protection from malaria. PLOS Comput Biol 2017;13:e1005812. [CrossRef]
- Wilson RC, Shenhav A, Straccia M, Cohen JD. The Eighty Five Percent Rule for optimal learning. Nat Commun 2019;10. [CrossRef]
- O’Leary NA, Wright MW, Brister JR, Ciufo S, Haddad D, McVeigh R, et al. Reference sequence (RefSeq) database at NCBI: current status, taxonomic expansion, and functional annotation. Nucleic Acids Res 2016;44:D733–45. [CrossRef]
- Karlin S, Altschul SF. Applications and statistics for multiple high-scoring segments in molecular sequences. Proc Natl Acad Sci U S A 1993;90:5873–7. [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J Stat Softw 2008;28:1–26. [CrossRef]
- Robin X, Turck N, Hainard A, Tiberti N, Lisacek F, Sanchez JC, et al. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinformatics 2011;12:1–8. [CrossRef]
- Wright MN, Ziegler A. ranger: A Fast Implementation of Random Forests for High Dimensional Data in C++ and R. J Stat Softw 2017;77:1–17. [CrossRef]
- Polley E, LeDell E, Kennedy C, van der Laan M. SuperLearner: Super Learner Prediction 2021.
- Tengvall K, Huang J, Hellström C, Kammer P, Biström M, Ayoglu B, et al. Molecular mimicry between Anoctamin 2 and Epstein-Barr virus nuclear antigen 1 associates with multiple sclerosis risk. Proc Natl Acad Sci U S A 2019;116:16955–60. [CrossRef]
- Sepúlveda, N. Impact of genetic variation on the molecular mimicry between Anoctamin-2 and Epstein-Barr virus nuclear antigen 1 in Multiple Sclerosis. Immunol Lett 2021;238:29–31. [CrossRef]
- O’Neal AJ, Glass KA, Emig CJ, Vitug AA, Henry SJ, Shungu DC, et al. Survey of Anti-Pathogen Antibody Levels in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Proteomes 2022, Vol 10, Page 21 2022;10:21. [CrossRef]
- Bjornevik K, Cortese M, Healy BC, Kuhle J, Mina MJ, Leng Y, et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Science (80- ) 2022;375:296–301. [CrossRef]
- Jason L, Ohanian D, Brown A, Sunnquist M, McManimen S, Klebek L, et al. Differentiating Multiple Sclerosis from Myalgic Encephalomyelitis and Chronic Fatigue Syndrome. Insights Biomed 2017;2:11. [CrossRef]
- Ohanian D, Brown A, Sunnquist M, Furst J, Nicholson L, Klebek L, et al. Identifying Key Symptoms Differentiating Myalgic Encephalomyelitis and Chronic Fatigue Syndrome from Multiple Sclerosis. Neurol 2016;4:41.
- Domingues TD, Malato J, Grabowska AD, Lee JS, Ameijeiras-Alonso J, Biecek P, et al. Association analysis between symptomology and herpesvirus IgG antibody concentrations in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) and multiple sclerosis. Heliyon 2023;9. [CrossRef]
- Fluge Ø, Rekeland IG, Lien K, Thürmer H, Borchgrevink PC, Schäfer C, et al. B-Lymphocyte Depletion in Patients With Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Ann Intern Med 2019;170:585–93. [CrossRef]
- Bastiaan Holwerda SJ, de Laat W. CTCF: the protein, the binding partners, the binding sites and their chromatin loops. Philos Trans R Soc B Biol Sci 2013;368. [CrossRef]
- Dehingia B, Milewska M, Janowski M, Pe Z Kowska A. CTCF shapes chromatin structure and gene expression in health and disease. EMBO Rep 2022;23:e55146. [CrossRef]
- DiSpirito JR, Zemmour D, Ramanan D, Cho J, Zilionis R, Klein AM, et al. Molecular diversification of regulatory T cells in nonlymphoid tissues. Sci Immunol 2018;3:5861. [CrossRef]
- Chandrasekaran V, Oparina N, Garcia-Bonete MJ, Wasén C, Erlandsson MC, Malmhäll-Bah E, et al. Cohesin-Mediated Chromatin Interactions and Autoimmunity. Front Immunol 2022;13:840002. [CrossRef]
- Kerr, JR. Gene profiling of patients with chronic fatigue syndrome/myalgic encephalomyelitis. Curr Rheumatol Rep 2008;10:482–91. [CrossRef]
- Kaushik N, Fear D, Richards SCM, McDermott CR, Nuwaysir EF, Kellam P, et al. Gene expression in peripheral blood mononuclear cells from patients with chronic fatigue syndrome. J Clin Pathol 2005;58:826–32. [CrossRef]
- Sweetman E, Ryan M, Edgar C, Mackay A, Vallings R, Tate W. Changes in the transcriptome of circulating immune cells of a New Zealand cohort with myalgic encephalomyelitis/chronic fatigue syndrome. Int J Immunopathol Pharmacol 2019;33:1–8. [CrossRef]
- Almenar-Pérez E, Ovejero T, Sánchez-Fito T, Espejo JA, Nathanson L, Oltra E. Epigenetic Components of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Uncover Potential Transposable Element Activation. Clin Ther 2019;41:675–98. [CrossRef]
- De Vega WC, Herrera S, Vernon SD, McGowan PO. Epigenetic modifications and glucocorticoid sensitivity in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). BMC Med Genomics 2017;10:11. [CrossRef]
- Lorusso L, Mikhaylova S V., Capelli E, Ferrari D, Ngonga GK, Ricevuti G. Immunological aspects of chronic fatigue syndrome. Autoimmun Rev 2009;8:287–91. [CrossRef]
- Majdalawieh AF, Massri M, Ro HS. AEBP1 is a Novel Oncogene: Mechanisms of Action and Signaling Pathways. J Oncol 2020;2020. [CrossRef]
- Angwin C, Ghali N, van Dijk FS. Case report: Two individuals with AEBP1-related classical-like EDS: Further clinical characterisation and description of novel AEBP1 variants. Front Genet 2023;14. [CrossRef]
- Ritelli M, Cinquina V, Venturini M, Pezzaioli L, Formenti AM, Chiarelli N, et al. Expanding the Clinical and Mutational Spectrum of Recessive AEBP1-Related Classical-Like Ehlers-Danlos Syndrome. Genes (Basel) 2019;10. [CrossRef]
- Castori M, Celletti C, Camerota F, Grammatico P. Chronic fatigue syndrome is commonly diagnosed in patients with Ehlers-Danlos syndrome hypermobility type/joint hypermobility syndrome. Clin Exp Rheumatol 2011;29:597–8.
- Hakim A, De Wandele I, O’Callaghan C, Pocinki A, Rowe P. Chronic fatigue in Ehlers-Danlos syndrome-Hypermobile type. Am J Med Genet C Semin Med Genet 2017;175:175–80. [CrossRef]
- Nacul L, Authier FJ, Scheibenbogen C, Lorusso L, Helland IB, Martin JA, et al. European Network on Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (EUROMENE): Expert Consensus on the Diagnosis, Service Provision, and Care of People with ME/CFS in Europe. Medicina (Kaunas) 2021;57:510. [CrossRef]
- Schlauch KA, Khaiboullina SF, De Meirleir KL, Rawat S, Petereit J, Rizvanov AA, et al. Genome-wide association analysis identifies genetic variations in subjects with myalgic encephalomyelitis/chronic fatigue syndrome. Transl Psychiatry 2016;6:e730. [CrossRef]
- Dibble JJ, McGrath SJ, Ponting CP. Genetic risk factors of ME/CFS: A critical review. Hum Mol Genet 2020;29:R118–25. [CrossRef]
- Herrera S, de Vega WC, Ashbrook D, Vernon SD, McGowan PO. Genome-epigenome interactions associated with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Epigenetics 2018;13:1174–90. [CrossRef]
- Hajdarevic R, Lande A, Mehlsen J, Rydland A, Sosa DD, Strand EB, et al. Genetic association study in myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) identifies several potential risk loci. Brain Behav Immun 2022;102:362–9. [CrossRef]
- Das S, Taylor K, Kozubek J, Sardell J, Gardner S. Genetic risk factors for ME/CFS identified using combinatorial analysis. J Transl Med 2022;20. [CrossRef]
- Haffke M, Freitag H, Rudolf G, Seifert M, Doehner W, Scherbakov N, et al. Endothelial dysfunction and altered endothelial biomarkers in patients with post-COVID-19 syndrome and chronic fatigue syndrome (ME/CFS). J Transl Med 2022;20. [CrossRef]
- Scherbakov N, Szklarski M, Hartwig J, Sotzny F, Lorenz S, Meyer A, et al. Peripheral endothelial dysfunction in myalgic encephalomyelitis/chronic fatigue syndrome. ESC Hear Fail 2020;7:1064–71. [CrossRef]
- Blauensteiner J, Bertinat R, León LE, Riederer M, Sepúlveda N, Westermeier F. Altered endothelial dysfunction-related miRs in plasma from ME/CFS patients. Sci Rep 2021;11:10604. [CrossRef]
- Wang J, Jelcic I, Mühlenbruch L, Haunerdinger V, Toussaint NC, Zhao Y, et al. HLA-DR15 Molecules Jointly Shape an Autoreactive T Cell Repertoire in Multiple Sclerosis. Cell 2020;183:1264-1281.e20. [CrossRef]
- Pradeu T, Cooper EL. The danger theory: 20 years later. Front Immunol 2012;3. [CrossRef]
- Gallucci S, Matzinger P. Danger signals: SOS to the immune system. Curr Opin Immunol 2001;13:114–9. [CrossRef]
- Cohen IR, Young DB. Autoimmunity, microbial immunity and the immunological homunculus. Immunol Today 1991;12:105–10. [CrossRef]
- Quintana FJ, Mimran A, Carmi P, Mor F, Cohen IR. HSP60 as a target of anti-ergotypic regulatory T cells. PLoS One 2008;3. [CrossRef]
- Jammes Y, Steinberg JG, Delliaux S. Chronic fatigue syndrome: acute infection and history of physical activity affect resting levels and response to exercise of plasma oxidant/antioxidant status and heat shock proteins. J Intern Med 2012;272:74–84. [CrossRef]
- Thambirajah AA, Sleigh K, Stiver HG, Chow AW. Differential heat shock protein responses to strenuous standardized exercise in chronic fatigue syndrome patients and matched healthy controls. Clin Investig Med 2008;31. [CrossRef]
- Clayton, EW. Beyond Myalgic Encephalomyelitis/Chronic Fatigue Syndrome: An IOM Report on Redefining an Illness. JAMA 2015;313:1101–2. [CrossRef]
- Elfaitouri A, Herrmann B, Bölin-Wiener A, Wang Y, Gottfries CG, Zachrisson O, et al. Epitopes of microbial and human heat shock protein 60 and their recognition in myalgic encephalomyelitis. PLoS One 2013;8. [CrossRef]
- Delneste Y, Herbault N, Galea B, Magistrelli G, Bazin I, Bonnefoy J-Y, et al. Vasoactive Intestinal Peptide Synergizes with TNF-α in Inducing Human Dendritic Cell Maturation. J Immunol 1999;163:3071–5. [CrossRef]
- Gomariz RP, Juarranz Y, Abad C, Arranz A, Leceta J, Martinez C. VIP-PACAP system in immunity: new insights for multitarget therapy. Ann N Y Acad Sci 2006;1070:51–74. [CrossRef]
- Staines, DR. Is chronic fatigue syndrome an autoimmune disorder of endogenous neuropeptides, exogenous infection and molecular mimicry? Med Hypotheses 2004;62:646–52. [CrossRef]
- Brenu EW, van Driel ML, Staines DR, Ashton KJ, Ramos SB, Keane J, et al. Immunological abnormalities as potential biomarkers in Chronic Fatigue Syndrome/Myalgic Encephalomyelitis. J Transl Med 2011;9. [CrossRef]
- Gonzalez-Rey E, Delgado M. Vasoactive intestinal peptide and regulatory T-cell induction: a new mechanism and therapeutic potential for immune homeostasis. Trends Mol Med 2007;13:241–51. [CrossRef]
- Blundell S, Ray KK, Buckland M, White PD. Chronic fatigue syndrome and circulating cytokines: A systematic review. Brain Behav Immun 2015;50:186–95. [CrossRef]
- Carlo-Stella N, Badulli C, De Silvestri A, Bazzichi L, Martinetti M, Lorusso L, et al. A first study of cytokine genomic polymorphisms in CFS: Positive association of TNF-857 and IFNγ874 rare alleles. Clin Exp Rheumatol 2006;24:179–82.
- Steiner S, Becker SC, Hartwig J, Sotzny F, Lorenz S, Bauer S, et al. Autoimmunity-Related Risk Variants in PTPN22 and CTLA4 Are Associated With ME/CFS With Infectious Onset. Front Immunol 2020;11:578. [CrossRef]
Figure 1.
Analysis of all ME/CFS patients versus HCs. (A) Density plot of the antibody’s average importance distribution obtained by the RF with the top 10 most important antibodies highlighted. (B) Accuracy of the SL classifier in the train (purple) and test (orange) datasets as a function of number of antibodies included. The black and blue horizontal dashed lines indicate the 85% (target accuracy). The best classifier is highlighted with a black dot. (C) Accuracy of the different classifiers assembled by the SL in the train dataset. (D) Sensitivity and specificity of the SL classifiers in the test dataset as a function of number of antibodies included.
Figure 1.
Analysis of all ME/CFS patients versus HCs. (A) Density plot of the antibody’s average importance distribution obtained by the RF with the top 10 most important antibodies highlighted. (B) Accuracy of the SL classifier in the train (purple) and test (orange) datasets as a function of number of antibodies included. The black and blue horizontal dashed lines indicate the 85% (target accuracy). The best classifier is highlighted with a black dot. (C) Accuracy of the different classifiers assembled by the SL in the train dataset. (D) Sensitivity and specificity of the SL classifiers in the test dataset as a function of number of antibodies included.
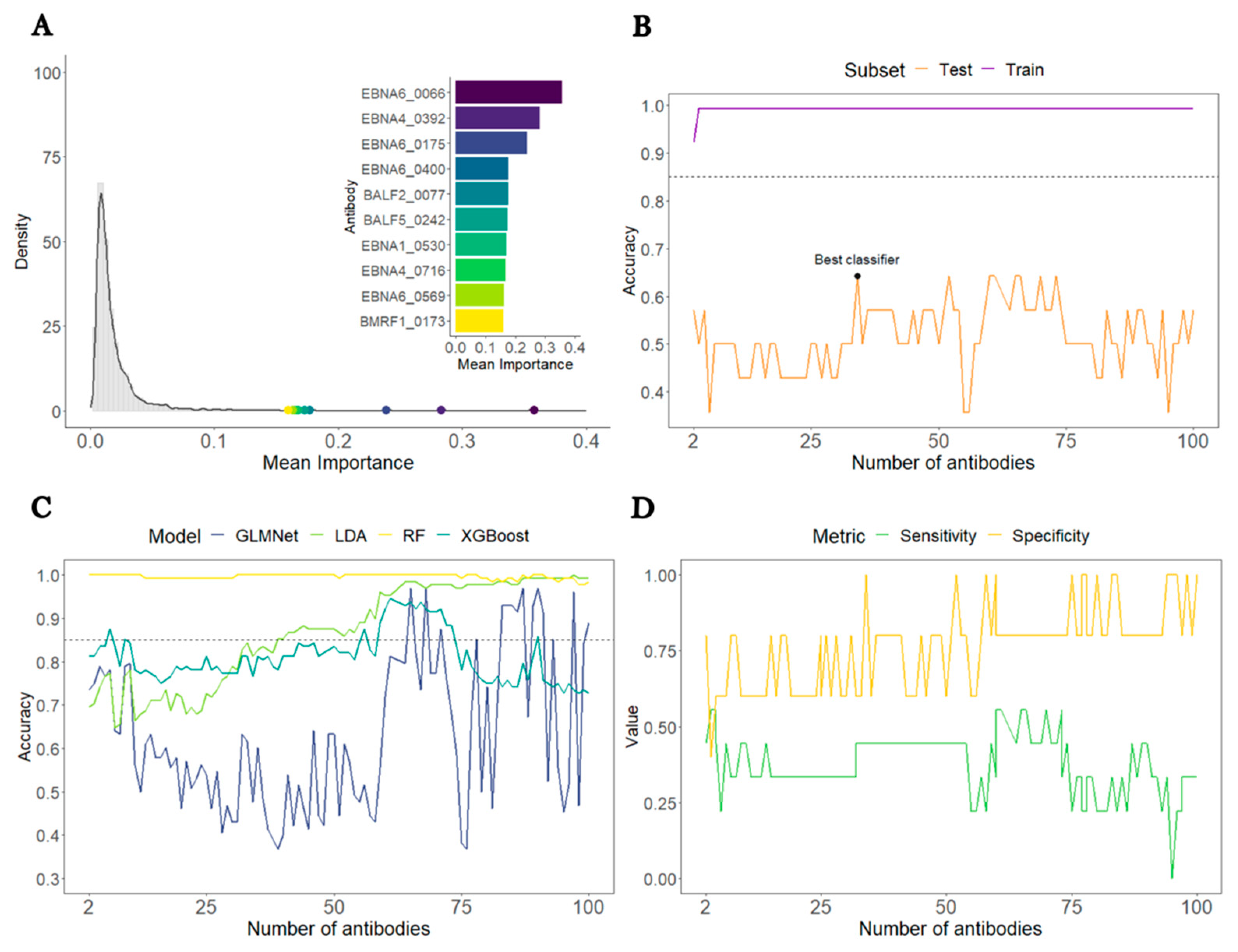
Figure 2.
Analysis of ME/CFS patients with non-infectious or unknown disease trigger against HCs. (A) Density plot of the antibody’s average importance distribution obtained by the RF with the top 10 most important antibodies highlighted. (B) Accuracy of the SL classifier in the train (purple) and test (orange) datasets as a function of number of antibodies included. The black and blue horizontal dashed lines indicate the 85% (target accuracy). The best classifier is highlighted with a black dot. (C) Sensitivity and specificity of the SL classifiers in the test dataset as a function of number of antibodies included. .
Figure 2.
Analysis of ME/CFS patients with non-infectious or unknown disease trigger against HCs. (A) Density plot of the antibody’s average importance distribution obtained by the RF with the top 10 most important antibodies highlighted. (B) Accuracy of the SL classifier in the train (purple) and test (orange) datasets as a function of number of antibodies included. The black and blue horizontal dashed lines indicate the 85% (target accuracy). The best classifier is highlighted with a black dot. (C) Sensitivity and specificity of the SL classifiers in the test dataset as a function of number of antibodies included. .
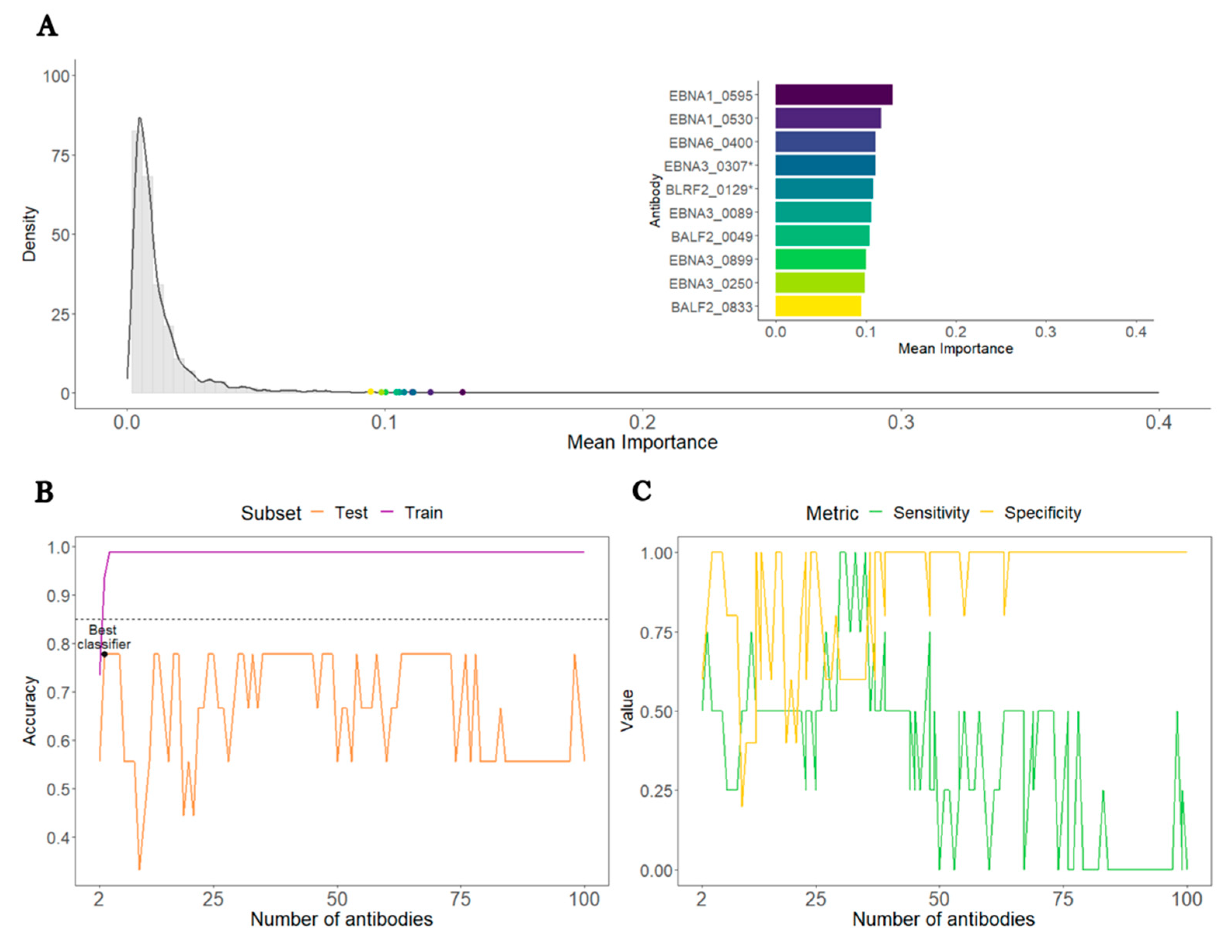
Figure 3.
Analysis of ME/CFS patients with an infectious disease trigger against HCs. (A) Density plot of the antibody’s average importance distribution obtained by the RF with the top 10 most important antibodies highlighted. (B) Accuracy of the SL classifier in the train and test datasets as a function of number of antibodies included. The black and blue horizontal dashed lines indicate the 85% (target accuracy). The best classifier is highlighted with a black dot. (C) Boxplots of the log10-levels of the selected 26 antibodies in HCs and ME/CFS patients. (D) Confusion matrix concerning the optimal classifier performance on the test set. (E) ROC curve of the optimal classifier for both train and test datasets.
Figure 3.
Analysis of ME/CFS patients with an infectious disease trigger against HCs. (A) Density plot of the antibody’s average importance distribution obtained by the RF with the top 10 most important antibodies highlighted. (B) Accuracy of the SL classifier in the train and test datasets as a function of number of antibodies included. The black and blue horizontal dashed lines indicate the 85% (target accuracy). The best classifier is highlighted with a black dot. (C) Boxplots of the log10-levels of the selected 26 antibodies in HCs and ME/CFS patients. (D) Confusion matrix concerning the optimal classifier performance on the test set. (E) ROC curve of the optimal classifier for both train and test datasets.
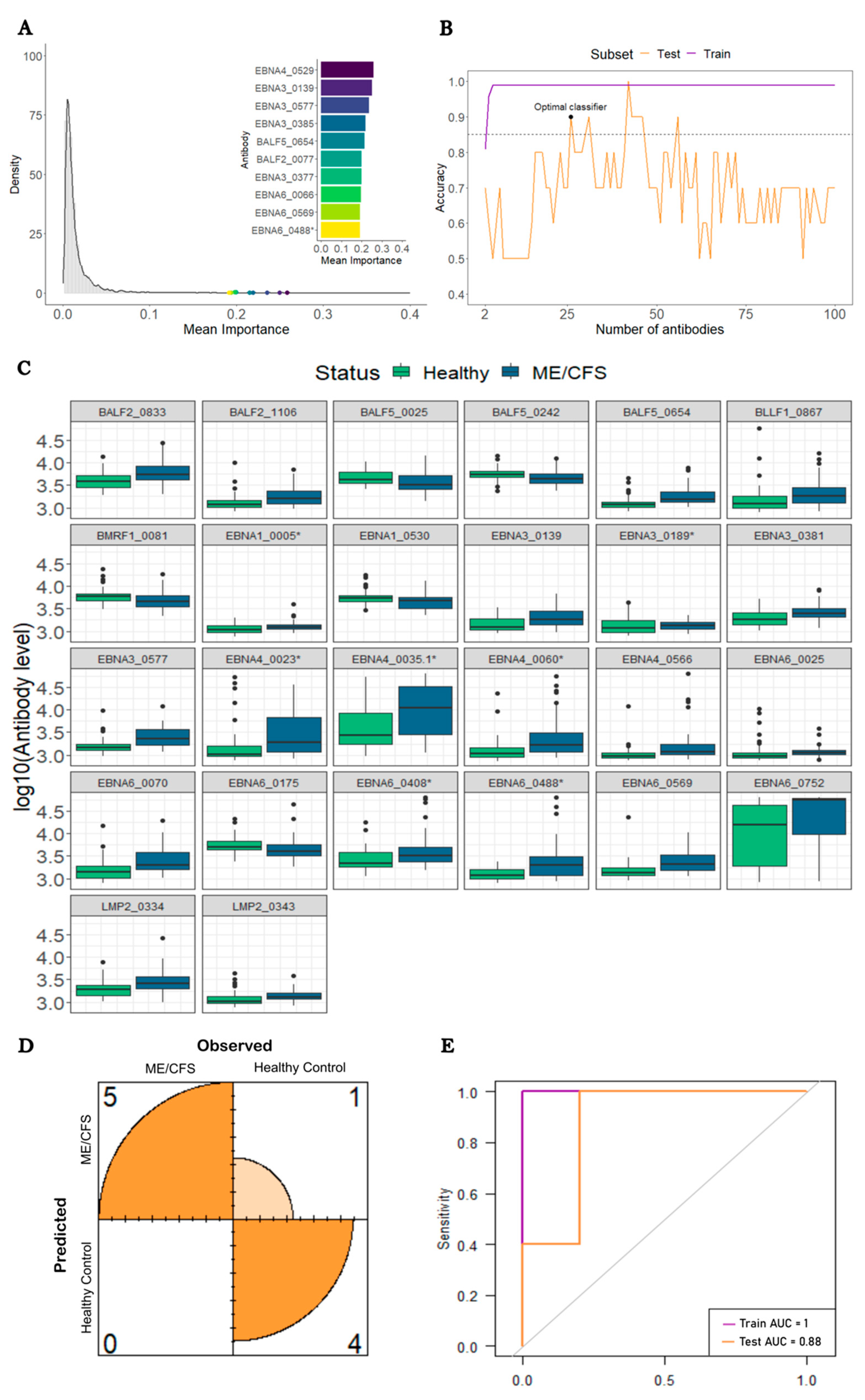
Figure 4.
Bioinformatic analysis of the EBV peptides associated with the 26 antibodies for predicting ME/CFS patients with an infectious disease trigger. A) Alignments between EBNA6_0070 and the human proteins HOXA-9A and ADRA1B_371 with the corresponding amino-acid coordinates within brackets. B) Alignments between EBNA6_0488* and the human proteins CTCF and AEBP1 with the corresponding protein coordinates within brackets. C). Scatterplot between the average importance of each EBV peptide and the maximum E-score alignment score with human proteins using the RefSeq reference protein database where R is the Spearman’s correlation coefficient with the respective 95% confidence interval in brackets.
Figure 4.
Bioinformatic analysis of the EBV peptides associated with the 26 antibodies for predicting ME/CFS patients with an infectious disease trigger. A) Alignments between EBNA6_0070 and the human proteins HOXA-9A and ADRA1B_371 with the corresponding amino-acid coordinates within brackets. B) Alignments between EBNA6_0488* and the human proteins CTCF and AEBP1 with the corresponding protein coordinates within brackets. C). Scatterplot between the average importance of each EBV peptide and the maximum E-score alignment score with human proteins using the RefSeq reference protein database where R is the Spearman’s correlation coefficient with the respective 95% confidence interval in brackets.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



