
Submitted:
22 November 2023
Posted:
26 November 2023
You are already at the latest version
Abstract
Despite great strides in diagnosis and treatment, cardiac diseases remain the number one cause of mortality in the developed world, with rates catching up at an alarming rate in the developing world. Targeted delivery of therapeutics, specifically to cardiomyocytes, would open up new frontiers. Our prior work using a combinatorial in vitro and in vivo phage display methodology identified a 12-amino acid long peptide that targets cardiomyocytes selectively after an intravenous injection in as little as 5 mins, and hence was termed cardiac targeting peptide (CTP: APHLSSQYSRT). CTP has been used to deliver imaging agents, small drug molecules, photosensitizing nanoparticles, exosomes, and even miRNA to cardiomyocytes. As a natural extension for development of CTP as a clinically viable cardiac vector, we now present toxicity studies performed on the peptide.
In vitro viability studies were performed in a human left ventricular myocyte cell line with 10µM of Cyanine-5.5 labeled CTP (CTP-Cy5.5). In vitro ion channel profiles were completed for CTP followed by more extensive studies performed by Eurofins in stably transfected cell lines for an extensive list of GPCR-coupled receptors. Positive data for GPCR-coupled receptors was interrogated further with RT-qPCRs performed on mouse heart tissue. In vivo studies consisted of pre- and post-blood pressure monitoring acutely after a single CTP (10 mg/Kg) injection. Further in vivo toxicity studies consisted of injecting 60, 6-week-old, wild-type CD1, male/female mice (1:1) injected with CTP (150 µg/Kg) intravenously. Mice were weighed immediately before injections and daily thereafter. Cohorts of mice were euthanized on day 0, 1, 2, 7 and 14 with inhalational CO2, followed by opening of the chest cavity, collection of blood via cardiac puncture, complete blood count analysis, metabolic profiling, and finally, liver, renal and thyroid studies. Lastly, mouse cardiac MRI was performed immediately before and after injections of CTP (150 µg/Kg) to assess changes in cardiac size or function.
Human left ventricular cardiomyocytes showed no decrease in viability after a 30 min incubation with CTP-Cy5.5. In the Eurofins Cardiac Profiler Qube, no significant activation or inhibition of any of seventy-eight protein channels was observed other than OPRM1 and COX2 at the highest tested concentration, neither of which were expressed in mouse heart tissue as assessed by RT-qPCR. There was no change in blood pressures before and after injections with CTP at 10 mg/Kg. Blood counts and chemistries showed no evidence of significant hematological, hepatic or renal toxicities. Lastly, there was no significant difference in cardiac function, size or mass before and after CTP injections.
Our studies with CTP showed no activation or inhibition of GPCR-associated receptors in vitro. We found no signals indicative of toxicity in vivo. Most importantly, cardiac functions remained unchanged acutely in response to CTP uptake. Further studies, in good laboratory practices fashion, are needed with more prolonged, chronic administration of CTP conjugated to a specific cargo of choice before human studies can be contemplated.
Keywords:
Cardiac Magnetic Resonance
; Cardiac Targeting Peptide
; Human Cardiomyocyte Cells
; Cytotoxicity
; Cell Penetrating Peptides
Introduction
Advances in the development of novel technologies in pharmaceutics today necessitate the development of safe and effective targeted delivery method for cargoes such as nanoparticles, siRNA, drugs, and more. Cell penetrating peptides (CPPs), which are small, positively charged peptides capable of crossing the cell membrane barrier, provide a promising method for intracellular delivery which is highly efficient and minimally cytotoxic [1]. TAT (HIV Trans-Activator of Transcription) was the first CPP to be discovered to enter the cell and induce the expression of viral genes [2,3], but given its nature to ubiquitously target all cell types, and even cross the blood-brain barrier, the quest to discover different cell-specific CPPs has become a major endeavor. Among other strategies, phage display libraries have been used successfully to identify peptide sequences which have cell-penetrating properties [3,4,5,6,7], leading to the discovery of many cell-specific CPPs over time. Such CPPs could be utilized for targeted delivery of both diagnostics as well as therapeutics [8]. Our previous work with phage display led to the identification of one such CPP, which we termed cardiac targeting peptide (CTP: APHLSSQYSRT), due to its ability to transduce cardiomyocytes in vivo in as little as 15 minutes after a peripheral intravenous injection [4,9].
Heart disease and cardiac illness continues to be a leading cause of death in the US [10], and as such, novel treatments for increasingly prevalent conditions such as heart failure and atrial fibrillation are sorely needed. CTP has shown promise as a vector for both therapeutics and diagnostic agents [11], and is capable of penetrating cardiomyocytes both in vivo in mice and in vitro in human cardiomyocytes (hCMCs) [4], exhibiting cardiac-specific transduction which is seemingly not species limited as it has also shown efficacy in transducing both sheep and rat models [12], as well as induced pluripotent human cells differentiated into beating cardiomyocytes [9]. CTP appears to primarily transduce the heart and is not taken up by the lungs, spleen, or other muscle subtypes, and does not cross the blood-brain barrier [9]. The cell-specific transduction ability of CTP has been validated several times by independent laboratories as being specific to cardiomyocytes and has been shown to be effective at delivering a variety of cargoes, including photosensitizers [13], exosomes [12], miRNA for the treatment of heart failure [14] and amiodarone [15] at significantly reduced dose, suggesting a potential for reduction in off-target side effects while retaining amiodarone functionality [16].
In conclusion, CTP appears to be a novel non-viral vector, specifically targeting cardiomyocytes, and has shown promise as an innovative technology that can be utilized to deliver a myriad of different cargos. While CTP has shown no outwardly toxic effects, the authors present here systematic studies showing CTP’s lack of toxicity both in vitro and in vivo. We first present in vitro studies in stably transformed cell lines showing no effects of CTP on important receptors like calcium, sodium, potassium, or HERG2 receptors. In vivo studies show a lack of overt hepatic or renal toxicity with normal biomarkers, and no acute effect of intravenous delivery on cardiac function in mice as assessed by cardiac MRI studies. Hence, CTP shows great promise as a cardiomyocyte specific, non-toxic CPP, opening the doors to targeted delivery of established or novel therapies development of which has been hindered by a lack of delivery to the heart.
Results
In vitro studies
FACS: Studies were conducted in biological triplicates and technical quadruplicates (n=4), showing that cell viability did not change significantly after treatment with 10µM of CTP compared to no treatment (live-dead stain only), vehicle, or random peptide (Figure 1).
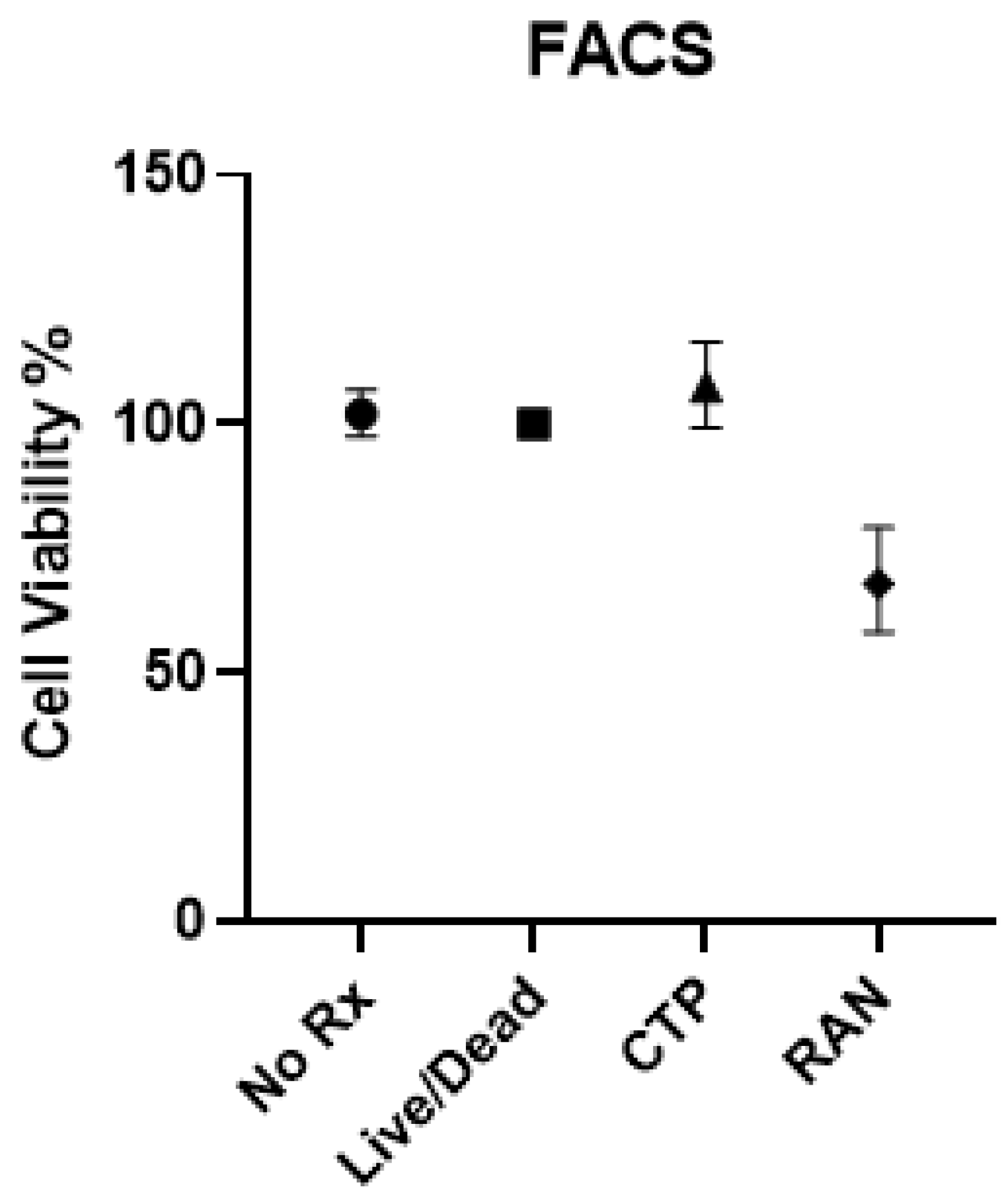
Figure 1.
Fluorescence activated cell sorting (FACS) assay of human ventricular myocyte cell line. Results of FACS show no significant reduction in cell viability between cells treated with live/dead stain only, CTP, random peptide (RAN), or no treatment as assessed by live-dead stain uptake using Fortessa X-20 for cell sorting.
Figure 1.
Fluorescence activated cell sorting (FACS) assay of human ventricular myocyte cell line. Results of FACS show no significant reduction in cell viability between cells treated with live/dead stain only, CTP, random peptide (RAN), or no treatment as assessed by live-dead stain uptake using Fortessa X-20 for cell sorting.
- Eurofins Toxicology Study
IonChannelProfiler CardiacProfiler Panel: Across all 7 ion channels tested, no inhibition/activation was observed. No significant inhibition was noted as compared to the control substances for any of the 7 ion channels (Figure 2), and therefore IC50 values were not calculable (Supplemental Table S1).
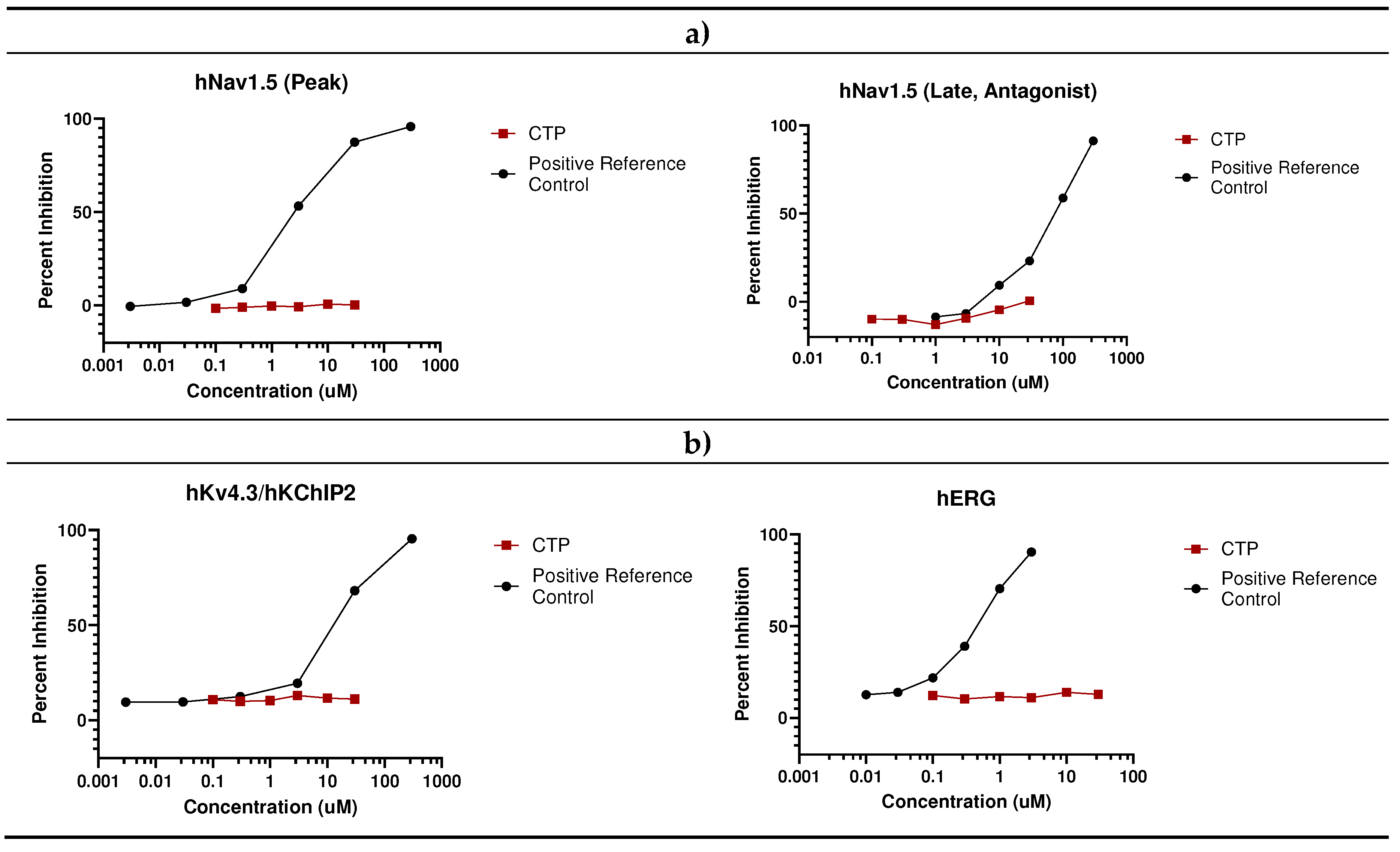
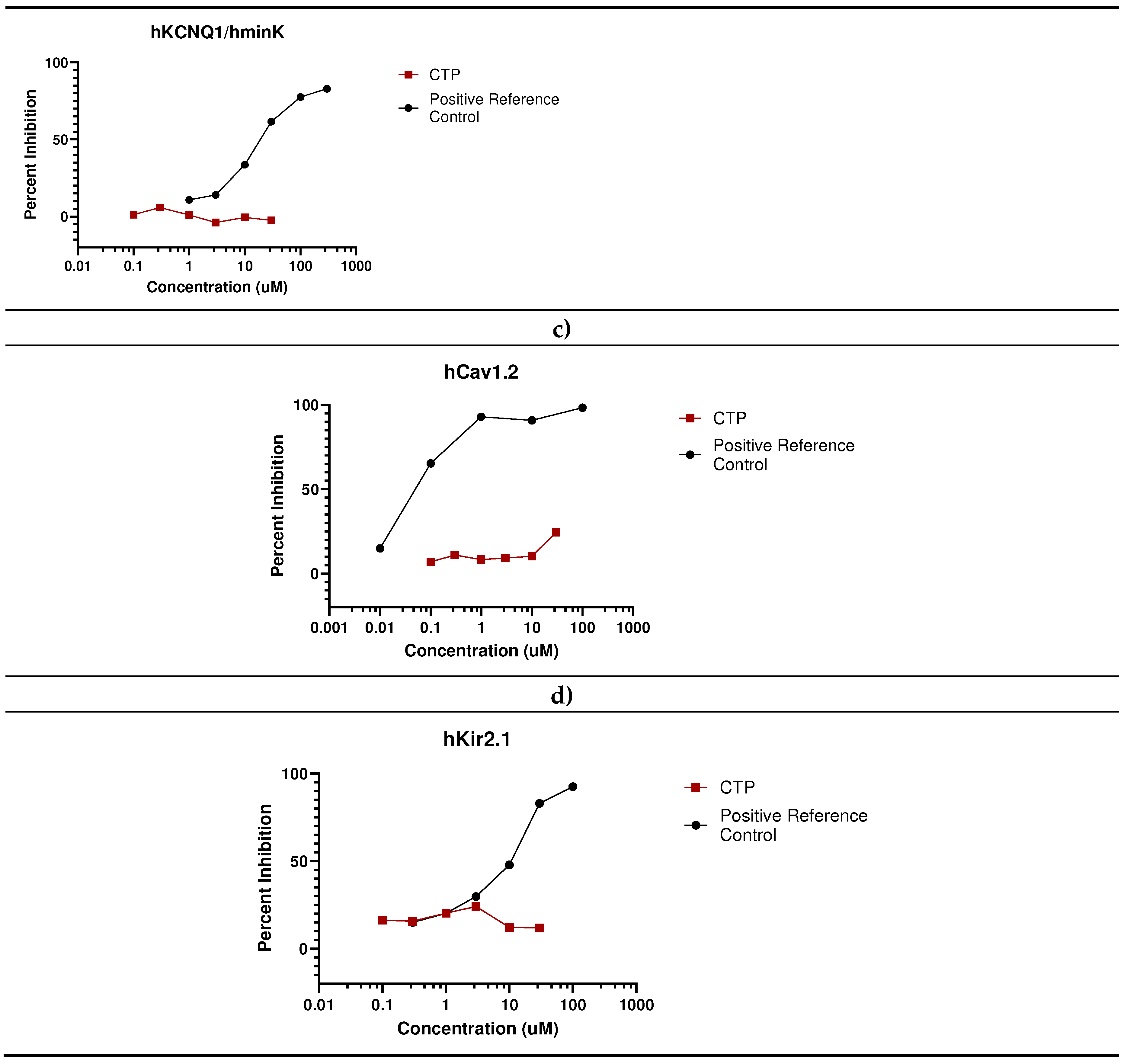
Figure 2.
Eurofins ion channel profiler results. CTP did not inhibit the activation of any tested a) voltage-gated sodium channels, b) voltage gated potassium channels, c) voltage gated calcium channels, or d) inward-rectifying voltage gated potassium channels. IC50 were incalculable for all results.
Figure 2.
Eurofins ion channel profiler results. CTP did not inhibit the activation of any tested a) voltage-gated sodium channels, b) voltage gated potassium channels, c) voltage gated calcium channels, or d) inward-rectifying voltage gated potassium channels. IC50 were incalculable for all results.
SAFETYscan E/IC50 ELECT Service: Across all seventy-eight assays, only two showed significant interactions between CTP and the assayed protein. In ion channel assays, CTP showed no significant blocking or activation of hERG, NAV1.5, or KvLQT1/minK, nor any significant opening action for KvLQT1/minK (Figure 3). In GPCR cAMP Modulation assays, CTP showed no significant action as either an agonist or antagonist of ADRA2A or ADRB1 as compared to the controls (Figure 3). In similar assays performed using OPRM1, CTP did show agonist action with an EC50 of 24.975μM, although no antagonist action of OPRM1 was noted (Figure 3). Enzymatic assays performed noted some inhibition of COX1, although it did not reach the set significant threshold. Significant inhibition of COX2 was noted with an IC50 of 16.869μM. Results of all seventy-eight assays are shown (Supplemental Figure S1). IN order to study whether activation of these two receptors was clinically relevant, we performed RT-qPCR on RNA extracted from wild-type mouse hearts. Results were normalized to GAPDH expression. Neither COX2 nor OPRM1 showed significant expression in heart tissue with COX2 expressed at a high level in lung and liver, and OPRM1 expressed in brain, lung and liver (Figure 4).
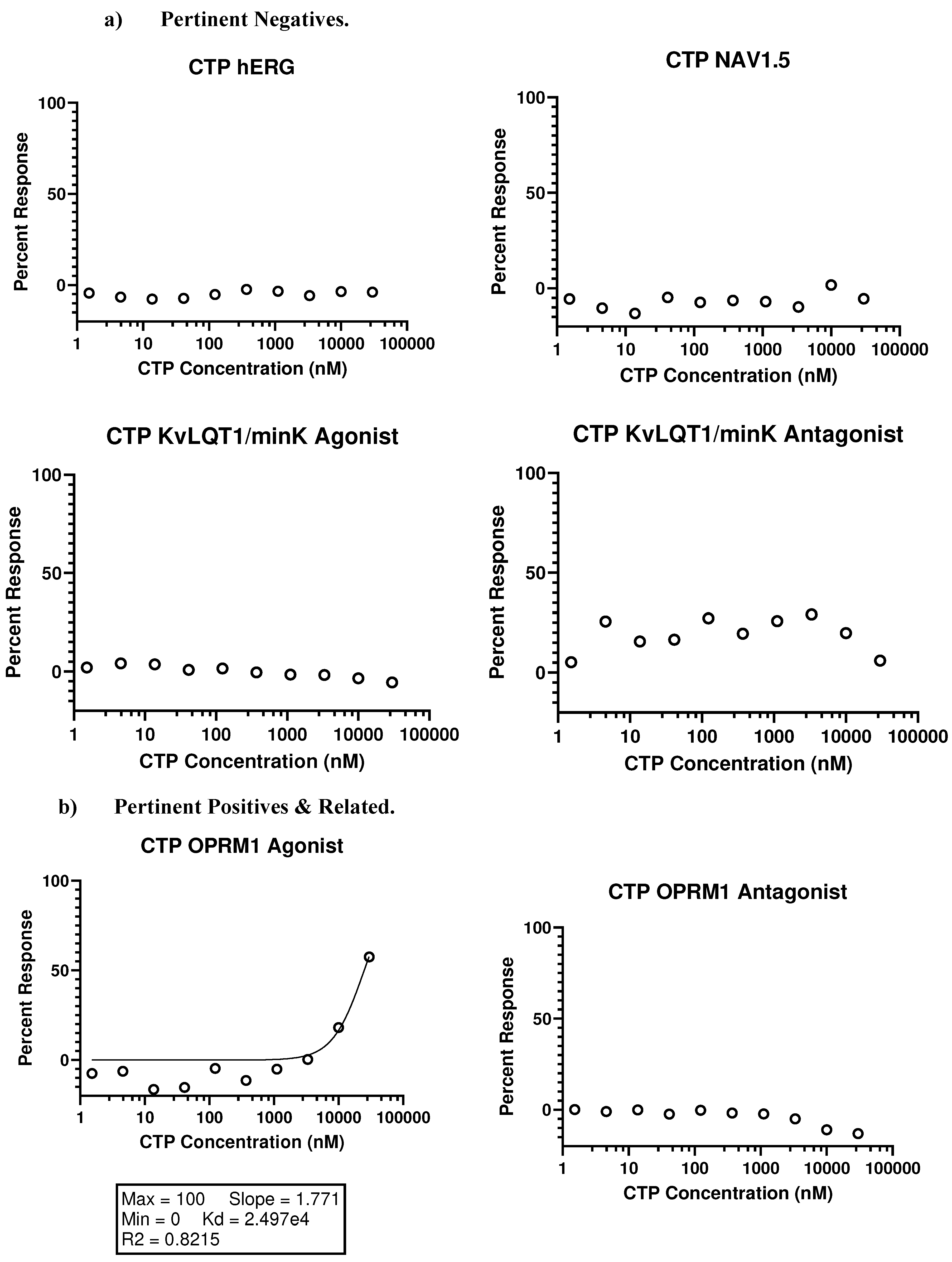
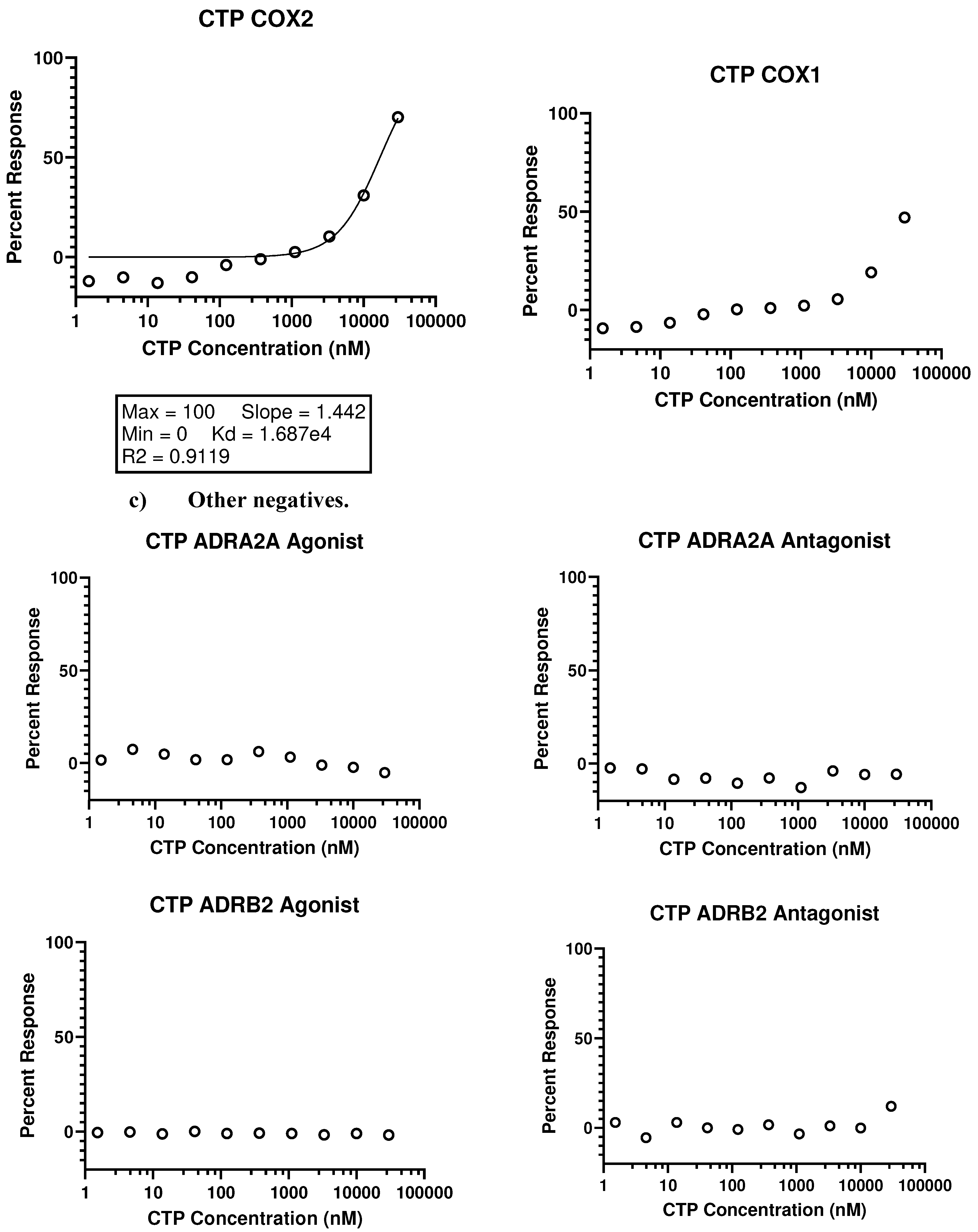
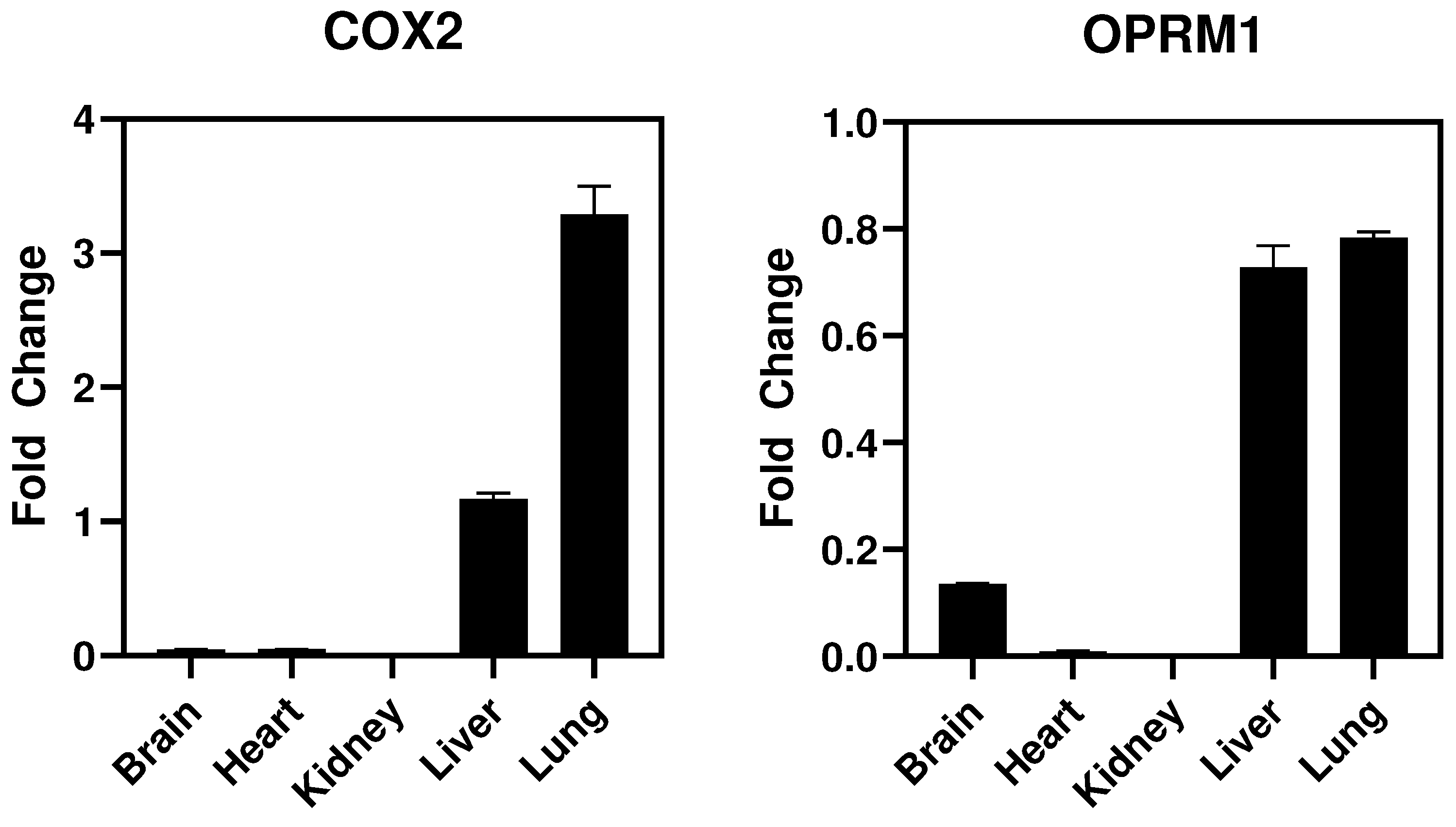
Figure 3.
Eurofins CTP SAFETY Scan results. a) high-lighted pertinent negative results, b) pertinent positives and related negatives, and c) other negative results. Data shown was normalized to the maximal and minimal response observed in the presence of control ligand and vehicle respectively (y-axis) and is plotted against the corresponding compound concentration in nM in log10 scale (x-axis).
Figure 3.
Eurofins CTP SAFETY Scan results. a) high-lighted pertinent negative results, b) pertinent positives and related negatives, and c) other negative results. Data shown was normalized to the maximal and minimal response observed in the presence of control ligand and vehicle respectively (y-axis) and is plotted against the corresponding compound concentration in nM in log10 scale (x-axis).
Figure 4.
RT-qPCR Results for COX2, and OPRM1. There is no significant expression at mRNA level of COX2 or OPRM1 in the mouse heart. Values are normalized to GAPDH expression.
Figure 4.
RT-qPCR Results for COX2, and OPRM1. There is no significant expression at mRNA level of COX2 or OPRM1 in the mouse heart. Values are normalized to GAPDH expression.
- In vivo studies
Blood Pressures Pre- and Post-Injection: No significant difference was measured in systolic or diastolic blood pressures between pre and post injection measurements (Figure 5). According to Janssen [17], average blood pressure values in wild type mice range from ~90 to 115 mmHg.
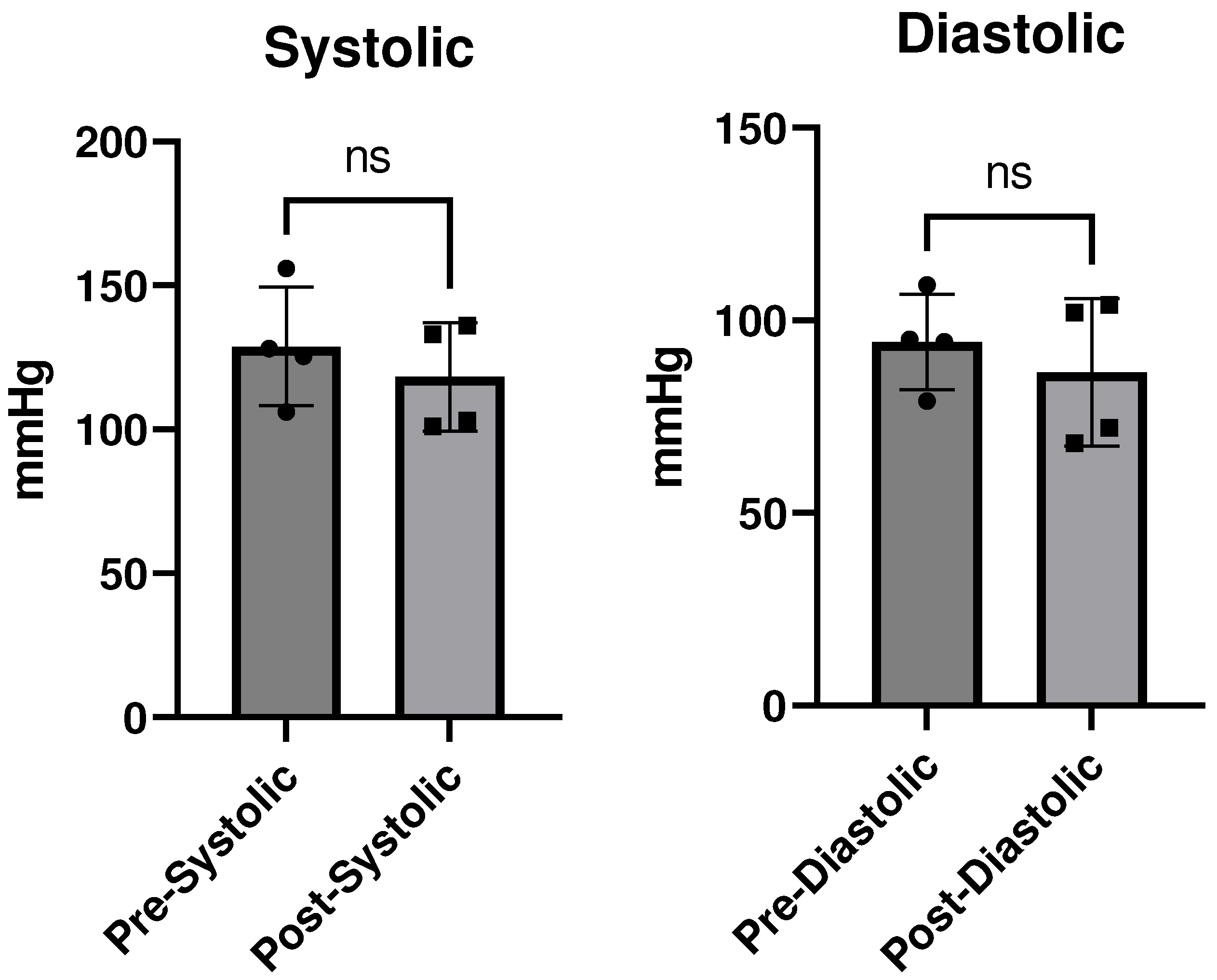
Figure 5.
Blood pressure pre and post CTP injections. There was no difference noted in systolic (a) or diastolic pressures (b) after a single CTP injection (10mg/Kg).
Figure 5.
Blood pressure pre and post CTP injections. There was no difference noted in systolic (a) or diastolic pressures (b) after a single CTP injection (10mg/Kg).
Toxicology Studies: There was no significant difference in weights during the two-week course of the study in male or female mice (Figure 6). In male mice, there was no difference noted in hemoglobin, white blood cell or platelet count, alanine aminotransferase, aspartate aminotransferase, alkaline phosphatase, total bilirubin, or phosphorous levels. Kidney function studies in male mice (blood urea nitrogen and creatinine) were significantly lower on Day 0-2 than the control mice ruling out overt kidney damage. Calcium levels increased marginally only on Day 0, normalizing at later time-points.
Female mice showed similar results. There was no significant difference overall between treated groups and control group in hemoglobin, alanine aminotransferase, aspartate aminotransferase, blood urea nitrogen or creatinine levels. A significant decrease in bilirubin, white blood cell count, and platelet count was noted only on Day 0, which recovered by Day 1 for both. The only consistent change noted was a rise in alkaline phosphatase for female mice (Figure 6). However, despite the rising alkaline phosphatase, the mean values of alkaline phosphatase level did not cross the upper limit of normal (140 IU/L) for any of the groups, and only 2 mice had borderline elevated levels (151 and 166) on day 0, with all levels returning to baseline by Day 2.
Cardiac MRI showed no significant change in heart rate, left ventricular mass, ejection fraction or cardiac output post CTP injection in male or female mice (Figure 7).
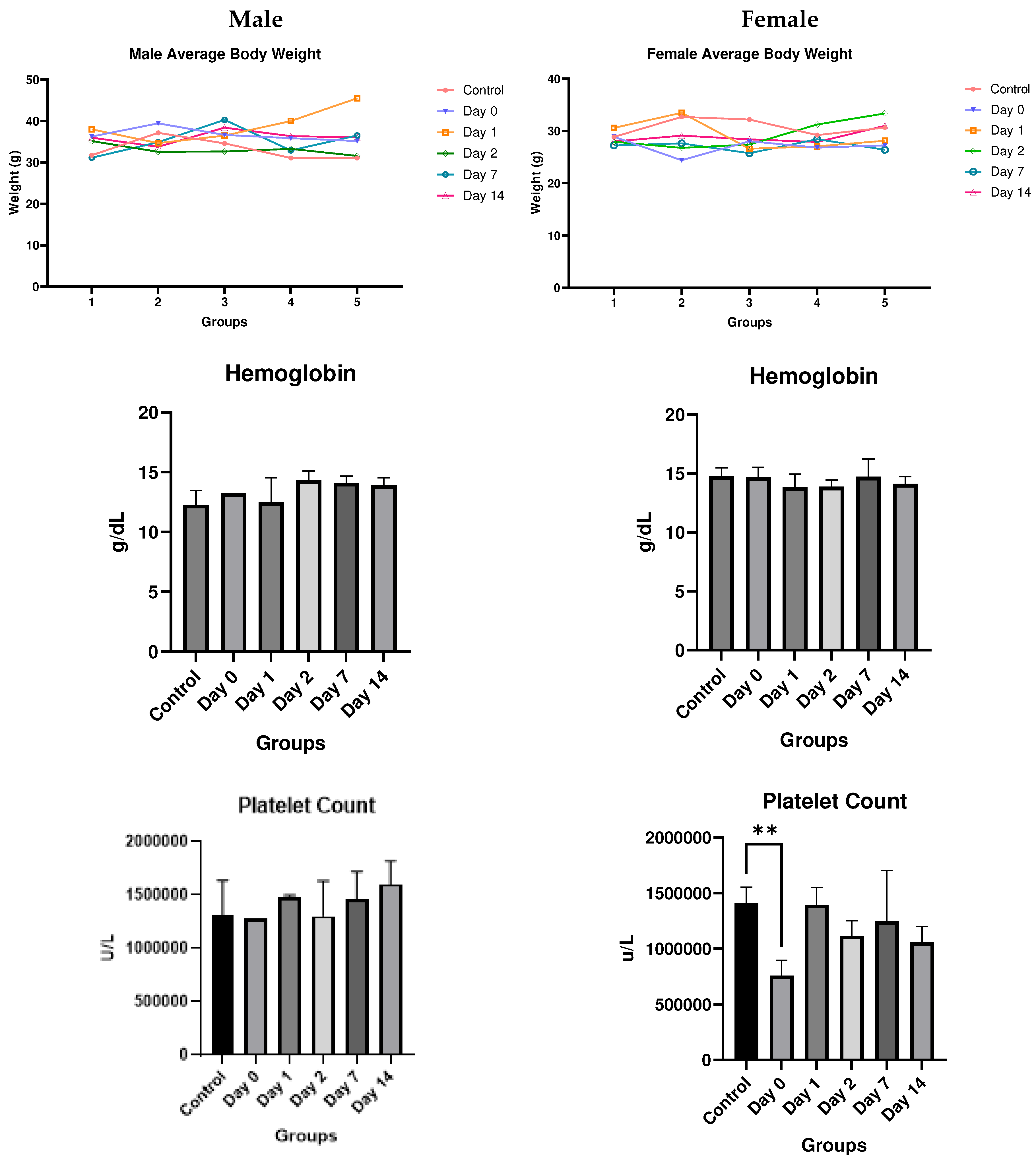
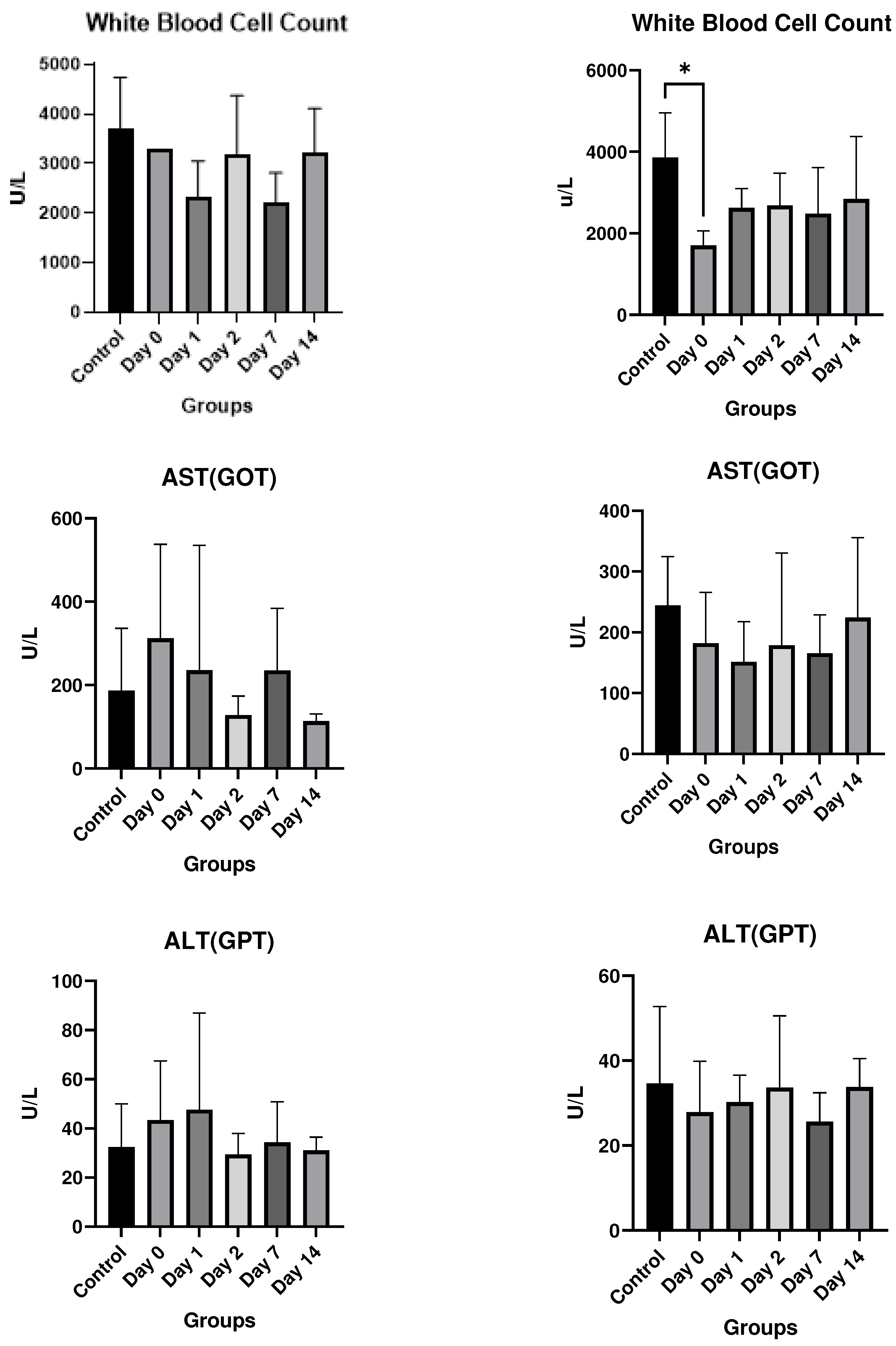
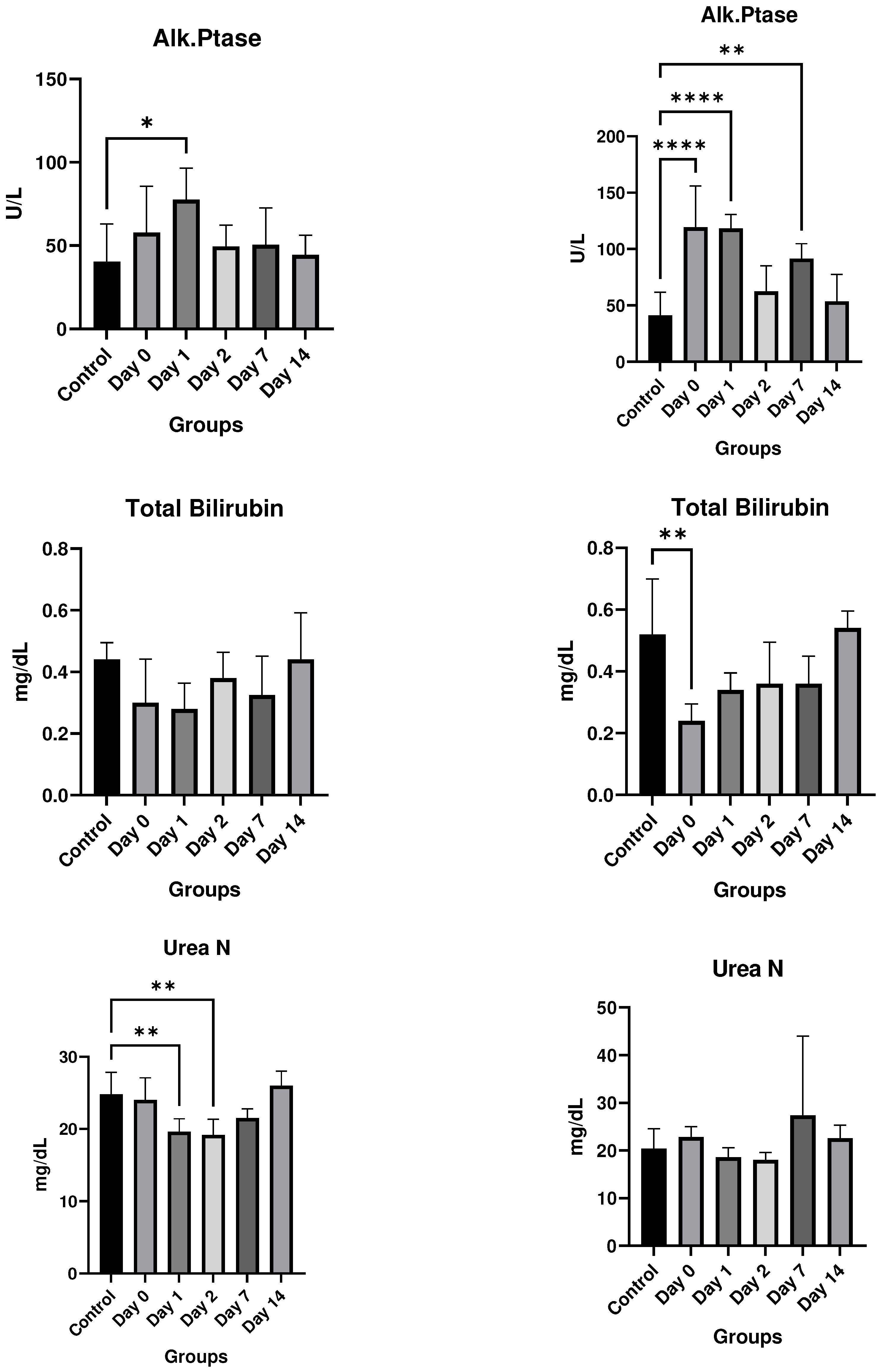
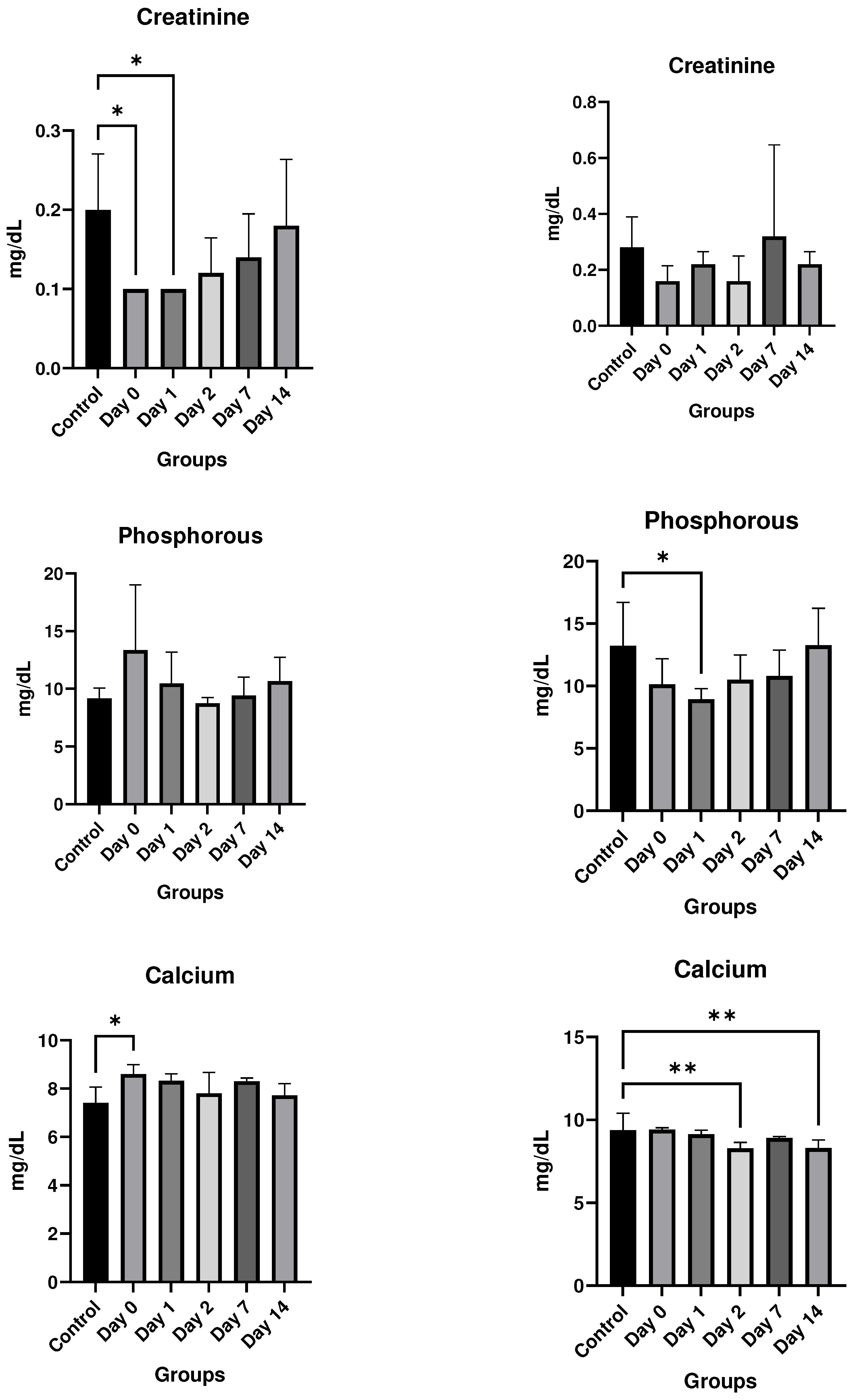
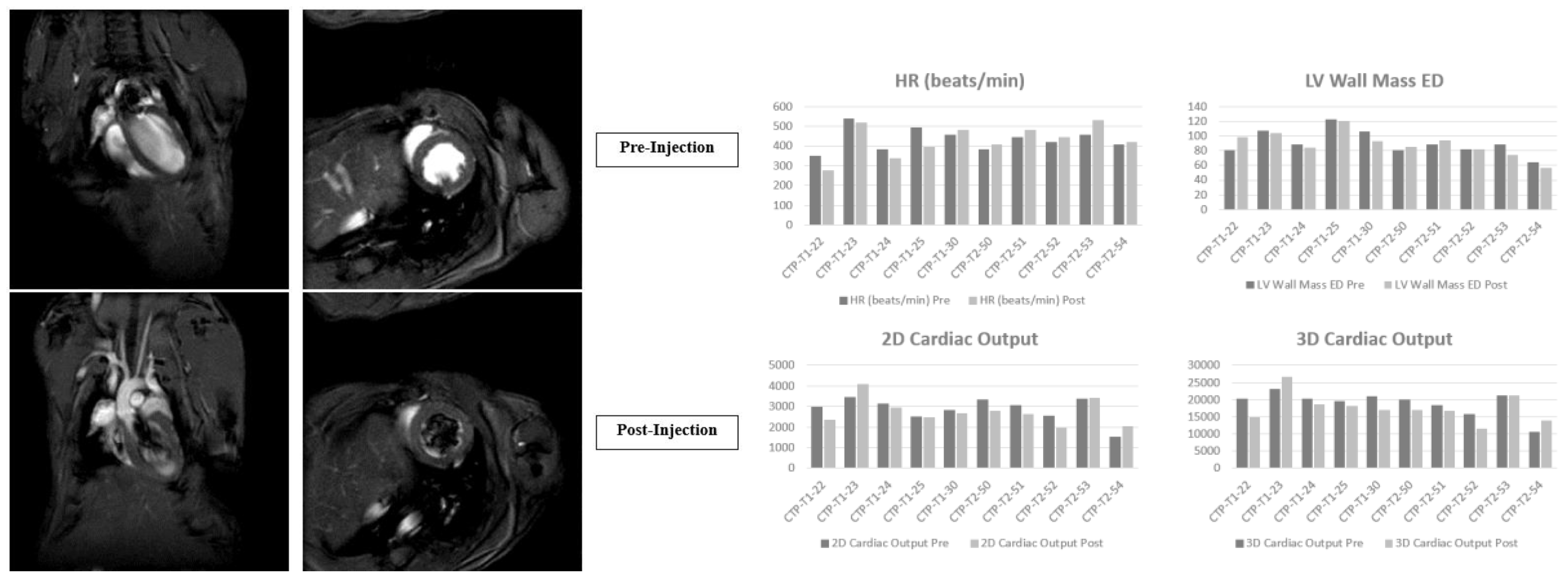
Figure 6.
In-vivo Toxicity Results in a) male and b) female mice. There was no significant change in weight, blood chemistries, liver function tests, renal function with CTP injection except for a temporary rise in alkaline phosphatase that returned to baseline by Day 1 (males) or Day 2 (females), that did not reach clinical significance of >3x upper limit of normal. Blood urea nitrogen and creatinine decreased in male mice and white blood cell count and platelets decreased in female mice, both temporary effects that could be secondary to vascular volume dilution secondary to the intravenous injections of the peptide. .
Figure 6.
In-vivo Toxicity Results in a) male and b) female mice. There was no significant change in weight, blood chemistries, liver function tests, renal function with CTP injection except for a temporary rise in alkaline phosphatase that returned to baseline by Day 1 (males) or Day 2 (females), that did not reach clinical significance of >3x upper limit of normal. Blood urea nitrogen and creatinine decreased in male mice and white blood cell count and platelets decreased in female mice, both temporary effects that could be secondary to vascular volume dilution secondary to the intravenous injections of the peptide. .
Figure 7.
Cardiac MRIs of pre and post injection of CTP. Heart beats/min, left ventricular (LV) walls mass, 2-D and 3-D cardiac outputs did not show any change in any parameter immediately after CTP injection.
Figure 7.
Cardiac MRIs of pre and post injection of CTP. Heart beats/min, left ventricular (LV) walls mass, 2-D and 3-D cardiac outputs did not show any change in any parameter immediately after CTP injection.
Discussion
The initial report of Tat’s ability to deliver beta-galactosidase, a payload much larger than Tat, in an intact, functional form was met with a lot of excitement [18]. However, Tat technology did not translate into a clinical application due to the ubiquitous transduction of multiple tissue types, including neuronal tissue with crossing of the blood-brain barrier. These hurdles were initially ameliorated by topical use of Tat conjugated to a therapeutic [19,20,21], or in a compartment-restricted manner such as intra-articular [22], or intra-ocular administration [23,24,25]. Another use of non-specific CPP was use of activatable peptides given a certain tissue milieu in oncological applications [26]. Phage display to identify specific tumor or tissue targeting CPPs was yet another strategy in the attempts to harness CPP’s transduction abilities for clinical application [7,27,28,29]. The road from bench to bedside is a long and arduous one with many pitfalls. Toxicity of CPPs, or the combination of a particular CPP with its cargo, is another major hurdle.
Our work with phage display led to the identification of our 12-amino acid long cardiomyocyte targeting peptide, CTP. It is a non-naturally occurring peptide with little homology to any naturally occurring peptides/proteins, and unlike non-cell specific CPPs is not composed of predominantly cationic amino acids. Our previous work has shown that it is taken up by cardiomyocytes in as little as 15 minutes after a peripheral intravenous injection [4,9]. Our studies and multiple investigators have shown that it targets cardiomyocytes specifically while sparing the other cells (fibroblasts, endothelial cells, myofibroblasts) in the heart [13]. To further CTP along in its journey towards becoming a clinically useful cardiomyocyte-specific vector, we performed the current set of in vitro and in vivo toxicity studies.
In the current body of work, we show that CTP did not activate or inhibit multiple different, key channels like HERG2, sodium, potassium or calcium channels that cardiomyocytes are particularly rich in. Activation or inhibition could lead to significant arrhythmias, an unacceptable and potentially life-threatening toxicity. We also show that CTP did not change the cell viability of a human myocyte cell line. For our in vivo toxicities, using a non-trivial dose of CTP (10 mg/Kg) we show no change in blood pressures. Cardiac function assessed by the most reliable imaging method available (cardiac MRI) showed no significant difference in cardiac function either in immediate post-treatment images. We also performed toxicity studies after a single dose in keeping with timelines and doses suggested by the FDA for “microdose” imaging studies. We chose this path to take as CTP is platform technology and can have both diagnostic and therapeutic applications. Using peptides for imaging applications involves a single dose, to be limited to no more than 100µg in a 70Kg human (approx. 1.4 µg/Kg), with toxicities performed at 100x the maximum allowed dose. This led to our current testing dose of 150µg/Kg. At that dose, there were no significant toxicity signals noted except for a borderline increase in alkaline phosphatase levels in female mice on Day 0 and 1, with recovery to control levels by Day 2. This statistical significance did not reach clinical significance thresholds, where a >3x the upper limit of normal for alkaline phosphatase is considered a meaningful elevation. In female mice, the white blood cell and platelet count decreased on Day 0 likely due to volume expansion associated with the injection. Male mice showed a decrease in blood urea nitrogen and creatinine levels which goes against any nephrotoxicity being associated with CTP.
Our results with CTP are consistent with reports in the literature on other CPPs. Obviously, the toxicity profile will vary from one peptide to the next, and depend on the dose tested. However, published literature suggests that toxicities remain low [30,31,32,33]. Saar and colleagues investigated membrane toxicity in cancer cell lines of 5 well characterized, non-cell specific CPPs: Ant, Tat, pVEC, MAP, and transportan 10, and found that the first three caused minimal membrane leakage of lactate dehydrogenase, whereas the latter two (MAP, transportan 10) caused significant leakage. However, none of the tested peptides had a hemolytic effect on bovine red blood cells. Another study evaluated the immunogenicity of transportan 10 and its chemical modified derivatives, PepFects, in a leukemia and peripheral blood monocyte cell line by evaluating release of multiple cytokines and apoptosis in response to incubation with these peptides at 5 and 10µM concentrations and found no increase in cytokine levels [33]. Additionally, in vivo toxicity was evaluated by injecting immunocompetent mice with 5mg/Kg of the peptides and blood collected at 24 and 48 hours for ELISA-based estimation of IL-1β and TNF-α [34]. There was no enhancement of any of these cytokine levels in response to peptide injections. Changes to cell metabolism in response to 5 CPPs (transportan, Ant, Tat, nona-arginine, MAP) were tested in Chinese hamster ovarian cell line. Analysis of the cell lysates was performed using liquid chromatography-mass spectrometry and showed the most significant changes in response to transportan, followed by Tat and MAP. Transportan affected cellular redox potential, depleted energy, and the pools of purines/pyrimidines [35]. Cells could recover from these effects at 5µM of transportan [34], but not at higher treatment concentrations. Clearly, toxicities are going to be dependent on the type of CPP being studied and the concentrations utilized in these studies [35].
Methods
In Vitro Studies
Fluorescence-Activated Cell Sorting Assays: CTP was tested in a human left ventricular myocyte cell line (Celprogen; cat #36044-15) for uptake and assessment of its effects on cell viability. Cells were plated onto 6-well plates at a density of 100,000 cells per well and incubated overnight at 37°C/5% CO2. The following day media was replaced with fresh media containing 10µM CTP-Cy5.5, or a random (RAN-CY5.5) peptide. A yellow Live-Dead stain (Invitrogen; cat #L34968) was added to all wells, including the negative control or vehicle only treated controls, and cells were incubated for 30 minutes, after which media aspirated, cells washed 3x with pre-warmed phosphate-buffered saline (PBS), trypsinized, and collected. After collection, cells were washed once with PBS, fixed with 2% paraformaldehyde at room temperature for 10 mins, washed once again and resuspended in 1000 µl of PBS. Fluorescence activated cell sorting (FACS) was performed using lasers selected for live-dead and Cy5.5 fluorophores, and 10,000 cells counted in Fortessa. The data generated was gated on the presence of the live-dead stain providing the percent viability of cells [(live/(live+dead cells))*100] after treatment with various peptides.
Eurofins Toxicology Study: In vitro toxicology studies were performed by Eurofins Scientific through both their Ion Channel Profiler and Cardiac Profiler Panel and their DiscoverX’s SAFETYscan E/IC50 SELECT service. Increasing concentrations of CTP (0.1µM to 30µM) was tested against various ion channels and a total of seventy-eight assays performed utilizing various output reads, including GPCR cAMP modulation assays, calcium mobilization assays, nuclear hormone receptor assays, KINOMEscan binding assays, ion channel assays, transporter assays, and enzymatic assays. All methods described below were performed, described, and reported by Eurofins Scientific in their study report (Supplemental Methods 1 & 2).
- Eurofins Ion Channel Cardiac Profiler Panel:
Detailed methods can be found in Supplemental Methods 1. Electrophysiological assays were conducted on four types of channels: Voltage gated sodium channels HEK-Nav1.5 (peak), and HEK-Nav1.5 (late, antagonist); voltage gated potassium channels HEK-Kv4.3/KChIP2, CHO-hERG, and CHO-KCNQ1/minK; voltage gated calcium channel HEK-Cav1.2; and inward-rectifying voltage gated potassium channel HEK-Kir2.1. IC50 values following CTP treatment at concentrations ranging from 0.1 to 30 µM were calculated using non-linear, least squares regression analyses, and compared to reference standards.
- Eurofins SAFETYscan E/IC50 ELECT Service:
GPCR cAMP modulation assays, calcium mobilization assays, nuclear hormone receptor assays, KINOMEscan binding assays, ion channel assays, transporter assays, and enzymatic assays were performed by Eurofins under GLP standards. Simplified methods are provided below; detailed methods can be found in Supplemental Methods 2.
GPCR cAMP Modulation Assays: cAMP Hunter cell lines were grown, plated, and subjected to Eurofins’ DiscoverX HitHunter cAMP XS+ assay. Gs agonist activity, Gi agonist activity, and antagonist activity induced assays were read using chemiluminescent signal detection. Percentage activity was calculated for Gs and Gi agonist assays, and percentage inhibition calculated for Gs antagonist assays.
Calcium Mobilization Assays: PathHunter cell lines were grown, plated, and subjected to calcium mobilization assays using the DiscoverX Calcium No WashPLUS kit. Agonist and antagonist activity of CTP was performed and measured on a FLIPR Tetra using a five second baseline read followed by 2 minutes of calcium mobilization monitoring. The area under the FLIPR curve and percentage activity was calculated to determine agonist activity, and percentage inhibition was calculated to determine antagonist activity.
Nuclear Hormone Receptor Assays: PathHunter NHR cell lines were grown, plated, and subjected to nuclear hormone receptor assays using the PathHunter Detection reagent cocktail. Chemiluminescent signals were read using a PerkinElmer Envision instrument, and percent activation and percent inhibition were calculated to determine agonist and antagonist activity, respectively.
KINOMEscan Binding Assays: Kinases were produced by the infection of BL21 E. coli using kinase tagged T7 phage strains. Cells were grown, lysed, centrifuged, filtered, and the remaining kinases produced in HEK-293 cells were tagged using DNA for detection using qPCR. Affinity resins for the kinase assays were created using Streptavidin-coated magnetic beads treated with biotinylated small molecule ligands. Kinases, beads, and CTP were combined in 1x binding buffer to perform the assay. Kinase concentrations were measured using qPCR, and percent response and binding constants were calculated.
Ion Channel Assays: Cell lines were grown, plated, and loaded with 1x loading buffer containing 1x dye and 2.5 mM Probenecid freshly prepared. Assays were performed, and activity was measured using a FLIPR Tetra. Percent activation and percent inhibition were calculated to determine agonist and antagonist activity, respectively.
Transporter Assays: Cell lines were grown, plated, and incubated first with CTP, then with 1x loading buffer containing 1x dye with 1x HBSS and 20mM HEPES. Fluorescence signals were detected using a PerkinElmer Envision, and percentage inhibition assessed.
Enzymatic Assays: AChE, COX1, COX2, MAOA, PDE3A and PDE4D2 activity assays were performed to determine CTP effects on each individual enzyme. Fluorescence was read using a PerkinElmer Envision, and percentage inhibition and compound activity calculated.
In Vivo Studies
All animal protocols were approved by the institutional animal care and committee of the University of Pittsburgh and Mayo Clinic. CTP was synthesized by the University of Pittsburgh using standard solid-state synthesis, followed by HPLC purification. MALDI LC/MS characterization of the final peptide was performed prior to use.
Blood Pressures Pre and Post CTP Injection: Six B6-wildtype mice were obtained from the Jackson Laboratory. Blood pressures were taken using a non-invasive blood pressure system from Kent Scientific. Mice were anesthetized using 2.5% isoflurane exposure for 5 minutes, placed in a tube holder, and warmed using a warming blanket for approximately 15 minutes before their tails were fitted with blood pressure cuffs. 21 blood pressure readings were taken, and average systolic and diastolic blood pressure calculated. After baseline blood pressure was established, mice were injected intravenously with 10mg/kg of CTP, and blood pressures were measured again 6 hours post-injection.
Mouse Toxicity Studies: Sixty CD1-wildtype mice (1:1 male to female ratio) were obtained from Charles River and divided into 6 groups of 10, 5 females and 5 males each. Ten control mice were injected with 1X PBS. Treated mice were injected intravenously with a single dose of CTP 150ug/Kg. Groups of 10 treated mice (5 male, 5 female) were euthanized on day 0 (immediately after injection), 1, 2, 7, and 14 post-injection using inhalational CO2. After euthanasia, chest cavity was opened, and blood drawn via cardiac puncture for complete blood count and serum chemistries. Following the blood draw, the right atrium was nicked and the whole animal perfusion fixed using 3ml of formalin injected into the left ventricular apex. The control group was euthanized on day 2. A complete blood count, metabolic profile and thyroid studies were performed.
Cardiac Function Studies, Pre and Post Injections: Six CD1, wild type, 6–8-week-old adult mice were obtained from Charles River and used for cardiac MRI studies. Mice were weighed, anesthetized with 2.5% isoflurane, and ear tags removed before being inserted into the MRI scanner. A baseline cardiac MRI was performed, followed by intravenous CTP injection (150 µg/Kg). Post-injection, a second cardiac MRI was performed.
Real Time quantitative PCR: Based on the results of the Eurofins Toxicology Study (Figure 3), targets for qPCR were identified for expression of the activated receptors, OPRM1, COX1, and COX2 in mouse hearts. Heart, brain, kidney, liver, and lungs were harvested from an untreated, wild-type CD1, adult mouse, and snap frozen in liquid nitrogen. RNA extraction was performed using the Qiagen RNeasy Micro Kit and cDNA libraries prepared using the high-capacity RNA to cDNA kit (Applied Biosciences). RT-qPCR was performed (ABI Quant) using PowerUp SYBR Green Master Mix (Applied Biosciences). Results were normalized to GAPDH expression.
Conclusion
CTP is a safe and effective new technology for the delivery of therapeutic cargoes to cardiomyocytes of the heart. CTP showed no overt toxicities, and caused no major changes in hematological counts, blood chemistries or MRI-based cardiac function. In vitro cell assays showed an activation of OPRM1 and COX 2 receptors, neither of which is expressed in the normal murine heart, or shown to express in large proteomics based datasets of the human heart. CTP is an extremely promising, novel drug delivery method, and studies to deliver diagnostic imaging agents and therapeutics to the heart are ongoing.
Our study has several limitations worth noting. The studies were done in a non-GLP environment, with a single dose of CTP studied in vivo. Although we did not find any toxicities associated with CTP, the same cannot be said of CTP conjugated to a specific cargo, or repeated dose administrations. Toxicity studies with a specific payload will need to be repeated in two vertebrate animal species, at least one of which must be a non-rodent one, in a longer timeline fashion, and in a GLP environment before CTP can make it as a viable vector to its first Phase I human study.
Supplemental Material Descriptions
Supplemental Methods 1: Full and detailed methods of the Eurofins Ion Channel Cardiac Profiler Panel as performed and reported by Eurofins. Supplemental Methods 2: Full and detailed methods of the Eurofins SAFETYscan E/IC50 ELECT Service, including principles and methods, as performed and reported by Eurofins. Supplemental Table S1: IC50 Values for CTP as performed and reported by Eurofins in their Ion Channel Cardiac Profiler Panel. Supplemental Figure S1: Test dose response curves for all 78 assays reported by Eurofins in their SAFETYscan E/IC50 ELECT Service.
Author Contributions
Conceptualization, MZ; Methodology, MZ; Software, KF; Validation, JL; Formal Analysis, DS, JL, KF; Investigation, KF, NP; Resources, MZ; Data Curation, NP, KF; Writing – Original Draft Preparation, DS, JL; Writing – Review & Editing, MZ; Visualization, DS, JL; Supervision, MZ; Project Administration, MZ; Funding Acquisition, MZ.
Funding
This work was supported by the Pitt Innovation Challenge, a program of the Clinical and Translational Science Institute at the University of Pittsburgh. DS, JL and MZ are supported by NIH grant R01HL153407,both awarded to MZ.
Conflict of Interest:
MZ holds a patent on use of cardiac targeting peptide for cardiac delivery along with Dr. Paul Robbins and University of Pittsburgh, PA.
References
- Derakhshankhah, H.; Jafari, S. Cell penetrating peptides: A concise review with emphasis on biomedical applications. Biomed Pharmacother 2018, 108, 1090–1096. [Google Scholar] [CrossRef]
- Frankel, A.D.; Pabo, C.O. Cellular uptake of the tat protein from human immunodeficiency virus. Cell 1988, 55, 1189–1193. [Google Scholar] [CrossRef]
- Green, M.; Loewenstein, P.M. Autonomous functional domains of chemically synthesized human immunodeficiency virus tat trans-activator protein. Cell 1988, 55, 1179–1188. [Google Scholar] [CrossRef]
- Zahid, M.; Phillips, B.E.; Albers, S.M.; Giannoukakis, N.; Watkins, S.C.; Robbins, P.D. Identification of a cardiac specific protein transduction domain by in vivo biopanning using a M13 phage peptide display library in mice. PLoS One 2010, 5, e12252. [Google Scholar] [CrossRef]
- Pasqualini, R.; Koivunen, E.; Ruoslahti, E. A peptide isolated from phage display libraries is a structural and functional mimic of an RGD-binding site on integrins. J Cell Biol 1995, 130, 1189–1196. [Google Scholar] [CrossRef]
- Rajotte, D.; Arap, W.; Hagedorn, M.; Koivunen, E.; Pasqualini, R.; Ruoslahti, E. Molecular heterogeneity of the vascular endothelium revealed by in vivo phage display. J Clin Invest 1998, 102, 430–437. [Google Scholar] [CrossRef]
- Arap, W.; Pasqualini, R.; Ruoslahti, E. Cancer treatment by targeted drug delivery to tumor vasculature in a mouse model. Science 1998, 279, 377–380. [Google Scholar] [CrossRef]
- Zahid, M.; Robbins, P.D. Protein transduction domains: applications for molecular medicine. Curr Gene Ther 2012, 12, 374–380. [Google Scholar] [CrossRef]
- Zahid, M.; Feldman, K.S.; Garcia-Borrero, G.; Feinstein, T.N.; Pogodzinski, N.; Xu, X.; Yurko, R.; Czachowski, M.; Wu, Y.L.; Mason, N.S.; et al. Cardiac Targeting Peptide, a Novel Cardiac Vector: Studies in Bio-Distribution, Imaging Application, and Mechanism of Transduction. Biomolecules 2018, 8. [Google Scholar] [CrossRef]
- Ahmad, F.B.; Anderson, R.N. The Leading Causes of Death in the US for 2020. JAMA 2021, 325, 1829–1830. [Google Scholar] [CrossRef]
- Pandey, M.; Gundam, S.R.; Bansal, A.; Kethamreddy, M.; Lowe, V.; Zahid, M. Synthesis and evaluation of novel [68Ga]Ga-NOTA-CTP in normal CD-1 mice as a myocardial perfusion imaging PET probe. Journal of Nuclear Medicine 2023, 64, P1066–P1066. [Google Scholar]
- Kim, H.; Mun, D.; Kang, J.Y.; Lee, S.H.; Yun, N.; Joung, B. Improved cardiac-specific delivery of RAGE siRNA within small extracellular vesicles engineered to express intense cardiac targeting peptide attenuates myocarditis. Mol Ther Nucleic Acids 2021, 24, 1024–1032. [Google Scholar] [CrossRef]
- Avula, U.M.; Yoon, H.K.; Lee, C.H.; Kaur, K.; Ramirez, R.J.; Takemoto, Y.; Ennis, S.R.; Morady, F.; Herron, T.; Berenfeld, O.; et al. Cell-selective arrhythmia ablation for photomodulation of heart rhythm. Sci Transl Med 2015, 7, 311ra172. [Google Scholar] [CrossRef]
- Gallicano, G.I.; Fu, J.; Mahapatra, S.; Sharma, M.V.R.; Dillon, C.; Deng, C.; Zahid, M. Reversing Cardiac Hypertrophy at the Source Using a Cardiac Targeting Peptide Linked to miRNA106a: Targeting Genes That Cause Cardiac Hypertrophy. Pharmaceuticals (Basel) 2022, 15. [Google Scholar] [CrossRef]
- Yurko, R.; Islam, K.; Weber, B.; Salama, G.; Zahid, M. Conjugation of amiodarone to a novel cardiomyocyte cell penetrating peptide for potential targeted delivery to the heart. Front Chem 2023, 11, 1220573. [Google Scholar] [CrossRef]
- Zahid, M.; Weber, B.; Yurko, R.; Islam, K.; Agrawal, V.; Lopuszynski, J.; Yagi, H.; Salama, G. Cardiomyocyte-Targeting Peptide to Deliver Amiodarone. Pharmaceutics 2023, 15. [Google Scholar] [CrossRef]
- Janssen, B.J.; Smits, J.F. Autonomic control of blood pressure in mice: basic physiology and effects of genetic modification. Am J Physiol Regul Integr Comp Physiol 2002, 282, R1545–1564. [Google Scholar] [CrossRef]
- Schwarze, S.R.; Ho, A.; Vocero-Akbani, A.; Dowdy, S.F. In vivo protein transduction: delivery of a biologically active protein into the mouse. Science 1999, 285, 1569–1572. [Google Scholar] [CrossRef]
- Manosroi, J.; Lohcharoenkal, W.; Gotz, F.; Werner, R.G.; Manosroi, W.; Manosroi, A. Transdermal absorption and stability enhancement of salmon calcitonin by Tat peptide. Drug Dev Ind Pharm 2013, 39, 520–525. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Lee, K.H.; Huang, Y.; Shin, M.C.; Park, Y.S.; Kim, H.; Moon, C. Topical Delivery of Cell-Penetrating Peptide-Modified Human Growth Hormone for Enhanced Wound Healing. Pharmaceuticals (Basel) 2023, 16. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Rao, P.; Bradshaw, J.; Weller, R. Topical application of superoxide dismutase mediated by HIV-TAT peptide attenuates UVB-induced damages in human skin. Eur J Pharm Biopharm 2016, 107, 286–294. [Google Scholar] [CrossRef]
- Mailhiot, S.E.; Thompson, M.A.; Eguchi, A.E.; Dinkel, S.E.; Lotz, M.K.; Dowdy, S.F.; June, R.K. The TAT Protein Transduction Domain as an Intra-Articular Drug Delivery Technology. Cartilage 2021, 13, 1637S–1645S. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, Y.; Lin, T.; Yin, H.; Pan, Y.; Zhu, M.; Zhang, M. A cell-permeable peptide inhibitor of p55PIK signaling alleviates suture-induced corneal neovascularization and inflammation. Heliyon 2023, 9, e14869. [Google Scholar] [CrossRef]
- Ozaki, T.; Nakazawa, M.; Yamashita, T.; Sorimachi, H.; Hata, S.; Tomita, H.; Isago, H.; Baba, A.; Ishiguro, S. Intravitreal injection or topical eye-drop application of a mu-calpain C2L domain peptide protects against photoreceptor cell death in Royal College of Surgeons' rats, a model of retinitis pigmentosa. Biochim Biophys Acta 2012, 1822, 1783–1795. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Gomara, M.J.; Haro, I. Atorvastatin-loaded peptide amphiphiles against corneal neovascularization. Nanomedicine (Lond) 2023, 18, 1095–1108. [Google Scholar] [CrossRef]
- Olson, E.S.; Aguilera, T.A.; Jiang, T.; Ellies, L.G.; Nguyen, Q.T.; Wong, E.H.; Gross, L.A.; Tsien, R.Y. In vivo characterization of activatable cell penetrating peptides for targeting protease activity in cancer. Integr Biol (Camb) 2009, 1, 382–393. [Google Scholar] [CrossRef]
- Whitney, M.; Crisp, J.L.; Olson, E.S.; Aguilera, T.A.; Gross, L.A.; Ellies, L.G.; Tsien, R.Y. Parallel in vivo and in vitro selection using phage display identifies protease-dependent tumor-targeting peptides. J Biol Chem 2010, 285, 22532–22541. [Google Scholar] [CrossRef]
- Koivunen, E.; Arap, W.; Valtanen, H.; Rainisalo, A.; Medina, O.P.; Heikkila, P.; Kantor, C.; Gahmberg, C.G.; Salo, T.; Konttinen, Y.T.; et al. Tumor targeting with a selective gelatinase inhibitor. Nat Biotechnol 1999, 17, 768–774. [Google Scholar] [CrossRef]
- Pasqualini, R.; Ruoslahti, E. Organ targeting in vivo using phage display peptide libraries. Nature 1996, 380, 364–366. [Google Scholar] [CrossRef]
- Jones, S.W.; Christison, R.; Bundell, K.; Voyce, C.J.; Brockbank, S.M.; Newham, P.; Lindsay, M.A. Characterisation of cell-penetrating peptide-mediated peptide delivery. Br J Pharmacol 2005, 145, 1093–1102. [Google Scholar] [CrossRef]
- Patel, S.G.; Sayers, E.J.; He, L.; Narayan, R.; Williams, T.L.; Mills, E.M.; Allemann, R.K.; Luk, L.Y.P.; Jones, A.T.; Tsai, Y.H. Cell-penetrating peptide sequence and modification dependent uptake and subcellular distribution of green florescent protein in different cell lines. Sci Rep 2019, 9, 6298. [Google Scholar] [CrossRef]
- Hoffmann, K.; Milech, N.; Juraja, S.M.; Cunningham, P.T.; Stone, S.R.; Francis, R.W.; Anastasas, M.; Hall, C.M.; Heinrich, T.; Bogdawa, H.M.; et al. A platform for discovery of functional cell-penetrating peptides for efficient multi-cargo intracellular delivery. Sci Rep 2018, 8, 12538. [Google Scholar] [CrossRef]
- Saar, K.; Lindgren, M.; Hansen, M.; Eiriksdottir, E.; Jiang, Y.; Rosenthal-Aizman, K.; Sassian, M.; Langel, U. Cell-penetrating peptides: a comparative membrane toxicity study. Anal Biochem 2005, 345, 55–65. [Google Scholar] [CrossRef]
- Suhorutsenko, J.; Oskolkov, N.; Arukuusk, P.; Kurrikoff, K.; Eriste, E.; Copolovici, D.M.; Langel, U. Cell-penetrating peptides, PepFects, show no evidence of toxicity and immunogenicity in vitro and in vivo. Bioconjug Chem 2011, 22, 2255–2262. [Google Scholar] [CrossRef]
- Kilk, K.; Mahlapuu, R.; Soomets, U.; Langel, U. Analysis of in vitro toxicity of five cell-penetrating peptides by metabolic profiling. Toxicology 2009, 265, 87–95. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



