Submitted:
23 November 2023
Posted:
24 November 2023
You are already at the latest version
Abstract
As a result of climate change, the Mediterranean Sea has been exposed to an increase in frequency and intensity of marine heat waves in the last decades, some of which caused mass mortality events of benthic invertebrates, including sponges. Sponges are an important component of benthic ecosystems and can be the dominant group in some rocky shallow-water areas in the Mediterranean Sea. In this study, we exposed the common shallow-water Mediterranean sponge Chondrilla nucula (Demospongiae: Chondrillidae) to six different temperatures for 24 h, ranging from temperatures experienced in the field during the year (15, 19, 22, 26, 28 °C) and above normal temperatures (32 °C) and metabolic (respiration rate) and behavioural (clearance rate) were measured. Both respiration and clearance rates were affected by temperature. Respiration rates increased at higher temperatures but were similar between 26 and 32 °C treatments. Clearance rates decreased at temperatures > 26 °C, indicating a drop in food intake that was not reflected by respiration rates. This decline in feeding, while maintaining high respiration rates, may indicate a negative energy balance that could affect this species under chronic or repeated thermal stress exposure. C. nucula will probably be a vulnerable species under climate change conditions, affecting its metabolic performance, ecological functioning and the ecosystem services it provides.

Keywords:
porifera
; mediterranean
; warming
; stressor
; respiration
; clearance
1. Introduction
The Mediterranean Sea is considered a biodiversity hotspot [1] as it hosts a rich biota that includes cold-temperate and subtropical species [2]. It is also expected to be one of the most vulnerable regions to climate change (CC) drivers, particularly ocean warming [3]. The frequency of extreme weather events such as droughts and heat waves has increased since 1950 [4] and their occurrence is expected to keep increasing in the future [5]. In the Mediterranean Sea, even short-duration temperature anomalies had detrimental effects on marine diversity. In the summers of 1999, 2003, 2008, and 2017 above normal surface water temperatures led to disease outbreaks and mass mortality events that affected numerous benthic species [6], including porifers [7,8,9,10,11,12,13], with up to 90 % sponge mortality in some locations [10].
Sponges are key components of benthic habitats worldwide as they contribute in several ways to ecosystem functioning [14]. Their key roles include substrate consolidation, habitat provision, bentho-pelagic energy transfer and seawater filtration [15,16,17,18]. Sponges can be the dominant fauna in shallow tropical and temperate reef habitats, where they can be found in high densities [19,20,21]. In temperate rocky communities, they are strong competitors for space with other invertebrates [22]. Because of their important roles, reductions in the abundance, biomass, and species richness can result in cascading impacts on marine ecosystems structure and functioning [14,23].
The effects of elevated temperature on sponges’ performance are thought to be species-specific [24]. For example, the sponges Cliona celata and Mycale grandis have been reported to be resistant to temperature of up to 4-5 °C higher than ambient temperatures increase [25,26]. There is also evidence that some sponges can even become dominant in some marine tropical environments as a result of extreme thermal events [27,28]. In the Caribbean Sea, for example, the sponges belonging the genus Chondrilla became the dominant species in the reef after thermal anomalies due to the 1998 El Niño event that had caused severe bleaching and mass mortality of corals [29]. On the contrary, other species present relatively low thermal tolerance, such as the Mediterranean sponges Crambe crambe and Petrosia ficiformis whose upper thermal limit has been determined at 26 ºC [30]. Although sponges have been proposed as putative winners under climate change scenarios [28,31], many species experience physiological stress when exposed to elevated temperatures [24].
Studies assessing the effects of elevated temperatures on sponges through controlled, manipulative experiments are lacking, and they are mostly limited to tropical sponges [32,33,34]. The lethal effects of high temperatures include extensive bleaching and disease, loss of symbionts and increase necrosis and mortality [33,34,35,36]. The sub-lethal effects of elevated temperatures include decreased growth and bioerosion rates [36], increased metabolic rates [33,34] and reduced filtering efficiency and pumping rates [32].
In the Mediterranean Sea, sponges can dominate some shallow rocky areas (Strano et al., 2020). Despite the importance of sponges in this region, to date no manipulative studies have been carried to investigate how elevated temperature might affect the metabolic machinery of this benthic group. Here, we used Chondrilla nucula (Scmidt, 1862) as a model species to explore the role of increasing temperature on one among the most common sponge in the Mediterranean Sea. It is a photophylous encrusting sponge living in shallow waters, from the surface up to ~ 30 m depth, forming large patches on well-lit hard substrata. It is a strong competitor for space, feature that makes it a dominant species in some benthic habitats [37]. Chondrilla nucula is also of high commercial interest for the presence of bioactive compounds, which make it an eligible candidate for bioremediation [38]. Due to its shallow distribution, it is potentially exposed to temperature spikes, marine heat waves effects and sea-surface temperature increase under CC scenarios. Thus, we tested the species response (in terms of respiration and clearance rates) to short-term exposure to different increasing/decreasing water temperatures (ranging from 15 to 32 °C) simulating different temperature spikes conditions.
2. Materials and Methods
2.1. Sponge Collection and Preparation
In October 2009, around 100 specimens of C. nucula (fragments of ~ 4 x 4 cm in size) were collected from 5 to 10 m depth by SCUBA divers from the Northern coast of Sicily, Italy. Upon collection, sponges were transported to the Ecology Laboratory at the University of Palermo, Italy, and maintained in aerated aquaria with seawater maintained at the same temperature of the collection site (22 °C). Each aquarium (8 l) contained 4–5 sponge samples. Epibionts were carefully removed from the sponge surfaces so that the successive measures (respiration and clearance rates) would not be compromised. Sponges were then randomly assigned to six temperature treatments: 15, 19, 22, 26, 28 and 32 °C (n = 8 for each one, divided in two aquaria for each treatment level). After one day of acclimation in the aquaria at 22 °C, the water temperatures were slowly changed (rate of 0.25 ºC per hour based on [39,40,41,42] and unpublished data on study site thermal series) to the experimental temperatures, at which sponges were exposed for further 24 h simulating short-intense temperature spikes [41,43,44]. At the end of this period, sponges were sampled for respiration rates and clearance rates measurements. Sponges were handled underwater during all the stages to avoid stress associated with air exposure.
2.2. Respiration Rates
Respiration rates were measured on 8 replicates for each temperature treatment according to a well-tested experimental protocol adopted in several companion experiments in the last decade (e.g. (Bosch-Belmar et al., 2021; Cheng et al., 2018; Sarà et al., 2013, 2021). To be sure that there was not any stressful interference due to manipulation, we tested if sponges were actively pumping with fluorescein dye before placing them into individual respiration chambers. Sponges were placed individually into 500 ml oxygen-saturated respiration chambers with filtered (Whatman GF/C 0.45 μm) seawater and acclimated in the dark for 20 minutes prior to start the Dissolved Oxygen (DO) readings. Water inside the chambers was mixed through magnetic stir bars at the bottom of a compartment on which each sponge was sitting. Respiration chambers were kept in a water bath to maintain constant water temperature. DO was measured with PyroScience Firesting O2 oxygen fibreglass sensors (four sensor connections per logger) and continuously recorded for 1.5 h in the dark, to avoid any potential oxygen production by photosynthetic symbionts. Blank incubations (n = 2 for each temperature treatment) were carried to adjust actual respiration measurements for any background microbial respiration in the seawater.
Respiration rate [RR, μmol O2 h−1g-1 AFDW] was calculated as:
according to Sarà et al. (2013), where Ct0 was the oxygen concentration at the beginning of the measurement, Ct1 was the oxygen concentration at the end of the measurement, Volr was the volume of water in the respirometric chamber and 60 refers to the measurement time. Respiration rates were standardized for the sponge ash free dry weight (AFDW) after drying sponge samples at 60 °C until constant weight, followed by ashing at 500 °C for 5 h to obtain the organic weight.
RR= (Ct0 - Ct1) • Volr • 60 • (t1-t0)-1
2.3. Clearance Rates
Clearance rates were measured on 8 replicates for each temperature treatment which were not the same of those used for the respiration measures. Sponges were placed individually into one-litre chambers and acclimated for 20 min. Feeding chambers were maintained in a water bath to maintain constant water temperature. Water mixing within the chambers was achieved using magnetic stir bars. After acclimation, Isocrysis galbana cells were added to the feeding chambers to obtain a concentration of 2.5 x 104 cells ml−1. Water samples (20 ml) were taken immediately after microalgae addition (T0) and every 30 min for two hours (T1, T2,…Tend) . Control feeding chambers (n = 2 for each temperature) were used to correct for any drop in cell concentration. Water samples were analysed with a Coulter Counter to assess particle concentration. Clearance rates [CR, l h−1] were calculated after Sarà et al. (2009) using the following formula:
where C1 and C2 are the cells concentration at the beginning and at the end of each time interval. Clearance rates were standardized for the sponge AFDW, following the methods described above
CR = (ln C1 − ln C2) / time
2.4. Statistics and Modelling
Statistical Analysis
The effect of temperature on respiration rates was tested with a one-way ANOVA. Respiration rate data was log-transformed to meet the normality of the residuals assumption. Tukey post-hoc pairwise comparisons were conducted to assess whether significant differences between treatments occurred. Clearance rate data was square-root transformed to meet the normality of the residuals assumption. The homogeneity of variance assumption was not met, even after data transformation, therefore the effect of temperature on clearance rates was tested with a one-way Welch’s ANOVA [49], which is not sensitive to unequal variance. Games-Howell post-hoc pairwise comparisons [50] were conducted to assess where significant differences between treatments occurred. Normality of the residuals and homogeneity of variance were tested with the Shapiro-Wilk’s and Levene test (Levene, 1960; Shapiro & Wilk, 1965), respectively. Statistical analyses and plots were performed in R version 4.1.1 [53].
Thermal Risk Map
According to obtained results from RR and CR experiments it was possible to identified a temperature threshold at which the species presents a performance impairment. Such temperature was used to create a thermal risk map for C. nucula at Mediterranean scale. Monthly mean temperature from 2019 to 2022 was download from Copernicus Marine Service Information at high spatial resolution (0.08 x 0.08 decimal degrees) for the whole Mediterranean basin. Temperature raster layer was created and the mean annual number of days exceeding the thermal threshold was calculated at pixel level.
3. Results
3.1. Respiration Rates
Temperature had a significant effect on C. nucula respiration rates (F(5,42)= 82.76, p = < 0.001). Sponge respiration rates increased with temperature (Figure 1). The lowest respiration rates were measured in the 15 °C treated sponges, while the highest respiration rates were observed in the 26 °C treatment group (2.73 ± 0.59 vs 75.1 ± 29.93 µmol O2 h–1 g (AFDW)–1, mean ± 95 % CI). Sponge respiration rates under 15°C treatment were significantly lower than those of all the other treatments; while respiration rates in the 26, 28 and 32 °C treatments (with non-significant differences among them) were significantly higher than respiration rates measured in the 19 and 22 °C treatments (Figure 1).
3.2. Clearance Rates
Temperature had a significant effect on C. nucula clearance rates (F(5,18) = 25.235, p < 0.001) (Figure 2, and Supplementary Material Figure S1). The lowest clearance rates were measured in the 15 °C treatment, whereas the highest clearance rates were measured in the experimental sponges belonging to the 26 °C treatment (0.13 ± 0.035 vs 0.5 ± 0.192 l h–1 g(AFDW)–1, mean ± 95 % CI; Figure 2). Clearance rates in the 15 °C treatment were significantly lower than those of the 19 and 22 ºC treatments. Clearance rates decreased in the 28 and 32 °C treatment compared to values recorded in sponges in the 19−26 °C treatments, however this difference was statistically significant only for the 28 °C treatment.
3.3. Thermal Risk Map
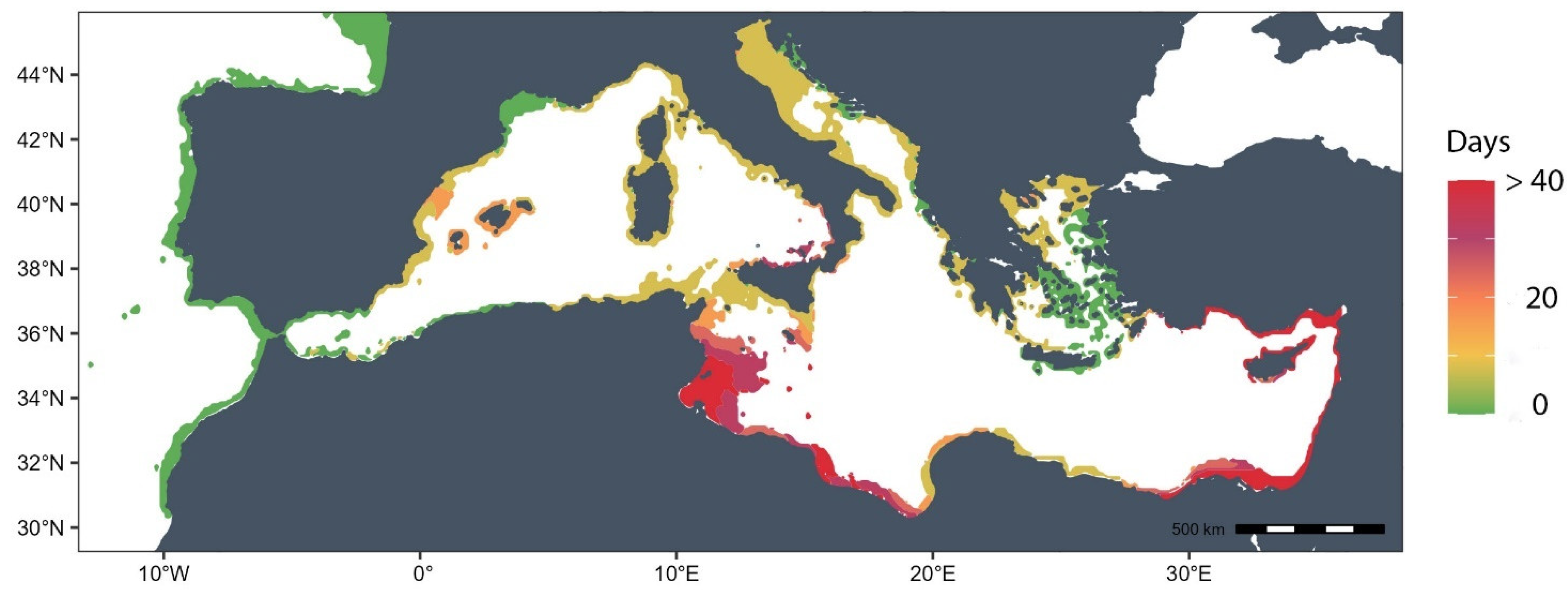
The lowest respiration rates were observed at 15 °C, aligning with the lowest clearance rates. Conversely, the highest respiration rates were associated with the maximum feeding rates in the 26 °C treatment. Despite the higher respiration rates recorded at 28 °C and 32 °C, they do not signify elevated metabolic rates linked to feeding activity. Instead, they likely reflect a response to stressful environmental conditions. Based on these findings, a temperature of 28ºC has been selected as the thermal threshold for creating a risk map.
The thermal map reveals that over 75% of the Mediterranean coast experiences 20 or more days per year with temperatures exceeding 28ºC. The southern coast of the basin, especially the Tunisian and eastern Mediterranean coast, exhibits the highest incidence of extreme temperatures. Conversely, the southern Aegean Sea, Croatian coast, the Gulf of Lion, and the Alboran Sea maintain annual temperatures below this threshold. The remaining coastline, with a particular emphasis on northern Sicily, registers between 15 and more than 40 days of elevated temperatures.
4. Discussions
Using a controlled laboratory experiment we investigated, for the first time, the responses of the Mediterranean shallow-water sponge Chondrilla nucula to a range of temperatures, including stressful thermal conditions projected under climate change scenarios. Respiration rates significantly decreased at the lowest tested temperature (15 ºC); while increased at higher temperatures, even if did not vary significantly between 26, 28 and 32 ºC treatments (being the last above normal seawater temperature, 4 °C higher than the maximum seawater temperature recorded in summer 2009). On the contrary, clearance rates decreased at the two highest temperatures (28 ºC and 32 ºC), indicating possible negative consequences on the energy balance of this species at temperatures equal or higher than 28 °C.
Higher respiration rates in sponges have been reported in summer seasons, from in situ measurements, both in tropical and temperate regions [54,55] showing that temperature exerts strong control over sponge metabolic rates. Different manipulative experiments focused on long and short-term exposure to thermal stressors equally observed increasing sponge respiration rates at the highest temperature treatments (most of them mirroring ocean warming conditions). Beepat et al. (2020) reported increased respiration rates in the sponges Neopetrosia exigua between 26 °C (control temperature) and 30 °C (CC projection), and in Amphimedon navalis and Spheciospongia vagabunda between 26 and 28 °C, after two weeks of thermal stress exposure. Similarly, Bennett et al. (2017) reported significant higher respiration rates in the tropical sponges Carteriospongia foliascens, Rhopaloeides odorabile and Cymbastela corallophila at 31.5 and 30 °C compared to the control temperature 28.5 °C. Beepat et al. (2021) observed higher oxygen consumption rates of three different sponge species after short-term exposure to 26 ºC, 28 ºC and 30 ºC temperature treatments. Only few studies have investigated the effects of temperature on sponge clearance rates, with contrasting results. Reduced clearance rates in response to elevate temperatures have been reported in the tropical sponge Rhopaloeides odorabile after exposure to temperatures 3 °C higher than control, under laboratory manipulation [32]. On the contrary, an increase in clearance rates was reported in the temperate sponge Halichondria panicea at 12 °C compared to 6 °C, in laboratory conditions [56]. Finally, an in situ study did not report altered clearance rates in Mediterranean sponges as a function of temperature, under seasonal temperature ranges [57]. While the response of sponge feeding to thermal anomalies is probably species specific, future increased sea surface temperature is likely to reduce the ability of sponges to actively pump water.
Sponge ability to feed, as well as respiration, is directly linked to sponge pumping rates [58]. Massaro et al. (2012), who reported a decline in clearance rates in sponges exposed to increasing temperatures, also reported a decrease in pumping rates. Beepat et al. (2021) reported increased pumping rates along with increased respiration rates in tropical sponges exposed to thermal stress. As respiration rates and clearance rates are linked to pumping rates, we expected these two traits should follow similar patterns in response to different temperatures. While the lowest respiration rates corresponded to the lowest clearance rates at 15 °C, and the highest respiration rates corresponded to the highest feeding rates in the 26 °C treatment, this correlation ceased at temperatures > 26 °C. The higher respiration rates at 28 °C and 32 °C do not indicate elevated metabolic rates related to feeding activity but it probably is a response to stressful environmental conditions.
Respiration rates represent a measure of the part of the food intake required to provide energy to support life processes [59]. In our study, the contrasting response of two related functional traits may indicate a greater energy expense than the energy gained through food as an immediately response to short-duration thermal stressor. Such response may indicate the proximity of the upper thermal tolerance limit for the species, triggering an energy imbalance under stressful conditions. The effects under recurrent and chronic stressor exposure could be more severe, as less energy might be directed towards processes related to organism life-history traits, such as growth and reproduction [60], dealing with consequences at population level.
Mechanisms to adapt to thermal stress have been studied in different sponges’ species confirming certain ability to recover after stressful conditions [61]. Even if larger degree of stress may cause the metabolic defence/compensation system to collapse some sponge species have showed survival ability by reducing cellular activity under intense thermal stress [62], indicating some adaptability to recover from significant heat stressful conditions [63]. Nevertheless, in the current and future scenarios of CC, temperature spikes and heat waves are predicted to increase in frequency and intensity [13], influencing species responses, acclimation and adaptation mechanisms. There is scientific evidence of massive mortality events on marine benthic environments due to stressful thermal events [9,64,65]. Sponges responses to these conditions are different (in particular those presenting photosymbionts) with some species showing bleaching and necrotic tissue, while others remained unaffected [34,66,67]. Much species living close to their upper thermal limits would not resist or be able to compensate lasting thermal stressful conditions, that interacting with other local stressors may result in high impacts at population and community levels. Even if current thermal conditions seem not to negatively affect C. nucula in the Mediterranean Sea, future scenarios of climate change may jeopardise the species functioning and occurrence, as well as the ecosystem services it provides, over the basin.
5. Conclusions
This study demonstrates that the Mediterranean shallow-water sponge Chondrilla nucula has a rapid response to change in temperature, with altered respiration and clearance rates. Respiration rates increased with temperature, while clearance rates decreased significantly at temperatures higher than 26 °C. The different patterns of respiration and clearance rates suggest greater energy expenses than energy gained through food intake at higher temperatures in response to acute short-term thermal stressor. Although sponges have been proposed as putative winners under climate change scenarios, in the Mediterranean Sea, even short-duration temperature anomalies may have detrimental effects on sponges’ communities and overall benthic biodiversity. Further research focuses on the species thermal tolerance thresholds is needed to better understand its potential response to chronic or repeated thermal stress exposure that could negatively affect species populations of such key taxa in the ecosystem functioning.
Author Contributions
MBB and VM analysed the data and wrote the first draft of this manuscript; MM and AS carried the experiment and collected the data, GS conceived and funded the study. All authors reviewed the manuscript.
Acknowledgments
This research was funded by the InterMED project.
References
- Coll, M.; Piroddi, C.; Albouy, C.; Ben Rais Lasram, F.; Cheung, W.W.L.; Christensen, V.; Karpouzi, V.S.; Guilhaumon, F.; Mouillot, D.; Paleczny, M.; et al. The Mediterranean Sea under Siege: Spatial Overlap between Marine Biodiversity, Cumulative Threats and Marine Reserves. Glob. Ecol. Biogeogr. 2012, 21, 465–480. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea Biodiversity between the Legacy from the Past and a Future of Change. Life Mediterr. Sea: A Look. Habitat 2012,, 1–55. [Google Scholar]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate Change and Interconnected Risks to Sustainable Development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef]
- Baldi, M.; Dalu, G.A.; Maracchi, G.; Pasqui, M.; Cesarone, F. Heat Waves in the Mediterranean: A Local Feature or a Larger-scale Effect? Int. J. Climatol. 2006, 26. [Google Scholar] [CrossRef]
- IPCC Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V.P., Zhai, A., Pirani, S.L., Connors, C., Péan, S., Berger, N., Caud, Y., Chen, L., Goldfarb, M.I., Gomis, M., Huang, K., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021. [Google Scholar]
- Sarà, G.; Giommi, C.; Giacoletti, A.; Conti, E.; Mulder, C.; Mangano, M.C. Multiple Climate-Driven Cascading Ecosystem Effects after the Loss of a Foundation Species. Sci. Total Environ. 2021, 770, 144749. [Google Scholar] [CrossRef] [PubMed]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Cattaneo-Vietti, R.; Bava, S.; Morganti, C.; Morri, C.; Picco, P.; Sara, G.; Schiaparelli, S.; et al. A Catastrophic Mass-Mortality Episode of Gorgonians and Other Organisms in the Ligurian Sea (North-Western Mediterranean), Summer 1999. Ecol Lett 2000, 3, 284–293. [Google Scholar] [CrossRef]
- Perez, T.; Garrabou, J.; Sartoretto, S.; Harmelin, J.G.; Francour, P.; Vacelet, J. Massive mortality of marine invertebrates: an unprecedented event in northwestern Mediterranean. C R. Acad. Sci. III 2000, 323, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Garrabou, J.; Coma, R.; Bensoussan, N.; Bally, M.; Chevaldonné, P.; Cigliano, M.; Diaz, D.; Harmelin, J.G.; Gambi, M.C.; Kersting, D.K.; et al. Mass Mortality in Northwestern Mediterranean Rocky Benthic Communities: Effects of the 2003 Heat Wave. Glob Chang Biol 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Klautau, M.; Russo, C.A.M.; Lazoski, C.; Boury-Esnault, N.; Thorpe, J.P.; Solé-Cava, A.M. Does Cosmopolitanism Result from Overconservative Systematics? A Case Study Using the Marine Sponge Chondrilla Nucula. Evolution 1999, 53, 1414–1422. [Google Scholar] [CrossRef] [PubMed]
- Cebrian, E.; Uriz, M.J.; Garrabou, J.; Ballesteros, E. Sponge Mass Mortalities in a Warming Mediterranean Sea: Are Cyanobacteria-Harboring Species Worse Off? PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Ereskovsky, A.; Ozerov, D.A.; Pantyulin, A.N.; Tzetlin, A.B. Mass Mortality Event of White Sea Sponges as the Result of High Temperature in Summer 2018. Polar Biol 2019, 42, 2313–2318. [Google Scholar] [CrossRef]
- Garrabou, J.; Gómez-Gras, D.; Medrano, A.; Cerrano, C.; Ponti, M.; Schlegel, R.; Bensoussan, N.; Turicchia, E.; Sini, M.; Gerovasileiou, V.; et al. Marine Heatwaves Drive Recurrent Mass Mortalities in the Mediterranean Sea. Glob. Chang. Biol. 2022, 5708–5725. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.J. The Functional Roles of Marine Sponges. Estuar. Coast. Shelf Sci. 2008, 79, 341–353. [Google Scholar] [CrossRef]
- Reiswig, H.M. In Situ Pumping Activities of Tropical Demospongiae. Mar. Biol. 1971, 9, 38–50. [Google Scholar] [CrossRef]
- Reiswig, H.M. Water Transport, Respiration and Energetics of Three Tropical Marine Sponges. J. Exp. Mar. Biol. Ecol. 1974, 14, 231–249. [Google Scholar] [CrossRef]
- Ribeiro, S.M.; Omena, E.P.; Muricy, G. Macrofauna Associated to Mycale Microsigmatosa (Porifera, Demospongiae) in Rio de Janeiro State, SE Brazil. Estuar. Coast. Shelf Sci. 2003, 57, 951–959. [Google Scholar] [CrossRef]
- Pile, A.J.; Young, C.M. The Natural Diet of a Hexactinellid Sponge: Benthic–Pelagic Coupling in a Deep-Sea Microbial Food Web. Deep. Sea Res. Part. I: Oceanogr. Res. Pap. 2006, 53, 1148–1156. [Google Scholar] [CrossRef]
- Wilkinson, C.R.; Evans, E. Sponge Distribution across Davies Reef, Great Barrier Reef, Relative to Location, Depth, and Water Movement. Coral Reefs 1989, 8, 1–7. [Google Scholar] [CrossRef]
- Murillo, F.J.; Muñoz, P.D.; Cristobo, J.; Ríos, P.; González, C.; Kenchington, E.; Serrano, A. Deep-Sea Sponge Grounds of the Flemish Cap, Flemish Pass and the Grand Banks of Newfoundland (Northwest Atlantic Ocean): Distribution and Species Composition. Mar. Biol. Res. 2012, 8, 842–854. [Google Scholar] [CrossRef]
- Maldonado, M.; Aguilar, R.; Bannister, R.J.; Bell, J.J.; Conway, K.W.; Dayton, P.K.; Díaz, C.; Gutt, J.; Kelly, M.; Kenchington, E.L.R.; et al. Sponge Grounds as Key Marine Habitats: A Synthetic Review of Types, Structure, Functional Roles, and Conservation Concerns BT - Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., Orejas Saco del Valle, C., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–39. ISBN 978-3-319-17001-5. [Google Scholar]
- Bell, J.J.; Barnes, D.K.A. The Importance of Competitor Identity, Morphology and Ranking Methodology to Outcomes in Interference Competition between Sponges. Mar. Biol. 2003, 143, 415–426. [Google Scholar] [CrossRef]
- Peterson, B.; Chester, C.; Jochem, F.; Fourqurean, J. Potential Role of Sponge Communities in Controlling Phytoplankton Blooms in Florida Bay. Mar. Ecol. Prog. Ser. 2006, 328, 93–103. [Google Scholar] [CrossRef]
- Bell, J.J.; Bennett, H.M.; Rovellini, A.; Webster, N.S. Sponges to Be Winners under Near-Future Climate Scenarios. Bioscience 2018, 68, 955–968. [Google Scholar] [CrossRef]
- Miller, A.N.; Strychar, K.B.; Shirley, T.C.; Rützler, K. Effects of Heat and Salinity Stress on the Sponge Cliona Celata. Int. J. Biol. 2010, 2, 2–16. [Google Scholar] [CrossRef]
- Duckworth, A.R.; West, L.; Vansach, T.; Stubler, A.; Hardt, M. Effects of Water Temperature and PH on Growth and Metabolite Biosynthesis of Coral Reef Sponges. Mar. Ecol. Prog. Ser. 2012, 462, 67–77. [Google Scholar] [CrossRef]
- Bell, J.J.; Smith, D. Ecology of Sponge Assemblages (Porifera) in the Wakatobi Region, South-East Sulawesi, Indonesia: Richness and Abundance. J. Mar. Biol. Assoc. United Kingd. 2004, 84, 581–591. [Google Scholar] [CrossRef]
- Kelmo, F.; Bell, J.J.; Attrill, M.J. Tolerance of Sponge Assemblages to Temperature Anomalies: Resilience and Proliferation of Sponges Following the 1997-8 El-Niño Southern Oscillation. PLoS ONE 2013, 8, e76441. [Google Scholar] [CrossRef] [PubMed]
- Aronson, R.; Precht, W.; Toscano, M.; Koltes, K. The 1998 Bleaching Event and Its Aftermath on a Coral Reef in Belize. Mar Biol 2002, 141, 435–447. [Google Scholar] [CrossRef]
- Gómez-Gras, D.; Linares, C.; de Caralt, S.; Cebrian, E.; Frleta-Valić, M.; Montero-Serra, I.; Pagès-Escolà, M.; López-Sendino, P.; Garrabou, J. Response Diversity in Mediterranean Coralligenous Assemblages Facing Climate Change: Insights from a Multispecific Thermotolerance Experiment. Ecol. Evol. 2019, 9, 4168–4180. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.J.; Davy, S.K.; Jones, T.; Taylor, M.W.; Webster, N.S. Could Some Coral Reefs Become Sponge Reefs as Our Climate Changes? Glob. Chang. Biol. 2013, 19, 2613–2624. [Google Scholar] [CrossRef] [PubMed]
- Massaro, A.J.; Weisz, J.B.; Hill, M.S.; Webster, N.S. Behavioral and Morphological Changes Caused by Thermal Stress in the Great Barrier Reef Sponge Rhopaloeides Odorabile. J. Exp. Mar. Biol. Ecol. 2012, 416–417, 55–60. [Google Scholar] [CrossRef]
- Beepat, S.S.; Davy, S.K.; Woods, L.; Bell, J.J. Short-Term Responses of Tropical Lagoon Sponges to Elevated Temperature and Nitrate. Mar. Env. Res. 2020, 157, 104922. [Google Scholar] [CrossRef]
- Bennett, H.M.; Altenrath, C.; Woods, L.; Davy, S.K.; Webster, N.S.; Bell, J.J. Interactive Effects of Temperature and PCO(2) on Sponges: From the Cradle to the Grave. Glob. Chang. Biol. 2017, 23, 2031–2046. [Google Scholar] [CrossRef]
- Webster, N.S. Sponge Disease: A Global Threat? Env. Microbiol. 2007, 9, 1363–1375. [Google Scholar] [CrossRef] [PubMed]
- Achlatis, M.; Van Der Zande, R.M.; Schönberg, C.H.L.; Fang, J.K.H.; Hoegh-Guldberg, O.; Dove, S. Sponge Bioerosion on Changing Reefs: Ocean Warming Poses Physiological Constraints to the Success of a Photosymbiotic Excavating Sponge. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Strano, F.; Micaroni, V.; Costa, G.; Bertocci, I.; Bertolino, M. Shallow-Water Sponge Grounds along the Apulian Coast (Central Mediterranean Sea). Mar. Biodivers. 2020, 50, 7. [Google Scholar] [CrossRef]
- Chelossi, E.; Pantile, R.; Pronzato, R.; Milanese, M.; Hentschel, U. Bacteria with Antimicrobial Properties Isolated from the Mediterranean Sponges Chondrilla Nucula and Petrosia Ficiformis. Aquat. Microb. Ecol. 2007, 49, 157–163. [Google Scholar] [CrossRef]
- Prusina, I.; Sarà, G.; De Pirro, M.; Dong, Y.W.; Han, G.D.; Glamuzina, B.; Williams, G.A. Variations in Physiological Responses to Thermal Stress in Congeneric Limpets in the Mediterranean Sea. J. Exp. Mar. Biol. Ecol. 2014, 456, 34–40. [Google Scholar] [CrossRef]
- Montalto, V.; Bagarella, R.; Rinaldi, A.; Sarà, G.; Mirto, S. Thermal Adaptation and Physiological Responses to Environmental Stress in Tunicates. Aquat. Biol. 2017, 26, 179–184. [Google Scholar] [CrossRef]
- Alsuwaiyan, N.; Vranken, S.; Filbee-Dexter, K.; Cambridge, M.; Coleman, M.; Wernberg, T. Genotypic Variation in Response to Extreme Events May Facilitate Kelp Adaptation under Future Climates. Mar. Ecol. Prog. Ser. 2021, 672. [Google Scholar] [CrossRef]
- Bosch-Belmar, M.; Giacoletti, A.; Giommi, C.; Girons, A.; Milisenda, G.; Sarà, G. Short-Term Exposure to Concurrent Biotic and Abiotic Stressors May Impair Farmed Molluscs Performance. Mar. Pollut. Bull. 2022, 179. [Google Scholar] [CrossRef]
- Hobday, A.J.; Alexander, L.V.; Perkins, S.E.; Smale, D.A.; Straub, S.C.; Oliver, E.C.J.; Benthuysen, J.A.; Burrows, M.T.; Donat, M.G.; Feng, M.; et al. A Hierarchical Approach to Defining Marine Heatwaves. Prog. Ocean. 2016, 141, 227–238. [Google Scholar] [CrossRef]
- Juza, M.; Fernández-Mora, À.; Tintoré, J. Sub-Regional Marine Heat Waves in the Mediterranean Sea from Observations: Long-Term Surface Changes, Sub-Surface and Coastal Responses. Front. Mar. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Sarà, G.; Palmeri, V.; Montalto, V.; Rinaldi, A.; Widdows, J. Parameterisation of Bivalve Functional Traits for Mechanistic Eco-Physiological Dynamic Energy Budget (DEB) Models. Mar. Ecol. Prog. Ser. 2013, 480, 99–117. [Google Scholar] [CrossRef]
- Cheng, M.C.F.; Sarà, G.; Williams, G.A. Combined Effects of Thermal Conditions and Food Availability on Thermal Tolerance of the Marine Bivalve, Perna Viridis. J. Therm. Biol. 2018, 78, 270–276. [Google Scholar] [CrossRef]
- Bosch-Belmar, M.; Giommi, C.; Milisenda, G.; Abbruzzo, A.; Sarà, G. Integrating Functional Traits into Correlative Species Distribution Models to Investigate the Vulnerability of Marine Human Activities to Climate Change. Sci. Total Environ. 2021, 799, 149351. [Google Scholar] [CrossRef]
- Sarà, G.; Milisenda, G.; Mangano, M.C.; Bosch-belmar, M. The Buffer Effect of Canopy-Forming Algae on Vermetid Reefs ’ Functioning : A Multiple Stressor Case Study. Mar. Pollut. Bull. 2021, 171, 112713. [Google Scholar] [CrossRef]
- Welch, B. On the Comparison of Several Mean Values: An Alternative Approach. Biometrika 1951, 38, 330–336. [Google Scholar] [CrossRef]
- Games, P.A. Howell, J.F. Pairwise Multiple Comparison Procedures with Unequal N’s and/or Variances: A Monte Carlo Study. J. Educ. Behav. Stat. 1976, 1, 113–125. [Google Scholar]
- Levene, H. Robust Tests for Equality of Variances. In Contributions to Probability and Statistics, Stanford University Press; Olkin, I., Ed.; Palo Alto: 1960; pp. 278–292.
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing 2021.
- Cheshire, A.C.; Butler, A.; Westphalen, G.; Rowland, B.C.; Stevenson, J.; Wilkinson, C.R. Preliminary Study of the Distribution and Photophysiology of the Temperate Phototrophic Sponge Cymbastela Sp. from South Australia. Mar. Freshw. Res. 1995, 46, 1211–1216. [Google Scholar] [CrossRef]
- Coma, R.; Ribes, M.; Gili, J.M.; Zabala, M. Seasonality of in Situ Respiration Rate in Three Temperate Benthic Suspension Feeders. Limnol. Ocean. 2002, 47, 324–331. [Google Scholar] [CrossRef]
- Riisgard, H.U.; Thomassen, S.; Jakobsen, H.; Weeks, J.M.; Larsen, P.S. Suspension Feeding in Marine Sponges Halichondria Panicea and Haliclona Urceolus: Effects of Temperature on Filtration Rate and Energy Cost of Pumping. Mar. Ecol. Prog. Ser. 1993, 96, 177–188. [Google Scholar] [CrossRef]
- Ribes, M.; Coma, R.; Gili, J.M. Natural Diet and Grazing Rate of the Temperate Sponge Dysidea Avara (Demospongiae, Dendroceratida) throughout an Annual Cycle. Mar. Ecol. Prog. Ser. 1999, 176, 179–190. [Google Scholar] [CrossRef]
- Gerrodette, T.; Flechsig, A.O. Sediment-Induced Reduction in the Pumping Rate of the Tropical Sponge Verongia Lacunosa. Mar. Biol. 1979, 55, 103–110. [Google Scholar] [CrossRef]
- Kooijman. Dynamic Energy Budget Theory for Metabolic Organisation, 3rd ed.; Cambridge University Press, 2010. [Google Scholar]
- Whalan, S.; Ettinger-Epstein; Battershill; de Nys, R. Larval Vertical Migration and Hierarchical Selectivity of Settlement in a Brooding Marine Sponge. Mar. Ecol. Prog. Ser. 2008, 368, 145–154. [Google Scholar] [CrossRef]
- López-Legentil, S.; Song, B.; McMurray, S.E.; Pawlik, J.R. Bleaching and Stress in Coral Reef Ecosystems: Hsp70 Expression by the Giant Barrel Sponge Xestospongia Muta. Mol. Ecol. 2008, 17, 1840–1849. [Google Scholar] [CrossRef] [PubMed]
- Pantile, R.; Webster, N. Strict Thermal Threshold Identified by Quantitative PCR in the Sponge Rhopaloeides Odorabile. Mar. Ecol. Prog. Ser. 2011, 431, 97–105. [Google Scholar] [CrossRef]
- Aguilar-Camacho, J.M.; McCormack, G. Molecular Responses of Sponges to Climate Change. In Climate Change, Ocean Acidification and Sponges: Impacts Across Multiple Levels of Organization; Carballo, J.L., Bell, J.J., Eds.; Springer International Publishing, 2017; pp. 1–452. ISBN 9783319590080. [Google Scholar]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global Warming and Recurrent Mass Bleaching of Corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, C.; Hadjioannou, L.; Petrou, A.; Nikolaidis, A.; Evriviadou, M.; Lange, M.A. Mortality of the Scleractinian Coral Cladocora Caespitosa during a Warming Event in the Levantine Sea (Cyprus). Reg. Env. Change 2016, 16, 1963–1973. [Google Scholar] [CrossRef]
- González-Aravena, M.; Kenny, N.J.; Osorio, M.; Font, A.; Riesgo, A.; Cárdenas, C.A. Warm Temperatures, Cool Sponges: The Effect of Increased Temperatures on the Antarctic Sponge Isodictya Sp. PeerJ 2019, 7, e8088. [Google Scholar] [CrossRef]
- Perkins, N.R.; Monk, J.; Soler, G.; Gallagher, P.; Barrett, N.S. Bleaching in Sponges on Temperate Mesophotic Reefs Observed Following Marine Heatwave Events. Clim. Change Ecol. 2022, 3, 100046. [Google Scholar] [CrossRef]
Figure 1.
Respiration rates of Chondrilla nucula measured at six different temperatures. Bars show mean values ± 95% CI. N = 8. Letters indicate significant differences among temperature treatments.
Figure 1.
Respiration rates of Chondrilla nucula measured at six different temperatures. Bars show mean values ± 95% CI. N = 8. Letters indicate significant differences among temperature treatments.
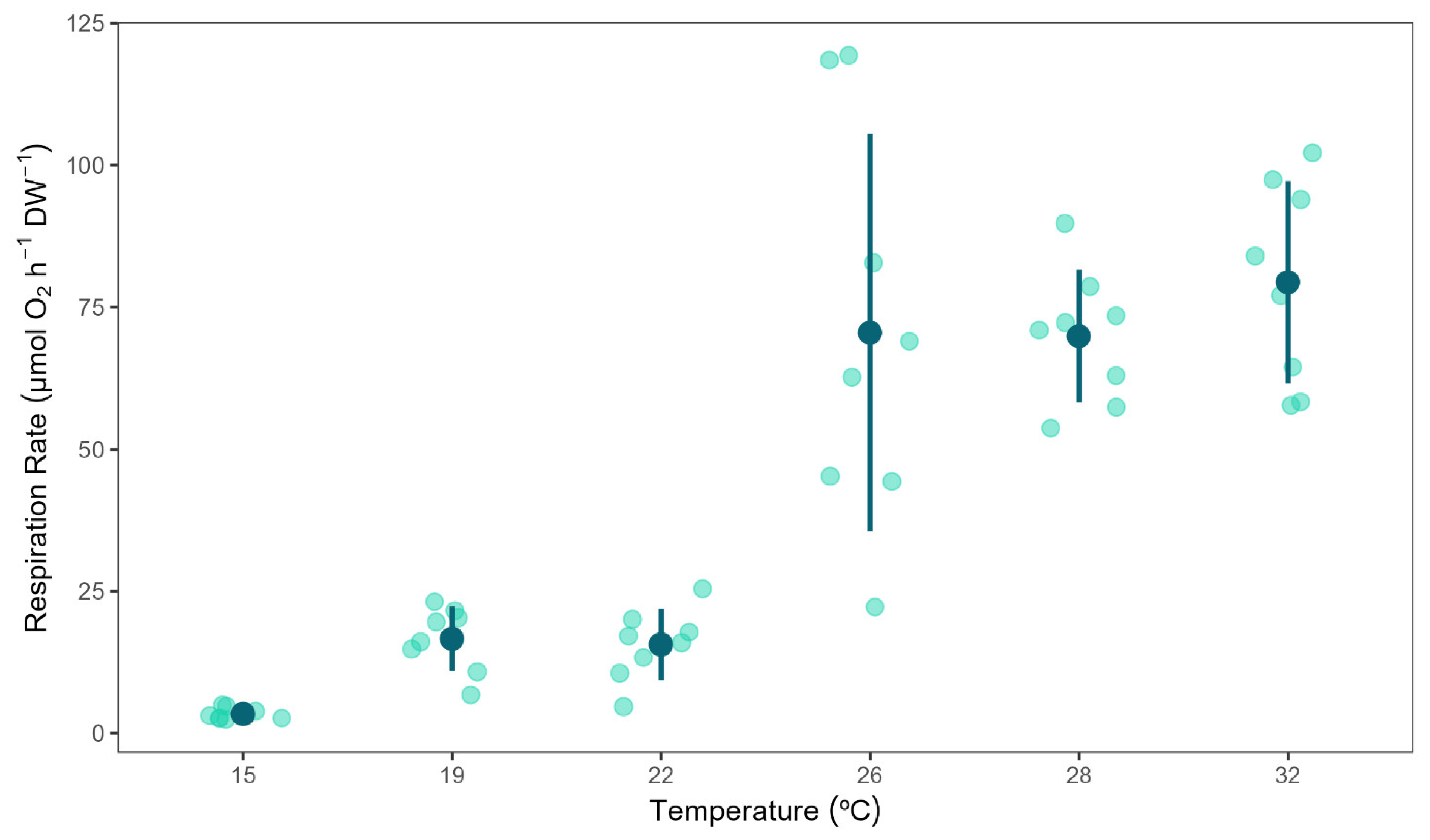
Figure 2.
Clearance rates of Chondrilla nucula measured at six different temperatures. Bars show mean values ± 95% CI . N = 8. Letters indicate significant differences among temperature treatments.
Figure 2.
Clearance rates of Chondrilla nucula measured at six different temperatures. Bars show mean values ± 95% CI . N = 8. Letters indicate significant differences among temperature treatments.
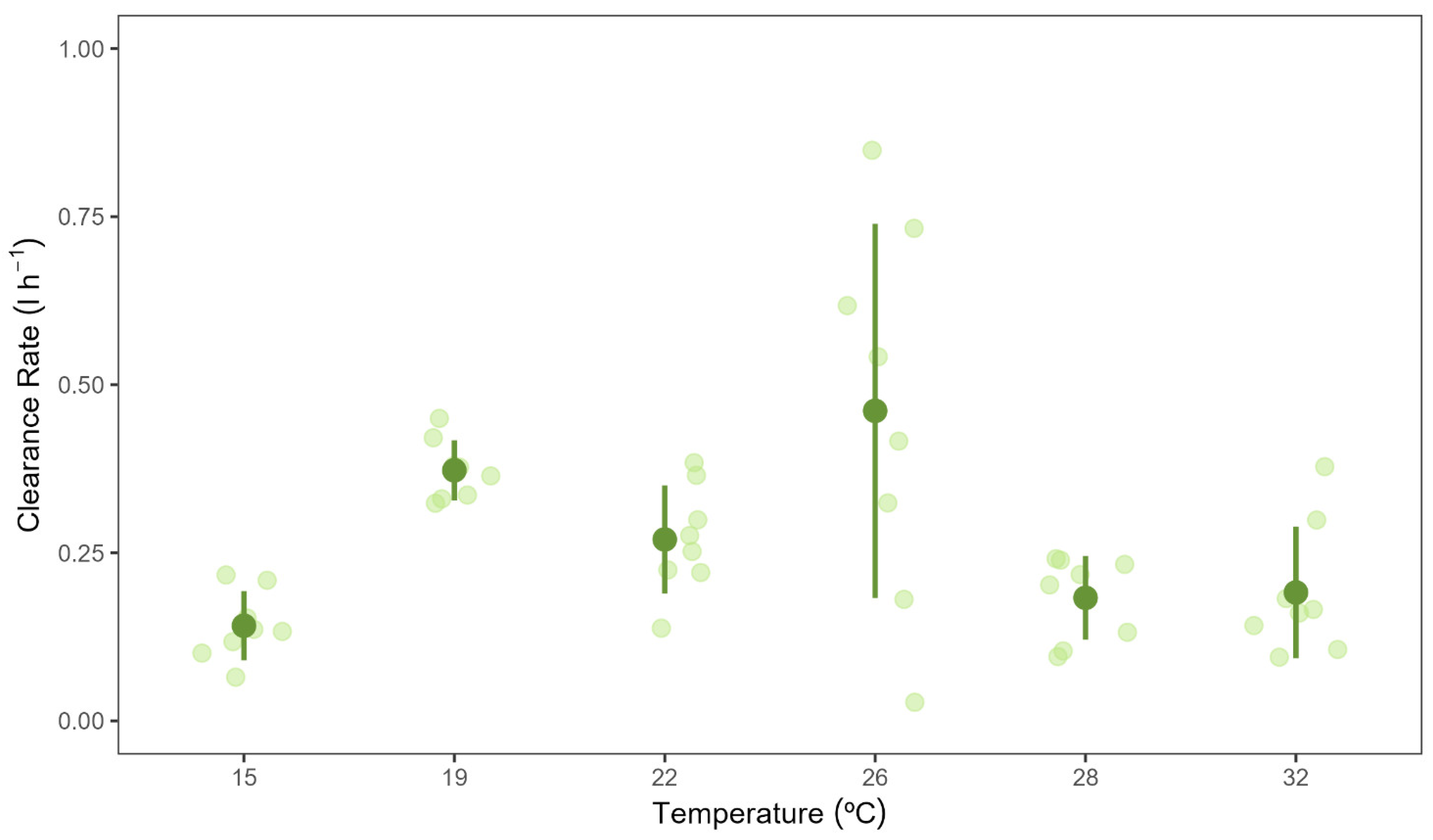
Figure 3.
Risk Map showing the number of days per year when temperature exceeds the temperature threshold of 28ºC in the Mediterranean Sea. Scale ranges from 0 to 1, where 0 represents any day of temperature above 28ºC and 1 more than 40 days exceeding that temperature.
Figure 3.
Risk Map showing the number of days per year when temperature exceeds the temperature threshold of 28ºC in the Mediterranean Sea. Scale ranges from 0 to 1, where 0 represents any day of temperature above 28ºC and 1 more than 40 days exceeding that temperature.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



